
cardiovascular, metabolic, endocrine and behavioral aspects
281
CARDIOVASCULAR, METABOLIC, ENDOCRINE AND BEHAVIORAL ASPECTS OF DEVELOPMENT IN POSTNATAL LAMBS IN RELATION TO AGE, SEX, LAMB NUMBER AND ACUTE FLUOXETINE ADMINISTRATION by Tuan-Anh Thi Nguyen M.D., University Training Center for Health Care Professional, HoChiMinh City, Vietnam, 2001 MSc., The University of British Columbia, 2008 A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in THE FACULTY OF GRADUATE STUDIES (REPRODUCTIVE AND DEVELOPMENTAL SCIENCES) THE UNIVERSITY OF BRITISH COLUMBIA (Vancouver) July 2013 © Tuan-Anh Thi Nguyen, 2013
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of cardiovascular, metabolic, endocrine and behavioral aspects

CARDIOVASCULAR, METABOLIC, ENDOCRINE AND BEHAVIORAL ASPECTS OF DEVELOPMENT IN POSTNATAL LAMBS IN RELATION TO AGE, SEX,
LAMB NUMBER AND ACUTE FLUOXETINE ADMINISTRATION
by
Tuan-Anh Thi Nguyen
M.D., University Training Center for Health Care Professional, HoChiMinh City, Vietnam, 2001
MSc., The University of British Columbia, 2008
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
in
THE FACULTY OF GRADUATE STUDIES
(REPRODUCTIVE AND DEVELOPMENTAL SCIENCES)
THE UNIVERSITY OF BRITISH COLUMBIA
(Vancouver)
July 2013
© Tuan-Anh Thi Nguyen, 2013

ii
Abstract
Human newborns exposed in utero to maternally administered SSRIs such as
fluoxetine (FX) have an increased risk of adverse pregnancy outcomes including poor
neonatal adaptation. This comprises respiratory difficulty, jitteriness, cyanosis when feeding
and persists for several days after birth. Several potential mechanisms underlying these
symptoms have been proposed: 1) acute toxicity to the drugs (i.e. serotonin syndrome), 2)
withdrawal syndrome due to the sudden discontinuation of maternal-fetal placental drug
transfer at birth or 3) an SSRIs-elicited alteration in fetal brain development. However, the
actual mechanism has not been elucidated. In addition, the safety of SSRIs for the treatment
of depression in children and adolescents is uncertain. Thus, we conducted experiments in
which acute i.v FX (1mg/kg) and sterile water were given to FX and control groups in the
lambs at ~ 3,10 days, 1,3,6 and 12 months of age. In another cohort, daily 50mg FX was
given i.v to 5 pregnant ewes via implanted catheters during late gestation (131-132d, term =
147d) for 2 weeks. Plasma FX concentrations in the newborn acute FX group were within the
range measured in human infants at the same age. There was a lack of significant acute FX
effects on cardiovascular-respiratory, behavioral and endocrine functions in ~ 4 day old
lambs. In the lambs exposed to FX prenatally, plasma FX at birth and postnatal day (PND) 2
were low and undetectable on PND 5,10,14. However, prenatal FX-exposed lambs were
hyperactive during PND 4 to14 and their heart rate variability (HRV) was significantly lower
than control lambs on PND 2. Results suggested that SSRI toxicity and withdrawal are
unlikely to be the mechanism underlying poor neonatal adaptation in human exposed to FX
in utero. However, acute FX effects were seen in some, but not all, age groups. No acute FX
effects were observed in the young lambs (10d and 3 months of age). However, hypoactivity,

iii
transient hypoxemia, hypertension and increased HRV occurred after acute FX
administration in the older lambs (1,6 and 12 months of age). Hypoxemia and hypertension
effects were more profound in males. No changes in HPA axis function were observed.

iv
Preface
I wrote this thesis with direction and input from Dr. Dan Rurak and my supervisory
committee members: Drs. Tim Oberlander, Ken Lim, Wayne Riggs and Anthony Perks.
Surgeries in the ewes and lambs were performed by Dr. Rurak or a clinical veterinarian, Dr.
Bev Chua with assistance from myself and Tim Chow, whose doctoral thesis related to the
determination of the disposition of FX in the postnatal lambs. (Chow, 2013) Daily catheter
flushing and lamb weights were done by myself and Tim. The experiments on the lamb
included in Chapters 3 to 5 were conducted by myself and Tim Chow with occasional
support from our supervisor, Dr. Dan Rurak. I was responsible for collecting heart rate, blood
pressure, heart rate variability data and blood gas samples. Blood gas samples were run by
myself and Dr. Rurak. I was also responsible for collecting plasma samples for ACTH and
cortisol. Tim Chow was responsible for collecting plasma samples for FX measurement and
urine samples as well as developing and running the LC/MS/MS assay. (Chow et al, 2011)
Thus the plasma FX concentration versus time curve in figure 4.1 is from Tim’s thesis.
However, although Tim analyzed the plasma FX concentrations in Chapter 6, these data are
not part of his PhD thesis, I did all the remaining data analysis from actiwatch, Digital Video
Recording (DVR), power lab, ECG, blood gas, ACTH and cortisol assay.
All experiments, sample collection and data analysis in the ewes and lambs included
in Chapter 6 were done by myself. The catheter implantation procedure in the pregnant ewes
was performed by Dr. Bev Chua with assistance from myself, Dr. Rurak and the staff in the
Centre of Comparative Medicine (CCM). FX measurement in this chapter was also run by
Tim Chow.

v
Manuscripts will be prepared for future publication based on results from Chapters 3-
6. Manuscripts will include relevant sections found in Chapters 1-7 including introduction,
methods, results and discussions.
This thesis was conducted under animal care approval from the University of British
Columbia Animal Research Ethics (Certificate number: A07-0302)

vi
Table of Contents
Abstract .................................................................................................................................... ii
Preface ..................................................................................................................................... iv
Table of Contents ................................................................................................................... vi
List of Tables .......................................................................................................................... ix
List of Figures .......................................................................................................................... x
List of Symbols and Abbreviations .................................................................................... xvi
Acknowledgements ............................................................................................................ xviii
Dedication ............................................................................................................................. xix
1. Introduction ......................................................................................................................... 1 1.1 Postnatal Lamb Development: .............................................................................................. 1
1.1.1 Cardiovascular Function: .................................................................................................. 1 1.1.2 Homeostasis And Metabolism: ......................................................................................... 2 1.1.3 Behavior: Maternal-Infant Bonding: ................................................................................ 4 1.1.4 Hypothalamic-Pituitary-Adrenal (HPA) Axis Function: .................................................. 6 1.1.5 Renal Function: ................................................................................................................. 7
1.2 Depression: Symptoms And Diagnosis:................................................................................ 8 1.3 Pathophysiology Of Depression: ........................................................................................ 10 1.4 Depression In Pregnancy, Postpartum And In Children: .................................................... 16
1.4.1 Incidence: ........................................................................................................................ 16 1.4.2 Risk Factors Of Depression: ........................................................................................... 17 1.4.3 Effects Of Maternal Depression On Fetal Neurobehavioral Development: ................... 19 1.4.4 Treatment Of Depression: ............................................................................................... 20
1.5 Selective Serotonin Reuptake Inhibitors (SSRIs): .............................................................. 21 1.5.1 Clinical Uses: .................................................................................................................. 21 1.5.2 Mechanism Of Action: ................................................................................................... 21 1.5.3 Pharmacokinetics: ........................................................................................................... 24 1.5.4 Side Effects: .................................................................................................................... 26
1.6 Fluoxetine (FX): .................................................................................................................. 27 1.6.1 Pharmacokinetics: ........................................................................................................... 28
1.6.1.1 Absorption: ............................................................................................................ 28 1.6.1.2 Distribution: ........................................................................................................... 28 1.6.1.3 Metabolism: ........................................................................................................... 28 1.6.1.4 Elimination: ............................................................................................................ 29
1.7 FX Effects On Newborns And Infants: ............................................................................... 29 1.7.1 FX Effects On Cardiovascular Functions: ...................................................................... 29 1.7.2 FX Effects On The Homeostatic And Respiratory Functions: ....................................... 32 1.7.3 FX Effects On Birth Outcomes: ..................................................................................... 32 1.7.4 FX Effects On Neonatal And Child Behaviors: .............................................................. 34 1.7.5 FX Effects On The Hypothalamic-Pituitary-Adrenal (HPA) Axis: ................................ 37
1.8 Rationale: ............................................................................................................................ 39 1.9 Objectives: .......................................................................................................................... 41 1.10 Hypotheses: ......................................................................................................................... 42
2. Materials and Methods ..................................................................................................... 43

vii
2.1.1 Animals: .......................................................................................................................... 43 2.1.2 Surgical Preparation: ...................................................................................................... 44 2.1.3 Experimental Protocol: ................................................................................................... 46 2.1.4 Sample Collection:.......................................................................................................... 48 2.1.5 Digital Video Recording And Behavioral Evaluation: ................................................... 49 2.1.6 Actiwatch Rest/Activity Monitoring: ............................................................................. 50 2.1.7 Powerlab Data Acquisition System: ............................................................................... 51 2.1.8 Arterial Blood Gases Analysis: ...................................................................................... 52 2.1.9 Adrenocorticotropin Hormone Assay: ............................................................................ 52 2.1.10 Cortisol Assay: ........................................................................................................... 53 2.1.11 FX and NFX assay: .................................................................................................... 53 2.1.12 Statistical Analysis: .................................................................................................... 54
3. Postnatal Lamb Development and Sex Differences ....................................................... 55 3.1 Introduction: ........................................................................................................................ 55 3.2 Methods: ............................................................................................................................. 57
3.2.1 Experimental Protocol: ................................................................................................... 57 3.2.2 Statistical Analysis: ........................................................................................................ 57
3.3 Results: ................................................................................................................................ 58 3.3.1 The Development Of Physiological Variables And Sex Differences In Development From The Newborn Period Up To 1 Year Of Age: ..................................................................... 58 3.3.2 Neonatal Behavioral Development: ................................................................................ 72 3.3.3 The Development Of Circadian And Ultradian Rhythms. The Ultradian 1 h Rhythm Is Associated With Suckling: .......................................................................................................... 74 3.3.4 Weaning And Changes In Physiological Variables: ....................................................... 80
3.4 Discussion: .......................................................................................................................... 83
4. Acute Fluoxetine Effects In The Newborn...................................................................... 99 4.1 Introduction: ........................................................................................................................ 99 4.2 Methods: ........................................................................................................................... 102
4.2.1 Experimental Protocol: ................................................................................................. 102 4.2.2 Statistical Analysis: ...................................................................................................... 102
4.3 Results: .............................................................................................................................. 103 4.3.1 Plasma FX Concentration And Weight Gain: ............................................................... 103 4.3.2 Rest Activity Cycles: .................................................................................................... 105 4.3.3 Cardiovascular Variables (Heart Rate, Arterial Pressure, Heart Rate Variability): ...... 105 4.3.4 Arterial Blood Gases: ................................................................................................... 109 4.3.5 Plasma Cortisol Concentration: .................................................................................... 112
4.4 Discussion: ........................................................................................................................ 113 4.4.1 Acute FX Effects On Cardiovascular –Respiratory Functions: .................................... 115 4.4.2 Acute FX Effects On HPA Axis Functions: ................................................................. 118 4.4.3 Underlying Mechanisms Of Poor Neonatal Adaptation: .............................................. 119
5. Acute Fluoxetine Effects In The Postnatal Lambs (From 10 Days To 1 Year Of Age)............................................................................................................................................... 123
5.1 Introduction: ...................................................................................................................... 123 5.2 Methods: ........................................................................................................................... 125 5.3 Results: .............................................................................................................................. 127
5.3.1 Acute FX Effects On Cardiovascular-Respiratory Functions: ...................................... 127 5.3.2 Acute FX Effects On Homeostatic And Respiratory Functions: .................................. 131 5.3.3 Acute FX Effects On Behavior: .................................................................................... 141
5.3.3.1 Behavioral Activities In The Lambs At 1 Month Of Age: ................................... 141

viii
5.3.3.2 Behavioral Activities In The Lambs At 6 Months Of Age: ................................. 148 5.3.3.3 Acute FX Effects On Cortisol Level: ................................................................... 153
5.4 Discussion: ........................................................................................................................ 156
6. Postnatal Outcomes in Lambs Exposed Antenatally to FX ........................................ 167 6.1 Introduction: ...................................................................................................................... 167 6.2 Methods: ........................................................................................................................... 169
6.2.1 Animal And Catheter Implantation: ............................................................................. 169 6.2.2 Experimental Protocol: ................................................................................................. 170 6.2.3 Digital Video Recording And Behavioral Evaluation: ................................................. 171 6.2.4 Actiwatch Rest/Activity Monitoring: ........................................................................... 172 6.2.5 Powerlab Data Acquisition System: ............................................................................. 172 6.2.6 Pulse Oximeter: Tissue SpO2 And Heart Rate:............................................................. 172 6.2.7 FX And NFX Concentration: ........................................................................................ 173 6.2.8 Statistical Analysis: ...................................................................................................... 173
6.3 Results: .............................................................................................................................. 174 6.4 Discussion: ........................................................................................................................ 187
7. Summary and Conclusions............................................................................................. 194
REFERENCES .................................................................................................................... 215
Appendices ........................................................................................................................... 252 Appendix 1: Actiwatch Variables Data In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age: ................................................................................................................................................ 252 Appendix 2: Heart Rate, Arterial Pressure And HRV In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection: ............................................................................................... 255 Appendix 3: Arterial Blood Gases And Body Temperature In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection: .................................................................................. 258 Appendix 4: Glucose, Lactate Concentration And Electrolite In The Lambs At~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection: .................................................................................. 261

ix
List of Tables
Table 1.1: DSM-IV-TR criteria for major depressive episode ............................................... 10
Table 2.1: Time points at which blood samples were collected for arterial blood gases, FX, ACTH and cortisol measurement. SW = sterile water. .......................................................... 48
Table 3.1: The time window of weaning in lambs was defined as when lambs started to first consume solid food to when 1h rhythm due to suckling disappeared .................................... 82
Table 3.2: Break points of cardiovascular, metabolic, endocrine and renal variables. ........... 82
Table 6.1: Gestational age and birth weight in the control and antenatally FX-exposed lambs............................................................................................................................................... 175
Table 6.2: Mean ± SEM of HRV values for the first 30 mins of the experiment in the lambs on day 2 and the changes from day 5, 10, 14 compared with the values on day 2 ............... 186
Table 7.1: Outcomes of chronic antenatal and acute postnatal FX exposure in fetal and postnatal lambs ...................................................................................................................... 206
Table 7.2: Outcomes of antenatal SSRI exposure in human (exposure for most or all of pregnancy) ............................................................................................................................. 207
Table 7.3: Postnatal outcomes of antenatal and postnatal SSRI exposure in rodents ........... 212

x
List of Figures
FIG. 1.1: Biosynthesis and metabolism of serotonin. ............................................................. 15
FIG. 1.2: Schematic diagram of monoaminergic synapse and steps involved in synaptic transmission: Monoamine neurotransmitters (1) are synthesized within neurons from common amino acid precursors, which are Tryptophan and 5-Hydroxytryptophan (5-HT) in the case of serotonin, (2) are taken up into synaptic vesicles via a vesicular monoamine transporter, (3) are released into the synaptic cleft upon stimulation, (4) interact with postsynaptic receptors to alter the excitability of postsynaptic cells, (5) interact with presynaptic autoreceptors located on the nerve terminal to suppress further release. (6) Released monoamines may also be taken back up from the synaptic cleft into the nerve terminal by plasma membrane transporter proteins. (7) Once taken up, monoamine may be subject to enzymatic degradation or may be protected from degradation by uptake into vesicles. MAO: monoamine oxidase ...................................................................................... 22
FIG. 3.1: Postnatal lamb arterial pressure (A) and heart rate (B) in all (open bar), male (black bar) and female (gray bar) lambs at 3 days (n = 14), 10 days (n = 18), 1 month (n = 15), 3 months (n = 17), 6 months (n = 11) and 1 year (n = 10) of age. ............................................. 60
FIG. 3.2: Time domain of heart rate variability: SDNN (standard deviation of NN interval) (A), SD Delta NN (Standard Deviation of the differences between adjacent NN intervals) (B), RMSSD (Square root of the mean of the squared differences between adjacent NN intervals) (C) in lambs at 3 days (n = 11), 10 days (n = 18), 1 month (n = 15), 3 months (n = 17), 6 months ( n = 11) and 1 year ( n = 10) of age. ............................................................... 61
FIG. 3.3: Frequency domain of heart rate variability: LF (Low Frequency) power (A), HF (High Frequency) power (B), LF/HF ratio (Low Frequency/High Frequency Ratio) (C) in lambs at 3 days (n = 11, male n = 4, female n = 7 ); 10 days (n= 18, male n = 9, female n = 9), 1 month (n= 15, male n = 8, female n = 7); 3 months (n= 17, male n = 9, female n = 8), 6 months (n = 11, male n = 6, female n = 5), 1 year (n = 10, male n = 6, female n = 4) of age. 62
FIG. 3.4: The relationship between SDNN (msec) and heart rate (bpm) in the lambs from 3 day to 1 year of age ................................................................................................................. 63
FIG. 3.5: Mean pH value in total (open bar), male (black bar) and female (gray bar) lambs at 3 days (n = 13, male n = 6, female n = 7 ); 10 days (n= 17, male n = 8, female n = 9), 1 month (n= 15, male n = 8, female n = 7); 3 months (n= 17, male n = 9, female n = 8), 6 months (n = 10, male n = 5, female n = 5), 1 year (n = 10, male n = 6, female n = 4) of age. 65
FIG. 3.6: Mean pCO2 (A), pO2 (B) and O2 sat (C) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. ....... 66
FIG. 3.7: Mean HCO3 (A), Base Excess (B) and Hemoglobin (C) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. .......................................................................................................................................... 67

xi
FIG. 3.8: Mean Glucose (A) and Lactate (B) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. ................. 68
FIG. 3.9: Mean urine flow rate in lambs at 3 days (n = 10); 10 days (n= 10), 1 month (n= 9); 3 months (n= 12); 6 months (n = 7) and 1 year (n = 10) of age. ............................................. 69
FIG. 3.10: Mean cortisol concentration in fetal lambs from preinfusion to infusion day 2 and from infusion day 5 to 8 adapted from Morrison et al, 2004 and in postnatal lambs prior surgery (n = 11), at 3 days (n = 14); 10 days (n= 12), 1 month (n= 12); 3 months (n= 12); 6 months (n = 8) and 1 year (n = 8) of age. ............................................................................... 70
FIG. 3.11: Mean ACTH concentration in fetal lambs from preinfusion to infusion day 2 and from infusion day 5 to 8 adapted from Morrison et al, 2004 and in postnatal lambs prior surgery (n = 11), at 3 days (n = 2); 10 days (n= 2), 1 month (n= 2); 3 months (n= 14); 6 months (n = 3) and 1 year (n = 4) of age. ............................................................................... 71
FIG.3.12: Cortisol concentration during the day in the lambs at 10 days, 1 month and 1 year of age. ...................................................................................................................................... 73
FIG. 3.13: Weight gain (A) and total suckling time (B) in singleton (n = 6), twin (n =14 for weight gain and n = 10 for total suckling time) and triplets (n = 3) in lambs on postnatal day 2-3 (N = 22). ........................................................................................................................... 76
FIG. 3.14: Plots of daily total activity score (a), activity bouts/day (b), mean score in active period (c), mean episode duration (d), % time moving (e) and % time immobile (f) against postnatal age in lambs. (n=21) ................................................................................................ 77
FIG. 3.15: (A) The average development of 24, 8, 6 and 1-2 hour activity-rest rhythms. The lines with diamond, square, triangle and x symbols indicate the 24h circadian, the 8 hour, the 6 hour and 1 hour ultradian rhythms, respectively. Levels of rhythm dominance of 1 to 4 indicate first to fourth most dominant rhythms. A level of rhythm dominance of greater than 4 indicates that this rhythm is not one of the top four rhythms (n=22). (B) The daily mean total activity score. (n= 21) ..................................................................................................... 78
FIG. 3.16: (A). Relationship between the day that the 1h rhythm disappears and the day of the lowest total activity (n = 21). (B) Relationship between the day that the 6h rhythm disappears and the first day lamb eating hay (n = 21) ............................................................ 79
FIG. 3.17: Relationship between suckling frequency on postnatal day 2-3 from DVR observation and calculated from the actiwatch FFT (n = 21) ................................................. 80
FIG. 4.1: (A) Plasma FX concentration verus time curve in the 3 day-old lambs (closed symbols and solid line) and in 13 individual human infants (open symbols) sampled at postnatal day 2 following in utero exposure to the drug. Human data was taken from the dissertation of Dr. John Kim, 2000 and the lambs data was taken from the dissertation of Dr. Tim Chow, 2013. (B) Weight gain on ~ postnatal day 4 among the normal term lambs (n = 11, open bar), control group (n = 9, black bar), FX group (n = 12, gray bar) and post FX day 1 (n = 12, light gray bar) ....................................................................................................... 104

xii
FIG. 4.2: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in the normal ~ 4 day-old term lambs (n = 10), control = on day of control experiment (n=6), and FX lambs on the day of FX experiment (n = 11) followed up to post FX day 1 (n = 11), post FX day 2 (n = 11) and post FX day 3 (n = 11). No significant changes were found between activities after FX administration and sterile water injection or normal day without experiments. ................... 106
FIG. 4.3: Arterial pressure and Heart rate (at 1 min intervals for the first 5 min after dosing and at 5 minute interval for the whole 2.5h of experiment) in the ~ 4 days old lamb with sterile water injection (control group) and FX injection (FX group). ................................... 107
FIG. 4.4: Time domain of HRV (SDNN, SD Delta NN, RMSSD) and frequency domain (LF power, HF power, LF/HF ratio) at 30 min intervals in the newborn lambs at ~ 4 days old in the control group (open diamond and dotted line) and FX group (close circle and solid line)............................................................................................................................................... 108
FIG 4.5: The relationship between changes in heart rate at 2 mins following FX (delta HR) and changes in time domain of HRV (SDNN, SD Delta NN, RMSSD) (i.e Delta SDNN, Delta SD Delta NN, Delta RMSSD) at 1 min following FX in the lambs at ~ 4 days of age................................................................................................................................................ 110
FIG. 4.6: Arterial blood gases (pH, O2 saturation, pO2, pCO2, HCO3, Base Excess) in the newborn lambs (~ 4 days old) in the control group (open diamond and dotted line, n = 9) and FX group (close circle and solid line, n = 11) ...................................................................... 111
FIG. 4.7: Glucose, Lactate and Hb concentration in the newborn lambs (~ 4 days old) in the control group (open diamond and dotted line, n = 9) and FX group (close circle and solid line, n = 11) ........................................................................................................................... 112
FIG. 4.8: Plasma cortisol concentration in the ~ 4 days old lambs in the control group with sterile water injection (n = 8, open bar) and the FX group with FX injection (n = 5, close bar) at 30 and 15 minutes before injection and 0.5, 1, 2, 6, 12 hours after injection ................... 113
FIG 5.1: Arterial pressure and heart rate changes in 1 year old lambs with acute FX administration ....................................................................................................................... 128
FIG 5.2: HRV (SDNN, SD Delta NN, RMSSD, LF power, HF power, LF/HF ratio) changes with acute FX administration in 1 year old lambs. ............................................................... 130
FIG 5.3: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 1 month of age. ....................................................................................................... 132
FIG 5.4: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 6 months of age. ..................................................................................................... 133
FIG 5.5: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 1 year of age. .......................................................................................................... 136

xiii
FIG 5.6: (A) The relationship between changes in pO2 at 5 mins and changes in arterial pressure (AP) at 2 mins following FX injection in all lambs at 1 year of age (square symbol, solid line, n = 8); (B) The relationship between changes in pO2 at 5 mins and changes in AP at 2 mins following FX injection in male (square symbols, dotted line, n = 4) and female (circle symbols, solid line, n = 4) at 1 year of age and (C) The relationship between changes in O2 saturation at 5 mins and changes in AP at 2 mins following FX injection in male (square symbols, dotted line, n = 4) and female (circle symbols, solid line, n = 4) at 1 year of age. ........................................................................................................................................ 137
FIG 5.7: (A) The relationship between changes in pO2 at 5 mins and log FX concentration at 5mins following FX injection in all lambs at 1 year of age (square symbol, solid line, n = 8) (B) The relationship between changes in pO2 at 5 mins and log FX concentration at 5mins following FX injection in male (triangle symbols, dotted line, n= 4) and female (circle symbols, solid line, n = 4) lambs at 1 year of age; and (C) The relationship between changes in O2 saturation at 5 min and log FX concentration at 5mins following FX injection in male (triangle symbols, dotted line, n= 4) and female (circle symbol, solid line, n = 4) lambs at 1 year of age. ............................................................................................................................ 138
FIG 5.8: PCO2 , base excess (BE), HCO3 concentrations with sterile water and FX administration in male and female lambs at 1 year of age. ................................................... 139
FIG 5.9: Glucose, lactate, Hb (Hemoglobulin) concentrations with sterile water and FX administration in male and female lambs at 1 year of age. ................................................... 140
FIG. 5.10: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in 1 month-old lambs with the average of the 4 normal days in the pen before the experiments (n = 12, open bar), on control experiment day (n=9, black bar), and on FX experiment day (n = 12, dark gray), post FX day 1 (n = 11, light gray), post FX day 2 (n = 11) and post FX day 3 (n = 11). ......................... 144
FIG. 5.11: (A) Percentage of time lambs spent for each activity from DVR observation and (B) the intensity of each activity when matching DVR observation with actiwatch score data in lambs at 1 month of age. Normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). ................................................................................................. 145
FIG. 5.12: (A) Percentage of time lambs spent for standing activity from DVR observation and (B) the intensity of standing quiet and standing moving when matching DVR observation with actiwatch score data in lambs at 1 month of age. Normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar)........................................................................... 146
FIG 5.13: Weight gain (mean ± SE) in the lambs during their normal days, control experiment with distilled water injection, FX experiment with FX injection and 3 continuous days after the FX experiment day around 1 month of age. ................................................... 147
FIG. 5.14: Intensity of standing and playing activity from DVR observation in the 1 month-old male and female lambs on normal day (day lambs in the pen without any experiments,

xiv
gray bar, n = 3), on Control experiment day (n=6, open bar) and FX experiment day (n=6, black bar). ............................................................................................................................. 147
FIG. 5.15: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in 6 month-old lambs with average of 4 normal days in the pen before the experiments (n = 8, open bar), on control experiment day (n=8, black bar), and on FX experiment day (n = 8, dark gray bar), post FX day 1 (n = 8), post FX day 2 (n = 8) and post FX day 3 (n = 8). ................................................................. 150
FIG. 5.16: (A) Percentage of time lambs spent for each activity from DVR observation and (B) the intensity of each activity when matching DVR observation with actiwatch score data in lambs at 6 month of age. The normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). ................................................................................................. 151
FIG. 5.17: (A) Percentage of time lambs spent on standing (quiet and moving) activity from DVR observation and (B) the intensity of standing quiet and standing moving when matching DVR observation with actiwatch score data in lambs at 6 month of age. The normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). ....................... 152
FIG 5.18: Weight gain (mean ± SE) in the lambs during their normal days, control experiment with distilled water injection, FX experiment with FX injection and 2 continuous days after the FX experiment day at around 6 months of age. ............................................. 153
FIG. 5.19: Cortisol and ACTH concentration in the lambs at 3 months of age at 15, 30 minutes before and 0.5, 1, 2, 6 and 12 h after sterile water injection (control group) and FX injection ( FX group) ............................................................................................................ 154
FIG. 5.20: Cortisol and ACTH concentration in the lambs at 1 year of age at 15, 30 minutes before and 0.5, 1, 2, 6 and 12 h after sterile water injection (control group) and FX injection (FX group) ............................................................................................................................ 155
FIG. 6.1: (A) Daily weight in the control (n=7) and prenatal FX-exposed lambs (n=7) from postnatal day 1 (day of birth) to postnatal day 14. ................................................................ 175
FIG.6.2: FX, (R)-FX, (S)-FX and NFX, (R-)NFX, (S)-NFX concentrations in the prenatally FX-exposed ewes (n= 5) and lambs (n=7). Samples were taken at birth in both ewes and lambs and on postnatal day 2,5,10 and 14 only in lambs. .................................................... 177
FIG 6.3: The relationship between the lamb:ewe plasma FX concentration ratio and the time from the last FX dose (hours) in the ewe. ............................................................................. 178
FIG. 6.4: Daily (A) tissue spO2 and (B) heart rate measured by pulse oximeter in the control (n=2) and prenatal FX-exposed lambs (n =7) from postnatal day 1 (day of birth) to postnatal day 14. ................................................................................................................................... 178

xv
FIG. 6.5: Neonatal behavior (first time to stand, walk, suck immediately after birth) in the control lambs (n = 37) and prenatal FX-exposed lambs (n =7). ........................................... 181
FIG. 6.6: Daily activity variables (active bouts per days, total activity score, mean score in active, episode duration, % time mobile, % time immobile) obtained from the actiwatches in the control (opened, diamond symbols) and prenatal FX-exposed lambs (closed, circle symbols) from postnatal day 2 (the first full day) to postnatal day 14. ................................ 183
FIG. 6.7: Daily sleep, suckling and non-suckling activity bouts obtained from DVR observations in the control (open, diamond symbols) and prenatal FX-exposed lambs (closed, circle symbols) from postnatal day 2 (the first full day) to postnatal day 10. ...................... 184
FIG. 6.8: HRV (time domain: SDNN, SD Delta NN, RMSSD; frequency domain: total power, LF power, HF power, LF/HF) on postnatal day 2 in control group (diamond symbol, dotted line, n = 8) and prenatal FX-exposed lambs (circle symbol, solid line, n = 7). ......... 185

xvi
List of Symbols and Abbreviations 5-HIAA 5-hydroxyindoleacetic acid 5-HTP 5-hydroxy-tryptophan 5-HT 5-hydroxy-tryptamine/ Serotonin ACTH Adrenocorticotrophin ANS Autonomic Nervous System AUC Area Under the Curve BE Base Excess CBT Cognitive-Behavioral Therapy CCM Centre of Comparative Medicine CRH Corticotropin-Releasing Hormone CYP Cytochrome P450 DVR Digital Video Recording FDA Food and Drug Administration FFA Free Fatty Acid FFT Fast Fourier Transformation FX Fluoxetine GA Gestational Age GFR Glomerular Filtration Rate GH Growth Hormone GLM General Linear Model HF power High Frequency power HPA Hypothalamic-Pituitary-Adrenal HRV Heart Rate Variability IGF-1 Insulin-like Growth Factor-1 LF power Low Frequency power LF/HF ratio Low Frequency/High Frequency power ratio MAOI Monoamine Oxidase Inhibitors MCA Middle Cerebral Artery MDD Major Depressive Disorder msec Millisecond NaSSA Noradrenergic and Specific Serotonergic Antidepressant NDRI Norepinephrine/Dopamine Reuptake Inhibitor NFX Norfluoxetine nu normalized unit OCD Obsessive Compulsive Disorder PMSG Pregnant Mare Serum Gonadotropin PND Postnatal Day PPHN Persistent Pulmonary Hypertension RCT Randomized Controlled Trial REM Rapid Eye Movement RMSSD Square root of the mean of the squared differences between adjacent
NN intervals. SARI Serotonin Antagonist Reuptake Inhibitor

xvii
SD Delta NN Standard Deviation of the differences between adjacent NN intervals SDNN Standard Deviation of the NN intervals SEM Standard Error of Mean SNRIs Serotonin-Norepinephrine Reuptake Inhibitors sO2 Oxygen saturation SSRIs Selective Serotonin Reuptake Inhibitors TCA Tricyclic Antidepressant/Tricyclic Acids VFA Volatile Fatty Acid WHO World Health Organization

xviii
Acknowledgements
I would like to express my deepest gratitude to my supervisor and mentor, Dr.Dan
Rurak for his guidance, dedication, and support over the course of my PhD study. I would
also like to thank my supervisory committee: Drs.Wayne Riggs, Tim Oberlander, Ken Lim
and Anthony Perks for their encouragement and valuable advice for my project; and my
special thanks to Tim Chow for his invaluable assistance in the lab experiments. I would also
appreciate Dr. Bev Chua for her friendship and her dedicated care toward the sheep. Thanks
to all the staffs in the Child and Family Research Institute (CFRI) and Centre of Comparative
Medicine (CCM): Catherine, Edwards, Leika, Claire, Gordon and Kelly for their animal care
knowledge. I would also like to thank Roshni Nair for her continuous support and
compassion towards me during my entire graduate program. Thanks to my friends in the
Reproductive and Developmental Sciences program for their positive engagement.
My warmest thanks to Rev. Joshep Nguyen; all seniors from the St. Andrew’ parish
as well as my friends for their help and making my life more functional outside of the school.
Most of all, I would like to sincerely thank my brother, Sean Nguyen, whom I am
most grateful for his sacrifice helping out with the family needs allowing me to focus on my
study. Above all I want to thank my mom and late dad for teaching me the value of
education. Without your unconditional and steadfast love, I cannot be who I am today.
A final thanks to all the funding sources for the project and my studentship (CIHR,
IWRH, CFRI).

xix
Dedication
To the memories of my father
To my beloved mom and brother

1
1. Introduction
1.1 Postnatal Lamb Development:
1.1.1 Cardiovascular Function:
Birth is an event which involves remarkable changes in the environment surrounding
the fetus. Moving from a comfortable warm environment, filled with fluid and maternal soft
tissue as a cushion, and readily available nutrients and oxygen supply via the placenta in
utero to an extra utero life, the newborn for the first time is exposed to cold challenge,
gravity, air, and hard surfaces. In order for a successful transition to be accomplished, it is
required that the newborn undergo a series of physiological changes (such as air breathing,
shift in cardiovascular patterns, thermoregulation, alteration of oxygen transport (Lister et al.,
1984) and behavioral adaptations (e.g: becoming aroused and aware of the surrounding
environment, being responsive and interactive in order to start establishing bonding with the
mother, and initiating suckling (Mellor & Gregory, 2003; Mellor, 1988).
After birth, the high relative cardiac output observed in the fetus is still maintained in
the newborn lambs up to 1 month of age (Minoura & Gilbert, 1987). One of the reasons
explained for the high cardiac output in the newborn lambs is that more O2 needs to be
transported to the tissues since O2 extraction is limited by the avid binding of O2 to
hemoglobin (Lister et al, 1979). As there is an increasing oxygen consumption in the
newborn lamb in response to cold exposure after birth (Alexander & Williams, 1968; Sidi et
al., 1983), it is vital for them to maintain a high cardiac output. The subsequent decline in
heart rate over the first 2 months of life is proportionate to that of cardiac output to keep the
stroke volume relatively constant in relation to the body weight (Lister et al., 1979). In

2
lambs, cardiac sympathetic innervations and parasympathetic influences on the heart are
incomplete at birth. However, the sympathetic nervous system appears to be predominant in
the lamb less than 1 week of age whereas parasympathetic becomes dominant in lambs older
than 3 months. It is believed that the sympathetic activity and circulating catecholamines help
to maintain such high cardiac output in 1-3 day old lambs (Minoura & Gilbert, 1987). Siimes
et al have reported that the beat-to-beat heart rate variability (HRV) increased in lambs after
3 weeks of age (Siimes et al., 1984). This increasing trend of HRV with advancing age is
similar to that observed in human infants (Harper et al., 1976; Wheeler et al., 1979). In
addition, HRV varies in relation to sleep state. It is highest during waking and active sleep
and lowest during quiet sleep (Siimes et al., 1984)
1.1.2 Homeostasis And Metabolism:
Since nutrients are ingested orally after birth, before the newborns are able to nurse
from their mother and before maternal milk production increases, they have to use their own
energy reserves to produce heat against the cold stress. In newborn lambs, both shivering
(muscle glycogenolysis) and non-shivering metabolism (or brown adipose tissue
thermogenesis) are equally important in cold-induced thermogenesis. In brown adipose tissue
thermogenesis, the brown adipocyte was stimulated by noradrenaline via ß-adrenergic
receptors. This gives rise to a series of actions with a final release of free fatty acid (FFA)
within the cell. Even though these FFA might be released to the circulation, a significant
amount of these FFA remain in the cell and is transferred to the mitochondria where they are
oxidized to produce heat via the electron transport change and action of brown adipose
specific uncoupling protein (Symonds et al., 2012). With advancing age, the non-shivering
contribution declines rapidly while shivering metabolism increases (Alexander & Williams,

3
1968; Alexander et al., 1968, Symonds et al., 2012). However, lambs exposed to cold
showed a much greater increase in free fatty acid concentration in those older than 20 days
than those at 1-3 days old (Alexander et al., 1968). As the authors explained, this is because
the proportion of brown adipose tissue, which can re-utilize fatty acids from triglyceride
lipolysis, decreases while white adipose tissue increases over age, thus during cold exposure
in the older lambs, the rate of lipolysis exceeds the rate of FFA recycling within the adipose
cell (Alexander et al., 1968).
In the transition from prenatal life, in which nutrient supply and growth are
increasingly constrained by the placenta, to postnatal life with major changes in the quantity
and composition of nutrient supply, there is a requirement for metabolic adaptations. This
includes colonization of the digestive system, which begins soon after birth (Morrison et al.,
2009). The composition of nutrients in utero is primarily glucose and amino acids whereas a
diet after birth may contain a high level of fat and less carbohydrate (Girard et al., 1997;
Greenwood et al., 2002). In sheep, with the dietary switch from milk-fed to solid feed at
weaning, the morphology and metabolic aspects of the ruminant digestive system also
change accordingly. The rumen increases in size, the intraruminal papillae increase in length
and the epithelium lining the rumen becomes keratinized (Lake et al., 1982; Lane, Baldwin,
& Jesse, 2000; Lavker, Chalupa, & Opliger, 1969). In the presence of solid feed, ruminal
microorganisms produce volatile fatty acids (VFA) via fermentation of the feed. VFA
exposure appears to be necessary to stimulate rumen metabolic development. Examining
rumen development in lambs with and without solid feed in their diet, Lane et al have
concluded that the presence of solid food is needed for rumen metabolic maturation (Lane et
al., 2000).

4
In a study of blood composition in normal and growth retarded lambs over the first 24
hour after birth, Mellor & Pearson (1977) found that plasma glucose and IgG increase a few
hours after birth, which is most likely attributed to the absorption of colostrum from the gut.
In contrast, lactate concentration decreases sharply after birth. Plasma fructose concentration
was decreased progressively in late gestation fetal lambs till birth and had disappeared from
the lamb’s plasma by the first day of birth (Comline & Silver, 1970).
A recent study in our lab has found that arterial Po2, pH, BE, So2 decrease and Pco2
and hemoglobin concentration increase in the fetal lambs with advancing gestational age
(Rurak & Bessette, 2012). However, to date there is limited information on the changes in
arterial blood gas values of newborn and postnatal lambs in response to cardiovascular-
respiratory development as well as other physiological processes.
1.1.3 Behavior: Maternal-Infant Bonding:
Different from rodents, which are altricial at birth, lambs are precocial and born at an
advanced stage of physical development. In addition, all sense organs (tactile, olfactory,
auditory and visual) of the lamb are fully-developed and ready to function from birth (Fraser
& Broom, 1997). This allows mutual recognition between ewes and lambs, which happens
very early following birth, and in which visual appearance of the lamb appears to be an
important cue (Walser & Alexander, 1980). After the first two hours of parturition, cross
fostering lambs is no longer possible, even when the alien lambs were masked by their foster
mother’s amniotic fluid (Walser & Alexander, 1980). Indeed, maternal-infant bonding is first
established immediately after delivery by a coordinated expression of behaviors between
both the ewe and lambs. Together with the ewe’s behavior at birth such as licking or

5
grooming, rumbles (frequent low-pitched bleats made by the ewes as she licks the lamb dry)
and udder acceptance, it is also vital for the lamb to successfully accomplish a sequence of
behaviors directed toward standing, finding the udder and suckling during this immediate
postpartum period (Fraser & Broom, 1997). These behaviors from the ewe serve not only for
the formation of a strong maternal-infant attachment with her own lamb but also as an
expression of nurturing behavior to facilitate the successful transition from the pre to
postnatal life for the lambs (Dwyer, 2008a). Normally, the ewe licks to dry the lamb, to clear
placental membranes from the lamb’s nose and mouth, to stimulate activity and respiration
and to form an olfactory memory for the lamb. In addition, a rumbling sound made by the
ewe when she licks the lamb may help to reduce stress in the lamb (Dwyer et al., 1998) and
form the auditory memory for later maternal recognition (Walser & Alexander, 1980). These
sound cues also strongly influence the lamb’s behavior. It helps the lambs to orientate to the
ewe and keep them near to the ewe, especially in the dark or when mingling with other ewes
and lambs. At the same time, in response to maternal care, lambs perform a series of
behaviors after birth initiated by shaking the head, rolling onto the sternum, bleating, pushing
up on the knees and then attempting to stand (Dwyer, 2003; Dwyer, 2008a; Fraser & Broom,
1997). Most newborn lambs are able to stand within the first half hour following birth and
sucking occurs within the first 2 hours of birth in normal cases (Dwyer, 2003). Studies have
shown that the rate of lamb survival is higher in lambs that stand and suckle quickly after
birth (Dwyer et al., 2005). However, the lamb’s first attempts to suck are usually
unsuccessful. There are a number of known factors affecting the behavior in the lamb
including breed, sex, litter size, prenatal nutrition, placental insufficiency, the ewe’s parity
and the birth process (Dwyer, 2003; Dwyer et al., 2005). Male lambs tend to be slower than

6
females to rise on their knees and to attempt to suck, whereas triplets are slower than twin or
single lambs to suck (Dwyer, 2003; Dwyer et al., 2005). Likewise, using an actigraphy
method, a previous study in our lab also found that from postnatal day 2 to 4 male lambs
seem to be less active than females but the situation was reversed from day 5 to 9 (Rurak et
al., 2008). Bonding is also developed between twins or triplet lambs. They seem to stay
closer to each other than to any alien lambs. Such attachment is established through visual
cues and voice recognition (Fraser & Broom, 1997).
1.1.4 Hypothalamic-Pituitary-Adrenal (HPA) Axis Function:
The fetal HPA axis plays an essential role in fetal development, maturation,
homeostasis and even neonatal survival. Studies have shown that the timing of HPA axis
maturation is species-specific and depends on the timing of the brain growth spurt. In
precocious animals such as sheep, the brain growth spurt, which is associated with a large
proportion of neuroendocrine maturation (including corticosteroid receptor development),
takes place in utero (Dobbing & Sands, 1979; Matthews et al., 2002).
It had been well established that the plasma concentration of cortisol, a final product
of the HPA axis, increases markedly during the last 5-10 days of pregnancy and can actively
initiate the onset of parturition by changes in the progesterone/estrogen ratio (Challis et al.,
2000; Mellor, Matheson, & Small, 1977; Norman et al.,1985; Thorburn & Challis, 1979).
Therefore, fetal hypophysectomy or adrenalectomy prolong pregnancy (Liggins, Holm, &
Kennedy, 1966), whereas infusions of adrenocorticotrophin (ACTH) or glucocorticoids can
cause premature delivery (Liggins, 1968; Thorburn & Challis, 1979). The prepartum surge of

7
cortisol is considered as a preparation for the fetus to face with stressful events at birth and
enable them to adapt to extrauterine life (Mastorakos & Ilias, 2003).
Postnatally, plasma cortisol concentration decreases markedly during the first 1-2
hours after birth and continues to decline for 24h thereafter (Mellor & Pearson, 1977). The
circadian rhythm of plasma cortisol does not appear until the third week of age in sheep. This
appearance of a 24 h rhythm of cortisol is not influenced by the presence of a light: dark
cycle since it appears at the same time in newborn lambs raised both under 12:12 hour light:
dark or constant light condition (Parraguez et al., 1989).
Sex differences in the HPA axis function has been observed and attributed to the
differences in the levels of gonadal hormones such as estrogens, progesterone, testosterone
(Carey et al., 1995; Viau & Meaney, 1991; Young, 1995; Young, 1996). This could
contribute, at least in part, to the higher prevalence of depression in females than in adult
males (Brummelte & Galea, 2009). In rats, estrogen enhances HPA axis responses (Viau &
Meaney, 1991), whereas androgen depresses HPA axis activity (Viau, 2002). In sheep, sex
differences in cortisol response only depend on the type of acute stressors but not the gonadal
status (Turner et al., 2002).
1.1.5 Renal Function:
Previous studies have shown that renal function changes from the fetal to the
newborn period in lambs, which indicates the rapid adjustment of the kidney to the changes
in fluid and electrolyte supply. The glomerular filtration rate (GFR) increases from 4.59 ±
0.27ml/min to 6.94 ± 1.0 ml/min, whereas urine flow, sodium and osmolar excretion
decreased and urinary osmolarity increased (more concentrated urine) in newborn lambs 24 h

8
following birth compared to fetal lambs (Smith & Lumbers, 1989). Renal adaptations in the
newborn lambs appear to be independent of their gestational age (Berry et al., 1995). The
mechanisms responsible for the increase in GFR in the postnatal lambs include an elevated
net filtration pressure and ultrafiltration coefficient (Turner et al, 2008). In fetal sheep, acute
stress due to surgery can reduce urine flow (Gresham et al, 1972), which can also be the case
with birth events. Given that urine flow is influenced by water and fluid intake, GFR and
sodium reabsorption, dietary changes from milk-fed to solid intake at the time of weaning are
hypothesized to cause a decline in urine flow rate.
1.2 Depression: Symptoms And Diagnosis:
Depression is often referred to as being one specific form of a mental health problem;
however there are actually several categories of depressive illnesses. The most common form
of depression is Major Depression or Major Depressive Disorder. This form of depression
comprises a combination of symptoms that interferes with one’s ability to work, study, sleep,
eat and enjoy activities that are pleasurable. A less severe form of depression is dysthymia, in
which the long-lasting symptoms do not seriously disable one, but keep one from functioning
well or feeling good. And another form of depressive illness is Bipolar Disorder, which is
characterized by cycling mood changes: high (manic episodes) and low (depressive episodes)
(Dubovsky et al., 2003).
The persistence of symptoms is a key to diagnosis. Many of us might feel blue or
down several times in our life, especially, for example when we lose a loved one. Sometimes
we may even have passing thoughts of suicide. However, in order to differentiate between
real depression and these more commonplace situations, we need to consider the time frame

9
of those feelings or symptoms. Based on the DSM-IV, diagnosis for major depression
disorder is made if symptoms are present for at least 2-week duration (table 1, criteria for
diagnosis of depression, DSM-IV). Conversely, dysthymia is considered as chronic
depression in which the symptoms must persist for at least 2 years to meet the diagnostic
criteria.
Assessing depression in children and youth is different from adults because the
symptoms may be atypical in these populations; for example, depressed mood in children and
adolescents can present as an irritable mood, or the weight loss criteria queried in adults can
be reconsidered as a failure to make the expected weight gain in children. In dysthymic
disorder, the time period for diagnosis is also shorter for children, at least one year (instead of
2 years in adults).

10
Table 1.1: DSM-IV-TR criteria for major depressive episode
A. Patients must have experienced at least five of the nine symptoms below, for the
same 2 weeks or more, for most of the time almost every day. One of the symptoms must
be either (a) depressed mood, or (b) loss of interest.
a. Depressed mood
b. A significantly reduced level of interest or pleasure in most or all activities.
c. A considerable loss or gain of weight (5% or more change of weight in a
month when not dieting). This may also be an increase or decrease in appetite
d. Difficulty falling or staying asleep (insomnia), or sleeping more than usual
(hypersomnia)
e. Behaviour that is agitated or slowed down.
f. Feeling fatigued, or diminished energy
g. Thoughts of worthlessness or extreme guilt (not about being ill)
h. Ability to think, concentrate, or make decisions is reduced
i. Frequent thoughts of death or suicide (with or without a specific plan), or
attempt of suicide
Adapted from Quick reference to the Diagnostic criteria from DSM-IV-TR, American
Psychiatric Association (Mood disorders. 2000).
1.3 Pathophysiology Of Depression:
Many theories have been proposed for the pathophysiology of affective illness. In
1965, extrapolating the indirect evidence from the pharmacological studies, Schildkraut

11
proposed the catecholamine hypothesis of affective disorders. This hypothesis states that
depression is associated with a decrease in catecholamines, particularly norepinephrine,
available at central adrenergic receptor sites (Schildkraut, 1965). On the other hand, the
indoleamine theory postulates that it is not only noradrenaline but also a serotonin deficiency
that contributes to the disrupted regulation of basic behaviors observed in depressed patients.
In addition, studies have shown that an intact serotonin system is necessary for optimal
functioning of noradrenergic neurons (Price et al., 1990). Subsequently, the roles of
cholinergic-noradrenaline systems in depression were emphasized in the work of David
Janowsky et al (Janowsky et al., 1972). Dopamine also appears to play a role in the
pathophysiology of depression. Decreased dopamine transmission has been associated with
depression in post-mortem and animal studies (Dunlop & Nemeroff, 2007; Nutt et al., 2006).
Collectively, these theories are termed the monoamine hypothesis, as they involve
deficiencies in norepinephrine, dopamine and serotonin.
Altered brain serotoninergic function has been implicated as an underlying cause of
not only depression but also of other psychiatric disorders including anxiety, obsessive-
compulsive disorder, eating disorders and substance dependence (Bellivier et al., 1998;
Lucki, 1998; Mann et al., 2001). Evidence that supports the serotonin theory includes: (a)
significantly lower plasma tryptophan levels in subjects with major depression as compared
to healthy controls (Coppen et al., 1973; Cowen et al., 1989), (b) lower cerebrospinal fluid
concentrations of 5-hydroxyindoleacetic acid (5-HIAA), a major metabolite of 5-hydroxy-
tryptophan (5-HT), in untreated depressed patients (Owens & Nemeroff, 1994), (c) lower
concentrations of 5-HT and 5-HIAA in postmortem brain tissue of depressed and/or suicidal
subjects (Owens & Nemeroff, 1994), (d) a decrease in the number of 5-HT transporter or

12
binding sites in postmortem brain tissues of depressed patients and suicide victims and in
platelets of untreated depressed patients (Owens & Nemeroff, 1994). In addition, depriving
the brain of tryptophan, which leads to a central serotonin deficiency, can induce depression
within hours (Lam et al., 1996) or a profound relapse in remitted depressed patients who
previously responded to serotonergic antidepressant therapy (Owens & Nemeroff, 1994).
It is well-known that the essential amino acid tryptophan is a precursor for serotonin
(5-HT) synthesis in the human body. Once ingested, tryptophan is converted to 5-HT through
a series of reactions. However, the majority of tryptophan is converted to kynurenine
following the pathway as described in Fig. 1.1. A portion of serotonin is also converted to
melatonin. (Fig. 1.1) (Jonnakuty & Gragnoli, 2008; Kopin et al, 1961; Myint et al., 2007;
Watts et al, 2012)
5-HT is found in both the central nervous system, mostly in the brain-stem neurons of
the raphe nuclei, and in the peripheral system, mostly in the enterochromaffin cells in the
gastrointestinal tract crypts (Furness & Costa, 1982; Maurer-Spurej et al., 2004). After
release into the blood circulation from the enterochromaffin cells, 5-HT is rapidly taken up
by platelets via the 5-HT transporter and is stored in platelet dense granules. Therefore,
peripherally, platelets store almost all the total body circulating serotonin (Maurer-Spurej et
al., 2004). Since the blood brain barrier is impermeable to peripheral 5-HT, central 5-HT
synthesis depends on the amount of tryptophan or 5-hydroxy-L tryptophan available
peripherally to cross the blood brain barrier. While tryptophan has to compete with other
amino acids such as valine, leucine and isoleucine for a carrier protein in order to cross the
blood brain barrier, 5-hydroxy-L tryptophan can easily cross the blood brain barrier since it
does not require a specific transport protein (Rahman et al., 1982; Yuwiler et al., 1977).

13
Centrally, 5-HT functions at the neuron synapses and influences a variety of behavioral,
physiological and cognitive functions such as memory, mood, emotions, wakefulness, sleep,
appetite and temperature regulation (Jacobs & Azmitia, 1992).
In the periphery, traditionally as its name suggests, serotonin is a vasoconstrictor
causing hypertension (Maurer-Spurej et al., 2004). On the other hand, it also mediates nitric
oxide release and functions as an endothelium-derived relaxing factor (Maurer-Spurej et al.,
2004). It exerts its action through many subtypes of 5-HT receptors present on vascular
smooth muscle cells and endothelial cells (5-HT1B, 5-HT 2A, 5-HT 2B, 5-HT 4 and 5-HT 7)
(Nilsson et al., 1999). 5-HT is metabolized primarily in the liver and only a small fraction
escapes from this primary metabolism and is metabolized in the endothelium of lung
capillaries. Finally, the product of 5-HT metabolism, 5-Hydroxyindoleacetic acid (5-HIAA),
from either central or peripheral sources, is excreted in the urine (Green & Curzon, 1968).
In addition to the indolamine theory of depression, collective evidence has suggested
a corticosteroid receptor hypothesis in depression (Holsboer et al., 1984; Holsboer, 1999;
Holsboer, 2000). Overproduction of corticotrophin releasing hormone which causes an
excess activity of the HPA axis with hypercortisolemia is also observed in many depressed
patients (Gillespie & Nemeroff, 2005; Vreeburg et al., 2009). This prolonged or excessive
secretion of glucocorticoids may lead to suppression of neurogenesis and hippocampal
atrophy (Belmaker & Agam, 2008; Heim & Nemeroff, 2001; Sapolsky, 2000).
In parallel, the leptin hypothesis of depression has also been proposed based on
supporting evidence from animal and human studies. (Lu et al., 2007) The hypothesis of
leptin insufficiency in depression is mainly supported by data from a rodent study. Decreased

14
plasma leptin is associated with behavioral changes in chronically stressed mice or rats and
systemic administration of leptin can reverse the chronic stress-induced behavior changes or
depressive-like behaviors (i.e a decrease in sucrose preference or reduced duration of
immobility in force swim test or tail suspension test) together with an increase in c-fos
expression in the hippocampus. (Lu et al., 2006; Lu et al., 2007) Thus, leptin acts as an
antidepressant in the rat model. However, in the human, leptin insufficiency seems to occur
in only a subpopulation of depressed patients and it alone cannot explain the association
between obesity and depression. Thus, it raised a complementary hypothesis of leptin
resistance, which is similar to insulin resistance in type 2 diabetic patients. In addition, since
leptin is also capable of modulating HPA axis function, the leptin hypothesis for depression
might be complementary to the aboved mention HPA hypothesis of depression. (Lu et al.,
2007)

15
FIG. 1.1: Biosynthesis and metabolism of serotonin.
(Jonnakuty & Gragnoli, 2008; Kopin et al, 1961; Myint et al., 2007; Watts et al, 2012)
Kynurenine
N-acetylserotonin
Melatonin
Indoleamine diooxygenase/Tryptophan dioxygenase Tryptophan (TP)
5-hydroxy-tryptophan (5-HTP)
5-hydroxy-tryptamine (5-HT)
5-hydroxyindole acetaldehyde
5-hydroxyindole acetic acid (5-HIAA)
Tryptophan hydroxylase
L-amino acid decarboxylase
Monoamine oxidase
Aldehyde dehydrogenase
Hydroxyindole-O-methyltransferase

16
1.4 Depression In Pregnancy, Postpartum And In Children:
1.4.1 Incidence:
Even though depression can occur in both genders, it affects more women than men
with a female to male ratio of 1.7-2.7 (Bhatia & Bhatia, 1999; Burt & Stein, 2002; Kessler et
al, 1993). It is believed that biologic factors such as endocrine, genetic or social features
contribute to this gender difference (Nonacs & Cohen, 2003; Weissman & Olfson, 1995).
Studies have shown that acute depletion of tryptophan has a more profound impact on mood
in women as compared to men. This implies that women and men are different in terms of
the magnitude and functional activity of their central serotonergic system (Nishizawa et al.,
1997). Additionally, tryptophan pyrrolase, an enzyme that reduces blood tryptophan levels,
which is an amino acid precursor of serotonin, may be stimulated by circulating
concentrations of estrogen and progesterone (Khan et al.,, 2005). This may, in part, account
for the greater frequency of depression among women during their childbearing years.
Whatever the cause might be, women are much more likely to suffer from depression at any
stage of their lifetime, especially during their childbearing and child-rearing years. The
incidence of clinical depression during pregnancy is about 10-15% (Bhatia & Bhatia, 1999;
Burt & Stein, 2002; Nonacs & Cohen, 2003). Postpartum depression occurs in 10-22% of
women (Burt & Stein, 2002), with symptoms beginning two weeks after delivery including
tearfulness, despondency, labile mood, feelings of inadequacy and inability to cope,
particularly with the baby (Cooper & Murray, 1998; Hopkins et al., 1984; Murray, 1992;
O'Hara, 1997).

