
Effects of crop thinning and reduced grain numbers per ear on grain size in two winter wheat...
11
Ann. appl. Biol. (1979), 92,383-393 Printed in Great Britain 383 Effects of crop thinning and reduced grain numbers per ear on grain size in two winter wheat varieties given different amounts of nitrogen BY R. MARTINEZ-CARRASCO* AND GILLIAN N. THORNE Rothamsted Experimental Station, Harpenden, Herts, AL5 2JQ (Accepted 14 May 1979) SUMMARY To study the importance for final grain size in wheat (Triticum aestiuum, L.) of assimilate supply and the storage capacity of the grain, two field experiments were done. In 1976 nitrogen was applied in the range from none to 180 kg ha-', part of the crop was thinned, and the top halves of some ears of the short variety Hobbit and of the tall variety Maris Huntsman were removed soon after anthesis. In 1977 ears of Maris Huntsman were halved 5 days after anthesis or at 30 days after anthesis when grain volume was maximum. Thinning the crop from 360 to 180 ear-bearing shoots m-2 30 days before anthesis increased the number of grains per ear, except in the absence of nitrogen fertiliser, but did not increase grain size, grain dry weight per ear or total dry weight per culm. Removing the upper half of ears of Hobbit 5 days after anthesis increased dry weight per grain, but when this treatment was applied to Maris Huntsman either 5 days after anthesis in 1976 and 1977, or when grain volume was maximal in 1977, the grains failed to increase in dry weight. Non-grain dry weight of both varieties was increased by halving the ear. In both varieties the maximum volume of grains in halved ears was larger than in intact ears. Grain dry weight increased relatively less than volume after halving the ear of Hobbit, and the decrease in volume up to maturity was greater in halved than intact ears of both varieties. The larger grain volume in halved ears of Maris Huntsman in 1977 was associated with more endosperm cells. INTRODUCTION Total dry weight of a wheat crop is determined mainly by the size of the photosynthetic system, but grain yield depends on the distribution of dry matter, i.e. on the proportion of the total dry weight that is found in the economically useful part. Grain yield will not increase with increase in total dry weight if the storage capacity of the ear is limited (Thorne, 1971). Little is known about the physiological determinants of grain size, which is one of the components of the storage capacity of the ear. There is a good deal of evidence that, as well as assimilate supply, the capacity of the grain to accumulate dry matter may control final grain size (Bingham, 1967; Welbank, Witts & Thorne, 1968; Spiertz, 1974; Sofield, Evans, Cook & Wardlaw, 1977). Watson (197 1) suggested that the relative importance of these two factors varies with stage of grain growth: early in grain filling, when leaf area is near maximal, the movement of photosynthate into the grains would be limited by their sink strength; later sink strength would be less important than the supply of photosynthate. Bingham (1969) found that final grain weight depended to a large extent on grain dry weight soon after anthesis, and, according to Brocklehurst (1977), the rate of accumulation of dry matter in the grain is governed by the number of endosperm cells formed, which in turn is regulated by the supply of assimilate to the * Present address: Centro de Adafologia y Biologia Aplicada, Salamanca. Spain.
Transcript of Effects of crop thinning and reduced grain numbers per ear on grain size in two winter wheat...

Ann. appl. Biol. (1979), 92,383-393 Printed in Great Britain
383
Effects of crop thinning and reduced grain numbers per ear on grain size in two winter wheat varieties given different amounts of
nitrogen
BY R. MARTINEZ-CARRASCO* AND GILLIAN N. THORNE Rothamsted Experimental Station, Harpenden, Herts, AL5 2JQ
(Accepted 14 May 1979)
S U M M A R Y
To study the importance for final grain size in wheat (Triticum aestiuum, L.) of assimilate supply and the storage capacity of the grain, two field experiments were done. In 1976 nitrogen was applied in the range from none to 180 kg ha-', part of the crop was thinned, and the top halves of some ears of the short variety Hobbit and of the tall variety Maris Huntsman were removed soon after anthesis. In 1977 ears of Maris Huntsman were halved 5 days after anthesis or at 30 days after anthesis when grain volume was maximum. Thinning the crop from 360 to 180 ear-bearing shoots m-2 30 days before anthesis increased the number of grains per ear, except in the absence of nitrogen fertiliser, but did not increase grain size, grain dry weight per ear or total dry weight per culm.
Removing the upper half of ears of Hobbit 5 days after anthesis increased dry weight per grain, but when this treatment was applied to Maris Huntsman either 5 days after anthesis in 1976 and 1977, or when grain volume was maximal in 1977, the grains failed to increase in dry weight. Non-grain dry weight of both varieties was increased by halving the ear. In both varieties the maximum volume of grains in halved ears was larger than in intact ears. Grain dry weight increased relatively less than volume after halving the ear of Hobbit, and the decrease in volume up to maturity was greater in halved than intact ears of both varieties. The larger grain volume in halved ears of Maris Huntsman in 1977 was associated with more endosperm cells.
