
Importance of mesophyll diffusion conductance in estimation ...

Physiologia Plantarum 149: 515–527. 2013 © 2013 Scandinavian Plant Physiology Society, ISSN 0031-9317
Ability of leaf mesophyll to retain potassium correlateswith salinity tolerance in wheat and barleyHonghong Wu, Lana Shabala, Karen Barry, Meixue Zhou and Sergey Shabala∗
School of Agricultural Science and Tasmanian Institute of Agriculture, University of Tasmania, Hobart, Tasmania 7001, Australia
Correspondence*Corresponding author,e-mail: [email protected]
Received 11 January 2013;revised 11 March 2013
doi:10.1111/ppl.12056
This work investigated the importance of the ability of leaf mesophyllcells to control K+ flux across the plasma membrane as a trait conferringtissue tolerance mechanism in plants grown under saline conditions. Fourwheat (Triticum aestivum and Triticum turgidum) and four barley (Hordeumvulgare) genotypes contrasting in their salinity tolerance were grown underglasshouse conditions. Seven to 10-day-old leaves were excised, and net K+
and H+ fluxes were measured from either epidermal or mesophyll cells uponacute 100 mM treatment (mimicking plant failure to restrict Na+ deliveryto the shoot) using non-invasive microelectrode ion flux estimation (theMIFE) system. To enable net ion flux measurements from leaf epidermalcells, removal of epicuticular waxes was trialed with organic solvents. Aseries of methodological experiments was conducted to test the efficiencyof different methods of wax removal, and the impact of experimentalprocedures on cell viability, in order to optimize the method. A strongpositive correlation was found between plants’ ability to retain K+ in salt-treated leaves and their salinity tolerance, in both wheat and especially barley.The observed effects were related to the ionic but not osmotic componentof salt stress. Pharmacological experiments have suggested that voltage-gatedK+-permeable channels mediate K+ retention in leaf mesophyll upon elevatedNaCl levels in the apoplast. It is concluded that MIFE measurements of NaCl-induced K+ fluxes from leaf mesophyll may be used as an efficient screeningtool for breeding in cereals for salinity tissue tolerance.
Introduction
Salinity affects over 800 million hectares of agriculturalland, which impacts crop production across the globe(Rengasamy 2010). To match the predicted populationgrowth with food supply salt-tolerant crops have to bebred and recommended to growers.
Salinity stress tolerance is a complex polygenic traitthat is conferred by multiple physiological mechanisms(Flowers 2004, Munns and Tester 2008, Shabala and
Abbreviations – BSM, basic salt media; BSTFA, N,O-bis-trimethylsilylacetamide; MAS, marker-assisted selection; MIFE,microelectrode ion flux estimation; MP, membrane potential; NSCC, non-selective cation channels; PCD, programmed celldeath; PSII, photosystem II; TEA, tetraethylammonium chloride; TMS, tri-methyl-silyl; TUNEL, terminal deoxynucleotidyltransferase dUTP nick end labeling; Vanadate, sodium orthovanadate.
Cuin 2008). Because of this, pyramiding physiologicaltraits contributing to salinity tolerance are arguably theonly practical way to improve salinity stress tolerancein crops. This also poses a question of what specifictrait should be used to identify the source of salttolerance genes to be used in marker-assisted selection(MAS)-based breeding programs. Various agronomictraits such as germination, survival and growth rates,the extent of leaf injury, biomass production (Manoand Takeda 1997, Eleuch et al. 2008, Xu et al. 2012,
Physiol. Plant. 149, 2013 515

Zhou et al. 2012) and whole-plant physiologicalcharacteristics such as CO2 assimilation, chlorophyllcontent, shoot Na+ and K+ content (James et al. 2002,Chen et al. 2005, Munns et al. 2006) have beenfrequently advocated. To a large extent, the choicehas been determined by the convenience (e.g. its cost-and time-efficiency) of the measuring procedure. Thisis not ideal as the screening technique should target themechanism directly contributing toward salinity stresstolerance. Striking the right balance is not an easy task.
Until now, researchers have focused predominantlyon traits related to sodium, such as Na+ exclusion fromuptake, control of xylem Na+ loading, Na+ retrieval fromthe shoot or vacuolar Na+ sequestration (Blumwald et al.2000, Horie and Schroeder 2004, Munns and Tester2008, Plett et al. 2010, Cuin et al. 2011). However, itis not Na+ but the K+/Na+ ratio in the cytosol whichdetermines plant performance under saline conditions(Maathuis and Amtmann 1999, Shabala and Cuin 2008).K+ is recognized as a rate-limiting factor for cropyield and quality, directly contributing to cell turgorand hence growth (Dreyer and Uozum 2011). At thecellular level, maintaining intracellular K+ homeostasisis essential for enzyme activation, stabilization of proteinsynthesis, neutralization of negatively charged proteins,formation of membrane potential (MP) and maintenanceof cytosolic pH homeostasis (Shabala 2003, Dreyerand Uozum 2011). Potassium also plays an importantrole in controlling apoptosis in animal tissues (Hughesand Cidlowski 1999), and a direct causal link betweenplant ability to control fluxes of K+ across the plasmamembrane and programmed cell death (PCD) wasreported for a range of abiotic stress conditions (Shabalaet al. 2007, Demidchik et al. 2010), including salinity.
