
Effect of hydrogen peroxide on catalase gene expression, isoform activities and levels in leaves of...
14
CSIRO PUBLISHING www.publish.csiro.au/journals/fpb Functional Plant Biology, 2005, 32, 707–720 Effect of hydrogen peroxide on catalase gene expression, isoform activities and levels in leaves of potato sprayed with homobrassinolide and ultrastructural changes in mesophyll cells Jos´ e M. Almeida A,B , Fernanda Fidalgo A,B , Ana Confraria A,B , Arlete Santos A,B , Helena Pires A and Isabel Santos A,B,C A Institute for Molecular and Cellular Biology, University of Porto, Rua do Campo Alegre, 823, 4150-180 Porto, Portugal. B Botany Department, School of Sciences, University of Porto, Rua do Campo Alegre, 1191, 4150-180 Porto, Portugal. C Corresponding author. Email: [email protected] Abstract. The effect of hydrogen peroxide (H 2 O 2 ) on catalase (CAT) isoform activities and amounts and on mRNA levels was studied in leaves from potato plants untreated and treated with homobrassinolide (HBR). Northern blot analysis revealed that 100 mM H 2 O 2 supplied through the leaf petiole for 4 h did not induce CAT expression. In contrast, CAT1 and CAT2 responded differently to longer treatment, as CAT2 transcript levels increased markedly whereas CAT1 transcript levels remained unchanged. Western blot analysis showed disparity between the level of CAT1 transcript and CAT1 amount, which actually decreased after 28 h. CAT2 amount correlated well with transcript accumulation and CAT2 activity as visualised by zymogram analysis. H 2 O 2 modified the relative importance of CAT isoforms. After 4 h, CAT1 was prevalent in untreated and H 2 O 2 -treated leaves. After 28 h, CAT2 was prevalent in H 2 O 2 -treated leaves; therefore, the quantified increase in total CAT activity in these leaves was due to the rise in CAT2. HBR pre-treatment increased CAT2 basal level not changing the pattern of CAT responses to H 2 O 2 , only lowering its amplitude. Even so, ultrastructural studies showed that HBR significantly reduced H 2 O 2 negative effects on cellular sub-structures, allowing better recovery of affected structures and reducing the macroscopic injury symptoms on leaves, thus data point to a HBR protective role. Keywords: brassinosteroids, catalase gene expression, catalase isozymes, hydrogen peroxide, oxidative stress. Introduction Plants produce reactive oxygen species (ROS) as by- products of normal cellular metabolism. However, several unfavourable environmental conditions increase the production and accumulation of ROS, which are potentially harmful for all cellular components. Deleterious effects of ROS include damage to proteins and DNA, and lipid peroxidation. Fortunately, to prevent the cytotoxic effects of ROS, plant cells possess enzymatic and non- enzymatic protective mechanisms that can scavenge ROS, keeping their levels under control and maintaining cellular redox balance. H 2 O 2 is one of the ROS produced in plants under both biotic and abiotic stresses. Catalase is one of the Abbreviations used: BRs, brassinosteroids; CAT, catalase; Et, ethanol; GST, glutathione S-transferase; HBR, homobrassinolide; ROS, reactive oxygen species; Wt, deionised water. main H 2 O 2 -scavenging enzymes in plants, and consequently plays a crucial role in plant defence. Higher plants have three main CAT isoforms that are distinguished on the basis of organ specificity and responses to stresses (Willekens et al. 1994a, b, c). CAT1 is highly expressed in photosynthetic tissues and is specifically involved in the scavenging of photorespiratory H 2 O 2 ; CAT2 is preferentially expressed in vascular tissues and its physiological roles are still unclear; CAT3 is abundant in seeds and young plants and is correlated with glyoxysomal activity (Willekens et al. 1994a, 1997). CAT converts H 2 O 2 to H 2 O and O 2 , without requiring reducing power and, although it shows poor affinity for H 2 O 2 , it has an extremely high reaction rate, providing the cell with a very energy-efficient © CSIRO 2005 10.1071/FP04235 1445-4408/05/080707
Transcript of Effect of hydrogen peroxide on catalase gene expression, isoform activities and levels in leaves of...

CSIRO PUBLISHING
www.publish.csiro.au/journals/fpb Functional Plant Biology, 2005, 32, 707–720
Effect of hydrogen peroxide on catalase gene expression, isoform activitiesand levels in leaves of potato sprayed with homobrassinolide
and ultrastructural changes in mesophyll cells
Jose M. AlmeidaA,B, Fernanda FidalgoA,B, Ana ConfrariaA,B, Arlete SantosA,B,Helena PiresA and Isabel SantosA,B,C
AInstitute for Molecular and Cellular Biology, University of Porto, Rua do Campo Alegre, 823,4150-180 Porto, Portugal.
BBotany Department, School of Sciences, University of Porto, Rua do Campo Alegre, 1191,4150-180 Porto, Portugal.
CCorresponding author. Email: [email protected]
Abstract. The effect of hydrogen peroxide (H2O2) on catalase (CAT) isoform activities and amounts and on mRNAlevels was studied in leaves from potato plants untreated and treated with homobrassinolide (HBR). Northern blotanalysis revealed that 100 mM H2O2 supplied through the leaf petiole for 4 h did not induce CAT expression.In contrast, CAT1 and CAT2 responded differently to longer treatment, as CAT2 transcript levels increased markedlywhereas CAT1 transcript levels remained unchanged. Western blot analysis showed disparity between the level ofCAT1 transcript and CAT1 amount, which actually decreased after 28 h. CAT2 amount correlated well with transcriptaccumulation and CAT2 activity as visualised by zymogram analysis. H2O2 modified the relative importance ofCAT isoforms. After 4 h, CAT1 was prevalent in untreated and H2O2-treated leaves. After 28 h, CAT2 was prevalentin H2O2-treated leaves; therefore, the quantified increase in total CAT activity in these leaves was due to the risein CAT2. HBR pre-treatment increased CAT2 basal level not changing the pattern of CAT responses to H2O2,only lowering its amplitude. Even so, ultrastructural studies showed that HBR significantly reduced H2O2 negativeeffects on cellular sub-structures, allowing better recovery of affected structures and reducing the macroscopicinjury symptoms on leaves, thus data point to a HBR protective role.
Keywords: brassinosteroids, catalase gene expression, catalase isozymes, hydrogen peroxide, oxidative stress.
Introduction
Plants produce reactive oxygen species (ROS) as by-products of normal cellular metabolism. However, severalunfavourable environmental conditions increase theproduction and accumulation of ROS, which are potentiallyharmful for all cellular components. Deleterious effectsof ROS include damage to proteins and DNA, andlipid peroxidation. Fortunately, to prevent the cytotoxiceffects of ROS, plant cells possess enzymatic and non-enzymatic protective mechanisms that can scavenge ROS,keeping their levels under control and maintaining cellularredox balance.
H2O2 is one of the ROS produced in plants underboth biotic and abiotic stresses. Catalase is one of the
Abbreviations used: BRs, brassinosteroids; CAT, catalase; Et, ethanol; GST, glutathione S-transferase; HBR, homobrassinolide; ROS, reactive oxygenspecies; Wt, deionised water.
main H2O2-scavenging enzymes in plants, and consequentlyplays a crucial role in plant defence. Higher plantshave three main CAT isoforms that are distinguishedon the basis of organ specificity and responses to stresses(Willekens et al. 1994a, b, c). CAT1 is highly expressedin photosynthetic tissues and is specifically involved in thescavenging of photorespiratory H2O2; CAT2 is preferentiallyexpressed in vascular tissues and its physiological rolesare still unclear; CAT3 is abundant in seeds and youngplants and is correlated with glyoxysomal activity (Willekenset al. 1994a, 1997). CAT converts H2O2 to H2O andO2, without requiring reducing power and, althoughit shows poor affinity for H2O2, it has an extremely highreaction rate, providing the cell with a very energy-efficient
© CSIRO 2005 10.1071/FP04235 1445-4408/05/080707

708 Functional Plant Biology J. M. Almeida et al.
mechanism to remove H2O2 (Scandalios et al. 1997). CAT isimportant because of this direct antioxidant ability and alsobecause of its ability to affect signal transduction pathwaysthat entail H2O2 as signal (Willekens et al. 1994a). H2O2 hasbeen the focus of much research, and several studies havebeen shown that, aside from the destructive effects of H2O2,plants can also use this molecule in a beneficial way (Corpaset al. 1999). Indeed, H2O2 can have a dual function dependingon its cellular concentration (Dat et al. 2000). Recently,a transcriptome analysis using catalase-deficient tobaccoplants (CAT1AS) confirmed the dual function of H2O2,which induced defence responses at low concentrationsbut caused cell death at high levels (Dat et al. 2003;Vandenabeele et al. 2003).
H2O2 acts as a signalling molecule, mediating plantresponses to biotic and abiotic stresses as well as to planthormones and gravity (Neill et al. 2002a), and it was recentlydemonstrated that H2O2 is a common signal mediating bothdark- and ABA-induced stomatal closure in pea (Desikanet al. 2004).