I N T R O D U C T I O N
Total dry weight of a wheat crop is determined mainly by the size of the photosynthetic system, but grain yield depends on the distribution of dry matter, i.e. on the proportion of the total dry weight that is found in the economically useful part. Grain yield will not increase with increase in total dry weight if the storage capacity of the ear is limited (Thorne, 1971). Little is known about the physiological determinants of grain size, which is one of the components of the storage capacity of the ear. There is a good deal of evidence that, as well as assimilate supply, the capacity of the grain to accumulate dry matter may control final grain size (Bingham, 1967; Welbank, Witts & Thorne, 1968; Spiertz, 1974; Sofield, Evans, Cook & Wardlaw, 1977). Watson (197 1) suggested that the relative importance of these two factors varies with stage of grain growth: early in grain filling, when leaf area is near maximal, the movement of photosynthate into the grains would be limited by their sink strength; later sink strength would be less important than the supply of photosynthate. Bingham (1969) found that final grain weight depended to a large extent on grain dry weight soon after anthesis, and, according to Brocklehurst (1977), the rate of accumulation of dry matter in the grain is governed by the number of endosperm cells formed, which in turn is regulated by the supply of assimilate to the
* Present address: Centro de Adafologia y Biologia Aplicada, Salamanca. Spain.

384 R . M A R T I N E Z - C A R R A S C O A N D G . N . T H O R N E
grains soon after fertilisation. However, in another experiment (Martinez-Carrasco & Thorne, 1979), dry matter accumulation late in the grain filling period did not appear to be related to the physical capacity of the grain, which is determined at an early stage. It is therefore possible that factors other than endosperm cell development control the sink capacity of the grains.
The effects on grain weight of factors which might alter assimilate supply and also the storage capacity of the grain were investigated in two field experiments done at Rothamsted in 1976 and 1977. In 1976 nitrogen was applied in the range from none to 180 kg ha-' to alter the supply of assimilate to the grains, and the number of tillers was decreased before anthesis, to improve early ear growth of the remaining shoots and to decrease competition for light after anthesis. To distinguish between the pre- and post-anthesis effects, carbohydrate supply per grain was increase.d still further by removing the upper half of some ears a few days after anthesis, when grain set was probably complete. These treatments were studied in the tall variety Maris Huntsman and in the short variety, Hobbit, to show whether the factors determining size are similar in contrasting genotypes. The results of this experiment indicated that grains of Hobbit, but not those of Maris Huntsman, increased in weight in response to increased assimilate supply. The results also suggested that, as previously (Martinez-Carrasco & Thorne, 1979), the physical capacity of the grain, as determined by volume, did not limit the response of grain weight to assimilate supply. The 1977 experiment was intended to study further the factors limiting grain weight in Maris Huntsman. To test whether the importance for final grain size of assimilate supply and the storage capacity of the grain varied during development, ears of Maris Huntsman halved early in grain growth, while endosperm cells were still dividing, were compared with ones halved when grain volume was maximal.
M E T H O D S
Arrangement of experiments 1976 experiment. This was part of an experiment (subsequently called the main experiment)
comparing the effects of several amounts of nitrogen on two varieties of winter wheat (Triticum aestiuum L.) (Pearman, Thomas & Thorne, 1978). The 32 treatment combinations of four rates of nitrogen fertiliser and two varieties, two shoot densities and two sizes of the ear were replicated twice in a randomised block factorial design. Variety and nitrogen were main plots and shoot density and ear size were sub-plots. The nitrogen amounts were 0, 60, 120 and 180 kg ha-' applied on 6 May (35 days before anthesis). The two varieties were the semi-dwarf Hobbit and the taller Maris Huntsman. Thirty days before anthesis, when the ear was showing stamen primordia (about the score 20 of Friend, Fisher & Helson, 1963), the smaller shoots were removed in a 50 cm length of seven adjacent rows (row width = 13 cm), to leave approximately 180 main stems m-2. This treatment will be referred to as thinning. The remaining six rows in that 50 cm strip were left untreated, the number of ears averaging 350 m-*. Tillers which grew subsequently on thinned plots were removed 7 days before anthesis. On three adjacent rows of the unthinned part of the strip and on three and a half adjacent rows of its thinned part, the top halves of the ears were removed 5 days after anthesis, leaving intact the ears in the other rows. This treatment will be referred to as halving. The two innermost rows, one thinned and the other unthinned, and the outer row on each side of each plot were kept as guard rows. Groups of rows for the different treatments were selected at random from the 13 rows of each plot.