17
Depression in children and adolescents is an important issue as depressed adolescents
frequently grow up to be depressed adults (Emslie et al., 1997; Harrington et al., 1990;
Lewinsohn et al., 1999; Rao et al., 1999; Weissman et al., 1999). They have more social
problems later in life at work and with their families, attempt suicide more often and require
more medical and psychiatric attention (Birmaher et al., 1996; Brent et al., 1988; Rohde et
al., 1994). Suicide is the fourth leading cause of death in 10 to 14 year old children and rises
to the third leading cause of death in adolescents (Anderson, 2002). Statistics in Canada
reported that the suicide deaths in 1996 have increased three fold in 10-14 age group as
compared to the rate in 1971 (Houle & Wilkins, 2010). In addition, there is a subsequent
increase in the likelihood of tobacco use, involvement in deviant activities and accidents and
impaired relationship with parents and partners, and also impaired academic function in these
depressed adolescents (Birmaher et al., 1996; Kandel & Davies, 1986; Rohde et al., 1994).
Childhood depression is more common than it used to be thought. In fact, depression
can be diagnosed in children as young as 3 years old (Dubovsky et al., 2003). In general,
depression occurs in 2% of prepubertal children and in 3-8% of adolescents (Birmaher et al,
1998; Costello et al., 1996; Lewinsohn et al., 1994).
1.4.2 Risk Factors Of Depression:
Generally, multiple factors may contribute to the occurrence of depression including
marital status, socioeconomic factors, residence, seasonal and geographic factors, social
stressors, lack of social support and the presence of comorbidity (Kaplan & Sadock's
comprehensive textbook of psychiatry2005). Omega-3-fatty acid, which reduces the turnover
of membrane phospholipids, is considered as an adjunctive mood stabilizer (Dubovsky et al.,

18
2003). However, there is evidence that a diet low in omega 3 fatty acids or fish consumption
is not associated with depressed mood, major depression or suicide (Hakkarainen et al.,
2004).
Early experience of loss or trauma and stress can initiate or exacerbate neurochemical
imbalance in vulnerable subjects. This idea is supported by studies in animals in which early
separation from peers and inescapable situations profoundly alters the turnover of their
biogenic amines and postsynaptic receptor sensitivity (Gilmer & McKinney, 2003). In
humans, a polymorphism of the serotonin transporter gene can identify who among
traumatized children will develop adult depression (Caspi et al., 2003; Kendler &
Karkowski-Shuman, 1997; Lesch et al., 1996; Ogilvie et al., 1996; Ogilvie & Harmar, 1997).
There are many factors that can play a role in the occurrence of postpartum
depression. Even though there are no firm causal links, the significant changes in hormone
levels of progesterone, estrogen, cortisol, neurosteroids and β-endorphins may lead to
depression in this period (McCoy et al., 2003; Weissman & Olfson, 1995). Additional
predictive factors are obstetric pregnancy complications, infant irritability and poor motor
function in the early neonatal period (Murray, 1993; Murray et al., 1996; Waterstone et al.,
2003). Besides these, probably the most attributed risks are psychiatric and psychosocial
factors including prior history of mood disorder, low socioeconomic status, lack of partner or
family support, an unwanted pregnancy and young maternal age (Burt & Stein, 2002;
Hopkins et al., 1984; Murray, 1993; Warner et al., 1996).
When left untreated, depression during pregnancy can lead to changes in maternal
behaviours such as decreased appetite, poor pregnancy-related weight gain, increased

19
likelihood of smoking and/or alcohol and drug consumption and even a risk of suicide in
severe cases (Ahluwalia et al., 2004; Bhatia & Bhatia, 1999; Nonacs & Cohen, 2003). There
is also evidence that these poor maternal health behaviours or depression per se are
associated with an increased risk of ischemic heart disease (Glassman & Shapiro, 1998).
These place both the mother and fetus at risk.
1.4.3 Effects Of Maternal Depression On Fetal Neurobehavioral Development:
Studies have shown that depression per se, without poor maternal behavior, can result
in pregnancy complications such as preterm birth and/or small for gestational age, and
increased rates of maternal obstetric complications (Anderson, 2004; Andersson et al., 2004a;
Andersson et al., 2004b; Dole et al., 2003; Hoffman & Hatch, 1996; McCormick et al., 1990;
Orr & Miller, 1995; Orr et al., 2002; Peacock et al., 1995; Steer et al.,, 1992; Weissman &
Olfson, 1995). It is not yet clear via which mechanisms depression can lead to preterm birth
and fetal growth restriction. One explanation for this relationship is that the elevated plasma
concentrations of maternal cortisol and catecholamines associated with depression lead to a
decrease in uterine blood flow (Hoffman & Hatch, 1996; Lake et al., 1982; Linkowski, 2003;
Weissman & Olfson, 1995; Rurak, 1995), and thus reduce nutrient supply to the fetus and
impair fetal growth (Lang et al., 2000). Moreover, the elevated endogenous maternal
corticosteroids and catecholamine concentrations can affect the fetal central nervous system
causing infant irritability such as excessive crying, difficulty in consoling, which is a
predictive factor of postpartum depression (Murray, 1993; Welberg et al., 2000; Zuckerman
et al.,1990). As a result, maternal depression during pregnancy can extend into the
postpartum period via altered neonatal behaviour (Zuckerman et al., 1990). This, in turn,
could further impair neurological development of the offspring. Indeed, some evidence has

20
indicated that maternal depression can adversely affect temperament and cognitive
development in the infant. Similarly, depressed mothers of preschoolers have more negative
perceptions of and interactions with their children (Beck, 1996; Hay & Kumar, 1995; Lang et
al., 1996).
1.4.4 Treatment Of Depression:
Given the negative impact of maternal depression in pregnancy on both the mother
and offspring, treatment for this condition is essential. Due to the general concern about
exposure of the fetus to drugs, psychotherapy seems to be a preferable choice of treatment.
However, there are few studies that assess the efficacy of psychotherapy during pregnancy.
Drug therapy is a norm and antidepressants are the preferred course of treatment even though
these drugs are not approved for use in pregnancy (Altshuler et al., 2001; Bhatia & Bhatia,
1999; Nonacs & Cohen, 2003; Weissman & Olfson, 1995). Selective serotonin reuptake
inhibitors (SSRIs) were first introduced into the market in the late of 1980s (DeVane, 1999),
and by 1998 they largely replaced the older classes of antidepressants for the treatment of
depression in pregnancy (Altshuler et al., 2001; Goldstein, 1998; Goodnick & Goldstein,
1998b; Misri et al., 2000; Nonacs & Cohen, 2003; Simon et al., 2002; Skop et al., 1994) due
to their lower incidence of side effects and greater safety in terms of drug overdose
(Goodnick & Goldstein, 1998b; Jones & Blackburn, 2002). One recent study has reported an
increasing use of antidepressants in pregnancy from 2% in 1996 to 7.6% of deliveries in
2004-2005, and in which SSRI use increased 4.9% from 1996 to 2004 (Andrade et al., 2008).
There are similar reports that SSRI accounts for more than 70% of prescribed antidepressant
as a pharmacotherapy in pregnant women (Cooper et al., 2007; Simon et al., 2002). In
addition, these agents have also been used to treat other psychotic disorders in pregnancy,

21
which are associated with a dysfunctional serotonin system, such as anxiety and obsessive-
compulsive disorders (OCD) (Levine et al., 2003; Vaswani et al., 2003).
1.5 Selective Serotonin Reuptake Inhibitors (SSRIs):
1.5.1 Clinical Uses:
SSRIs are considered as a second generation of antidepressant after the first
generation of monoamine oxidase inhibitors (MAOI) and tricyclic antidepressants (TCA). In
general, SSRIs are widely used in psychiatry. Since SSRIs are relatively safe in terms of
overdose and have a more benign side-effect profile than the older generation agents, they
offer a greater tolerability and compliance to the treatment. (Lane et al., 1995; Masand &
Gupta., 1999) Besides their main indication for the treatment of major depressive disorders,
SSRIs are also prescribed as a first-line pharmacological treatment for other disorders such as
bipolar disorder, social anxiety disorder, generalized anxiety disorder, obsessive-compulsive
disorder (in combination with cognitive-behavioral therapy), panic disorder, eating disorder
(bulimia), premenstrual dysphoric disorder, and posttraumatic stress disorder. (Boyer et al.,
1992, Masand & Gupta., 1999) In addition, they are also used to control mood disorders in
special subsets of the population including post stroke patients, and terminally-ill patients
(cancer, HIV/ADIS) (Franco-Bronson et al., 1996; Sauer et al., 2001) A typical response to
treatment occurs within four to six weeks of SSRIs treatment.
1.5.2 Mechanism Of Action:
Monoamine transmitter activity may be regulated at many different levels including
synthesis, packaging and storage of neurotransmitters in synaptic vesicles, release, reuptake,

22
metabolism and postsynaptic receptor responsiveness (Fig. 1.3) (Kaplan & Sadock's
comprehensive textbook of psychiatry 2005).
FIG. 1.2: Schematic diagram of monoaminergic synapse and steps involved in synaptic transmission: Monoamine neurotransmitters (1) are synthesized within neurons from common amino acid precursors, which are Tryptophan and 5-Hydroxytryptophan (5-HT) in the case of serotonin, (2) are taken up into synaptic vesicles via a vesicular monoamine transporter, (3) are released into the synaptic cleft upon stimulation, (4) interact with postsynaptic receptors to alter the excitability of postsynaptic cells, (5) interact with presynaptic autoreceptors located on the nerve terminal to suppress further release. (6) Released monoamines may also be taken back up from the synaptic cleft into the nerve terminal by plasma membrane transporter proteins. (7) Once taken up, monoamine may be subject to enzymatic degradation or may be protected from degradation by uptake into vesicles. MAO: monoamine oxidase Adapted from Kaplan and Sadock’s comprehensive textbook of Psychiatry 2005.
1
2
3
4
5 6
7
MAO
amino acidprecursor
monoamine neurotransmitter

23
The goal of drug therapy in the treatment of depression is to maintain the circulating
neurotransmitter concentration in the synapse at a certain level, either by increasing the
amount of neurotransmitter produced for release or the duration of time the neurotransmitter
spends in the synapse. Antidepressants are classified based on the type of monoamine
neurotransmitter (dopamine, noradrenaline or serotonin), being affected at the synapse and
the specific mechanisms being impacted such as synthesis, release, reuptake, and
metabolism. There are three major classes of antidepressants. Monoamine oxidase inhibitors
(MAOI) prevent the degradation of noradrenaline and serotonin in the presynaptic cell.
Tricyclic acids (TCA) block the reuptake of noradrenaline and serotonin to varying degrees
at the presynaptic cell. SSRIs selectively inhibit the reuptake of serotonin (5-HT) by
presynaptic serotoninergic neurons and peripheral serotonin containing cells, particularly
platelets, via effects on the serotonin transporter (Anderson, 2004; Goodnick & Goldstein,
1998a; Stahl, 1998; Wright & Angus, 1989). In addition, there are sub-classes of dual
inhibitors of the three neurotransmitters mentioned above that have been developed,
including norepinephrine/dopamine reuptake inhibitor (NDRI), serotonin/norepinephrine
reuptake inhibitor (SNRI), noradrenergic and specific serotonergic antidepressant (NaSSA)
and serotonin antagonist reuptake inhibitor (SARI) (Stahl, 1998).
The various SSRIs (sertraline, fluoxetine, paroxetine, citaloprame, fluvoxamine)
differ in their potency and inhibitory effects on reuptake of other monoamine
neurotransmitters (Kaplan & Sadock's comprehensive textbook of psychiatry 2005).
However, comparisons of these agents in acute treatment have shown that they have similar
efficacies, probably due to the parallel differences in dose and plasma concentration
(DeVane, 2003; Hiemke & Hartter, 2000).

24
After SSRI treatment, the inhibition of serotonin reuptake usually occurs immediately
whereas the onset of the antidepressant effect is delayed for about 2-6 weeks (Goodnick &
Goldstein, 1998b). It is generally accepted that the initial rise in extracellular 5-HT in the
somatodendritic regions of the dorsal and median raphe nuclei activates somatodendritic 5-
HT1A autoreceptors which are responsible for the inhibition of the firing of 5-HT neurons
and thus reducing 5-HT release. Yet, when given chronically, SSRI will down-regulate the 5-
HT1A autoreceptors and increase serotonergic neurotransmission (Celada et al, 2004;
Hensler, 2003; Hjorth, 1996; Stahl, 1998). In addition, other neurotransmitters can also be
involved in the control of the firing rate and serotonin release (Kalsner, 2000; Kalsner &
Abdali, 2002).
1.5.3 Pharmacokinetics:
The different SSRIs vary in their half-lives and other pharmacokinetic variables.
Fluoxetine (FX) has the longest half-life, whereas the values for the other SSRIs are shorter
(Hiemke & Hartter, 2000; Vaswani et al., 2003). The wide difference in the half-lives among
SSRIs account for the differences not only in the time to reach the steady state, the time to
clear the drug from the body after discontinuation but also in the potential for withdrawal
symptoms after discontinuing the SSRIs. Concomitant use of SSRIs and other agents
metabolized by the cytochrome P450 (CYP) isozymes can lead to drug-drug interactions,
because SSRIs inhibit the activity of some of the isozymes. Among SSRIs, FX and
paroxetine are strong CYP2D6 inhibitors, and the others are weak CYP2D6 inhibitors. FX is
also a moderate CYP2C19 inhibitor (Harvey & Preskorn, 2001). Additionally, FX is also a
substrate of CYP2C19 and CYP2D6 (Jeppesen et al., 1996; Ring et al., 2001; Spina et al.,
2003, Harvey & Preskorn, 2001)

25
Genetic polymorphisms have been identified in two CYP isoenzymes, CYP2D6 and
CYP2C19, and that divides the population into several groups including ultra extensive,
extensive, intermediate and poor metabolizers. The poor metabolizers with nonfunctional
allele of CYP2D6 gene have a severely impaired catalytic function of the isoenzymes. The
clinical impact of poor metabolizers is that they are unable to metabolize the drugs, which are
substrates of these enzymes; therefore the sub-population will have a higher risk of adverse
effects, especially when no other metabolic route is available for the drugs or when multiple
drugs share the same CYP in their metabolism (Vaswani et al., 2003). This becomes more
important when switching medications having a long half live (eg: FX) to other medications
metabolized by the same cytochrome since the risk of toxicity still persists long after the
drugs were discontinued.
Among the SSRIs, paroxetine, sertraline and fluoxetine are highly bound to plasma
proteins and only a small fraction of the drugs stays in the form of free drug. Thus,
coadministration of the SSRIs and another highly-protein-bound drug might elevate the free
concentrations of the other drug, potentially resulting in adverse events.
All of the SSRIs can readily cross the placenta in humans, rats and sheep because
they are highly lipophilic (Hendrick et al., 2003; Kim et al.,2004; Kim et al., 2006; Pohland
et al., 1989; Rampono et al., 2009). SSRIs are also excreted in breast milk. Despite this fact,
the concentrations of the drugs in an infant are much less than those in a fetus, because of the
much lower amount of drug acquired by the infant via breast milk compared to that acquired
by the fetus via placental transfer and also because of the progressive development of
metabolic capacity in the infant following birth (Kim et al., 2006). Hence, the exposure of a
fetus to the drugs during prenatal period is more likely to be the cause of any long-term

26
effects of the drugs in compared to the exposure of an infant to the drugs during the postnatal
period.
1.5.4 Side Effects:
As a class, SSRIs, clearly demonstrate superiority in terms of safety and tolerability
compared to the older antidepressant medications. A meta-analysis of 84 double-blind
Randomized Controlled Trials (RCT) of the adverse effects of antidepressants, particularly
SSRIs and TCA has found that all antidepressants cause adverse effects (Trindade et al.,
1998). Typical anticholinergic side effects, including dry mouth, constipation, blurred vision,
urinary retention and postural hypotension, which are usually reported in tricylic
antidepressants, occur less frequently with SSRIs. Serotonin has been implicated as playing
roles in gastrointestinal motility, appetite control, sexual behavior, sleep regulation, and
cerebral vasomotor tone; therefore SSRIs are predicted to cause nausea, appetite loss, sexual
dysfunction, and sleep disturbances (Dubovsky et al., 2003). In fact, there is evidence that
fluoxetine, fluvoxamine, paroxetine and sertraline have been associated with nausea,
diarrhea, insomnia, nervousness, agitation and anxiety. Even though paroxetine was found to
induce less nausea than the other SSRIs, the difference was not significant (Trindade et al.,
1998). SSRI-associated adverse effects seem to be related to drug dose (Beasley et al.,1991).
Together with the introduction of SSRIs and the increased incidence of depression in
children and adolescents, SSRIs is also used to treat children with this condition. SSRIs were
found to be associated with a higher rate of agitation and suicidal thoughts, or both, from
clinical trials comparing SSRIs treatment and placebo in children and adolescents with major
depressive disorder. Thus, a warning for a close observation of signs or symptoms of

27
worsening depression, including suicidal ideation, early on in therapy with these medications
or on titration of dosing, was required to be included in their labels and product inserts
(Bridge et al., 2007; Hammad et al., 2006). In the meta-analyses, SSRI treatment had a
benefit compared to placebo treatment, but also was associated with significant risks for
adverse events, including suicidal behaviour (except for FX) (Jureidini et al., 2004;
Whittington et al., 2004)
1.6 Fluoxetine (FX):
FX is an antidepressant that belongs to the SSRI class. It was one of the first SSRIs to
be developed. FX (trade name Prozac) was first introduced into the US market in 1988
(Borys et al., 1990). It has been shown to effectively relieve the symptoms of major
depression and is better tolerated than the older classes of antidepressant such as TCA (Cohn
& Wilcox, 1985). Currently, FX also has received approved indications for OCD, bulimia
nervosa and premenstrual dysphoric disorder. In 2003, the FDA approved the combination of
FX and olanzapine (trade name Symbyax) for treatment-resistant depression (unresponsive to
two separate trials of different antidepressants). Despite the fact that many new SSRI agents
have been introduced into the market after FX was, FX remains very popular. In 2008, over
23 million prescriptions of FX were filled in the US, an increase of 4.5% from 2007. Also, it
was ranked the second most prescribed antidepressant (Top 200 generic drugs by total
prescriptions 20082009).

28
1.6.1 Pharmacokinetics:
1.6.1.1 Absorption:
The bioavailability of FX is relatively high, about 70 to 90 percent. This
bioavailability results from an almost complete absorption and a limited hepatic first-pass
effect. The time to maximum concentration (tmax) of FX is about 6-8 hours. And the peak
plasma concentration (Cmax) of FX is about 15-55 ng/ml (Altamura et al., 1994). Food does
not decrease the absorption, but may delay the time to reach the peak concentration by 1-2
hours (Benfield et al., 1986).
1.6.1.2 Distribution:
FX is one of the SSRIs highly bound to plasma proteins, approximately 95%. The
volume of distribution (Vd) of FX and its active metabolite, norfluoxetine (NFX), falls in the
range from 12 to 45 L/kg (Benfield et al., 1986).
1.6.1.3 Metabolism:
FX is a 1:1 racemic mixture of two optical isomers. Both S- and R- enantiomers have
equivalent pharmacologic activity, although the S-enantiomer is eliminated more slowly and
therefore presents predominantly in the plasma at steady state (Wong, Fuller, & Robertson,
1990). FX is metabolized by the cytochrome (CYP) P450 isozymes in the liver. FX is
demethylated to its major active metabolite, norfluoxetine (NFX) and several inactive
metabolites. The two clinically important CYP P450 isozymes involving the metabolism of
FX are CYP2C19 and CYP2D6 (Stevens & Wrighton, 1993). CYP2D6 catalyzed the
metabolism of R & S FX and S-NFX but not of R-NFX (Fjordside et al., 1999).

29
1.6.1.4 Elimination:
Hepatic metabolism of FX to its metabolites then renal excretion is the primary route
of elimination. The half-life (t1/2) values of fluoxetine following an acute and chronic
administration are 1-3 days and 4-6 days respectively. The major active metabolite, NFX, has
a longer half-life, which ranges from 4 to 16 days, independent of length of treatment. The
relatively slow elimination of FX and NFX ensure that they will remain in the body for
weeks after discontinuation. Hepatic impairment will prolong the half-life elimination of FX
and NFX.
1.7 FX Effects On Newborns And Infants:
1.7.1 FX Effects On Cardiovascular Functions:
There are a few case reports in adults of adverse cardiac events after starting FX
therapy including sinus bradycardia, supraventricular tachycardia, atrial fibrillation, atrial
flutter and ST segment depression. (Ahmed et al., 1993; Gardner et al., 1991; Isbister et al.,
2004; McAnally et al., 1992; Riddle et al., 1989; Roberge & Martin, 1994). One case report
of FX poisoning in which the patient ingested 12g of FX and developed bradycardia,
progressing to ventricular fibrillation and asystole (Compton et al., 2005). Another case
report of QT prolongation to 600msec and torsades de point in a 74 year-old woman occurred
3 weeks after starting FX therapy (Appleby et al., 1995).
QTc interval prolongation was also reported in a newborn after in utero exposure to
FX. To note, a prolonged QTc interval (> 560 msec) predisposes the sufferers to torsade de
pointes, an intermittent ventricular dysrhythmia which, in turn, can lead to a potentially life-

30
threatening monomorphic ventricular tachycardia or fibrillation. A subsequent retrospective
cohort study comparing 52 SSRIs exposed newborns with 52 unexposed controls matched by
gestational age also confirmed that the exposed newborns had a longer QTc interval than the
unexposed controls (409 vs 392 msec, respectively). However, there were no cases of
arrhythmia reported. The SSRIs used in the study included paroxetine, citalopram,
fluoxetine, fluvoxamine and venlafaxine and there was no drug-specific evaluation or serum
electrolytes measurement (Dubnov-Raz et al., 2008). In agreement with these findings,
prolongation of QTc interval of no greater than 450 msec existed in 62% of FX overdose
patients in one study, but none developed ventricular dysrhythmia (Phillips et al., 1997).
Nevertheless, of the SSRIs, FX appears to have minimal cardiotoxicity.
Effects of SSRIs exposure on cardiovascular function are likely to originate in utero.
Recently our group has found a reduced fetal middle cerebral artery (MCA) flow resistance
and cross sectional MCA area and fetal heart rate variability in the SSRIs-exposed fetus,
even after controlling for maternal mood. Importantly, despite a reduced MCA flow
resistance, MCA volume flow was not increased, and this was associated with a reduced
cross sectional MCA area (Rurak et al., 2011).
Chambers et al., (2006) was one of the first papers that reported an increased risk of
persistent pulmonary hypertension (PPHN) development (adjusted OR = 6.1, 95% CI 2.2-
16.8) in newborns whose mothers were taking SSRIs during late gestation. Even though there
were some methodological limitations in the study such as retrospective design and potential
recall bias, her finding was confirmed by a subsequent study by Kallen & Olausson, (2008).
Despite the fact that persistent pulmonary hypertension is a rare condition, these findings
prompted the FDA to issue a public health advisory to add a warning label of potential risk of

31
PPHN in 3rd trimester SSRI-exposed infants (Public health advisory: Treatment challenges
of depression in pregnancy and possibility of persistent pulmonary hypertension in
newborns.2006). However, two following studies, which involved retrospective examinations
of administrative health records, failed to demonstrate this increased risk of PPHN in SSRIs-
exposed infants (Andrade et al., 2009; Wichman et al., 2009). However, both studies were
underpowered to detect small differences in the incidence of PPHN, which itself is a rare
event. In another human study (Wilson et al, 2011), several potential factors, including use of
SSRIs after 20 weeks gestation, were examined in 20 case of primary PPHN compared to
120 controls. The only significant association was between cesarean delivery before labor
onset and PPHN and the authors noted that the 2 studies reporting an association between
antenatal SSRI exposure and PPHN (Chambers et al., 2006; Kallen et al., 2008) did not
control for the mode of delivery. A very recent study from our group measuring fetal right
pulmonary artery resistance, flow and vessel cross sectional area by Doppler ultrasound also
did not support an association between SSRIs exposure and PPHN in the infants even though
the authors observed a significantly higher rate of right pulmonary artery flow in the SSRI-
exposed fetuses with respiratory difficulties (Lim et al., 2012).
In a study in pregnant rats involving daily administration of fluoxetine by gastric
gavage from day 11 to 21 of pregnancy (Fornaro et al., 2007), the offspring from the
fluoxetine treated group exhibited an increase in pulmonary arterial medial thickness and in
the right/left ventricular weight ratio. In vitro, fluoxetine at lower concentrations increased
proliferation of fetal but not adult pulmonary arterial smooth muscle cells, whereas at higher
concentrations the drug inhibited proliferation of both the fetal and adult cells.

32
In sheep, 8 days of late gestational FX infusion in pregnant ewes did not result in any
changes in the daily average maternal arterial pressure and heart rate nor in fetal arterial
pressure. However, there was a transient decrease of fetal heart rate on both the FX exposed
group (infusion d2 and 6) and the control group (infusion day 4 and 8) (Morrison et al.,
2002).
1.7.2 FX Effects On The Homeostatic And Respiratory Functions:
Extensive research focused on brainstem mechanisms underlying sudden infant death
syndrome has shown the essential role of 5-HT neurons in the modulation of the respiratory
response and thus homeostatic functions (Kinney, 2009; Kinney et al., 2009). In vitro studies
have indicated that the midbrain 5-HT neurons, which serve as an arousal chemoreceptor,
and the medullary 5-HT neurons, which function as respiratory chemoreceptor, increase their
firing rate 3-fold on average in response to hypercarnia and a decrease in pH from 7.4 to 7.2
(Buchanan & Richerson, 2010; Richerson, 2004; Wang et al., 1998). In addition, acute
hypercapnia can also activate the limbic systems causing dyspnea and panic which are
usually observed in panic disorders (Gorman et al., 1994; von Leupoldt & Dahme, 2005).
Clearly, given that 5-HT neurons play an important role in homeostatic function, changes in
the serotonergic system either by depression itself or pharmacologic intervention may lead to
changes in homeostatic systems.
1.7.3 FX Effects On Birth Outcomes:
A majority of research regarding the effects of SSRI on birth and pregnancy
outcomes have found that late pregnancy SSRI exposure increases the risk of preterm birth,
shorter gestational length, lower birth weight, smaller than gestational age, lower APGAR

33
score at birth, admission to NICU and reduced postnatal weight gain (Chambers et al., 1996;
Einarson et al., 2010; Kallen, 2004; Lewis et al., 2010; Maschi et al., 2008; Oberlander et al.
2006; Oberlander et al. 2008; Pearson et al., 2007; Simon et al., 2002; Suri et al., 2007; Wen
et al., 2006; Wisner et al., 2009; Zeskind & Stephens, 2004). However, others have failed to
confirm these effects (Casper et al., 2003; Hendrick et al., 2003; Kulin et al., 1998; Lund et
al. 2009; Malm et al., 2005; Suri et al., 2004). There are conflicting findings regarding the
impact of timing and duration of SSRI exposure during pregnancy on perinatal outcomes.
Chambers et al., (1996) found that preterm birth, low birth weight and poor neonatal
adaptation were significantly more common in a late gestational exposure group than in an
early exposure group. Conversely, Oberlander et al., (2008) reported early and late exposure
groups had the same neonatal outcomes while an increase in days of exposure was
significantly associated with low birth weight and low gestational age, even after controlling
for maternal illness and medication dose. Still, others found no association between late
gestation exposure and adverse neonatal outcomes (Cohen et al., 2000; Malm et al., 2005).
The potential reason for the lack of consistency in these findings is the differences in doses
among studies. Recently, it has been reported that much higher preterm birth rate and lower
gestational age were observed in the higher FX dose group, even after controlling for length
of exposure (Roca et al., 2011). The mechanisms underlying the association between prenatal
SSRI exposure and the shortening of gestational length are not yet been clear. However, it
has been proposed that SSRI exposure might promote adverse birth outcome through the
dysregulation of the fetal HPA system or through the vasoconstrictor effects of serotonin
which could decrease maternal placental blood flow, especially in the 3rd trimister (Morrison
et al., 2002; Roca et al., 2011). SSRIs-induced fetal HPA axis disturbances were evidenced

34
by a previous sheep study in our lab showing an increase in the fetal plasma prepartum
cortisol surge and apparent increased sensitivity of the fetal adrenal gland to ACTH after 8 d
maternal FX administration (Morrison et al., 2004). In addition, a relationship between
elevated placental corticotropin-releasing hormone (CRH) levels, which also regulates
cortisol secretion, and a higher risk of preterm birth and fetal growth restriction has been
established in human pregnancies associated with antenatal SSRI therapy (Wadhwa et al.,
2004). Other potential mechanisms concerning the effect of SSRIs exposure on lower birth
weight or smaller than gestational age include a SSRIs-induced alteration in the fetal IGF-
1/GH axis (Davidson et al., 2009) or a reduction in uterine blood flow by 5-HT, a
vasoconstrictor agent, which limits oxygen and nutrient supply to the fetus (Morrison et al.,
2002) or a direct SSRIs-effect on the growing skeleton by exerting their effect on osteoclast
differentiation and formation (Battaglino et al., 2004; Warden et al., 2005).
1.7.4 FX Effects On Neonatal And Child Behaviors:
Accumulating data from case reports and cohort studies have also indicated that late
gestation exposure to SSRIs increases the risk of a variety of neonatal complications termed
“neonatal behavioral syndrome” or “poor neonatal adaptation” (Chambers et al., 1996;
Cohen et al., 2000; Kwon & Lefkowitz, 2008; Mhanna et al., 1997; Oberlander et al., 2004).
According to some studies, the risk can increase up to 3 times (Chambers et al., 1996; Cohen
et al., 2000; Oberlander et al., 2004). Among SSRIs, most reported cases involved FX and
paroxetine exposure. Behavioral and neurologic symptoms vary from mild central nervous
system, motor, respiratory and gastrointestinal signs and metabolic dysfunction (including
irritability, persistent crying, shivering, tremor, restlessness, feeding difficulties, sleep
disturbances) to severe symptoms (including seizures, dehydration, excessive weight loss,

35
hyperpyrexia and the need for intubation). These symptoms usually disappear by the second
week after birth (Moses-Kolko et al., 2005). These findings led Health Canada and the FDA
in 2004 to include a warning in the drug information labeling for SSRI/SNRIs that neonates
exposed to these agents in the late third trimester have developed adverse events (Summary
minutes of the pediatrics subcommittee of the anti-infective drugs advisory committee.2004).
These symptoms are observed in around 10-30% of the exposed newborns (Koren et al.,
2009). Unfortunately, the mechanisms that may account for these complications have yet to
be elucidated. They may be multifactorial, with the severity of the maternal mental illness
playing a major role.
In fact, one of the big challenges to this type of research in humans is to distinguish
between the drug effects and the impact of maternal psychiatric disorders on the occurrence
of neonatal behavioral syndrome. Even though some studies have enrolled mothers with
untreated depression in their studies to adjust for the possible indication confounders, those
are more likely to have less severe psychiatric problems than mothers with treated
depression. Using a propensity score matching approach, Oberlander et al., (2006) have
found that the incidence of symptoms of behavioral syndrome from SSRI exposed infants is
similar to non-exposed infants born to depressed mothers.
Since the symptoms observed in adults with SSRI discontinuation syndrome overlap
with those seen in neonates born to mothers treated with SSRIs, one of the mechanisms
suggested for poor neonatal adaptation syndrome is a withdrawal syndrome. On the other
hand, some symptoms in the neonates are similar to those seen as side effects of SSRIs or
SSRI toxicity associated with elevated 5-HT levels (serotonin syndrome) (Birmes et al.,
2003; Ener et al., 2003). Therefore, it remains unclear whether neonatal symptoms are due to

36
SSRI withdrawal or directly due to SSRI toxicity (Koren et al., 2009; Moses-Kolko et al.,
2005). However, since theoretically the management for these mechanisms is clearly
opposite, there is a compelling need to identify which is the true case. Currently, the
management for SSRI/SNRI poor neonatal adaptation syndrome is mainly symptomatic, with
supportive care only (Koren et al., 2009).
In parallel with the mechanisms suggested from human studies, previous studies on
fetal lambs with chronically 8-day FX-exposure in late gestation in our lab (Morrison et al.,
2001; Morrison et al., 2002; Morrison et al., 2004; Morrison et al., 2005) have suggested
another mechanism: that late gestational SSRIs exposure could alter fetal brain development,
which relates to the importance of serotonin in the maturation of brain pathways. During the
period of FX administration, the incidence of fetal rapid-eye-movement (REM) sleep was
significantly decreased (Morrison et al., 2001). In addition, a study of human fetal behavioral
development also demonstrated an abnormaly high rate of continual bodily activity during
non-REM sleep in SSRI-exposed fetuses (Mulder et al., 2011). This mechanism is further
supported by a subsequent study from our group which showed additive negative effects in
infants of depressed mothers treated with SSRIs as compared to untreated depressed mothers
(Oberlander et al., 2006). Our recent findings of a decreased MCA flow resistance without
increasing MCA volume flow in the fetus exposed to SSRIs (Rurak et al., 2011) adds more
evidence to support this proposed mechanism. Changes in tissue volume in the brain region
perfused by the MCA, which is secondary to changes in brain blood flow, could be due to a
SSRIs-mediated alteration of serotonergic tone. However, whether the disturbed serotonergic
system places any long-term sequelae during postnatal development beyond the exposure
period still needs further investigation. To date, only two studies have reported significant

37
difference in psychomotor development in SSRIs-exposed versus non-exposed infants
(Casper et al., 2003; Mortensen et al., 2003).
1.7.5 FX Effects On The Hypothalamic-Pituitary-Adrenal (HPA) Axis:
HPA axis activity is proposed to be involved in the pathophysiology of depression
since altered levels of corticotropin-releasing factor in the cerebrospinal fluid (Arborelius et
al., 1999) and hypercortisolemia are observed in depressed patients (Holsboer et al., 1995)
and this was normalized with antidepressants (Holsboer & Barden, 1996) possibly by
increasing hippocampal glucocorticoid receptor expression (Yau et al., 2002). The function
of the HPA axis and serotonergic system are highly interrelated. Hypothalamic CRH might
modulate the serotonergic system (Boadle-Biber et al., 1993; Fuller, 1996a). Conversely,
serotonergic neurons in the raphe nucleus can directly stimulate HPA axis function via 5-
HT1A and 5-HT2A receptors in the paraventricular nucleus of the hypothalamus (Fuller,
1996b). Therefore, increases in extracellular 5-HT levels acutely and in serotonergic
neurotransmission chronically by treatment with SSRIs, such as FX, in depressed mothers
could have an impact on fetal HPA activity. As mentioned previously, in our sheep studies,
8-day late gestational FX i.v infusion in pregnant ewes resulted in an augmented prepartum
rise in fetal ACTH and cortisol levels and reduced low voltage rapid eye movement
behavioral state in the fetus (Morrison et al., 2004). Although the mechanisms are not fully
understood, alterations in HPA axis activity in the fetus might be one of the possibilities
underlying negative perinatal outcomes observed in human studies. In addition, this is
particularly important in the context of fetal exposure to excess glucocorticoids either by
prenatal stress or maternal illness (e.g: depression, anxiety disorders) or pharmacologic
agents effects (eg: SSRIs, synthetic glucocorticoids for lung maturation in preterm labor),

38
which can program the HPA axis in ways that it will have negative impacts on mental,
metabolic and cardiovascular health later in life (Lupien et al., 2009; Seckl, 1997; Tamashiro
& Moran, 2010). Interestingly, a recent study from our group, which further examined the
long-term effects of prenatal SSRIs (fluoxetine) on the HPA axis in the postnatal period,
have shown an association between SSRI exposure and reduced basal cortisol levels as well
as an alteration in the HPA stress response pattern (Oberlander et al., 2008). A lower cord
blood cortisol level in SSRIs-exposed neonates was also reported in a subsequent study
(Davidson et al., 2009). Conflicting findings from the sheep and human studies could be, in
part, attributed to the differences in the timing and duration of exposure which, determine the
nature of the offspring adaptive responses and the physiological disturbances in later life
(Chadio et al., 2007).
Sex differences in the HPA axis normalization in response to antidepressant treatment
have been observed (Binder et al., 2009). In rodents, there is a differential HPA axis response
between males and females following acute SSRIs (citalopram). Female mice had a greater
corticosterone response after citalopram administration than males and testosterone treatment
in ovariectomized females eliminated the sex differences (Goel & Bale, 2010). Similar
differences were also reported with sertraline treatment in sheep (Broadbear et al., 2004).
ACTH and cortisol secretion significantly increased in female but not in male after sertraline
s.c injection, independent of circulating steroid hormones. These studies indicate that the
HPA axis of the female is more sensitive to the stimulatory effects of serotonin than that of
the male.

39
1.8 Rationale:
The incidence of clinical depression during pregnancy is about 10-15% (Bhatia &
Bhatia, 1999; Nonacs & Cohen, 2003). Postpartum depression occurs in 10-22% of women,
with symptoms beginning two weeks after delivery (Burt & Stein, 2002). Depression during
pregnancy can place both the mother and fetus at risk (Ahluwalia et al., 2004; Orr & Miller,
1995; Zuckerman et al., 1989), and there is abundant human evidence for adverse effects of
maternal postpartum depression on the cognitive, motor and emotional development of the
offspring (Kurstjens & Wolke, 2001; Murray & Cooper, 1997; Murray et al., 2001). Since
their introduction in the late 1980’s, SSRIs remain a first-line pharmacological treatment for
both depression in pregnancy and in the postpartum period (Altshuler et al., 2001; Misri et
al., 2000). However, numerous studies in humans have shown that infants delivered from
women taking SSRIs during pregnancy have a higher risk of a number of adverse pregnancy
outcomes, including premature birth, low birth weight, decreased APGAR scores at birth,
admission to special-care nurseries, reduced postnatal weight gain, persistent pulmonary
hypertension and poor neonatal adaptation (Chambers et al., 1996; Chambers et al., 1999;
Lattimore et al., 2005; Oberlander et al., 2006). Human studies have found that prenatal SSRI
exposure results in blunted infant heart rate and facial responses to a painful stimulus (heel
prick) at 2 days and 2 months after birth (Oberlander et al., 2002; Oberlander et al., 2005)
and altered hypothalamic-pituitary-adrenal (HPA) axis function at 3 months of age
(Oberlander et al., 2008).
The most commonly reported consequence of prenatal SSRI exposure is poor
neonatal adaptation (Laine et al., 2003; Oberlander et al., 2004; Sanz et al., 2005; Simon et
al., 2002; Zeskind & Stephens, 2004). This is a self-limited phenomenon usually lasting a

40
few days and comprising respiratory difficulty, cyanosis on feeding and jitteriness. There are
several possible mechanisms for these effects. One is an SSRI-elicited alteration in fetal brain
development and/or maturation. This is suggested by the results from our previous studies,
which involved 8-day administration of fluoxetine to chronically instrumented pregnant ewes
in late gestation. The fetuses of these ewes exhibited acute changes in fetal cardio-respiratory
function (Morrison et al., 2002), behavior (Morrison et al., 2001) and in HPA axis function
(Morrison et al., 2004). In addition, there was a transient reduction of postnatal weight gain,
similar to the human findings. The altered fetal behavior comprised a reduction in REM sleep
and increase in quiet sleep, while the altered HPA axis function comprised an increase in the
fetal prepartum cortisol surge and an apparent increase in the sensitivity of the adrenal gland
to ACTH. These prenatal alterations could thus contribute to the poor neonatal adaptation, as
well as some of the other adverse postnatal findings associated with prenatal SSRI exposure
in the human. However, other explanations for the poor neonatal adaptation have been
proposed. These include an SSRI withdrawal phenomenon in the postnatal period or a
persistence of drug in the newborn, leading to drug-related toxicity, i.e., the serotonin
syndrome (Andrade et al., 2006; Hendrick et al., 2001; Isbister et al., 2001; Koren, 2004;
Ruchkin & Martin, 2005). Moreover, FX and SSRIs are also used in children and adolescents
for treatment of depression and there are raising concerns about the safety of this therapy
(Sharp & Hellings, 2006; Usala et al., 2008). Therefore, we suggest that fluoxetine injection
to the newborn lambs and later in life at different ages is a useful model to test the hypothesis
which cannot be implemented in human because of ethical issues. In addition, as the lambs
were to be studied from birth to one year of age, longitudinal studies on cardiovascular,

41
metabolic and behavioral, and endocrine functions could be studied in relation to age, sex
and FX exposure.
1.9 Objectives:
To determine the ontogenetic changes in various physiologic, behavioral and
endocrine functions in lambs from birth to 1 year of age in relation to sex and acute postnatal
fluoxetine administration. The study involved 2 groups of lambs: one group received a bolus
FX injection at approximately 2, 10, 30 days and at about 3, 6 and 12 months of age and a
control group that was studied at the same time periods. In addition, postnatal outcomes in
the prenatally FX-exposed lambs were investigated with 2 groups of lambs: one group
maternally exposed to FX for 14 days of late gestation and one control group with no
prenatally FX exposure.
Specific objectives:
1. To examine the behavioral functions in newborn and postnatal lambs.
2. To examine cardiovascular function including blood pressure, heart rate and heart
rate variability in newborn and postnatal lambs.
3. To determine blood gas, oxygen saturation, pH, glucose, lactate in newborn and
postnatal lambs.
4. To investigate HPA axis function by measuring plasma cortisol and ACTH
concentrations in newborn and postnatal lambs.

42
5. To determine the effects of acute fluoxetine administration on the above variables
at 2, 10 and 30 days and 3, 6 and 12 months of age.
6. To investigate the postnatal outcomes in the lambs antenatally exposed to FX in
late gestation.
1.10 Hypotheses:
1. The major postnatal changes in behavior, cardiovascular and HPA axis
functions in postnatal lambs will be temporally related to weaning.
2. FX injection will cause transiently agitation in the newborn and postnatal
lambs, disrupt their feeding activities and thus affect their ultradian and circadian rhythms.
3. FX injection in the newborn lambs will transiently decrease their heart rate
and increase their arterial pressure, changing their heart rate variability.
4. FX injection will cause transient hypoxemia with the decrease in blood
oxygen saturation and Po2.
5. FX injection will alter the activity of HPA axis and lead to an increase in
cortisol and ACTH plasma levels.
6. Antenatally FX-exposed lambs will have adverse outcomes: lower birth
weight, and poor neonatal adaptation.

43
2. Materials and Methods
All procedures conducted on the animals were approved by the UBC Committee on
Animal Care and conformed to the guidelines of the Canadian Council on Animal Care. This
chapter describes the general materials and methods for Chapter 3 to 5
2.1.1 Animals:
Sheep were bred following a protocol in our lab to provide us with lambs for studying
all year round. The estrous cycle was synchronized via progestin containing vaginal pessaries
(Pharmplex, Milton NSW, Australia) for 14 days. Ovulation was then induced with an
intramuscular injection of pregnant mare serum gonadotropin (PMSG) (Wyeth, Ontario,
Canada). Ewes were then placed with a ram for natural mating. Subsequently, conception
was confirmed by measurement of plasma progesterone by the Gynecologic Endocrinology
laboratory in the Division of Reproductive Endocrinology and Infertility, UBC at 19 days
after pessary removal and gonadotropin injection. An ultrasound was performed at ~90 days
gestation to confirm a successful pregnancy. Pregnant ewes (Dorset/Suffolk) were
transported to the Child & Family Research Institute from Three Gates Farm, Gabriola
Island, at least 3 weeks before their due date (term = 147 days) to allow them to acclimate
with the new environment before their deliveries. They were housed singly in 2.4 x 1.2 m
pens or in groups of 2 in 2.4 x 2.4 m pens. Sheep were kept under a 12 hour light: 12 hour
dark cycle with scheduled lights on at 6:00 am and off at 6:00pm. The feeding regimen
comprised 0.6 kg grain and 0.7 kg local hay at 0730 am and 0.7 kg local and 0.7 kg alfalfa
hay at ~1430 pm. The grain ration was increased to 0.8 kg after delivery. Water was available
ad lib. Signs of imminent parturition in the ewe (bilateral concave flanks, restlessness,

44
pawing at the pen floor, moving in circles and looking toward the back) and the lambs’ birth
were revealed by digital video recording (DVR) from infrared cameras which will be
described in more detail in Section 2.2.5 (DVR and behavioral evaluation). Ewes were
allowed to deliver without assistance except in cases of malposition of the fetus. After the
lambs were born, they were dried with a towel, sexed, weighed and had an actiwatch placed
around their neck (more details about actiwatch in Section 2.2.6: Actiwatch/Rest activity
monitoring). Surgeries were conducted on the lambs on the first day after birth. Lambs were
housed in the same pen with their mothers until the weaning period. The study involved 2
groups of lambs: one group received a bolus fluoxetine injection at ~ 2, 10, 30 days and at ~
3, 6 and 12 months of age and a control group that was studied at the same time periods.
2.1.2 Surgical Preparation:
Surgeries were done on the lambs on postnatal day 1.5 ± 0.1 (for those who started
the day 3 experiment) or on postnatal day 9 ± 1.1 (for those who started the approximately
day 10 experiment). Lamb was anesthetized with 4% isoflurane and 40% NO delivered via a
face mask and then intubated with a cuffed endotracheal tube (3.5 to 5.0 mm O.D, depending
on the size of the lamb). Anesthesia was maintained with 1.5 to 2% isoflurane, 40% NO in
oxygen. The lambs were ventilated at a rate of with a frequency and tidal volume appropriate
for their weight and postnatal age (Moss et al., 1995). The eyes were lubricated with Refresh
Eye Lube. Bupivacaine hydrochloride (0.5%) was infiltrated subcutaneously at the base of
the ear before an ear tag was applied and at the surgical sites. Meloxicam (0.2 mg/kg) was
also given subcutaneously for analgesia and 250 mg ampicillin intramuscularly for antibiotic
prophylaxis. The lamb was placed on a heating pad during the surgery and a thermistor rectal
probe connected to a Yellow Springs Instruments Tele-Thermometer was used to monitor the

45
body temperature continuously. Heart rate was monitored intermittently via auscultation or
later in the study with a pulse oximeter (Nonin Medical Inc, Plymouth, MN, USA). Aseptic
surgical techniques were employed. All surgical instruments were autoclaved before surgery.
The skin area to be operated on the neck was shaved and washed with antiseptic soap, then
sprayed with 70% alcohol and Proviodine solution (USP 10%) with the combination repeated
3 times. A sterile surgical drape was used to cover the lamb with the surgical opening
positioned appropriately.
Access to the carotid artery and jugular vein was gained via an oblique transverse
skin incision in the neck from the sternomastoid muscle (from ~ 1cm lateral to just above the
jugular vein). Heparin-bonded 3.5 or 5 French polyurethane catheters (Instech Solomon,
Plymouth Meeting, PA) were filled with sterile heparinized 0.9% NaCl solution (12U
heparin/ml) and were implanted into the right carotid artery and jugular vein. The catheters
were secured in place with 2-0 silk non-reabsorbable sutures and then the catheters were
tunneled subcutaneously and exteriorized through a small skin incision at the dorsal neck.
These catheters were available for drug administration and blood sampling in the subsequent
series of experiments, while the arterial catheter was employed to measure arterial pressure.
The transverse and dorsal neck skin incisions were closed with 2-0 Polysorb reabsorbable
(Syneture, Canada) interrupted sutures. The skin and the outer surfaces of the catheters were
then cleaned with 70% alcohol. Then the distal ends of the catheters were sealed with sterile,
metal pins and the catheters were placed in a small zip-lock bag. The skin incisions were
covered with sterile gauze pads, over which was placed in a zip lock bag into which the
catheters were placed. An elastic bandage was then placed loosely around the neck and
secured with a safety pin.

46
To prevent the risk of fly strike when the lamb gets older, all newborn lambs also had
their tails docked. The section of the tail to be cut was first shaved, disinfected, infiltrated
with 0.5% bupivicaine and then cut and cauterized using a Supervet Tail Dock
Cutter/Cauterizing Instrument (Syrvet Inc., Iowa, USA). Following this, the skin margins of
the tail stump were opposed using a continuous 2-0 Polysorb suture.
Lambs were kept in the operating room until they could breathe spontaneously and
were conscious. They were then returned to their mother. The total surgery time from
removing the lamb from its mother to returning the lamb post-surgery averaged 2.2±0.01 h.
The lambs were weighed daily in the morning. Catheters were flushed daily with sterile
heparinized 0.9% NaCl solution (12U heparin/ml) to maintain their functioning until the last
experiment (1 year of age). If catheters stopped functioning, another surgery was performed
to implant other appropriate catheters at least 2 days before the experiments were started.
2.1.3 Experimental Protocol:
The experiments on the lambs began on the day following surgery. A sterile silicone
urinary catheter was placed in the urethra of the female lambs and secured with tissue
adhesive and a 2-0 silk suture tied to the wool in the groin area on the day before FX
experiments. Since in male lambs, the urethra makes a hairpin turn, it is impossible to insert a
urethral catheter. A strap-like urine collection device, which was successfully developed by
Tim Chow, another PhD candidate in our lab, was placed on the lamb’s penis, wrapped
around the abdomen and tied securely on their back about 30 minutes before the FX
experiments. This served for urine collection mainly for Tim Chow’s project which involves
developmental fluoxetine pharmacokinetics and metabolism. The urine volume data was also

47
used in chapter 3 to determine the development of urine flow rate in the lambs with
advancing age. For the experiments on the 2, 10 and 30 day old lambs, two experiments were
carried out on the same day, starting at 8:30am for the first lamb and 11:30am for the second.
The lambs were placed in a cloth sling set up in the pen that contained the lamb’s mother or
the biggest pen. The lambs were suspended above the floor in the sling and normally lay
quietly or slept. They were only in the sling for the first two and a half hours of the
experiment, and then were returned to their mothers, so as not to disrupt the normal suckling
regimen. Lambs at 10 days and 1 month of age were bleating and jumping sometimes
especially toward the end of the 2.5h duration. For the experiments conducted at 3 months
and beyond, the lambs were placed in a small monitoring pen (120 X 65 cm) set up in one
corner of the holding pen. They remained in this pen for the duration of each experiment
(24h for the control experiment and 72h for the FX experiment) and had access to food and
water. Since lambs started having solid food around 2 months old, experiments on lamb at 3
months and above were started at 9:00am (30 min after lambs finished their feeding) to
eliminate the affect of eating on changes in heart rate and blood pressure (Jones, Langille,
Frise, & Adamson, 1993).
Each experiment involved i.v administration of sterile water (0.1ml/kg, equal to the
volume of 10mg/ml FX given in FX experiment) or fluoxetine (1mg/kg) followed by serial
collection of arterial blood samples (~ 3 ml) at -30, -15, 5, 15, 30, 45 and 60 minutes and 2,
4, 6, 9, 12, 24, 36, 48, 60 and 72 h (more specifics in table 2). The total volume of blood
collected (42ml) represents ~ 8% of the total blood volume in the newborn lamb and much
less in the older lambs.

48
FX Blood gases ACTH, cortisol -30min -15min SW/FX i.v 5min 15min 30min 60min 2h 4h 6h 9h 12h 24h 36h 48h 60h 72h
Table 2.1: Time points at which blood samples were collected for arterial blood gases, FX, ACTH and cortisol measurement. SW = sterile water.
The symbols indicate when the samples were taken for the corresponding variables
2.1.4 Sample Collection:
Blood samples for hormone and fluoxetine measurements were collected through the
carotid artery catheter and placed in a 4ml EDTA K2 and Lithium Heparin (68USP units)
vacutainer tubes (BD Vacutainer, NJ, USA) and centrifuged for 20 min at 4000 rpm in a
Centrifuge 5810R (Eppendorf, ON, Canada). Plasma was then transferred to polyvinyl
Eppendrorf tubes for hormone samples and glass tubes for fluoxetine samples and stored at -
20oC until analysis. Arterial pressure was obtained from the arterial catheter using a
disposable pressure transducer and Powerlab data acquisition system, which also calculated a
heart rate from the pulse pressure. The ECG was recorded using surface neonatal ECG

49
electrodes and the Powerlab Heart Rate Variability Module was used to assess heart rate
variability (HRV).
Blood samples were also collected before the surgery, on postnatal day 2.3 ± 0.2 (n =
11), for ACTH and cortisol measurements. Lambs were taken to the operating room and their
wool along the jugular vein was shaved. The skin was disinfected with alcohol 70% and
applied with local anesthetic Xylocain 2%. Approximately 2ml of blood was taken to the
EDTA K2 and Lithium Heparin (68USP units) vacutainer tubes via venipuncture. Lambs
usually slept while blood was collected and were returned to their mother immediately.
Samples were centrifuged and plasma was transferred and stored as mentioned above.
2.1.5 Digital Video Recording And Behavioral Evaluation:
One infrared video camera were installed on the wall of each pen in the sheep holding
room and connected to a digital video recording system on a computer in the lab, which was
directly adjacent to the sheep room. Another mobile infrared camera was put outside the pen
or above the monitoring pen to allow for the recording and observations of the lambs from a
direct view, as the lambs sometimes could not be observed by the fixed cameras because of
the ewe and lamb’s position within the pen. On the day of experiment the mobile camera was
also be used to record behavioral changes immediately after FX administration. The infrared
function allowed the recording and monitoring the ewe and lamb’s behavior at night without
interrupting their circadian rhythm. These cameras could also be accessed remotely from
home and the video records could be analyzed off-line.
Lamb’s activities were categorized into lying down sleep, lying down awake,
standing (still or moving), walking, suckling attempt, successful suckling and playing.