There are several reports describing the protectiverole of H2O2 in plants under different stress situations(Van Breusegem et al. 2001; Neill et al. 2002a, b). Maizecoleoptiles treated with H2O2 are protected from subsequentchilling stress (Prasad et al. 1994a, b). Thermotolerancein potato microplants was induced by H2O2 or byacetylsalicylic acid treatment (Lopez-Delgado et al. 1998),and in mustard seedlings thermotolerance induced bysalicylic acid or heat acclimation was correlated with anincrease in H2O2 (Dat et al. 1998). H2O2 treatment ofArabidopsis leaves induced protection against a subsequenthigh light stress (Karpinski et al. 1999). Gechev et al. (2002)reported that spraying tobacco plants with H2O2 inducesprotection against oxidative stress through the enhancementof the activities of a set of antioxidant enzymes, amongstthem catalase. Moreover, several studies have shown thatH2O2 modulates gene expression during defence responses.In soybean, H2O2 induced the expression of genes encodingglutathione S-transferase (GST) and glutathione peroxidase(Levine et al. 1994) and in Arabidopsis thaliana (L.) Heynh.suspension cultures, H2O2 induced the expression of GSTand genes encoding phenylalanine ammonia-lyase (Desikanet al. 1998). The expression of genes encoding ascorbateperoxidase in germinating rice embryos (Morita et al. 1999)and in leaves of Arabidopsis was induced by H2O2 (Karpinskiet al. 1999). A direct signalling action of H2O2 in theregulation of expression of the three CAT genes and GST1gene of maize was demonstrated by Polidoros and Scandalios(1999). In maize, further studies showed induction of CAT1expression, which was related with the production of H2O2in response to wounding (Guan and Scandalios 2000). Toour knowledge, the effect of H2O2 on the CAT system hasbeen the subject of very few studies, and the evaluationshave only been conducted by following changes in total
catalase activity (Gechev et al. 2002). Indeed, the effect onCAT isoforms has not been studied to date. Additionally,the regulatory role of H2O2 on the expression of CATgenes has been studied only in maize, a monocot with a C4photosynthetic pathway.
Several studies have shown that brassinosteroids (BRs) areinvolved in protecting plants from a wide spectrum of stressessuch as low and high temperatures, drought, high salt andpathogen attack (references in Mussig and Altmann 1999;Castle et al. 2003; Krishna 2003). Despite this, only fewstudies aimed at understanding the mechanism by which BRspromote stress resistance have been undertaken (see Mussigand Altmann 1999; Krishna 2003). Studies aiming tounderstand the BRs signal transduction pathway and BRsgene-regulating properties indicate that there is cross-talkbetween BRs and abscisic acid, jasmonic acid and ethylene,hormones with established roles in plant defence responses(see Krishna 2003). Recent studies revealed that the BRreceptor BRI1 in tomato perceives both BRs and the systemin,a molecule that mediates systemic responses to woundingby insect pests (see Wang and He 2004). This informationraised the possibility of interaction among the BR-signalling,defence response and systemin-signalling pathways. Studiesin tobacco and rice showed that BRs enhance resistanceto a broad range of diseases independently of salicylicacid, without cross-talk between systemic acquired resistanceand wound-inducible disease resistance (Nakashita et al.2003). The authors speculate that the resistance mechanismsmediated by BRs are distinct from other known mechanisms,suggesting that brassinosteroid-mediated disease resistancefunctions in the innate immunity system of higher plants.Further studies are needed to gain a deeper insight intothe biochemical and molecular mechanisms underlying theprotective ability of BRs.
The influence of BRs in the response of the antioxidantenzymes of plants under stress conditions has been studiedrecently. Data available show that the changes inducedin the activity of antioxidant enzymes by BRs, differedwith plant species and with stress conditions (Mazorraet al. 2002; Nunez et al. 2003, 2004; Vardhini and Rao2003; Ozdemir et al. 2004). However, in all the citedstudies the effect of BRs on CAT was evaluated throughchanges in total activity as the response of CAT isoformswas unknown.
Bearing in mind the reports referring the protective roleof both H2O2 and BRs against stresses and the importantrole of CAT in plant defence, the purpose of our workwas to examine the effects of H2O2 on the activity andamount of CAT isoforms and on CAT genes expression inpotato, a dicotyledonous with C3 photosynthesis, to findout whether this oxidant acts at a transcription or post-transcription level and to know how CAT isoforms respondto H2O2. A further aim was to examine whether the pre-treatment of potato plants with a brassinosteroid affected

H2O2 and catalase gene expression of potato leaves Functional Plant Biology 709
the way CAT isoforms respond to H2O2. To this end, theultrastructural changes in mesophyll cells induced by H2O2in both untreated and homobrassinolide-treated leaves werestudied to better understand the magnitude of the response ofpotato leaves to H2O2.
Materials and methodsPlant material
Potato plants (Solanum tuberosum L. cv. Desiree) were grown in potscontaining soil : turf mixture (1 : 1, v / v), under field conditions. A veryhomogeneous emergence of the plants was observed, and 9 d afteremergence, plants with a similar vegetative growth were divided intothree sets. One set was sprayed with 1 µM 22(S),23(S)-homobrassinolide(HBR) in 10% (v / v) ethanol, and the other two sets were used ascontrols; one sprayed with the solvent of the HBR (10% ethanol, Et), andthe other sprayed with deionised water (Wt), the latter being the controlfor both HBR and Et plants. After 3 d, all sets of plants were sprayedagain with the respective solution. 100 mL of each spraying solutionwas uniformly applied over the three plants in each pot, assuring that allleaves were well moistened.
Treatment of leaves with H2 O2
Fully expanded leaves at the fourth leaf node from the apex wereharvested from each plant of each group of plants (HBR-, Et- andWt-sprayed plants) 6 d after the second spraying. H2O2 (100 mM) wassupplied to detached leaves from these three groups of plants through cutpetioles. Deionised water was supplied to equivalent groups of leavesand used as controls.
The six sets of leaves were maintained in a growth chamber under60 µmol m−2 s−1 photosynthetically active radiation (PAR). Terminalleaflets were collected 4 and 28 h after the beginning of H2O2 treatment,immediately frozen in liquid nitrogen and stored at –80◦C until requiredfor further molecular and biochemical studies. To minimise plant-to-plant variation, at least four terminal leaflets were sampled andpooled. The biochemical results presented represent the average overthree independent experiments. Leaf samples were also prepared forultrastructural studies.
Catalase extraction and activity assay
Catalase (CAT; EC 1.11.1.6) extraction was basically performed aspreviously described (Santos et al. 2004) with the extraction mediumsupplemented with 5 mM L (+)-ascorbic acid (Merck, WhitehorseStation, NJ). After the supernatant had been conditioned withDTT and glycerol (Santos et al. 2004), CAT activity was assayedspectrophotometrically by monitoring the decrease in absorbance at240 nm due to H2O2 consumption, at room temperature according toRao et al. (1996). One unit of CAT activity was defined as the amountof enzyme that decomposes 1 µmol H2O2 min−1 at 25◦C.
Catalase staining activity and western blotting
Catalase isozymes were separated on discontinuous PAGE under non-denaturing conditions, as described previously (Santos et al. 2004).Two gels were performed for each independent experiment and therepresentative results from six gels obtained with material from thethree experiments are presented.
For western blotting, the gels were incubated for 1 h in 187.5 mM
Tris–HCl (pH 8.8) with 0.3% (w / v) SDS and 5% (v / v) mercaptoethanolwith mild agitation, at room temperature. The proteins were thenelectrotransferred overnight (20 mA) to nitrocellulose membrane(0.45 µm pore; Schleicher and Schuell, Dassel, Germany) in a HoeferTE22 Mini Transphor apparatus (Hoefer Inc., San Francisco, CA). Thetransfer buffer was 50 mM Tris–HCl, 380 mM glycine (pH 8.3) with
20% (v / v) methanol and 0.1% (w / v) SDS. On the following day thecurrent was raised to 200 mA, for 1 h.
To immunodetect CAT, the membrane was blocked for 1 h in5% (w / v) non-fat milk powder in PBS-T (phosphate bufferedsaline–Tween 20; Sambrook et al. 1989) and then washed with1% (w / v) powdered milk in PBS-T with mild agitation, at roomtemperature. Incubation with primary antibodies against CAT2 or CAT1,in PBS-T with 0.3% (w / v) BSA, was conducted overnight at4◦C. Both polyclonal antibodies were raised in rabbit againsttwo peptides, selected as antigenic regions from the amino acidsequences deduced from Solanum tuberosum L. cDNAs for CAT2 (NCBIaccess no. AY500290) and for CAT1 (NCBI access no. AY442179),both isolated in our laboratory. The membrane was washed as previouslydescribed and incubated at room temperature for 1 h, with anti-rabbitIgG peroxidase conjugate (Sigma, St Louis, MO), diluted 1 : 1000 inPBS-T with 0.3% (w / v) BSA followed by several washes in PBS-T.Detection was made using an enhanced chemiluminescence method(Amersham Biosciences, Buckinghamshire, UK), and visualised witha photographic film (Ortho CP-G Plus AGFA, Mortsel, Belgium).Controls were prepared by substituting the primary antibody with thesame amount of pre-immune rabbit serum.
Densitometric analysis of the gels and of western blots wasperformed. Gels stained for CAT activity and western blot films werescanned with a computer-assisted GS-800 densitometer (Bio-Rad,Hercules, CA) and the CAT band density (the total intensity ofall the pixels in the volume divided by the area of the volume)quantified with the Quantity One Software (Bio-Rad). Densitometricreadings were performed using an equal area over each of all thebands resolved in the same gel and in the same blot film and localbackground subtracted.
RNA isolation and northern blot analysis
RNA from frozen leaf tissue was extracted with Trizol reagent accordingto the manufacturer’s recommendations (Invitrogen, Carlsbad, CA).RNA was quantified by gel electrophoresis with ethidium bromidestaining as well as through spectrophotometry. Total RNA (20 µg)from each sample was separated on a 1.5% (w / v) agarose gel,containing 0.42% (v / v) formaldehyde, 40% (w / v) Mops buffer (10×)and 66.6% (v / v) formamide, at 80 V, and blotted onto Hybond-N+
nylon filters (Amersham Biosciences) according to the method bySambrook et al. (1989). The RNA was fixed to the membrane byUV cross-linking. The probes used for transcript detection werea 294-bp fragment of a cDNA encoding CAT1 from potato (NCBIaccess no. AY442179) and a 307-bp fragment of a cDNA encodingCAT2 from potato (NCBI access no. AY500290) obtained by PCRusing specific primers for CAT1 (5′-CACTTATGAAATACGCAG-3′
and 5′-GCTTCACTCATTCATTCATTCA-3′), and for CAT2 (5′-GTGGTCAGAAGGTTGCTTCTCGTCT-3′ and 5′-GATGATGTTCTTCAAGCACCAAAGTG-3′). The template cDNAs were isolated inour laboratory by 3′ RACE using a primer with a poly-T adaptor,according to the method by Frohman et al. (1988) and a specificone for CAT1 (5′-GTCAAGTGCTTTCAATTCACCATTCTGT-3′)and another for CAT2 (5′-CCATGCATCCGTCTAAGTAT-3′).Hybridisation probes were generated from the gel-purified cDNAfragments by labelling with [α-32P] dCTP using random primingprocedures [Ready-To-Go DNA labelling beads (-dCTP) (AmershamBiosciences)]. Probes were purified with ProbeQuant G50 MicroColumns (Amersham Biosciences). Hybridisations were performedovernight at 65◦C in 1 M NaCl, 10% (w / v) dextran sulfate,1% (w / v) SDS and 50 µg mL−1 denatured salmon sperm DNA, afterprehybridisation, at 65◦C for 4 h, in the same solution without labelledprobe. Filters were washed twice for 15 min at 42◦C in 10% (v / v)SSPE (standard saline phosphate-EDTA; Sambrook et al. 1989)and 0.5% (w / v) SDS. A third wash was performed at room temperature

710 Functional Plant Biology J. M. Almeida et al.
in 10% (v / v) SSPE. Membranes were exposed to a MolecularDynamics (Sunnyvale, CA) storage phosphor screen and scanned withTyphoon 8600. The rRNA (5S) band was used as control for equalloading of RNA.