We have previously reported a strong correlationbetween plant ability to retain K+ and salinity tolerancein both barley (Chen et al. 2005, 2007b, 2007c) andwheat (Cuin et al. 2008, 2012), as well as providedstrong evidence for the heritability of this trait (Chenet al. 2007a, Cuin et al. 2012). All these studies weredone in roots. However, it is ultimately a plant’sability (or inability) to control the K+/Na+ ratio inphotosynthetically active leaf tissues that determinesits photosynthetic competence (and hence growth andyield) under saline conditions. Indeed, each of theabove Na+-related mechanisms (e.g. exclusion fromuptake; restricted loading into xylem and retrieval fromthe shoot) has potential limitations and cannot operateindefinitely in a broad range of salinities. Exposure ofthe plant to longer durations and more severe salt stressincreases the chance that one of these mechanismswill fail and high amounts of Na+ will be deliveredto the shoot with a transpiration flow. This will result
in significant membrane depolarization and a massiveK+ leak from leaf mesophyll (Shabala and Cuin 2008).The consequence of this will be reduced metabolic(e.g. Rubisco; Osaki et al. 1993) activity, reduced cellturgor, impaired Na+ sequestration in the vacuole(resulting from K+ requirements for vacuolar PPaseactivity; Davies et al. 1992) and a danger of undertakingPCD resulting from increased activities of proteases andendonucleases at low cytosolic K+ (Demidchik et al.2010). Thus, the ability of leaf mesophyll cells to controlK+ flux across the plasma membrane may be a veryessential component of tissue tolerance mechanisms,contributing to overall plant performance under salineenvironment. However, to the best of our knowledge,no studies have been conducted so far looking at theextent of genetic variability in this trait. Thus, the aim ofthis study was two-fold: (1) to determine to what extentsalinity tolerance in wheat and barley is related to theability of mesophyll tissue to retain K+ when exposed toelevated NaCl levels in the apoplast and (2) to developefficient screening protocols allowing rapid assessmentof a large number of samples for the above trait.
Materials and methods
Plant material
Four wheat (Triticum aestivum cultivars Kharchia 65 andBaart 46, and Triticum turgidum spp. durum cultivarsWollaroi and Tamaroi) and four barley (Hordeumvulgare cultivars CM 72, Gairdner, TX9425 and NasoNijo) varieties, contrasting in their salinity tolerance,were used in this study. Among wheats, bread wheatvariety Kharchia and durum wheat variety Wollaroiwere previously reported as being tolerant, while Baartand Tamaroi were sensitive to salt (Cuin et al. 2008).Among barley varieties, CM72 and TX9425 weredeemed tolerant, while Gairdner and Naso Nijo werehighly sensitive (Chen et al. 2007b, Xu et al. 2012).Salt tolerance was defined as a genotype’s ability tomaintain biomass growth and grain yield under salineconditions and was correlated with better K+/Na+ inthe shoot, both in wheat (Cuin et al. 2008) and barley(Chen et al. 2005, 2007c). All seeds of these varietieswere obtained from various sources in the past andmultiplied in our laboratory. Plants were grown usingUniversity of Tasmania glasshouse facilities essentiallyas described in Chen et al. (2007c). Two- to 3-week-oldplants were used for measurements.
Sample preparation and wax removal trials
Seven to 10-day-old leaves were excised by a razorblade and brought into the laboratory. The presence of
516 Physiol. Plant. 149, 2013

epicuticular waxes poses a methodological problem forion flux measurements (Shabala and Newman 1999) incereals and removing these without altering cell functionwas required to develop an efficient screening protocol.Leaf cuticular wax removal was trialed with differentsolvents (either 25 or 96% methanol or analytical gradehexane), application methods (immersion or rubbing)and duration of application (for the rubbing procedure).The effect of these procedures on efficiency of waxremoval, ability to measure K+ flux and toxicity to theplant was determined, as outlined below.
After wax removal, leaves were immediately andthoroughly rinsed in running distilled water for severalminutes. Small leaf segments (approximately 5 × 8 mm)were cut and left floating (rubbed surface down) onexperimental basic salt media (BSM) solution (0.1 mMCaCl2 and 0.5 mM KCl; pH 5.7 non-buffered) overnightin the dark to minimize possible confounding effectsof tissue damage on ion fluxes (Shabala and Newman1999). Net ion fluxes were then measured from theepidermal leaf surface on which cuticular wax wasremoved. To measure net fluxes of K+ and H+ fromleaf mesophyll, a cross-sectional cut was made acrossthe middle part of the leaf blade exposing leaf mesophylltissue. Cut samples were then placed onto the surface ofBSM solution and kept in the dark until measured.