The experiment was sown at 200 kg ha-' on 22 October 1975. Anthesis and maturity dates were 10 June and 19 July respectively.
1977 experiment. This experiment compared shoots with intact ears with shoots whose ears were halved either 5 days after anthesis or at 30 days when grain volume was approximately maximal. The largest shoots of each plot were selected for this experiment. There were two

Control of grain size in winter wheat 385
blocks, each of three plots. Individual plots had an area of 2.3 mz, with eight rows approximately 17.5 cm apart, surrounded by a 0.5 m strip of non-experimental wheat.
Winter wheat var. Maris Huntsman was sown at a rate of 200 kg ha-'; 94 kg N ha-' and ample phosphorus and potassium fertiliser was applied.
Observations The date when half the ears flowered was determined by regularly counting the ears showing
anthers in samples of 20 ear-bearing shoots per plot. Twenty and 28 days after anthesis in the 1976 experiment, and at 3- or 4-day intervals (or longer after maximum grain volume was attained) in the 1977 experiment, samples were taken for growth measurements. In 1976 the samples consisted of five adjacent ears in a row from every sub-plot. The stems of these ears were not removed so that competition was changed minimally. In 1977 two groups of five adjacent ear-bearing shoots were harvested on each occasion and the sub-samples pooled: 29 days after anthesis the sample consisted of 20 shoots. At maturity, 25 adjacent shoots were harvested in 1976 and two groups of 10 adjacent shoots in 1977. The two basal grains in the four uppermost spikelets of the lower half of the ear, excluding the top spikelet, were selected to determine grain volume, fresh and dry weights and water content. Additional samples of similar grains were taken 12 days after anthesis in 1977 to count the number of cells in the endosperm. In 1976 the development of the selected grains was examined at intervals in the control ears. Measurements of growth of the grains and the rest of the shoot, and determination of cell numbers, were the same as those described by Martinez-Carrasco & Thorne (1979).
RESULTS
1976 experiment Unthinned plants with intact ears, which were comparable to the plants in the main
experiment, had slightly larger ears with more and larger grains than reported by Pearman et al. (1978) (Fig. 1). In both sets of data, Hobbit had smaller grains than Maris Huntsman when given ample nitrogen (Fig. lc), but our measurements did not detect the large number of grains in the ears of Hobbit reported by Pearman et al. ( I 978). Averaged over all nitrogen treatments,
0.9 L I I I I 1 I I I
0 30 6 0 90 I20150180210 kg N ha-l
0 A
46 (c)
44 F mg 40 421
38 t
/- I 0 A A
0 A A 36
I I I I I I I I
0 30 60 90 120150 180210
kg N ha-' Fig. 1 . 1976 experiment. Effect of nitrogen fertiliser on (a) grain dry weight per ear; (b) number of grains per ear, and (c) dry weight per grain. Closed symbols (our experiment) unthinned plots with intact ears; open symbols, main experiment. Circles, Hobbit. Triangles, Maris Huntsman. Vertical bars in this and the other figures represent least significant differences ( P = 0.05).

386
70
60 - - 7 d
5 0 -
3 > 5 - 3 4 0 - E
2 3 0 -
.-, L. 0 3
- v F 5 20
6 x
10
0
-
-
-
-
I 1 I 1
10 20 30 40
Table 1. 1976 Experiment. ESfects of nitrogen fertiliser and crop thinning on the volume, fresh and dry weights and water content of the selected grains during development.
(Mean of cvs Maris Huntsman and Hobbit)
Days after anthesis . . . 20 28 39
Volume (PI)
F. wt (mg)
D. wt (mg)
Water content (mg)
+Ah kgNha- ' U T U T U T
0 66.4 63.5 65.6 63.1 42.1 39.0 60 66.2 67.8 70.6 68.7 40.0 40.5
120 63.0 64.6 68.5 69.7 40.1 43.7 180 60.5 66.7 66.5 68.9 41.4 41.2
S.E.D. 3.25 2.73 1.36 0 72.6 66.5 77.8 75.1 54.6 50.2
60 72.7 73.1 82.9 80.4 53.3 52.9 120 68.4 70.4 81.1 81.7 52.5 56.5 180 66.0 73.1 79.0 80.7 53.9 53.7
S.E.D. 3.89 3.11 1.58
0 29.5 26.8 43.6 40.0 47.2 43.8 60 30.2 30.2 46.2 43.7 46.4 45.1
120 28.2 29.3 44.9 45.1 45.7 46.4 180 27.4 30.1 43.7 42.8 45.6 45.8
S.E.D. 1.75 1.69 1.04 0 43.0 39.6 34.2 35.1 7.4 6.3
60 42.6 42.8 36.7 36.1 6.9 7,2 120 40.2 41.1 36.2 36.6 6.8 10.1 180 38.6 43.0 35.3 37.9 8.3 7.9
S.E.D 2.56 1.75 1.43
U: unthinned; T: thinned.