Electron microscopy and mesophyll cell ultrastructure observation
Samples from three terminal leaflets from each of the six sets of leavesunder study were collected. Lamina region from the middle of theleaflets was selected to obtain samples with the required size to beprepared for electron microscopy studies. Leaf pieces were fixed in2.5% (v / v) glutaraldehyde followed by 2% (w / v) osmium tetroxide,using NaOH–PIPES buffer (Salema and Brandao 1973), dehydrated inacetone and routinely embedded in Epon. To assure random pictures, foreach situation, ultra thin sections were obtained from different blocks,mounted in grids, contrasted with uranyl acetate and lead citrate andthen analysed with a Zeiss EM C10 transmission electron microscope(Zeiss, Gottingen, Germany).
Quantification of the cellular structural changes induced byH2O2 was performed. Morphometric determinations of fractionalvolume (Vv) of starch / chloroplast were performed by recordingthe number of hits of a 0.5 cm2 grid on the starch and on thechloroplasts, using for each set of leaves, at least, 10 micrographstaken at random and enlarged to the same final magnification(10 000×). The same micrographs were used for the quantification ofthe average number of damaged mitochondria, of chloroplasts withswelled thylakoids, and of peroxisomes with paracrystalline inclusions.The micrographs were digitalised and in each of them, six cell areasof 2.54 cm2 were randomly chosen and the area of the osmiophilicdeposits measured using the Adobe Acrobat 6.0 Professional software(Adobe, San Jose, CA).
Statistical analysis
Sample variability is given as the standard error of the mean (SE).The significance of differences was determined by the Student’stwo-tailed t-test for comparison of means, and P<0.05 wereconsidered significant.
Results
Total CAT activity and isoform pattern in native gels
Hydrogen peroxide supplied to detached leaves from HBR-,Et- and Wt-sprayed plants increased the extractable activityof CAT after 4 and 28 h of treatment. As shown in Table 1,after 4 h, the level of total CAT activity was significantlyhigher in control leaves from Wt-sprayed plants than incontrols of the other two sets of leaves, between which
Table 1. Effect of H2O2 treatment on CAT activity in potato leavesThe data are mean values ± SE (n = 6). Different superscript letters in the samecolumn indicate values that are significantly different according to Student’s t-testat P<0.05. Wt, water; Et, ethanol; HBR, homobrassinolide; Ø, control supplied
with deionised water
CAT activity (µmol H2O2 min−1 mg−1 protein)Plant set Treatment 4 h 28 h
Wt Ø 144.30 ± 2.45a 75.27 ± 1.78a
H2O2 159.64 ± 2.43b 121.95 ± 1.54b
Et Ø 134.57 ± 3.53ce 82.26 ± 2.01c
H2O2 149.37 ± 3.9d 113.52 ± 2.80d
HBR Ø 129.04 ± 1.49c 98.91 ± 2.21e
H2O2 136.78 ± 2.05e 112.88 ± 1.93d
CAT activity did not differ significantly. In comparisonwith the controls, H2O2 for 4 h significantly increased totalCAT activity in leaves from both Et- and Wt-sprayed plantssimilarly, with a rise by ∼11% and in leaves from HBR-sprayed plants, H2O2 only caused a rise of about 6% (Table 1).From 4 to 28 h, the CAT activity decreased in controlsas well as in H2O2-treated leaves. The highest decline(∼47%) occurred in controls from Wt-sprayed plants andthe lowest decrease (∼23%) in leaves from HBR-sprayedplants. In H2O2-treated leaves the decrease in CAT activitywas also lesser in leaves from HBR-sprayed plants (∼17%)than in the two other sets of leaves in which the decrease wassimilar. After 28 h, CAT activity continued to be significantlyhigher in all H2O2-treated leaves than in controls. However,owing to the decrease, from 4 to 28 h in CAT activity inboth controls and treated leaves, the difference in CATactivity between treated leaves and the respective controlsafter 28 h, become larger than after 4 h. In fact, after 28 h,CAT activity was ∼62, 38 and 14% higher in H2O2-treatedleaves from Wt- Et- or HBR-sprayed plants, respectively,when compared with the respective controls (Table 1). Theanalysis of CAT polymorphism in native gels showed thatH2O2 did not change the isozyme pattern, since two CATisoforms were detected in all studied situations. In gelsloaded with an equal amount of proteins extracted from allsets of leaves, after 4 h, the isoform with lower mobility(CAT1) was the prevalent one and appeared with higherstaining intensity, i.e., higher activity, than the isoform withhigher mobility (CAT2), which appeared as a very faint band(Fig. 1b). Densitometric analysis of CAT2 band densityrevealed that CAT2 activity in control leaves from HBR-sprayed plants was significantly higher than in control leavesfrom Wt- and Et-sprayed plants (Fig. 1a) and that H2O2caused a significant increase in the activity of this isoformin leaves from Wt- and HBR-sprayed plants. No significantdifferences were found among CAT1 density of thesix sets of leaves.
Although CAT isozyme pattern was the same in responseto H2O2 after 4 and 28 h, the relative abundance ofthe two isoforms was markedly different as is evidenced

H2O2 and catalase gene expression of potato leaves Functional Plant Biology 711
a
b
c
d
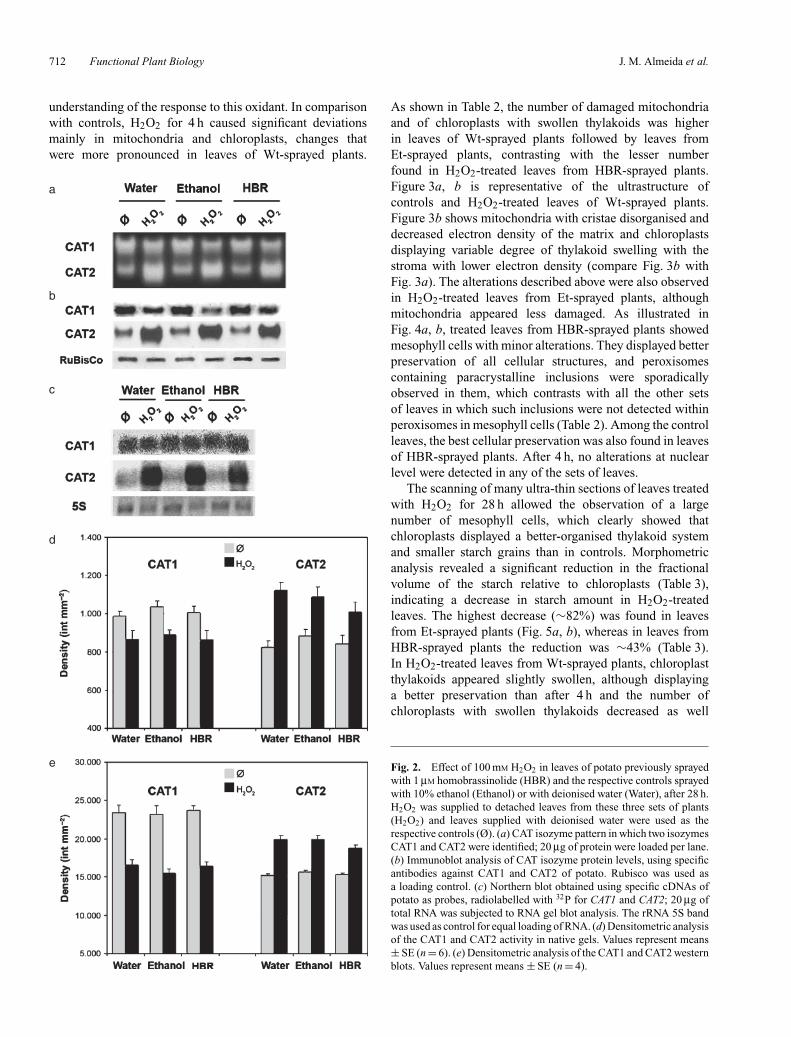
Fig. 1. Effect of 100 mM H2O2 in leaves of potato previously sprayedwith 1 µM homobrassinolide (HBR) and the respective controls sprayedwith 10% ethanol (Ethanol) or with deionised water (Water), after 4 h.H2O2 was supplied to detached leaves from these three sets of plants(H2O2) and leaves supplied with deionised water were used as therespective controls (Ø). (a) Densitometric analysis of CAT2 activityin native gels (I) and of the CAT2 western blot (II). Values representmeans ± SE (I, n = 6; II, n = 4). (b) CAT isozyme pattern in whichtwo isozymes CAT1 and CAT2 were identified; 20 µg of protein wereloaded per lane. (c) Immunoblot analysis of CAT isozyme protein levels,using specific antibodies against CAT1 and CAT2 of potato. Rubiscowas used as a loading control. (d) Northern blot obtained using specificcDNAs of potato as probes, radiolabelled with 32P for CAT1 and CAT2;20 µg of total RNA was subjected to RNA gel blot analysis. The rRNA5S band was used as control for equal loading of RNA.
by the comparison of the 4-h isozyme pattern in Fig. 1bwith the pattern at 28 h illustrated in Fig. 2a. After 28 h,a significant increase in CAT2 activity and a decrease inCAT1 were observed in response to H2O2. CAT2 was the
prevalent isoform in all H2O2 treated leaves, whereas inrespective controls, CAT1 was the isoform with highestactivity (Fig. 2a). Densitometric analysis (Fig. 2d) revealedthat in H2O2-treated leaves from Wt-sprayed plants theincrease in CAT2 activity was more pronounced than in thetwo other groups. The decrease in CAT1 activity in treatedleaves from Wt-sprayed plants was lower than in leaves fromboth Et- and HBR-sprayed plants (Fig. 1d). The CAT isozymepattern and the densitometric analysis showed that the higherlevel of total CAT activity, quantified by spectrophotometry,in H2O2-treated leaves was because of the increase inactivity of CAT2.