50
Playing activities included running, jumping, climbing on their mother’s back, nudging their
mother, nosing and playing with hay. These activities were collected for the dominant
activity in 15 second epochs. The collected activities from the DVR were then matched with
the actiwatch data (see below) to obtain an equivalent score from the actiwatch record for the
activity identified from DVR in the same epoch. These scores were used to determine the
intensity of each kind of activity. After each type of activity was identified as mentioned
above from DVR observations, only standing activity was further analyzed using a cut-off
point of 60 on actiwatch score to separate standing still from standing moving. In other
words, standing still was defined as standing activity from DVR observation and
corresponding actiwatch score lower than 60. Similarly, standing mobile was standing
activity from DVR and corresponding actiwatch score equal or greater than 60. The
percentage of time per 24 hour lambs spent for each activity was also calculated.
2.1.6 Actiwatch Rest/Activity Monitoring:
An Actiwatch Activity Monitor (AW64, Mini Mitter Inc., Bend, OR, USA), which is
a small wrist watch sized device, was enclosed in a protective metal case and placed around
the neck of the lamb with a dog collar from the first day after birth until at least 90 days after
birth to measure rest-activity cycles and sleep patterns. The actiwatch contains an
omnidirectional accelerometer which detects and stores the occurrence and intensity of any
movement with a force of at least 0.01g. The activity data was stored in 15s epochs and
downloaded every 10 days using an Actiwatch reader and analyzed using an Actiware-Sleep
version 3.5 software (Mini Mitter). The analyzed variables included total daily activity score
(total number of movement counts in each 24h period), active bouts/day (= mean activity
score, i.e magnitude of activity per epoch), mean movement score in active (the average

51
number of movement counts during bouts of activity), episode duration (= moving minutes/
mean activity score), % time moving and % time immobile.
The ultradian and circadian rhythms in the lambs were determined using Fast Fourier
Transformation (FFT) analysis from the Actiware software. The four most prominent
frequencies in the FFT analysis were selected.
2.1.7 Powerlab Data Acquisition System:
The lamb’s carotid artery catheter was connected to a disposable pressure transducer
(DTXTM Plus TNF-R; Becton Dickinson, Singapore), which was connected to a polygraph
recorder (model TA-4000, Gould Instrument Systems Inc, Valley View, OH) and to a
Powerlab data acquisition system (AD instrument, Mountain View, CA, USA) to
continuously record heart rate and arterial pressure throughout the first 2h of the experiments
in 4, 10 and 30 day old lambs and for the entire 24h or 72h of the experiments at 3, 6 and 12
month old lambs. The transducer was kept on the platform which was adjusted to the lambs’
heart level when they stood or were in the sling. Three ECG snap electrodes were attached to
the skin on the back and bilaterally on the chest and secured in place by adhesive bandages.
During the experiments these electrodes were connected to the Powerlab system by three
snap leads to record the electrocardiogram. The Powerlab Heart Rate Variability Module was
used to assess and analyze heart rate variability (HRV) including estimates of
parasympathetic and sympathetic tone.

52
2.1.8 Arterial Blood Gases Analysis:
Blood samples were collected at -15, 5, 15, 30, 60 and 120 minutes to measure acid-
base balance (pH), base excess (BE), bicarbonate (HCO3-), arterial blood gases (pO2, pCO2),
electrolyte (Na+, K+ and iCa2+) using an I-STAT-1 analyzer (Abbott, ON, Canada) and CG8+
cartridges (Abaxis, Union City, CA, USA). Body temperature was measured continuously
with the thermistor rectal probe (YSI Instruments, Yellow Springs, OH) over the first 2h of
experiment or later by a regular digital thermometer and blood gas results were corrected
with the appropriate temperature when the sample was collected. Hemoglobin concentration
and oxygen saturation were measured with a Radiometer OSM3 Hemoximeter (Radiometer,
Copenhagen, Denmark). Glucose and lactate were determined with a YSI glucose-lactate
analyzer (YSI Inc., Yellow Springs, OH).
2.1.9 Adrenocorticotropin Hormone Assay:
ACTH level was measured using a commercially available human ACTH ELISA kit
(Calbiotech Inc., CA, USA) that detects ACTH1-24. The kit has previously been validated for
use in sheep (You et al., 2008). 200µl of plasma was added together with 25 µl of
biotinylated antibody and 25µl of enzyme labeled antibody into each well of the microplate.
The microplate then was covered with aluminum foil to prevent light exposure and was
shaken at 170 ±10 rpm for 4.5 hour at room temperature. After the remaining antibodies were
washed with wash solution five times, 150 µl of TMB substrate was added into each well and
the plates shaken at the above speed for 30 minutes at room temperature. 100 µl of stopping
solution was then added to terminate the reaction. ACTH level was determined by an ELISA
reader (initially by Dynex Technologies and later by Wallac Victor3V 1420) set at 450nm.

53
Each sample was measured in duplicate. The sensitivity of the assay was < 1 pg/ml. The
intra- and interassay coefficients of variation were 7.27% and 7.08%, respectively.
2.1.10 Cortisol Assay:
Commercially available sheep cortisol ELISA kits (Calbiotech Inc., USA) was used
to determine cortisol concentrations. 40µl of sample in duplicate and 200µl of Cortisol
enzyme conjugate were added into each well coated with anti-cortisol monoclonal antibody
for competitive binding. The microplate was then incubated for 1 hour at room temperature
with shaking. The unbound hormone and conjugate was washed off with washing solution 3
times. 100µl of TBM substrate was added and incubated for 15 minutes in the same previous
condition. The reaction was brought to end by adding 50 µl of stop solution in each well. The
cortisol level was determined by ELISA reader (initially by Dynex Technologies and later by
Wallac Victor3V 1420) at 450nm. The sensitivity of the assay was 1.38 ng/ml. The intra- and
interassay coefficients of variation for low control (78 ± 3 ng/ml) were 10.7% and 3.2%, and
for high control (225 ± 5 ng/ml) were 7.2% and 6.7%, respectively.
2.1.11 FX and NFX assay:
FX and NFX concentrations were measured by Dr.Tim Chow from Dr. Wayne Riggs’ lab,
Faculty of Pharmaceutical Science, UBC using a validated liquid chromatography with
tandem mass spectrometry method (LC/MS/MS) (Chow et al., 2011). The sample volume
used for each enantiomer (R)-, (S)- fluoxetine and (R)-, (S)- norfluoxetine was 250 µl. The
limit of quantitation was 1 ng/ml.

54
2.1.12 Statistical Analysis:
All data were analyzed using the Sigma Plot 11 statistics software (Systat Software,
Inc; San Jose, CA, USA). A 2 way General Linear Model (GLM) for repeated measures
ANOVA or a three way ANOVA followed by the Bonferroni t-test for multiple comparisons
was performed to determine the effects of age, sex and FX treatment on the interested
variables. P < 0.05 defines significant differences. Data are presented as mean ± SEM. Other
statistical methods that were used in particular results chapters are described in those
chapters.

55
3. Postnatal Lamb Development and Sex Differences
3.1 Introduction:
The geometry of the circulation in the newborn alters remarkably at birth, due to the
loss of the umbilical circulation, onset of pulmonary ventilation and increase in pulmonary
blood flow. With the onset of air breathing, the pulmonary vascular resistance decreases and
the surface area for gas exchange increases. Hemodynamic changes result in the closure of
fetal circulatory shunts – formamen ovale and ductus arteriosus. At birth, cardiovascular
function also alters significantly with an increase in arterial pressure, heart rate and cardiac
output. Much research has been done regarding the cardiovascular and hemodynamic
changes immediately following birth. (Klopfenstein & Rudolph, 1978; Lister et al., 1979;
Minoura & Gilbert, 1987; Woods et al., 1977) However, little has been known about these
physiological developments (heart rate, blood pressure, HRV) in lambs after the initial
postnatal period. In lambs, heart rate and blood pressure increase with feeding (Jones et al.,
1993) but whether there is any effect of weaning on the development of these variables is
unknown. Studies in goats found an elevated cortisol and B-endorphin concentration during
suckling in kids but no significant changes in heart rate or blood pressure related to suckling
(von Walter et al., 2010).
Heart rate variability (HRV) represents the oscillation of the intervals between
consecutive heart beats. It is in part determined by influences of the autonomic nervous
system on the heart. Since HRV is a non-invasive method to measure the autonomic
impulses, monitoring HRV variables are used clinically as an evaluation of fetal well being
in the human fetus (Alfirevic et al., 2006; Graatsma et al., 2009; Maso et al., 2012; Van

56
Leeuwen et al., 1999) and also in the neonate in neonatal intensive care (Fairchild & O'Shea,
2010). They are also used in a number of conditions in the adult, including coronary artery
disease, systemic hypertension and myocardial infarction (Charkoudian & Rabbitts, 2009;
Huikuri, 1995; Pagani & Lucini, 2001). However, there is less information about changes in
HRV characteristics during development, including the changes which occur around
weaning. Even though high HRV usually indicates a good adaptation of an effective
autonomic system in a healthy individual while low HRV often indicates the presence of
physiological malformation, the threshold is age-dependent. HRV has been shown to be
lower in healthy women than in men (Saleem et al., 2012); however, it is not known if the
same situation exists in sheep. There are established data on the differences in the prevalence,
characteristics and response to treatment of cardiovascular diseases (Dunlay & Roger, 2012).
After adjustments for the increased longevity in women, age-specific stroke incidence is
generally higher in men than women (Katsiki et al., 2011). Gender differences in cardiac
autonomic control might occur early in life and could be one of the potential mechanisms for
the observed differences in cardiovascular disease between males and females.
Birth also results in changes in the surrounding environment. The newborn has to
adapt with and become entrained to the circadian rhythm of the light dark cycle. Melatonin,
which is secreted from the suprachiasmatic nucleus under the influence of light from the
environment, is not present in human infants until 9-15 weeks of age (Kennaway et al., 1996)
and in lambs at 3-4 weeks after birth (Kennaway et al., 1996; Nowak et al., 1990). A
disrupted circadian rhythm may play a role in the pathogenesis of certain illnesses such as
coronary heart disease, breast cancer, gastrointestinal disturbances, sudden infant death
syndrome even though the mechanisms involved have not yet been established (Mosendane

57
et al., 2008; Lohr & Siegmund, 1999). In a previous study using actigraphy in newborn
lambs, our lab found a short (~ 1 h) activity rhythm not present in adult sheep, in which the
circadian rhythm was the most dominant. Only the lactating ewes had a short (~1 h) rhythm
as the second, third or fourth most dominant rhythms. Also, the total daily activity level in
the lambs over the first ~3 weeks was significantly higher than in adult sheep (Rurak et al.,
2008). However, it is still not yet clear when the short term ultradian rhythm in the lamb
disappears and when the circadian rhythm becomes dominant as in the adult sheep. In the
human, babies who are breast-fed for the first 2 years of life wake up more often at night and
have shorter sleep bouts instead of having a long unbroken night sleep (Elias et al., 1986).
Thus the short 1 h rhythm observed in the lambs may be the result of suckling activity.
3.2 Methods:
3.2.1 Experimental Protocol:
As described in chapter 2, the lambs in this study were used in a longitudinal study of
the disposition of the antidepressant, fluoxetine (FX), from birth to one year of age. All the
data used in this chapter was from the values of control experiments with sterile water
injection or from the control time point of FX experiments (i.e: 30 mins before FX injection)
except for the urine volume data and rest-activity cycles. The animal housing, surgical,
experimental and analytical methods are described in Chapter 2.
3.2.2 Statistical Analysis:
A 2 way General Linear Model (GLM) for repeated measures ANOVA followed by
the Bonferroni t-test was used to determine the effect of age and sex in cardiovascular,

58
metabolic and endocrine functions. The relationship between postnatal age and the
cardiovascular, metabolic, endocrine and urinary variable was analyzed using piecewise
linear regression with 2 elements. The area under the curve (AUC) of the activity variables
(total activity score, mean activity score, mean score in active, episode duration, % time
mobile, % time immobile, circadian and ultradian rhythms) was calculated for each lamb and
analyzed by 2 way ANOVA to determine the effect of age and gender in their rest-activity
cycle.
3.3 Results:
For the newborn lambs, the average age of the lambs on the control and FX
experiments is 3.2 ± 0.5 and 4 ± 0.4 postnatal days, respectively
3.3.1 The Development Of Physiological Variables And Sex Differences In
Development From The Newborn Period Up To 1 Year Of Age:
The lambs’ heart rate fell dramatically over the first month of life from 220.4 ± 6.3
bpm on postnatal day 3 to 155.2 ± 8 bpm at 1 month of age. A further decrease in heart rate
occurred thereafter to 94 ± 3.4bpm at 1 year of age. There was a significant difference in the
values between 10 days, 1 month, 3 months and 6 months. There was also a sex difference in
heart rate development. At 6 months, heart rate in female lambs was significantly lower than
male lambs (96 ± 7.8 bpm in female vs 115 ± 7.7 bpm in male, (p = 0.016) while the
situation seemed to be reversed at 1 month of age (167 ± 10.5 bpm in female and 155 ± 8bpm
in male) even though the difference was not significant at 1 month. However, they both
reached a similar heart rate at 1 year of age (97 ± 3.9 bpm in female and 93 ± 5.2bpm in
male). Female lambs tended to reach the adult heart rate value earlier than males, at 6 months

59
of age. Arterial pressure development was similar in male and female lambs. Arterial
pressure gradually increased from 66 ± 1.5 mmHg on day 3 to 82.9 ± 1.1 mmHg at 1 year of
age and the value was significantly different at 1 month and 1 year-old against 3 days of age.
(Fig. 3.1)
In terms of heart rate variability, a sex difference was only found in Low Frequency
(LF) power. At 3 day-old, LF power in male was significantly higher than that in female
(52.4 ± 6.2 nu in male vs 25.7 ± 6.5 nu in female, p = 0.036). In female, LF power increased
to peak at 10 day-old (61.5 ± 5 nu) and then decreased to a plateau from 3 months to 1 year
of age (range: 38 ± 6.8 – 43.4 ± 4.9nu). In males, LF power decreased from 52.4 ± 6.2 nu on
day 4 to a plateau between 4 and 12 weeks (range: 35.8 ± 3.8 – 46.9 ± 5.9 nu ), then
increased at 6 months (56.1 ± 4.2 nu), followed by a decrease at 1 year of age (Fig. 3.3A).
The time domain variables of HRV (including SDNN, SD Delta NN, RMSSD) were
significantly different between the 3-day, 10 day-old and the remaining ages (1, 3, 6 months
and 1 year old) with the lower values at 3 days of age (22 ± 1.7 msec for SDNN, 19.4 ± 3.4
for SD Delta NN and RMSSD) and higher values at 1 year of age (102.8 ± 12.4 for SDNN,
80.5 ± 9.8 for SD Delta NN and RMSSD) (Fig. 3.2). High Frequency (HF) power tended to
increase from 18.4 ± 2 nu on day 3 to 28.3 ± 2.2 nu at 1 year of age with the lowest value at
10 days of age (16.4 ± 1 nu). Since HF power did not change much over the time, changes in
LF/HF ratio were due mainly to LF power alterations. The LF/ HF ratio peaked at day 10
(3.7 ± 0.48) and was significantly higher than that at 3 months (1.8 ± 0.3) and 1 year of age
(1.6 ± 0.2). (Fig. 3.3)
Fig. 3.4 show an inverse relationship between SDNN (msec) and heart rate in the
lambs from 3 days to 1 year of age with r2 = 0.55, p < 0.001

60
FIG. 3.1: Postnatal lamb arterial pressure (A) and heart rate (B) in all (open bar), male (black bar) and female (gray bar) lambs at 3 days (n = 14), 10 days (n = 18), 1 month (n = 15), 3 months (n = 17), 6 months (n = 11) and 1 year (n = 10) of age. Different letters (a,b,c and d) indicate significant differences between ages. Asterisk indicates significant difference between male and female, (p < 0.05). 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

61
FIG. 3.2: Time domain of heart rate variability: SDNN (standard deviation of NN interval) (A), SD Delta NN (Standard Deviation of the differences between adjacent NN intervals) (B), RMSSD (Square root of the mean of the squared differences between adjacent NN intervals) (C) in lambs at 3 days (n = 11), 10 days (n = 18), 1 month (n = 15), 3 months (n = 17), 6 months ( n = 11) and 1 year ( n = 10) of age. Different letters (a,b, and c) indicate significant differences between ages, (p < 0.05). 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year
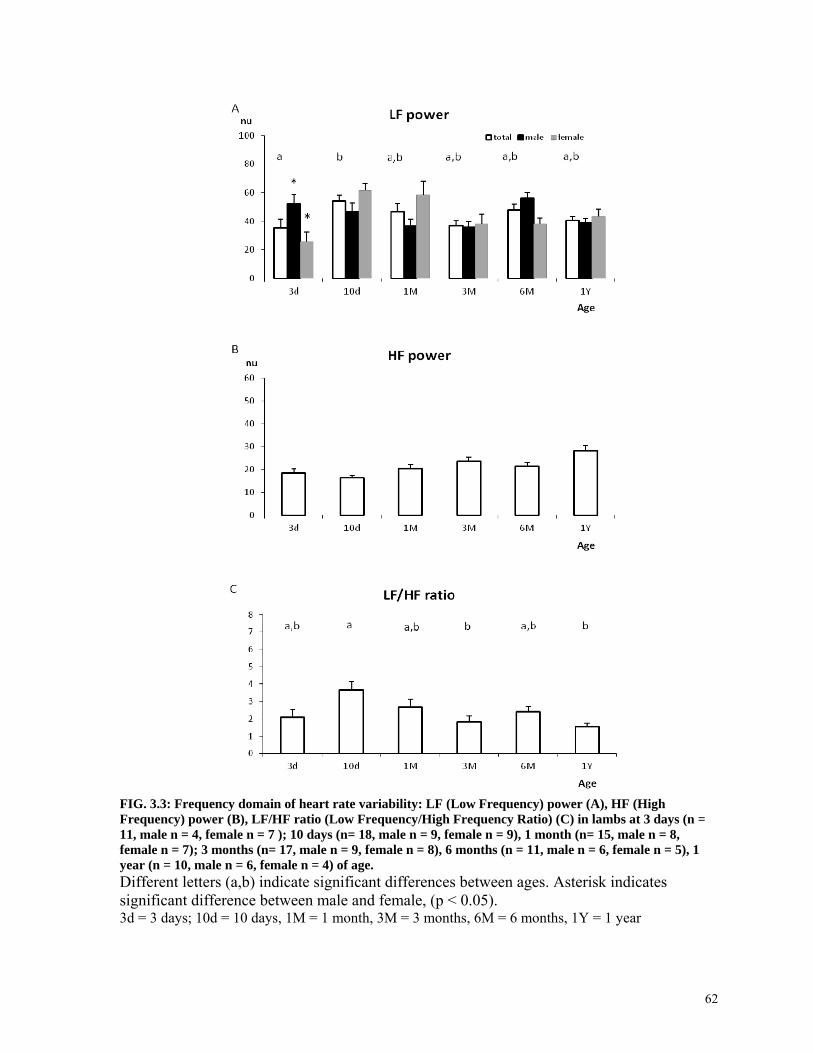
62
FIG. 3.3: Frequency domain of heart rate variability: LF (Low Frequency) power (A), HF (High Frequency) power (B), LF/HF ratio (Low Frequency/High Frequency Ratio) (C) in lambs at 3 days (n = 11, male n = 4, female n = 7 ); 10 days (n= 18, male n = 9, female n = 9), 1 month (n= 15, male n = 8, female n = 7); 3 months (n= 17, male n = 9, female n = 8), 6 months (n = 11, male n = 6, female n = 5), 1 year (n = 10, male n = 6, female n = 4) of age. Different letters (a,b) indicate significant differences between ages. Asterisk indicates significant difference between male and female, (p < 0.05). 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

63
FIG. 3.4: The relationship between SDNN (msec) and heart rate (bpm) in the lambs from 3 day to 1 year of age
Among the blood chemistry variables collected (glucose, lactate, O2 sat, pO2, pCO2,
pH, BE, HCO3, Hb), only the pH value at day 3 was different between male and female (Fig.
3.5), with the value in males (7.379 ± 0.0016) being significantly lower than in females
(7.457 ± 0.018), p < 0.0001). Following birth, Po2 and pH increased from 86.6 ± 3.4 mmHg
and 7.421 ± 0.016 on postnatal day 3 to 110.7 ± 3.27mmHg and 7.489 ± 0.009 at 1 year of
age, while Pco2 decreased from 41.9 ± 1.01 mmHg on postnatal day 3 to 35.57 ±
0.653mmHg at 1 year of age, with the main changes occurring over the first 12 weeks. Base
excess tended to increase from 3 ± 1.019 mmol/l on postnatal day 3 to peak 5.647 ± 0.342
mmol/l at 3 months of age and slightly decrease thereafter. Both glucose and lactate
concentrations decreased from 5.39 ± 0.215 mmol/l and 1.028 ± 0.055 mmol/l on postnatal
day 3 to 2.91 ± 0.12 mmol/l and 0.527 ± 0.042 mmol/l at 1 year of age, respectively, again
with most of the change occurring in the first 12 weeks, although lactate decreased further at

64
6 months. Hemoglobin concentration decreased more rapidly, from 12.2 ± 0.34 g/l at 3 days
of age to low value 8.729 ± 0.418 g/l at week 4, and was relatively constant thereafter
(Fig.3.6-3.8). These suggested that the newborn lambs appeared to be more hypoxemic and
acidemic than the adult.
Urine flow rate (ml/min.kg) was based on the mean of urine flow rate at different
intervals of the FX experiment and was normalized for the weight of each lamb at the
corresponding age. Urine flow rates at 2 and 10 days of age were significantly higher than
the values at 3, 6 and 12 months of age. There was no sex difference in urine flow rate at any
age. (Fig. 3.9)
The cortisol concentration was not significantly different between day 3 (day of
control experiment) and ~ day 2.3 ± 0.2 before the surgery to implant the catheters for the
subsequent experimental sampling.
In fact, combining the mean cortisol values in the fetus lamb in late gestation from a
previous paper in our lab (Morrison et al., 2004) suggested the lambs’ prior surgery cortisol
value was consistent with the cortisol surge which usually occurs right before birth in the
sheep. After the day 3 experiment, cortisol concentrations at 10 days and 1 month gradually
decreased, but are significantly higher than the values at 3 and 6 months. At 3 months, the
cortisol concentration became stable through to 1 year of age. (Fig. 3.10) There were no
statistical significant changes in ACTH concentration from before surgery up to 1 year of
age. (Fig. 3.11)

65
FIG. 3.5: Mean pH value in total (open bar), male (black bar) and female (gray bar) lambs at 3 days (n = 13, male n = 6, female n = 7 ); 10 days (n= 17, male n = 8, female n = 9), 1 month (n= 15, male n = 8, female n = 7); 3 months (n= 17, male n = 9, female n = 8), 6 months (n = 10, male n = 5, female n = 5), 1 year (n = 10, male n = 6, female n = 4) of age. Different letters (a,b,c) indicate significant differences between ages. Asterisk indicates significant difference between male and female, (p < 0.05). The fetal value at 147 days of gestational age (at term) is taken from Rurak & Bessette, (2012) and was not included in the statistical test. 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

66
FIG. 3.6: Mean pCO2 (A), pO2 (B) and O2 sat (C) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. Different letters (a, b, c, d) indicate significant differences between ages, (p < 0.05). The Fetal values at 147 days of gestational age (at term) are taken from Rurak & Bessette, 2012 and was not included in the statistical test. 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

67
FIG. 3.7: Mean HCO3 (A), Base Excess (B) and Hemoglobin (C) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. Different letters (a,b)indicate significant differences between ages, (p < 0.05). The Fetal values at 147 days of gestational age (at term) are taken from Rurak & Bessette, 2012 and was not included in the statistical test 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

68
FIG. 3.8: Mean Glucose (A) and Lactate (B) in lambs at 3 days (n = 13); 10 days (n= 17), 1 month (n= 15); 3 months (n= 17); 6 months (n = 10) and 1 year (n = 10) of age. Different letters (a ≠ b ≠ c≠ d) indicate significant differences between ages, (p < 0.05). The Fetal values at 147 days of gestational age (at term) are taken from Rurak & Bessette, 2012 and was not included in the statistical test 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

69
FIG. 3.9: Mean urine flow rate in lambs at 3 days (n = 10); 10 days (n= 10), 1 month (n= 9); 3 months (n= 12); 6 months (n = 7) and 1 year (n = 10) of age. Different letters (a ≠ b) indicate significant differences between ages, (p < 0.05). Fetal value at 130-140 days of gestational age is for reference and was not included in the statistical test. The value is taken from (Turner et al., 2008) GA = gestational age, 3d = 3 days; 10d = 10 days, 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year

70
FIG. 3.10: Mean cortisol concentration in fetal lambs from preinfusion to infusion day 2 and from infusion day 5 to 8 adapted from Morrison et al, 2004 and in postnatal lambs prior surgery (n = 11), at 3 days (n = 14); 10 days (n= 12), 1 month (n= 12); 3 months (n= 12); 6 months (n = 8) and 1 year (n = 8) of age. Different letters (a ≠ b ≠ c ≠ d ≠ e ≠ f) indicate significant differences between ages, (p < 0.05)

71
FIG. 3.11: Mean ACTH concentration in fetal lambs from preinfusion to infusion day 2 and from infusion day 5 to 8 adapted from Morrison et al, 2004 and in postnatal lambs prior surgery (n = 11), at 3 days (n = 2); 10 days (n= 2), 1 month (n= 2); 3 months (n= 14); 6 months (n = 3) and 1 year (n = 4) of age.

72
Cortisol, ACTH rhythm in the newborn and postnatal lambs:
At 3 day, 3, 6 month and 1 year–old experiment, the cortisol level at 30min, 1h and
2h tend to be higher than that at other time points during the day but the differences were not
statistically significant. However at 10 days and 1 month, respectively the higher cortisol
value at 30 min (at 10:00am) (201 ±64.9; 145.6 ± 24.8 ng/ml), 1 h (at 10:30am) (209 ±76.6;
178.1 ± 35.6ng/ml), and 2 h (at 11:30 am) (231.8 ± 72.5; 183.5 ± 46 ng/ml) were
significantly different from that at 6 h (at 15:30 pm) (85.7 ± 25.6; 68.2 ± 13 ng/ml) and 12 h
(at 21:30 pm) (78.1 ± 17.3; 73.4 ± 19.8 ng/ml), respectively. (Fig. 3.12)
From 3 month onward, lambs were put in the monitoring pen during the whole period
of the experiment (4 days). The cortisol level did not change much over the sampling time
point during the day with placebo injection at 3 month, 6 month and 1 year-old experiment,
but there was a sex difference in cortisol concentration at 30 min (10:00 am), 1 h (10:30 am),
6 h (15:30 pm) and 12 h (21:30 pm) samples when the lambs were 1 year old. Cortisol levels
were siginificantly higher in males than females at 1 year of age (p = <0.001).
3.3.2 Neonatal Behavioral Development:
From the DVR observations on 35 lambs following birth, we found the average times
that it took for the lambs to first stand, walk and attempt to suckle were 33 ± 3, 45 ± 4 and 57
± 5 minutes, respectively. Our results are similar to what have been shown in the literature
(Dwyer, 2003; Fraser & Broom, 1997). However, we did not see any effects of sex or lamb
number on the time that lambs took to first stand, walk or suckle. Also, we did not see any
significant correlation between these first time activities and birth weight.

73
FIG.3.12: Cortisol concentration during the day in the lambs at 10 days, 1 month and 1 year of age. Samples were collected at -30, -15 min before sterile water injection and 30 min, 1,2,6 and 12 hour after the injection (10 days group, n= 9; 1 month group, n = 11; 1 year group, n = 8). Different letters (a ≠ b) indicates significant differences between the time points. Asterisk indicates significant differences between male and female

74
Activities on postnatal day 2-3 were observed from a smaller group of normal term
lambs (n= 11) who were not subjected for any experiments or surgery at this age. The results
showed lambs at this age spend most of the time at rest or asleep (~ 74.4%). Ewes of twin
and triplets appear more frequently to initiate suckling than their lambs at this age. In
general, only 33 ± 7% the number of bouts was initiated by the lamb itself. In terms of infant-
maternal bonding, we observed that lambs sleep close to either their mom or with their
siblings most of the time. Only one single female lamb showed less attachment to her
mother.
In addition, I compared the time spent on suckling on postnatal day 2-3 in single
(n=6), twin (n= 14) and triplet (n=3) lambs. The suckling time included both suckling
attempts and successful suckling. The triplets spent significantly more time suckling than the
singles and the twins. However, they gained less weight than the singleton and twins
although the differences were not significant (p = 0.064) (Fig. 3.13)
3.3.3 The Development Of Circadian And Ultradian Rhythms. The Ultradian 1 h
Rhythm Is Associated With Suckling:
Since during the period of the circadian rhythm study (i.e from birth to postnatal day
90), the lambs used in this study were also subjected to acute FX exposure intermittently at
approximately day 4, 10 and 30, it is important to note that the FX administration did not
result in any obvious physiological or behavioral changes in the lambs at day 4 and 10. Since
the activity was lower on FX day experiments in lambs at 1 month of age, those who were
subjected to the 1 month FX experiment were excluded from the regression analysis of this
circadian rhythm study. The results of FX administration are not included here.

75
The daily total activity score up to postnatal day 90 of the rest activity cycle is shown
in Fig. 3.14. There were no differences in the rest activity cycle development in male and
female lambs (data not shown). The development of four most dominant FFT rhythms for 21
lambs for 90 days after birth are shown in Fig. 3.15. The results indicate that in the newborn
the 1-2 hour ultradian rhythm is the most dominant. About 3 weeks later, the ~ 1 hour rhythm
disappears and the circadian rhythm becomes dominant. Subsequently, an 8 hour ultradian
rhythm becomes secondarily dominant. A third rhythm, a 6 hour ultradian rhythm also
developed at ~ 20 postnatal day, and by about postnatal day 65, this rhythm was consistently
the third or fourth most dominant rhythm. No sex differences were found in circadian and
ultradian rhythm development (data not shown).
The 1-2 h rhythm in the lambs, which was not observed in the adult sheep,
disappeared around 3 weeks after birth. At the same time, the total activity also decreased
and became stable (Fig. 3.15). Regression analysis of the day that the 1h rhythm disappeared
against the day that the lambs had the lowest activity during the first 90 postnatal days
indicated that there was a significant correlation between the ~ 1-2 h rhythm and total
activity (r² = 0.5375, p < 0.0001) (Fig. 3.15). In addition, the correlation between sucking
frequency from DVR observations and that from FFT in 21 individual lambs on postnatal day
2 and 3 was also significant (r² = 0.1947, p = 0.045) (Fig. 3.17). This suggests that the 1-2 h
rhythm only seen in the lambs is associated with suckling.
The correlation between the first day the 6h rhythm appeared and the first day lambs
ate significant amounts of hay was also statistically significantly (r² = 0.4973, p = 0.00107)
which indicated the link between their rumen development and the 6h rhythm (Fig. 3.16).

76
FIG. 3.13: Weight gain (A) and total suckling time (B) in singleton (n = 6), twin (n =14 for weight gain and n = 10 for total suckling time) and triplets (n = 3) in lambs on postnatal day 2-3 (N = 22). Asterisk indicates significant difference between singleton and triplets, (p < 0.05)

77
FIG. 3.14: Plots of daily total activity score (a), activity bouts/day (b), mean score in active period (c), mean episode duration (d), % time moving (e) and % time immobile (f) against postnatal age in lambs. (n=21)
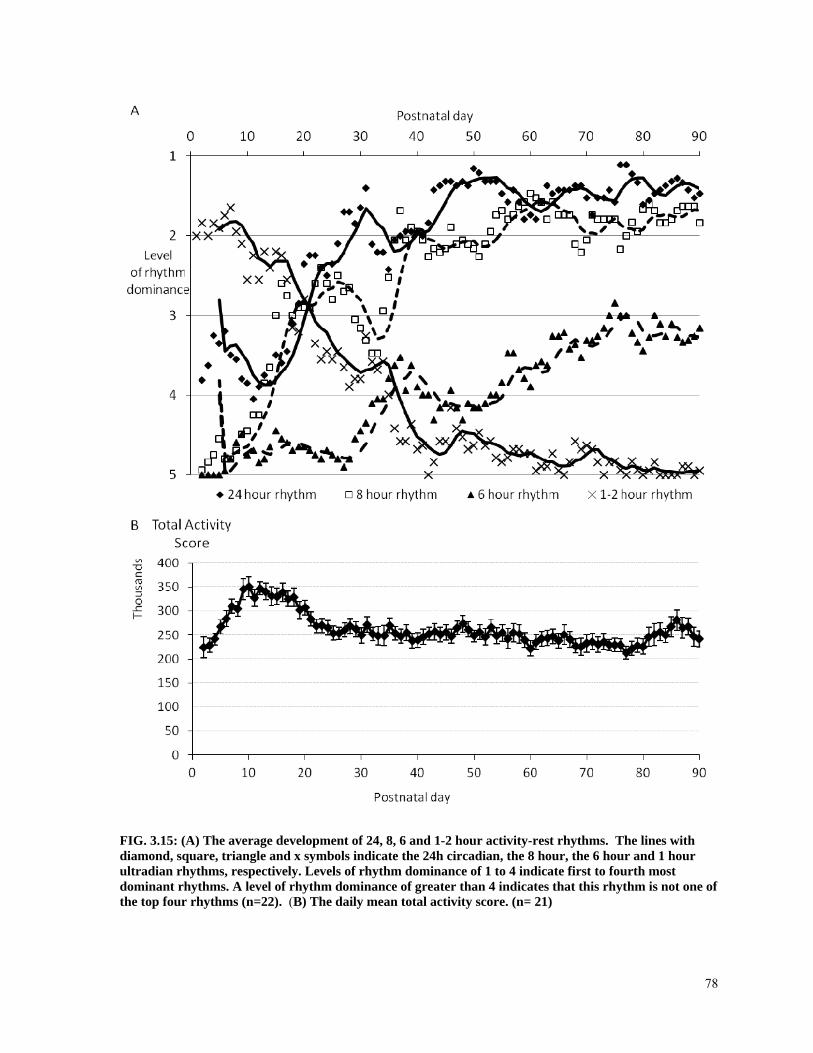
78
FIG. 3.15: (A) The average development of 24, 8, 6 and 1-2 hour activity-rest rhythms. The lines with diamond, square, triangle and x symbols indicate the 24h circadian, the 8 hour, the 6 hour and 1 hour ultradian rhythms, respectively. Levels of rhythm dominance of 1 to 4 indicate first to fourth most dominant rhythms. A level of rhythm dominance of greater than 4 indicates that this rhythm is not one of the top four rhythms (n=22). (B) The daily mean total activity score. (n= 21)

79
FIG. 3.16: (A). Relationship between the day that the 1h rhythm disappears and the day of the lowest total activity (n = 21). (B) Relationship between the day that the 6h rhythm disappears and the first day lamb eating hay (n = 21)

80
FIG. 3.17: Relationship between suckling frequency on postnatal day 2-3 from DVR observation and calculated from the actiwatch FFT (n = 21)
The growth rate was significantly different between the 1st and 2nd of the 25-day
intervals and the 3rd, 4th of the 25-day intervals (p< 0.001). The slope of the daily weight
gain decreased from 279.0 ± 1.86 g/d for the first 25 days to 209.3 ± 3.03, 142.7 ± 1.95,
143.6 ±7.54 and 122.1 ± 13.2 g/d for the subsequent 25 day intervals.
3.3.4 Weaning And Changes In Physiological Variables:
In our study, the time of weaning was defined as a window between when lambs
started to first consume solid food (mostly hay) to when the 1h rhythm due to suckling
disappeared (i.e., from 29.6 ± 1.1 to 50.9 ± 4.8 days). We have used piecewise linear
regression with 2 elements to determine the break point which is the point where the slope of
the regression curve for the variable versus age plot changed abruptly. The break points of
the significant changes in heart rate, SDNN, glucose, pO2 and urine flow rate were at 41.1 ±

81
5.5 (p <0.0001); 51.6 ± 13.3 (p = 0.0002); 61.1 ± 7.9 (p <0.0001); 52.3 ± 22.3 (p = 0.0213)
and 38.4 ± 8.7 (p < 0.001) days of age, equivalent to 5.87 ± 0.78; 7.37 ± 1.9; 8.72 ± 1.12;
7.47 ± 3.18 and 5.48 ± 1.2 weeks of age, respectively. However, arterial pressure, LF/HF
ratio, Lactate, Hb and cortisol changed significantly with the break points at 16 ± 5 (p =
0.002); 5 ± 2 (p = 0.014); 6.5 ± 2.4 (p = 0.0084); 12 ± 1.7637 (p <0.001) and 12.2 ± 2.4 (p <
0.0001) days of age, equivalent to 2.28 ± 0.71; 0.71 ± 0.28; 0.92 ± 0.34; 1.71 ± 0.25 and 1.74
± 0.34 weeks of age, respectively (Table 3.2). Taken together, the changes in heart rate,
SDNN, glucose, pO2 and urine flow rate occurred around the window of weaning time,
suggesting a relationship between weaning and these physiological changes.

82
Variables Mean age (days) ± SE 1h rhythm lost 50.9 ± 4.8
Lowest actiwatch total activity 56.6 ± 4 Solid food was first consumed 29.6 ± 1.1
Table 3.1: The time window of weaning in lambs was defined as when lambs started to first consume solid food to when 1h rhythm due to suckling disappeared
Variables Breakpoint (days) ± SE (p) r2 Heart rate (bpm) 41.1 ± 5.5 (<0.0001) 0.6914
Arterial pressure (mmHg) 16 ± 5 (0.002) 0.2345 SDNN (msec) 51.6 ± 13.3 (0.0002) 0.473
SD Delta NN (msec) 60 ± 26.2 (0.0247) 0.3296 RMSSD 60 ± 26.2 (0.0247) 0.3296
LF power (nu) 3.6 ± 0.54 (<0.0001) 0.1578 HF power (nu) 2 ± α (1.0) 0.2588
LF/HF ratio 5 ± 2 (0.014) 0.1417 Glucose (mmol/l) 61.1 ± 7.9(<0.0001) 0.8184 Lactate (mmol/l) 6.5 ± 2.4 (0.0084) 0.2375
O2 sat (%) 99 ± 29.5 (0.0012) 0.3133 pO2 (mmHg) 52.3 ± 22.3 (0.0213) 0.2738
pCO2 (mmHg) 12 ± 6.4 (0.066) 0.1119 pH 55.5 ± 80.7 (0.494) 0.0223
HCO3 (mmol/l) 99 ± 68.4 (0.1522) 0.0426 BE (mmol/l) 73.6 ± 38.5 (0.0595) 0.1609
Hb (g/l) 12 ± 1.7637 (<0.001) 0.2814 Cortisol (ng/ml) 12.2 ± 2.4 (<0.0001) 0.414
Urine flow rate (ml/min/kg) 38.4 ± 8.7 (<0.0001) 0.5 Table 3.2: Break points of cardiovascular, metabolic, endocrine and renal variables. Break point is the point (day of age) where the slope of the plots of the variable versus age changes.

83
3.4 Discussion:
The main findings of this study are that there were sex differences in pH value and LF
power at 3 days of age and in heart rate at 6 months of age. Cardiovascular function in the
lambs changed over time with a decrease in heart rate and increase in arterial pressure
together with the maturation of the autonomic system (i.e., increase in the time domain of
HRV and a reduced influence of the sympathetic system on the heart as indicated by a
decrease of LF power and therefore LF/HF ratio). The relation between HRV frequency
domain variables and autonomic function will be discussed further below.
In addition, we also found the acid base balance shifted toward the alkaline range
over age with a decline in pCO2, and increase in pH. The efficiency of pulmonary oxygen
uptake indicated by arterial PO2 was progressively increased after birth until 3 months of ages
and slowly increased thereafter which suggested that a process of improvements in
pulmonary gas exchange still continued at least for the first 3 months of life. Glucose, lactate
concentration, urine flow rate and Hb decreased with increasing age, while a slight anemia
occurred at 1 month of age. The cortisol concentration was attenuated a few days before
birth, decreased rapidly for the first 10 days of age and gradually thereafter.
The 1h rhythm that exists in the lambs but not in the adult sheep is associated with
suckling. The disappearance of 1h rhythm was associated with the changes in growth rate
and the decline in total activity. The break points of some physiological variables (including
changes in heart rate, SDNN, glucose, pO2 and urine flow rate) occurred around the window
of weaning time suggests the effects of weaning and thus the beginning of solid food intake.

84
Indexes for HRV analysis are usually obtained by linear methods (including time
domain and frequency domain analyses) or nonlinear methods. In this study we used the
linear method with statistical indexes of the time domain (SDNN, SD Delta NN and
RMSSD) and frequency domain indexes. Compared to the time domain, the frequency
domain provides better information of the sympathetic-vagal balance on the heart since the
SDNN of time domain represents both sympathetic and parasympathetic activity, but does
not allow for identification of changes in HRV elicited by autonomic system. Frequency
domain analysis, on the other hand, is composed of several oscillatory components reflecting
the autonomic system: high frequency (HF), low frequency (LF) and very low frequency
(VLF) components. HF covers a frequency from 0.15 to 0.4 Hz and is an indicator of the
influence of the vagus nerve on the heart. LF, ranging from 0.04 and 0.15 Hz, represents both
vagal and sympathetic components on the heart, with a predominance of the sympathetic
system. Therefore, the LF/HF ratio reflects the relative changes between sympathetic and
parasympathetic systems, and hence represents the sympathetic-vagal balance acting on the
heart. VLF does not have important role in indicating the function of the autonomic system
(Vanderlei et al., 2009).
The possibility that the surgery could affect my measurements of cardiovascular
function, in particular the postoperative autonomic nervous system hyperactivity (i.e.
increased LF power), was considered. However, I do not think that this is the case for several
reasons. First, the surgery for catheter implantation in our lambs was a simple procedure
performed under both general anesthesia and local analgesia. Second, the cortisol
concentration measured on day 3 (i.e day after surgery) tended to be lower than that prior to
surgery, suggesting that the surgery was not too stressful event compared to birth itself.

85
Third, the values of heart rate, arterial pressure and HRV in our study were similar to those
reported in other studies which focused on the postnatal changes in cardiovascular function
in lambs. Consistent with our results, other studies also found a similar high heart rate after
birth, a decrease of heart rate and increase of arterial pressure at rest in lambs from 1 day to 6
months of age (Klopfenstein & Rudolph, 1978; Minoura & Gilbert, 1987). The increasing
trend of beat to beat HRV in my study from postnatal day 2-3 to 1 year of age was similar to
other findings in lambs from 1 to 10 week-old even though the HRV in lambs in the study of
Siimes et al seemed to plateau earlier (after 6 weeks of age) (Siimes et al., 1984). In the
human, it has also been shown that HRV increases with gestational age, postnatal age and
with maturity (Harper et al., 1976; Katona et al., 1980; Wheeler et al., 1979).
The decline in heart rate values with ages observed from our study may result from an
increase of parasympathetic activity or a decrease in sympathetic activity or from
simultaneous changes in both regulatory systems. The latter is most likely the case, since the
two branches of the autonomic nervous systems do not function on a continuum to regulate
cardiac activity, thus the effect of each system on the heart cannot be simply determined by
addition or subtraction of the relative components (Hainsworth, 1995). Instead, heart rate is
affected by the net interaction between vagal and sympathetic regulation. This can explain
the unsynchronized changes among LF power, HF power and heart rate over ages in our
lambs. The sympathetic system appears to be a major regulator of vascular function up to 8
weeks of life and by 3 months of age, vagal regulation becomes dominate (Gronlund et al.,
1989; Woods et al., 1977). Our results represent the normal postnatal ANS maturation in the
term lambs with a progressive decrease in heart rate and increase in HRV which is, in part,
consistent with findings in human newborns. (Longin et al., 2005)

86
Changes in the time domain of HRV was inversely related to changes in heart rate.
(Fig. 3.4) When heart rate decreased, the time intervals between the single beats had to
become longer, thus, as expected, time domain of HRV increased. However, LF power which
mainly indicates the influence of the sympathetic system on the heart altered independently
to the changes of heart rate. The increase to peak of LF power at ~ 10 days of age in the
lambs was sex specific, due to significantly increased LF power in female from 3 days to 10
days of age but not in male. Also, this peak LF power at ~ 10 days of age occurred around
the same time with the peak of total activity score (at ~ 15 days of age) observed from the
actiwatch while heart rate slightly decreased at this age. It has been known that physical
activity strongly influences cardiac activity and is frequently characterized by decreasing
vagal tone and increasing sympathetic influences on the heart (Nakamura et al., 1993; Voss
et al., 2002; Warren et al., 1997). This was shown in the work of Voss et al in which heart
rate and HRV were evaluated in seven horses at rest and during six incremental levels of
exercise from walking on a dry treadmill to trotting in water above the elbow. Heart rate
increased together with a decrease in overall variability (SDNN, withdrawal of vagal), LF
power increased (higher influence of sympathetic nervous system) and HF power decreased
(decrease in parasympathetic activity) during exercise compared to rest. (Voss et al., 2002)
It was also found that LF power in male lambs was significantly higher than in female
lambs at 3 days of age, but there was no difference in heart rate. This highlighted the greater
sensitivity of HRV in detecting changes in sympathovagal balance in the absence of apparent
changes in heart rate or respiratory rate. Indeed, Sleigh and Henderson found HF power,
which represented vagal activity, reduced in relation to pre-surgery anxiety, whereas no
effect was seen on heart rate. (Sleigh & Henderson, 1995) The sex difference in LF power

87
could be due to an influence of sex hormones on the ANS. Testosterone is present in the
plasma of male lambs by 36h after birth and increases to a peak at 3 days of age (Savoie et
al., 1981). The higher LF power in male lambs could also be a very first sign of sympathetic
nervous system response to an unfavorable environment. This could be related to the greater
degree of acidemia (lower pH) in the males than females at this age. In this case, the high
HRV response in males may reflect an appropriate adaptation. On the other hand, it could
also reflect a higher innate sympathetic drive in the male which leads to their higher risk of
metabolic syndrome, a situation observed in human adults. (Chang et al., 2010; Licht et al.,
2010) However, this is not likely to be the case, since insulin resistance, a key phenomenon
in the pathogenesis of many metabolic cardiovascular disorders, results from a diminished
regional blood flow and tissue glucose delivery as a consequence of vasoconstriction due to
an enhanced sympathetic drive (Esler et al., 2001) and we do not see any gender differences
in arterial pressure in lambs at this age and even later in life. Another possibility, even though
less likely, of high LF power in male lambs is due to the high body fat. In human, obesity is
associated with increased sympathetic outflow to the kidneys and skeletal muscle vasculature
(Esler et al., 2001) and body fat is the main source of sympathetic discharge. Even though
male lambs usually have a higher carcass weight than female lambs at the same age, this
difference is attributed to a heavier fat-free weight only (Bennett et al., 1991).
Interestingly, our results showed that male lambs had a lower pH value than females
at 3 days of age. Although the extent of acidosis in the males was not great, this could be one
of the predisposing factors that lead to a higher mortality rate in male than female neonates
(Khoury et al., 1985), especially related to respiratory distress syndrome (RDS) (Ingemarsson
et al., 1997; Ingemarsson, 2003; Khoury et al., 1985). Ishak et al (2012) have recently

88
reported in the fetal lambs delivered by Cesaeren section at 133 days, arterial pH lower in the
male lambs than in female lambs in the initial postnatal period, with the sex difference (~0.15
pH units) being greater than the difference in the 3 day old lambs in my study (~0,08 pH
units). This suggests that an attenuated sex difference in arterial pH is present even in term
lambs. A study in Sweden analyzed acid- base balance in 15,480 human cord arterial blood
samples in term infants and showed a higher rate of pronounced acidaemia (pH < 7) in male
newborns (64%) than female newborns (36%) (Ingemarsson et al., 1997). In our study, the
earliest pH measurement obtained from the lambs was at 3 d, thus the male pH value might
have been even more acidotic at birth. This is especially a serious problem if we consider the
higher rate of preterm birth in humans, which is usually associated with pulmonary
immaturity in male infants compared to females (Ingemarsson, 2003). In addition, there is a
delay of lung maturation in male fetuses (Torday et al., 1981) and antenatal betamethasone
administration is not as effective in preventing RDS in male as in female. (Ballard et al.,
1980; Papageorgiou et al., 1981)
A sex difference in heart rate was also observed in lambs at 6 month of age. Heart
rate was significantly higher in male than female. Experiments of this age group in our study
were conducted on postnatal day 195.2 ± 8.6 and 214.6 ± 4.2 in females and males,
respectively. Female lambs raised under natural light can reach puberty as early as 153 ± 7.1
days in Dorset (Fitzgerald & Butler, 1982) or up to 234 days in Dorset and 245 days in
Suffolk ewe lambs (Quirke et al., 1985). Male lambs (Dorset x Leicester x Suffolk) raised
indoor under natural light conditions, on the other hand, enter puberty a bit earlier between
150 to 200 days of age as defined by an increase in plasma testosterone concentration from
1ng/ml at 150 days of age to 8 ng/ml at 200 days of age (Yarney & Sanford, 1989). Since our

89
lambs were raised under a 12 hour light/dark cycle, their sexual maturation could be delayed,
but in any case male lambs reach puberty earlier than females. Therefore, the differences in
their heart rate might be a result of sex steroid hormone influences on the cardiovascular
system. With the growing rates of cardiovascular disease and metabolic syndrome around the
world, they could become the epidemics of the century (Banos et al., 2011; Iyer et al., 2011,
Barouki et al., 2012). There will no doubt be increasing attention paid to the effects of sex
hormones on the occurrence of these diseases. It appears that androgen and estrogen exhibit
differential effects on the pathophysiology of the vascular beds in male and female. So far,
whether testosterone is protective or harmful to the development of those chronic diseases is
still a matter of debate (Corona et al., 2011; Traish & Kypreos, 2011). In my study, it is
uncertain if a gender difference in heart rate would have any long-term effects on the lambs
given that there were no accompanying significant differences in other systemic
cardiovascular variables such as blood pressure and heart rate variability between genders at
this age.
The rapid increase in arterial PO2 in the postnatal lambs compared to the fetus at term
is the result of a transition from gas exchange via the placenta in the fetus to air-breathing in
the newborn. As the lung takes over the role of placenta in gas exchange, it is necessary that
the lung fluid is cleared from the airway and pulmonary vascular resistance decreases around
birth (Hooper & Harding, 2001). However, my results showed that after birth, arterial PO2
continues to increase slowly with age. This increase in PO2 is consistent with a slow increase
in O2 saturation and slow decrease in PCO2. These changes could reflect slow changes in
alveolar O2 and CO2 concentrations in the lung. If this is the case, the lambs must be
hypoventilated initially but increase thereafter due to a slow shift in the set point for

90
ventilatory control in the carotid body regulating PO2. Alternatively, these changes could
reflect slow changes in the alveolar-arterial gas differences. In that case, the lambs are
ventilated adequately at first but there is a diffusion limitation that is slowly corrected over
time. This is most likely to be the case since it has been shown that in sheep even though the
blood air distance becomes quite thin after birth (~0.2µm), it is only at focal points initially
and it is not until they reach adulthood that the basement membrane fusion has spread
enough to cover more than 50% of the epithelial surface (Hooper & Harding, 2001).
The level of activity in the lambs increased progressively right after birth, reached a
peak level at ~13-15 postnatal days and then started to decline, becoming stable around
postnatal day 30. The activity results for the first 10 days in these newborn lambs is similar to
the activity level that was found in term lambs in a previous study (Rurak et al., 2008). Over
the 90 days of monitoring, even when lamb’s activity decreased to a basal level, the lowest
average values of their total activity score (212473 ± 23298) which is on postnatal day 60
was still higher than the value reported in adolescent sheep (100927 ± 7189) or in adult ewes
(134808 ± 14644) held in the same conditions (Rurak et al., 2008). To determine when the
level of activity decreases to the level of adolescent or adult sheep will require a longer
monitoring period.
For the first 30 days, the increase in daily total activity score, active bouts per day and
the intensity of activity to the peak on postnatal day 13-15 of is probably a sign of the
maturation in their bone and skeletal muscle after birth (Rurak et al, 2008). The pattern is
reversed for the duration of activity. The shifts in activity pattern may reflect the suckling
pattern in lambs which goes from less frequent and intensive for a longer period of time at
birth to more frequent and intensive but for a shorter periods of time around postnatal day 15,

91
and then suckling less and less until eventually the lamb was weaned and eating solid food,
which they can do for longer periods of time. The underlying mechanism of this pattern of
activity may involve the changes in the hormonal levels, such as insulin-like growth factor I
which peaks at around the same time (i.e postnatal day 13-15) (Greenwood et al., 2002),
since from direct observation, lambs start to be interested in grain at around this period.
Indeed, a study that was focused on the effects of birth weight and postnatal nutrition on
neonatal lambs has shown that the IGF-I concentration is maximum on days 9-11 in high
birth weight lambs and on days 11 to 13 in low birth weight lambs (Greenwood et al., 2002).
Another explanation for the shifts in lambs’ activity could be due to the changes in milk
production from the ewe over time. When the milk production from the ewes is at its peak,
the lamb’s activity was also highest. Milk production of 2 year old ewes rearing twin lambs
and 1-2 year old ewes rearing singletons peaked at 21 day of lactation and 27 to 30 of
lactation, respectively (Cardellino & Benson, 2002). However, the time of the peak of milk
production in our ewes could have been shorter, at around 15 day of lactation, since most of
them werer older than 2 years.
Using FFT we found the initial rhythms present after birth in lambs was dominated by
the short 1-2 hour rhythms, which corresponded to suckling patterns from our DVR
observations. Over the next four weeks, these rhythms decreased in dominance, matching the
decrease in frequency and duration of suckling. When lambs’ nutritional requirement
changed from the solely milk regimen to solid food supplement, reflected by the
disappearance of the short 1-2 hour rhythm due to suckling, their activity also declined.
Interestingly, around the time when the 1-2 hour rhythms disappeared and their activity
declined, their growth rate was also reduced.

92
In human infants, there are ultradian rhythms of 3h, 4h and 6h as sleep, wake and
food intake behavioral cycles (Menna-Barreto et al., 1993). The time course of the
development of a permanent circadian rhythm for lambs seems to be similar to that found in
humans. In humans, Tomioka & Tomioka found that the circadian rhythm of sleep-wake
cycle appeared in the first week of life in 3 infants and became dominant in the 3rd and 5th
week of life (Tomioka & Tomioka, 1991). However, there was a large interindividual
variability of both the ultradian rhythms and the development of a 24 h pattern, whose
appearance varied considerably between the 6th and the 16th week of life (Lohr &
Siegmund, 1999). In sheep, the dominant circadian rhythm is in synchrony with the
development of a postnatal melatonin rhythm in lambs at 3-4 weeks (Nowak et al., 1990). In
some lambs in our study, the circadian rhythm existed for 1-2 days after birth and
disappeared for almost a month before it became persistently dominant. The existence of the
circadian rhythm for the first few days after birth in the lambs could be an influence of the
remaining melatonin level from the ewe after birth (McMillen et al., 1989).
Similar to infants, lambs have 2 developmental processes in the development of their
circadian rhythm: a decreased ultradian and increased circadian components and the process
of synchronization with external time cues or social factors such as light/dark cycle, meal
feeding, experiments, ect. The development of circadian rhythm appears to be important,
since its existence plays an important role in the well being of both animals and humans,
because it guarantees optimal functioning of biological systems with maximum efficiency.
Once the circadian rhythm appears, it lasts for a considerable length of time, but begins to
break down during illness or old age (Weinert, 2000).