Wax analysis
To compare the efficiency of wax removal methods,epicuticular wax was extracted from segments of wheatusing dichloromethane with docosane (C-22 alkane)as an internal standard (2 mg docosane was dilutedin 100 ml dichloromethane). The C28 wax speciesare dominant in wheat where it contributes to over2/3 of all waxes (e.g. Koch et al. 2006); this isalso consistent with our own findings. Accordingly,we considered the C28 species representative for thepurposes of this article. Leaf segments of the samethickness and surface area were soaked in 10 ml ofsolvent for 5 min with gentle stirring. This solution wasdecanted, taken to dryness under a stream of N2, thenderivatized with a double TMS (tri-methyl-silyl) method.Briefly, 10 μl of pyridine and 40 μl of BSTFA (N,O-bis-trimethylsilylacetamide) were added, samples were thenheated at 80◦C for 20 min, then dried. A second smallervolume of pyridine and BSTFA was added; samples wereheated at 80◦C for 15 min, and then dried again. Sampleswere analyzed by combined gas chromatography–massspectrometry (GC–MS) on a Hewlett-Packard 5890 Gaschromatograph coupled to a Hewlett-Packard 5970Bmass selective detector (Hewlett-Packard Australia PtyLtd, Melbourne). One microliter injections were made
in the splitless mode, with an injection temperatureof 300◦C and transfer line temperature of 290◦C.The column was a 25 m × 0.32 mm HP1 (0.17 μmfilm thickness), with a temperature program of 60◦C(held for 1 min) to 300◦C at 10◦C min−1, with an 8-min hold time at the final temperature. Mass spectrawere collected over the range m/z 40–550 at twoscans per second. Total wax was expressed as theratio of C28 alcohols (the TMS derivatives) to the C22internal standard.
Non-invasive ion flux measurements
Net K+ and H+ fluxes were measured using thenon-invasive microelectrode ion flux estimation (MIFE)technique (UTas Innovation Ltd., Hobart, Tasmania)essentially as described previously (Shabala and Shabala2002, Tegg et al. 2005). The theory of MIFE measure-ments is available elsewhere (Newman 2001). Briefly,microelectrodes were pulled from borosilicate glass cap-illaries (GC150-10; Clark Electrochemical instruments,Pangbourne, Berks, UK), dried in the oven at 225◦Covernight, and then silanized with tributylchlorosilane(catalog no. 90796; Fluka, Busch, Switzerland). Driedand cooled microelectrodes were backfilled withappropriate solutions (15 mM NaCl + 40 mM KH2PO4;pH 6.0 adjusted using NaOH for H+ or 200 mM KClfor K+). Then, the tip of the electrodes was filledwith commercially available ionophore cocktails (K+,60031; H+, 95297, Fluka, Busch, Switzerland) andprepared microelectrodes were calibrated using a setof appropriate standards before and after use. Onlyelectrodes with a slope of above 50 mV per decade andcorrelation 0.999 or better were used.
Experimental procedure
Leaf samples were mounted in a Perspex holder andplaced into measuring chambers filled with measuringsolution (0.1 mM CaCl2 and 0.5 mM KCl). Electrodeswere mounted on a 3D-micromanipulator (MMT-5;Narishige, Tokyo, Japan), tips were positioned closetogether and at 40 μm from the leaf sample. Duringmeasurements, the electrodes were moved by acomputer-controlled hydraulic manipulator betweentwo positions, 40 and 110 μm from the leaf surface, in a12-s square-wave cycle. Net ion fluxes were calculatedfrom recorded electrical potential differences usingcylindrical diffusion geometry (Newman 2001). Net ionfluxes were measured for 5–10 min under control (BSM)conditions, before 100 mM NaCl was added to the bathfollowed by another 50–60 min of measurements. Insome experiments, 170 mM mannitol treatment (isotonicto 100 mM NaCl) was used of NaCl.
Physiol. Plant. 149, 2013 517

Membrane potential measurements
Conventional KCl-filled Ag/AgCl microelectrodes withtip diameter 0.5 μm were used to measure the MP ofwheat leaf segments. Measurements were taken fromat least four individual plants for each treatment, withfour to five measurements taken from each leaf segment.MPs were recorded for at least 30 s after the potentialstabilized following cell penetration. Measurementswere carried out on epidermal cells treated with themost efficient method of wax removal (as described inthe section Results).
Chlorophyll fluorescence
The maximal photochemical efficiency of PhotosystemII (PSII) was estimated by measuring the chlorophyllfluorescence Fv/Fm ratio (Smethurst and Shabala 2003)of the leaf segments that were subjected to different waxremoval methods.