Immunoblot analysis
In parallel to the CAT activity data, western blot analysesrevealed that the level of the two CAT isoforms wasdifferently affected by H2O2. After 4 h, CAT1 was moreabundant than CAT2, a result in accordance with the activitypattern visualised in gels (Fig. 1b, c). Densitometric analysis(Fig. 1a) showed that the CAT2 amount in leaves fromHBR-sprayed plants was significantly higher than in leavesfrom Wt- and from Et-sprayed plants. H2O2 treatmentcaused a significant increase in CAT2 in leaves from bothWt- and HBR-sprayed plants. After 28 h, H2O2 causeda significant decrease in CAT1 amount, and increasedsignificantly the level of CAT2 compared with the controls(Fig. 2b). Densitometric analysis (Fig. 2e) of bands showed,in H2O2-treated leaves, a significant decrease in CAT1 andan increase in CAT2 that was more pronounced in leavesfrom Wt-sprayed plants. The results obtained with westernblot analysis are in accordance with the isozyme patterns(Fig. 2a, b, d, e).
Analysis of transcript levels
Northern blot analysis showed that CAT1 and CAT2expression was differently affected by H2O2, as alreadysuggested by isozyme pattern and western blot analyses.There was no induction of CAT expression within 4 h ofH2O2 treatment. At this time, transcript levels of CAT1 andCAT2 did not reveal significant differences between H2O2-treated leaves and respective controls (Fig. 1d). After 28 h,northern blot analysis showed that CAT1 and CAT2 respondto H2O2 in a different way (Fig. 2c). CAT1 expression wasnot affected and the level of transcripts appeared similarto that of 4 h. In contrast, H2O2 induced the expressionof CAT2, as illustrated in Fig. 2c; CAT2 mRNA level wasmarkedly enhanced in comparison to the controls. Northernblot analyses showed that the response of the isoforms toH2O2 was not affected by the previous plant spraying withHBR, Et or Wt.
Ultrastructural studies
The effect of H2O2 treatment on mesophyll cells wasevaluated at the ultrastructural level to gain a better
712 Functional Plant Biology J. M. Almeida et al.
understanding of the response to this oxidant. In comparisonwith controls, H2O2 for 4 h caused significant deviationsmainly in mitochondria and chloroplasts, changes thatwere more pronounced in leaves of Wt-sprayed plants.
a
b
c
d
e
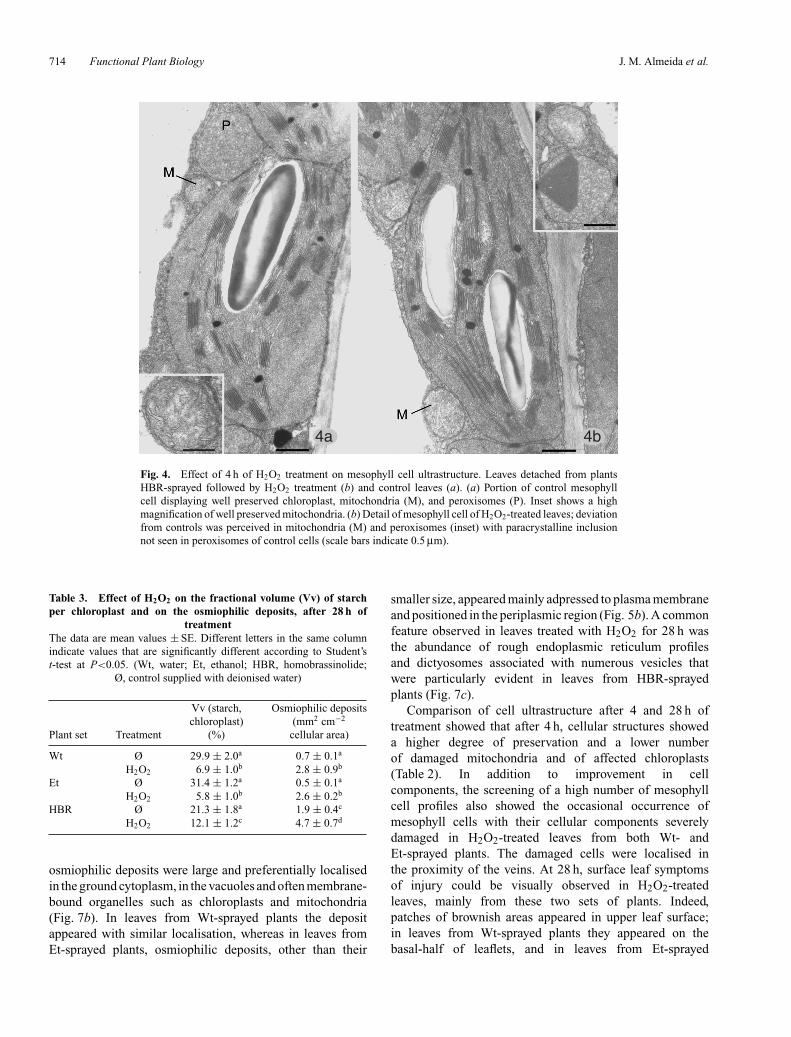
As shown in Table 2, the number of damaged mitochondriaand of chloroplasts with swollen thylakoids was higherin leaves of Wt-sprayed plants followed by leaves fromEt-sprayed plants, contrasting with the lesser numberfound in H2O2-treated leaves from HBR-sprayed plants.Figure 3a, b is representative of the ultrastructure ofcontrols and H2O2-treated leaves of Wt-sprayed plants.Figure 3b shows mitochondria with cristae disorganised anddecreased electron density of the matrix and chloroplastsdisplaying variable degree of thylakoid swelling with thestroma with lower electron density (compare Fig. 3b withFig. 3a). The alterations described above were also observedin H2O2-treated leaves from Et-sprayed plants, althoughmitochondria appeared less damaged. As illustrated inFig. 4a, b, treated leaves from HBR-sprayed plants showedmesophyll cells with minor alterations. They displayed betterpreservation of all cellular structures, and peroxisomescontaining paracrystalline inclusions were sporadicallyobserved in them, which contrasts with all the other setsof leaves in which such inclusions were not detected withinperoxisomes in mesophyll cells (Table 2). Among the controlleaves, the best cellular preservation was also found in leavesof HBR-sprayed plants. After 4 h, no alterations at nuclearlevel were detected in any of the sets of leaves.
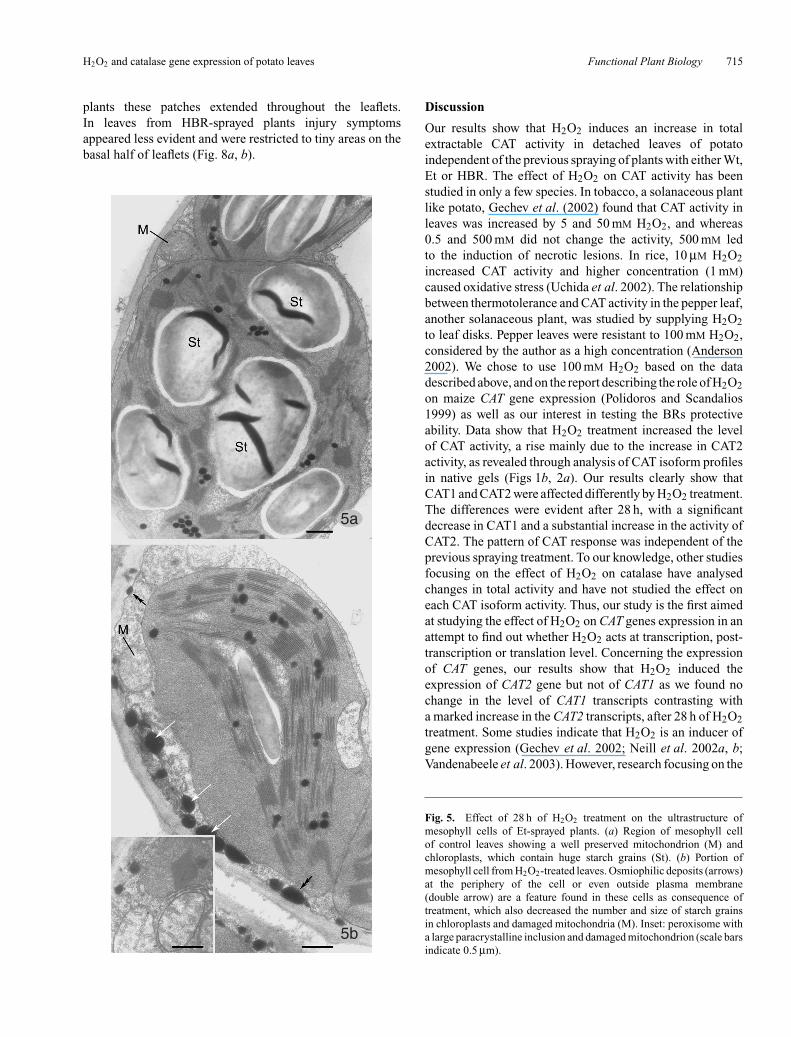
The scanning of many ultra-thin sections of leaves treatedwith H2O2 for 28 h allowed the observation of a largenumber of mesophyll cells, which clearly showed thatchloroplasts displayed a better-organised thylakoid systemand smaller starch grains than in controls. Morphometricanalysis revealed a significant reduction in the fractionalvolume of the starch relative to chloroplasts (Table 3),indicating a decrease in starch amount in H2O2-treatedleaves. The highest decrease (∼82%) was found in leavesfrom Et-sprayed plants (Fig. 5a, b), whereas in leaves fromHBR-sprayed plants the reduction was ∼43% (Table 3).In H2O2-treated leaves from Wt-sprayed plants, chloroplastthylakoids appeared slightly swollen, although displayinga better preservation than after 4 h and the number ofchloroplasts with swollen thylakoids decreased as well
Fig. 2. Effect of 100 mM H2O2 in leaves of potato previously sprayedwith 1 µM homobrassinolide (HBR) and the respective controls sprayedwith 10% ethanol (Ethanol) or with deionised water (Water), after 28 h.H2O2 was supplied to detached leaves from these three sets of plants(H2O2) and leaves supplied with deionised water were used as therespective controls (Ø). (a) CAT isozyme pattern in which two isozymesCAT1 and CAT2 were identified; 20 µg of protein were loaded per lane.(b) Immunoblot analysis of CAT isozyme protein levels, using specificantibodies against CAT1 and CAT2 of potato. Rubisco was used asa loading control. (c) Northern blot obtained using specific cDNAs ofpotato as probes, radiolabelled with 32P for CAT1 and CAT2; 20 µg oftotal RNA was subjected to RNA gel blot analysis. The rRNA 5S bandwas used as control for equal loading of RNA. (d) Densitometric analysisof the CAT1 and CAT2 activity in native gels. Values represent means± SE (n = 6). (e) Densitometric analysis of the CAT1 and CAT2 westernblots. Values represent means ± SE (n = 4).