93
The eight hour ultradian rhythm emerged as a dominant rhythm around 2 weeks after
birth and is thought to be due to the daily scheduled feeding times, which occured twice a
day at 7:30-8:00 am and ~ 2:30-3:00 pm, an approximately eight hour interval. Even though
the young lambs only fed on their mothers’ milk, they became more active as their mothers
were being fed. To further confirm that the 8 hour ultradian rhythm is the result of the
feeding time entrainment, further study is needed to determine if this ultradian rhythm exists
in the lambs that develop under field conditions where they can graze ad libitum.
The 6h rhythm seemed to become the third most dominant rhythm when the lamb was
about two months of age. This 6 hour rhythm was most likely a result of maturation of the
lambs’ rumen and reticulum, which were stimulated by the intake of solid food, as we found
a correlation between the first time lambs eat significant amounts of hay, and the day this 6 h
rhythm became dominant. This is supported by research which showed that β-
hydroxybutyrate production from butyrate in isolated sheep ruminal cells increased 10 fold
from day 42 to day 56, as compared to the rate at birth; this marks a shift in the ability to
synthesize ketones in a mature rumen, and the development of the rumen microbiota
(Baldwin & Jesse, 1992). Lane et al also observed changes in the morphology of the rumen
epithelium (fewer and larger intraluminal papillea per cm2) on day 84 from lambs fed with
solid food after postnatal day 49 (weaned lambs), as compared to epithelium of lambs fed
solely with milk replacer (non-weaned lambs) (Lane et al., 2000)
The suckling frequency was similar in singleton, twins or triplet lambs. However,
triplets seemed to have longer feeding bouts which might be secondary to the restriction of
the available milk supply in triplet pregnancies. As a result, twins or triplets spent more time
in competing with their siblings to obtain milk, which led to an increase in their feeding

94
bouts,but it did not appear to improve the efficiency of their suckling, as reflected in the
tendency for a lower weight gain in triplets and twins, as compared to the singleton, even
though this difference was not significant. This could be due to my small sample of triplets.
Results arising from a much larger study, comprising approximately 3000 singleton, 1500
twins and 200 multiple lambs have shown that singleton lambs gained significantly more
weight than twins and multiples. As a result, singleton lambs were heavier than those born as
twins and multiples, both at birth and at weaning (Carrillo & Segura, 1993).
The short 1h rhythm was not observed in adult sheep and only the lactating ewes have
the short ~ 1h rhythm, but none of them had this as the first dominant rhythm. This suggested
that the 1h rhythm is the rhythm that the ewes share with their offspring. It has been known
that suckling serves as a mechanical stimulus to inhibit the HPA axis in the ewe to regulate
the estrus cycle. Depending on breed, time of the year and degree of mammary stimulation,
the duration of lactational anestrus varies considerably (Fletcher, 1971; Gordon & Siegmann,
1991; Mallampati et al., 1971). Usually after the termination of lactation, the mother and
lamb relationship does not fade away completely. Indeed, lambs remain associated with their
mothers for several weeks or months (Hinch et al., 1990). Therefore, beside measurement of
maternal prolactin level, the waning of this 1h rhythm in the ewe, which can be non-
invasively obtained by actigraphy, could be an alternative indicator that the ewe was free
from the lactational inhibition of reproduction.
Our results provided a normative data set of cortisol concentration in the normal
lambs at different ages under basal conditions. Cortisol level on postnatal day 2 ± 0.2 (i.e
prior surgery) were higher than that measured in the fetus at 7.1 ± 1.6 days before delivery
(at gestational age = 135.4 ± 0.7 days). This was consistent with the cortisol surge resulting

95
from activation of the fetal HPA axis at ~ 125-130 of gestation which is required for the
initiation of parturition (Wood, 1994, Norman et al., 1985). Cortisol concentration declined
significantly thereafter during the first 10 days of life and was returned to the similar level
observed in the fetus on gestational day 135.4 ± 0.7, and further decreased after 1 month of
age to reach the adult value at 3 months of age. However, plasma ACTH concentration was
not well-correlated with the postnatal plasma cortisol changes. ACTH was low on postnatal
day 2 ± 0.2, probably due to a refunctioning of cortisol negative feedback after birth, which
was desensitized at the end of gestation. In addition, the high cortisol concentration without
an adequate increase of ACTH could be a result of increasing cortisol binding protein near
term, which exaggerates the total cortisol level but suppresses the efficient negative feedback
of cortisol on the pituitary, as well as hypothalamus (Challis et al., 1995). The ACTH level
remained lower at other ages as well. It was not clear why there was dissociation between
plasma cortisol and ACTH concentration even at these later ages.
Cortisol concentration was significantly higher in the morning (at 10:00, 10:30 and
11:30 am) than in the afternoon (at 3:30 pm) or in the evening (at 9:30 pm) in the lambs at 10
days and 1 month of age which suggested that there was diurnal rhythm of cortisol in the
lamb at these ages. Such a rhythm was not found at other ages (3 day, 3, 6 and 12 month-old)
which is in agreement with the findings from the work of Bell et al, who reported there were
no endogenous circadian or diurnal rhythms in ACTH or cortisol secretion in pregnant, non
pregnant cycling, non cycling or ovariectomized adult ewes (Bell et al. 1991). In our study,
the samples were collected for only a 12h period; therefore we could not make a firm
conclusion about a circadian rhythm of these hormones, which appear to be present in lambs
after 3 weeks of age (Parraguez et al., 1989). There were some other factors that might have

96
contributed to the higher level of cortisol in the morning. First, it has been known that the
cortisol level increases after feeding in pregnant ewes (Slater & Mellor, 1981) and in the
human (Follenius et al., 1982). Lambs around 10 days to 1 month old had a high activity of
suckling. In addition, from direct observations we noticed that after the first week lambs
started to participate in nozing the ewe’s daily feed. However, if the higher cortisol level was
a response to feeding, the cortisol level measured at the earlier time (around 8:30 am) should
have been higher, and also higher at the pm feeding time (at 3:30 pm), which was not the
case. Thus, it is more likely that the high cortisol level for the 2 h of experiment after dosing
is a result of the lamb’s response to the social cue of the investigators’ close proximity for the
2 h duration when there was frequent sample collection. This social cue disappeared at later
ages when lambs become habituated to the investigators.
Although there was no rhythm in the older lambs, we did find a sex difference in the
cortisol level in lambs at 1 year of age at 0.5 h (10:00 am), 1 h (10:30 am), 6 h (15:30 pm)
and 12 h (21:30 pm) sample. Studies that have examined the influence of sex hormones on
the cortisol level have demonstrated estrogen-enhanced HPA axis function in adult animals
under both basal and stress conditions (Bell et al., 1991; Carey et al., 1995; Wood et al.,
2001). However, my results showed that basal cortisol concentration was lower in the female
than male lambs, which suggests a reduced HPA axis activity in the female at this age.
Nevertheless, since cortisol levels vary during the estrous in sheep (Bell et al., 1991), it is
possible that the cortisol levels in the female were measured in noncycling animals or during
anestrus cycles which have no estrogen effects.
In our study the lambs were not weaned abruptly. In studies focused on the effect of
diet and weaning on growth, the time of weaning can be as early as 28 days of age (Lane &

97
Albrecht, 1991) or as late as 42 days, by which time the rumen is believed to be already
functional (Holcombe et al., 1992). The earliest our lambs were separated with their mothers
was at 47 days of age and in some cases they remained with their mothers for up to 80 days.
However, the lambs were gradually weaned by the ewes refusing to let them suckle and
walking away from them, even though the lambs still tried to nurse sometimes. In the context
of our study, the time of weaning was defined as the time window when lambs started to first
consume solid food (hay) to when the 1h rhythm due to suckling disappeared.
The blood glucose concentration, which is related to rumen development (Holcombe
et al., 1992) decreased gradually with age, and the value at 1 month was significantly
different from that at other ages. This alteration in glucose concentration were consistent with
our observations of the time lambs first started to consume solid food (29.6 ± 1.1 days of
age) which indicated a functional forestomach already developed. Similar results were also
reported in the work of Holcombe et al., (1992). Our results, taken with the data from the
Holcombe et al study, suggest that the window of weaning is around 1 month of age. It
appears that the greatest changes (i.e break points on piecewise linear regression) of most
physiological variables (heart rate, SDNN, Glucose, pO2, urine flow rate) occurred around
the window of weaning time (1 to 2 months of age), so that by 3 months of age they are
similar to their levels at more mature ages.
A decrease in urine flow rate around the time of weaning was reasonable as the lambs
changed from a predominantly milk diet to a solid food diet. In addition, it has been known
that feeding activity has effects on the cardiovascular system; thus it was not surprising to
observe heart rate and HRV (SDNN) were significantly altered at the time of weaning.

98
Overall, our results revealed a data set of normal development of physiological,
endocrine and behavior characteristics in the lambs in relation to several milestones of the
developmental process (birth, weaning and puberty) under basal conditions. This will provide
reference information of these variables for future studies related to any relevant disorders.

99
4. Acute Fluoxetine Effects In The Newborn
4.1 Introduction:
Major depression is probably one of the most burdensome disorders in the world. In
the report of World Health Organization (WHO) in 1996, it was estimated that by 2020
depression will rank second only after cardiovascular disease in terms of its disability-
associated burden (Murray & Lopez, 1996). Depression affects at least about 15% of women
in their lifetime (Burt & Stein, 2002; Kessler et al., 2003). The risk is even higher during the
reproductive years. Thus there is a significant risk that one might become depressed during
pregnancy or even get pregnant while being depressed or on antidepressant therapy. Recent
studies have failed to confirm the previous belief that the state of pregnancy is protective
against mood disorders (O'Hara et al., 1990). Moreover, it has been shown that untreated
maternal depression leads to long-term neurobehavioral consequences in the offspring,
including poor cognitive function, and diminished language and psychomotor development
(Deave et al., 2008; DiPietro et al., 2006; Hay et al., 2008; Murray & Cooper, 1997; Murray
et al., 2001; Nulman et al., 2002). Therefore, pharmacologic treatment is recommended for
pregnant women who have moderate to severe symptoms.
In addition, depression during pregnancy is in turn a risk factor for postpartum
depression, affecting 10% to 22% of women (Burt & Stein, 2002). Since the postpartum
period is the most vulnerable time for serious symptoms and their short-term and long-term
consequences to not only the mother but also the entire family unit, this has resulted in
postpartum depression becoming a significant health care problem. Moreover, it has been
shown that the risk of relapse increases 5 times in women who discontinued medication

100
during their pregnancy compared with those who maintained their medication (Cohen et al.,
2006). Thus continuing medication in the third trimester is recommended (Toohey, 2012).
Among the classes of antidepressants, SSRIs have been widely used in pregnant women due
to their relatively safe side effects profile. In fact, 33% of pregnant women with clinical
depression were prescribed an antidepressant (Oberlander et al., 2006), in which SSRIs
accounted for 75% of the cases (Simon et al., 2002). Recent studies have reported an
increasing use (4.9- 6.2% increase over 9-14 years) of SSRIs during pregnancy (Andrade et
al., 2008; Cooper et al., 2007; Simon et al., 2002; Wichman et al., 2008). However, third
trimester use of SSRIs, unfortunately, was also reported to increase the risk of a variety of
neonatal complications termed “neonatal behavioral syndrome” or “poor neonatal
adaptation” after birth (Chambers et al., 1996; Cohen et al., 2000; Kwon & Lefkowitz, 2008;
Mhanna et al., 1997; Oberlander et al., 2004; Oberlander et al., 2006), a phenomenon
observed in 10% to 30% of infants exposed in utero to SSRIs (Koren et al, 2009). The
presenting symptoms include irritability, persistent crying, shivering, tremor, restlessness,
feeding difficulties, sleep disturbances or even more severe symptoms, such as seizures,
dehydration, excessive weight loss, hyperpyrexia and the need for intubation (Moses-Kolko
et al., 2005). Even though this syndrome is self-limiting, it results in longer hospitalization
and led to a warning from Health Canada and FDA about the adverse events when using
SSRI/SNRIs in pregnant women during the third trimester (Summary minutes of the
pediatrics subcommittee of the anti-infective drugs advisory committee.2004).
There are several suggested mechanisms for the poor neonatal adaptation. First is a
withdrawal syndrome due to the discontinuation of maternal-fetus placental transfer right
after birth. Second is a serotonin toxicity mechanism induced by the high circulating SSRI

101
concentrations in the newborn at birth, due to their low capability of drug clearance. Third is
an SSRIs-elicited alteration in fetal brain development. Findings supporting the latter
mechanism arise from the work of our group (Morrison et al, 2001; Morrison et al., 2002;
Morrison et al., 2004; Morrison et al., 2005), which reported a decrease of fetal rapid-eye-
movement (REM) sleep in lambs whose mother were administered FX i.v. continuously for 8
days in late gestation. Similar findings were also demonstrated in human fetus with SSRI
exposure (Mulder et al, 2011). Further supporting evidence comes from the study of
Oberlander et al., 2006 and Rurak et al., 2011, who observed altered fetal middle cerebral
artery flow characteristics in 36 week human fetuses exposed in utero to SSRIs compared to
a control group.
However, which mechanism is responsible for the observed poor neonatal adaptation
is not yet clear. Currently, the management for these syndromes is simply symptomatic with
supportive care (Koren et al., 2009). Since the strategy of treatment could be completely
opposite for the first and second mechanisms mentioned above, it is important to elucidate
the underlying mechanism for the syndrome. For this reason, in this study we investigated the
effect of acute FX, one of the most prescribed SSRIs on growth, behavior, and
cardiovascular-respiratory, and endocrine functions in ~4 d old newborn lambs. The
hypothesis that was tested was that acute administration of FX to achieve plasma
concentrations similar to those in human newborns exposed to FX in utero would result in
effects similar to poor neonatal adaptation in human infants.

102
4.2 Methods:
4.2.1 Experimental Protocol:
21 lambs were included in the study reported in this chapter, of which 9 lambs were
in the control group with sterile water i.v. bolus administration (0.1ml/kg) and 12 lambs were
in the FX group with FX (1mg/kg) bolus i.v injection, respectively. The animal housing,
surgical, experimental and analytical methods are described in Chapter 2. Heart rate and
blood pressure data used in this chapter were averaged at 1 min intervals for 15 mins before
dosing and the first 10 mins after dosing and at 5 min intervals for the remaining time while
HRV data were averaged over 30 mins intervals and also averaged at 1 min intervals for 2
min before dosing and 5 mins after dosing. Blood gas samples were collected at -15, 5, 15,
30, 60, 120 mins as described above, however, another arterial blood gas sample was taken at
2 mins in 4 lambs in this study to characterize more fully the short term changes in blood gas
and acid-base status following water or FX administration. For the rest-activity analysis,
beside the control group with sterile water injection and FX group with FX injection, another
group of normal term lambs in which no surgeries or experiments occurred were included.
4.2.2 Statistical Analysis:
Activity data were analyzed using a 2 way GLM for repeated measures ANOVA
followed by the Bonferroni t-test for multiple comparisons versus the control group to
determine the effect of treatment and sex on changes in behavioral activities. Cardiovascular
and blood gas data were analyzed using a 3 way ANOVA followed by Bonferroni’s
correction for pairwise or control comparisons to determine the effects of treatment, time and
sex on the measured variables. Two way GLM for repeated measure ANOVA was used for

103
not-normally distributed variables (SDNN, SD Delta NN, RMSSD, LF power, HF power,
LF/HF ratio, O2 saturation, pCO2, glucose and Hb) to determine the effects of treatment and
time. Because plasma cortisol concentrations were not normally distributed, the non-
parametric Mann-Whitney U-test was used for the cortisol data to determine the effect of
treatment. P < 0.05 defines statistical significance. Data are presented as mean ± SEM.
4.3 Results:
On postnatal day 3.3 ± 0.4 and 4.2 ± 0.4 sterile water (0.1ml/kg, N=9) or FX
(1mg/kg, N=12) was acutely injected via the jugular vein catheter, respectively.
4.3.1 Plasma FX Concentration And Weight Gain:
The plasma FX concentration in the ~ 3 day old lambs over 72h of experiment and in
13 individual human infants sampled at postnatal day 2 following in utero exposure to the
drug cover a similar range (~ 10-100 ng/ml) (Fig. 4.1A). (Chow, 2013; Kim, 2000) There
were no significant differences in postnatal weight gain on postnatal day 4 between the
normal term lambs group who were not subjected to any experiments or surgery (0.327 ±
0.048 kg, n = 11), control group (0.411 ± 0.056 kg, n = 9), FX group (0.392 ± 0.045 kg, n =
12), respectively (Fig. 4.1B). This indicated FX does not have any effects on initial postnatal
weight gain in the neonatal lambs.

104
FIG. 4.1: (A) Plasma FX concentration verus time curve in the 3 day-old lambs (closed symbols and solid line) and in 13 individual human infants (open symbols) sampled at postnatal day 2 following in utero exposure to the drug. Human data was taken from the dissertation of Dr. John Kim, 2000 and the lambs data was taken from the dissertation of Dr. Tim Chow, 2013. (B) Weight gain on ~ postnatal day 4 among the normal term lambs (n = 11, open bar), control group (n = 9, black bar), FX group (n = 12, gray bar) and post FX day 1 (n = 12, light gray bar)
Time (h)
0 20 40 60 80
[FX
] (ng
/ml)
1
10
100
1000
3 d lambsHuman PKU [FX]

105
4.3.2 Rest Activity Cycles:
The number of active bouts per day, total activity score, the intensity and duration of
activity and the percent of time moving and percent immobile time were comparable between
the normal term lamb group (n = 10), the control group (n = 6) and the FX group (n = 11)
(Fig. 4.2). In addition, from the DVR observation we did not notice any obvious behavioral
changes or any symptoms representing serotonin toxicity with FX dosing. During the first
2.5h of experiment when lambs were kept in the sling, they just lay quiet or slept for most of
the time, and there were no obvious behavioral changes following FX administration.
4.3.3 Cardiovascular Variables (Heart Rate, Arterial Pressure, Heart Rate
Variability):
Heart rate, arterial pressure and heart rate variability (HRV) in the newborn lambs
with placebo and FX injection are illustrated in Figures 4.3 and 4.4, respectively. The time
points included in the 3 way ANOVA test (treatment x time x sex) were at -15 min, 1,2,3,4,5
and 10 min only, since the data was not normal distributed with additional time points.
Arterial pressure were significantly different between the control and FX group at 1, 2, 3, 4, 5
and 10 minutes post-FX (p<0.001), but not at the control time point (-15 minutes). The
maximum increase in arterial pressure in the FX group was 8.5 ± 1.1 mmHg at 4 minutes
following FX injection compared to values before FX injection. Heart rate was lower in the
FX group than the control group only at 2 minutes post-FX (p = 0.034), the difference was 38
± 9 bpm. However, these effects were very transient and self-limited since heart rate returned
to normal values within 1 min later and by 15 minutes post-FX arterial pressure had also
returned to values similar to those observed before FX injection.

106
FIG. 4.2: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in the normal ~ 4 day-old term lambs (n = 10), control = on day of control experiment (n=6), and FX lambs on the day of FX experiment (n = 11) followed up to post FX day 1 (n = 11), post FX day 2 (n = 11) and post FX day 3 (n = 11). No significant changes were found between activities after FX administration and sterile water injection or normal day without experiments.

107
FIG. 4.3: Arterial pressure and Heart rate (at 1 min intervals for the first 5 min after dosing and at 5 minute interval for the whole 2.5h of experiment) in the ~ 4 days old lamb with sterile water injection (control group) and FX injection (FX group). a indicates significant differences between variables at -15 min vs 1,2,3,4,5,10 min after FX within the FX group b indicates significant differences between control and FX at the corresponding time
a a a a ab b b b b b
b
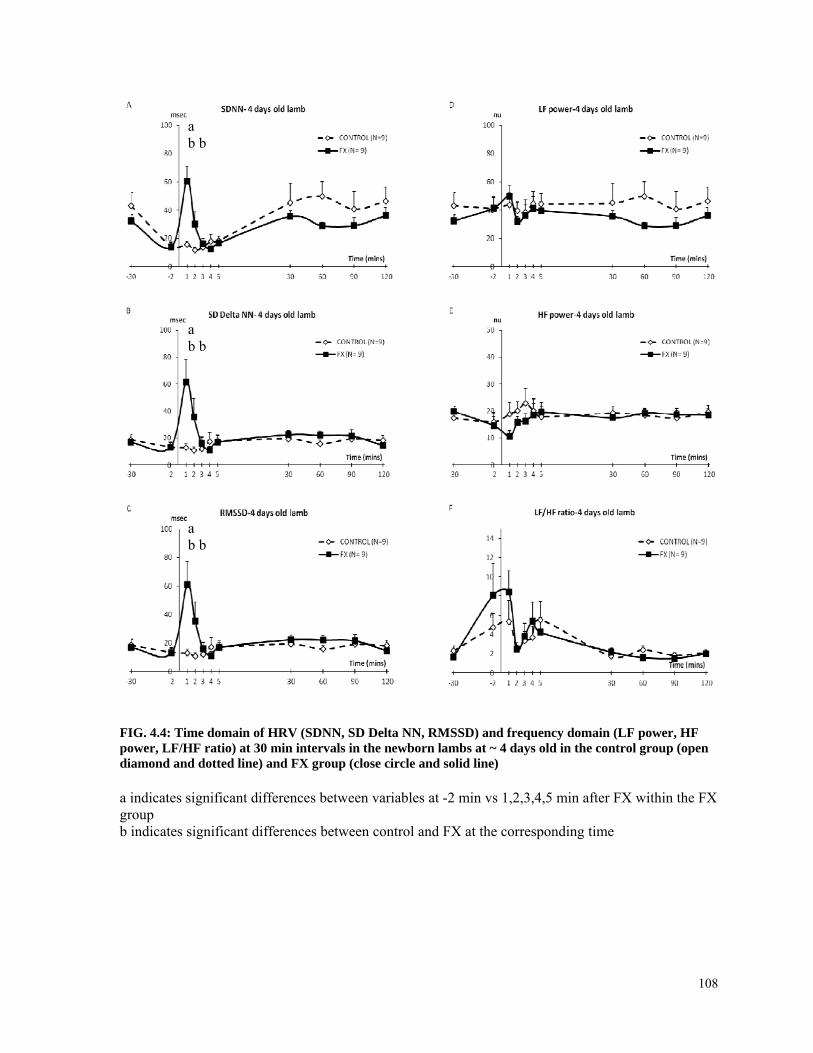
108
FIG. 4.4: Time domain of HRV (SDNN, SD Delta NN, RMSSD) and frequency domain (LF power, HF power, LF/HF ratio) at 30 min intervals in the newborn lambs at ~ 4 days old in the control group (open diamond and dotted line) and FX group (close circle and solid line) a indicates significant differences between variables at -2 min vs 1,2,3,4,5 min after FX within the FX group b indicates significant differences between control and FX at the corresponding time
a b b
a b b
a b b

109
The time domain of HRV including SDNN, SD Delta NN, RMSSD, which is related
to the RR interval variability was significantly higher in the FX group than that in the control
group at 1 and 2 min after FX administration but were comparable between the two groups at
other time points. Indeed, there were significant relationships between the changes in SDNN,
SD Delta NN, RMSSD at 1 min following FX injection and the changes in heart rate at 2
mins following FX with r2 = 0.8, p = 0.001 and r2 = 0.57, p = 0.0017, respectively (Fig. 4.5)
However, the frequency domain of HRV including LF power, HF power and LF/HF ratio,
which indicate parasympathetic and sympathetic influences on the heart were not different
between the control and FX group and also did not change significantly over time. (Fig. 4.4)
Sex differences were observed with SD Delta NN, RMSSD (within the control group, at - 30
min) and with LF power and LF/HF ratio (at - 30, 60 and 120 min) but the differences were
not related to treatment (data not shown).
4.3.4 Arterial Blood Gases:
Figure 4.6 and 4.7 illustrate the changes in arterial blood gas variables (O2 sat, pO2,
pCO2, pH, BE, HCO3), and lactate, glucose and Hb concentrations in newborn lambs with
placebo and FX injection. There were no consistent changes in these arterial blood gas
variables in either control or FX group with time. Arterial pH tended to decline in both
groups with treatment but this was not statistically significant. A paired-t test was performed
to further compare Po2 and O2 saturation at -15 minute before and 2 minute after FX injection
in a smaller group of lambs (n= 4) but no statistical significance was reached. There was a
sex difference in pH, HCO3 and lactate but this did not depend on the treatment. Hb was
slightly lower, but significantly on the day of FX than the control day. This could due to the
blood volume lost for sampling.

110
FIG 4.5: The relationship between changes in heart rate at 2 mins following FX (delta HR) and changes in time domain of HRV (SDNN, SD Delta NN, RMSSD) (i.e Delta SDNN, Delta SD Delta NN, Delta RMSSD) at 1 min following FX in the lambs at ~ 4 days of age.

111
FIG. 4.6: Arterial blood gases (pH, O2 saturation, pO2, pCO2, HCO3, Base Excess) in the newborn lambs (~ 4 days old) in the control group (open diamond and dotted line, n = 9) and FX group (close circle and solid line, n = 11)
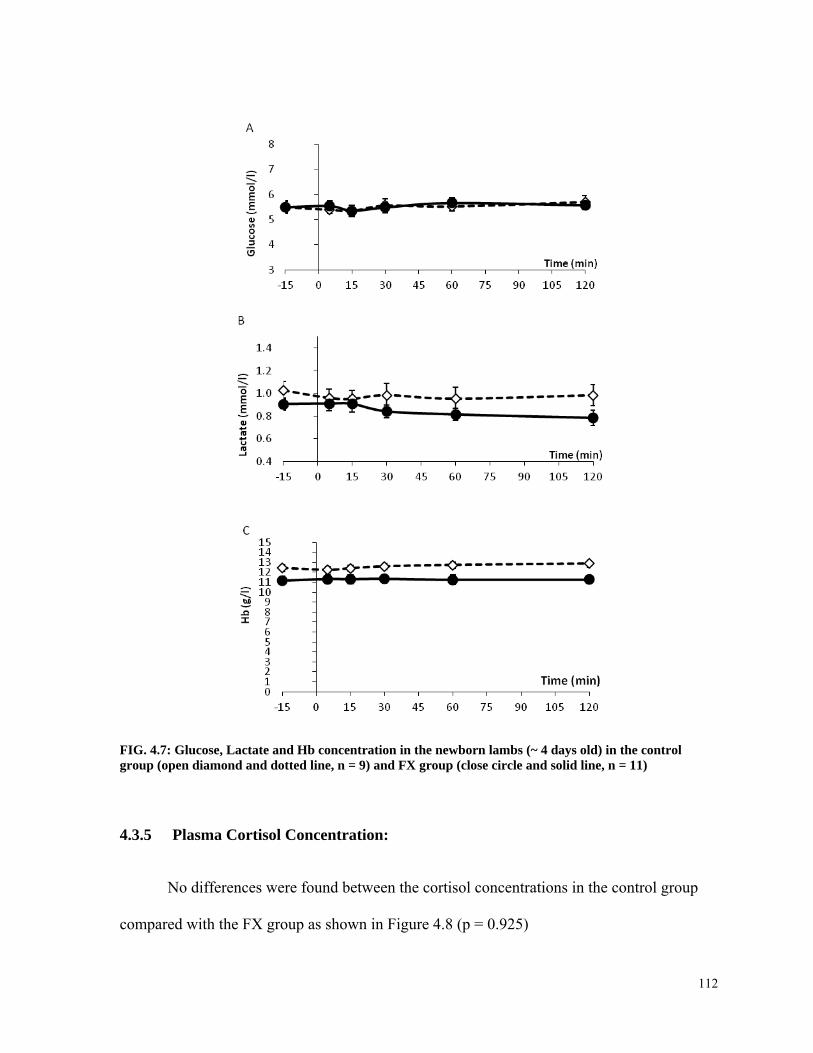
112
FIG. 4.7: Glucose, Lactate and Hb concentration in the newborn lambs (~ 4 days old) in the control group (open diamond and dotted line, n = 9) and FX group (close circle and solid line, n = 11)
4.3.5 Plasma Cortisol Concentration:
No differences were found between the cortisol concentrations in the control group
compared with the FX group as shown in Figure 4.8 (p = 0.925)

113
FIG. 4.8: Plasma cortisol concentration in the ~ 4 days old lambs in the control group with sterile water injection (n = 8, open bar) and the FX group with FX injection (n = 5, close bar) at 30 and 15 minutes before injection and 0.5, 1, 2, 6, 12 hours after injection
4.4 Discussion:
We have found that acute FX administration did not cause any significant effects on
the feeding behavior, rest-activity cycle, cardiovascular-respiratory functions and the cortisol
level in the approximately 4 day old postnatal lambs. Mild and transient increase in arterial
blood pressure and decrease in heart rate occurred immediately after FX administration and
recovered within 15 minutes without any significant changes in HRV or arterial blood gases.
The plasma FX concentration time curve in our studied lambs over 72h of experiment
fell in the range of 10-100ng/ml, which is similar to the plasma FX level found in 13
individual human infants sampled at postnatal day 2 following in utero exposure to the drug
(Kim et al,. 2006). The wide range of plasma FX concentration in human infants could be
due to the differences in maternal FX doses or to cytochrome P450 polymorphisms in the

114
infants, which could affect their ability to metabolize the drug, a phenomenon that has been
reported in adults (Bradford, 2002; Fjordside et al., 1999; Sachse et al, 1997; Scordo et al,
2005)
Acute FX effects on weight gain (feeding behavior):
Studies in humans regarding postnatal weight gain in infants breastfed by mothers
taking FX are inconsistent. While Chambers et al., (1999) reported a diminished postnatal
weight gain in nursing infants of mother treated with FX, a subsequent study from Hendrick
et al, (2003) failed to confirm this finding. In our study, the postnatal weight gain was
comparable between the acute FX exposed and the normal term lamb group on postnatal day
4. The activity bouts per day were also not different between the two groups. As was
demonstrated in chapter 3, this variable at this postnatal age was always associated with
suckling or suckling attempts. Collectively, this suggests that acute FX administration did not
affect feeding behavior in the lambs. However, it must be kept in mind that in our study, FX
was given acutely single dose, while in the human studies, the infants were exposed to FX
chronically from the prenatal to postnatal period. Acute and chronic FX exposure might exert
different effects on weight gain and feeding behavior. There are unchanged plasma
concentrations of leptin, a feeding-inhibiting hormone, in response to acute administration of
the serotonergic agent, clomipramine, in both normal weight and obese women (Oppert et al.,
1997). However, a study in rats showed acute or chronic FX administration resulted in a
significant decrease in food intake and plasma leptin levels in lean and obese rats (Dryden et
al.,1999). The differences in those findings and ours might be attributed to the differences in
animal ages or the dose of SSRIs used (10mg/kg in rat study and 1mg/kg in ours)

115
In addition, there were also no significant differences in factors that might have
influences on postnatal weight gain in sheep such as birth weight, litter, breed, ewe-lamb
bonding and neonatal behaviors (Dwyer, 2003; Dwyer et al., 2005) between the normal term
lambs and the FX group. In addition, different from human studies, the effect of FX on
postnatal weight gain in sheep was not confounded by the severity of maternal depression
and other associated risk factors (eg: low socioeconomic status, family conflict, financial
stress, ect), which could presumably affect neonatal weight gain (Stewart, 2007; Wright et
al,2006; Zuckerman et al., 1989).
4.4.1 Acute FX Effects On Cardiovascular –Respiratory Functions:
FX and other SSRIs cause an acute increase in plasma serotonin levels through
inhibiting the serotonin reuptake by platelets (Maurer-Spurej et al., 2004). The effects of
serotonin on the cardiovascular system are contradictory between in vivo and in vitro studies,
at least in the rat (Watts & Davis, 2011). Serotonin has long been known as a vasoconstrictor
agent, which can lead to hypertension (Maurer-Spurej et al., 2004). However, it appears to
have divergent effects on the cardiovascular system. Some studies showed an increase in
blood pressure with acute FX injection (Anderson et al, 1996; Cavero et al., 1981;
Lazartigues et al., 2000) while others reported a decline in blood pressure (Callera et al,
2005; Fuller et al, 1979). In addition, the cardiovascular response to serotonin (i.v) is species-
specific (Watts & Davis, 2011): a decreased blood pressure was observed in broiler chickens
(Chapman & Wideman, 2002) while an increased blood pressure occurred in calves (Linden
et al., 1999). In conscious sheep, serotonin injection caused a dose-related increase in blood
pressure (Nelson et al., 1987). This is in agreement with our findings that arterial pressure
increased at 1-10 minutes after FX administration in the newborn lambs. Heart rate was also

116
declined at 2 minutes post-FX. However, these effects were very transient and self-limited
since heart rate returned to normal values within 1 min later and by 15 minutes post-FX
arterial pressure was also returned to values similar to those observed before FX injection.
Therefore, we think these transient changes did not have any clinical significance in the
newborns. Interestingly, we noted that cardiovascular responses to acute and chronic FX
were opposite. The previous study from our group with 8 day i.v FX infusion in the pregnant
ewes showed an increase in fetal heart rate during the first 6h of FX infusion and no changes
in daily maternal arterial pressure and heart rate nor fetal arterial pressure during the period
of 8 day infusion (Morrison et al., 2002). The possible explanation for the different
cardiovascular response is that serotonin response is different with acute and chronic FX
treatment. It was reported in human and mice that acute FX treatment increases plasma
serotonin concentration while chronic FX treatment causes a decrease in plasma and platelet
serotonin levels (Alvarez et al., 1999; Alvarez et al, 1999). Indeed, in the previous sheep
study in our lab with chronic FX i.v infusion, the average maternal plasma serotonin level
increased for the first hour (at 15, 30 and 60 minutes) but decreased at 2 and 6 hours
compared to values prior to FX infusion (Morrison, 2001).
The transient mild bradycardia could be due to either a direct effect of FX on the
cardiomyocytes via 5-HT4 receptors or an indirect effect of FX on blood pressure leading to
a baroreceptor reflex in the autonomic nervous system (ANS) to increase vagal tone (Watts
& Davis, 2011). This reflex bradycardia could be mediated by 5-HT3 receptors located on
afferent vagal and sympathetic neurons (Kaumann & Levy, 2006). Even though it has been
reported that FX can inhibit the L-type Ca2+ current, which plays a role in reducing cardiac
contractility (Pacher & Kecskemeti, 2004), we believe that the bradycardia observed in our

117
study is due to an ANS response on the heart rather than a negative inotropic effect of FX
since we did not notice any significant changes in the electrolyte concentration (i.e: K+, Ca2+,
Na+) at the corresponding time points (data in appendix 4).
Given the fact that bradycardia is a result of an ANS response, it is expected that
HRV changes accordingly. Indeed, the time domain of HRV including SDNN, SD Delta NN,
RMSSD was significantly higher in the FX group compared to the control group at 1 and 2
mins after FX administration. However, the effects appeared to be limited to only two
minutes after FX. As it was shown in Fig. 4.5, there was an inverse relationship between the
changes in heart rate and the changes of time domain of HRV. The greater the decrease in
heart rate at 2 min post-FX, the greater the increase in SDNN, SD Delta NN and RMSSD at 1
min post-FX. In addition, the frequency domain of HRV (LF power, HF power, LF/HF ratio)
was comparable between the two groups at all time points. Acute and chronic SSRIs
exposure might exhibit different mechanisms. A recent longitudinal observation human study
from our group has found a reduced fetal heart rate variability (decreased number of
accelerations) in the fetuses exposed to SSRIs throughout most or all of gestation, even after
controlling for maternal mood, which might reflect an alteration in prenatal cardiac
autonomic control (Rurak et al., 2011).
A FX-induced elevated arterial pressure and mild bradycardia at 2 minutes after FX
injection were associated with a very mild and transient hypoxemia reflected by lower
arterial Po2 and O2 saturation values observed at 2 minutes post-FX in a smaller group of
lambs (n = 4) , but these changes were not statistically significant. However, there were no
significant changes in arterial pH, Pco2 nor lactate concentration at 2 minutes following FX
injection. Moreover, Po2 and O2 saturation at 5 minutes post-FX and later time point were

118
comparable with values observed before FX injection. Thus, transient hypoxemia with acute
FX injection was too minor to cause any respiratory acidemia. Collectively, respiratory
difficulties, a common feature of poor neonatal adaptation in human infants exposed to
SSRIs in utero, did not occur in the lambs with acute FX administration.
4.4.2 Acute FX Effects On HPA Axis Functions:
Similarly, FX did not cause significant changes in plasma cortisol concentrations in
the newborn lambs in our study. Also, no sex differences were found. To our knowledge,
there is limited information about the acute SSRIs effect on the neonate’s HPA axis as well
as the influence of maternal SSRI treatment on the fetal HPA axis. HPA axis function and the
central serotonergic system are known to be highly interrelated (Boadle-Biber et al., 1993;
Fuller, 1996a; Fuller, 1996b). In our previous sheep studies, 8-day late gestational FX i.v
infusion in the pregnant ewes resulted in an augmented prepartum rise in fetal ACTH and
cortisol levels and reduced low voltage rapid eye movement behavioral state in the fetus
(Morrison et al., 2004). In contrast, another human study from our group, which further
examined the long-term effects of prenatal SSRIs (fluoxetine) on the HPA axis in the
postnatal period, has shown an association between SSRI exposure and reduced basal cortisol
levels as well as an alteration in the HPA stress response pattern in the offspring at 3 months
of age (Oberlander et al., 2008). A lower cord blood cortisol level in SSRIs-exposed neonates
was also reported in a subsequent study (Davidson et al., 2009). Conflicting findings from
the sheep and human studies could be, in part, attributed to the differences in the timing and
duration of SSRI exposure. Even though there was no stressor stimulation included in our
study, it is unclear why the cortisol level in the lambs did not increase with acute FX
treatment as expected. It has been reported that acute administration of SSRIs increased the

119
secretion of several hormones including cortisol and ACTH (Raap & Van de Kar, 1999)
while chronic treatment with SSRI (citalopram) resulted in a decrease (Jongsma et al, 2005).
Although the mechanisms are not fully understood, alterations in HPA axis activity in the
fetus might be one of the possibilities underlying negative perinatal outcomes observed in
human studies. In addition, this is particularly important in the context of fetal exposure to
excess glucocorticoids either by prenatal stress or maternal illness (e.g: depression, anxiety
disorders) or pharmacologic agents effects (eg: SSRIs, synthetic glucocorticoids for lung
maturation in preterm labor), which can program the HPA axis in ways that it will have
negative impacts on mental, metabolic and cardiovascular health later in life (Lupien et al.,
2009; Seckl, 1997; Tamashiro & Moran, 2010).
4.4.3 Underlying Mechanisms Of Poor Neonatal Adaptation:
There is an ongoing debate in the literature regarding the appropriate mechanism for
the observed poor neonatal adaptation, whether the symptoms reflect a true withdrawal
syndrome after abrupt discontinuation of transplacental transfer of maternal SSRI at birth or
are a result of serotonin toxicity due to SSRI exposure in the newborn (Haddad et al, 2005;
Isbister et al., 2001; Stiskal et al, 2001). Clearly, it is difficult to differentiate the syndrome
based simply on clinical signs and symptoms since the symptoms appear to be overlapped
between an SSRI withdrawal syndrome and serotonin toxicity. Thus, measurement of the
plasma concentrations of the administered SSRIs at delivery and at the time the neonate was
symptomatic would help in determining whether it is a withdrawal or toxicity. For example, a
high level of SSRIs concentration at the time of peak symptoms and a decline thereafter
would suggest a toxicity phenomenon. In contrast, if drug concentration is undetectable at the
time the symptoms are present, it is more likely a case of withdrawal syndrome. There is

120
evidence supporting a withdrawal syndrome in the neonates after in utero SSRIs exposure
(Jaiswa et al., 2005). However, most of the withdrawal syndromes occurred more often with
paroxetine use than with other SSRIs (Jaiswal et al., 2003; Sanz et al., 2005), likely due to its
short half life and no active metabolite (Hiemke & Hartter, 2000). Since FX has a very long
elimination half life, it is is unlikely that there is a withdrawal syndrome with FX use. In
addition, there is no evidence for an ability to metabolize FX in the fetal lamb and human and
most importantly, it has been shown that the plasma FX concentration measured in human
infants at day 2 following birth is the same as that in cord blood at delivery (Laine et al.,
2003,Kim et al., 2006). Hence, it is less likely, we believe, that a withdrawal syndrome is an
underlying mechanism for poor neonatal adaptation after in utero FX exposure.
On the other hand, there is also evidence supporting the serotonin toxicity mechanism
in the “poor neonatal adaptation” in neonates who were prenatally exposed to SSRIs (Jessel
& Stiskal, 2004; Knopper et al.,2006; Kwon & Lefkowitz, 2008; Laine et al., 2003; Rampono
et al.,2004). These studies reported measureable concentrations of SSRIs (PX, FX and NFX)
in cord blood at birth and/or when the symptoms were present during the neonatal period. In
our study, we used a different approach to determine if serotonin toxicity is a mechanism
responsible for poor neonatal adaptation in the neonates. The study allowed us to reproduce
in the newborn lambs a similar drug level observed in the human neonates and the clinical
effects were recorded. As described above, the FX level in the lambs and in human infants
cover the same range. However, no symptoms related to poor neonatal adaptation (feeding
behavior, rest- activities cycle, irritability, respiratory difficulty) or other serotonergic
symptoms (restlessness, myoclonus, tremor, shivering) were observed in our exposed lambs.

121
Thus, our results do not favor serotonin toxicity as an underlying mechanism for poor
neonatal adaptation.
To date, it appears that there is a growing body of evidence supporting the view that
SSRI-elicited alterations in fetal brain development may be responsible for poor neonatal
adaptation. As were mentioned above, supporting evidence include the initial study in sheep
from our group and subsequent findings in human which showed a decrease of fetal REM
sleep, an important determinant of brain development, in FX exposed fetuses (Morrison et
al., 2001; Mulder et al., 2011; Richardson, 1994). In addition, a recent study demonstrated
that the risk of respiratory distress, a common symptoms in poor neonatal adaptation, was
reduced by stopping SSRI treatment group before the end of pregnancy compared to the
group in which SSRI therapy continued until delivery, after controlling for maternal illness
severity, suggesting that the mechanism is less likely due to either withdrawal or toxicity
syndromes, (Warburton et al., 2010). Furthermore, it is known that serotonin plays an
important role in the regulation of early brain development. It autoregulates serotonergic
neuron growth in culture and the development of related neural systems (Whitaker-Azmitia,
2001). Thus, alteration the fetal serotonin level in the CNS secondary to prenatal SSRI
exposure could potentially influence serotonin signaling pathways in a developing brain,
affecting the fetal serotonergic tone and finally lead to changes in fetal/neonatal behavior
expressing as poor neonatal adaptation (Oberlander, 2012).
In conclusion, overall our results have shown that there is a lack of significant FX
effects on cardiovascular-respiratory, behavioral and endocrine functions in ~ 4–day old
postnatal lambs with acute FX administration. Thus SSRI toxicity is unlikely to be the
mechanism underlying poor neonatal adaptation in human infants exposed to this drug in

122
utero. Further investigation is needed to determine the exact mechanism to provide more
sufficient information regarding the safety of FX and similar drugs in pregnancy

123
5. Acute Fluoxetine Effects In The Postnatal Lambs (From 10 Days To 1
Year Of Age)
5.1 Introduction:
There is increasing awareness that children and adolescences also suffer from
depressive disorders, which were formerly thought only to occur in adults. In fact, the
prevalence of depression ranges from 0.4 to 2.5% in children and 0.4-8.3% in adolescents
(Birmaher et al., 1996; Fleming & Offord, 1990). And the cumulative incidence of major
depressive disorder (MDD) during adolescence is similar to the lifetime prevalence of MDD
in the adult population, which ranges from 15-20% (Birmaher et al., 1996). In addition,
teenage girls are at higher risk for developing depression than teenage boys, with a male:
female ratio of 1:2, similar to adults (Kessler et al., 1993; Cohen et al., 1993; Nolen-
Hoeksema & Girgus, 1994). These data support the view that depression in adults might
actually originate during the adolescent period (Lewinsohn et al., 1999; Rao et al., 1999).
Depression that occurs during such a critical period of development can lead to impaired
school functioning, disturbed peer and parent relationships and increased rates of health
problems or somatic complaints, as well as long-term serious consequences due to an
increase risk of engaging in high-risk behaviors such as smoking, alcohol and other substance
abuses and even suicide attempts (Asarnow et al., 2005; Bernstein et al., 1997; Deykin et al.,
1992; Egger et al.,1999). Since SSRIs were introduced into the market, due to their safety
profile and benign adverse effects compared to the earlier antidepressants, they have also
been used in the treatment of depression in children and adolescents. However, SSRI side
effects testing has mostly been done in adults; thus their safety in the pediatric population is

124
not well-established. In their meta-analyses, Whittington et al., (2004) and Jureidini et al.,
(2004) concluded that the benefits from SSRIs treatment in children and adolescents are
offset by the significant risks of serious adverse events (i.e suicidal behavior), except for FX.
Thus, cognitive-behavioral therapy (CBT) appeared to be a better approach in regard to
efficacy and safety (Harrington et al.,1998). Nevertheless, some subsequent studies have
shown that it is more efficious to treat MDD in adolescent with CBT and FX in combination
compared to either therapy alone.(Goodyer et al., 2007; J. March et al., 2004; March et al.,
2007; Pampallona et al., 2004). However, the availability of CBT is limited in some areas,
thus SSRIs can be prescribed in these circumstances. Currently, only FX and escitalopram
are approved by the FDA for the treatment of depression in adolescents (Hosenbocus &
Chahal, 2011; Ronsley et al., 2010). In Europe, FX is approved by the European Medicine
Agency for children and adolescents with moderate to severe symptoms but only after failure
to respond to 4-6 sessions of psychotherapy (Eaton, 2006). In Canada, Health Canada has not
yet approved any SSRIs for use in children and adolescents because of the concerns about the
adverse effects of these agents, particularly suicidal thoughts and behaviors. Hence, any use
of SSRIs in this population is off-label (Voysey, 2004). Even though FX and escitalopram
were approved for use in the US, the FDA did issue a warning regarding the use of SSRIs in
the treatment of depression in adolescents (2007 > FDA proposes new warnings about
suicidal thinking, behavior in young adults who take antidepressant medications; Public
health advisories (drugs) > suicidality in children and adolescents being treated with
antidepressant medications). Besides the potential increased risk of suicidaity associated
with the use of these agents in children and adolescents, another lethal adverse reaction is the
serotonin syndrome. This syndrome is a relatively rare but lethal condition and is a result of

125
overstimulation of peripheral and central serotonergic systems. It can occur at high SSRI
doses, overdose or even at therapeutic dose when combined with other serotonergic
medications (eg: TCA, MAOIs, dextromethorphan, lithium) (Nelson et al., 2007). As
antidepressants are used in patients at high risk for suicide, the risk of intentional overdosing
of the drugs is high. In fact, it was reported that among prescription drug overdoses managed
by poison centers in the US, antidepressant overdoses are the most common (Nelson et al.,
2007). In addition, children less than 6 years of age accounted for 17% of all cases of
inappropriate SSRI ingestion. (Nelson et al., 2007) Since there is still limited information on
adverse effects (other than suicide behaviors) of SSRIs use in children and adolescences,
especially when the drug is used off-label, there is a compelling need for the study of the
safety of SSRIs use in such a special population. Therefore, in this study our aim is to
investigate the acute effect of FX on a variety of physiological, behavioral and endocrine
variables in the lambs at different ages from 10 days to 12 months old.
5.2 Methods:
Animal housing, surgical procedure, experimental protocol and methods are
described in detail in chapter 2.
In this chapter, an additional behavior study was performed in the lambs at 6 months
of age in which lambs were not restricted to the monitoring pen for sample collection.
Instead, lambs were allowed to move freely in the pen with their penmates (peers or siblings)
and have access to food and water ad libitum. Sterile water or FX 1mg/kg were given via
their jugular vein catheter or by venouspuncture around 10am and their behavior were
observed by a continuous DVR system to look for any abnormal behaviors. Through DVR

126
observations, their activities were divided into lying down asleep, lying down awake,
standing moving, standing quiet, walking and headbutting. The activities were collected from
the DVR system in 15 second epochs and then matched with the activity score from the
lamb’s actiwatch in the same epoch to provide the intensity for each corresponding activity.
In the lambs from 10 days old onward, cortisol and ACTH measurements in the FX
group were measured in the 3 month and 1 year old groups only.
Statistical analysis:
Differences in the effects of treatment and sex on behavior activities and weight gain
were determined using two-way repeated measure ANOVA. In general, three way ANOVA
was used to determine the effects of treatment, time and sex on cardiovascular-respiratory,
metabolic, endocrine variables. Two-way repeated measured ANOVA or Mann Whitney U
test were used for the non-normally distributed variables to determine the effect of treatment
and time. Delta O2 saturation, which is the difference between O2 saturation at 5 min after
dosing and that at -15 min before sterile water or FX administration, was calculated for each
animal in the control and FX group. Then delta O2 saturation between the control and FX
group was compared using the Mann Whitney U test (in the 1 and 6 months old lambs) or 2
way ANOVA (in the 1 year old lambs). For the data presented as percentage, statistical tests
were run on the raw data, not on % data, except for O2 saturation. P < 0.05 defines significant
differences. Data are presented as mean ± SEM. All data were analyzed using the Sigma Plot
11 statistics software (Systat Software, Inc; San Jose, CA, USA).

127
5.3 Results:
Actiwatch activities, arterial pressure, heart rate, heart rate variability, arterial blood
gases (O2 sat, pO2, pCO2, pH, BE, HCO3), lactate, glucose and Hb concentration in FX
experiments in the lambs at all age groups are summarized in the table in the appendices.
5.3.1 Acute FX Effects On Cardiovascular-Respiratory Functions:
There were no significant changes and no gender differences in arterial pressure,
heart rate and HRV variables with acute FX administration in the lambs at 10 days, 1, 3 and 6
months of age even though arterial pressure tended to increase for the first 5 min post-FX in
the lambs at these ages. (data in appendices)
However, in the 1 year old lamb, arterial pressure significantly increased after FX
administration and this effect lasted for about 15-30 mins (Fig. 5.1). Mean arterial pressure
were significantly different between the control and FX group at 1,2,3,4,5, 15 and 30 minutes
post-FX (p<0.001), but not at the control time point (-15 minutes). Pressure increased from
85.1 ± 2.2 mmHg before FX to peak value of 106.6 ± 4.1 mmHg at 2 mins after FX
administration, thus the maximum increase was 21.5 ± 4 mmHg. It is noted that the increase
in mean arterial pressure was more profound and lasted longer in males than in females.
Statistical analysis indicated that the difference in arterial pressure at 2 and 3min after
dosing vs -15min before dosing were only significant in male lambs (p = < 0.001 and p =
0.034). At the 2 min sample, the mean arterial pressure increase was significantly higher in
males than in females (to 112.6 ± 4.1 mmHg in male vs 99.1 ± 5.2 mmHg in female; p =
0.012), with a maximum increase of 25.8 ± 4.4 mmHg in male compared to 16 ± 6.8 mmHg
in female.

128
FIG 5.1: Arterial pressure and heart rate changes in 1 year old lambs with acute FX administration a indicates significant differences between variables at -15 min vs 1,2,3,4,5,10,15 min after FX within the FX group b indicates significant differences between control and FX at the corresponding time c indicates significant differences between male and female at the corresponding time within FX group
a a b b b
a a a a a
b b b b b b b
c

129
There was a transient decrease in heart rate at 2 min after FX (p = 0.026), the
difference was 30 ± 4 bpm but it quickly returned to normal within 1 min. No sex difference
was found in the heart rate response. (Fig 5.1)
In terms of HRV, we did not find any significant difference between male and female
lambs at 1 year of age. Except for the LF/HF ratio, all other variables (SDNN, SD Delta NN,
RMSSD, LF power (ms2), HF power (ms2) significantly increased at 30 min after FX dosing
in the FX group compared with the control group (SDNN: p = 0.013, SD Delta NN: p =
0.002, RMSSD: p = 0.002; LF power: p = 0.01; HF power: p = 0.002). The difference
persisted for 60 min for some variables (SD Delta NN, RMSSD, HF power). (Fig 5.2)
In 4 male lambs at 1 year of age, delta arterial pressure (i.e the difference between
arterial pressure value at 2 min post-FX and that at -15 min pre-FX) and plasma log FX
concentration at 5 mins followed a linear regression with r2 = 0.84, p = 0.08

130
FIG 5.2: HRV (SDNN, SD Delta NN, RMSSD, LF power, HF power, LF/HF ratio) changes with acute FX administration in 1 year old lambs. a indicates significant differences between variables at -30 min vs 30,60,90 and 120 min after FX within the FX group b indicates significant differences between control and FX at the corresponding time
b b
b b
b b
a b
a b b

131
5.3.2 Acute FX Effects On Homeostatic And Respiratory Functions:
There was no FX effects on respiratory and metabolic function in the lambs at 10
days and 3 months of age even though Po2 tends to be declined at 5 min post FX in the 3
month-old lambs but this is not significant. (data in appendices)
However, Po2 significantly decreased at 5 min after FX injection in the 1 month-old
lambs (p < 0.001). The FX effect was significantly different between male and female (p =
0.002). In males, Po2 decreased from 92.4 ± 4.4 at 15 min pre-FX to 67.2 ± 4.1 mmHg 5 min
post-FX while in females it fell from 97.6 ± 2.5 at 15 min pre-FX to 92.5 ± 4.5 mmHg 5 min
post-FX. (Fig 5.3) There was also a statistically significant difference in delta O2 sat between
the control group (median = 0.625%) and the FX group (median = -3.35%) (p = 0.031) with
the Mann-Whitney U test. This indicates that FX caused a transient hypoxemia in lambs at 1
month of age and it was more apparent and severe in male than female lambs. Other blood
gas variables (pH, pCO2, HCO3-) and glucose and lactate concentrations were similar between
the control and FX groups in this age group.
Similarly, in the 6 month-old lambs, 2 way (treatment x time) repeated measure
ANOVA revealed a significant decrease of pO2 at 5 min post-FX compared to control (p =
0.035). Po2 fell from 111.9 ± 5.4mmHg at 15 min pre-FX to 95 ± 7.7mmHg at 5 min post-
FX. (Fig 5.4) Delta O2 sat was significantly different between the control group (median =
0.65%) and the FX group (median = -1.05%) (p = 0.004) with Mann-Whitney U test.
However, there were no significant differences in hypoxemia response to acute FX
administration between males and females. Other variables were also not significantly
different between the control and FX groups.

132
FIG 5.3: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 1 month of age. a indicates significant differences between -15 min and 5 min after FX within the FX group b indicates significant differences between control and FX within the corresponding time c indicate significant differences between male and female within the corresponding time Asterisk (*) indicates significant differences in delta O2 sat (i.e differences between values at 5 mins and at -15 mins) between control and FX group using Mann-Whitney u test
*
a b c

133
FIG 5.4: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 6 months of age. a indicates significant differences between -15 min and 5 min after FX within the FX group b indicates significant differences between control and FX within the corresponding time Asterisk (*) indicates significant differences in delta O2 sat (i.e differences between values at 5 mins and at -15 mins) between control and FX group using Mann-Whitney u test
*
a b

134
In the 1 year-old lambs, a transient decrease in Po2 occurred at 5 min after FX
injection and returned to normal by the time of 15min post- FX. Po2 fell from 110.5 ± 2.5
mmHg at 15 min pre-FX to 94 ± 5.8 mmHg at 5 min post-FX. Delta O2 sat was significantly
different between control and FX group and was more severe in male lambs (p = 0.037).
(Fig. 5.5) There was a significant inverse relationship between the changes in Po2 at 5 min
(delta Po2) and changes in arterial pressure at 2 min (delta AP) following FX injection with r2
= 0.54, p = 0.037, n = 8. The greater the increase in arterial pressure at 2 min post-FX, the
greater the decrease in Po2 at 5 min post-FX. (Fig. 5.6A) The significant relationship between
delta Po2 and delta arterial pressure is sex-specific, being significant in males (r2 = 0.93, p =
0.03, n = 4) but not in females (r2 = 0.66, p = 0.18, n = 4) (Fig. 5.6B). Similarly, changes in
O2 saturation (delta O2) at 5 min and delta AP were significantly correlated in male (r2 = 0.98,
p = 0.01, n = 4) but not in female (r2 = 0.27, p = 0.48, n = 4) (Fig. 5.6C). Also, there was a
significant relationship between delta pO2 (i.e. the difference between pO2 at 5 min post-FX
and that at -15 min pre-FX) and log plasma FX concentration at 5 min in lambs at 1 years of
age (r2 = 0.81, p =0.002, n = 8) (Fig. 5.7A). However, the relationship between delta pO2 and
log plama FX concentration at 5 min was not statistically significant in males (r2 = 0.75, p =
0.13, n = 4) and in females (r2 = 0.69, p = 0.16, n = 4) (Fig. 5.7B). Nevertheless, there was a
significant relationship between delta O2 saturation (i.e. the difference between O2 saturation
at 5 min post-FX and that at -15 min pre-FX) and log plasma FX concentration at 5 min in
both males and females with r2 = 0.92, p =0.040, n = 4 and r2 = 0.97, p =0.016, n = 4,
respectively. The higher the plasma FX concentration at 5 min, the greater the decrease in O2
saturation at 5 min following FX injection (Figure 5.7C).