Control of grain size in winter wheat 387
Table 2. 1976 Experiment. EfSects of nitrogen fertiliser and crop thinning on growth attributes per culm at maturity. (Mean of cvs Maris Huntsman and Hobbit)
Kg N ha-'. . . 0 60 120 180 **** Thinning . . . U T U T U T U T S.E.D.
D.wtpergrain(mg) 46.4 41.0 43.3 42.1 42.5 42.9 42.2 43.3 1.37 No. grains 21.0 19.0 24.3 27.8 25.9 30.7 26.5 30.8 2.12 Grain d.wt (g) 0.95 0.79 1.05 1.16 1.11 1.32 1.11 1.33 0.102 Chaff d.wt (8) 0.27 0.23 0.29 0.31 0.30 0.35 0.30 0.34 0.023 Shoot d.wt (g) 1.06 0.87 1.05 1.06 1.10 1.17 1.10 1.18 0.082 Total d.wt (g) 2.28 1.90 2.50 2.53 2.51 2.84 2.50 2.85 0.195
U: unthinned: T: thinned.
grain dry weight per ear of Hobbit and Maris Huntsman were 1.31 and 1.40 g (s.E.D. 0.105) respectively in our experiment. These compare with values of 1.14 and 1.09 g (s.E.D. 0.033) respectively for the two varieties in the main experiment. The difference between the two sets of
45
G 35 E
5 6 25 3
4 c
L aJ u
15
5
( a ) Hobbit Maris Huntsman 80
70
- 60 3
50 2 0
5
40
30
k I I
10 20 30 40 10 20 30 40 Days after anthesis
Fig. 3. 1976 experiment. Effect of removing the top half of the ear on (a) the volume, (b) water content and (c) dry weight of the selected grains of Hobbit (left) and Maris Huntsman (right) during development. Open symbols represent intact ears and closed symbols halved ears. Values are per grain (means for 0, 60, 120 and 180 kg N ha-').
388 R. M A R T I N E Z - C A R R A S C O A N D G . N . T H O R N E
measurements may be attributed to the composition of the samples; very small ears were excluded in our experiment.
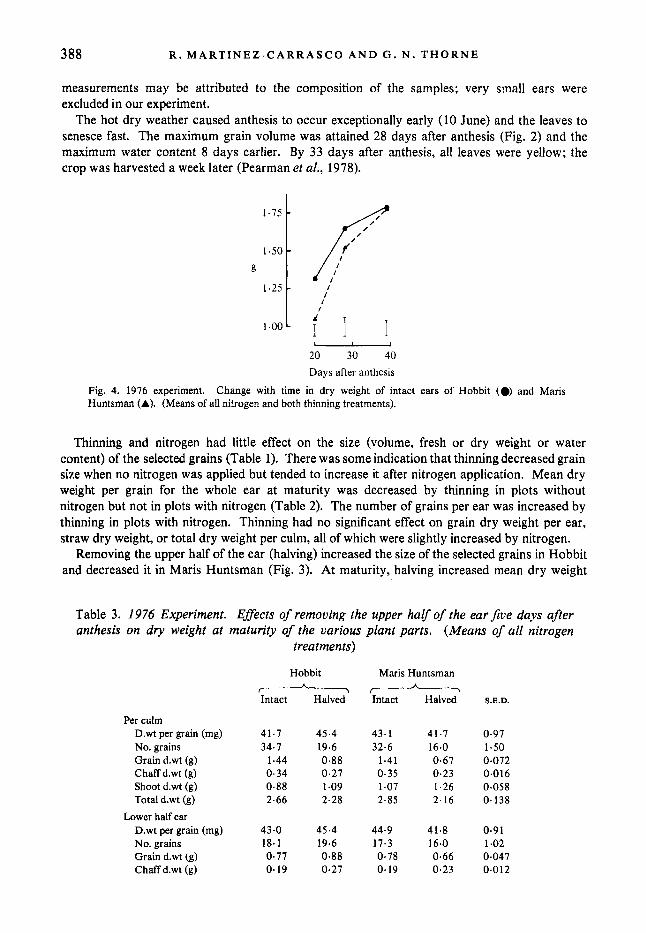
The hot dry weather caused anthesis to occur exceptionally early (10 June) and the leaves to senesce fast. The maximum grain volume was attained 28 days after anthesis (Fig. 2) and the maximum water content 8 days earlier. By 33 days after anthesis, all leaves were yellow; the crop was harvested a week later (Pearman et al., 1978).