H2O2 and catalase gene expression of potato leaves Functional Plant Biology 713
Table 2. Alterations induced by H2O2 at subcellular level after 4 and 28 h of treatmentMean values are expressed as percentage ± SE. Different superscript letters in the same column indicate values that are significantly different
according to Student’s t-test at P<0.05. (Wt, water; Et, ethanol; HBR, homobrassinolide; Ø, control supplied with deionised water)
Chloroplasts with Peroxisomes withDamaged mitochondria swollen thylakoids paracrystalline inclusion
Plant set Treatment 4 h 28 h 4 h 28 h 4 h 28 h
Wt Ø 0a 19.8 ± 8.1a 18.3 ± 6.9a 0a 0a 0a
H2O2 98.5 ± 1.5b 26.0 ± 9.6b 53.9 ± 7.9b 19.9 ± 5.9b 0a 66.7 ± 14.2b
Et Ø 13.9 ± 5.4c 0c 12.2 ± 2.2a 0a 0a 0a
H2O2 48.5 ± 12.4d 22.0 ± 9.8ab 20.0 ± 4.5a 0a 0a 62.5 ± 15.7b
HBR Ø 0a 0c 6.6 ± 2.4c 0a 0a 0a
H2O2 14.0 ± 4.4c 0c 7.5 ± 3.2c 0a 9.6 ± 1.1b 88.9 ± 11.1c
3a 3b
Fig. 3. Effect of 4 h of H2O2 treatment on mesophyll cell ultrastructure. Leaves detached from Wt-sprayedplants and then supplied with H2O2 (b) and control leaves (a). (a) Region of mesophyll cell of control leavesdisplaying well preserved peroxisomes (P), mitochondria (M) and chloroplast. (b) Portion of mesophyll cellof treated leaves, showing negative effects on mitochondria (M) cristae and matrix and on chloroplast stromaand thylakoids some of them appearing swollen (arrows). Peroxisomes (P) are devoid of paracrystallineinclusion in both controls and treated leaves (scale bars indicate 0.5 µm).
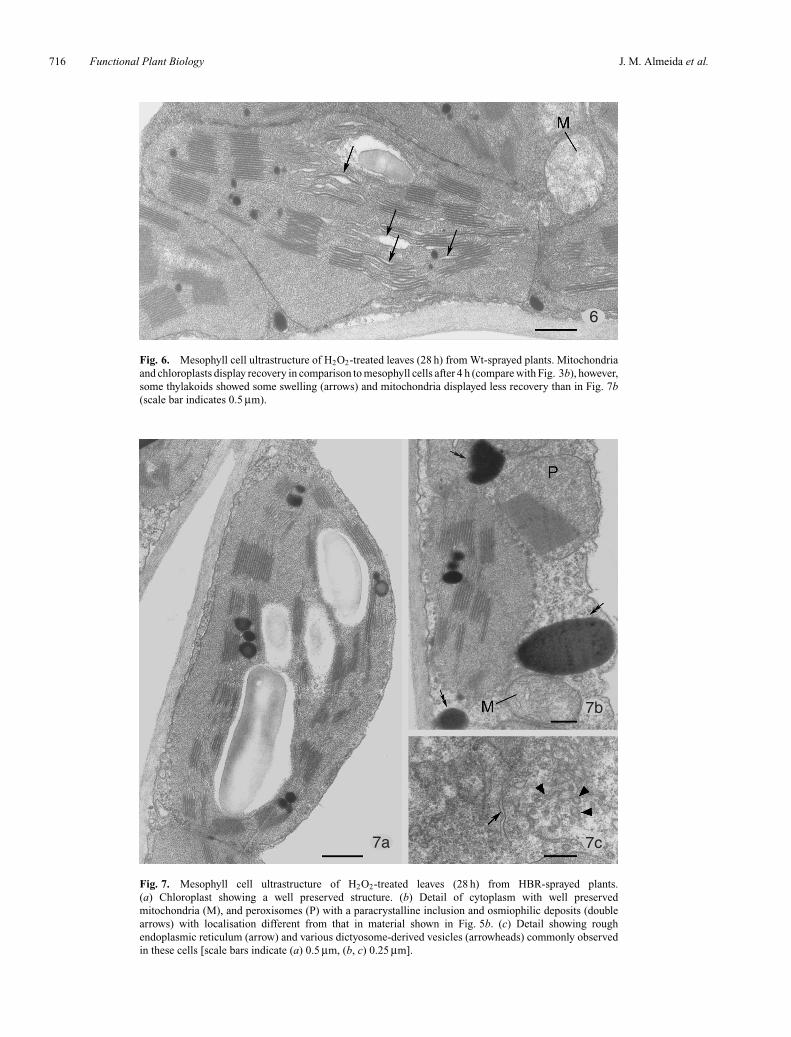
(compare Fig. 6 with Fig. 3b; Table 2). Mitochondria showeda better structural organisation in all leaves treated withhydrogen peroxide after 28 h than after 4 h, althoughthe best situation was found in leaves previously treatedwith HBR in which no damaged mitochondria were found(compare Figs 5b, 6, and 7; Table 2). After 28 h, in allH2O2-treated leaves, peroxisomes containing paracrystallineinclusions were observed in mesophyll cells, mainly inHBR-sprayed leaves (Fig. 7b), in which the number of
peroxisomes with the inclusion was significantly higher thanin the other two leaf sets (Table 2). In cells of all H2O2-treatedleaves, a significant increase in osmiophilic deposits wasobserved (Table 3). However, quantification of these depositsper cellular area revealed that control leaves from HBR-treated plants have significantly more osmiophilic depositsthan the other control leaves. After H2O2-treatment, thehighest value for osmiophilic deposits was also found inleaves from HBR-sprayed plants (Table 3). In these leaves, the
714 Functional Plant Biology J. M. Almeida et al.
4a 4b
Fig. 4. Effect of 4 h of H2O2 treatment on mesophyll cell ultrastructure. Leaves detached from plantsHBR-sprayed followed by H2O2 treatment (b) and control leaves (a). (a) Portion of control mesophyllcell displaying well preserved chloroplast, mitochondria (M), and peroxisomes (P). Inset shows a highmagnification of well preserved mitochondria. (b) Detail of mesophyll cell of H2O2-treated leaves; deviationfrom controls was perceived in mitochondria (M) and peroxisomes (inset) with paracrystalline inclusionnot seen in peroxisomes of control cells (scale bars indicate 0.5 µm).
Table 3. Effect of H2O2 on the fractional volume (Vv) of starchper chloroplast and on the osmiophilic deposits, after 28 h of
treatmentThe data are mean values ± SE. Different letters in the same columnindicate values that are significantly different according to Student’st-test at P<0.05. (Wt, water; Et, ethanol; HBR, homobrassinolide;
Ø, control supplied with deionised water)
Vv (starch, Osmiophilic depositschloroplast) (mm2 cm−2
Plant set Treatment (%) cellular area)
Wt Ø 29.9 ± 2.0a 0.7 ± 0.1a
H2O2 6.9 ± 1.0b 2.8 ± 0.9b
Et Ø 31.4 ± 1.2a 0.5 ± 0.1a
H2O2 5.8 ± 1.0b 2.6 ± 0.2b
HBR Ø 21.3 ± 1.8a 1.9 ± 0.4c
H2O2 12.1 ± 1.2c 4.7 ± 0.7d
osmiophilic deposits were large and preferentially localisedin the ground cytoplasm, in the vacuoles and often membrane-bound organelles such as chloroplasts and mitochondria(Fig. 7b). In leaves from Wt-sprayed plants the depositappeared with similar localisation, whereas in leaves fromEt-sprayed plants, osmiophilic deposits, other than their
smaller size, appeared mainly adpressed to plasma membraneand positioned in the periplasmic region (Fig. 5b). A commonfeature observed in leaves treated with H2O2 for 28 h wasthe abundance of rough endoplasmic reticulum profilesand dictyosomes associated with numerous vesicles thatwere particularly evident in leaves from HBR-sprayedplants (Fig. 7c).