135
However, pH value did not change significantly over time and was comparable
between the control and FX group. Pco2 (at -15 min), BE and HCO3 were significantly
different between the control and FX experiments and this occurred only in female lambs
(Fig. 5.8). Similarly, lactate (at 5 min post FX) and Hb (at 5 and 15 min post-FX) were
significantly higher with FX administration compare to placebo only in females (Fig. 5.9).
Glucose concentration was significantly different between male and female lambs in both
control and FX group (Fig. 5.9). There were no significant changes in temperature after FX
administration. (data in appendices)
Arterial blood gas samples were also taken at 2 mins after FX in 5 out of the 10 lambs
in this 1 year age group. Except for pO2 and O2 sat, which were also decreased as shown at 5
mins after FX (Fig. 5.5), there were no significant differences in other blood gas variables at
2 mins after FX compared with those at -15 mins before FX. However, pCO2 tended to
decrease and pH tended to increase at 2 mins post-FX compare with -15 min pre-FX (32.6 ±
1.6 vs 36.8 ± 2.1 mmHg; 7.531 ± 0.027 vs 7.486 ± 0.027, n = 5, respectively) although they
are not statistically significant.

136
FIG 5.5: pO2 and O2 saturation with sterile water and FX administration in male and female lambs at 1 year of age. a indicates significant differences between -15 min and 5 min after FX within the FX group b indicates significant differences between control and FX within the corresponding time Asterisk (*) indicates significant differences in delta O2 sat (i.e differences between values at 5 mins and at -15 mins) between control and FX group within male, and between male and female within FX group.
*
a b
A

137
FIG 5.6: (A) The relationship between changes in pO2 at 5 mins and changes in arterial pressure (AP) at 2 mins following FX injection in all lambs at 1 year of age (square symbol, solid line, n = 8); (B) The relationship between changes in pO2 at 5 mins and changes in AP at 2 mins following FX injection in male (square symbols, dotted line, n = 4) and female (circle symbols, solid line, n = 4) at 1 year of age and (C) The relationship between changes in O2 saturation at 5 mins and changes in AP at 2 mins following FX injection in male (square symbols, dotted line, n = 4) and female (circle symbols, solid line, n = 4) at 1 year of age.
A
B
C

138
FIG 5.7: (A) The relationship between changes in pO2 at 5 mins and log FX concentration at 5mins following FX injection in all lambs at 1 year of age (square symbol, solid line, n = 8) (B) The relationship between changes in pO2 at 5 mins and log FX concentration at 5mins following FX injection in male (triangle symbols, dotted line, n= 4) and female (circle symbols, solid line, n = 4) lambs at 1 year of age; and (C) The relationship between changes in O2 saturation at 5 min and log FX concentration at 5mins following FX injection in male (triangle symbols, dotted line, n= 4) and female (circle symbol, solid line, n = 4) lambs at 1 year of age.
A
B
C

139
FIG 5.8: PCO2 , base excess (BE), HCO3 concentrations with sterile water and FX administration in male and female lambs at 1 year of age.

140
FIG 5.9: Glucose, lactate, Hb (Hemoglobulin) concentrations with sterile water and FX administration in male and female lambs at 1 year of age.

141
5.3.3 Acute FX Effects On Behavior:
There were no differences in activity variables between the average of 3 normal days,
control group and the FX group in the ~ 10 day-old group. In contrast, activities significantly
decreased on the FX experimental day compare to both control experimental day and normal
day in the 1 month-old lambs. Starting from 3 months old, the lambs were kept in the
monitoring pen for 4 days during the control and FX experiments. Thus, their activities
decreased on both control and FX experiment day due mainly to the restraint. Summarized
data for all age groups are in tables 1, 2 in the appendices. To investigate the actual FX
effects on the older lambs (after 3 months old) we conducted a behavior study on the 6
month-old lambs in which they were housed in their regular holding pen with their sibling or
peers as usual, as described in the method section. The activity data of this behavior study is
presented in figures 5.15-5.17. Note that activity data of the 6 month-old group presented in
the appendices are the activities when the lambs were kept in the monitoring pen.
5.3.3.1 Behavioral Activities In The Lambs At 1 Month Of Age:
Active bouts per day, mean score in active and total activity score were significantly
lower on FX experimental day as compared to the average of 4 normal days (p<0.001) while
they were comparable between the other days (control experiment day, post FX day 1, post
FX day 2) and the average of 4 days. In contrast, episode duration was significantly higher on
the FX day compared to the average of the 4 normal days. Even though the percentage of
moving time was lower and the percentage of time immobile were higher on the FX day
compared to the average of 4 normal days, respectively, they were not statistically
significant. (Fig 5.10) There were no sex differences in these actiwatch variables.

142
Results from DVR observations showed that the intensity of suckling attempts
determined by actigraphy was significantly higher on the FX day compared to the normal day
(FX day: 327 ± 21 counts/15s versus normal day: 247 ± 28 counts/15s). The lambs always
attempted to suckle when they were returned to their mothers after every blood sampling but
the ewes almost always refused them. Thus, it is reasonable that the intensity of suckling
attempts was higher on the FX days as lambs tried harder. Importantly, there were no
significant changes in the percentage of time lamb spent on suckling attempts on the FX days
compared to normal days, which indicates that blood sampling did not interfere with their
feeding behavior. However, the amount of time lambs spent on successful suckling was
significantly lower on the day of the FX experiment (FX day: 0.7 ± 0.14 %/24h versus the
normal day: 1.1 ± 0.22 %/24h). In addition, the amount of time that lambs spent walking
significantly decreased on the FX experiment day (3.6 ± 0.2%/24h) vs the control day (4.9 ±
0.4 %/24h) (Fig 5.11A). Even though the time lambs spent standing was not significantly
different between the days, when dividing the standing activity into standing quiet (defined
by standing from DVR observation and actiwatch score less than 60) and standing mobile
(defined by standing activity from DVR observation and actiwatch score equal or greater
than 60), there was a significant decrease in the amount of time lambs spent standing moving
on the FX experiment day (8.9 ± 0.9 %/24h) compared to the control experiment day (13.7
±1.5 %/24h) or normal day (13.7 ± 1.3 %/24h), (p = 0.013). Moreover, the intensity of
standing quiet was also significantly lower on the day of FX experiment (28 ± 2.8
counts/15s) versus control day (34 ± 2.3 counts/15s) (Fig 5.12). Although the time lambs
spent for experiments was significantly higher on the control and FX day compared to
normal day, it was negated by the significantly lower intensity of activity during the

143
experiment on experiment days compared to normal days (Fig 5.11). In fact, lambs slept or
lay awake most of the time during the experiments. Collectively, the results suggest that 1
month-old lambs were less successful at suckling and less active (walking less, moving less
and more still while standing) with FX administration. However, their weight gain was
comparable between the average of normal days, control, FX experiment and post FX days.
(Fig 5.13)
Regardless of the day, there were a sex difference in the intensity of standing and
playing activity. Female lambs had a higher average score of standing activity (normal day:
102 ± 2.2, control day: 110 ± 11, FX day: 93 ± 16 counts/15s) than the male (normal day: 74
± 6.6, control day: 75 ± 8.2, FX day: 63 ± 3 counts/15s). Similarly, there was higher playing
activity in females than in males (normal day: 272 ± 33 vs 178 ± 44; control day: 232 ± 43 vs
164 ± 18, FX day: 265 ± 15 vs 174 ± 31 counts/day) (Fig 5.14). In general, around 1 month
of age female lambs were more active than male lambs but this was independent of FX
administration.
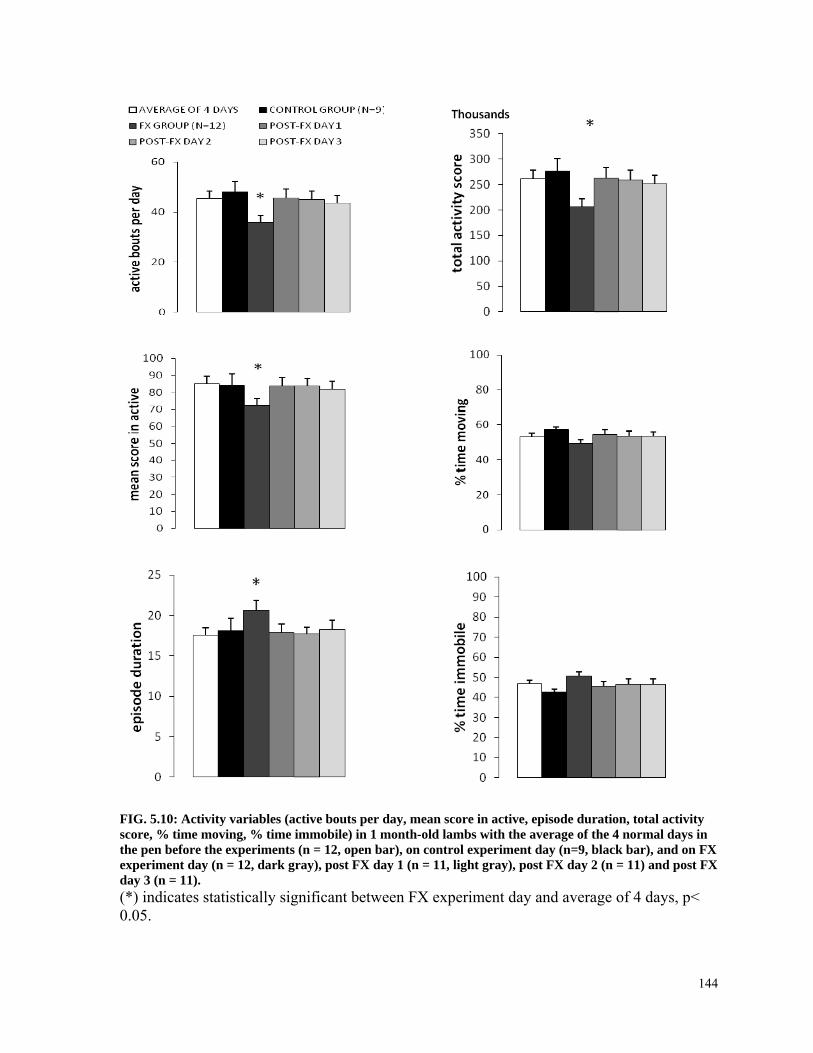
144
FIG. 5.10: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in 1 month-old lambs with the average of the 4 normal days in the pen before the experiments (n = 12, open bar), on control experiment day (n=9, black bar), and on FX experiment day (n = 12, dark gray), post FX day 1 (n = 11, light gray), post FX day 2 (n = 11) and post FX day 3 (n = 11). (*) indicates statistically significant between FX experiment day and average of 4 days, p< 0.05.

145
FIG. 5.11: (A) Percentage of time lambs spent for each activity from DVR observation and (B) the intensity of each activity when matching DVR observation with actiwatch score data in lambs at 1 month of age. Normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). (^) indicates significant differences between control experiment day and normal day (+) indicates significant differences between FX experiment day and control experiment day (*) indicates significant differences between FX experiment day and normal day, p < 0.05.
A
B

146
FIG. 5.12: (A) Percentage of time lambs spent for standing activity from DVR observation and (B) the intensity of standing quiet and standing moving when matching DVR observation with actiwatch score data in lambs at 1 month of age. Normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). (^) indicates significant differences between control experiment day and normal day (+) indicates significant differences between FX experiment day and control experiment day (*) indicates significant differences between FX experiment day and normal day, p < 0.05.
A
B

147
FIG 5.13: Weight gain (mean ± SE) in the lambs during their normal days, control experiment with distilled water injection, FX experiment with FX injection and 3 continuous days after the FX experiment day around 1 month of age.
FIG. 5.14: Intensity of standing and playing activity from DVR observation in the 1 month-old male and female lambs on normal day (day lambs in the pen without any experiments, gray bar, n = 3), on Control experiment day (n=6, open bar) and FX experiment day (n=6, black bar). (a) indicates significant differences in standing activity on each corresponding day between male and female. (b) indicates significant differences in playing activity on each corresponding day between male and female. p < 0.05.

148
5.3.3.2 Behavioral Activities In The Lambs At 6 Months Of Age:
Active bouts per day, total activity score and % time moving were significantly lower
on the FX experimental day and post FX day 1 as compared to the average of 4 normal days
(p = 0.012, p = 0.007, respectively) while they were comparable between the other days
(control experiment day and post FX day 2) and the average of 4 days (Fig. 5.15). In contrast,
% time immobile was higher on the FX day and post-FX day 1 compared to the average of 4
normal days. Mean score in active tends to be lower and episode duration higher on the day
of FX experiment but these changes were not statistically significant. (Fig 5.15) There were
no sex differences in these activity variables from the actiwatch measurements.
The results from the DVR observations showed that there was a small but significant
increase in the amount of time lambs spent on headbutting on the day of the FX experiment
(0.8 ± 0.17 %/24h) compared to the day of control experiment (0.39 ± 0.09 %/24h) (p =
0.036) but not significant to the normal day (0.47 ± 0.09 %/24h) (Fig 5.16A). Even though
the amount of time spent for standing was not significantly different between the days, when
dividing the standing activity into standing quiet (defined by standing from DVR observation
and actiwatch score less than 60) and standing mobile (defined by standing activity from
DVR observation and actiwatch score equal or greater than 60), there was a significant
decrease in the amount of time lambs spent for standing moving (FX: 14.4 ± 1.51%/24h,
control: 21.2 ± 2.9 %/24h; normal: 21.2 ± 3.06 %/24h) and an increase in the time standing
quiet (FX: 18.2 ± 1.53 %/24h, control: 12.2 ± 1.44 %/24h, normal: 13.6 ± 2.02 %/24h) on the
FX experiment day compared to the control experiment day or normal day (p = 0.027 and p =
0.005, respectively) (Fig. 5.17). In addition, the intensity of standing activity was
significantly lower on the day of FX experiment (68 ± 7.2 counts/15s) compare to the normal

149
day (102 ± 15 counts/15s) (p = 0.042) (Fig. 5.17B) and this difference was due to the
significant differences in the intensity of standing quiet (FX day: 12 ± 1.4 counts/15s, control
day: 18 ± 1.9 counts/15s, normal day: 18.9 ± 1.9 counts/15s) (Fig 5.17). These results
suggested that after FX administration the lambs’ behavior were shifted toward two extremes
of activity: they were less active, stood very still for longer time, but were also more
aggressive indicated by an increase in headbutting activity. It was noted from DVR
observations that two lambs (one male and one female) presented with jerky movements
from around 20.5h after FX administration. These jerky movements occurred intermittently
in the lambs for ~10h during the day. However, no differences in the intensity and the % time
of each activity were found between male and female lambs. Unfortunately, no blood
samples were available for the measurement of FX concentration in these lambs.
Interestingly, at 6 months of age, acute FX administration resulted in a significant
weight loss in the lambs (-0.7 ± 0.19 kg, n = 8) as compared to an average of daily weight
gain of 0.1 ± 0.05 kg at their age (p = 0.011). (Fig 5.18) This FX effect on bodyweight gain
was not observed in the younger lambs.

150
FIG. 5.15: Activity variables (active bouts per day, mean score in active, episode duration, total activity score, % time moving, % time immobile) in 6 month-old lambs with average of 4 normal days in the pen before the experiments (n = 8, open bar), on control experiment day (n=8, black bar), and on FX experiment day (n = 8, dark gray bar), post FX day 1 (n = 8), post FX day 2 (n = 8) and post FX day 3 (n = 8). Asterisk (*) indicates statistically significant between FX day or post FX day 1 and average of 4 normal days, p< 0.05.

151
FIG. 5.16: (A) Percentage of time lambs spent for each activity from DVR observation and (B) the intensity of each activity when matching DVR observation with actiwatch score data in lambs at 6 month of age. The normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). (+) indicates significant differences between FX experiment day and control experiment day (*) indicates significant differences between FX experiment day and normal day, p < 0.05.
A
B

152
FIG. 5.17: (A) Percentage of time lambs spent on standing (quiet and moving) activity from DVR observation and (B) the intensity of standing quiet and standing moving when matching DVR observation with actiwatch score data in lambs at 6 month of age. The normal day was a day lambs were in the pen without either control or FX experiment (n = 6, gray bar). Control experiment (n=6, open bar), FX experiment (n=6, black bar). (+) indicates significant differences between FX experiment day and control experiment day (*) indicates significant differences between FX experiment day and normal day, p < 0.05.
A
B

153
FIG 5.18: Weight gain (mean ± SE) in the lambs during their normal days, control experiment with distilled water injection, FX experiment with FX injection and 2 continuous days after the FX experiment day at around 6 months of age. (*) indicates significant differences between FX experiment day and the normal average of 4 days, p < 0.05.
5.3.3.3 Acute FX Effects On Cortisol Level:
No differences were found between the cortisol concentrations in the control group
compared with the FX group at 3 month and 1 year old, nor ACTH concentration. (Figs 5.19
& 5.20)
There were also no differences in cortisol level between sexes.

154
FIG. 5.19: Cortisol and ACTH concentration in the lambs at 3 months of age at 15, 30 minutes before and 0.5, 1, 2, 6 and 12 h after sterile water injection (control group) and FX injection ( FX group)

155
FIG. 5.20: Cortisol and ACTH concentration in the lambs at 1 year of age at 15, 30 minutes before and 0.5, 1, 2, 6 and 12 h after sterile water injection (control group) and FX injection (FX group)

156
5.4 Discussion:
In this study which included lambs at 10 days, 1, 3, 6 month and 1 year of age, we
found that acute i.v. FX administration did not have any significant effects in the lambs at 10
days and 3 months of age. However, transient hypoxemia occurred in the 1 month old lambs
with acute FX administration and it was more profound in males. Lambs were also less active
and altered their feeding behavior on the day after FX administration. These FX effects on
respiration and behavior were also observed in the lambs at 6 months of age but there were
no sex differences. At 1 year of age, acute FX administration resulted in an increase in
arterial pressure, to a greater extent in the male than the female, and a transient decrease in
heart rate. HRV was also altered with FX administration for about 30-60 minutes in both
sexes. No changes in cortisol level occurred with acute FX administration.
The daily therapeutic FX dosing for children from 6 to 18 years old ranges from 10 to
20 mg but no mg/kg dose has been provided (Eaton, 2006). However, according to a
reference used by poison control centers in the US, a minimum toxic dose of FX in children
less than 6 years of age is 60 mg or 5mg/kg (Nelson et al., 2007). In our study 1mg/kg FX
dose was used, which is at the high margin of the therapeutic range but does not reach a
minimal toxic dose. Moreover, the plasma FX concentrations achieved in the lamb
experiments were in the therapeutic FX concentration range in humans (Chow, 2013).
In our study, increase in arterial pressure and decrease in heart rate after acute FX
administration were observed in the older lambs. As the mechanism of SSRI group, plasma
5-HT concentration is expected to increase after FX bolus by the inhibition of 5-HT reuptake
by platelets, the main reservoir of peripheral 5-HT (Maurer-Spurej et al., 2004; Ortiz &

157
Artigas, 1992). 5-HT has been known as a vasoconstrictor agent, its increase would lead to
an elevated blood pressure (Maurer-Spurej et al., 2004). Even though FX seems to have
divergent effects on blood pressure, (Lazartigues et al., 2000; Tsai & Lin, 1986) did report
that central acute administration of FX caused hypertension in rats in a dose-dependent
fashion. In humans FX is usually administered for longer time durations (several weeks) per
os and an increase in blood pressure was also observed in those subjects who had a normal
pretreatment mean diastolic pressure (Amsterdam et al., 1999). The diversity of 5-HT effects
on blood pressure in the literature could be attributed to many factors such as differences in
species, route of administration, doses, and 5-HT receptor subtypes. However, elevated
arterial pressure after acute FX injection in the older lambs in our study was consistent with a
report by (Nelson et al., 1987) in which increase in blood pressure was observed with 5-HT
injection in conscious sheep.
However, our study is probably one of the first to examine blood pressure responses
to FX administration over age in relation to gender. The effect of FX on arterial pressure
elevation only became statistically significant in lambs at 1 year of age and it also increased
higher in males compared to females. It is not yet clear why the FX effects on blood pressure
was only significant at 1 year of age. However, one explanation could be due to either a
lower level of platelets, which are the main reservoir of peripheral 5-HT, in the younger
lambs or a lesser extent of a functioning 5-HT receptor subtypes that mediate 5-HT effects.
Platelet concentrations are lower in human newborns and children compared to adults
(Merritt & Davidson, 1933) and in addition, the platelet 5-HT concentration in newborns is
only ~42% of the value in children and adults (Flachaire et al., 1990). If a similar situation
occurs in sheep, the transient increase in blood 5-HT following SSRI administration may

158
have been less in the younger lambs and not sufficient to elicit effects on arterial pressure and
blood oxygenation. Measurement of platelet concentrations, platelet 5-HT concentration and
the magnitude of the transient increase in blood 5-HT concentration following SSRI
administration in sheep as a function of age could confirm this hypothesis.
In terms of the sex differences in the FX effects on arterial pressure in the year old
lambs, the lung is a major site of clearance of plasma 5-HT, with ~95% of 5-HT being
removed from plasma by the lung via a first-pass effect (Gillis & Pitt, 1982). However, a
small change in the 5-HT clearance rate by the lung can dramatically affect of the systemic
levels of 5-HT, for example increasing pulmonary extraction from 95 to 97.5% would reduce
plasma 5-HT concentration in the systemic circulation by 50%. (Gillis & Pitt, 1982) Estrogen
(17ß estradiol) or progesterone appears to increase 5-HT metabolism in lung homogenates
prepared from female rats (Bakhle & Ben-Harari, 1979). By one year of age, male and
female lambs who are raised indoors under the same condition as ours have undergone
puberty (Suttie et al., 1991) and the females have thus initiated estrous cycles (Ryan et al.,
1991; see Chapter 3), with pulsatile increases in blood estrogen and progesterone levels.
Thus it is possible that an effect of ovarian steroids on increasing 5-HT clearance by the lung
is responsible for the observed smaller increase in the arterial pressure response to FX
administration in the 1 year old female lambs compared to male lambs, because of a lesser
transient increase in blood 5-HT levels following FX injection. It has been reported that the
rate of sustained hypertension is higher in adults who underwent 12 weeks of venlafaxine (a
serotonin and noradrenaline reuptake inhibitor) treatment for depression compare to placebo
group (Feighner, 1995). In general, SSRIs are not considered to be cardiotoxic, in
comparison to the classical antidepressants (TCA, MAOI) (Lane et al., 1995). However,

159
hypertension is one of the symptoms included in serotonin syndrome (Boyer & Shannon,
2005) and the serotonin syndrome has been reported in pediatric and adolescent populations
after SSRI administration (Asch & Parker, 1988; Gill et al., 1999). However, we did not
observe hyperthermia after FX injection in the lambs in our study (data in appendices), and
hyperthermia is also a symptom of the serotonin syndrome (Boyer & Shannon, 2005).
In the 1 year old male lambs the magnitude of increase in arterial pressure at 2 min
following FX injection (i.e: peak value) and plasma FX concentration at 5 mins followed a
linear regression analysis. Even though the relationship was not statistically significant (p =
0.08) because of the small sample size (n = 4), it to some extent implies a correlation
between FX concentration and its effect.
Interestingly, we found HRV variables (SDNN, LF power, SD Delta NN, RMSSD,
HF power) increased for the first 30 mins after acute FX administration in the older lambs. In
the literature, the effects of SSRIs on HRV are still a matter of debate. It has been established
that HRV decreases in depressed patients, which increases the risk of cardiovascular disease
and mortality in these patients (Carney et al., 2005; Grippo & Johnson, 2002; Thayer &
Lane, 2007). It has also been shown that TCA treatment exacerbated the decreased HRV in
depressed patient which could contribute to the adverse cardiovascular side effect of TCA
(van Zyl et al., 2008). However, there is more controversy when it comes to SSRIs effect on
HRV. Recent publications reported an “unfavorable” decrease (Licht et al., 2008; Licht et
al.,2010) or unchanged of HRV (Kemp et al., 2010) with SSRIs treatment in depressed
patients. However, in a meta-analysis done by van Zyl et al., (2008), SSRI use was
associated with a slight decrease in heart rate and increase in SDNN. Among SSRIs included
in this meta-analysis, FX was investigated in only one study by Khaykin et al, which reported

160
an increase of SDNN (Khaykin et al., 1998; van Zyl et al., 2008). This finding is consistent
with our HRV results, at least in terms of the SDNN variable. In addition, we did observe
increases in other HRV variables (SD Delta NN, RMSSD, HF power, LF power) after acute
FX administration. However, in the present study, it is unclear why both LF power and HF
power increase at the same time. One possibility is that FX might increase sympathetic tone
but, in parallel, increase vagal tone and thus minimize the heart rate changes.
However, the significant increase in HRV was only observed in the lambs at 1 year of
age and not in the younger lambs. Given the developmental aspects of the lambs, they were
deemed to be an adult at 1 year old. Therefore our results suggested the HRV response to
acute FX is age-dependent in parallel with the maturation of the autonomic system and
adolescents might respond differently to the drug as compared to the adult.
In vivo studies have shown an opposite respiratory response to acute and chronic
SSRI treatment. While acute SSRIs (FX, PX) administration caused a reduction in respiratory
rate and minute volume in alert, freely moving rats; chronic treatment for more than 23 days
in those rats led to an increase in respiratory rate (Annerbrink et al., 2010). In our study, we
did not record respiratory rate, instead the effect of FX on respiratory system and
homeostasis was observed through the changes in the lambs’ arterial blood gas status. We
found a decline in pO2 and O2 saturation in the lambs at 1, and 6 months and 1 year of age at
5 mins after FX administration. However, there were no significant accompanying changes in
pCO2, pH, HCO3- and lactate concentration after FX. FX and other SSRIs agents when given
acutely will lead to an acute increase in synaptic levels of serotonin by blocking serotonin
reuptake (Stahl, 1998). There is evidence to suggest that serotonin might exert both
stimulatory and inhibitory influences on respiration. (Annerbrink et al., 2003; Mulkey et al.,

161
2004) Thus the observed reduction in pO2 and O2 sat after FX administration could be a result
of respiratory inhibition due to an increase in synaptic serotonin concentrations. Although
due to technical difficulties we did not succeed in an attempt to measure plasma serotonin
following FX administration in the lambs in our study, a study in rat has shown that after
acute SSRI (FX) administration plasma serotonin increased 520% as compared to the control
(Ortiz & Artigas, 1992). The increase in serotonin also occurred in previous study in our lab
in which plasma serotonin level increase for the first hour after the bolus of loading FX dose
in 3 adult non-pregnant ewes (Morrison, 2001). In addition, in the present study, we found
that there was a significant relationship between the level of hypoxemia at 5 mins post-FX
and the magnitude of increase in plasma FX concentration at 5 mins in both male and female
lambs at 1 year of age. The higher the level of FX, the greater the decrease in O2 sat or
hypoxemia in the 1 year old lambs. This again confirms the direct effects of FX on
physiological changes. As it was mentioned above, a study in rats showed that the effect of
acute FX on reduced respiratory rate only occurred at the dose of 10mg/kg to achieve mean
plasma FX = 1092.8 ± 315.9mmol/L, NFX = 1203.2 ± 308.2 mmol/L and a more reduced
respiratory rate at a higher dose of 30mg/kg while there was no FX effect at the dose of 1
mg/kg. (Annerbrink et al., 2010) Similarly, in a human study, SSRI effects on fetal behavior
were only observed in the fetuses exposed in utero to the drug at standard and high doses in a
dose-dependent manner but not in the low dose group (Mulder et al., 2011). Roca et al.,
(2011) also reported an increased risk of preterm birth mainly occurred in women treated
with high dose SSRIs during pregnancy.
Another possibility is that hypoxemia occurred as a result of acute increase in
serotonin which caused vasoconstriction and sudden increase in blood pressure. This could

162
result in a decrease in cardiac output due to an afterload effect, which would reduce
pulmonary blood flow to result in the decrease in arterial pO2. In fact, our data have shown
that the magnitude of increase in arterial pressure at 2 min following FX injection was
significantly related to the magnitude of hypoxemia reflected by a decrease in Po2 at 5 min
following FX in the 1 year old lambs supporting for this hypothesis. The relationship was
stronger in males than in females. In addition, an experimental study in dogs showed that
pulmonary arterial pressure increased after 20-30 mins of i.v serotonin infusion (Sackner,
Will, & DuBois, 1966), which suggests pulmonary vasoconstriction. The decrease in pO2
could via chemoreceptor activation lead to hyperventilation leading to a decrease in arterial
pCO2 and increase in pH. This is consistent with the trend of pCO2 and pH changes observed
at the 2 min sample in the 1 year old lambs. However, we did not record respiratory rate and
so could not determine if in fact hyperventilation did occur, and thus explain the unchanged
pCO2 and pH at the 5 min sample.
It is unclear why hypoxemia after FX injection only occurred in the lambs at 1, 6, 12
months of age but not at the other ages (3, 10 days and 3 months of age) and why it was more
severe in male than female at 1 month of age. However, it appeared that FX-induced
hypoxemia was coincident with milestone events in their development (i.e weaning, puberty).
Other factor(s) which are associated with these events might contribute to the observed FX
effects on respiratory and homeostasis in the lambs.
Behavior studies in which lambs were not restrained to the monitoring pen were
performed in lambs at 3, 10 days and 1, 6 months of age. In general, at 1 and 6 months of
age, lambs were significantly less active on the day after FX administration as shown from
the actiwatch data (Figs 5.9 & 5.14). The reduction in their overall activity could be due to

163
either the fall in the intensity of high score activities (such as standing moving, walking,
suckling and playing or headbutting) and/or a reduction in the time they spent for these high
score activities over the day. Using a combination of DVR observations and actiwatch score
data, we found that the reduction in the 1 month old lambs after FX injection was a result of
both the reduction in the intensity of standing quiet and a decline in the time lambs spent for
standing moving, walking and successful suckling. One question is whether this reduction in
activity following FX administration at 1 month has any relation to the normal reduction in
lambs’ activity observed around 1 month of age due to weaning onset. Our developmental
study in chapter 3 showed that the 1h rhythm due to suckling is a prominent rhythm in lambs
for the first month of life. That means their main state for this first month of life was simply
either sleeping or suckling. Our results suggested acute FX administration did not disrupt
their feeding activity before 1 month since there were no significant differences in activities
in experiments at 4 and 10 days of age. In parallel of the disappearance of the 1h rhythm
around 1 month, a circadian rhythm and other ultradian rhythms (8h, 6h) emerged and
became dominant (Fig 3.14). Since when this 1h rhythm disappeared, their total activity also
decreased, it is tempting to postulate that this led to a decrease in activity in the FX
experiments at 1 month. If this is the case, one would expect to see the same level of activity
on the days after FX compared to that on the day of FX experiment. However, based on the
pattern of decreased activity illustrated in Fig 5.8, this is less likely the case since the activity
dropped sharply on the day of FX compared to the normal day and recovered quickly right on
the next day after FX back to their previous higher level of activity and remain at that level
days after. In addition, given the fact that lambs were disturbed more by blood sample
collection on the day of control/FX experiment, their activity might have been expected to be

164
higher on those days compared to normal days rather than exhibiting a decrease in activity.
However, the reduction in the lamb activity did not affect weight gain, probably because
lambs at this age have already started to consume solid food and although they were less
successful suckling on the day of FX experiment, they did attempt to suckle harder. Taken
together, the effect of FX on the lambs’ activity at 1 month of age does not appear to be
simply due to weaning although it could be a predisposing factor. Instead, the mechanism for
the decreased locomotor activity in the lambs might be located at the neurotransmitter level.
However, since FX has an antidepressant effect, the finding that activity decreased with FX
administration was unexpected. Nevertheless, in clinical practice, it usually requires at least
two weeks of treatment before FX can exert its antidepressant effect (Stahl, 1998). Thus,
acute administration could elicit a paradoxical decrease in activity through an activation of 5-
HT autoreceptor which causes an initial silencing of serotonergic neuron activity (Stahl,
1998). In addition, in our study FX was administered to healthy lambs, which might result in
different effects on serotonergic tone compared to individuals with depressive symptoms.
At 6 months of age, a decrease in the lamb activity was a result of a shift in the time
lambs spent standing moving to standing quiet and a reduction in the intensity of standing
quiet activity. However, the FX exposed lambs were headbutting more on the day of FX
experiment. That suggests that these lambs were quieter but also more aggressive with acute
FX administration. In addition, we also observed jerky movements, which could represent for
one of the symptoms of serotonin syndrome (eg: tremor or akathisia) (Boyer & Shannon,
2005), in two of the eight lambs in this study. Blood samples for FX measurement were not
available for those lambs but it is possible that the effects of FX on the lambs’ behavior was
due to an acute increase in 5-HT available at the synapse on serotonergic neurons which

165
regulate the affective behaviors and motor tone (Boyer & Shannon, 2005). Interestingly, the
decrease in activity following acute FX administration in the lambs at 6 months of age was
associated with a bodyweight loss. As a vast majority of their usual standing moving at this
age is attributed to food intake activity at fixed and ad libitum meals, their weight loss
appeared to be consistent with DVR observations showing a decline in the time they spent
with standing moving after FX injection. However, whether the FX-induced decrease in
activity interfered with eating and led to a loss in their body weight or alternatively whether
FX caused a reduction in pre-meal appetite and food intake which in turn led to a decrease in
activity remains unclear. Evidence from human and rodent studies have shown that FX has
an inhibitory role on feeding behavior. (Clifton et al,. 1989; Lawton et al,.1995). Elevated
serotonin at the synapse following FX administration might interact with other neuropeptide
systems (neuropeptide Y, orexin, melanocortin) to alter feeding behaviors (Halford et
al,.2007).
Few studies have examined the HPA axis response following acute FX
administration. In rats, acute FX administration caused an increase in CRH, AVP, ACTH and
corticosterone (Fuller et al., 1976; Fuller et al., 1996; Gibbs & Vale, 1983). Similarly,
cortisol level increased following i.v infusion of citalopram (Bhagwagar et al., 2002) or after
a single dose of FX (von Bardeleben et al., 1989) in healthy humans. In sheep, Broadbear et
al., (2004) found an increase in ACTH and cortisol for 40 mins following acute subcutaneous
sertraline injection in gonadectomized ewes. However, we did not observe any significant
changes in ACTH and cortisol in lambs at 3 months and 1 year of age following acute FX
administration. The differences in our result and the Broadbear et al study could be due to the
route of administration and the sampling period. FX was given intravenously and the earliest

166
sample of ACTH and cortisol after dosing was at 30 mins in our study while sertraline was
given subcutaneously and blood was sampled every 10 mins in the other study. In addition,
sex hormones could be a contributing factor in different results since in the subsequent study,
Broadbear et al.,(2004) did show that the increase in ACTH and cortisol in gonadectomized
sheep was abolished by sex hormone treatment. Moreover, it is believed that the weak
response of the HPA axis to acute SSRI administration is due to the activation of the negative
feedback on the 5-HT1A autoreceptors (Raap & Van de Kar, 1999). The midbrain
serotonergic system and HPA axis are highly interrelated; this is mediated by 5-HT1A
receptors. (Porter et al., 2004) An overactive HPA axis which is not suppressed by
dexamethasone administration is a potential long-term suicide predictor (Pompili et al.,
2010). However, whether the finding that no changes in basal cortisol and ACTH level after
acute FX administration in our lambs can indirectly imply that FX does not have effects on
suicidal behavior in the adolescent humans is uncertain.
In conclusion, acute FX effects on cardiovascular-respiratory function and behavior
were seen in some, but not all age groups. The effect tended to occur at milestones events of
development such as weaning, puberty and adulthood. Gender difference in FX effect on the
cardiovascular-respiratory function was observed in the older lambs with more profound FX
effects in males.

167
6. Postnatal Outcomes in Lambs Exposed Antenatally to FX
6.1 Introduction:
Up to 20% of pregnant women might experience mood disturbances at any stage of
their pregnancy and thus might require antidepressant therapy (Burt & Stein, 2002). SSRIs
have been widely used as a pharmacological treatment for depression during pregnancy with
moderate to severe symptoms thanks to their relatively benign side effects profile compared
to other earlier antidepressants (MAOIs, TCA). Indeed, over the last decade, SSRI use during
pregnancy has increased from 1.5% in 1996 to 6.4% in 2004 (Andrade et al., 2008). A
similar increase to approximately 7% was also reported by Wichman et al., (2008). As the
name suggests, SSRIs exert their actions by inhibiting serotonin recycling through the
serotonin transporter, thereby increasing the amount of serotonin available in the synaptic
cleft, and altering serotonergic transmission (Oberlander et al., 2009). It has been well-
established that in adults serotonin is a neurotransmitter which plays a key role in modulating
emotion, cognition, attention, arousal and stress responses. In the developing brain, central
serotonin also acts as a growth factor regulating serotonergic neuronal growth and the
development of other related neural systems (Whitaker-Azmitia, 2001). Since SSRIs are
capable of crossing the placenta (Hendrick et al., 2003; Kim et al., 2004; Kim et al., 2006;
Rampono et al., 2009), there is a significant risk for the fetus from exposure to SSRIs during
such critical phases of their neurodevelopment. Studies in human and sheep have
demonstrated SSRIs effects on the fetuses. Mulder et al reported a disrupted quiet sleep
reflected by the abnormal phenomenon of continual bodily activity during non-REM sleep
periods in human fetuses in utero exposed to SSRIs (Mulder et al., 2011). Similarly,

168
Salisbury et al (2009) demonstrated increased jerky movements and decreased fetal breathing
movements in SSRIs-exposed fetuses at 26 and 36 weeks gestational age as compared to the
non exposed group. This is in agreement with previous findings from our group which
showed an alteration of fetal behavioral state and neuroendocrine function in fetal lambs with
chronic FX infusion to the mothers during late gestation (Morrison et al., 2001; Morrison,
2001; Morrison et al., 2004). Moreover, a recent study also reported an alteration of fetal
middle cerebral artery flow characteristics and blunted fetal heart rate variability across the
day in 36 week human fetuses exposed to SSRIs as compared to the control group (Rurak et
al., 2011). These findings raise an important question whether these fetal neurobehavioral
effects persist beyond the perinatal period into childhood, forming a concept of behavioral
teratology in the neonate resulting from fetal exposure to SSRIs. A blunted pain response in
terms of facial characteristics and cardiac responses in prenatally SRIs-exposed neonates has
been reported by Oberlander et al., (2002). This alteration in pain reactivity was still apparent
at 2 months of age in the fetal SSRIs-exposed infants, even after controlling for drug level
and maternal mood (Oberlander et al., 2005). In addition, a lower basal cortisol level in the
evening was also observed at 3 month of age in prenatal SSRIs-exposed infants (Oberlander
et al., 2008)
Yet, it is still inconclusive regarding the long-term effects of prenatal SSRI exposure
on neurobehavioral development in early childhood. Some studies showed a delayed in
psychomotor and behavioral development in prenatally SSRIs-exposed children (Casper et
al., 2003; Pedersen et al., 2010) while others failed to observe these differences (Heikkinen et
al.,2002; Heikkinen et al.,2003; Nulman et al., 1997; Nulman et al., 2002). Nevertheless, a
challenge for this type of research in human is that it is impossible to distinguish between the

169
effects of SSRIs and the effects of maternal psychiatric illness on the outcomes. Moreover,
the postnatal social environment of a depressed mother could also have negative influences
on child development after SSRI exposure (Oberlander et al., 2008). These issues might be
resolved with animal studies. Since lambs are precocious at birth and have a very similar
physiological characteristics to the human, it appears to be a good model to control for the
confounding factors present in human studies. In this study, we chose to investigate the
postnatal outcomes including birth weight, postnatal weight gain, behavioral development
(neonatal behavior, sleep-rest cycles/activities, feeding activity) and cardiovascular changes
(heart rate, HRV, sPO2) in lambs exposed to FX during late gestation.
6.2 Methods:
6.2.1 Animal And Catheter Implantation:
Seven time-bred pregnant ewes (Dorset/Suffolk Cross) were bred following the same
protocol in our laboratory described previously in Chapter 2. Ewes were moved to Centre of
Comparative Medicine (CCM), UBC 2-3 weeks before their due date (term = 147 days) to
allow for acclimation.
They were housed in groups of two or three in the holding pens of approximately
10x12 feet and were able to go outdoors through a small door in their pen until the end of
their pregnancy. Approaching their due date, ewes were kept indoors under natural light
conditions. Ewes were fed following the protocol in the CCM: 300 g grain + 450 g alfalfa
cubes at 7:00 to 8:00 am, 2 flakes of hay at 8:00 to 9:00 am and 2 flakes of hay at 2:00 pm.
The study involved 2 groups of lambs: one group of 7 lambs who were exposed to FX
antenatally and another control group of 2 lambs born in CCM and 10 control lambs

170
previously born in the CFRI without any perinatal FX exposure for the first 11 days after
birth.
A heparin-bonded 5.0 French polyurethane catheter (Instech Solomon, Plymouth
Meeting, PA) was percutaneously implanted to the ewe’s jugular vein using a 13G sterile
stainless steel needle (Instech Solomon, Plymouth Meeting, PA) under local analgesia with
Lidocaine HCl 2% (Bimeda-MTC, Animal Health Inc, Cambridge, Ontario). The catheter
was implanted at an average gestational age of 131 ± 1 d in 5 pregnant ewes. The catheter
was inserted approximately 20cm and proper functioning of the catheter was ensured before
it was secured to the skin by a 2-0 Nylon suture. The external part of the catheter was closed
with a sterile metal pin at the end and kept in a small ziplock bag and safety pinned under a
bandage around the animal’s neck. The catheters were flushed every day with heparinized
saline and served for subsequent daily FX injection and blood samplings at delivery.
6.2.2 Experimental Protocol:
Beginning one day after the catheter insertion, the pregnant ewes were given daily 50
mg of FX (5 ml of 10 mg/ml) via the inserted catheter. FX was injected every day in the
morning around 10:00-11:00 am for 14 days or until the ewes gave birth, whichever came
earlier. Signs of imminent parturition in the ewe (bilateral concave flanks, restlessness,
pawing at the pen floor, moving in circles and looking toward the back) and the lambs’ birth
were observed by DVR from infrared cameras which will be described more detail in the
section DVR and behavioral evaluation. Every attempt was made to measure the lambs’
weight, head circumference, tissue spO2 and to obtain a jugular vein blood sample (via
percutaneous puncture) as soon as they were born. They were dried with a towel, sexed,

171
weighed and had on actiwatch around their neck right after birth (more details about
actiwatch in the section Actiwatch/Rest activity monitoring). Lambs were monitored for 14
days and daily weight, tissue spO2, head circumference and abdominal circumference were
measured. They were housed in a smaller pen with their mothers (8x8 feet) to facilitate ewe-
lamb bonding until the end of the monitoring period. On postnatal day 2, 5, 10 and 14, ECG
measurements and blood samplings for FX, NFX concentration were performed (more details
in the sections below).
6.2.3 Digital Video Recording And Behavioral Evaluation:
Two infrared video cameras were installed at two corners of each pen in three pens in
the sheep holding room and connected to a digital video recording system on a computer
which was located on the hallway just outside the sheep pen. Another mobile infrared camera
was used to directly observe and record the lambs’ behavior while they were on the sling for
ECG measurement. The infrared function allowed the recording and monitoring of the ewe
and lamb’s behavior at night without interrupting their circadian rhythm. These cameras
could also be accessed remotely from home and the video records could be analyzed off-line.
Lamb’s activities were monitored continuously at least 14 days to determine any
abnormal behavior changes in the lambs. The number of daily sleep and suckling bouts were
identified from the DVR observations for each lamb from postnatal day 2 until day 10. The
total activity bout was defined as a sum of suckling bout and active bout without suckling
during 24h of daily observation.

172
6.2.4 Actiwatch Rest/Activity Monitoring:
A similar method and analysis were used as described in chapter 2. The lambs’
activities were recorded for at least 14 days after birth.
6.2.5 Powerlab Data Acquisition System:
ECG measurement was conducted on the lambs for 1 hour on postnatal day 2, 5, 10
and 14. Lambs were taken to the procedure room for ~ 20 minutes to glue on surface ECG
electrodes. Three ECG snap electrodes were attached to the skin on the back and bilaterally
on the chest and secured in place by adhesive bandages. Lambs were put in the cloth sling,
which located right in front of the holding pen from where the lamb came in full view of the
mother, and the ECG electrodes were connected to a polygraph recorder (model TA-4000,
Gould Instrument Systems Inc, Valley View, OH) and to a Powerlab data acquisition system
(AD instrument, Mountain View, CA, USA) by three snap leads to record their
electrocardiograph. HRV data were analyzed using the same method as described in chapter
2.
6.2.6 Pulse Oximeter: Tissue SpO2 And Heart Rate:
Daily tissue spO2 and heart rate were measured using a pulse oximeter (Nonin
Medical Inc, Plymouth, MN, USA), with the oximeter sensor clipped on the ear of the lamb.
The lamb’s ear was shaved for a better signal. The value is an average of 5 minutes
monitoring.

173
6.2.7 FX And NFX Concentration:
Blood samples were also collected after birth from the ewes and lambs and on
postnatal day 2, 5, 10 and 14 from the lambs for fluoxetine measurements. Samples were
taken from the ewes via the inserted jugular vein catheters. Lambs were taken to the
procedure room and the wool along the jugular vein was shaved. The skin was disinfected
with alcohol 70% and Xylocain 2% gel was applied. The jugular vein was accessed by
percutaneous puncture. Approximately 4 ml of blood was collected in Lithium Heparin
(68USP units) vacutainer tubes (BD Vacutainer, NJ, USA) and centrifuged for 20 min at the
speed of 4000rpm in a Centrifuge 5810R (Eppendorf, ON, Canada). Plasma was then
transferred to glass tubes for fluoxetine samples and stored at 0oC until analysis. Lambs
usually slept or lay quiet while blood was collected and were returned to their mother
immediately.
FX and NFX concentrations were measured using a validated liquid chromatography
with tandem mass spectrometry method (LC/MS/MS) (Chow et al., 2011) as described in
Chapter 2. The sample volume used for each enantiomer (R)-, (S)- fluoxetine and (R)-, (S)-
norfluoxetine was 250 µl. The limit of quantitation was 1 ng/ml.
6.2.8 Statistical Analysis:
The areas under the curve (AUC) were calculated for daily variables (weight, sPO2,
heart rate) by applying the formula described by (Altman, 1991). Then the AUC of each
variable were compared between the control and FX group using the unpaired t-test. The
effect of FX exposure and sex on gestational age, birth weight and the time for the lambs to
first stand, walk and suckle were determined using 2 way ANOVA. Two-way ANOVA with

174
repeated measures was used to determine the effect of FX exposure and sex on activities
(actiwatch and DVR observations) for the first 10 days of life and on HRV variables on
postnatal day 2,5,10 and 14 in the lambs with multiple comparison correction (Bonferoni t-
test). All data were analyzed using the Sigma Plot 11 statistics software (Systat Software,
Inc; San Jose, CA, USA).
6.3 Results:
In the FX group, ewes were given daily 50mg FX i.v injection for approximately 2
weeks before their due date. The average duration of antenatal FX exposure in the lambs was
12 ± 1.3 days.
Table 6.1 describes the gestational age (GA) and birth weight of the lambs included
in the study in this chapter. There were no significant differences in GA at delivery between
the control and FX-exposed group. However, it was noticed that 2 ewes that delivered
singleton lambs in the FX group had a shorter gestational length (141 and 142 days) as
compared to the normal GA of 147 days at term. In the control group, two lambs from the
triplet also had a shorter gestational length (142 days). There was also no significant
difference in birth weight between control and FX exposed lambs.
Figure 6.1 illustrates daily postnatal weight gain for the first two weeks of life in the
control and FX groups. No significant differences in postnatal weight gain were found
between the two groups.

175
Control lamb
GA at birth
(days)
Birth weight (gram)
Lamb number
FX-exposed Lamb
GA at birth
(days)
Birth weight (gram)
Lamb number
L620F 146 5830 Single L756M1 147 4700 Twin L371M 148 6250 Single L752M1 148 4400 Twin L310M1 142 3300 Triplet L752M2 148 5000 Twin L310M3 142 3500 Triplet L577M 141 4100 Single L825M1 145 4900 Twin L630F 142 5000 Single L825M2 145 5300 Twin L756F2 147 3900 Twin L752F 144 5200 Single L643F 148 7000 Single Mean 145 4897 146 4871 SEM 1 421 1.1 388
Table 6.1: Gestational age and birth weight in the control and antenatally FX-exposed lambs M = male, F = female, 1,2,3 after M,F indicates the order of delivery in twin and triplet lambs
FIG. 6.1: (A) Daily weight in the control (n=7) and prenatal FX-exposed lambs (n=7) from postnatal day 1 (day of birth) to postnatal day 14.
Plasma FX, norfluoxetine (NFX) enantiomer concentrations in the ewes (n = 5) at
delivery and in the lambs (n = 7) at birth, on postnatal day 2, 5, 10 and 14 are presented in
Figure 6.2. At delivery, the mean maternal FX and NFX level were 627.9 ± 249 and 66.6 ±
14.4 ng/ml, respectively. Mean plasma FX level was quite low in the lambs at birth, with an
average of 17.3 ± 10.9 ng/ml and decreased progressively thereafter. It became undetectable

176
after postnatal day 5. NFX was undetectable in the lambs from birth to postnatal day 14,
which suggests that lambs of this age are unable to metabolize the drug. The relationship
between the lamb: ewe ratio for FX concentration and the duration from the last maternal FX
dose is shown in figure 6.3. The ratio was highest in the lambs that had the shortest duration
from the last FX dose, within 8 hours and it fell rapidly thereafter.
Figure 6.4 illustrates the daily tissue sPO2 and heart rate values obtained from pulse
oximeter from postnatal day 1 to 14 in two control lambs and seven FX-exposed lambs. The
results were similar between the two groups. sPO2 over 14 postnatal days fluctuated within
the normal range 90-100% while heart rate decreased progressively in both control and FX
lambs from an average of 234 ± 17 bpm and 230 ± 19 bpm on postnatal day 2 to 187± 6 bpm
and 200 ± 8 bpm on postnatal day 14, respectively.

177
FIG.6.2: FX, (R)-FX, (S)-FX and NFX, (R-)NFX, (S)-NFX concentrations in the prenatally FX-exposed ewes (n= 5) and lambs (n=7). Samples were taken at birth in both ewes and lambs and on postnatal day 2,5,10 and 14 only in lambs. PND = Postnatal day [NFX] levels in the lambs were lower than the limit of quantitation (1ng/ml)

178
FIG 6.3: The relationship between the lamb:ewe plasma FX concentration ratio and the time from the last FX dose (hours) in the ewe.
FIG. 6.4: Daily (A) tissue spO2 and (B) heart rate measured by pulse oximeter in the control (n=2) and prenatal FX-exposed lambs (n =7) from postnatal day 1 (day of birth) to postnatal day 14.

179
The lambs with prenatal FX exposure were more active than the control lambs, which
was reflected by faster first time activities (stand-walk-suck) and an increase in subsequent
activities obtained from actiwatch and DRV observations. (Figs. 6.5-6.7) It took the FX
lambs (n = 7) only half the amount of time as compared to the control lambs (n = 37) to
achieve their first time activities. While the control lambs started to stand at 32 ± 2 mins
after birth, the FX-exposed lambs did this at 17 ± 3 mins, p = 0.017. In general, the control
lambs attempted to suck within the first one hour of life (56 ± 5 mins) whereas the FX-
exposed lambs did this within the first half hour (29 ± 6 mins), p = 0.006. There was no sex
difference in the first time activities.
Figure 6.6 shows the activity variables from actiwatch monitoring from postnatal day
2 (the first full day after birth) to postnatal day 14 in seven control and seven FX-exposed
lambs. The lambs’ activity increased over days in both control and FX groups, but it was
increasingly greater in the FX group than the control group. Total activity score and active
bout per days were significantly higher in the FX-exposed group from postnatal day 4 to 14
as compared to that on each corresponding day in the control group, while they were
comparable between the 2 groups on postnatal day 2 to 3. Mean score in active periods were
also higher in the FX group vs control group, but not significantly different. In contrast,
episode duration was significantly lower in the FX group compare to the control on postnatal
day 5, 9, 10 and 12. The percentage of time moving was significantly higher in the FX group
compare to control on postnatal day 12. A reverse situation occurred with the percentage of
time immobile.
From DVR observations, sleep, suckling and non-suckling activity bouts were
determined from postnatal day 2 to day 10 in the same seven control and FX-exposed lambs

180
and are shown in figure 6.7. The number of total activity bouts is the sum of suckling bouts
and non-suckling activity bouts. Prenatal FX-exposed lambs appeared to have a significantly
lower number of suckling bouts than the control lambs on postnatal day 2, 5 and 6. However,
the AUC of non-suckling activity bouts for 10 postnatal days was significantly higher in the
FX-exposed lambs compared to the control. Therefore, the number of total activity bouts (=
suckling bouts + non-suckling active bouts) was comparable between the two groups until
postnatal day 9 and so was the number of sleep bouts. On postnatal day 10, the number of
total activity bouts was significantly lower in the FX group than the control, and this
difference is due to the significantly lower number of sleep bouts in the FX group than the
control group. There were no significant differences in the number of suckling bouts between
the two groups on postnatal day 10. Collectively, the results suggested that lambs exposed to
FX prenatally were more active than the control group, but the increase in their activity did
not contribute to their feeding behavior and thus, their postnatal weight gain. In addition, we
did not observe any other abnormal behaviors in the FX-exposed lambs for the first 14 days
of life.
Interestingly, the high level of activity appeared to exist in the FX-exposed lambs
even at a low level of FX and undetectable NFX level and this high level of activity still
persisted on postnatal day 10, 14 when the level of FX was no longer detectable. (Figs. 6.2 &
6.6)
HRV was only significantly different on postnatal day 2 and was comparable on
postnatal days 5, 10 and 14 between FX and control groups. The time domain variables of
HRV (SDNN, SD Delta NN, RMSSD) was significantly lower in the FX-exposed lambs than
the control lambs at both 30 and 60 mins of the recording period. Similarly, the total power

181
was significantly lower in the FX group but no significant differences were found with LF
power, HF power and LF/HF ratio. (Fig. 6.8) Total activity score from actiwatch during the
1h period of ECG monitoring also showed a lower activity level in the FX group compared to
the control group, however the difference in total activity score between the two groups was
not statistically significant. (Fig. 6.8)
FIG. 6.5: Neonatal behavior (first time to stand, walk, suck immediately after birth) in the control lambs (n = 37) and prenatal FX-exposed lambs (n =7). Asterisk (*) indicates significant differences between the control (open bars) and FX group (black bars), p < 0.05.