20 30 40 Days after anthesis
Fig. 4. 1976 experiment. Huntsman (A). (Means of all nitrogen and both thinning treatments).
Change with time in dry weight of intact ears of Hobbit (0) and Maris
Thinning and nitrogen had little effect on the size (volume, fresh or dry weight or water content) of the selected grains (Table 1). There was some indication that thinning decreased grain size when no nitrogen was applied but tended to increase it after nitrogen application. Mean dry weight per grain for the whole ear at maturity was decreased by thinning in plots without nitrogen but not in plots with nitrogen (Table 2). The number of grains per ear was increased by thinning in plots with nitrogen. Thinning had no significant effect on grain dry weight per ear, straw dry weight, or total dry weight per culm, all of which were slightly increased by nitrogen.
Removing the upper half of the ear (halving) increased the size of the selected grains in Hobbit and decreased it in Maris Huntsman (Fig. 3). At maturity, halving increased mean dry weight
Table 3. 1976 Experiment. Effects of removing the upper half of the ear five days a f e r anthesis on dry weight at maturity of the various plant parts. (Means of all nitrogen
treatments)
Per culm D.wt per grain (mg) No. grains Grain d.wt (9) Chaff d.wt (9) Shoot d.wt (g) Total d.wt (g)
Lower half ear D.wt per grain (me) No. grains Grain d.wt (g) Chaff d.wt (9)
Hobbit & Intact Halved
41.7 45.4 34.7 19.6
1 *44 0.88 0.34 0.27 0.88 1-09 2.66 2.28
43.0 45.4 18.1 19.6 0.77 0.88 0.19 0.27
Maris Huntsman - Intact Halved
43.1 41.7 32.6 16.0
1.41 0.67 0.35 0.23 1.07 1.26 2.85 2.16
44.9 41.8 17.3 16.0 0.78 0.66 0.19 0.23
S.E.D.
0.97 1.50 0.072 0.016 0.058 0.138
0.91 1.02 0,047 0.012

Control of grain size in winter wheat 389
80 =‘ P v 5 6 0 .
4 0 ’
- 0 >
per grain and total grain dry weight in the remaining half of ears of Hobbit and decreased them in Maris Huntsman (Table 3). The number of grains in the lower half of the ear was not altered. Halving increased the weight of the remaining chaff more in Hobbit than in Maris Huntsman and increased shoot dry weight similarly in the two varieties.
Twenty days after anthesis the intact ears of Hobbit weighed more than those of Maris Huntsman, but the difference was not significant at the 28th day and disappeared by maturity (Fig. 4). The number of grains per ear was similar in the two varieties (Table 3). There were no significant differences between varieties in dry weight per grain, grain dry weight per ear, or dry weight of the chaff in intact ears at maturity (Table 3). The shoot of these ears weighed more in the taller Maris Huntsman than in Hobbit, but total dry weight per culm varied little between varieties.
’
6
1977 experiment The volume, fresh weight and water content of the selected grains attained their maximum
values between the 29th and the 33rd day after anthesis (Fig. 5). Grain dry weight at that time was about 75% of the final value, reached at 66 days after anthesis. After the 36th day, the smooth ontogenetic trends in grain volume, fresh weight and water content were obscured by fluctuations associated with rain. After the 50th day the rain caused some lodging and sprouting. These factors slightly decreased the dry weight at maturity of the selected grains in ears halved early. They made sampling difficult and caused considerable variation in total grain weight per ear late in grain growth, which data are therefore not presented.
Halving the ear five days after anthesis increased the maximum volume, fresh weight and water content of the selected grains, but not their dry weight (Fig. 5). There was no consistent
loo I
20 L 3 0 L d
- - Days after anthesis
0 20 40 60 0 20 40 60
Fig. 5 . 1977 experiment. Effect of removing the top half of the ear on (a) the volume, (b) fresh weight, (c) dry weight and (d) water content of the selected grains of Maris Huntsman during development. 0- - ~ 0 intact ears; t-0 ears halved five days after anthesis; A. . . * A ears halved 30 days after anthesis. Least significant differences are for samplings between 26 and 4 1 days after anthesis.