Comparison of cell ultrastructure after 4 and 28 h oftreatment showed that after 4 h, cellular structures showeda higher degree of preservation and a lower numberof damaged mitochondria and of affected chloroplasts(Table 2). In addition to improvement in cellcomponents, the screening of a high number of mesophyllcell profiles also showed the occasional occurrence ofmesophyll cells with their cellular components severelydamaged in H2O2-treated leaves from both Wt- andEt-sprayed plants. The damaged cells were localised inthe proximity of the veins. At 28 h, surface leaf symptomsof injury could be visually observed in H2O2-treatedleaves, mainly from these two sets of plants. Indeed,patches of brownish areas appeared in upper leaf surface;in leaves from Wt-sprayed plants they appeared on thebasal-half of leaflets, and in leaves from Et-sprayed
H2O2 and catalase gene expression of potato leaves Functional Plant Biology 715
plants these patches extended throughout the leaflets.In leaves from HBR-sprayed plants injury symptomsappeared less evident and were restricted to tiny areas on thebasal half of leaflets (Fig. 8a, b).
5a
5b
Discussion
Our results show that H2O2 induces an increase in totalextractable CAT activity in detached leaves of potatoindependent of the previous spraying of plants with either Wt,Et or HBR. The effect of H2O2 on CAT activity has beenstudied in only a few species. In tobacco, a solanaceous plantlike potato, Gechev et al. (2002) found that CAT activity inleaves was increased by 5 and 50 mM H2O2, and whereas0.5 and 500 mM did not change the activity, 500 mM ledto the induction of necrotic lesions. In rice, 10 µM H2O2increased CAT activity and higher concentration (1 mM)caused oxidative stress (Uchida et al. 2002). The relationshipbetween thermotolerance and CAT activity in the pepper leaf,another solanaceous plant, was studied by supplying H2O2to leaf disks. Pepper leaves were resistant to 100 mM H2O2,considered by the author as a high concentration (Anderson2002). We chose to use 100 mM H2O2 based on the datadescribed above, and on the report describing the role of H2O2on maize CAT gene expression (Polidoros and Scandalios1999) as well as our interest in testing the BRs protectiveability. Data show that H2O2 treatment increased the levelof CAT activity, a rise mainly due to the increase in CAT2activity, as revealed through analysis of CAT isoform profilesin native gels (Figs 1b, 2a). Our results clearly show thatCAT1 and CAT2 were affected differently by H2O2 treatment.The differences were evident after 28 h, with a significantdecrease in CAT1 and a substantial increase in the activity ofCAT2. The pattern of CAT response was independent of theprevious spraying treatment. To our knowledge, other studiesfocusing on the effect of H2O2 on catalase have analysedchanges in total activity and have not studied the effect oneach CAT isoform activity. Thus, our study is the first aimedat studying the effect of H2O2 on CAT genes expression in anattempt to find out whether H2O2 acts at transcription, post-transcription or translation level. Concerning the expressionof CAT genes, our results show that H2O2 induced theexpression of CAT2 gene but not of CAT1 as we found nochange in the level of CAT1 transcripts contrasting witha marked increase in the CAT2 transcripts, after 28 h of H2O2treatment. Some studies indicate that H2O2 is an inducer ofgene expression (Gechev et al. 2002; Neill et al. 2002a, b;Vandenabeele et al. 2003). However, research focusing on the
Fig. 5. Effect of 28 h of H2O2 treatment on the ultrastructure ofmesophyll cells of Et-sprayed plants. (a) Region of mesophyll cellof control leaves showing a well preserved mitochondrion (M) andchloroplasts, which contain huge starch grains (St). (b) Portion ofmesophyll cell from H2O2-treated leaves. Osmiophilic deposits (arrows)at the periphery of the cell or even outside plasma membrane(double arrow) are a feature found in these cells as consequence oftreatment, which also decreased the number and size of starch grainsin chloroplasts and damaged mitochondria (M). Inset: peroxisome witha large paracrystalline inclusion and damaged mitochondrion (scale barsindicate 0.5 µm).
716 Functional Plant Biology J. M. Almeida et al.
6
Fig. 6. Mesophyll cell ultrastructure of H2O2-treated leaves (28 h) from Wt-sprayed plants. Mitochondriaand chloroplasts display recovery in comparison to mesophyll cells after 4 h (compare with Fig. 3b), however,some thylakoids showed some swelling (arrows) and mitochondria displayed less recovery than in Fig. 7b(scale bar indicates 0.5 µm).
7a
7b
7c
Fig. 7. Mesophyll cell ultrastructure of H2O2-treated leaves (28 h) from HBR-sprayed plants.(a) Chloroplast showing a well preserved structure. (b) Detail of cytoplasm with well preservedmitochondria (M), and peroxisomes (P) with a paracrystalline inclusion and osmiophilic deposits (doublearrows) with localisation different from that in material shown in Fig. 5b. (c) Detail showing roughendoplasmic reticulum (arrow) and various dictyosome-derived vesicles (arrowheads) commonly observedin these cells [scale bars indicate (a) 0.5 µm, (b, c) 0.25 µm].

H2O2 and catalase gene expression of potato leaves Functional Plant Biology 717
a b
Fig. 8. Symptoms of injury on leaf surface caused by 28 h of H2O2
treatment. Leaves from HBR-sprayed plants (a) and from Et-sprayedplants (b). Brownish areas throughout the leaflet surface illustrated (b)contrast with the damage shown (a) in which it appeared restricted totiny areas to the basal-half of the leaflet.
role of H2O2 in the regulation of the expression of CAT genesis limited. Polidoros and Scandalios (1999) investigated therole of H2O2 in the modulation of the expressionof the three maize CAT genes. These authors reported thatthe response of CAT genes to H2O2 was dependent onconcentration and on the duration of treatment. Since thesestudies were conducted with maize and ours with potato,comparison of results must take into consideration the basicnomenclature proposed for CAT isoforms (Willekens et al.1994a). In accordance to this nomenclature, CAT2 of potatoand CAT3 of maize are included in the class II catalases.All the isoforms included in this class are mainly found invascular tissues and, although their biological role is stillunclear, may play a role in lignification (Van Breusegem et al.2001). CAT1 of potato and CAT2 of maize are included inthe class I catalases and are considered responsible for theremoval of H2O2 during photorespiration. Despite of theiridentical physiological role, they respond differently to H2O2in each species. In maize, 60 mM or 150 mM H2O2 inducedCAT2 expression, after 24 h (Polidoros and Scandalios 1999),whereas in potato 100 mM H2O2 did not affect CAT1expression after 28 h. Although the maintenance of CAT1transcript levels, CAT1 activity and protein amount declinedin H2O2-treated leaves, suggesting that the effect of H2O2on CAT1 is downward of transcription. A decline in CAT1may be related to protein degradation caused by changes inprotective compound content or a consequence of mesophyllcellular changes that disable the cells for post-transcriptionprocessing of CAT1 mRNA. H2O2 caused a marked increasein CAT2 transcript abundance, and in CAT2 protein andactivity, so H2O2 induced CAT2 transcription and did notprevent the cells from producing and maintaining this catalaseisoform active. The regulatory role of H2O2 in the expressionof potato CAT2 is in accordance with the role reported
in maize CAT3 gene expression (Polidoros and Scandalios1999). The different response of CAT1 gene of potato to H2O2compared with that of CAT2 in maize could be related withthe different photosynthetic pathways these plants perform,and also due to the differential localisation of antioxidants inmaize leaves (Doulis et al. 1997). However, the possibilityof species-specific regulation of CAT expression and CATactivity must not be ruled out.
In our experiments, H2O2 supplied through the cut petiolemoved upwards through xylem and a high level could beaccumulated earlier in the phloem cells as well as in theperiveinal mesophyll cells, and then also in both palisadeand spongy parenchyma cells. Our previous work showedthat in potato leaves, CAT2 is localised in the paracrystallineinclusions in the peroxisomes (Santos et al. 2004). Thistype of peroxisomes appeared restricted to perivascularcells in non-stressed potato plants, but under UV-B stress,peroxisomes with paracrystalline inclusions appeared inmesophyll and in epidermal cells paralleled to an increase inCAT2 activity (Santos et al. 2004). The present work showsthat in H2O2-treated leaves the increase in CAT2 was alsoparalleled by the presence of this type of peroxisomes inmesophyll cells besides perivascular cells. The role of CAT2is still unclear; however, data available has led to the ideathat this isoform might play a particularly important role instress protection (Willekens et al. 1994b; Santos et al. 2004).Data from the present study represent an additional supportto the involvement of CAT2 in stress defence, as it shows thatH2O2 induced CAT2 expression and increased CAT2 activityin potato plants.
Brassinosteroids have been associated with the protectionof plants from different stresses (Krishna 2003) although themechanisms underlying this protection are not yet completelyunderstood. The influence of BRs on CAT activity hasbeen studied in only a few species. In a salt-sensitive ricecultivar, under salinity stress, 24-epibrassinolide did notchange CAT activity (Ozdemir et al. 2004), whereas ina different cultivar of rice an increase in activity was inducedby a BR analogue (Nunez et al. 2003, 2004). An increasein CAT activity has been reported in different varieties ofsorghum (Vardhini and Rao 2003), and in tomato, the increasein CAT activity depended on the structure of BRs, dosesand temperature (Mazorra et al. 2002). Except for the studyin leaf disks of tomato, all these studies were conductedwith monocotyledonous plants. Our study was conductedwith a dicotyledonous plant, and HBR was supplied byspraying. In leaves from HBR-sprayed plants, the responseof CAT1 and CAT2 to H2O2 followed the same pattern asencountered in leaves from Wt- and HBR-sprayed plants,after 4 h as well as after 28 h. However, the differencebetween controls and H2O2-treated leaves was smaller inleaves from HBR-sprayed plants. Because HBR is dissolvedin ethanol (10%), the response in both, Et- and HBR-sprayedleaves must be compared to access the changes due to