182

183
FIG. 6.6: Daily activity variables (active bouts per days, total activity score, mean score in active, episode duration, % time mobile, % time immobile) obtained from the actiwatches in the control (opened, diamond symbols) and prenatal FX-exposed lambs (closed, circle symbols) from postnatal day 2 (the first full day) to postnatal day 14. * indicates significant differences in the corresponding values between the control and FX group, + indicates significant differences from the day 2 within the control group, ^ indicates significant differences from the day 2 within the FX group. Stats: 2 way ANOVA repeated measures with multiple comparison correction Bonferoni t-test

184
FIG. 6.7: Daily sleep, suckling and non-suckling activity bouts obtained from DVR observations in the control (open, diamond symbols) and prenatal FX-exposed lambs (closed, circle symbols) from postnatal day 2 (the first full day) to postnatal day 10. * indicates significant differences in the corresponding values between the control and FX group, + indicates significant differences from the day 2 within the control group, ^ indicates significant differences from the day 2 within the FX group. AUC non-suckling activity bout were significantly higher in the FX group (p = 0.04)

185
FIG. 6.8: HRV (time domain: SDNN, SD Delta NN, RMSSD; frequency domain: total power, LF power, HF power, LF/HF) on postnatal day 2 in control group (diamond symbol, dotted line, n = 8) and prenatal FX-exposed lambs (circle symbol, solid line, n = 7). * indicates significant differences between the corresponding values in the control and FX groups. + indicates significant differences between the values at 30 and 60 min within the control group, ^ indicates significant differences between the values at 30 and 60 mins within the FX group, p < 0.05. Stats: 2 way ANOVA repeated measures with multiple comparison correction Bonferoni t-test

186
Day 2 Day 5 Day 10 Day 14
Heart Rate
Control Mean 220 4.0 ‐17.8 ‐29.7
SEM 4.3 12.5 4.8 2.4
FX Mean 225 8.3 ‐22.4 ‐26.3
SEM 6.0 0.2 0.6 ‐0.1
SDNN
Control Mean 22.5 ‐2.6 5.9 7.1
SEM 2.3 ‐0.2 0.9 2.2
FX Mean 15.5*
7.2 13.1 10.3
SEM 0.8 2.9 0.9 1.5
SD Delta NN
Control Mean 18.8 ‐7.5 ‐2.8 ‐4.6
SEM 4.8 ‐2.4 ‐1.2 0.2
FX Mean 7.2*
12.0 6.0 2.0
SEM 1.4 5.5 1.9 1.3
RMSSD
Control Mean 18.8 ‐7.5 ‐2.8 ‐4.6
SEM 4.8 ‐2.4 ‐1.2 0.2
FX Mean 7.2* 12.0 6.0 2.0
SEM 1.4 5.5 1.9 1.3
LF Power (ms²)
Control Mean 73.8 24.6 96.6 197.2
SEM 11.5 14.0 15.3 24.0
FX Mean 37.5 52.2 114.3 170.6
SEM 4.7 11.7 18.6 24.4
HF Power (ms²)
Control Mean 51.4 ‐25.0 3.0 33.1
SEM 13.4 ‐8.8 ‐1.7 11.1
FX Mean 19.2 35.0 35.2 25.0
SEM 4.8 16.9 8.6 2.6
LF/HF
Control Mean 2.2 1.5 2.0 2.1
SEM 0.6 0.1 0.3 0.4
FX Mean 2.6 ‐0.2 0.7 2.9
SEM 0.5 ‐0.1 0.1 0.5
Control N = 8 5 12 5
FX N = 7 7 7 7 Table 6.2: Mean ± SEM of HRV values for the first 30 mins of the experiment in the lambs on day 2 and the changes from day 5, 10, 14 compared with the values on day 2 (*) indicates significant differences between FX and control group

187
6.4 Discussion:
In this study we found that the lambs with prenatal FX exposure were more active
than the control lambs. This was indicated by a faster first time activities (stand-walk-suck)
which represent important neonatal behaviors in sheep and by an increase in subsequent
activities obtained from actiwatch and DRV observations. HRV variables on postnatal day 2
were lower in the FX-exposed lambs than the control lambs. There were no significant
differences in gestational age and birth weight between the two groups. Daily postnatal
weight gain, tissue sPO2 and heart rate obtained from pulse oximeter were also comparable
between the two groups.
Maternal FX level at delivery in our study was higher than that reported in the human
(Kim et al., 2006) and in a previous sheep study in our lab (Morrison, 2001; Morrison et al.,
2002). As was discussed in previous papers (Alfaro et al, 2000; Kim et al., 2004), since
CYP2D6 is a key enzyme involved in FX metabolism, the maternal FX concentration might
be a result of FX-mediated CYP2D6 inhibition. From the literature, the level of FX in the
neonates varies between studies and depends on the postnatal day on which FX was
measured (Kim et al., 2006; Kwon & Lefkowitz, 2008; Rampono et al., 2004; Spencer,
1993). One case report gave a FX concentration at delivery of 26 ng/ml and undetectable
levels at 96h thereafter in an infant, which is similar to our current study (Spencer, 1993).
The FX level in our lambs at birth (mean = 17.3 ± 10.9ng/ml) was lower than that measured
in umbilical cord vein at delivery (mean = 41.3, 95%CI = 16.9, 59.8) from 9 pregnant women
on FX therapy in another study (Kim et al., 2006). Interestingly, NFX was also not detectable
at birth and the days after, which suggests that fetal and newborn lambs are unable to
metabolize the drug. Our result is in agreement with previous study in our lab in which NFX

188
was also undetectable in the lamb fetuses after fetal FX administration, in contrast with the
mother (Kim et al., 2004). Since the lamb was not able to metabolize the drug, one would
expect to see a higher level of FX in the lambs than it was. However, we also found that there
was a relationship between the ratio of lamb:ewe FX concentration at delivery and the
duration from maternal the last FX dose (Fig. 6.3), indicating a link between maternal and
fetal FX level. This suggests that following the period of maternal FX administration, the
elimination of FX from the fetus was largely the result of transfer of the drug back to the
mother and maternal elimination.
In the human newborn, motor activity is one of the indicies of biobehavioral
assessment. In sheep, neonatal behavior is one of the many factors contributing to lamb
survival. Lambs that are slow in standing and suckling after birth have a lower survival
chance. (Dwyer et al., 2005) Moreover, first time activities such as standing, walking and
suckling are indicators of neonatal adaptation to postnatal life (Dwyer, 2008b). In the present
study, we found that FX-exposed lambs were faster to stand, walk and suckle as compared to
the control lambs. This suggests that FX-exposed newborn lambs do not appear to have any
difficulty in neonatal adaptation. Our result was different from a human study reported by
(Pedersen et al., 2010) in which the first time to sit and walk was delayed in infants exposed
to antidepressants during second and third trimester of gestation. However, in their study
different classes of antidepressants (SSRIs, TCAs) were included.
For the first 2 weeks of life, there was a progressive increase in the level of lambs’
activity in both the FX-exposed and control groups, which is similar to our developmental
study presented in chapter 3 and the results reported by Rurak et al., (2008). However, FX-
exposed lambs were more active than the control lambs, reflected in significantly higher

189
values for both the number of active bouts per day and total activity score between FX and
control groups from postnatal day 4 to 14. Although the FX-exposed lambs were more active,
it appeared that their movement bouts were shorter than the control lambs, which was
reflected by significantly lower values for episode duration in the FX group on postnatal day
5,9,10 and 12. Similar to our findings, there is one study in human infants that also found
prenatally SSRIs exposed infants increase their motor activity during sleep compared to the
control (Zeskind & Stephens, 2004), however the difference in activities between the groups
in their study was attributed to the shorter gestational age in the SSRIs-exposed infants. In
our present study, mean gestational age at delivery was similar between the two groups,
suggesting that the increase in the level of activity in the FX-exposed lambs is independent of
gestational age. Moreover, in our study changes in activity in FX-exposed lambs were not
confounded by additive effects of maternal depressive status or stress during gestation as in
human study. Instead, in sheep ewe and lamb bonding after birth is the key factor affecting
behavior (Dwyer, 2008b) and it appeared that all the ewe-lamb bonding in our sheep was
successfully established soon after delivery. As it was shown in chapter 3, the more vigorous
activity of the lambs for the first month of life was due to their feeding activity. However,
observation from DVR revealed that the increase in the level of activity in FX-exposed lambs
was not due to the increase in suckling activity, which is the major activity in lambs in the
first month of life. Instead, the increase in the level of activity in FX-exposed lambs was due
to the higher in the number of non-suckling activity bouts in the FX group reflected by
significant differences in AUC of non-suckling activity bouts between FX and control
groups. This is consistent with our finding that there was no significant difference in
postnatal weight gain between the two groups. From DVR observation, on postnatal day 10,

190
FX-exposed lambs had significantly lower number of sleep bouts as compared to the control,
suggesting they sleep for longer duration than the control. However, on this postnatal day 10,
FX-exposed lambs had a shorter episode duration from actiwatch measurement. This
suggested that even though the FX-exposed lambs slept more, their sleep was more disrupted
than the control. However we did not observe abnormal behavior in the prenatal FX-exposed
lambs during the first 10 postnatal days.
As was mentioned previously, the mechanisms of poor neonatal adaptation in
newborns exposed to SSRIs during gestation are still a matter of debate. From our study it is
apparent that the high level of activity in our FX-exposed lambs was not due to FX toxicity
since the average of FX level in the lambs at delivery was lower than the minimum range of
adult human therapeutic level, which is 40-250 ng/ml (Krogh CME, 1995) and NFX, a
pharmacologically active metabolite, was also undetectable. The question is whether this is a
result of a withdrawal effect. In adults with opioid-withdrawal syndrome, the symptoms can
last for weeks after abrupt discontinuation of the drug involved. Methadone appears to have a
longest duration withdrawal symptoms among opioids after acute discontinuation with the
severity of symptoms and can last up to 16 days after the last dose. (Kosten & O'Connor,
2003) The argument is that in my study, the increased activity in the lambs in the FX groups
was observed up to 2 weeks after birth, which can be similar the duration of opioid-
withdrawal syndrome in the human. Longer follow-up of the increased activity in the
antenatally FX-exposed lambs will help to rule out the mechanism. That is, if the increased
activity in the lambs still persisted after 2 weeks, it is less likely due to the withdrawal
syndrome.

191
However, I believe that the increased in activity in the FX-exposed lambs is less
likely due to withdrawal syndrome. Rather, it is due to a change in the fetal brain
development since evidence from human and sheep studies have shown that the alteration in
activity resulting from antenatal SSRI exposure seems to appear in the fetuses before the
drug exposure ended and continue until the postnatal period. Mulder et al., (2011) reported
an abnormal phenomenon of continual bodily activity during non-REM sleep in SSRIs-
exposed fetuses. Similarly, another study found SSRIs-exposed fetuses at 26 weeks of
gestation had more jerky movements than the control and MDD/no SSRIs exposure groups
(Salisbury et al., 2006). In sheep, there were also increased eye movement and fetal breathing
movement with 8 day FX i.v infusion. (Morrison et al, 2001) Collectively, these findings
from our study and others support the hypothesis that a possible underlying mechanism for
the high level of activity in the FX-exposed lambs is SSRIs-induced changes in fetal brain
development. Further evidence for this mechanism comes from studies showing a lower cord
blood level of calcium-binding protein S100B, a biomarker of early brain development and
central serotonergic function, in SSRI-exposed neonates (Pawluski et al., 2009). It is known
that the astroglial 5-HT 1A receptor is activated by an increase of synaptic 5-HT after SSRI
administration, and this will stimulate a release of S100B protein, which in turn mediates the
growth and survival of neurons (Whitaker-Azmitia et al., 1990; Whitaker-Azmitia, 2001).
In our study we also found that the time domain of HRV (SDNN, SD Delta NN,
RMSSD) and total power were significantly lower in the prenatally FX-exposed lambs on
postnatal day 2 compared to the control lambs. It is known that changes in HRV are usually
associated with physical activity (Nakamura et al, 1993; Voss et al, 2002; Warren et al,
1997). On postnatal day 2, the 24 h total activity score were comparable between control and

192
FX groups. Further analysis of the total activity score for during the 1h of HRV measurement
on postnatal day 2 did not show significant differences in activity score between the two
groups even though the activity score during this 1h of HRV measurement was lower in the
FX-exposed group (Fig. 6.8). Thus, it is uncertain whether activity plays a role in the lower
level of HRV in the FX-exposed group. There are limited data on HRV changes in the
newborns after prenatal SSRIs exposure in the literature, although two studies did report a
similar trend of lower HRV rhythms in the newborns or a blunted facial and HRV response
to pain in 2 month-old infants after prenatal SSRI exposure (Oberlander et al., 2005; Zeskind
& Stephens, 2004). Since changes in HRV in our study only occurred in the time domain but
not in frequency domain indices, it is uncertain if the changes come from sympathetic or
parasympathetic systems. Changes in HRV also occurred in SSRI-exposed human fetuses. A
recent study has reported that short and long-term variation of fetal heart rate, accelerations
and duration of high variability episodes did not change during the day and were lower in the
SSRIs-exposed fetuses compared to a significant increase during the day in the control group.
(Rurak et al., 2011) Taken together, our results and the other findings showed a similar low
HRV or blunted HRV response in the SSRIs-exposed infants which originate from the fetal
period with SSRI exposure.
The major effects of prenatal FX exposure on the postnatal lambs in the present study
consisting of an increase in activity at birth and from postnatal day 4 to 14 and low HRV on
postnatal day 2. There was a lack of FX effects on gestational length, birth weight, postnatal
weight gain. Heart rate and tissue SpO2 were also not different between the control and FX
groups. Our results are different from other previous human studies which reported an
increase in preterm delivery, shorter gestational length, lower birth weight and postnatal

193
weight gain in third trimester FX exposure fetuses. (Chambers et al., 1996; Goldstein, 1995;
Hendrick et al., 2003; Lewis et al., 2010; Oberlander et al., 2006; Suri et al., 2007). However,
our results were consistent with those found in previous sheep study in our lab (Morrison,
2001; Morrison et al., 2002) except for postnatal weight gain which was not obtained in
Morrison el al study. As was discussed before, different results between human studies and
ours are probably due to the presence of confounding factors in human studies but not in
animal studies, for example the maternal depressive illness and other associated risk factor
such as smoking and alcohol abuse. Even though maternal illness severity was accounted for
in the Oberlander et al and Suri et al studies, it is possible that the cumulative effect of FX
exposure over most or all of pregnancy in the human studies, compared to the short-term late
gestation sheep studies, resulted in the observed lower birth weight and gestation length.
In summary, from this study we found that the newborn lambs were not able to
metabolize FX, thus FX metabolise and clearance in the fetal lambs were likely achieved via
the mother. FX-exposed lambs were more active than the control lambs at birth and from
postnatal day 4 to 14 and had a lower HRV on postnatal day 2. Together with the FX
measurement after birth and on postnatal day 2, 5, 10, 14, the increase in activity in the
postnatal lambs exposed to the drug in utero suggests the most likely mechanism for these
effects is alteration in fetal brain development, due to the importance of serotonin in brain
maturation. However, DVR observations showed the increase in activity score was not due to
suckling activity but due to a decrease and disrupted sleep activity. Consistently, there was
no difference in postnatal weight gain in the two groups. And there was also a lack of FX
effect in gestational age, birth weight, heart rate and tissue SpO2.

194
7. Summary and Conclusions
Depression has been recognized as one of the most burdensome disorders during
pregnancy, affecting at least 10-15% of pregnant women (Bhatia & Bhatia, 1999; Nonacs &
Cohen, 2003). Pharmacological treatment is a norm. And one third of pregnant women
suffering from depression take SSRIs as antidepressants (Oberlander et al., 2006). There is an
increasing use of FX and other SSRIs during pregnancy (Andrade et al., 2008). A
considerable amount of attention has been paid in the literature to the assessment of birth
outcomes following in utero FX exposure during late gestation. However, studies that focus
on the mechanisms underlying the observed outcomes are relatively sparse. Several
mechanisms have been proposed but there is a lack of supporting evidence. Previous
experiments in our lab were conducted with 8 day maternal FX i.v infusion to expose late
gestation sheep fetuses to FX and primarily focused on the fetal responses and the underlying
mechanisms. (Morrison et al., 2001; Morrison, 2001; Morrison et al., 2002; Morrison et al.,
2004; Morrison et al., 2005). The current study is in part a continuation of the previous
studies, focusing on the effects of maternal FX treatment on postnatal lambs and the
underlying mechanisms of poor neonatal adaptation. As noted elsewhere in this thesis, this
syndrome occurs in 30% of newborns prenatally exposed to SSRIs (Koren et al., 2009). In an
attempt to elucidate the underlying mechanisms for poor neonatal adaptation, I set out to
determine if SSRI toxicity is the responsible mechanism, FX was given to the newborn lambs
acutely on approximately postnatal day 4 in an attempt to reach the plasma concentration
levels observed in human infants at the same age. This was successful in that the level of FX
in the newborn lambs fell within the range of plasma FX concentration measured in human
infants sampled on postnatal day 2 following in utero exposure to the drug. However, our

195
protocol of acute, single dose FX administration differs from the clinical setting since human
infants are exposed to FX throughout all or most of pregnancy. Similarly, when FX is
prescribed to treat depression in children and adolescents, it is given chronically with
multiple, daily doses. However, in my subsequent study to investigate the effects of maternal
FX treatment on the postnatal lambs, FX was given to the pregnant ewes for the last ~2
weeks of pregnancy, but again for a relatively short period of gestation. The lambs were
exposed in utero to FX at clinically relevant concentrations, followed by evaluation of birth
weight, postnatal weight gain, behavioral development and cardiovascular function. In the
previous studies in our lab, FX was administered to the ewes using infusion pump and thus
required surgical instrumentation of the ewes and fetuses, and the restraint of the ewes in a
monitoring pen during the infusion and monitoring period (Morrison et al., 2001; Morrison et
al., 2002; Morrison et al., 2004; Morrison et al., 2005). Thanks to the procedure to implant
jugular vein catheter subcutaneously in the pregnant ewes, we were able to successfully
administer FX allowing the ewes to moving freely in their pen. FX was given i.v to the
pregnant ewes once daily to mimic the human dosing with pulsatile FX treatment. Oral
administration of the drug, which is the norm in humans, was not attempted in this and
previous studies because of the concern about the impact of ruminant nutrition on absorption
of orally administered drugs in sheep.
The following conclusions can be made from the results of the current study:
1. The results from the developmental study showed a decrease in heart rate,
increase in arterial pressure and maturation of the Autonomic Nervous System (ANS)
system as the lambs aged. pH, pO2, O2 saturation increase, while pCO2, glucose, lactate,
Hb, urine flow rate decrease with age. The plasma cortisol level is high around the time

196
of birth and decreases progressively thereafter. The 1h activity rhythm, which only exists
and is dominant over the first month of life is associated with suckling. The changes in
some of the physiological variables including heart rate, SDNN, glucose, pO2, urine flow
rate occurred around the window of weaning time. Weaning was defined in our study as
the time window between when lambs started to first consume solid food to when the 1h
rhythm due to suckling disappeared. Alternatively, the weaning time could be determined
by measuring the level of plasma ketone bodies (acetoacetic acid and 3-hydroxybutyric
acid) in the lambs. Even though the neonatal rumen epithelium is not ketogenic, the
ruminal epithelium in a mature ruminant animal is able to produce ketone bodies, which
is supplied by the microbial fermentation of feed (Lane et al., 2002). In addition, the
developmental data also showed that at 3 days of age male lambs are slightly more
acidotic than female lambs. This is similar to the findings of Ishak et al in lambs
delivered preterm at 133 d, although in this study male lambs exhibited other differences
from female lambs, including a higher pCO2, blood lactate and glucose concentrations
and higher arterial pressure (Ishak et al., 2012). In preterm lambs, this reduced postnatal
cardio-respiratory adaptation in male lambs could contribute to a higher mortality rate in
male than female neonates, especially related to respiratory distress syndrome
(Ingemarsson et al., 1997; Ingemarsson, 2003; Khoury et al., 1985). The current data
suggest even with delivery at term, respiratory function or acid-base regulatory
mechanisms in male lambs are less effective than in females, although the sex differences
are much less than with preterm birth. At 3 days of age, LF power were also higher in
male than female lambs, possibly due to an influence of testosterone, which present in the

197
male lambs by 36h after birth (Savoie et al., 1981), on the ANS or due to an ANS
response to an unfavorable environment (i.e. lower pH level).
However, one limitation of the developmental study is that the variables were not
measured on a daily basis, and thus cannot provide an exact time when the changes occur
as well as the pattern of changes between the experimental intervals. In addition, the
lambs included in the developmental study were also used for a longitudinal study of FX
effects, although only as control animals.
2. Acute FX administration did not cause any significant effects on the feeding
behavior, rest-activity cycles, cardiovascular and respiratory functions and the cortisol
level in the ~ 4 day old postnatal lambs. Even though arterial blood pressure increased for
the first 10 mins and heart rate decreased at 2 mins after FX injection, the changes were
too minor and transient to have any clinical or physiological significance. Therefore, it is
less likely that SSRIs toxicity is the mechanism underlying poor neonatal adaptation in
human infants exposed to this drug and other SSRIs in utero. This is further confirmed by
the subsequent results.
3. Acute FX administration did not cause any significant effects in the lambs at
10 days and 3 months of age. However, at 1 month of age, transient hypoxemia occurred
at 5 mins after FX injection. The hypoxemia was more severe in males than females.
Lambs were also less active on the day of FX administration at this age. Similarly, at 6
months of age, transient hypoxemia also occurred at 5 mins after FX injection but there
was no sex difference. Behavioral observations showed lambs were less active but more
irritable following FX administration. At 1 year of age, arterial pressure increased

198
significantly for 30 mins following FX injection. The increase was more profound and
lasted longer in males. HRV increased at 30 mins after FX and hypoxemia also occurred
at 5 mins after FX injection. However, no changes in the HPA axis function were
observed. FX effects on arterial pressure and blood oxygenation only occurred in the
older lambs but not in the younger lambs potentially due to the lower level of platelets,
which are the main reservoir of peripheral 5-HT in the younger lambs. In humans, it has
been known that the platelet concentrations are lower in newborns and children than in
adults (Merritt & Davidson, 1933) and platelet 5-HT concentration in newborn is only ~
42% of the value in children and adults (Flachaire et al., 1990). Measurement of platelet
concentration, platelet 5-HT concentration and the magnitude of transient increase in
blood 5-HT concentration following FX administration in sheep as a function of age
could confirm this hypothesis. However, analysis of serotonin from plasma samples is
technically difficult. In this regard, I did attempt to measure plasma 5-HT concentration
after FX injection in several lambs at 3 months of age but the results were very variable.
This was likely due to platelet/aggregation during sample collection, resulting in release
of serotonin from the platelets into blood. The sex differences in FX effects on arterial
pressure in the older lambs could be due to the effect of ovarian steroids on increasing 5-
HT clearance by the lung in the females, leading to a lesser transient increase in blood 5-
HT levels following FX injection (Bakhle & Ben-Harari, 1979; Gillis & Pitt, 1982). In
addition, our data showed that there was a relationship between FX concentration and the
changes in respiratory effects in the 1 year old lambs. The higher the FX concentration,
the greater decrease in O2 saturation. Dose-related effects of SSRIs have also been
observed in human and animal studies. In particular, increased risk of preterm birth was

199
mainly found in women treated with high dose SSRIs during pregnancy (Roca et al.,
2011). Increased fetal behaviors were also observed at standard and high SSRIs doses,
but not at the low dose and the effect at higher doses was greater than at standard doses.
(Mulder et al., 2011) In a rat study, respiratory rate reduced at standard doses and further
decreased at the higher doses of FX, while no changes occurred at the low dose of FX.
(Annerbrink et al., 2010) Moreover, in our study cardiovascular and respiratory effects of
FX were also significantly related. The greater the increased arterial pressure, the greater
the decreased pO2 concentration. As the mechanism of action of FX in inhibiting
serotonin reuptake by platelets leads to an acute increase in plasma serotonin, and since
serotonin is a potent vasoconstrictor, it can cause a sudden increase in blood pressure.
This could result in a decrease in cardiac output due to an afterload effect, which would
reduce pulmonary blood flow and result in the decrease in arterial pO2.
4. Postnatal lambs who were prenatally exposed to FX showed an increase in
activity for the first 2 weeks of life, and a lower HRV (time domain indexes: SDNN, SD
Delta NN, RMSSS) on postnatal day 2. Measures of birth and postnatal outcomes such as
gestational length, birth weight, postnatal weight gain, tissue sPo2 were not affected by
late gestation FX exposure. The increase in activity in the FX-exposed lambs existed in
the absence of increase in suckling activity, which is consistent with no difference in
postnatal weight gain between FX-exposed and control lambs. Measurement of a low
level of plasma FX concentration at birth and on the following postnatal day 2,5,10 and
14 confirmed that FX toxicity is less likely to be the underlying mechanism for poor
neonatal adaptation observed in human infants. Since the significant increase in activity
in the FX-exposed lambs existed until postnatal day 14, while FX level was low from

200
postnatal day 2, a withdrawal syndrome is also less likely to be the underlying
mechanism. Thus this leaves the mechanism of an SSRI-induced alteration in fetal brain
development to be related to the importance of serotonin in brain maturation pathways as
the most likely candidate. Indeed, an increase in fetal activity and a lower HRV through
the day have been observed in the human fetuses exposed to SSRIs in utero (Mulder et
al., 2011; Rurak et al., 2011). Measurement of serotonin concentrations in the
cerebrospinal fluid would provide direct evidence for the effects of FX on the lambs’
serotonin brain levels. Alternatively, S100B protein has been used as a biomarker for
early brain development and central serotonergic function (Pawluski et al., 2009). An
increase in synaptic 5-HT after SSRIs administration activates the astroglial 5-HT1A
receptor, which leads to a release of S100B protein which mediates the growth and
survival of neurons (Whitaker-Azmitia et al., 1990; Whitaker-Azmitia, 2001). Therefore,
in the absence of cerebrospinal fluid samples, determination of S100B protein could
provide a marker for changes in extracellular serotonin caused by FX administration. The
possibility of measuring plasma S100B concentration in my study arose late in the study,
after completion of the developmental studies. Unfortunately, due to the lack of
remaining blood samples in the control group, S100B protein analysis was not performed
in our studies.
In addition, from our studies it appeared that acute postnatal and long-term
prenatal FX exposure exert opposite effects. While acute FX injection caused a reduction
in activity, subchronic prenatal FX exposure resulted in hyperactivity in the lambs. Since
clinically it usually takes about 2 to 4 weeks for an SSRIs fully work, acute FX
administration could elicit a paradoxical antidepressant effect (i.e. sedation rather than

201
hyperactivity), which is different from the long-term exposure. There was a case report of
a paradoxical sedative effects at the onset of FX treatment, but this effect disappeared
after a few weeks of continuous FX therapy (Gupta et al., 1994). Moreover, it has been
documented that FX treatment is associated with a significant increase in the rate of both
activating adverse events including insomnia, agitation, anxiety, nervousness, and
sedating events such as somnolence or asthenia, as compared to placebo treatment in
patients with different baseline psychomotor activity status (agitated, retarded or neither).
Interestingly, 10% of patients that experienced adverse events actually reported both
effects, either simultaneously or serially. (Beasley et al., 1991)
Alternatively, it has been shown in a previous study in our lab that an enhanced
prepartum cortisol surge is associated with long-term prenatal FX-exposure in the fetal
lambs (Morrison et al., 2004). This could result in an early maturation of the musculo-
skeletal system, and thus explain the observed hyperactivity in the postnatal lambs
exposed to FX prenatally in our current study.
In summary, my study is the first to investigate postnatal outcomes in lambs exposed
to FX in utero or postnatally. In this study, we have two study cohorts: one was exposed to
FX acutely after birth and the other was exposed prenatally to FX for 2 weeks of late
gestation, which is equivalent to the last month of third trimester in human pregnancy. Acute
FX administration on postnatal day 4 did not cause any significant effects other than slight
and transient changes in heart rate and arterial pressure. In contrast, in the ~ 2 weeks
gestational FX exposure lambs, we observed a decreased time required for initial lamb
milestones (i.e. first time to stand, to walk and to suckle), a reduced HRV on postnatal day 2,
a hyperactivity not associated with suckling. (see Table 7.1; A vs C)

202
Human studies (Table 7.2 B & C) have shown that antenatal SSRIs exposure results
in many adverse birth outcomes in the offspring such as shorter gestational age, increased
risk of preterm birth, lower birth weight, small for gestational age, reduced postnatal weight
gain, poor neonatal adaptation (respiratory difficulty, cyanosis on feeding, irritability), lower
APGAR score, increased admission rate to NICU and reduced endocrine and facial response
to pain stimulus (via vaccination) at 1 and 3 months of age (Chambers et al., 1996; Costei et
al., 2002; Goldstein et al., 1995; Kallen, 2004; Lattimore et al., 2005; Levinson-Castiel et al.,
2006; Lewis et al., 2010; Lund et al., 2009; Maschi et al., 2008; Oberlander et al., 2005;
Oberlander et al., 2006; Oberlander et al., 2008; Roca et al., 2011; Simon et al., 2002; Toh et
al., 2009; Wen et al., 2006; Wisner et al., 2009; Zeskind & Stephens, 2004). However, we
did not observe any differences in the corresponding outcomes reported in human studies
such as gestational age, birth weight, postnatal weight gain in the lambs whose mothers were
administered FX i.v for ~ 12 days late gestation (see Table 7.1 C vs Table 7.2 B).
Differences observed in my study and human studies could be due to several reasons.
In human studies, the infants were exposed to the drugs for all or most of pregnancy, whereas
in my study, the exposure period was limited for ~ 12 days late gestation. In addition, women
who were treated with antidepressants were also clinically depressed; depression per se, and
its associated risk factors such as smoking, alcohol and drug abuse could be major
confounders for adverse effects in the offspring. As far as we know, our sheep were not
depressed. In the current study, we only investigate one subtype of SSRIs, fluoxetine, while
in human studies, many subtype of SSRIs (paroxetine, citalopram, sertraline, fluvoxamine,
etc.) and other antidepressants (TCA, SNRIs) were included.

203
Similar to our results, in one rodent study, there were no differences in body weight at
postnatal day 1, 8, 15, 22 between pups exposed to saline or FX during pregnancy and
lactation (from GA 0 to PND 21) (Lisboa et al., 2007). Other studies in rodents have also
shown changes in postnatal behavior (long-term effects) in mice/rats exposed to SSRIs
postnatally over the range from postnatal day 0 to 21 (see Table 7.3B) (Ansorge et al., 2004;
Hansen et al., 1997; Lee, 2009; Lisboa et al.,2007; Maciag et al., 2006; Manhaes de Castro et
al., 2001; Mirmiran et al., 1981; Popa et al., 2008). For instance, Lisboa et al, 2007 reported
that female FX-exposed mice had an increased immobility in forced swimming tests on PND
30 and 70 while male FX-exposed mice decreased their impulsivity on PND 40 and 70
(Lisboa et al., 2007). In the rat, neonatal exposure to citalopram (SSRI) from postnatal day 8
to 21 resulted in a decreased synthesis of tryptophan hydroxylase in the dorsal raphe and 5-
HT transporter expression in the cortex, increased locomotor activity and decreased sexual
behavior (male) in adult rats. (Maciag et al., 2006). The observed hyperactivity in our
prenatally FX-exposed lambs is similar to the increased locomotor activity in the Maciag’
study but different from the remaining rodent studies which reported an increased immobility
following SSRI exposure. (see Table 7.3 B). Despite this, the results of chronic FX or SSRI
exposure in my study, human studies (Table 7.2 B &C) and rodent studies (Table 7.3 A &B)
showed that there are long-term effects in the offspring exposed to SSRI (FX). These long-
term effects following FX exposure has been identified to be mediated by 5-HT transporter
blockade in rodent models that were exposed to FX early ex utero (PND 4-21) (Ansorge et
al, 2004). Interestingly, the effect of antenatal SSRIs exposure in human and sheep studies
actually appears to emerge in the fetal period before the treatment was ended. Indeed, a
blunted fetal heart rate response during the day, a reduced MCA pulsatile index, increased

204
activities in human fetuses on non-REM sleep, increased jerky movement and lower
reactivity to the vibroacoustic stimulus were observed (Mulder et al., 2011; Rurak et al.,
2011; Salisbury et al., 2006) (see Table 7.2A). In the sheep fetuses, 8 day continuous
maternal i.v FX infusion led to a reduced REM sleep, increase quiet sleep, enhanced
prepartum ACTH and cortisol surge and a transient decrease in uterine artery blood flow
(Morrison et al., 2001; Morrison, 2001; Morrison et al., 2002; Morrison et al., 2004;
Morrison et al., 2005). Taken together, the results from this study contribute to elucidating
the mechanisms of underlying poor neonatal adaptation in humans exposed to SSRI in utero.
So far, data from the current study and other relevant studies have pointed to the mechanism
of SSRIs-elicited altered fetal brain development. Particularly, prenatal SSRIs exposure
might alter 5-HT signaling in utero. This is important in the context of a “fetal programming”
impact on adult health. We hope the data gained from the present study contributes valuable
knowledge, and could assist physicians and pregnant women in the assessment of the safety
of the drugs, and to make the best informed decisions based on the risks and benefits of
antidepressant treatment during pregnancy.
In this study, the postnatal follow-up of prenatal FX exposure was limited only to 2
weeks following birth. Considering the increased activity in the newborn lambs is equivalent
to jitteriness, one of the symptoms of poor neonatal adaptation in human exposed in utero to
the drug, it would be very interesting to follow up their behaviors at longer time beyond this
2 week period, and at milestones of development, such as weaning (at 2 months), puberty (at
6 months) and adulthood (at 12 months) by actiwatch and DVR observations. A study in
human has reported that increased aggressiveness scores in the 4-year old children exposed
to SSRIs during gestation is associated with a history of poor neonatal adaptaion, however,

205
current maternal mood and parental stress appear to be the best predictors for the
externalizing behaviors than a history of prenatal depression or SSRIs exposure. (Oberlander
et al, 2007) The challenge to distinguish between the effects of the drug exposure and a
negative influence of postnatal social environment with depressed mothers on neurocognitive
development in the children in human study might be resolved using our sheep model.
Further studies to confirm the hypothesis of in utero altered 5-HT signaling in
prenatal SSRI exposure could be accomplished by investigating the fetal brain development
in SSRIs exposure subjects. It would be ideal to be able to assess fetal brain structure
development by invasive methods such as Magnetic Resonance Imaging (MRI), Positron
Emission Tomography (PET) and follow-ups evaluating the structure of brain development
and function after birth and up to adulthood. Alternatively, the effect of SSRIs on fetal brain
development could be evaluated by serial measuring plasma S100B protein, a biomarker of
early brain development and central serotonergic function, in the sheep fetuses exposed to
SSRIs (FX) following up to postnatal period into adulthood. In addition, it would be
interesting to investigate the transgenerational effects on fetal programming of altered 5-HT
signaling pathways in utero. Given the time required for a gestation of 145 days and
additional 1-2 years for the female lambs to be able to reproduce, the sheep model is
probably not an appropriate model for these long-term studies. Rodents or guinea pigs might
be better models for this type of study.

206
Table 7.1: Outcomes of chronic antenatal and acute postnatal FX exposure in fetal and postnatal lambs
Sheep studies Agents Outcomes
(A) Present study Acute FX i.v (1mg/kg) on PN
day 4
No differences in feeding behavior, postnatal weight gain, no respiratory difficulties, no
differences in cortisol level
Slight and transient heart rate and arterial pressure changes
(A) Present study Acute FX i.v (1mg/kg) at ~ 10
days, 1,3,6 and 12 months of age
Increased arterial pressure in the 1 year old lambs
Transient hypoxemia in the older lambs (1,6 and 12 months of age)
Decrease activities (1 & 6 months of age)
No changes in HPA axis function
(B) Morrison et al, 2001
(B) Morrison et al, 2002
(B) Morrison et al, 2004
Maternal FX i.v infusion x 8
days (~125-140 GA)
(98.5µg/kg.d)
Altered fetal behavioral state: Increased quiet sleep (low-voltage ECoG activity), eye movements
and fetal breathing movements. Decreased REM sleep (high-voltage ECoG activity)
Transient decrease in uterine artery blood flow
Transient fetal hypoxemia and acidosis for the first 24h of infusion
Increased prepartum ACTH & cortisol surges
(C) Present study Maternal daily FX (50mg) i.v x
~12 days (~131-143 GA)
No differences in gestational age, birth weight
Decreased time required for initial lamb milestones (to stand, to walk, to suckle)
Reduced HRV on PN day 2
PN 2- 14: hyperactivity not associated with suckling, no differences in postnatal weight gain, no
respiratory difficulties

207
Table 7.2: Outcomes of antenatal SSRI exposure in human (exposure for most or all of pregnancy)
(A) Human studies Agents Outcomes
Mulder et al, 2011
Prospective study
SSRIs throughout gestation Fetuses:
Disrupted non-REM sleep
Increase bodily activity during non-REM sleep
Rurak et al, 2011
Longitudinal observational study
SSRIs throughout gestation Fetuses (36 weeks gestation)
Blunted fetal HRV
Lower MCA pulsatility index: MCA mean flow velocity, MCA volume flow, MCA cross-sectional area
Salisbury et al, 2006
Prospective study
SSRIs + no MDD vs MDD
+ no SSRIs vs MDD +
SSRIs vs control
At 26 weeks GA:
Less movement, more jerky movements, lower reactivity to vibroacoustic stimulus
At 36 weeks GA:
SSRI or MDD fetuses: increased reactivity to vibroacoustic stimulus
SSRI + MDD fetuses: decreased reactivity to vibroacoustic stimulus
Newborns:
Increased tremor, startles, hypotonia and lowest quality of movement scores

208
(B) Human studies Agents Outcomes (short-term effects)
Chambers et al, 1996
Prospective/ retrospective reports
FX
Early vs late gestation
exposure
Newborn:
Increase risk of poor neonatal adaptation
Increase risk of special- nursery admission
Simon et al, 2002
Retrospective records
SSRIs
TCA
Shorter gestational age in SSRIs exposed infants
Lower birth weight
Lower Apgar scores (3rd trimester SSRI exposure)
Kallen et al, 2004
Prospective cohort study
SSRIs (Citalopram, FX, PX,
Sertraline)
Increased risk of respiratory distress
Increased risk of low Apgar score
Increased risk of convulsions
Wen et al, 2006
Retrospective cohort study
SSRIs Increased risk of low birth weight, preterm birth, fetal death, and seizures
Wisner et al, 2009
Prospective observational
SSRIs Increased risk of preterm birth in continuously SSRIs exposure across gestation

209
(B) Human studies Agents Outcomes (short-term effects)
Oberlander et al, 2006
Retrospective record-linkage
study
SSRIs (1st trimester to
delivery)
Increased risk of low birth weight
Increased risk of respiratory distress
Roca et al. 2011
Case-control study
SSRIs Increased risk of preterm birth (mainly in high dose)
Lund et al, 2009
Prospective cohort study
SSRIs during pregnancy Increased risk of preterm birth, lower 5-min Apgar score and NICU admission
Lewis et al, 2010
Prospective case-control study
Antidepressants Increased rate of preterm birth, lower birth weight, smaller head circumference
Toh et al. 2009
Retrospective cohort study
SSRIs vs non-SSRIs
antidepressants
Increased rate of smaller than gestational age in late gestational SSRIs exposure
Increased rate of preterm birth and smaller than gestational age in non-SSRIs antidepressant exposure
Levinson-Castiel et al, 2006
Cohort study
SSRIs (PX, FX, Citalopram,
Sertraline, Venlafaxine)
Neonatal abstinence syndrome in 30% SSRIs exposed neonates (Finnegan score)

210
(B) Human studies Agents Outcomes (short-term effects)
Goldstein et al, 1995
Prospective & retrospective
study
FX (3rd trimester) Incidence of poor neonatal adaptation to FX exposure: 13.4%
Increased risk of respiratory signs
Increased risk of sleep problems
Maschi et al, 2008
Prospective cohort study
PX, FX, other
antidepressants
Increased incidence of poor neonatal adaptation but not statistical significant
Costei et al, 2002
Prospective study
Paroxetine (early vs late
gestational vs no exposure
Increased risk of neonatal complications (respiratory distress, hypoglycemia, bradycardia, suckling
problems, prolonged hospitalization)
Increased risk of respiratory difficulties
Zeskind et al. 2004
Prospective study
SSRIs (citalopram, FX, PX,
sertraline)
During pregnancy
Shorter gestational age
Increase motor activity (*) and tremulous
Fewer rhythms in HRV (*)
Fewer changes in behavioral state, fewer different behavioral states and lower peak behavioral state
Increased REM sleep
(*) no longer significant after adjusted for gestational age

211
(C) Human studies Agents Outcomes (long-term effects)
Lewis et al, 2010
Prospective case-control study
Antidepressants At 1 month, lower body weight in exposed infants
Oberlander et al, 2005
Prospective study
SSRIs (FX, PX, sertraline)
(prenatal and postnatal
exposure)
At 2 months of age:
Blunted facial response to heel lance in prenatal SSRI exposure infants
Reduced parasympathetic withdrawal
Increased parasympathetic cardiac modulation during recovery in pre and postnatal SSRI exposure
infants
Oberlander et al, 2008
Prospective study
SSRIs Reduced early evening basal cortisol level
At 3 months of age:
Higher post-stress cortisol levels in SSRIs exposed infants.

212
Table 7.3: Postnatal outcomes of antenatal and postnatal SSRI exposure in rodents
(A) Rodent studies Agents Outcomes
Forcelli et al, 2008 FX
Prenatal infusion for 14 days
PN 30: decreased conflict-exploratory behavior
PN 60: increased place preference
PN 90: increased extinction response rate
PN 120: decreased nucleus accubens cell count & serotonin transporter-like in raphe nucleus
(B) Rodent studies Agents Outcomes
Lisboa et al, 2007 FX
GA 0-PN 21 to dams
7.5mg/kg gavage
No differences in body weight @ PN day 1,8,15,22
PN 30, 70: Increased immobility (female)
PN 40, 70: decreased impulsivity (male)
Lee et al, 2009 FX
PN 0-6
10mg/kg s.c
PN day 7-35:
Neuronal structure deformation in the somatosensory cortex
PN day 30-35
Higher thermal threshold (hotplate test)
Impaired whisker-specific tactile function (gap-crossing test)
Altered exploratory behavior (open field test)

213
(B) Rodent studies Agents Outcomes
Ansorge et al, 2004 FX
PN 4-21
10mg/kg i.p
At 12 weeks of age:
Decreased exploratory behavior in open field and elevated plus maze test
Prolonged latency to begin feeding in novelty-suppressed feeding test
Prolonged latency to escape a foot-shock in shock-escape paradigm
Mashaes de Castro et al,
2001
Citalopram
PN 1-19
20mg/kg s.c every 3 days
PN day 90-120:
Decreased aggressive behavior in footshock test
Hansen et al, 1997 SSRIs (LU)
PN 8-21
2.5-15mg/kg bid
At 4 months:
No difference in open field test & social interaction test
Increased immobile time in forced swim test
Maciag et al, 2006 Citalopram
PN 8-21
5mg/kg x 2 s.c
Adult:
Decreased synthesis of tryptophan hydroxylase in dorsal raphe & 5-HTT expression in cortex
Increased locomotor activity
Decreased sexual behavior

214
(B) Rodent studies Agents Outcomes
Popa et al, 2008 Escitalopram
PN 5
10mg/kg s.c x 2 weeks daily
Sleep anomalies
Anhedonia
Increased helplessness
Increased response to acute stress
Increased serotonergic autoinhibitory feedback
Mirmiran et al, 1981 Chlorimipramine
PN 8-21
x 2 daily
Decreased active sleep, increased quiet sleep
Decreased exploratory behavior in open field test
Decreased temporal learning task
Increased spatial task

215
REFERENCES
2007 > FDA proposes new warnings about suicidal thinking, behavior in young adults who take antidepressant medications Retrieved 11/13/2012, 2012, from
http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/2007/ucm108905.htm
Ahluwalia, I. B., Mack, K. A., & Mokdad, A. (2004). Mental and physical distress and high-risk behaviors among reproductive-age women. Obstetrics and Gynecology, 104(3), 477-483.
Ahmed, I., Dagincourt, P. G., Miller, L. G., & Shader, R. I. (1993). Possible interaction between fluoxetine and pimozide causing sinus bradycardia. Canadian Journal of Psychiatry.Revue Canadienne De Psychiatrie, 38(1), 62-63.
Alexander, G., Mills, S. C., & Scott, T. W. (1968). Changes in plasma glucose, lactate and free fatty acids in lambs during summit metabolism and treatment with catecholamines. The Journal of Physiology, 198(2), 277-289.
Alexander, G., & Williams, D. (1968). Shivering and non-shivering thermogenesis during summit metabolism in young lambs. The Journal of Physiology, 198(2), 251-276.
Alfaro, C. L., Lam, Y. W., Simpson, J., & Ereshefsky, L. (2000). CYP2D6 inhibition by fluoxetine, paroxetine, sertraline, and venlafaxine in a crossover study: Intraindividual variability and plasma concentration correlations. Journal of Clinical Pharmacology, 40(1), 58-66.
Alfirevic, Z., Devane, D., & Gyte, G. M. (2006). Continuous cardiotocography (CTG) as a form of electronic fetal monitoring (EFM) for fetal assessment during labour. Cochrane Database of Systematic Reviews (Online), (3)(3), CD006066.
Altamura, A. C., Moro, A. R., & Percudani, M. (1994). Clinical pharmacokinetics of fluoxetine. Clinical Pharmacokinetics, 26(3), 201-214.
Altman, D. (1991). Some common problems in medical research. Practical statistics for medical research (pp. 426-439). London: Chapman & Hall.
Altshuler, L. L., Cohen, L. S., Moline, M. L., Kahn, D. A., Carpenter, D., Docherty, J. P., et al. (2001). Treatment of depression in women: A summary of the expert consensus guidelines. Journal of Psychiatric Practice, 7(3), 185-208.
Amsterdam, J. D., Garcia-Espana, F., Fawcett, J., Quitkin, F. M., Reimherr, F. W., Rosenbaum, J. F., et al. (1999). Blood pressure changes during short-term fluoxetine treatment. Journal of Clinical Psychopharmacology, 19(1), 9-14.
Anderson, G. M. (2004). Peripheral and central neurochemical effects of the selective serotonin reuptake inhibitors (SSRIs) in humans and nonhuman primates: Assessing

216
bioeffect and mechanisms of action. International Journal of Developmental Neuroscience : The Official Journal of the International Society for Developmental Neuroscience, 22(5-6), 397-404.
Anderson, R. N. (2002). Deaths: Leading causes for 2000. National Vital Statistics Reports : From the Centers for Disease Control and Prevention, National Center for Health Statistics, National Vital Statistics System, 50(16), 1-85.
Andersson, L., Sundstrom-Poromaa, I., Wulff, M., Astrom, M., & Bixo, M. (2004a). Implications of antenatal depression and anxiety for obstetric outcome. Obstetrics and Gynecology, 104(3), 467-476.
Andersson, L., Sundstrom-Poromaa, I., Wulff, M., Astrom, M., & Bixo, M. (2004b). Neonatal outcome following maternal antenatal depression and anxiety: A population-based study. American Journal of Epidemiology, 159(9), 872-881.
Andrade, C., Bhakta, S. G., & Singh, N. M. (2006). Controversy revisited: Selective serotonin reuptake inhibitors in paediatric depression. The World Journal of Biological Psychiatry : The Official Journal of the World Federation of Societies of Biological Psychiatry, 7(4), 251-260.
Andrade, S. E., McPhillips, H., Loren, D., Raebel, M. A., Lane, K., Livingston, J., et al. (2009). Antidepressant medication use and risk of persistent pulmonary hypertension of the newborn. Pharmacoepidemiol Drug Saf, 18(3), 246-252.
Andrade, S. E., Raebel, M. A., Brown, J., Lane, K., Livingston, J., Boudreau, D., et al. (2008). Use of antidepressant medications during pregnancy: A multisite study. American Journal of Obstetrics and Gynecology, 198(2), 194.e1-194.e5.
Annerbrink, K., Olsson, M., Hedner, J., & Eriksson, E. (2010). Acute and chronic treatment with serotonin reuptake inhibitors exert opposite effects on respiration in rats: Possible implications for panic disorder. Journal of Psychopharmacology (Oxford, England), 24(12), 1793-1801.
Annerbrink, K., Olsson, M., Melchior, L. K., Hedner, J., & Eriksson, E. (2003). Serotonin depletion increases respiratory variability in freely moving rats: Implications for panic disorder. The International Journal of Neuropsychopharmacology / Official Scientific Journal of the Collegium Internationale Neuropsychopharmacologicum (CINP), 6(1), 51-56.
Ansorge, M. S., Zhou, M., Lira, A., Hen, R., & Gingrich, J. A. (2004). Early-life blockade of the 5-HT transporter alters emotional behavior in adult mice. Science, 306(5697), 879-881.
Appleby, M., Mbewu, A., & Clarke, B. (1995). Fluoxetine and ventricular torsade--is there a link? International Journal of Cardiology, 49(2), 178-180.

217
Arborelius, L., Owens, M. J., Plotsky, P. M., & Nemeroff, C. B. (1999). The role of corticotropin-releasing factor in depression and anxiety disorders. The Journal of Endocrinology, 160(1), 1-12.
Asarnow, J. R., Jaycox, L. H., Duan, N., LaBorde, A. P., Rea, M. M., Tang, L., et al. (2005). Depression and role impairment among adolescents in primary care clinics. The Journal of Adolescent Health : Official Publication of the Society for Adolescent Medicine, 37(6), 477-483.
Asch, D. A., & Parker, R. M. (1988). The libby zion case. one step forward or two steps backward? The New England Journal of Medicine, 318(12), 771-775.
Bakhle, Y. S., & Ben-Harari, R. R. (1979). Effects of oestrous cycle and exogenous ovarian steroids on 5-hydroxytryptamine metabolism in rat lung. The Journal of Physiology, 291, 11-18.
Barouki, R., Gluckman, P. D., Grandjean, P., Hanson, M., & Heindel, J. J. (2012). Developmental origins of non-communicable disease: Implications for research and public health. Environmental Health : A Global Access Science Source, 11, 42.
Battaglino, R., Fu, J., Spate, U., Ersoy, U., Joe, M., Sedaghat, L., et al. (2004). Serotonin regulates osteoclast differentiation through its transporter. Journal of Bone and Mineral Research : The Official Journal of the American Society for Bone and Mineral Research, 19(9), 1420-1431.
Beasley, C. M.,Jr, Sayler, M. E., Bosomworth, J. C., & Wernicke, J. F. (1991). High-dose fluoxetine: Efficacy and activating-sedating effects in agitated and retarded depression. Journal of Clinical Psychopharmacology, 11(3), 166-174.
Beck, C. T. (1996). A meta-analysis of the relationship between postpartum depression and infant temperament.
Bellivier, F., Leboyer, M., Courtet, P., Buresi, C., Beaufils, B., Samolyk, D., et al. (1998). Association between the tryptophan hydroxylase gene and manic-depressive illness. Archives of General Psychiatry, 55(1), 33-37.
Belmaker, R. H., & Agam, G. (2008). Major depressive disorder. The New England Journal of Medicine, 358(1), 55-68.
Benfield, P., Heel, R. C., & Lewis, S. P. (1986). Fluoxetine. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic efficacy in depressive illness. Drugs, 32(6), 481-508.
Bernstein, G. A., Massie, E. D., Thuras, P. D., Perwien, A. R., Borchardt, C. M., & Crosby, R. D. (1997). Somatic symptoms in anxious-depressed school refusers. Journal of the American Academy of Child and Adolescent Psychiatry, 36(5), 661-668.

218
Berry, L. M., Ikegami, M., Woods, E., & Ervin, M. G. (1995). Postnatal renal adaptation in preterm and term lambs. Reproduction, Fertility, and Development, 7(3), 491-498.
Bhagwagar, Z., Hafizi, S., & Cowen, P. J. (2002). Acute citalopram administration produces correlated increases in plasma and salivary cortisol. Psychopharmacology, 163(1), 118-120.
Bhatia, S. C., & Bhatia, S. K. (1999). Depression in women: Diagnostic and treatment considerations. American Family Physician, 60(1), 225-34, 239-40.
Binder, E. B., Kunzel, H. E., Nickel, T., Kern, N., Pfennig, A., Majer, M., et al. (2009). HPA-axis regulation at in-patient admission is associated with antidepressant therapy outcome in male but not in female depressed patients. Psychoneuroendocrinology, 34(1), 99-109.
Birmaher, B., Brent, D. A., & Benson, R. S. (1998). Summary of the practice parameters for the assessment and treatment of children and adolescents with depressive disorders. american academy of child and adolescent psychiatry. Journal of the American Academy of Child and Adolescent Psychiatry, 37(11), 1234-1238.
Birmaher, B., Ryan, N. D., Williamson, D. E., Brent, D. A., Kaufman, J., Dahl, R. E., et al. (1996). Childhood and adolescent depression: A review of the past 10 years. part I. Journal of the American Academy of Child and Adolescent Psychiatry, 35(11), 1427-1439.
Birmes, P., Coppin, D., Schmitt, L., & Lauque, D. (2003). Serotonin syndrome: A brief review. CMAJ : Canadian Medical Association Journal = Journal De l'Association Medicale Canadienne, 168(11), 1439-1442.
Boadle-Biber, M. C., Singh, V. B., Corley, K. C., Phan, T. H., & Dilts, R. P. (1993). Evidence that corticotropin-releasing factor within the extended amygdala mediates the activation of tryptophan hydroxylase produced by sound stress in the rat. Brain Research, 628(1-2), 105-114.
Borys, D. J., Setzer, S. C., Ling, L. J., Reisdorf, J. J., Day, L. C., & Krenzelok, E. P. (1990). The effects of fluoxetine in the overdose patient. Journal of Toxicology.Clinical Toxicology, 28(3), 331-340.
Boyer, E. W., & Shannon, M. (2005). The serotonin syndrome. The New England Journal of Medicine, 352(11), 1112-1120.
Boyer, W. F. (1992). Potential indications for the selective serotonin reuptake inhibitors. International Clinical Psychopharmacology, 6 Suppl 5, 5-12.
Bradford, L. D. (2002). CYP2D6 allele frequency in european caucasians, asians, africans and their descendants. Pharmacogenomics, 3(2), 229-243.