Pharmacology
Further insights into mechanisms underlying leaf abilityto retain K+ upon salinity treatment were obtainedby conducted a series of pharmacological experimentsusing barley variety Gairdner. In these pharmacologicalexperiments, leaf segments were pre-treated for 1 h witheither 20 mM tetraethylammonium chloride (TEA+; aknown blocker of K+ selective channels) or 1 mM sodiumorthovanadate (Na3VO4; a known inhibitor of P-typeH+-ATPase) before being exposed to NaCl treatment.All chemicals are from Sigma-Aldrich, St. Louis, MO.
Statistical analysis
Data was analyzed with SPSS 16.0 for windows (SPSS Inc.,Chicago, IL). All data are given as mean ± SE (n = samplesize). Significance was determined by one-way ANOVA
based on Duncan’s multiple range test. Significantdifferences (P ≤ 0.05) between treatments were denotedby different lower case letters.
Results
Methodological aspects of ion flux measurementsfrom leaf epidermis and requirement forwax removal
The primary aim of this work was to develop a highthroughput method for a rapid assessment of a largenumber (hundreds or even thousands) of specimens.From a technological point of view, this can be donemore easily when working with intact leaves (e.g. mea-suring fluxes from leaf surface). However, the leaf surface
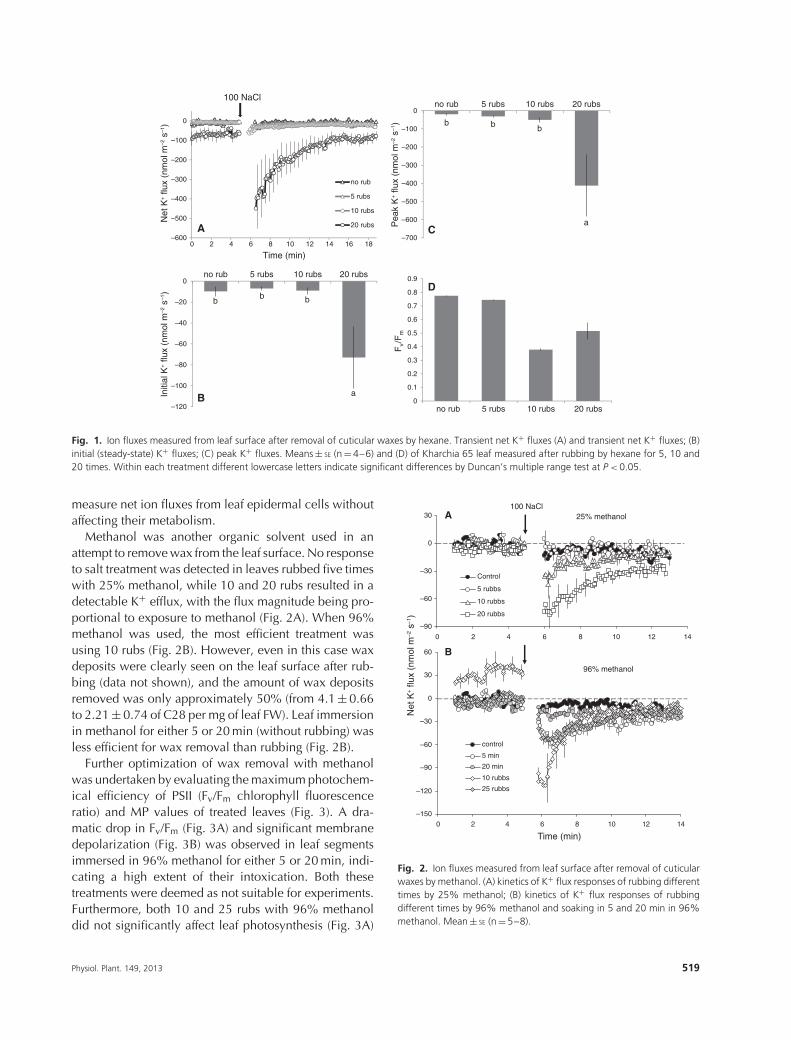
is known to be covered by a cuticle which is composedof a matrix of the biopolymer cutin and waxes of variouscomposition that are usually referred to as epicuticularwaxes (Bewick et al. 1993). Epicuticular waxes plays animportant role in plant function, including prevention ofnon-stomatal water loss, defending leaves from bacterialand fungal pathogens, alleviating detrimental effects ofabiotic stress such as UV damage, and playing a rolein the interaction between plants and insect herbivores(Bewick et al. 1993, Rhee et al. 1998). However, theirpresence makes the leaf surface highly hydrophobic andalmost impermeable to water and ion movement, posinga methodological problem for ion flux measurements(Shabala and Newman 1999). Both wheat and barleyhave significant wax deposits (data not show). Whenchallenged with stress factors, these species displayedlittle if any ion exchange across the leaf surface whenwax was present (Fig. 1A). The 100 mM NaCl treatmentcaused no detectable K+ flux signal when measuredfrom untreated leaf epidermis, with epicuticular waxespresent (Fig. 1A). This may be caused by either waxesrestricting Na+ uptake into the epidermal cells (thus, pre-venting NaCl-induced plasma membrane depolarizationand associated K+ efflux as reported for other non-cutinized leaf tissues; Shabala 2000, Shabala et al. 2006)or by preventing K+ leakage from beneath the epicuticu-lar barrier, or both. This poses a serious methodologicalissue and prevents studying the effect of salinity on ionicrelations in leaves with the MIFE technique.