390 R . M A R T I N E Z - C A R R A S C O A N D G . N . T H O R N E
* \
b--_ --O
I 0.15- 0 20 40 60 0 20 40 60
Days after anthesis
Fig. 6. 1977 experiment. Effect of removing the top half of the ear on (a) the dry weight of the shoot (top less ear), and (b) the chaff (glurnes and rachis) of the lower half of the ear of Maris Huntsman during development. Symbols as in Fig. 5 . The two vertical bars represent least significant differences between treatments for samplings between 8 and 29, and between 33 and 62 days after anthesis.
increase in grain size after halving the ear when volume was maximal. Early removal of the upper half of the ear increased the number of endosperm cells 12 days after anthesis from 2 12 to
The dry weight of the chaff in the lower part of the ear was increased by early treatment, and the dry weight of the shoot after grain volume was maximal by both treatments (Fig. 6). The mean number of grains in the intact ears was 40.5 and in the halved ears 22.3.
239 x 103 (s.E.D. 10 x 103).
D I S C U S S I O N
The study made of growth in the main experiment showed- that nitrogen fertiliser had some unusual effects in 1976 that were attributed to the exceptional weather (Pearman et al., 1978). In particular, nitrogen did not increase the number of ears, and increased harvest index because it increased the number of grains per ear and grain weight per ear, whilst scarcely affecting the dry weight of the rest of the shoot. The increased production of carbohydrate resulting from the increase in leaf area caused by additional nitrogen did not increase dry weight per grain because there was a more than equivalent increase in number of grains.
Except in plants that received no nitrogen, thinning increased the number of grains per ear, presumably because conditions for ear growth before anthesis were improved. But grain dry weight per ear and total dry weight per shoot were increased little, and only with ample nitrogen, indicating that thinning did not make much more carbohydrate available for growth. The number of ears per unit land area was already low, and further decrease in competition for light was unlikely to affect photosynthesis significantly. Consequently, thinning did not increase the size of the grains. Thinning plots without nitrogen decreased grain size and grain dry weight per ear, possibly because the remaining shoots were deprived of the nitrogen that dying tillers would have supplied (Austin & Jones, 1975).
Although the two varieties had similar dry weight per ear at maturity, they differed in ear growth rate. The dry weight of the ear 20 days after anthesis was greater for Hobbit than for Maris Huntsman, but the latter grew faster thereafter. The number of grains per ear was similar in the two varieties (Table 3), so the difference in ear growth rate was due to mean dry weight per grain. Possibly the ears of Hobbit were able to attract larger amounts of assimilate at an early stage, as Austin, Edrich, Ford & Blackwell (1977) have found using 14C as a tracer. The increase in assimilate supply provided by halving in Hobbit increased the relative growth rate of these ears after the 20th day from anthesis to values higher than, or similar to, those of intact ears of Maris Huntsman (Table 4). So dry weight accumulation in intact ears of Hobbit may

Control of grain size in winter wheat 39 1
have declined late in their growth because of insufficient photosynthate. Early in growth, carbohydrates were probably stored in the stem of Maris Huntsman because of the incapacity of the ear to absorb them, and they would be transported to the ear when leaf area rapidly decreased at the end of the growth period; this could account for the faster growth rate of ears of Maris Huntsman. The late movement of assimilates from the stem to the ear of Maris Huntsman has been reported by Austin et al. (1977) and is also indicated by the sudden decrease in dry weight of the shoot after the 29th day in 1977 (Fig. 6). The dry weight loss was smaller when the ear was halved, presumably because the halved ears did not remove as much carbohydrates from the stem as did the intact ears. So grain size in Maris Huntsman seems to be more sink limited than that of Hobbit, which in turn seems relatively more source controlled at the end than at the beginning of grain filling. Comparisons of the two varieties have shown that the proportion of 14C supplied to leaves or to whole plants that is recovered in the ear or the grains is greater for Hobbit (TL 363/30) than for Maris Huntsman (Ruckenbauer, 1975; Austin et al., 1977; Makunga, Pearman, Thomas & Thorne, 1978). It was suggested that this was mostly due to differences between varieties in mean numbers of grains per ear (Makunga et al., 1978); our results indicate that there may also be a larger storage capacity in individual grains of Hobbit.