219
Brent, D. A., Perper, J. A., Goldstein, C. E., Kolko, D. J., Allan, M. J., Allman, C. J., et al. (1988). Risk factors for adolescent suicide. A comparison of adolescent suicide victims with suicidal inpatients. Archives of General Psychiatry, 45(6), 581-588.
Bridge, J. A., Iyengar, S., Salary, C. B., Barbe, R. P., Birmaher, B., Pincus, H. A., et al. (2007). Clinical response and risk for reported suicidal ideation and suicide attempts in pediatric antidepressant treatment: A meta-analysis of randomized controlled trials. JAMA : The Journal of the American Medical Association, 297(15), 1683-1696.
Broadbear, J. H., Hutton, L. C., Clarke, I. J., & Canny, B. J. (2004). Sex differences in the pituitary-adrenal response following acute antidepressant treatment in sheep. Psychopharmacology, 171(4), 450-457.
Broadbear, J. H., Nguyen, T., Clarke, I. J., & Canny, B. J. (2004). Antidepressants, sex steroids and pituitary-adrenal response in sheep. Psychopharmacology, 175(2), 247-255.
Brummelte, S., & Galea, L. A. (2009). Depression during pregnancy and postpartum: Contribution of stress and ovarian hormones. Progress in Neuro-Psychopharmacology & Biological Psychiatry,
Buchanan, G. F., & Richerson, G. B. (2010). Central serotonin neurons are required for arousal to CO2. Proceedings of the National Academy of Sciences of the United States of America, 107(37), 16354-16359.
Burt, V. K., & Stein, K. (2002). Epidemiology of depression throughout the female life cycle. The Journal of Clinical Psychiatry, 63 Suppl 7, 9-15.
Cain, C. R., Hamilton, T. C., Norton, J., Petersen, E. N., Poyser, R. H., & Thormahlen, D. (1989). Relative lack of cardiotoxicity of paroxetine in animal models. Acta Psychiatr Scand Suppl, 350, 27-30.
Carey, M. P., Deterd, C. H., de Koning, J., Helmerhorst, F., & de Kloet, E. R. (1995). The influence of ovarian steroids on hypothalamic-pituitary-adrenal regulation in the female rat. The Journal of Endocrinology, 144(2), 311-321.
Carney, R. M., Blumenthal, J. A., Freedland, K. E., Stein, P. K., Howells, W. B., Berkman, L. F., et al. (2005). Low heart rate variability and the effect of depression on post-myocardial infarction mortality. Archives of Internal Medicine, 165(13), 1486-1491.
Casper, R. C., Fleisher, B. E., Lee-Ancajas, J. C., Gilles, A., Gaylor, E., DeBattista, A., et al. (2003). Follow-up of children of depressed mothers exposed or not exposed to antidepressant drugs during pregnancy. The Journal of Pediatrics, 142(4), 402-408.
Caspi, A., Sugden, K., Moffitt, T. E., Taylor, A., Craig, I. W., Harrington, H., et al. (2003). Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene. Science (New York, N.Y.), 301(5631), 386-389.

220
Celada, P., Puig, M., Amargos-Bosch, M., Adell, A., & Artigas, F. (2004). The therapeutic role of 5-HT1A and 5-HT2A receptors in depression. Journal of Psychiatry & Neuroscience : JPN, 29(4), 252-265.
Chadio, S. E., Kotsampasi, B., Papadomichelakis, G., Deligeorgis, S., Kalogiannis, D., Menegatos, I., et al. (2007). Impact of maternal undernutrition on the hypothalamic-pituitary-adrenal axis responsiveness in sheep at different ages postnatal. The Journal of Endocrinology, 192(3), 495-503.
Challis, J. R., Berdusco, E. T., Jeffray, T. M., Yang, K., & Hammond, G. L. (1995). Corticosteroid-binding globulin (CBG) in fetal development. The Journal of Steroid Biochemistry and Molecular Biology, 53(1-6), 523-527.
Challis, J. R. G., Matthews, S. G., Gibb, W., & Lye, S. J. (2000). Endocrine and paracrine regulation of birth at term and preterm. Endocrine Reviews, 21(5), 514-550.
Chambers, C. D., Anderson, P. O., Thomas, R. G., Dick, L. M., Felix, R. J., Johnson, K. A., et al. (1999). Weight gain in infants breastfed by mothers who take fluoxetine. Pediatrics, 104(5), e61.
Chambers, C. D., Hernandez-Diaz, S., Van Marter, L. J., Werler, M. M., Louik, C., Jones, K. L., et al. (2006). Selective serotonin-reuptake inhibitors and risk of persistent pulmonary hypertension of the newborn. N Engl J Med, 354(6), 579-587.
Chambers, C. D., Hernandez-Diaz, S., Van Marter, L. J., Werler, M. M., Louik, C., Jones, K. L., et al. (2006). Selective serotonin-reuptake inhibitors and risk of persistent pulmonary hypertension of the newborn. The New England Journal of Medicine, 354(6), 579-587.
Chambers, C. D., Johnson, K. A., Dick, L. M., Felix, R. J., & Jones, K. L. (1996). Birth outcomes in pregnant women taking fluoxetine. The New England Journal of Medicine, 335(14), 1010-1015.
Chow, T. W. (2013). The disposition of fluoxetine in newborn lambs up to one year of age. (Doctor of Philosophy, University of British Columbia).
Chow, T. W., Szeitz, A., Rurak, D. W., & Riggs, K. W. (2011). A validated enantioselective assay for the simultaneous quantitation of (R)-, (S)-fluoxetine and (R)-, (S)-norfluoxetine in ovine plasma using liquid chromatography with tandem mass spectrometry (LC/MS/MS). Journal of Chromatography.B, Analytical Technologies in the Biomedical and Life Sciences, 879(5-6), 349-358.
Clifton, P. G., Barnfield, A. M., & Philcox, L. (1989). A behavioural profile of fluoxetine-induced anorexia. Psychopharmacology, 97(1), 89-95.
Cohen, L. S., Altshuler, L. L., Harlow, B. L., Nonacs, R., Newport, D. J., Viguera, A. C., et al. (2006). Relapse of major depression during pregnancy in women who maintain or

221
discontinue antidepressant treatment. JAMA : The Journal of the American Medical Association, 295(5), 499-507.
Cohen, L. S., Heller, V. L., Bailey, J. W., Grush, L., Ablon, J. S., & Bouffard, S. M. (2000). Birth outcomes following prenatal exposure to fluoxetine. Biological Psychiatry, 48(10), 996-1000.
Cohen, P., Cohen, J., Kasen, S., Velez, C. N., Hartmark, C., Johnson, J., et al. (1993). An epidemiological study of disorders in late childhood and adolescence--I. age- and gender-specific prevalence. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 34(6), 851-867.
Cohn, J. B., & Wilcox, C. (1985). A comparison of fluoxetine, imipramine, and placebo in patients with major depressive disorder. The Journal of Clinical Psychiatry, 46(3 Pt 2), 26-31.
Comline, R. S., & Silver, M. (1970). Daily changes in foetal and maternal blood of conscious pregnant ewes, with catheters in umbilical and uterine vessels. The Journal of Physiology, 209(3), 567-586.
Compton, R., Spiller, H. A., & Bosse, G. M. (2005). Fatal fluoxetine ingestion with postmortem blood concentrations. Clinical Toxicology (Philadelphia, Pa.), 43(4), 277-279.
Cooper, P. J., & Murray, L. (1998). Postnatal depression. BMJ (Clinical Research Ed.), 316(7148), 1884-1886.
Cooper, W. O., Willy, M. E., Pont, S. J., & Ray, W. A. (2007). Increasing use of antidepressants in pregnancy. American Journal of Obstetrics and Gynecology, 196(6), 544.e1-544.e5.
Coppen, A., Eccleston, E. G., & Peet, M. (1973). Total and free tryptophan concentration in the plasma of depressive patients. Lancet, 2(7820), 60-63.
Costei, A. M., Kozer, E., Ho, T., Ito, S., & Koren, G. (2002). Perinatal outcome following third trimester exposure to paroxetine. Archives of Pediatrics & Adolescent Medicine, 156(11), 1129-1132.
Costello, E. J., Angold, A., Burns, B. J., Stangl, D. K., Tweed, D. L., Erkanli, A., et al. (1996). The great smoky mountains study of youth. goals, design, methods, and the prevalence of DSM-III-R disorders. Archives of General Psychiatry, 53(12), 1129-1136.
Cowen, P. J., Parry-Billings, M., & Newsholme, E. A. (1989). Decreased plasma tryptophan levels in major depression. Journal of Affective Disorders, 16(1), 27-31.

222
Davidson, S., Prokonov, D., Taler, M., Maayan, R., Harell, D., Gil-Ad, I., et al. (2009). Effect of exposure to selective serotonin reuptake inhibitors in utero on fetal growth: Potential role for the IGF-I and HPA axes. Pediatr Res, 65(2), 236-241.
Deave, T., Heron, J., Evans, J., & Emond, A. (2008). The impact of maternal depression in pregnancy on early child development. BJOG : An International Journal of Obstetrics and Gynaecology, 115(8), 1043-1051.
DeVane, C. L. (1999). Metabolism and pharmacokinetics of selective serotonin reuptake inhibitors. Cellular and Molecular Neurobiology, 19(4), 443-466.
DeVane, C. L. (2003). Immediate-release versus controlled-release formulations: Pharmacokinetics of newer antidepressants in relation to nausea. The Journal of Clinical Psychiatry, 64 Suppl 18, 14-19.
Deykin, E. Y., Buka, S. L., & Zeena, T. H. (1992). Depressive illness among chemically dependent adolescents. The American Journal of Psychiatry, 149(10), 1341-1347.
DiPietro, J. A., Novak, M. F., Costigan, K. A., Atella, L. D., & Reusing, S. P. (2006). Maternal psychological distress during pregnancy in relation to child development at age two. Child Development, 77(3), 573-587.
Dobbing, J., & Sands, J. (1979). Comparative aspects of the brain growth spurt. Early Human Development, 3(1), 79-83.
Dole, N., Savitz, D. A., Hertz-Picciotto, I., Siega-Riz, A. M., McMahon, M. J., & Buekens, P. (2003). Maternal stress and preterm birth. American Journal of Epidemiology, 157(1), 14-24.
Dubnov-Raz, G., Juurlink, D. N., Fogelman, R., Merlob, P., Ito, S., Koren, G., et al. (2008). Antenatal use of selective serotonin-reuptake inhibitors and QT interval prolongation in newborns. Pediatrics, 122(3), e710-5.
Dubovsky, S. L., Davies, R., & Dubovsky, A. N. (2003). Mood disorders. In S. C. Yudofsky (Ed.), Textbook of clinical psychiatry (4th ed., pp. 439-542). Washington, DC: Washington, DC : American Psychiatric Pub.
Dunlop, B. W., & Nemeroff, C. B. (2007). The role of dopamine in the pathophysiology of depression. Archives of General Psychiatry, 64(3), 327-337.
Dwyer, C. M. (2003). Behavioural development in the neonatal lamb: Effect of maternal and birth-related factors. Theriogenology, 59(3-4), 1027-1050.
Dwyer, C. M. (2008a). Genetic and physiological determinants of maternal behavior and lamb survival: Implications for low-input sheep management. Journal of Animal Science, 86(14 Suppl), E246-58.

223
Dwyer, C. M. (2008b). The welfare of the neonatal lamb. Small Ruminant Research, 76, 31-41.
Dwyer, C. M., Calvert, S. K., Farish, M., Donbavand, J., & Pickup, H. E. (2005). Breed, litter and parity effects on placental weight and placentome number, and consequences for the neonatal behaviour of the lamb. Theriogenology, 63(4), 1092-1110.
Dwyer, C. M., McLean, K. A., Deans, L. A., Chirnside, J., Calvert, S. K., & Lawrence, A. B. (1998). Vocalisations between mother and young in sheep: Effects of breed and maternal experience. Applied Animal Behaviour Science, 58(1–2), 105-119.
Eaton, L. (2006). European agency approves use of fluoxetine for children and teens. BMJ (Clinical Research Ed.), 332(7555), 1407.
Egger, H. L., Costello, E. J., Erkanli, A., & Angold, A. (1999). Somatic complaints and psychopathology in children and adolescents: Stomach aches, musculoskeletal pains, and headaches. Journal of the American Academy of Child and Adolescent Psychiatry, 38(7), 852-860.
Einarson, A., Choi, J., Einarson, T. R., & Koren, G. (2010). Adverse effects of antidepressant use in pregnancy: An evaluation of fetal growth and preterm birth. Depression and Anxiety, 27(1), 35-38.
Emslie, G. J., Rush, A. J., Weinberg, W. A., Gullion, C. M., Rintelmann, J., & Hughes, C. W. (1997). Recurrence of major depressive disorder in hospitalized children and adolescents. Journal of the American Academy of Child and Adolescent Psychiatry, 36(6), 785-792.
Ener, R. A., Meglathery, S. B., Van Decker, W. A., & Gallagher, R. M. (2003). Serotonin syndrome and other serotonergic disorders. Pain Medicine (Malden, Mass.), 4(1), 63-74.
Fairchild, K. D., & O'Shea, T. M. (2010). Heart rate characteristics: Physiomarkers for detection of late-onset neonatal sepsis. Clinics in Perinatology, 37(3), 581-598.
Feighner, J. P. (1995). Cardiovascular safety in depressed patients: Focus on venlafaxine. The Journal of Clinical Psychiatry, 56(12), 574-579.
Fjordside, L., Jeppesen, U., Eap, C. B., Powell, K., Baumann, P., & Brosen, K. (1999). The stereoselective metabolism of fluoxetine in poor and extensive metabolizers of sparteine. Pharmacogenetics, 9(1), 55-60.
Flachaire, E., Beney, C., Berthier, A., Salandre, J., Quincy, C., & Renaud, B. (1990). Determination of reference values for serotonin concentration in platelets of healthy newborns, children, adults, and elderly subjects by HPLC with electrochemical detection. Clinical Chemistry, 36(12), 2117-2120.

224
Fleming, J. E., & Offord, D. R. (1990). Epidemiology of childhood depressive disorders: A critical review. Journal of the American Academy of Child and Adolescent Psychiatry, 29(4), 571-580.
Forcelli, P. A., & Heinrichs, S. C. (2008). Teratogenic effects of maternal antidepressant exposure on neural substrates of drug-seeking behavior in offspring. Addict Biol, 13(1), 52-62.
Fornaro, E., Li, D., Pan, J., & Belik, J. (2007). Prenatal exposure to fluoxetine induces fetal pulmonary hypertension in the rat. American Journal of Respiratory and Critical Care Medicine, 176(10), 1035-1040.
Franco-Bronson, K. (1996). The management of treatment-resistant depression in the medically ill. The Psychiatric Clinics of North America, 19(2), 329-350.
Fraser, A. F., & Broom, D. M. (1997). Farm animal behaviour and welfare (3rd ed.). Wallingford, Oxon, UK ;New York, NY, USA: CAB International.
Fuller, R. W. (1996a). Mechanisms and functions of serotonin neuronal systems: Opportunities for neuropeptide interactions. Annals of the New York Academy of Sciences, 780, 176-184.
Fuller, R. W. (1996b). Serotonin receptors involved in regulation of pituitary-adrenocortical function in rats. Behavioural Brain Research, 73(1-2), 215-219.
Fuller, R. W., Perry, K. W., Hemrick-Luecke, S. K., & Engleman, E. (1996). Serum corticosterone increases reflect enhanced uptake inhibitor-induced elevation of extracellular 5-hydroxytryptamine in rat hypothalamus. The Journal of Pharmacy and Pharmacology, 48(1), 68-70.
Fuller, R. W., Snoddy, H. D., & Molloy, B. B. (1976). Pharmacologic evidence for a serotonin neural pathway involved in hypothalamus-pituitary-adrenal function in rats. Life Sciences, 19(3), 337-345.
Furness, J. B., & Costa, M. (1982). Neurons with 5-hydroxytryptamine-like immunoreactivity in the enteric nervous system: Their projections in the guinea-pig small intestine. Neuroscience, 7(2), 341-349.
Gardner, S. F., Rutherford, W. F., Munger, M. A., & Panacek, E. A. (1991). Drug-induced supraventricular tachycardia: A case report of fluoxetine. Annals of Emergency Medicine, 20(2), 194-197.
Gibbs, D. M., & Vale, W. (1983). Effect of the serotonin reuptake inhibitor fluoxetine on corticotropin-releasing factor and vasopressin secretion into hypophysial portal blood. Brain Research, 280(1), 176-179.

225
Gill, M., LoVecchio, F., & Selden, B. (1999). Serotonin syndrome in a child after a single dose of fluvoxamine. Annals of Emergency Medicine, 33(4), 457-459.
Gillespie, C. F., & Nemeroff, C. B. (2005). Hypercortisolemia and depression. Psychosomatic Medicine, 67 Suppl 1, S26-8.
Gillis, C. N., & Pitt, B. R. (1982). The fate of circulating amines within the pulmonary circulation. Annual Review of Physiology, 44, 269-281.
Gilmer, W. S., & McKinney, W. T. (2003). Early experience and depressive disorders: Human and non-human primate studies. Journal of Affective Disorders, 75(2), 97-113.
Girard, J., Chatelain, F., Boillot, J., Prip-Buus, C., Thumelin, S., Pégorier, J., et al. (1997). Nutrient regulation of gene expression. Journal of Animal Science, 75(Supplement 2), 46-57.
Glassman, A. H., & Shapiro, P. A. (1998). Depression and the course of coronary artery disease. The American Journal of Psychiatry, 155(1), 4-11.
Goel, N., & Bale, T. L. (2010). Sex differences in the serotonergic influence on the hypothalamic-pituitary-adrenal stress axis. Endocrinology, 151(4), 1784-1794.
Goldstein, B. J. (1998). SSRIs in depression: A decade of progress. Journal of Psychopharmacology (Oxford, England), 12(3 Suppl B), S3.
Goldstein, D. J. (1995). Effects of third trimester fluoxetine exposure on the newborn. Journal of Clinical Psychopharmacology, 15(6), 417-420.
Goodnick, P. J., & Goldstein, B. J. (1998a). Selective serotonin reuptake inhibitors in affective disorders--I. basic pharmacology. Journal of Psychopharmacology (Oxford, England), 12(3 Suppl B), S5-20.
Goodnick, P. J., & Goldstein, B. J. (1998b). Selective serotonin reuptake inhibitors in affective disorders--II. efficacy and quality of life. Journal of Psychopharmacology (Oxford, England), 12(3 Suppl B), S21-54.
Goodyer, I., Dubicka, B., Wilkinson, P., Kelvin, R., Roberts, C., Byford, S., et al. (2007). Selective serotonin reuptake inhibitors (SSRIs) and routine specialist care with and without cognitive behaviour therapy in adolescents with major depression: Randomised controlled trial. BMJ (Clinical Research Ed.), 335(7611), 142.
Gorman, J. M., Papp, L. A., Coplan, J. D., Martinez, J. M., Lennon, S., Goetz, R. R., et al. (1994). Anxiogenic effects of CO2 and hyperventilation in patients with panic disorder. The American Journal of Psychiatry, 151(4), 547-553.

226
Graatsma, E. M., Jacod, B. C., van Egmond, L. A., Mulder, E. J., & Visser, G. H. (2009). Fetal electrocardiography: Feasibility of long-term fetal heart rate recordings. BJOG : An International Journal of Obstetrics and Gynaecology, 116(2), 334-7; discussion 337-8.
Green, A. R., & Curzon, G. (1968). Decrease of 5-hydroxytryptamine in the brain provoked by hydrocortisone and its prevention by allopurinol. Nature, 220(5172), 1095-1097.
Greenwood, P. L., Hunt, A. S., Slepetis, R. M., Finnerty, K. D., Alston, C., Beermann, D. H., et al. (2002). Effects of birth weight and postnatal nutrition on neonatal sheep: III. regulation of energy metabolism. Journal of Animal Science, 80(11), 2850-2861.
Gresham, E. L., Rankin, J. H., Makowski, E. L., Meschia, G., & Battaglia, F. C. (1972). An evaluation of fetal renal function in a chronic sheep preparation. The Journal of Clinical Investigation, 51(1), 149-156.
Grippo, A. J., & Johnson, A. K. (2002). Biological mechanisms in the relationship between depression and heart disease. Neuroscience and Biobehavioral Reviews, 26(8), 941-962.
Gupta, S., & Rajaprabhakaran, R. (1994). A case report of paradoxical sedation with fluoxetine therapy. The Journal of Clinical Psychiatry, 55(3), 118.
Haddad, P. M., Pal, B. R., Clarke, P., Wieck, A., & Sridhiran, S. (2005). Neonatal symptoms following maternal paroxetine treatment: Serotonin toxicity or paroxetine discontinuation syndrome? J Psychopharmacol, 19(5), 554-557.
Hakkarainen, R., Partonen, T., Haukka, J., Virtamo, J., Albanes, D., & Lonnqvist, J. (2004). Is low dietary intake of omega-3 fatty acids associated with depression? The American Journal of Psychiatry, 161(3), 567-569.
Halford, J. C., Harrold, J. A., Boyland, E. J., Lawton, C. L., & Blundell, J. E. (2007). Serotonergic drugs : Effects on appetite expression and use for the treatment of obesity. Drugs, 67(1), 27-55.
Hammad, T. A., Laughren, T., & Racoosin, J. (2006). Suicidality in pediatric patients treated with antidepressant drugs. Archives of General Psychiatry, 63(3), 332-339.
Hansen, H. H., Sanchez, C., & Meier, E. (1997). Neonatal administration of the selective serotonin reuptake inhibitor lu 10-134-C increases forced swimming-induced immobility in adult rats: A putative animal model of depression? J Pharmacol Exp Ther, 283(3), 1333-1341.
Harper, R. M., Hoppenbrouwers, T., Sterman, M. B., McGinty, D. J., & Hodgman, J. (1976). Polygraphic studies of normal infants during the first six months of life. I. heart rate and variability as a function of state. Pediatric Research, 10(11), 945-948.

227
Harrington, R., Fudge, H., Rutter, M., Pickles, A., & Hill, J. (1990). Adult outcomes of childhood and adolescent depression. I. psychiatric status. Archives of General Psychiatry, 47(5), 465-473.
Harrington, R., Whittaker, J., Shoebridge, P., & Campbell, F. (1998). Systematic review of efficacy of cognitive behaviour therapies in childhood and adolescent depressive disorder. BMJ (Clinical Research Ed.), 316(7144), 1559-1563.
Harvey, A. T., & Preskorn, S. H. (2001). Fluoxetine pharmacokinetics and effect on CYP2C19 in young and elderly volunteers. Journal of Clinical Psychopharmacology, 21(2), 161-166.
Hay, D. F., Pawlby, S., Waters, C. S., & Sharp, D. (2008). Antepartum and postpartum exposure to maternal depression: Different effects on different adolescent outcomes. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 49(10), 1079-1088.
Hay, D. F., & Kumar, R. (1995). Interpreting the effects of mothers' post natal depression on children's intelligence: A critique and re-analysis. Child Psychiatry & Human Development, 25(3), 165-181.
Heikkinen, T., Ekblad, U., Kero, P., Ekblad, S., & Laine, K. (2002). Citalopram in pregnancy and lactation. Clin Pharmacol Ther, 72(2), 184-191.
Heikkinen, T., Ekblad, U., Palo, P., & Laine, K. (2003). Pharmacokinetics of fluoxetine and norfluoxetine in pregnancy and lactation. Clinical Pharmacology and Therapeutics, 73(4), 330-337.
Heim, C., & Nemeroff, C. B. (2001). The role of childhood trauma in the neurobiology of mood and anxiety disorders: Preclinical and clinical studies. Biological Psychiatry, 49(12), 1023-1039.
Hendrick, V., Smith, L. M., Hwang, S., Altshuler, L. L., & Haynes, D. (2003). Weight gain in breastfed infants of mothers taking antidepressant medications. The Journal of Clinical Psychiatry, 64(4), 410-412.
Hendrick, V., Stowe, Z. N., Altshuler, L. L., Hwang, S., Lee, E., & Haynes, D. (2003). Placental passage of antidepressant medications. The American Journal of Psychiatry, 160(5), 993-996.
Hendrick, V., Stowe, Z. N., Altshuler, L. L., Mintz, J., Hwang, S., Hostetter, A., et al. (2001). Fluoxetine and norfluoxetine concentrations in nursing infants and breast milk. Biological Psychiatry, 50(10), 775-782.
Hensler, J. G. (2003). Regulation of 5-HT1A receptor function in brain following agonist or antidepressant administration. Life Sciences, 72(15), 1665-1682.

228
Hiemke, C., & Hartter, S. (2000). Pharmacokinetics of selective serotonin reuptake inhibitors. Pharmacology & Therapeutics, 85(1), 11-28.
Hjorth, S. (1996). (-)-Pindolol, but not buspirone, potentiates the citalopram-induced rise in extracellular 5-hydroxytryptamine. European Journal of Pharmacology, 303(3), 183-186.
Hoffman, S., & Hatch, M. C. (1996). Stress, social support and pregnancy outcome: A reassessment based on recent research. Paediatric and Perinatal Epidemiology, 10(4), 380-405.
Holsboer, F. (1999). The rationale for corticotropin-releasing hormone receptor (CRH-R) antagonists to treat depression and anxiety. Journal of Psychiatric Research, 33(3), 181-214.
Holsboer, F. (2000). The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 23(5), 477-501.
Holsboer, F., & Barden, N. (1996). Antidepressants and hypothalamic-pituitary-adrenocortical regulation. Endocrine Reviews, 17(2), 187-205.
Holsboer, F., Gerken, A., Steiger, A., Benkert, O., Muller, O. A., & Stalla, G. K. (1984). Corticotropin-releasing-factor induced pituitary-adrenal response in depression. Lancet, 1(8367), 55.
Holsboer, F., Lauer, C. J., Schreiber, W., & Krieg, J. C. (1995). Altered hypothalamic-pituitary-adrenocortical regulation in healthy subjects at high familial risk for affective disorders. Neuroendocrinology, 62(4), 340-347.
Hooper, S., & Harding, R. (2001). Respiratory system. In R. Harding, & A. Bocking (Eds.), Fetal growth and development (1st ed., pp. 114-136). United Kingdom: Cambrigde University Press.
Hopkins, J., Marcus, M., & Campbell, S. B. (1984). Postpartum depression: A critical review. Psychological Bulletin, 95(3), 498-515.
Hosenbocus, S., & Chahal, R. (2011). SSRIs and SNRIs: A review of the discontinuation syndrome in children and adolescents. Journal of the Canadian Academy of Child and Adolescent Psychiatry = Journal De l'Academie Canadienne De Psychiatrie De l'Enfant Et De l'Adolescent, 20(1), 60-67.
Houle, C., & Wilkins, R. Health status of children. statistics canada: Canada's national statistical agency. Retrieved 3/7/2010, 2010, from http://www.statcan.gc.ca/start-debut-eng.html

229
Ingemarsson, I. (2003). Gender aspects of preterm birth. BJOG : An International Journal of Obstetrics and Gynaecology, 110 Suppl 20, 34-38.
Ingemarsson, I., Herbst, A., & Thorngren-Jerneck, K. (1997). Long term outcome after umbilical artery acidaemia at term birth: Influence of gender and duration of fetal heart rate abnormalities. British Journal of Obstetrics and Gynaecology, 104(10), 1123-1127.
Isbister, G. K., Bowe, S. J., Dawson, A., & Whyte, I. M. (2004). Relative toxicity of selective serotonin reuptake inhibitors (SSRIs) in overdose. Journal of Toxicology.Clinical Toxicology, 42(3), 277-285.
Isbister, G. K., Dawson, A., Whyte, I. M., Prior, F. H., Clancy, C., & Smith, A. J. (2001). Neonatal paroxetine withdrawal syndrome or actually serotonin syndrome? Archives of Disease in Childhood.Fetal and Neonatal Edition, 85(2), F147-8.
Ishak, N., Hanita, T., Sozo, F., Maritz, G., Harding, R., & De Matteo, R. (2012). Sex differences in cardiorespiratory transition and surfactant composition following preterm birth in sheep. American Journal of Physiology.Regulatory, Integrative and Comparative Physiology, 303(7), R778-89.
Jacobs, B. L., & Azmitia, E. C. (1992). Structure and function of the brain serotonin system. Physiological Reviews, 72(1), 165-229.
Jaiswal, S., Coombs, R. C., & Isbister, G. K. (2003). Paroxetine withdrawal in a neonate with historical and laboratory confirmation. European Journal of Pediatrics, 162(10), 723-724.
Janowsky, D. S., el-Yousef, M. K., Davis, J. M., & Sekerke, H. J. (1972). A cholinergic-adrenergic hypothesis of mania and depression. Lancet, 2(7778), 632-635.
Jeppesen, U., Gram, L. F., Vistisen, K., Loft, S., Poulsen, H. E., & Brosen, K. (1996). Dose-dependent inhibition of CYP1A2, CYP2C19 and CYP2D6 by citalopram, fluoxetine, fluvoxamine and paroxetine. European Journal of Clinical Pharmacology, 51(1), 73-78.
Jessel, N., & Stiskal, J. A. (2004). Neonatal encephalopathy after antidepressant exposure during pregnancy. herbst F, gortner L. paroxetine withdrawal syndrome as differential diagnosis of acute neonatal encephalophathy? Z geburtsh neonatol 2003; 207: 232-234. Z Geburtshilfe Neonatol, 208(2), 75-6; author reply 76.
Jones, B. J., & Blackburn, T. P. (2002). The medical benefit of 5-HT research. Pharmacology Biochemistry and Behavior, 71(4), 555-568.
Jones, S. A., Langille, B. L., Frise, S., & Adamson, S. L. (1993). Nonadrenergic noncholinergic autonomic mediation of pressor response to feeding in lambs. The American Journal of Physiology, 265(3 Pt 2), R530-6.

230
Jonnakuty, C., & Gragnoli, C. (2008). What do we know about serotonin? Journal of Cellular Physiology, 217(2), 301-306.
Jureidini, J. N., Doecke, C. J., Mansfield, P. R., Haby, M. M., Menkes, D. B., & Tonkin, A. L. (2004). Efficacy and safety of antidepressants for children and adolescents. BMJ (Clinical Research Ed.), 328(7444), 879-883.
Kallen, B. (2004). Neonate characteristics after maternal use of antidepressants in late pregnancy. Arch Pediatr Adolesc Med, 158(4), 312-316.
Kallen, B., & Olausson, P. O. (2008). Maternal use of selective serotonin re-uptake inhibitors and persistent pulmonary hypertension of the newborn. Pharmacoepidemiology and Drug Safety, 17(8), 801-806.
Kalsner, S. (2000). The question of feedback at the somadendritic region and antidepressant drug action. Brain Research Bulletin, 52(6), 467-473.
Kalsner, S., & Abdali, S. A. (2002). Rate-independent inhibition of 5-HT release by 5-HT in the somadendritic regions of raphe neurons. Brain Research Bulletin, 58(3), 289-294.
Kandel, D. B., & Davies, M. (1986). Adult sequelae of adolescent depressive symptoms. Archives of General Psychiatry, 43(3), 255-262.
Kaplan & sadock's comprehensive textbook of psychiatry(2005). In Sadock V. A. (Ed.), . Philadelphia: Philadelphia : Lippincott Williams & Wilkins.
Kaumann, A. J., & Levy, F. O. (2006). 5-hydroxytryptamine receptors in the human cardiovascular system. Pharmacology & Therapeutics, 111(3), 674-706.
Kemp, A. H., Quintana, D. S., Gray, M. A., Felmingham, K. L., Brown, K., & Gatt, J. M. (2010). Impact of depression and antidepressant treatment on heart rate variability: A review and meta-analysis. Biological Psychiatry, 67(11), 1067-1074.
Kendler, K. S., & Karkowski-Shuman, L. (1997). Stressful life events and genetic liability to major depression: Genetic control of exposure to the environment? Psychological Medicine, 27(3), 539-547.
Kessler, R. C., Berglund, P., Demler, O., Jin, R., Koretz, D., Merikangas, K. R., et al. (2003). The epidemiology of major depressive disorder: Results from the national comorbidity survey replication (NCS-R). JAMA : The Journal of the American Medical Association, 289(23), 3095-3105.
Kessler, R. C., McGonagle, K. A., Swartz, M., Blazer, D. G., & Nelson, C. B. (1993). Sex and depression in the national comorbidity survey I: Lifetime prevalence, chronicity and recurrence. Journal of Affective Disorders, 29(2-3), 85-96.

231
Khan, A., Brodhead, A. E., Schwartz, K. A., Kolts, R. L., & Brown, W. A. (2005). Sex differences in antidepressant response in recent antidepressant clinical trials. Journal of Clinical Psychopharmacology, 25(4), 318-324.
Khaykin, Y., Dorian, P., Baker, B., Shapiro, C., Sandor, P., Mironov, D., et al. (1998). Autonomic correlates of antidepressant treatment using heart-rate variability analysis. Canadian Journal of Psychiatry.Revue Canadienne De Psychiatrie, 43(2), 183-186.
Khoury, M. J., Marks, J. S., McCarthy, B. J., & Zaro, S. M. (1985). Factors affecting the sex differential in neonatal mortality: The role of respiratory distress syndrome. American Journal of Obstetrics and Gynecology, 151(6), 777-782.
Kim, J. (2000). Pharmacokinetics and pharmacodynamics of the selective serotonin reuptake inhibitors, fluoxetine and paroxetine, during pregnancy and the nursing period. (Doctor of Philosophy, University of British Columbia).
Kim, J., Riggs, K. W., Misri, S., Kent, N., Oberlander, T. F., Grunau, R. E., et al. (2006). Stereoselective disposition of fluoxetine and norfluoxetine during pregnancy and breast-feeding. British Journal of Clinical Pharmacology, 61(2), 155-163.
Kim, J., Riggs, K. W., & Rurak, D. W. (2004). Stereoselective pharmacokinetics of fluoxetine and norfluoxetine enantiomers in pregnant sheep. Drug Metabolism and Disposition: The Biological Fate of Chemicals, 32(2), 212-221.
Kinney, H. C. (2009). Brainstem mechanisms underlying the sudden infant death syndrome: Evidence from human pathologic studies. Developmental Psychobiology, 51(3), 223-233.
Kinney, H. C., Richerson, G. B., Dymecki, S. M., Darnall, R. A., & Nattie, E. E. (2009). The brainstem and serotonin in the sudden infant death syndrome. Annual Review of Pathology, 4, 517-550.
Knoppert, D. C., Nimkar, R., Principi, T., & Yuen, D. (2006). Paroxetine toxicity in a newborn after in utero exposure: Clinical symptoms correlate with serum levels. Ther Drug Monit, 28(1), 5-7.
Kopin, I. J., Pare, C. M., Axelrod, J., & Weissbach, H. (1961). The fate of melatonin in animals. The Journal of Biological Chemistry, 236, 3072-3075.
Koren, G. (2004). Discontinuation syndrome following late pregnancy exposure to antidepressants. Archives of Pediatrics & Adolescent Medicine, 158(4), 307-308.
Koren, G., Finkelstein, Y., Matsui, D., & Berkovich, M. (2009). Diagnosis and management of poor neonatal adaptation syndrome in newborns exposed in utero to selective seretonin/norepinephrine reuptake inhibitors. Journal of Obstetrics and Gynaecology Canada : JOGC = Journal d'Obstetrique Et Gynecologie Du Canada : JOGC, 31(4), 348-350.

232
Kosten, T. R., & O'Connor, P. G. (2003). Management of drug and alcohol withdrawal. The New England Journal of Medicine, 348(18), 1786-1795.
Krogh CME. (1995). Compendium of pharmaceuticals and specialties, (30th ed.). Ottawa, Ontario: Canadian Pharmaceutical Association.
Kulin, N. A., Pastuszak, A., Sage, S. R., Schick-Boschetto, B., Spivey, G., Feldkamp, M., et al. (1998). Pregnancy outcome following maternal use of the new selective serotonin reuptake inhibitors: A prospective controlled multicenter study. JAMA : The Journal of the American Medical Association, 279(8), 609-610.
Kurstjens, S., & Wolke, D. (2001). Effects of maternal depression on cognitive development of children over the first 7 years of life. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 42(5), 623-636.
Kwon, P., & Lefkowitz, W. (2008). Poor neonatal adaptation in term infant. fluoxetine toxicity. Pediatric Annals, 37(3), 131-133.
Laine, K., Heikkinen, T., Ekblad, U., & Kero, P. (2003). Effects of exposure to selective serotonin reuptake inhibitors during pregnancy on serotonergic symptoms in newborns and cord blood monoamine and prolactin concentrations. Archives of General Psychiatry, 60(7), 720-726.
Lake, C. R., Pickar, D., Ziegler, M. G., Lipper, S., Slater, S., & Murphy, D. L. (1982). High plasma norepinephrine levels in patients with major affective disorder. The American Journal of Psychiatry, 139(10), 1315-1318.
Lam, R. W., Zis, A. P., Grewal, A., Delgado, P. L., Charney, D. S., & Krystal, J. H. (1996). Effects of rapid tryptophan depletion in patients with seasonal affective disorder in remission after light therapy. Archives of General Psychiatry, 53(1), 41-44.
Lane, M. A., Baldwin, R. L.,4th, & Jesse, B. W. (2002). Developmental changes in ketogenic enzyme gene expression during sheep rumen development. Journal of Animal Science, 80(6), 1538-1544.
Lane, M. A., Baldwin, R. L., & Jesse, B. W. (2000). Sheep rumen metabolic development in response to age and dietary treatments. Journal of Animal Science, 78(7), 1990-1996.
Lane, R., Baldwin, D., & Preskorn, S. (1995). The SSRIs: Advantages, disadvantages and differences. Journal of Psychopharmacology (Oxford, England), 9(2 Suppl), 163-178.
Lang, C., Field, T., Pickens, J., Martinez, A., Bendell, D., Yando, R., et al. (1996). Preschoolers of dysphoric mothers. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 37(2), 221-224.

233
Lang, U., Baker, R. S., Khoury, J., & Clark, K. E. (2000). Effects of chronic reduction in uterine blood flow on fetal and placental growth in the sheep. American Journal of Physiology.Regulatory, Integrative and Comparative Physiology, 279(1), R53-9.
Lattimore, K. A., Donn, S. M., Kaciroti, N., Kemper, A. R., Neal, C. R.,Jr, & Vazquez, D. M. (2005). Selective serotonin reuptake inhibitor (SSRI) use during pregnancy and effects on the fetus and newborn: A meta-analysis. Journal of Perinatology : Official Journal of the California Perinatal Association, 25(9), 595-604.
Lavker, R., Chalupa, W., & Opliger, P. (1969). Histochemical observations on rumen mucosa structure and composition. Journal of Dairy Science, 52(2), 266-269.
Lawton, C. L., Wales, J. K., Hill, A. J., & Blundell, J. E. (1995). Serotoninergic manipulation, meal-induced satiety and eating pattern: Effect of fluoxetine in obese female subjects. Obesity Research, 3(4), 345-356.
Lazartigues, E., Brefel-Courbon, C., Bagheri, H., Costes, S., Gharib, C., Tran, M. A., et al. (2000). Fluoxetine-induced pressor response in freely moving rats: A role for vasopressin and sympathetic tone. Fundamental & Clinical Pharmacology, 14(5), 443-451.
Lee, L. J. (2009). Neonatal fluoxetine exposure affects the neuronal structure in the somatosensory cortex and somatosensory-related behaviors in adolescent rats. Neurotoxicity Research, 15(3), 212-223.
Lesch, K. P., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., et al. (1996). Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science (New York, N.Y.), 274(5292), 1527-1531.
Levine, R. E., Oandasan, A. P., Primeau, L. A., & Berenson, A. B. (2003). Anxiety disorders during pregnancy and postpartum. American Journal of Perinatology, 20(5), 239-248.
Levinson-Castiel, R., Merlob, P., Linder, N., Sirota, L., & Klinger, G. (2006). Neonatal abstinence syndrome after in utero exposure to selective serotonin reuptake inhibitors in term infants. Archives of Pediatrics & Adolescent Medicine, 160(2), 173-176.
Lewinsohn, P. M., Clarke, G. N., Seeley, J. R., & Rohde, P. (1994). Major depression in community adolescents: Age at onset, episode duration, and time to recurrence. Journal of the American Academy of Child and Adolescent Psychiatry, 33(6), 809-818.
Lewinsohn, P. M., Rohde, P., Klein, D. N., & Seeley, J. R. (1999). Natural course of adolescent major depressive disorder: I. continuity into young adulthood. Journal of the American Academy of Child and Adolescent Psychiatry, 38(1), 56-63.
Lewis, A. J., Galbally, M., Opie, G., & Buist, A. (2010). Neonatal growth outcomes at birth and one month postpartum following in utero exposure to antidepressant medication. The Australian and New Zealand Journal of Psychiatry, 44(5), 482-487.

234
Licht, C. M., de Geus, E. J., van Dyck, R., & Penninx, B. W. (2010). Longitudinal evidence for unfavorable effects of antidepressants on heart rate variability. Biological Psychiatry, 68(9), 861-868.
Licht, C. M., de Geus, E. J., Zitman, F. G., Hoogendijk, W. J., van Dyck, R., & Penninx, B. W. (2008). Association between major depressive disorder and heart rate variability in the netherlands study of depression and anxiety (NESDA). Archives of General Psychiatry, 65(12), 1358-1367.
Liggins, G. (1968). Premature parturition after infusion of corticotrophin or cortisol into foetal lambs. J Endocr, 42, 323-329.
Liggins, G., Holm, L., & Kennedy, P. (1966). Prolonged pregnancy following surgical lesions of the foetal lamb pituitary. J Reprod Fertil, 12, 419-422.
Lim, K., Sanders, A., Brain, U., Riggs, W., Oberlander, T. F., & Rurak, D. (2012). Third trimester fetal pulmonary artery doppler blood flow velocity characteristics following prenatal selective serotonin reuptake inhibitor (SSRI) exposure. Early Human Development,
Linkowski, P. (2003). Neuroendocrine profiles in mood disorders. The International Journal of Neuropsychopharmacology / Official Scientific Journal of the Collegium Internationale Neuropsychopharmacologicum (CINP), 6(2), 191-197.
Lisboa, S. F., Oliveira, P. E., Costa, L. C., Venancio, E. J., & Moreira, E. G. (2007). Behavioral evaluation of male and female mice pups exposed to fluoxetine during pregnancy and lactation. Pharmacology, 80(1), 49-56.
Lister, G., Moreau, G., Moss, M., & Talner, N. S. (1984). Effects of alterations of oxygen transport on the neonate. Seminars in Perinatology, 8(3), 192-204.
Lister, G., Walter, T. K., Versmold, H. T., Dallman, P. R., & Rudolph, A. M. (1979). Oxygen delivery in lambs: Cardiovascular and hematologic development. The American Journal of Physiology, 237(6), H668-75.
Lu, X. Y. (2007). The leptin hypothesis of depression: A potential link between mood disorders and obesity? Current Opinion in Pharmacology, 7(6), 648-652.
Lu, X. Y., Kim, C. S., Frazer, A., & Zhang, W. (2006). Leptin: A potential novel antidepressant. Proceedings of the National Academy of Sciences of the United States of America, 103(5), 1593-1598.
Lucki, I. (1998). The spectrum of behaviors influenced by serotonin. Biological Psychiatry, 44(3), 151-162.

235
Lund, N., Pedersen, L. H., & Henriksen, T. B. (2009). Selective serotonin reuptake inhibitor exposure in utero and pregnancy outcomes. Arch Pediatr Adolesc Med, 163(10), 949-954.
Lupien, S. J., McEwen, B. S., Gunnar, M. R., & Heim, C. (2009). Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nature Reviews.Neuroscience, 10(6), 434-445.
Maciag, D., Simpson, K. L., Coppinger, D., Lu, Y., Wang, Y., Lin, R. C., et al. (2006). Neonatal antidepressant exposure has lasting effects on behavior and serotonin circuitry. Neuropsychopharmacology, 31(1), 47-57.
Malm, H., Klaukka, T., & Neuvonen, P. J. (2005). Risks associated with selective serotonin reuptake inhibitors in pregnancy. Obstetrics and Gynecology, 106(6), 1289-1296.
Manhaes de Castro, R., Barreto Medeiros, J. M., Mendes da Silva, C., Ferreira, L. M., Guedes, R. C., Cabral Filho, J. E., et al. (2001). Reduction of intraspecific aggression in adult rats by neonatal treatment with a selective serotonin reuptake inhibitor. Braz J Med Biol Res, 34(1), 121-124.
Mann, J. J., Brent, D. A., & Arango, V. (2001). The neurobiology and genetics of suicide and attempted suicide: A focus on the serotonergic system. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 24(5), 467-477.
March, J., Silva, S., Petrycki, S., Curry, J., Wells, K., Fairbank, J., et al. (2004). Fluoxetine, cognitive-behavioral therapy, and their combination for adolescents with depression: Treatment for adolescents with depression study (TADS) randomized controlled trial. JAMA : The Journal of the American Medical Association, 292(7), 807-820.
March, J. S., Silva, S., Petrycki, S., Curry, J., Wells, K., Fairbank, J., et al. (2007). The treatment for adolescents with depression study (TADS): Long-term effectiveness and safety outcomes. Archives of General Psychiatry, 64(10), 1132-1143.
Masand, P. S., & Gupta, S. (1999). Selective serotonin-reuptake inhibitors: An update. Harvard Review of Psychiatry, 7(2), 69-84.
Maschi, S., Clavenna, A., Campi, R., Schiavetti, B., Bernat, M., & Bonati, M. (2008). Neonatal outcome following pregnancy exposure to antidepressants: A prospective controlled cohort study. BJOG : An International Journal of Obstetrics and Gynaecology, 115(2), 283-289.
Maso, G., Businelli, C., Piccoli, M., Montico, M., De Seta, F., Sartore, A., et al. (2012). The clinical interpretation and significance of electronic fetal heart rate patterns 2 h before delivery: An institutional observational study. Archives of Gynecology and Obstetrics,

236
Mastorakos, G., & Ilias, I. (2003). Maternal and fetal hypothalamic-pituitary-adrenal axes during pregnancy and postpartum. Annals of the New York Academy of Sciences, 997, 136-149.
Matthews, S. G., Owen, D., Banjanin, S., & Andrews, M. H. (2002). Glucocorticoids, hypothalamo-pituitary-adrenal (HPA) development, and life after birth. Endocrine Research, 28(4), 709-718.
Maurer-Spurej, E., Pittendreigh, C., & Solomons, K. (2004). The influence of selective serotonin reuptake inhibitors on human platelet serotonin. Thrombosis and Haemostasis, 91(1), 119-128.
McAnally, L. E., Threlkeld, K. R., & Dreyling, C. A. (1992). Case report of a syncopal episode associated with fluoxetine. The Annals of Pharmacotherapy, 26(9), 1090-1091.
McCormick, M. C., Brooks-Gunn, J., Shorter, T., Holmes, J. H., Wallace, C. Y., & Heagarty, M. C. (1990). Factors associated with smoking in low-income pregnant women: Relationship to birth weight, stressful life events, social support, health behaviors and mental distress. Journal of Clinical Epidemiology, 43(5), 441-448.
McCoy, S. J., Beal, J. M., & Watson, G. H. (2003). Endocrine factors and postpartum depression. A selected review. The Journal of Reproductive Medicine, 48(6), 402-408.
McMillen, I. C., & Nowak, R. (1989). Maternal pinealectomy abolishes the diurnal rhythm in plasma melatonin concentrations in the fetal sheep and pregnant ewe during late gestation. The Journal of Endocrinology, 120(3), 459-464.
Mellor, D. J. (1988). Integration of perinatal events, pathophysiological changes and consequences for the newborn lamb. The British Veterinary Journal, 144(6), 552-569.
Mellor, D. J., & Gregory, N. G. (2003). Responsiveness, behavioural arousal and awareness in fetal and newborn lambs: Experimental, practical and therapeutic implications. New Zealand Veterinary Journal, 51(1), 2-13.
Mellor, D. J., Matheson, I. C., & Small, J. (1977). Changes in the corticosteroid concentrations of plasma from single and twin fetuses during the last 3 weeks of pregnancy in sheep. Journal of Reproduction and Fertility, 50(2), 383-385.
Mellor, D. J., & Pearson, R. A. (1977). Some changes in the composition of blood during the first 24 hours after birth in normal and growth retarded lambs. Annales De Recherches Veterinaires.Annals of Veterinary Research, 8(4), 460-467.
Merritt, K., & Davidson, L. (1933). The blood during the first year of life. I. normal values for erythrocytes, hemoglobin, reticulocytes and platelets, and their relationship to neonatal bleeding and coagulation time. Arch Pediatr Adolesc Med, 46(5), 990.

237
Mhanna, M. J., Bennet, J. B.,2nd, & Izatt, S. D. (1997). Potential fluoxetine chloride (prozac) toxicity in a newborn. Pediatrics, 100(1), 158-159.
Minoura, S., & Gilbert, R. D. (1987). Postnatal change of cardiac function in lambs: Effects of ganglionic block and afterload. Journal of Developmental Physiology, 9(2), 123-135.
Mirmiran, M., van de Poll, N. E., Corner, M. A., van Oyen, H. G., & Bour, H. L. (1981). Suppression of active sleep by chronic treatment with chlorimipramine during early postnatal development: Effects upon adult sleep and behavior in the rat. Brain Res, 204(1), 129-146.
Misri, S., Kostaras, D., & Kostaras, X. (2000). The use of selective serotonin reuptake inhibitors during pregnancy and lactation: Current knowledge. Canadian Journal of Psychiatry.Revue Canadienne De Psychiatrie, 45(3), 285-287.
Mood disorders. (2000). In American Psychiatric Association (Ed.), Quick reference to the diagnostic criteria from DSM-IV-TR (pp. 168-169). Washington, D.C.: Washington, D.C. : American Psychiatric Association.
Morrison, J. L. (2001). Physiologic responses of the fetal lamb to eight day fluoxetine exposure during late gestation. (Doctor of Philosophy, University of British Columbia).
Morrison, J. L., Chien, C., Gruber, N., Rurak, D., & Riggs, W. (2001). Fetal behavioural state changes following maternal fluoxetine infusion in sheep. Brain Research.Developmental Brain Research, 131(1-2), 47-56.
Morrison, J. L., Chien, C., Riggs, K. W., Gruber, N., & Rurak, D. (2002). Effect of maternal fluoxetine administration on uterine blood flow, fetal blood gas status, and growth. Pediatric Research, 51(4), 433-442.
Morrison, J. L., Riggs, K. W., Chien, C., Gruber, N., McMillen, I. C., & Rurak, D. W. (2004). Chronic maternal fluoxetine infusion in pregnant sheep: Effects on the maternal and fetal hypothalamic-pituitary-adrenal axes. Pediatric Research, 56(1), 40-46.
Morrison, J. L., Riggs, K. W., & Rurak, D. W. (2005). Fluoxetine during pregnancy: Impact on fetal development. Reprod Fertil Dev, 17(6), 641-650.
Morrison, J. L., Rurak, D. W., Chien, C., Kennaway, D. J., Gruber, N., McMillen, I. C., et al. (2005). Maternal fluoxetine infusion does not alter fetal endocrine and biophysical circadian rhythms in pregnant sheep. Journal of the Society for Gynecologic Investigation, 12(5), 356-364.
Morrison, M., Pope, P. B., Denman, S. E., & McSweeney, C. S. (2009). Plant biomass degradation by gut microbiomes: More of the same or something new? Current Opinion in Biotechnology, 20(3), 358-363.

238
Mortensen, J. T., Olsen, J., Larsen, H., Bendsen, J., Obel, C., & Sorensen, H. T. (2003). Psychomotor development in children exposed in utero to benzodiazepines, antidepressants, neuroleptics, and anti-epileptics. European Journal of Epidemiology, 18(8), 769-771.
Moses-Kolko, E. L., Bogen, D., Perel, J., Bregar, A., Uhl, K., Levin, B., et al. (2005). Neonatal signs after late in utero exposure to serotonin reuptake inhibitors: Literature review and implications for clinical applications. JAMA : The Journal of the American Medical Association, 293(19), 2372-2383.
Moss, T. J., Jakubowska, A. E., McCrabb, G. J., Billings, K., & Harding, R. (1995). Ventilatory responses to progressive hypoxia and hypercapnia in developing sheep. Respiration Physiology, 100(1), 33-44.
Mulder, E. J., Ververs, F. F., de Heus, R., & Visser, G. H. (2011). Selective serotonin reuptake inhibitors affect neurobehavioral development in the human fetus. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 36(10), 1961-1971.
Mulkey, D. K., Stornetta, R. L., Weston, M. C., Simmons, J. R., Parker, A., Bayliss, D. A., et al. (2004). Respiratory control by ventral surface chemoreceptor neurons in rats. Nature Neuroscience, 7(12), 1360-1369.
Murray L, C. W. (1993). The role of obstetric factors in postpartum depression. J Reprod Infant Psychol, 11, 215-219.
Murray, C. J., & Lopez, A. D. (1996). Evidence-based health policy--lessons from the global burden of disease study. Science (New York, N.Y.), 274(5288), 740-743.
Murray, L. (1992). The impact of postnatal depression on infant development. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 33(3), 543-561.
Murray, L., & Cooper, P. J. (1997). Postpartum depression and child development. Psychological Medicine, 27(2), 253-260.
Murray, L., Stanley, C., Hooper, R., King, F., & Fiori-Cowley, A. (1996). The role of infant factors in postnatal depression and mother-infant interactions. Developmental Medicine and Child Neurology, 38(2), 109-119.
Murray, L., Woolgar, M., Cooper, P., & Hipwell, A. (2001). Cognitive vulnerability to depression in 5-year-old children of depressed mothers. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 42(7), 891-899.
Myint, A. M., Kim, Y. K., Verkerk, R., Scharpe, S., Steinbusch, H., & Leonard, B. (2007). Kynurenine pathway in major depression: Evidence of impaired neuroprotection. Journal of Affective Disorders, 98(1-2), 143-151.

239
Nakamura, Y., Yamamoto, Y., & Muraoka, I. (1993). Autonomic control of heart rate during physical exercise and fractal dimension of heart rate variability. Journal of Applied Physiology (Bethesda, Md.: 1985), 74(2), 875-881.
Nelson, L. S., Erdman, A. R., Booze, L. L., Cobaugh, D. J., Chyka, P. A., Woolf, A. D., et al. (2007). Selective serotonin reuptake inhibitor poisoning: An evidence-based consensus guideline for out-of-hospital management. Clinical Toxicology (Philadelphia, Pa.), 45(4), 315-332.
Nelson, M., Coghlan, J. P., Denton, D. A., Tresham, J. J., Whitworth, J. A., & Scoggins, B. A. (1987). Ritanserin and serotonergic mechanisms in blood pressure and fluid regulation in sheep. Clinical and Experimental Pharmacology & Physiology, 14(7), 555-563.
Nilsson, T., Longmore, J., Shaw, D., Pantev, E., Bard, J. A., Branchek, T., et al. (1999). Characterisation of 5-HT receptors in human coronary arteries by molecular and pharmacological techniques. European Journal of Pharmacology, 372(1), 49-56.
Nishizawa, S., Benkelfat, C., Young, S. N., Leyton, M., Mzengeza, S., de Montigny, C., et al. (1997). Differences between males and females in rates of serotonin synthesis in human brain. Proceedings of the National Academy of Sciences of the United States of America, 94(10), 5308-5313.
Nolen-Hoeksema, S., & Girgus, J. S. (1994). The emergence of gender differences in depression during adolescence. Psychological Bulletin, 115(3), 424-443.
Nonacs, R., & Cohen, L. S. (2003). Assessment and treatment of depression during pregnancy: An update. The Psychiatric Clinics of North America, 26(3), 547-562.
Norman, L. J., Lye, S. J., Wlodek, M. E., & Challis, J. R. (1985). Changes in pituitary responses to synthetic ovine corticotrophin releasing factor in fetal sheep. Canadian Journal of Physiology and Pharmacology, 63(11), 1398-1403.
Nulman, I., Rovet, J., Stewart, D. E., Wolpin, J., Gardner, H. A., Theis, J. G., et al. (1997). Neurodevelopment of children exposed in utero to antidepressant drugs. N Engl J Med, 336(4), 258-262.
Nulman, I., Rovet, J., Stewart, D. E., Wolpin, J., Pace-Asciak, P., Shuhaiber, S., et al. (2002). Child development following exposure to tricyclic antidepressants or fluoxetine throughout fetal life: A prospective, controlled study. The American Journal of Psychiatry, 159(11), 1889-1895.
Nutt, D. J., Baldwin, D. S., Clayton, A. H., Elgie, R., Lecrubier, Y., Montejo, A. L., et al. (2006). Consensus statement and research needs: The role of dopamine and norepinephrine in depression and antidepressant treatment. The Journal of Clinical Psychiatry, 67 Suppl 6, 46-49.