An efficient way to deal with the problem may beto remove wax deposits by some organic solvents, e.g.chloroform, methanol, ethanol and hexane (Bewick et al.1993, Shabala and Shabala 2002, Zivanovic et al. 2005,Agrawal et al. 2007). The extent of the wax removal willbe critically dependent on the concentration of solventused and the duration of exposure. While the longer andhigher treatments will certainly assure more effectivewax removal, leaf intoxication is likely to occur. Thus,the process of removal of epicuticular waxes needs tobe finely balanced.
We investigated the above problem first by usinganalytical grade hexane and rubbing the leaf surface withhexane-wetted cotton buds for various number of times(hence, different exposures). Rubbing the leaf surfacewith hexane 5 or 10 times did not enable measurement ofNaCl-induced K+ fluxes from epidermal cells under salttreatment (Fig. 1A–C). Longer exposures to hexane (20rubs) make NaCl-induced K+ flux detectable (Fig. 1A–C),however this treatment also caused significant leafintoxication as judged by a very significant declinein leaf chlorophyll fluorescence characteristics (Fv/Fm
values; Fig. 1D). Thus, it was concluded that hexanewas not suitable to be used for efficient wax removal to
518 Physiol. Plant. 149, 2013
–120
–100
–80
–60
–40
–20
0no rub 5 rubs 10 rubs 20 rubs
–600
–500
–400
–300
–200
–100
0
0 2 4 6 8 10 12 14 16 18
Time (min)
no rub
5 rubs
10 rubs
20 rubs
100 NaCl
–700
–600
–500
–400
–300
–200
–100
0no rub 5 rubs 10 rubs 20 rubs
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
no rub 5 rubs 10 rubs 20 rubsF
v/F
m
A
B
C
D
a
b bb
b b b
aNet
K+ fl
ux (
nmol
m–2
s–1
)
Pea
k K
+ fl
ux (
nmol
m–2
s–1
)
Initi
al K
+ fl
ux (
nmol
m–2
s–1
)
Fig. 1. Ion fluxes measured from leaf surface after removal of cuticular waxes by hexane. Transient net K+ fluxes (A) and transient net K+ fluxes; (B)initial (steady-state) K+ fluxes; (C) peak K+ fluxes. Means ± SE (n = 4–6) and (D) of Kharchia 65 leaf measured after rubbing by hexane for 5, 10 and20 times. Within each treatment different lowercase letters indicate significant differences by Duncan’s multiple range test at P < 0.05.
measure net ion fluxes from leaf epidermal cells withoutaffecting their metabolism.
Methanol was another organic solvent used in anattempt to remove wax from the leaf surface. No responseto salt treatment was detected in leaves rubbed five timeswith 25% methanol, while 10 and 20 rubs resulted in adetectable K+ efflux, with the flux magnitude being pro-portional to exposure to methanol (Fig. 2A). When 96%methanol was used, the most efficient treatment wasusing 10 rubs (Fig. 2B). However, even in this case waxdeposits were clearly seen on the leaf surface after rub-bing (data not shown), and the amount of wax depositsremoved was only approximately 50% (from 4.1 ± 0.66to 2.21 ± 0.74 of C28 per mg of leaf FW). Leaf immersionin methanol for either 5 or 20 min (without rubbing) wasless efficient for wax removal than rubbing (Fig. 2B).
Further optimization of wax removal with methanolwas undertaken by evaluating the maximum photochem-ical efficiency of PSII (Fv/Fm chlorophyll fluorescenceratio) and MP values of treated leaves (Fig. 3). A dra-matic drop in Fv/Fm (Fig. 3A) and significant membranedepolarization (Fig. 3B) was observed in leaf segmentsimmersed in 96% methanol for either 5 or 20 min, indi-cating a high extent of their intoxication. Both thesetreatments were deemed as not suitable for experiments.Furthermore, both 10 and 25 rubs with 96% methanoldid not significantly affect leaf photosynthesis (Fig. 3A)
–150
–120
–90
–60
–30
0
30
60
0 2 8 10 12 14
control
5 min
20 min
10 rubbs
25 rubbs
–90
–60
–30
0
30 A
B
0 2
4 6
4 6 8 10 12 14
Control
5 rubbs
10 rubbs
20 rubbs
25% methanol
96% methanol
100 NaCl
Net
K+ fl
ux (
nmol
m–2
s–1
)
Time (min)
Fig. 2. Ion fluxes measured from leaf surface after removal of cuticularwaxes by methanol. (A) kinetics of K+ flux responses of rubbing differenttimes by 25% methanol; (B) kinetics of K+ flux responses of rubbingdifferent times by 96% methanol and soaking in 5 and 20 min in 96%methanol. Mean ± SE (n = 5–8).