Table 4. 1976 Experiment. Relative growth rates (mg g-' d-I) of intact and halved ears of the varieties Hobbit and Maris Huntsman after the 20th day from anthesis
Days after anthesis . . . 20-28 28-39 ** Intact Halved Intact Halved
Hobbit 32.5 65.0 7.2 13.4 Maris Huntsman 57.5 53.7 14.3 16.7
In contrast to thinning and nitrogen application, removing the upper half of the ear soon after anthesis did not increase the number of grains in the remaining half of the ear, so that assimilate supply per grain should have increased, unless the rate of photosynthesis was decreased by a feed-back mechanism (King, Wardlaw & Evans, 1967). The increase in dry weight of the rest of the shoot after halving indicated that photosynthate continued to be available, as has been found in other experiments where all or some of the grains in the ear have been removed (Nosberger & Thorne, 1965; Lupton, 1968; Austin & Edrich, 1975). Bingham (1966) has shown that the response of grain size to assimilate supply varies with the genotype, and the different effect of halving on grain size in the two varieties is consistent with that finding. Hobbit responded positively to treatment, whereas in Maris Huntsman spikelet removal did not increase the dry weight per grain in 1977 and decreased it in 1976. Pinthus & Millet (1978) and Fischer & HilleRisLambers ( 1978) have observed differences between varieties in response to spikelet removal. Pinthus & Millet (1978). Bremner & Rawson (1978) and Lupton & Ali (1966) have reported examples of decreases in size of the remaining grains following removal of complete spikelets or single florets. The causes of such decreases and of the varietal differences in response to spikelet removal are obscure. The increase in grain size of Hobbit caused by halving supports the conclusion reached above that the supply of carbohydrate controlled dry weight per grain in this variety. But the increase in dry weight per grain amounted to only 4%. There- fore the capacity of the grain to accept assimilates also limited its dry weight. The sink capacity of the grain seemed to be the only factor limiting early and late grain growth and final grain weight in Maris Huntsman. Brocklehurst (1977) reported an increase in dry weight per grain in Maris Huntsman after removing all but the basal florets of the four central spikelets at anthesis. However, the grains increased in weight much less than those of another variety, and there was only a small increase when floret number was reduced 15 days after anthesis. The

392 R . M A R T I N E Z - C A R R A S C O A N D G . N . T H O R N E
number of ears per unit land area in 1976 was similar to that of Brocklehurst (1977), and in 1977 the crop was much denser than in the previous year. The number of grains per ear was similar in our 1977 experiment to that in the experiments of Brocklehurst, Moss & Williams (1978), in which halving the irradiance during the endosperm cell division phase decreased final dry weight per grain in Maris Huntsman. So, the lack of effect of halving on final dry weight per grain in Maris Huntsman is not likely to be due to less competition between grains or ears in our experiments, than in those of Brocklehurst (1977) and Brocklehurst et al. (1978). Interaction between grains within or between spikelets (Bremner & Rawson, 1978), or differences in time of degraining, could account for the difference between Brocklehurst’s results and ours. The increases in maximum volume after halving the ears of Hobbit in 1976 and Maris Huntsman in 1977 suggest that the supply of assimilates available to the grain, while this is expanding, regulated endosperm development and the volume of the grain. The grains from halved ears of Maris Huntsman in 1977 had more endosperm cells, so the larger endosperm size was due to increased cell division, in accord with the results of Brocklehurst (1977). But there was no corresponding increase in grain dry weight in Maris Huntsman, and dry weight increased relatively less than volume in Hobbit. The decrease in volume up to maturity was greater in halved than intact ears, as found previously (Martinez-Carrasco & Thorne, 1979). Thus, if potential grain size is measured by endosperm cell development and maximum grain volume, the gap between potential and actual size was relatively greater in halved than intact ears, even though more carbohydrate was available. Possibly some factor involved in the synthesis and deposition of starch limits the response of grain size to assimilate supply.
We thank the International Bank for Reconstruction and Development and the Consejo Superior de Investigaciones Cientificas for Fellowships to R. M.-C. during 1976 and 1977 respectively. We also thank D. J. Dunstan and M. Snow for help with measurements of growth.
R E F E R E N C E S
AUSTIN, R. B. & EDRICH, J. A. ( 1 9 7 5 ) . Effects of ear removal on photosynthesis, carbohydrate accumulation and on the distribution of assimilated 14C in wheat. Annals of Botany 39, 1 4 1 - 1 5 2 .
AUSTIN, R. B. JONES, H. G. ( 1 9 7 5 ) . The physiology of wheat. Plant Breeding Institute Annual Report for 1974,20-73.
AUSTIN, R. B., EDRICH, J. A., FORD, M. A. & BLACKWELL, R. D. ( 1 9 7 7 ) . The fate of the dry matter, carbohydrate and 14C lost from the leaves and stems of wheat during grain filling. Annals of Botanv 41, 1 3 0 9 - 1 3 2 1 .
BINGHAM, J. ( 1 9 6 6 ) . Paternal effect on grain size in wheat. Nature, London 209, 940-94 1. BINGHAM, J . ( 1 9 6 7 ) . Investigtitions on the physiology of yield in winter wheat, by comparison of varieties
and by artificial variation in grain number per ear. Journal of Agricultural Science, Cambridge 68,
BINGHAM, J. (1 969). The physiological determinants of grain yield in cereals. Agricultural Progress 44, 30-42.