240
Oberlander, T. F. (2012). Fetal serotonin signaling: Setting pathways for early childhood development and behavior. The Journal of Adolescent Health : Official Publication of the Society for Adolescent Medicine, 51(2 Suppl), S9-16.
Oberlander, T. F., Bonaguro, R. J., Misri, S., Papsdorf, M., Ross, C. J., & Simpson, E. M. (2008). Infant serotonin transporter (SLC6A4) promoter genotype is associated with adverse neonatal outcomes after prenatal exposure to serotonin reuptake inhibitor medications. Mol Psychiatry, 13(1), 65-73.
Oberlander, T. F., Eckstein Grunau, R., Fitzgerald, C., Ellwood, A. L., Misri, S., Rurak, D., et al. (2002). Prolonged prenatal psychotropic medication exposure alters neonatal acute pain response. Pediatr Res, 51(4), 443-453.
Oberlander, T. F., Gingrich, J. A., & Ansorge, M. S. (2009). Sustained neurobehavioral effects of exposure to SSRI antidepressants during development: Molecular to clinical evidence. Clinical Pharmacology and Therapeutics, 86(6), 672-677.
Oberlander, T. F., Grunau, R., Mayes, L., Riggs, W., Rurak, D., Papsdorf, M., et al. (2008). Hypothalamic-pituitary-adrenal (HPA) axis function in 3-month old infants with prenatal selective serotonin reuptake inhibitor (SSRI) antidepressant exposure. Early Human Development, 84(10), 689-697.
Oberlander, T. F., Grunau, R. E., Fitzgerald, C., Papsdorf, M., Rurak, D., & Riggs, W. (2005). Pain reactivity in 2-month-old infants after prenatal and postnatal serotonin reuptake inhibitor medication exposure. Pediatrics, 115(2), 411-425.
Oberlander, T. F., Misri, S., Fitzgerald, C. E., Kostaras, X., Rurak, D., & Riggs, W. (2004). Pharmacologic factors associated with transient neonatal symptoms following prenatal psychotropic medication exposure. The Journal of Clinical Psychiatry, 65(2), 230-237.
Oberlander, T. F., Reebye, P., Misri, S., Papsdorf, M., Kim, J., & Grunau, R. E. (2007). Externalizing and attentional behaviors in children of depressed mothers treated with a selective serotonin reuptake inhibitor antidepressant during pregnancy. Archives of Pediatrics & Adolescent Medicine, 161(1), 22-29.
Oberlander, T. F., Warburton, W., Misri, S., Aghajanian, J., & Hertzman, C. (2006). Neonatal outcomes after prenatal exposure to selective serotonin reuptake inhibitor antidepressants and maternal depression using population-based linked health data. Archives of General Psychiatry, 63(8), 898-906.
Oberlander, T. F., Warburton, W., Misri, S., Aghajanian, J., & Hertzman, C. (2008). Effects of timing and duration of gestational exposure to serotonin reuptake inhibitor antidepressants: Population-based study. Br J Psychiatry, 192(5), 338-343.

241
Ogilvie, A. D., Battersby, S., Bubb, V. J., Fink, G., Harmar, A. J., Goodwim, G. M., et al. (1996). Polymorphism in serotonin transporter gene associated with susceptibility to major depression. Lancet, 347(9003), 731-733.
Ogilvie, A. D., & Harmar, A. J. (1997). Association between the serotonin transporter gene and affective disorder: The evidence so far. Molecular Medicine (Cambridge, Mass.), 3(2), 90-93.
O'Hara, M. (1997). The nature of postpartum depressive disorders. In L. Murray, & P. Cooper (Eds.), Postpartum depression and child development (pp. 3-31). New York: The GUilford Press.
O'Hara, M. W., Zekoski, E. M., Philipps, L. H., & Wright, E. J. (1990). Controlled prospective study of postpartum mood disorders: Comparison of childbearing and nonchildbearing women. Journal of Abnormal Psychology, 99(1), 3-15.
Orr, S. T., James, S. A., & Blackmore Prince, C. (2002). Maternal prenatal depressive symptoms and spontaneous preterm births among african-american women in baltimore, maryland. American Journal of Epidemiology, 156(9), 797-802.
Orr, S. T., & Miller, C. A. (1995). Maternal depressive symptoms and the risk of poor pregnancy outcome. review of the literature and preliminary findings. Epidemiologic Reviews, 17(1), 165-171.
Ortiz, J., & Artigas, F. (1992). Effects of monoamine uptake inhibitors on extracellular and platelet 5-hydroxytryptamine in rat blood: Different effects of clomipramine and fluoxetine. British Journal of Pharmacology, 105(4), 941-946.
Owens, M. J., & Nemeroff, C. B. (1994). Role of serotonin in the pathophysiology of depression: Focus on the serotonin transporter. Clinical Chemistry, 40(2), 288-295.
Pampallona, S., Bollini, P., Tibaldi, G., Kupelnick, B., & Munizza, C. (2004). Combined pharmacotherapy and psychological treatment for depression: A systematic review. Archives of General Psychiatry, 61(7), 714-719.
Parraguez, V. H., Vergara, M., Riquelme, R., Raimann, R., Llanos, A. J., & Seron-Ferre, M. (1989). Ontogeny of the circadian rhythm of cortisol in sheep. Biology of Reproduction, 40(6), 1137-1143.
Pawluski, J. L., Galea, L. A., Brain, U., Papsdorf, M., & Oberlander, T. F. (2009). Neonatal S100B protein levels after prenatal exposure to selective serotonin reuptake inhibitors. Pediatrics, 124(4), e662-70.
Peacock, J. L., Bland, J. M., & Anderson, H. R. (1995). Preterm delivery: Effects of socioeconomic factors, psychological stress, smoking, alcohol, and caffeine. BMJ (Clinical Research Ed.), 311(7004), 531-535.

242
Pearson, K. H., Nonacs, R. M., Viguera, A. C., Heller, V. L., Petrillo, L. F., Brandes, M., et al. (2007). Birth outcomes following prenatal exposure to antidepressants. The Journal of Clinical Psychiatry, 68(8), 1284-1289.
Pedersen, L. H., Henriksen, T. B., & Olsen, J. (2010). Fetal exposure to antidepressants and normal milestone development at 6 and 19 months of age. Pediatrics, 125(3), e600-8.
Phillips, S., Brent, J., Kulig, K., Heiligenstein, J., & Birkett, M. (1997). Fluoxetine versus tricyclic antidepressants: A prospective multicenter study of antidepressant drug overdoses. the antidepressant study group. The Journal of Emergency Medicine, 15(4), 439-445.
Pohland, R. C., Byrd, T. K., Hamilton, M., & Koons, J. R. (1989). Placental transfer and fetal distribution of fluoxetine in the rat. Toxicology and Applied Pharmacology, 98(2), 198-205.
Pompili, M., Serafini, G., Innamorati, M., Moller-Leimkuhler, A. M., Giupponi, G., Girardi, P., et al. (2010). The hypothalamic-pituitary-adrenal axis and serotonin abnormalities: A selective overview for the implications of suicide prevention. European Archives of Psychiatry and Clinical Neuroscience, 260(8), 583-600.
Popa, D., Lena, C., Alexandre, C., & Adrien, J. (2008). Lasting syndrome of depression produced by reduction in serotonin uptake during postnatal development: Evidence from sleep, stress, and behavior. J Neurosci, 28(14), 3546-3554.
Porter, R. J., Gallagher, P., Watson, S., & Young, A. H. (2004). Corticosteroid-serotonin interactions in depression: A review of the human evidence. Psychopharmacology, 173(1-2), 1-17.
Price, L. H., Charney, D. S., Delgado, P. L., Goodman, W. K., Krystal, J. H., Woods, S. W., et al. (1990). Clinical data on the role of serotonin in the mechanism(s) of action of antidepressant drugs. The Journal of Clinical Psychiatry, 51 Suppl, 44-50; discussion 51-2.
Public health advisories (drugs) > suicidality in children and adolescents being treated with antidepressant medications Retrieved 11/13/2012, 2012, from http://www.fda.gov/Drugs/DrugSafety/PostmarketDrugSafetyInformationforPatientsandProviders/DrugSafetyInformationforHeathcareProfessionals/PublicHealthAdvisories/ucm161679.htm
Public health advisory: Treatment challenges of depression in pregnancy and possibility of persistent pulmonary hypertension in newborns. (2006). Retrieved 03/2012http://www.fda.gov/Drugs/DrugSafety/PostmarketDrugSafetyInformationforPatientsandProviders/DrugSafetyInformationforHeathcareProfessionals/PublicHealthAdvisories/ucm124348.htm

243
Raap, D. K., & Van de Kar, L. D. (1999). Selective serotonin reuptake inhibitors and neuroendocrine function. Life Sciences, 65(12), 1217-1235.
Rahman, M. K., Nagatsu, T., Sakurai, T., Hori, S., Abe, M., & Matsuda, M. (1982). Effect of pyridoxal phosphate deficiency on aromatic L-amino acid decarboxylase activity with L-DOPA and L-5-hydroxytryptophan as substrates in rats. Japanese Journal of Pharmacology, 32(5), 803-811.
Rampono, J., Proud, S., Hackett, L. P., Kristensen, J. H., & Ilett, K. F. (2004). A pilot study of newer antidepressant concentrations in cord and maternal serum and possible effects in the neonate. Int J Neuropsychopharmacol, 7(3), 329-334.
Rampono, J., Simmer, K., Ilett, K. F., Hackett, L. P., Doherty, D. A., Elliot, R., et al. (2009). Placental transfer of SSRI and SNRI antidepressants and effects on the neonate. Pharmacopsychiatry, 42(3), 95-100.
Rao, U., Hammen, C., & Daley, S. E. (1999). Continuity of depression during the transition to adulthood: A 5-year longitudinal study of young women. Journal of the American Academy of Child and Adolescent Psychiatry, 38(7), 908-915.
Richardson, B. S. (1994). Ontogeny of behavioral states in the fetus. In G. Thorburn, & R. Harding (Eds.), Textbook of fetal physiology (pp. 322-329). New York: Oxford University Press Inc.
Richerson, G. B. (2004). Serotonergic neurons as carbon dioxide sensors that maintain pH homeostasis. Nature Reviews.Neuroscience, 5(6), 449-461.
Riddle, M. A., Brown, N., Dzubinski, D., Jetmalani, A. N., Law, Y., & Woolston, J. L. (1989). Fluoxetine overdose in an adolescent. Journal of the American Academy of Child and Adolescent Psychiatry, 28(4), 587-588.
Ring, B. J., Eckstein, J. A., Gillespie, J. S., Binkley, S. N., VandenBranden, M., & Wrighton, S. A. (2001). Identification of the human cytochromes p450 responsible for in vitro formation of R- and S-norfluoxetine. The Journal of Pharmacology and Experimental Therapeutics, 297(3), 1044-1050.
Roberge, R. J., & Martin, T. G. (1994). Mixed fluoxetine/loxapine overdose and atrial flutter. Annals of Emergency Medicine, 23(3), 586-590.
Roca, A., Garcia-Esteve, L., Imaz, M. L., Torres, A., Hernandez, S., Botet, F., et al. (2011). Obstetrical and neonatal outcomes after prenatal exposure to selective serotonin reuptake inhibitors: The relevance of dose. Journal of Affective Disorders, 135(1-3), 208-215.
Rohde, P., Lewinsohn, P. M., & Seeley, J. R. (1994). Are adolescents changed by an episode of major depression? Journal of the American Academy of Child and Adolescent Psychiatry, 33(9), 1289-1298.

244
Ronsley, R., Elbe, D., Smith, D. H., & Garland, E. J. (2010). Do hospital and community SSRI usage patterns in children and adolescents match the evidence? Journal of the Canadian Academy of Child and Adolescent Psychiatry = Journal De l'Academie Canadienne De Psychiatrie De l'Enfant Et De l'Adolescent, 19(3), 218-226.
Ruchkin, V., & Martin, A. (2005). SSRIs and the developing brain. Lancet, 365(9458), 451-453.
Rurak, D., Lim, K., Sanders, A., Brain, U., Riggs, W., & Oberlander, T. F. (2011). Third trimester fetal heart rate and doppler middle cerebral artery blood flow velocity characteristics during prenatal selective serotonin reuptake inhibitor exposure. Pediatric Research, 70(1), 96-101.
Rurak, D. W. (1995). Anatomy and physiology of the placenta. In J. McDonald, & J. Bonica (Eds.), Principles and practice of obstetric analgesia and anesthesia (pp. 157-194). Baltimore, Md.: Baltimore, Md. : Williams & Wilkins.
Rurak, D. W., Fay, S., & Gruber, N. C. (2008). Measurement of rest and activity in newborn lambs using actigraphy: Studies in term and preterm lambs. Reproduction, Fertility, and Development, 20(3), 418-430.
Rurak, D. W., & Bessette, N. W. (2012). Deterioration in fetal lamb blood gas and acid-base status with advancing gestation [Abstract].
Ryan, K. D., Goodman, R. L., Karsch, F. J., Legan, S. J., & Foster, D. L. (1991). Patterns of circulating gonadotropins and ovarian steroids during the first periovulatory period in the developing sheep. Biology of Reproduction, 45(3), 471-477.
Sachse, C., Brockmoller, J., Bauer, S., & Roots, I. (1997). Cytochrome P450 2D6 variants in a caucasian population: Allele frequencies and phenotypic consequences. American Journal of Human Genetics, 60(2), 284-295.
Sackner, M. A., Will, D. H., & DuBois, A. B. (1966). The site of pulmonary vasomotor activity during hypoxia or serotonin administration. The Journal of Clinical Investigation, 45(1), 112-121.
Salisbury, A. L., DiPietro, J. A., Pearlstein, T., Wisner, K. L., Carr S, & Lester, B. M. (2006). Prenatal serotonin reuptake inhibitor exposure: Fetal and infant outcomes. Dev Psychobiol, 48, 624.
Salisbury, A. L., Ponder, K. L., Padbury, J. F., Carr, S., & Lester, B. M. (2009). Fetal effects of psychoactive drugs. Clinics in Perinatology, 36(3), 595-619.
Sanz, E. J., De-las-Cuevas, C., Kiuru, A., Bate, A., & Edwards, R. (2005). Selective serotonin reuptake inhibitors in pregnant women and neonatal withdrawal syndrome: A database analysis. Lancet, 365(9458), 482-487.

245
Sapolsky, R. M. (2000). Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Archives of General Psychiatry, 57(10), 925-935.
Sauer, W. H., Berlin, J. A., & Kimmel, S. E. (2001). Selective serotonin reuptake inhibitors and myocardial infarction. Circulation, 104(16), 1894-1898.
Savoie, S., Polychronakos, C., Forest, M. G., Haour, F., Collu, R., & Ducharme, J. R. (1981). Perinatal activity of the hypothalamic-pituitary-gonadal axis in the lamb. III. LH, testosterone and prolactin secretory pattern in newborn lambs. Hormone Research, 15(3), 167-178.
Schildkraut, J. J. (1965). The catecholamine hypothesis of affective disorders: A review of supporting evidence. The American Journal of Psychiatry, 122(5), 509-522.
Scordo, M. G., Spina, E., Dahl, M. L., Gatti, G., & Perucca, E. (2005). Influence of CYP2C9, 2C19 and 2D6 genetic polymorphisms on the steady-state plasma concentrations of the enantiomers of fluoxetine and norfluoxetine. Basic & Clinical Pharmacology & Toxicology, 97(5), 296-301.
Seckl, J. R. (1997). Glucocorticoids, feto-placental 11 beta-hydroxysteroid dehydrogenase type 2, and the early life origins of adult disease. Steroids, 62(1), 89-94.
Sharp, S. C., & Hellings, J. A. (2006). Efficacy and safety of selective serotonin reuptake inhibitors in the treatment of depression in children and adolescents: Practitioner review. Clinical Drug Investigation, 26(5), 247-255.
Sidi, D., Kuipers, J. R., Heymann, M. A., & Rudolph, A. M. (1983). Effects of ambient temperature on oxygen consumption and the circulation in newborn lambs at rest and during hypoxemia. Pediatric Research, 17(4), 254-258.
Siimes, A. S., Valimaki, I. A., Sarajas, H. S., Sakone, K., & Oja, R. T. (1984). Heart rate variation in relation to age and sleep state in neonatal lambs. Acta Physiologica Scandinavica.Supplementum, 537, 7-15.
Simon, G. E., Cunningham, M. L., & Davis, R. L. (2002). Outcomes of prenatal antidepressant exposure. The American Journal of Psychiatry, 159(12), 2055-2061.
Skop, B. P., Finkelstein, J. A., Mareth, T. R., Magoon, M. R., & Brown, T. M. (1994). The serotonin syndrome associated with paroxetine, an over-the-counter cold remedy, and vascular disease. The American Journal of Emergency Medicine, 12(6), 642-644.
Smith, F. G., & Lumbers, E. R. (1989). Comparison of renal function in term fetal sheep and newborn lambs. Biology of the Neonate, 55(4-5), 309-316.
Spencer, M. J. (1993). Fluoxetine hydrochloride (prozac) toxicity in a neonate. Pediatrics, 92(5), 721-722.

246
Spina, E., Scordo, M. G., & D'Arrigo, C. (2003). Metabolic drug interactions with new psychotropic agents. Fundamental & Clinical Pharmacology, 17(5), 517-538.
Stahl, S. M. (1998). Basic psychopharmacology of antidepressants, part 1: Antidepressants have seven distinct mechanisms of action. The Journal of Clinical Psychiatry, 59 Suppl 4, 5-14.
Steer, R. A., Scholl, T. O., Hediger, M. L., & Fischer, R. L. (1992). Self-reported depression and negative pregnancy outcomes. Journal of Clinical Epidemiology, 45(10), 1093-1099.
Stevens, J. C., & Wrighton, S. A. (1993). Interaction of the enantiomers of fluoxetine and norfluoxetine with human liver cytochromes P450. The Journal of Pharmacology and Experimental Therapeutics, 266(2), 964-971.
Stewart, R. C. (2007). Maternal depression and infant growth: A review of recent evidence. Maternal & Child Nutrition, 3(2), 94-107.
Stiskal, J. A., Kulin, N., Koren, G., Ho, T., & Ito, S. (2001). Neonatal paroxetine withdrawal syndrome. Archives of Disease in Childhood.Fetal and Neonatal Edition, 84(2), F134-5.
Stowe, Z. N., Hostetter, A. L., Owens, M. J., Ritchie, J. C., Sternberg, K., Cohen, L. S., et al. (2003). The pharmacokinetics of sertraline excretion into human breast milk: Determinants of infant serum concentrations. J Clin Psychiatry, 64(1), 73-80.
Summary minutes of the pediatrics subcommittee of the anti-infective drugs advisory committee. (2004). Retrieved 03/2012http://www.fda.gov/ohrms/dockets/ac/04/minutes/2004-4050M1.pdf
Suri, R., Altshuler, L., Hellemann, G., Burt, V. K., Aquino, A., & Mintz, J. (2007). Effects of antenatal depression and antidepressant treatment on gestational age at birth and risk of preterm birth. The American Journal of Psychiatry, 164(8), 1206-1213.
Suri, R., Altshuler, L., Hendrick, V., Rasgon, N., Lee, E., & Mintz, J. (2004). The impact of depression and fluoxetine treatment on obstetrical outcome. Archives of Women's Mental Health, 7(3), 193-200.
Suttie, J. M., Foster, D. L., Veenvliet, B. A., Manley, T. R., & Corson, I. D. (1991). Influence of food intake but independence of body weight on puberty in female sheep. Journal of Reproduction and Fertility, 92(1), 33-39.
Symonds, M. E., Pope, M., & Budge, H. (2012). Adipose tissue development during early life: Novel insights into energy balance from small and large mammals. The Proceedings of the Nutrition Society, 71(3), 363-370.
Tamashiro, K. L., & Moran, T. H. (2010). Perinatal environment and its influences on metabolic programming of offspring. Physiology & Behavior, 100(5), 560-566.

247
Thayer, J. F., & Lane, R. D. (2007). The role of vagal function in the risk for cardiovascular disease and mortality. Biological Psychology, 74(2), 224-242.
Thorburn, G. D., & Challis, J. R. (1979). Endocrine control of parturition. Physiological Reviews, 59(4), 863-918.
Toh, S., Mitchell, A. A., Louik, C., Werler, M. M., Chambers, C. D., & Hernandez-Diaz, S. (2009). Antidepressant use during pregnancy and the risk of preterm delivery and fetal growth restriction. Journal of Clinical Psychopharmacology, 29(6), 555-560.
Toohey, J. (2012). Depression during pregnancy and postpartum. Clinical Obstetrics and Gynecology, 55(3), 788-797.
Top 200 generic drugs by total prescriptions 2008(2009).
Trindade, E., Menon, D., Topfer, L. A., & Coloma, C. (1998). Adverse effects associated with selective serotonin reuptake inhibitors and tricyclic antidepressants: A meta-analysis. CMAJ : Canadian Medical Association Journal = Journal De l'Association Medicale Canadienne, 159(10), 1245-1252.
Tsai, M. L., & Lin, M. T. (1986). Hypertension and tachycardia produced by inhibition of reuptake of 5-hydroxytryptamine by fluoxetine in the rat. Neuropharmacology, 25(7), 799-802.
Turner, A. I., Canny, B. J., Hobbs, R. J., Bond, J. D., Clarke, I. J., & Tilbrook, A. J. (2002). Influence of sex and gonadal status of sheep on cortisol secretion in response to ACTH and on cortisol and LH secretion in response to stress: Importance of different stressors. The Journal of Endocrinology, 173(1), 113-122.
Turner, A. J., Brown, R. D., Carlstrom, M., Gibson, K. J., & Persson, A. E. (2008). Mechanisms of neonatal increase in glomerular filtration rate. American Journal of Physiology.Regulatory, Integrative and Comparative Physiology, 295(3), R916-21.
Usala, T., Clavenna, A., Zuddas, A., & Bonati, M. (2008). Randomised controlled trials of selective serotonin reuptake inhibitors in treating depression in children and adolescents: A systematic review and meta-analysis. European Neuropsychopharmacology : The Journal of the European College of Neuropsychopharmacology, 18(1), 62-73.
Van Leeuwen, P., Lange, S., Bettermann, H., Gronemeyer, D., & Hatzmann, W. (1999). Fetal heart rate variability and complexity in the course of pregnancy. Early Human Development, 54(3), 259-269.
van Zyl, L. T., Hasegawa, T., & Nagata, K. (2008). Effects of antidepressant treatment on heart rate variability in major depression: A quantitative review. BioPsychoSocial Medicine, 2, 12-0759-2-12.

248
Vanderlei, L. C., Pastre, C. M., Hoshi, R. A., Carvalho, T. D., & Godoy, M. F. (2009). Basic notions of heart rate variability and its clinical applicability. Revista Brasileira De Cirurgia Cardiovascular : Orgao Oficial Da Sociedade Brasileira De Cirurgia Cardiovascular, 24(2), 205-217.
Vaswani, M., Linda, F. K., & Ramesh, S. (2003). Role of selective serotonin reuptake inhibitors in psychiatric disorders: A comprehensive review. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 27(1), 85-102.
Viau, V. (2002). Functional cross-talk between the hypothalamic-pituitary-gonadal and -adrenal axes. Journal of Neuroendocrinology, 14(6), 506-513.
Viau, V., & Meaney, M. J. (1991). Variations in the hypothalamic-pituitary-adrenal response to stress during the estrous cycle in the rat. Endocrinology, 129(5), 2503-2511.
von Bardeleben, U., Steiger, A., Gerken, A., & Holsboer, F. (1989). Effects of fluoxetine upon pharmacoendocrine and sleep-EEG parameters in normal controls. International Clinical Psychopharmacology, 4 Suppl 1, 1-5.
von Leupoldt, A., & Dahme, B. (2005). Cortical substrates for the perception of dyspnea. Chest, 128(1), 345-354.
Voss, B., Mohr, E., & Krzywanek, H. (2002). Effects of aqua-treadmill exercise on selected blood parameters and on heart-rate variability of horses. Journal of Veterinary Medicine.A, Physiology, Pathology, Clinical Medicine, 49(3), 137-143.
Voysey, M. A. (2004). SSRI treatment for under-18s. CMAJ : Canadian Medical Association Journal = Journal De l'Association Medicale Canadienne, 170(12), 1771; author reply 1171-2.
Vreeburg, S. A., Hoogendijk, W. J., van Pelt, J., Derijk, R. H., Verhagen, J. C., van Dyck, R., et al. (2009). Major depressive disorder and hypothalamic-pituitary-adrenal axis activity: Results from a large cohort study. Archives of General Psychiatry, 66(6), 617-626.
Wadhwa, P. D., Garite, T. J., Porto, M., Glynn, L., Chicz-DeMet, A., Dunkel-Schetter, C., et al. (2004). Placental corticotropin-releasing hormone (CRH), spontaneous preterm birth, and fetal growth restriction: A prospective investigation. American Journal of Obstetrics and Gynecology, 191(4), 1063-1069.
Walser, E. S., & Alexander, G. (1980). Mutual recognition between ewes and lambs. Reproduction, Nutrition, Development, 20(3B), 807-816.
Wang, W., Pizzonia, J. H., & Richerson, G. B. (1998). Chemosensitivity of rat medullary raphe neurones in primary tissue culture. The Journal of Physiology, 511 ( Pt 2)(Pt 2), 433-450.

249
Warburton, W., Hertzman, C., & Oberlander, T. F. (2010). A register study of the impact of stopping third trimester selective serotonin reuptake inhibitor exposure on neonatal health. Acta Psychiatrica Scandinavica, 121(6), 471-479.
Warden, S. J., Robling, A. G., Sanders, M. S., Bliziotes, M. M., & Turner, C. H. (2005). Inhibition of the serotonin (5-hydroxytryptamine) transporter reduces bone accrual during growth. Endocrinology, 146(2), 685-693.
Warner, R., Appleby, L., Whitton, A., & Faragher, B. (1996). Demographic and obstetric risk factors for postnatal psychiatric morbidity. The British Journal of Psychiatry : The Journal of Mental Science, 168(5), 607-611.
Warren, J. H., Jaffe, R. S., Wraa, C. E., & Stebbins, C. L. (1997). Effect of autonomic blockade on power spectrum of heart rate variability during exercise. The American Journal of Physiology, 273(2 Pt 2), R495-502.
Waterstone, M., Wolfe, C., Hooper, R., & Bewley, S. (2003). Postnatal morbidity after childbirth and severe obstetric morbidity. BJOG : An International Journal of Obstetrics and Gynaecology, 110(2), 128-133.
Watts, S. W., Morrison, S. F., Davis, R. P., & Barman, S. M. (2012). Serotonin and blood pressure regulation. Pharmacological Reviews, 64(2), 359-388.
Weissman, M. M., & Olfson, M. (1995). Depression in women: Implications for health care research. Science (New York, N.Y.), 269(5225), 799-801.
Weissman, M. M., Wolk, S., Wickramaratne, P., Goldstein, R. B., Adams, P., Greenwald, S., et al. (1999). Children with prepubertal-onset major depressive disorder and anxiety grown up. Archives of General Psychiatry, 56(9), 794-801.
Welberg, L. A., Seckl, J. R., & Holmes, M. C. (2000). Inhibition of 11beta-hydroxysteroid dehydrogenase, the foeto-placental barrier to maternal glucocorticoids, permanently programs amygdala GR mRNA expression and anxiety-like behaviour in the offspring. The European Journal of Neuroscience, 12(3), 1047-1054.
Wen, S. W., Yang, Q., Garner, P., Fraser, W., Olatunbosun, O., Nimrod, C., et al. (2006). Selective serotonin reuptake inhibitors and adverse pregnancy outcomes. Am J Obstet Gynecol, 194(4), 961-966.
Wheeler, T., Cooke, E., & Murrills, A. (1979). Computer analysis of fetal heart rate variation during normal pregnancy. British Journal of Obstetrics and Gynaecology, 86(3), 186-197.
Whitaker-Azmitia, P. M. (2001). Serotonin and brain development: Role in human developmental diseases. Brain Research Bulletin, 56(5), 479-485.

250
Whitaker-Azmitia, P. M., Murphy, R., & Azmitia, E. C. (1990). Stimulation of astroglial 5-HT1A receptors releases the serotonergic growth factor, protein S-100, and alters astroglial morphology. Brain Research, 528(1), 155-158.
Whittington, C. J., Kendall, T., Fonagy, P., Cottrell, D., Cotgrove, A., & Boddington, E. (2004). Selective serotonin reuptake inhibitors in childhood depression: Systematic review of published versus unpublished data. Lancet, 363(9418), 1341-1345.
Wichman, C. L., Fothergill, A., Moore, K. M., Lang, T. R., Heise, R. H.,Jr, & Watson, W. J. (2008). Recent trends in selective serotonin reuptake inhibitor use in pregnancy. Journal of Clinical Psychopharmacology, 28(6), 714-716.
Wichman, C. L., Moore, K. M., Lang, T. R., St Sauver, J. L., Heise, R. H.,Jr., & Watson, W. J. (2009). Congenital heart disease associated with selective serotonin reuptake inhibitor use during pregnancy. Mayo Clin Proc, 84(1), 23-27.
Wisner, K. L., Sit, D. K., Hanusa, B. H., Moses-Kolko, E. L., Bogen, D. L., Hunker, D. F., et al. (2009). Major depression and antidepressant treatment: Impact on pregnancy and neonatal outcomes. Am J Psychiatry, 166(5), 557-566.
Wong, D. T., Fuller, R. W., & Robertson, D. W. (1990). Fluoxetine and its two enantiomers as selective serotonin uptake inhibitors. Acta Pharmaceutica Nordica, 2(3), 171-180.
Wright, C. E., & Angus, J. A. (1989). 5-carboxamidotryptamine elicits 5-HT2 and 5-HT3 receptor-mediated cardiovascular responses in the conscious rabbit: Evidence for 5-HT release from platelets. Journal of Cardiovascular Pharmacology, 13(4), 557-564.
Wright, C. M., Parkinson, K. N., & Drewett, R. F. (2006). The influence of maternal socioeconomic and emotional factors on infant weight gain and weight faltering (failure to thrive): Data from a prospective birth cohort. Archives of Disease in Childhood, 91(4), 312-317.
Yarney, T. A., & Sanford, L. M. (1989). Pubertal changes in the secretion of gonadotropic hormones, testicular gonadotropic receptors and testicular function in the ram. Domestic Animal Endocrinology, 6(3), 219-229.
Yau, J. L., Hibberd, C., Noble, J., & Seckl, J. R. (2002). The effect of chronic fluoxetine treatment on brain corticosteroid receptor mRNA expression and spatial memory in young and aged rats. Brain Research.Molecular Brain Research, 106(1-2), 117-123.
You, Q., Karrow, N. A., Cao, H., Rodriguez, A., Mallard, B. A., & Boermans, H. J. (2008). Variation in the ovine cortisol response to systemic bacterial endotoxin challenge is predominantly determined by signalling within the hypothalamic-pituitary-adrenal axis. Toxicology and Applied Pharmacology, 230(1), 1-8.

251
Young, E. A. (1995). The role of gonadal steroids in hypothalamic-pituitary-adrenal axis regulation. Critical Reviews in Neurobiology, 9(4), 371-381.
Young, E. A. (1996). Sex differences in response to exogenous corticosterone: A rat model of hypercortisolemia. Molecular Psychiatry, 1(4), 313-319.
Yuwiler, A., Oldendorf, W. H., Geller, E., & Braun, L. (1977). Effect of albumin binding and amino acid competition on tryptophan uptake into brain. Journal of Neurochemistry, 28(5), 1015-1023.
Zeskind, P. S., & Stephens, L. E. (2004). Maternal selective serotonin reuptake inhibitor use during pregnancy and newborn neurobehavior. Pediatrics, 113(2), 368-375.
Zuckerman, B., Amaro, H., Bauchner, H., & Cabral, H. (1989). Depressive symptoms during pregnancy: Relationship to poor health behaviors. American Journal of Obstetrics and Gynecology, 160(5 Pt 1), 1107-1111.
Zuckerman, B., Bauchner, H., Parker, S., & Cabral, H. (1990). Maternal depressive symptoms during pregnancy, and newborn irritability. Journal of Developmental and Behavioral Pediatrics : JDBP, 11(4), 190-194.

252
Appendices
Appendix 1: Actiwatch Variables Data In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age:
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. ~4 day 10 day 1M 3M 6M 1Y
N = 6-11 Mean
SEM
N = 9-10 Mean
SEM
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 4 Mean
SEM
N = 5-6 Mean
SEM
Active bout per day Normal day in pen 63.3 4.24 47.6 3.93 44.6 3.62 36.0 4.81 33.7 3.40 Average of 3 or 4 days 57.3 5.14 45.4 2.97 48.7 3.19 41.4 4.76 34.5 3.07 Control exp 40.2 3.49 61.3 5.84 48.1 4.28 33.7 2.81 24.9 4.88 20.8 2.64 FX exp 42.4 4.78 55.3 5.08 35.8 2.81 33.4 4.50 28.0 5.05 18.8 1.76 Post FX d1 47.5 3.22 60.2 5.89 45.7 3.57 37.8 4.80 31.1 5.39 23.8 2.67 Post FX day 2 49.7 3.97 58.5 4.60 45.0 3.45 35.4 3.85 24.8 4.16 20.0 2.09 Post FX day3 48.0 4.47 58.8 4.97 43.6 3.00 43.5 3.15 42.2 4.23 42.1 6.38 Mean score in active Normal day in pen 126.6 5.76 84.5 5.82 62.5 4.70 58.4 3.76 55.8 7.00 Average of 3 or 4 days 116.5 8.39 85.0 4.61 66.7 3.37 63.2 4.27 56.7 6.47 Control exp 95.2 8.51 118.7 8.84 84.0 6.85 52.1 4.17 45.3 4.77 47.8 8.47 FX exp 92.8 6.96 109.8 8.19 72.3 4.00 48.4 4.84 48.8 4.73 42.0 4.72 Post FX d1 108.2 5.26 117.0 8.09 83.6 4.95 53.3 5.35 51.4 4.97 49.1 3.73 Post FX day 2 109.1 6.78 114.1 8.40 83.8 4.29 52.0 5.51 45.2 5.82 43.9 4.33 Post FX day3 109.1 8.84 118.4 7.12 82.0 4.48 61.6 5.47 67.2 5.34 70.0 9.28

253
~ 4 day 10 day 1M 3M 6M 1Y
N = 6-11 Mean
SEM
N = 9-10 Mean
SEM
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 4 Mean
SEM
N = 5-6 Mean
SEM
Episode duration Normal day in pen 11.6 0.57 17.9 1.18 24.8 2.23 25.0 1.76 28.0 3.63 Average of 3 or 4 days 12.7 0.96 17.5 0.91 22.4 1.33 24.3 1.75 27.3 3.16 Control exp 15.6 1.17 12.7 1.00 18.1 1.52 30.6 3.57 33.1 4.24 34.2 6.02 FX exp 16.4 1.27 14.1 1.48 20.6 1.22 33.8 4.09 30.3 3.00 36.5 4.17 Post FX d1 13.6 0.62 12.8 0.90 17.9 1.06 31.9 5.51 28.8 2.77 30.1 2.09 Post FX day 2 13.7 0.90 13.4 1.19 17.7 0.88 32.3 4.51 33.6 4.67 34.3 3.17 Post FX day3 14.2 1.44 12.6 0.79 18.2 1.15 25.4 2.25 21.9 2.01 22.3 2.70 Total activity score Normal day in pen 364101 24379 273966 22628 256500 20844 207330 27707 194202 19554 Average of 3 or 4 days 329552 29573 261100 17066 280306 18370 238344 27423 198719 17684 Control exp 231622 20094 353036 33624 276761 24624 193704 16161 143300 28081 119890 15178 FX exp 244185 27519 318034 29246 206102 16201 192362 25905 160958 29069 108207 10106 Post FX d1 273458 18561 346625 33925 262755 20552 217733 27649 179190 31005 136994 15370 Post FX day 2 286283 22827 336898 26493 258849 19842 203499 22156 142906 23973 115289 12053 Post FX day3 276287 25708 338189 28610 250920 17267 250660 18128 243123 24369 242252 36725

254
~ 4 day 10 day 1M 3M 6M 1Y
N = 6-11 Mean
SEM
N = 9-10 Mean
SEM
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 4 Mean
SEM
N = 5-6 Mean
SEM
% time mobile Normal day in pen 49.8 1.73 56.1 2.10 72.4 3.97 61.9 7.19 62.3 4.58 Average of 3 or 4 days 48.8 1.40 53.3 1.96 73.4 3.64 65.1 5.07 62.5 4.42 Control exp 42.5 1.72 51.3 1.75 57.2 1.44 64.5 4.20 53.1 6.23 46.5 5.11 FX exp 45.7 4.02 49.8 1.72 49.3 2.01 65.2 5.04 55.8 6.46 47.0 5.52 Post FX d1 43.7 1.58 50.9 1.88 54.6 2.47 68.5 3.69 60.1 7.42 50.1 6.64 Post FX day 2 45.2 1.48 51.3 0.98 53.6 2.76 66.9 3.71 54.5 3.87 47.3 5.76 Post FX day3 43.9 1.60 49.1 1.72 53.4 2.59 71.8 3.33 63.4 5.33 59.6 3.89 % time immobile Normal day in pen 50.3 1.73 43.9 2.10 27.6 3.97 38.2 7.19 37.7 4.58 Average of 3 or 4 days 51.2 1.40 46.7 1.96 26.6 3.64 34.9 5.07 37.5 4.42 Control exp 55.3 3.78 48.7 1.75 42.8 1.44 35.5 4.20 46.9 6.23 53.5 5.11 FX exp 55.4 2.98 50.2 1.72 50.7 2.01 34.8 5.04 44.2 6.46 53.0 5.52 Post FX d1 56.3 1.58 49.1 1.88 45.4 2.47 31.5 3.69 39.9 7.42 50.0 6.64 Post FX day 2 54.8 1.48 48.7 0.98 46.4 2.76 33.1 3.71 45.5 3.87 52.8 5.76 Post FX day3 56.1 1.60 50.9 1.72 46.6 2.59 28.2 3.33 36.6 5.33 40.4 3.89

255
Appendix 2: Heart Rate, Arterial Pressure And HRV In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection:
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. 2.1: Heart Rate And Arterial Pressure: ~4 day 10 day 1M 3M 6M 1Y
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 5-8 Mean
SEM
N = 12 Mean SEM
N = 7 Mean
SEM
N = 9 Mean SEM
Heart rate -30 min 228 9.5 203 5.7 241 8.9 129 7.6 108 5.6 104 4.7 -15 min 228 9.1 196 6.9 241 7.2 125 6.0 106 5.9 115 6.3 5 min 195 9.3 179 7.6 210 5.2 123 4.7 107 9.3 102 6.6 15min 197 9.0 180 8.9 213 5.0 115 4.5 104 6.6 103 8.1 30min 199 9.5 179 7.9 215 6.1 108 3.3 100 4.8 98 5.4 60min 209 7.9 200 6.4 221 5.8 108 4.9 92 2.6 89 2.8 120min 223 9.7 206 11.6 230 12.4 107 4.0 95 3.9 90 4.1 Arterial pressure -30 min 67.8 1.4 73.5 3.1 68.3 2.1 75.5 3.0 71.5 3.6 85.0 1.9 -15 min 66.6 1.8 75.2 3.4 67.5 2.3 75.1 2.2 70.3 2.8 84.3 2.2 5 min 72.4 1.4 85.5 2.6 72.8 2.0 82.8 3.0 79.9 4.6 97.6 3.2 15min 70.2 1.8 81.3 2.6 71.5 2.1 80.2 1.9 75.9 3.6 90.1 1.7 30min 67.8 1.4 77.2 2.2 67.9 1.6 79.5 2.0 73.3 3.8 89.8 1.7 60min 68.7 1.6 79.9 3.2 69.0 2.0 77.5 1.9 74.2 3.5 88.3 1.9 120min 68.7 1.8 76.5 2.2 71.2 2.2 75.5 2.7 74.3 3.6 87.1 2.3

256
2.2: Time Domain Of HRV (SDNN, SD Delta NN, RMSSD): 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. ~ 4 day 10 day 1M 3M 6M 1Y
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 5-8 Mean
SEM
N = 12 Mean SEM
N = 7 Mean
SEM
N = 9 Mean SEM
SDNN -30 min 20.6 2.5 29.2 2.6 51.6 12.9 61.1 6.2 67.5 5.7 112.1 15.9 30min 28.9 3.7 43.8 4.9 72.4 13.9 72.8 6.3 96.1 13.1 140.2 14.5 60min 24.0 3.1 38.0 2.3 55.1 11.5 76.6 7.6 82.4 11.8 123.7 16.4 90 min 23.5 3.4 34.7 2.2 51.9 9.5 74.6 7.4 78.5 7.4 118.4 18.5 120 min 18.4 2.8 36.4 5.5 38.6 5.6 66.6 6.6 75.8 10.1 116.0 17.2 SD Delta NN -30 min 16.8 2.8 18.0 2.6 37.0 12.1 51.9 8.2 49.8 11.0 91.0 12.4 30min 22.4 3.2 30.7 4.7 57.9 14.4 57.2 7.6 73.1 18.1 110.8 15.5 60min 22.0 3.1 22.5 2.0 41.0 11.8 72.6 14.5 53.4 16.2 85.7 12.5 90 min 21.5 4.7 20.1 2.3 37.2 9.5 68.1 11.2 55.9 10.4 81.9 13.8 120 min 14.6 2.9 29.6 10.1 23.4 6.4 59.0 6.8 59.8 11.7 87.1 13.4 RMSSD -30 min 16.8 2.8 18.0 2.6 36.9 12.1 51.9 8.1 49.8 11.0 91.0 12.4 30min 22.4 3.2 30.7 4.7 57.9 14.4 57.2 7.6 73.1 18.1 110.8 15.5 60min 22.0 3.1 22.5 2.0 41.0 11.8 72.6 14.5 53.4 16.2 85.7 12.5 90 min 21.5 4.7 20.1 2.3 37.2 9.5 68.1 11.2 55.9 10.4 81.9 13.8 120 min 14.6 2.9 29.6 10.1 23.4 6.4 59.0 6.8 59.8 11.7 87.1 13.4

257
2.3: Frequency Domain Of HRV (LF Power, HF Power, LF/HF Ratio): 1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. ~ 4 day 10 day 1M 3M 6M 1Y
N = 9-12 Mean
SEM
N = 12 Mean
SEM
N = 5-8 Mean
SEM
N = 12 Mean SEM
N = 7 Mean
SEM
N = 9 Mean SEM
LF power -30 min 75.2 17.4 192.5 29.8 711.4 282.2 531.1 90.4 727.7 129.8 2410.1 665.4 30min 104.7 27.1 275.8 35.0 882.6 275.4 664.6 110.9 1250.7 524.0 3614.1 765.6 60min 94.2 21.5 263.3 43.1 642.3 203.3 675.4 118.2 1087.2 460.5 2150.2 531.9 90 min 73.3 15.9 261.6 57.5 500.3 111.7 632.4 92.8 944.8 241.3 2218.2 556.5 120 min 67.7 17.4 245.0 45.3 337.2 78.7 671.1 103.9 1094.7 306.5 2749.8 632.0 HF power -30 min 47.7 10.6 74.1 10.8 532.8 288.6 459.0 104.2 570.4 317.4 2176.1 535.9 30min 55.5 14.8 131.0 29.3 865.9 383.3 598.1 140.1 954.1 654.0 3585.3 866.0 60min 60.6 11.0 99.6 15.0 446.5 200.6 662.8 147.3 732.0 483.7 1950.0 521.5 90 min 56.7 15.7 89.5 18.8 310.9 130.3 618.1 124.1 653.9 336.5 1996.7 605.1 120 min 37.8 11.5 134.0 53.6 115.8 19.9 629.1 117.4 956.4 527.0 2478.0 630.2 LF/HF ratio -30 min 1.7 0.2 3.1 0.5 2.9 0.6 1.5 0.2 2.2 0.4 1.2 0.2 30min 2.2 0.4 2.9 0.5 2.1 0.5 1.5 0.2 2.6 0.7 1.2 0.2 60min 1.6 0.2 3.0 0.4 2.0 0.4 1.4 0.3 3.5 1.0 1.3 0.1 90 min 1.5 0.2 3.3 0.4 2.8 0.6 1.4 0.2 2.5 0.6 1.4 0.2 120 min 2.0 0.3 3.1 0.6 3.5 1.0 1.4 0.2 2.5 0.9 1.4 0.2

258
Appendix 3: Arterial Blood Gases And Body Temperature In The Lambs At ~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection:
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. ~4 day 10 day 1M 3M 6M 1Y
N = 11 N = 8-12 N = 6-10 N = 10-12 N = 7 N = 10
pH Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM -15 min 7.385 0.010 7.433 0.009 7.443 0.010 7.496 0.014 7.502 0.019 7.482 0.014 5 min 7.369 0.013 7.427 0.011 7.439 0.007 7.494 0.011 7.503 0.013 7.498 0.012 15 min 7.388 0.010 7.425 0.008 7.439 0.011 7.475 0.012 7.481 0.008 7.506 0.011 30 min 7.383 0.009 7.424 0.009 7.428 0.006 7.489 0.009 7.487 0.014 7.506 0.011 60 min 7.378 0.012 7.411 0.006 7.434 0.012 7.467 0.016 7.474 0.011 7.488 0.008 120 min 7.375 0.010 7.413 0.007 7.456 0.009 7.467 0.014 7.479 0.016 7.488 0.018 Temperature -15 min 39.8 0.1 39.6 0.2 39.7 0.2 39.6 0.1 39.6 0.1 39.3 0.2 5 min 39.5 0.2 39.7 0.2 39.5 0.1 39.7 0.1 39.6 0.1 39.3 0.2 15 min 39.3 0.3 39.6 0.2 39.6 0.1 39.7 0.1 39.5 0.1 39.4 0.1 30 min 39.4 0.2 39.9 0.2 39.7 0.1 39.8 0.1 39.5 0.1 39.2 0.2 60 min 39.4 0.2 39.9 0.2 39.7 0.1 40 0.2 39.5 0.1 39.3 0.1 120 min 39.5 0.2 39.7 0.1 39.5 0.2 39.9 0.1 39.4 0.1 39.3 0.1 O2 saturation -15 min 93.3 0.4 93.1 0.5 93.2 0.3 95.4 0.2 94.5 1.4 95.0 0.4 5 min 93.4 0.4 92.5 0.7 89.0 1.5 93.8 1.1 92.1 1.9 92.9 0.8 15 min 93.5 0.4 92.8 0.4 93.1 0.4 95.3 0.4 93.6 1.4 96.0 0.2 30 min 93.5 0.4 92.9 0.6 92.3 0.4 94.6 0.6 94.4 0.9 95.5 0.3 60 min 93.2 0.5 92.4 0.3 93.4 0.3 94.6 0.3 94.9 0.5 95.2 0.2 120 min 92.9 1.0 93.0 0.3 93.0 1.0 95.2 0.2 95.1 0.4 95.7 0.3

259
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. ~4 day 10 day 1M 3M 6M 1Y
N = 11 N = 8-12 N = 6-10 N = 10-12 N = 7 N = 10
PO2 Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM -15 min 88.0 3.8 95.0 3.9 95.3 2.5 104.7 2.9 111.9 5.4 110.5 2.5 5 min 89.6 2.8 93.7 3.2 81.0 5.0 96.2 3.5 95.0 7.7 94.0 5.8 15min 90.5 4.0 94.0 2.7 93.5 2.2 103.0 3.5 104.1 6.5 114.3 2.1 30min 86.9 3.4 94.4 4.0 92.6 1.8 99.9 3.4 106.6 5.5 108.6 3.2 60min 89.7 4.2 95.2 2.4 95.5 2.1 102.1 2.5 108.3 2.1 107.1 2.6 120min 91.9 3.5 96.7 1.7 94.4 3.3 103.4 2.4 106.3 3.7 111.8 2.1 PCO2
-15 min 42.2 1.0 40.9 0.7 41.5 1.2 36.6 1.2 36.2 1.2 37.6 1.1 5 min 42.8 1.6 41.7 1.1 40.8 1.0 36.8 0.7 36.6 0.9 36.4 0.8 15min 40.9 0.9 41.5 0.7 40.3 1.0 38.2 1.3 38.0 0.7 35.5 0.8 30min 40.9 0.7 41.9 0.9 41.4 0.8 37.9 0.6 37.7 0.9 36.0 0.9 60min 40.0 1.1 41.2 0.7 41.1 1.7 38.9 1.4 36.8 0.6 36.7 0.7 120min 39.3 0.8 40.9 0.6 39.1 1.0 38.5 1.6 37.2 0.3 36.1 1.2

260
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year.
~4 day 10 day 1M 3M 6M 1Y N = 11 N = 8-12 N = 6-10 N = 10-12 N = 7 N = 10
Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Mean SEMBE -15 min 0.27 0.75 3.09 0.62 4.18 0.60 5.17 0.81 5.43 1.59 4.70 0.62 5 min -0.73 0.73 3.00 0.47 3.55 0.68 5.17 0.89 5.86 1.03 5.10 0.57 15min -0.45 0.72 2.91 0.46 3.00 0.56 4.64 0.91 4.86 0.88 5.00 0.68 30min -0.73 0.71 2.91 0.49 3.00 0.45 5.58 0.92 5.14 1.20 5.60 0.65 60min -1.55 0.89 1.64 0.43 3.18 0.60 4.45 0.96 3.71 0.92 4.40 0.73 120min -2.09 0.51 1.64 0.49 3.82 0.75 4.00 0.93 4.43 1.23 4.00 0.86 HCO3 -15 min 24.6 0.7 26.7 0.5 27.5 0.6 27.6 0.7 27.9 1.3 27.6 0.5 5 min 24.1 0.6 26.8 0.4 26.9 0.7 27.8 0.7 28.2 0.8 27.7 0.4 15min 23.9 0.6 26.6 0.4 26.5 0.5 27.5 0.8 27.9 0.8 27.5 0.5 30min 23.8 0.6 26.7 0.4 26.6 0.5 28.2 0.8 28.0 1.0 28.0 0.6 60min 23.1 0.7 25.5 0.4 26.6 0.6 27.4 0.8 26.5 0.7 27.4 0.6 120min 22.5 0.4 25.4 0.4 26.8 0.7 27.1 0.8 27.2 0.9 26.8 0.6 Hb -15 min 11.2 0.4 9.0 0.4 8.2 0.6 9.6 0.2 9.2 0.2 9.6 0.2 5 min 11.3 0.4 9.1 0.4 8.9 0.6 9.9 0.2 9.8 0.2 10.6 0.2 15min 11.3 0.4 9.1 0.4 8.9 0.7 9.9 0.2 9.6 0.2 10.0 0.2 30min 11.3 0.4 9.2 0.4 8.8 0.7 9.7 0.2 9.3 0.2 9.8 0.2 60min 11.3 0.5 9.2 0.4 8.7 0.7 9.8 0.2 9.3 0.2 9.7 0.2 120min 11.3 0.4 9.2 0.5 8.5 0.7 9.7 0.2 9.4 0.2 9.9 0.2

261
Appendix 4: Glucose, Lactate Concentration And Electrolite In The Lambs At~ 4, 10 Days, 1, 3, 6 And 12 Months Of Age With FX Injection:
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year. Mean ~4 day 10 day 1M 3M 6M 1Y N = 11 N = 8-12 N = 6-10 N = 10-12 N = 7 N = 10
Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Glucose -15 min 5.492 0.249 5.446 0.158 4.155 0.126 3.166 0.137 2.950 0.135 2.936 0.103 5 min 5.539 0.213 5.451 0.127 4.049 0.143 3.099 0.137 3.047 0.119 2.818 0.085 15min 5.345 0.214 5.321 0.138 4.020 0.138 3.166 0.158 2.981 0.087 2.894 0.087 30min 5.483 0.214 5.450 0.172 4.321 0.189 3.232 0.137 3.063 0.093 2.974 0.097 60min 5.667 0.194 5.590 0.105 4.294 0.172 3.257 0.160 3.139 0.080 3.026 0.113 120min 5.574 0.174 5.791 0.104 4.474 0.213 3.413 0.169 3.242 0.090 3.156 0.130 Lactate -15 min 0.904 0.054 0.947 0.225 0.820 0.275 0.633 0.063 0.510 0.043 0.562 0.044 5 min 0.909 0.062 0.811 0.112 0.859 0.300 0.645 0.063 0.595 0.063 0.580 0.039 15min 0.905 0.071 0.834 0.106 0.806 0.230 0.625 0.057 0.669 0.090 0.571 0.046 30min 0.841 0.058 0.651 0.062 0.820 0.221 0.665 0.077 0.650 0.099 0.547 0.039 60min 0.814 0.053 0.862 0.190 0.841 0.261 0.728 0.101 0.662 0.098 0.606 0.041 120min 0.784 0.069 0.777 0.090 0.821 0.237 0.619 0.078 0.540 0.042 0.550 0.043

262
1M = 1 month, 3M = 3 months, 6M = 6 months, 1Y = 1 year.
Mean ~4 day 10 day 1M 3M 6M 1Y N = 11 N = 8-12 N = 6-10 N = 10-12 N = 7 N = 10
Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Mean SEM Na+ -15 min 141 0.5 143 0.6 143 0.4 146 0.5 146 0.5 147 0.4
5 min 140 0.6 143 0.5 143 0.5 146 0.4 147 0.5 147 0.4
15min 141 0.5 143 0.4 143 0.4 146 0.4 147 0.5 147 0.4
30min 141 0.6 143 0.4 143 0.4 146 0.4 146 0.6 147 0.5
60min 140 0.5 143 0.4 143 0.4 145 0.4 146 0.9 147 0.4
120min 140 0.7 143 0.5 143 0.5 145 0.4 146 0.8 146 0.3
K+ -15 min 4.05 0.12 3.85 0.11 4.09 0.11 4.02 0.09 3.83 0.10 4.01 0.08
5 min 4.29 0.14 3.89 0.11 4.12 0.11 4.18 0.10 3.96 0.10 4.06 0.05
15min 4.27 0.15 3.85 0.12 4.17 0.13 4.06 0.08 3.84 0.13 3.95 0.04
30min 4.28 0.13 3.99 0.14 4.14 0.09 4.13 0.07 3.81 0.10 3.95 0.05
60min 4.33 0.11 3.99 0.14 4.09 0.10 4.31 0.07 3.96 0.16 4.02 0.08
120min 4.40 0.13 4.02 0.14 4.12 0.13 4.31 0.12 4.17 0.09 4.19 0.07
iCa -15 min 1.51 0.03 1.48 0.02 1.42 0.02 1.22 0.02 1.23 0.04 1.20 0.02
5 min 1.49 0.03 1.48 0.02 1.44 0.01 1.23 0.02 1.23 0.03 1.20 0.02
15min 1.48 0.03 1.47 0.02 1.41 0.01 1.21 0.02 1.21 0.02 1.17 0.02
30min 1.52 0.03 1.49 0.02 1.43 0.01 1.22 0.02 1.20 0.02 1.17 0.02
60min 1.51 0.03 1.47 0.02 1.43 0.01 1.25 0.03 1.25 0.03 1.20 0.02
120min 1.50 0.03 1.47 0.01 1.40 0.01 1.28 0.02 1.28 0.03 1.23 0.02




