Physiol. Plant. 149, 2013 519
–150
–140
–130
–120
–110
–100
–90
–80
–70
–60
Wetcontrol
x 10(96%)
x 25(25%)
x 25(96%)
20 min(96%)
10 h(25%)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Drycontrol
Wetcontrol
x 10(96%)
x 25(96%)
5 min(96%)
20 min(96%)
Fv/
Fm
(ar
b. u
nits
)M
embr
ane
pote
ntia
l (m
V)
Treatment
A
B
Fig. 3. Assaying leaf viability after removal of cuticular waxesby methanol. (A) chlorophyll fluorescence Fv/Fm values. Mean ± SE
(n = 12–15); (B) MP values. Mean ± SE (n = 10–12).
but the latter treatment resulted in significant membranedepolarization (Fig. 3B). Two treatments, namely 10 rubswith 96% methanol, and 25 rubs with 25% methanol,did not result in membrane depolarization (Fig. 3B) andshowed optimal Fv/Fm values (Fig. 3A), suggesting theyare both suitable for the purpose of wax removal. Ofthese two, 10 rubs with 96% methanol resulted in themaximum strength of the K+ flux signal (Fig. 2). There-fore, this treatment was accepted as the most optimaland used in all further studies.
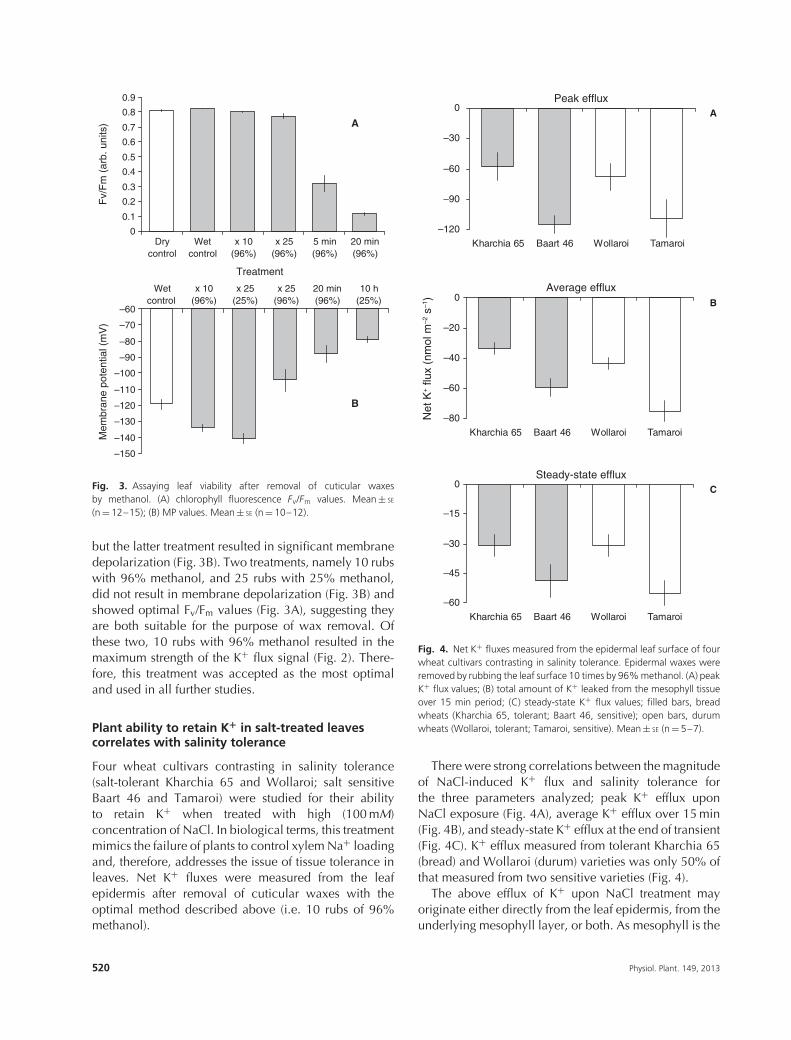
Plant ability to retain K+ in salt-treated leavescorrelates with salinity tolerance
Four wheat cultivars contrasting in salinity tolerance(salt-tolerant Kharchia 65 and Wollaroi; salt sensitiveBaart 46 and Tamaroi) were studied for their abilityto retain K+ when treated with high (100 mM)concentration of NaCl. In biological terms, this treatmentmimics the failure of plants to control xylem Na+ loadingand, therefore, addresses the issue of tissue tolerance inleaves. Net K+ fluxes were measured from the leafepidermis after removal of cuticular waxes with theoptimal method described above (i.e. 10 rubs of 96%methanol).