BREMNER, P. M. & RAWSON, H. M. ( 1 0 7 8 ) . The weights of individual grains of the wheat ear in relation to their growth potential, the supply of assimilate and interaction between grains. Australian Journal of Plant Physiology 5 , 6 1 - 7 2 .
4 1 1-422.
BROCKLEHURST, P. A. ( 1 9 7 7 ) . Factors controlling grain weight in wheat. Nature, London 266, 348-349. BROCKLEHURST, P. A., MOSS, J. P. & WILLIAMS, w. ( 1 9 7 8 ) . Effects of irradiance and water supply on grain
development in wheat. Annals of Applied Biology 90,265-276. FISCHER, R. A. & HILLERISLAMBERS, D. ( 1 9 7 8 ) . Effect of environment and cultivar on source limitation
to grain weight in wheat. Australian Journal of Agricultural Research 29,443-458. FRIEND, D. J. c., FISHER, J. E. & HEISON, v. A. ( 1 9 6 3 ) . The effect of light intensity and temperature on
floral initiation and inflorescence development of Marquis wheat. Canadian Journal of Botany 41, 1 6 6 3 - 1 674.

Control of grain size in winter wheat 393
KING, R. w., WARDLAW, I. F. EVANS, L. T. (1967). Effect of assimilate utilisation on photosynthetic rate in wheat. Planta 77, 261-276.
LUPTON, F. G. H. ( 1 9 6 8 ) . The analysis of grain yield of wheat in terms of photosynthetic ability and efficiency of translocation. Aiznals ofApplied Biology 61, 109-1 1 9 .
LUPTON, F. G. H. L ALI, M. A. M. (1966) . Studies on photosynthesis in the ear of wheat. Annals of Applied Biology 5 7 , 2 8 1-286.
MAKUNGA, 0. H. D., PEARMAN, I., THOMAS, s. M. & THORNE, G. N. (1978) . Distribution of photosynthate produced before and after anthesis in tall and semi-dwarf winter wheat, as affected by nitrogen fertiliser. Annals of Applied Biology 8 8 , 4 2 9 - 4 3 7 .
MARTINEZ-CARRASCO, R. ~r THORNE, G . N. ( 1 9 7 9 ) . Physiological factors limiting grain size in wheat. Journal of Experimental Botany (in press).
N ~ S B E R G E R , J. & THORNE, G . N. (1965). The effect of removing florets or shading the ear of barley on production and distribution of dry matter. Annals of Botany 29, 635-644.
PEARMAN, I., THOMAS, s. M. & THORNE, G. N. ( 1 9 7 7 ) . Effects of nitrogen fertiliser on growth and yield of spring wheat. Annals ofBotany 4 1 , 9 3 - 1 0 8 .
PEARMAN, I., THOMAS, s. M. g, THORNE, G. N. (1978). Effect of nitrogen fertiliser on growth and yield of semi-dwarf and tall varieties of winter wheat. Journal of Agricultural Science, Cambridge 91, 3 1-45.
PINTHUS, M. J. 6t MILLET, E. (1978). Interactions among number of spikelets, number of grains and grain weight in the spikes of wheat (Triticum aestivum L.). Annals of Botany 4 2 , 8 3 9 - 8 4 8 .
RUCKENBAUER, P. (1975). Photosynthetic and translocation pattern in contrasting winter wheat varieties. Annals ofApplied Biology 79, 35 1-359.
SOFIELD, I., EVANS, L. T., COOK, M. G. & WARDLAW, I. w. ( 1 9 7 7 ) . Factors influencing the rate and duration of grain filling in wheat. Australian Journal of Plant Physiology 4 , 1 8 5 - 7 9 8 .
SPIERTZ, J. H. J. ( 1 9 7 4 ) . Grain growth and distribution of dry matter in the wheat plant as influenced by temperature, light energy and ear size. Netherland Journal of Agricultural Science 2 2 , 2 0 7 - 2 2 0 .
THORNE, G. N. ( 1 9 7 1 ) . Physiological factors limiting yield of arable crops. In Potential Crop Production. (eds. J. P. Cooper and P. F. Wareing), 387 pp. London: Heinemann.
WATSON, D. J . (1971). Size, structure and activity of the productive system of crops. In Potential Crop Production, (eds. J. P. Cooper and P. F. Wareing), 387 pp. London: Heinemann.
WELBANK, P. J., WITTS, K. J. & THORNE, G . N. (1968). Effect of radiation and temperature on efficiency of cereal leaves during grain growth. Annals of Botany 32, 79-95 .
(Received 18 January 1979)




