718 Functional Plant Biology J. M. Almeida et al.
HBR itself. Leaves from Et-sprayed plants respond to H2O2after 4 h, with a 1.8-fold increase in CAT activity comparedwith that observed in leaves from HBR-sprayed plants, and,after 28 h, the difference was 2.7 times higher. These resultscould be interpreted as the antagonistic effects of ethanol andHBR on CAT activity. However, since leaves were suppliedwith H2O2 6 d after the second spraying, it seems moreplausible that spraying the plants with ethanol or with HBRdissolved in ethanol affected the metabolic status of leaves,leading them to respond differently to exogenous H2O2.Indeed, differences were found through biochemical andelectron microscopy studies between H2O2-treated leavesfrom HBR- and Et-sprayed plants. H2O2-treated leaves fromHBR-sprayed plants appeared cytologically less affected thanleaves from Et-sprayed plants (Tables 2, 3; Figs 4b, 5, 7).After 4 h, the number of damaged mitochondria and affectedchloroplasts was smaller in H2O2-treated leaves from HBR-sprayed leaves than in treated leaves from Et-sprayed plants.This type of mitochondrial change is similar to that observedin leaves of transgenic tobacco with reduced activity inCAT1 after exposure to high light for 4 h, treatment thatcaused mitochondria disruption (Dat et al. 2003), althoughmitochondria disruption was not seen in our material. Thismaintenance of mitochondria integrity might be an importantprerequisite for the recovery of the cell. In fact, from4 to 28 h, recovery occurred since in all leaves, as all cellcomponents appeared better preserved and the number ofdamaged mitochondria decreased. There are reports showingthat the structural damage of stressed cells can be repairedwithin minutes or hours (Ciamporova 1989). The rapidrepair of mitochondria was reported and considered essentialfor further recovery of other cell components (Noailles1978). Rough endoplasmic reticulum and dictyosomes withassociated vesicles were abundant and most conspicuousin HBR-sprayed leaves after 28 h in H2O2-treated leaves.These features are indicative of high metabolic activity.Considering the three groups of H2O2-treated leaves the mostnoticeable difference was the high amount of osmiophilicdeposits in leaves from HBR-sprayed plants. Additionally,in these leaves the deposits are localised in the cellprotoplasts contrasting with the localisation mainly outsidecell protoplasts, found in the Et-sprayed plants. Accumulationof tannin-like compounds and phenolics droplets has beenreported in plants under stress and is considered to beinvolved in stress defence (Paakkonen et al. 1998; Oksanenet al. 2001; Blokhina et al. 2003). Osmiophilic depositssimilar to the ones we observed were induced by low-temperatures in leaves of winter oilseed rape, in whichthey were attributed an antioxidant role (Stefanowska et al.2002). It can be considered that the osmiophilic depositslocalised within cells in H2O2-treated leaves from HBR-sprayed plants can perform a similar role, contributing toless damage of cellular sub-structures and to the restrictionof leaf surface injury. There are two reports on the effect of
BRs on the ultrastructure of cells. One studied the effect ofBRs on the ultrastructure of cells of barley leaf and reportedthat 24-epibrassinolide produced a protective effect on cellultrastructure in leaves under saline stress (Kulaeva et al.1991). The other focused on the effect of a BRs-analogue andhigh temperature stress on leaf ultrastructure of tomato, andreported that the changes provoked by heat shock were moremarked in cells treated with the brassinosteroid analogue(Sam et al. 2001). Our study shows that the homobrassinolidereduced the negative changes on cell ultrastructure inducedby H2O2 in potato leaves.
Despite the general cytological recovery after 28 h,severely damaged cells were found in the proximity of theveins, although only in Wt- and in Et-sprayed leaves. Thisresponse to H2O2 seems to be localised to microdomains,i.e. to the periveinal parenchyma cells. Cell death in thevicinity of veins has been reported in response to salt(Hernandez et al. 2001), ozone, pathogens, and in transgenicplants with reduced catalase activity (see Dat et al. 2003), andhas been correlated with high levels of H2O2. This localisedcell death has been interpreted as indicating that constitutiveor stress-induced protective enzymes may be differentlyexpressed within the leaf (Willekens et al. 1994a, b)and small clusters of cells in the proximity of the veins seemto have lower antioxidant capability to remove the cytotoxicmolecule. There are reports that the effects of exogenousH2O2 depend on its concentration at the site of its action(Neill et al. 2002b). Thus, the damage we observed canbe partially explained by lower H2O2-scavenging capabilityof the cells, mainly due to the low level of CAT2 in thephloem cells at the beginning of treatment, which allowedfor rapid transport of H2O2 to perivascular mesophyll cells,in which H2O2 reached phytotoxic levels and caused damage.In H2O2-treated leaves from HBR-plants, the higher level ofCAT2 at the beginning of the treatment and the high level ofosmiophilic deposits they accumulate certainly contributedto avoidance of severe damage of the perivascular cells.
In conclusion, our results show that in potato leaves,exogenous H2O2 induces CAT2 expression and increasesCAT2 protein and activity. In contrast, H2O2 does not changethe level of CAT1 transcripts, but decreases CAT1 activity andamount; data showing that CAT1 and CAT2 genes responddifferently to 100 mM H2O2. Moreover, our data show thatHBR spraying of potato increases the basal level of CAT2,does not change the isozyme pattern of catalase in responseto H2O2, but affects the magnitude of the increase of totalactivity. Our results also show that HBR treatment alleviatesthe negative effects of H2O2 on the leaf cell structures andallows better recovery of the cell components. All theseresults point to a protective role of HBR that would involvediverse mechanisms besides the antioxidant enzyme catalase.The response to oxidative stress involves various genesbesides those implicated in the control of the level of ROS(Vranova et al. 2002; Dutilleul et al. 2003), and, therefore,

H2O2 and catalase gene expression of potato leaves Functional Plant Biology 719
further studies are required to unravel the stress tolerancemechanisms activated by BRs in plants.
To gain further insights into the regulation of theexpression of CAT1 and CAT2 genes in potato by H2O2,further studies are needed, and time course experimentswith H2O2, at different concentrations, are underway in ourlaboratory.
Acknowledgments
The work was supported by the Fundacao para aCiencia e Tecnologia (FCT, Lisboa, Portugal — projectPOCTI / BME / 33044 / 2000).
References
Anderson JA (2002) Catalase activity, hydrogen peroxide contentand thermotolerance of pepper leaves. Scientia Horticulturae 95,277–284. doi: 10.1016/S0304-4238(02)00076-6
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidativedamage and oxygen deprivation stress: a review. Annals of Botany91, 179–194. doi: 10.1093/aob/mcf118
Castle J, Montoya T, Bishop GJ (2003) Selected physiological responsesof brassinosteroids: a historical approach. In ‘Brassinosteroids:bioactivity and crop productivity’. (Eds S Hayat, A Ahmad)pp. 45–68. (Kluwer Academic Publishers: Dordrecht)
Ciamporova M (1989) Recovery of ultrastructure in water stressed rootepidermal cells of Zea mays. In ‘Structural and functional aspectsof transport in roots’. (Ed. BC Loughman) pp. 263–267. (KluwerAcademic Publishers: London)
Corpas FJ, Palma JM, Sandalio LM, Lopez-Huertas E,Romero-Puertas MC, Barroso JB, Del Rio LA (1999) Purificationof catalase from pea leaf peroxisomes: identification of five differentisoforms. Free Radical Research 31, S235–S241.
Dat J, Vandenabeele S, Vranova E, Van Montagu M, Inze D,Van Breusegem F (2000) Dual action of the active oxygen speciesduring plant stress responses. Cellular and Molecular Life Sciences57, 779–795. doi: 10.1007/s000180050041
Dat JF, Lopez-Delgado H, Foyer CH, Scott IM (1998) Parallel changesin H2O2 and catalase during thermotolerance induced by salicylicacid or heat acclimation in mustard seedlings. Plant Physiology 116,1351–1357. doi: 10.1104/pp.116.4.1351
Dat JF, Pellinen R, Beeckman T, Van de Cotte B, Langebartels C,Kangasjarvi J, Inze D, Van Breusegem F (2003) Changes inhydrogen peroxide homeostasis trigger an active cell death processin tobacco. The Plant Journal 33, 621–632. doi: 10.1046/j.1365-313X.2003.01655.x
Desikan R, Reynolds A, Hancock JT, Neill SJ (1998) Harpin andhydrogen peroxide both initiate programmed cell death but havedifferential effects on defence gene expression in Arabidopsissuspension cultures. Biochemical Journal 330, 115–120.
Desikan R, Cheung M-K, Clarke A, Golding S, Sagi M, Fluhr R, Rock C,Hancock JT, Neill SJ (2004) Hydrogen peroxide is a common signalfor darkness- and ABA-induced stomatal closure in Pisum sativum.Functional Plant Biology 31, 913–920. doi: 10.1071/FP04035
Doulis AG, Debian N, Kingston-Smith AH, Foyer CH (1997)Differential localization of antioxidants in maize leaves. PlantPhysiology 114, 1031–1037.
Dutilleul C, Garmier M, Noctor G, Mathieu C, Chetrit P, Foyer CH,de Paepe R (2003) Leaf mitochondria modulate whole cellredox homeostasis, set antioxidant capacity, and determine stressresistance through altered signaling and diurnal regulation. The PlantCell 15, 1212–1226. doi: 10.1105/tpc.009464
Frohman MA, Dush MK, Martin GR (1988) Rapid production of full-length cDNAs from rare transcripts: amplification using a singlegene-specific oligonucleotide primer. Proceedings of the NationalAcademy of Sciences USA 85, 8998–9002.