–120
–90
–60
–30
0
Kharchia 65 Baart 46 Wollaroi Tamaroi
–80
–60
–40
–20
0
Kharchia 65 Baart 46 Wollaroi Tamaroi
–60
–45
–30
–15
0
Kharchia 65 Baart 46 Wollaroi Tamaroi
A
B
C
Net
K+ fl
ux (
nmol
m–2
s–1
)
Peak efflux
Average efflux
Steady-state efflux
Fig. 4. Net K+ fluxes measured from the epidermal leaf surface of fourwheat cultivars contrasting in salinity tolerance. Epidermal waxes wereremoved by rubbing the leaf surface 10 times by 96% methanol. (A) peakK+ flux values; (B) total amount of K+ leaked from the mesophyll tissueover 15 min period; (C) steady-state K+ flux values; filled bars, breadwheats (Kharchia 65, tolerant; Baart 46, sensitive); open bars, durumwheats (Wollaroi, tolerant; Tamaroi, sensitive). Mean ± SE (n = 5–7).
There were strong correlations between the magnitudeof NaCl-induced K+ flux and salinity tolerance forthe three parameters analyzed; peak K+ efflux uponNaCl exposure (Fig. 4A), average K+ efflux over 15 min(Fig. 4B), and steady-state K+ efflux at the end of transient(Fig. 4C). K+ efflux measured from tolerant Kharchia 65(bread) and Wollaroi (durum) varieties was only 50% ofthat measured from two sensitive varieties (Fig. 4).
The above efflux of K+ upon NaCl treatment mayoriginate either directly from the leaf epidermis, from theunderlying mesophyll layer, or both. As mesophyll is the
520 Physiol. Plant. 149, 2013

–350
–300
–250
–200
–150
–100
–50
0Kharchia 65 Baart 46 Wollaroi Tamaroi
100 NaCl
A
B
bb
a
a–160
–140
–120
–100
–80
–60
–40
–20
0Kharchia 65 Baart Wollaroi Tamaroi
–200
–150
–100
–50
0
50
100
Time (min)
Kharchia 65
Baart
Wollaroi
Tamaroi
100 NaCl
C
D
b
a
ab
a
–900
–700
–500
–300
–100
100
0 2 4 6 8 10 12 14 16 0 2 4 6 8 10 12 14 16
Kharchia 65
Baart 46
WollaroiTamaroi
Time (min)
Net
K+ fl
ux (
nmol
m–2
s–1
)A
vera
ge K
+ fl
ux (
nmol
m–2
s–1
)
Net
H+ fl
ux (
nmol
m–2
s–1
)P
eak
H+ fl
ux (
nmol
m–2
s–1
)
Fig. 5. Net K+ (A, B) and H+ (C, D) flux responses measured from the mesophyll tissue of four wheat cultivars contrasting in salinity tolerance (asabove). To expose mesophyll, the excised leaf (middle part) was cut to segments (roughly 5 × 8 mm) by a sharp blade angularly. (A) kinetics of K+ fluxresponses; (B) total amount of K+ leaked from the mesophyll tissue over 15-min period; (C) kinetics of H+ flux responses; (D) peak H+ flux values.Mean ± SE (n = 8–12). Within each treatment different low case letters indicate significant differences by Duncan’s multiple range test at P < 0.05.
most important metabolically active tissue in plants, itsability to retain K+ was expected to be more essential toleaf salinity tissue tolerance. Thus, the extent of NaCl-induced K+ leak from wheat and barley mesophyll wasalso investigated in direct experiments (Figs 5 and 6).
The observed results were consistent with our observa-tions for epidermal tissue. In wheat, the average K+ effluxfrom mesophyll (Fig. 5A, B) was three-fold to four-foldhigher compared with the flux from epidermal cells (Fig.4B). This is hardly surprising, given the lack of blockingeffect of the cuticle on ion fluxes measured from meso-phyll tissue. The qualitative patterns of genetic variabilityin K+ retention in mesophyll were generally consistentwith epidermal data (Fig. 5B). Similar to data from theepidermis, salt-sensitive durum wheat Tamaroi showeda two-fold higher K+ leak from its mesophyll, comparedwith tolerant durum wheat variety Wollaroi (significant atP < 0.05; Fig. 5B). The difference between bread wheatswas also consistent with epidermal data (Fig. 5B).
Previously, a genetic variability in K+ retention inbarley roots was attributed to better control of MP andhigher H+ pump activity (Chen et al. 2007b). In order toinvestigate the relationship between K+ and H+ responsein wheat leaf mesophyll cells under salt stress, NaCl-induced net H+ flux was also measured. Significantlymore H+ flux was pumped out from mesophyll cells ofthe two bread wheat varieties, as compared with durum
wheat (Fig. 5C, D). This is consistent with the generalconsensus that durum wheat is generally more sensitiveto salt than bread wheat (Munns and Tester 2008, Cuinet al. 2009).