Gechev T, Gadjev I, Van Breusegem F, Inze D, Dukiandjiev S,Toneva V, Minkov I (2002) Hydrogen peroxide protects tobacco fromoxidative stress by inducing a set of antioxidant enzymes. Cellularand Molecular Life Sciences 59, 708–714. doi: 10.1007/s00018-002-8459-x
Guan LM, Scandalios G (2000) Hydrogen peroxide-mediated catalasegene expression in response to wounding. Free Radical Biology andMedicine 28, 1182–1190. doi: 10.1016/S0891-5849(00)00212-4
Hernandez JA, Ferrer MA, Jimenez A, Barcelo AR, Sevilla F (2001)Antioxidant systems and O2
./ H2O2 production in the apoplast of
pea leaves. Its relation with salt-induced necrotic lesions in minorveins. Plant Physiology 127, 817–831. doi: 10.1104/pp.127.3.817
Karpinski S, Reynolds H, Karpinska B, Wingsle G, Creissen G,Mullineaux P (1999) Systemic signaling and acclimation in responseto excess excitation energy in Arabidopsis. Science 284, 654–657.doi: 10.1126/science.284.5414.654
Krishna P (2003) Brassinosteroid-mediated stress responses. Journalof Plant Growth Regulation 22, 289–297. doi: 10.1007/s00344-003-0058-z
Kulaeva OP, Burkhanova EA, Fedina AB, Khokhlova VA,Bokebayeva GA, Vorbrodt HM, Adam G (1991) Effect ofbrassinosteroids on protein synthesis and plant-cell ultrastructureunder stress conditions. In ‘Brassinosteroids: chemistry, bioactivity,and applications’. pp. 141–155. (American Chemical Society:Washington)
Levine A, Tenhaken R, Dixon R, Lamb C (1994) H2O2 fromthe oxidative burst orchestrates the plant hypersensitive diseaseresistance response. Cell 79, 583–593. doi: 10.1016/0092-8674(94)90544-4
Lopez-Delgado H, Dat JF, Foyer CH, Scott IM (1998) Inductionof thermotolerance in potato microplants by acetylsalicylicacid and H2O2. Journal of Experimental Botany 49, 713–720.doi: 10.1093/jexbot/49.321.713
Mazorra LM, Nunez M, Hechavarria M, Coll F, Sanchez-Blanco MJ(2002) Influence of brassinosteroids on antioxidant enzymes activityin tomato under different temperatures. Biologia Plantarum 45,593–596. doi: 10.1023/A:1022390917656
Morita S, Kaminaka H, Masumura T, Tanaka K (1999) Induction ofrice cytosolic ascorbate peroxidase mRNA by oxidative stress; theinvolvement of hydrogen peroxide in oxidative stress signalling.Plant and Cell Physiology 40, 417–422.
Mussig C, Altmann T (1999) Physiology and molecular mode ofaction of brassinosteroids. Plant Physiology and Biochemistry 37,363–372. doi: 10.1016/S0981-9428(99)80042-4
Nakashita H, Yasuda M, Nitt T, Asami T, Fujioka S, Arai Y, Sekimata K,Takatsuto S, Yamaguchi I, Yoshida S (2003) Brassinosteroidfunctions in a broad range of disease resistance in tobaccoand rice. The Plant Journal 33, 887–898. doi: 10.1046/j.1365-313X.2003.01675.x
Neill S, Desikan R, Hancock J (2002a) Hydrogen peroxide signalling.Current Opinion in Plant Biology 5, 388–395. doi: 10.1016/S1369-5266(02)00282-0
Neill SJ, Desikan R, Clarke A, Hurst RD, Hancock JT (2002b)Hydrogen peroxide and nitric oxide as signalling moleculesin plants. Journal of Experimental Botany 53, 1237–1247.doi: 10.1093/jexbot/53.372.1237
Noailles M-C (1978) Etude ultrastructurale de la recuperation hydriqueapres une periode de secheresse cez une hypnobryale: Pleuroziumschreberi (Willd.) Mitt. Annales des Sciences Naturelles —Botanique et Biologie Vegetale 19, 249–265.

720 Functional Plant Biology J. M. Almeida et al.
Nunez M, Mazzafera P, Mazorra LM, Siqueira WJ, Zullo MAT (2003)Influence of a brassinosteroid analogue on antioxidant enzymes inrice grown in culture medium with NaCl. Biologia Plantarum 47,67–70. doi: 10.1023/A:1027380831429
Oksanen E, Sober J, Karnosky DF (2001) Impacts of elevated CO2
and / or O3 on leaf ultrastructure of aspen (Populus tremuloids)and birch (Betula papyrifera) in the Aspen FACE experiment.Environmental Pollution 115, 437–446. doi: 10.1016/S0269-7491(01)00233-0
Ozdemir F, Bor M, Demiral T, Turkan I (2004) Effects of24-epibrassinolide on seed germination, seedling growth,lipid peroxidation, proline content and antioxidative system of rice(Oryza sativa L.) under salinity stress. Plant Growth Regulation 42,203–211. doi: 10.1023/B:GROW.0000026509.25995.13
Paakkonen E, Gunthardt-Georg M, Holopainen T (1998) Responses ofleaf processes in a sensitive birch (Betula pendula Roth) clone toozone combined with drought. Annals of Botany 82, 45–59.
Polidoros AN, Scandalios JG (1999) Role of hydrogen peroxide anddifferent classes of antioxidants in the regulation of catalase andglutathione S-transferase gene expression in maize (Zea mays L.).Physiologia Plantarum 106, 112–120. doi: 10.1034/j.1399-3054.1999.106116.x
Prasad TK, Anderson MD, Martin BA, Stewart CR (1994a) Evidencefor chilling-induced oxidative stress in maize seedlings anda regulatory role for hydrogen-peroxide. The Plant Cell 6, 65–74.doi: 10.1105/tpc.6.1.65
Prasad T, Anderson M, Steward C (1994b) Acclimation, hydrogenperoxide, and abscisic acid protect mitochondria against irreversiblechilling injury in maize seedlings. Plant Physiology 105,619–627.
Rao MV, Paliyath G, Ormrod DP (1996) Ultraviolet-B- andozone-induced biochemical changes in antioxidant enzymesof Arabidopsis thaliana. Plant Physiology 110, 125–136.doi: 10.1104/pp.110.1.125
Salema R, Brandao I (1973) The use of PIPES buffer in the fixationof plant cells for electron microscopy. Journal SubmicroscopyCytolology 5, 317–329.
Sam O, Nunez M, Ruiz-Sanchez MC, Dell’amico J, Falcon V,De La Rosa MC, Seoane J (2001) Effect of a brassinosteroidanalogue and high temperature stress on leaf ultrastructureof Lycopersicon esculentum. Biologia Plantarum 44, 213–218.doi: 10.1023/A:1010291124178
Sambrook J, Fritsch EF, Maniatis T (1989) ‘Molecular cloning:a laboratory manual.’ (2nd edn) (Cold Spring Harbor LaboratoryPress: Cold Spring Harbor, NY)
Santos I, Fidalgo F, Almeida JM, Salema R (2004) Biochemicaland ultrastructural changes in leaves of potato plants grownunder supplementary UV-B radiation. Plant Science 167, 925–935.doi: 10.1016/j.plantsci.2004.05.035
Scandalios JG, Guan L, Polidoros AN (1997) Catalases in plants:gene structure, properties, regulation and expression. In ‘Oxidativestress and the molecular biology of antioxidant defenses’.(Ed. JG Scandalios) pp. 343–406. (Cold Spring Harbor LaboratoryPress: Cold Spring Harbor, NY)
Stefanowska M, Kuras M, Kacperska A (2002) Lowtemperature-induced modifications in cell ultrastructure andlocalization of phenolics in winter oilseed rape (Brassicanapus L. var. oleifera L.) leaves. Annals of Botany 90, 637–645.doi: 10.1093/aob/mcf241
Uchida A, Jagendorf AT, Hibino T, Takabe T, Takabe T (2002) Effectsof hydrogen peroxide and nitric oxide on both salt and heat stresstolerance in rice. Plant Science 163, 515–523. doi: 10.1016/S0168-9452(02)00159-0
Van Breusegem F, Vranova E, Dat JF, Inze D (2001) The role of activeoxygen species in plant signal transduction. Plant Science 161,405–414. doi: 10.1016/S0168-9452(01)00452-6
Vandenabeele S, Van Der Kelen K, Dat J, Gadjev I,Boonefaes T, et al. (2003) A comprehensive analysis of hydrogenperoxide-induced gene expression in tobacco. Proceedings ofthe National Academy of Sciences USA 100, 16113–16118.doi: 10.1073/pnas.2136610100
Vardhini BV, Rao SSR (2003) Amelioration of osmotic stress bybrassinosteroids on seed germination and seedling growth ofthree varieties of sorghum. Plant Growth Regulation 41, 25–31.doi: 10.1023/A:1027303518467
Vranova E, Atichartpongkul S, Villarroel R, Van Montagu M,Inze D, Van Camp W (2002) Comprehensive analysis of geneexpression in Nicotiana tabacum leaves acclimated to oxidativestress. Proceedings of the National Academy of Sciences USA 99,10 870–10 875. doi: 10.1073/pnas.152337999
Wang Z-Y, He J-X (2004) Brassinosteroid signal transduction —choices of signals and receptors. Trends in Plant Science 9, 91–96.doi: 10.1016/j.tplants.2003.12.009
Willekens H, Chamnongpol S, Davey M, Schraudner M,Langebartels C, Van Montagu M, Inze D, Van Camp W(1997) Catalase is a sink for H2O2 and is indispensable forstress defence in C3 plants. EMBO Journal 16, 4806–4816.doi: 10.1093/emboj/16.16.4806
Willekens H, Langebartels C, Tire C, Van Montagu M, Inze D,Van Camp W (1994a) Differential expression of catalase genesin Nicotiana plumbaginifolia (L.). Proceedings of the NationalAcademy of Sciences USA 91, 10 450–10 454
Willekens H, Van Camp W, Van Montagu M, Inze D,Sandermann H, Langebartels C (1994b) Ozone, sulfur dioxide,and ultraviolet B have similar effects on mRNA accumulationof antioxidant genes in Nicotiana plumbaginifolia (L.). PlantPhysiology 106, 1007–1014.
Willekens H, Villarroel R, Van Montagu M, Inze D, Van Camp W(1994c) Molecular identification of catalases from Nicotianaplumbaginifolia (L.). FEBS Letters 352, 79–83. doi: 10.1016/0014-5793(94)00923-6
Manuscript received 11 December 2004, received in revised form12 April 2005, accepted 6 May 2005
http://www.publish.csiro.au/journals/fpb




