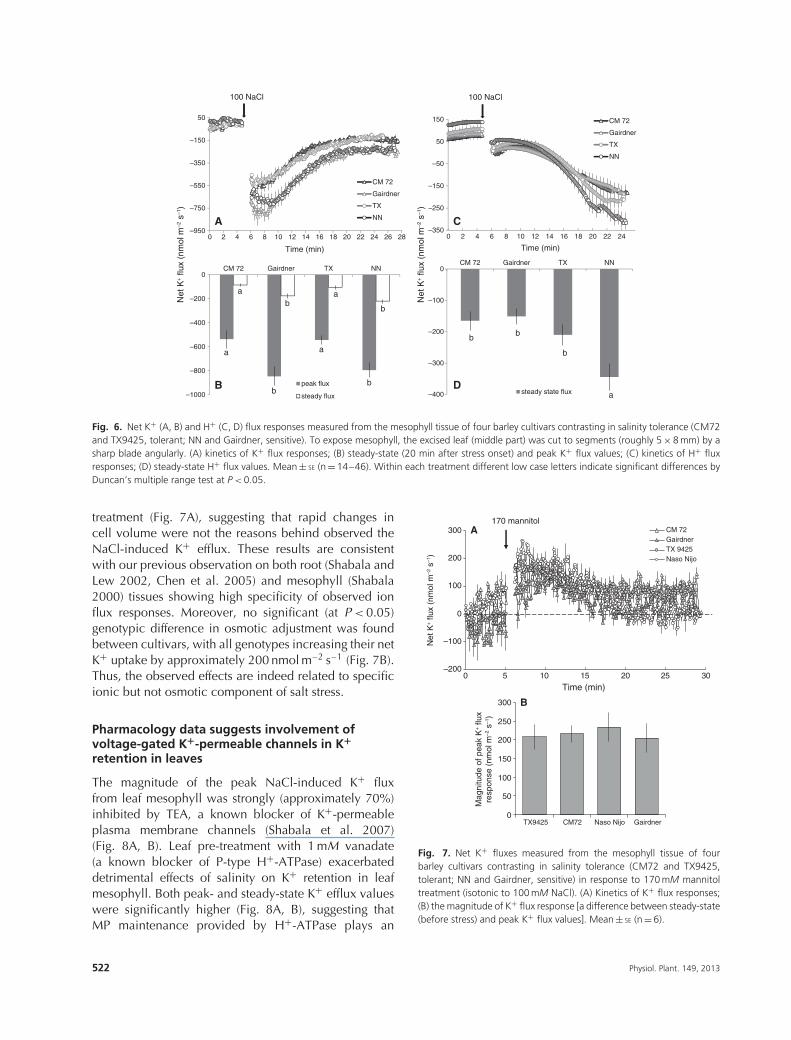
The strong correlation between the magnitude ofNaCl-induced K+ efflux and salinity tolerance was evenmore pronounced in barley (Fig. 6A, B). Both peak andsteady-state K+ fluxes were significantly higher in twosensitive varieties (Gairdner and Naso Nijo) comparedwith two tolerant varieties (CM 72and TX9425). NaCl-induced activation of H+ efflux was reported for allvarieties, but no clear trends were observed for H+ fluxdata (Fig. 6C, D).
The observed effects are related to specific ionicbut not osmotic component of salt stress
Acute 100 mM NaCl treatment causes rapid cellshrinking thus concentrating the internal K+ and othersolutes and creating a chemical gradient favoring passiveK+ leak from the cell. To ensure that the observeddifference in K+ retention is not attributed to geneticvariability in elasticity modules among genotypes but isindeed related to regulation of plasma membrane K+
transporters, additional experiments were undertakenusing 170 mM mannitol (a non-ionic osmolyte isotonicto 100 mM NaCl). Contrary to NaCl treatment, all plantstook up but not lost K+ in response to isotonic mannitol
Physiol. Plant. 149, 2013 521
–950
–750
–550
–350
–150
50
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28
Time (min)
CM 72
Gairdner
TX
NNA
–1000
–800
–600
–400
–200
0CM 72 Gairdner TX NN
peak flux
steady fluxB
100 NaCl 100 NaCl
–350
–250
–150
–50
50
150
0 2 4 6 8 10 12 14 16 18 20 22 24
Time (min)
CM 72
Gairdner
TX
NN
–400
–300
–200
–100
0CM 72 Gairdner TX NN
steady state fluxD
C
aa
a a
b
bb
b
b
b b
a
Net
K+ fl
ux (
nmol
m–2
s–1
)
Net
K+ fl
ux (
nmol
m–2
s–1
)Fig. 6. Net K+ (A, B) and H+ (C, D) flux responses measured from the mesophyll tissue of four barley cultivars contrasting in salinity tolerance (CM72and TX9425, tolerant; NN and Gairdner, sensitive). To expose mesophyll, the excised leaf (middle part) was cut to segments (roughly 5 × 8 mm) by asharp blade angularly. (A) kinetics of K+ flux responses; (B) steady-state (20 min after stress onset) and peak K+ flux values; (C) kinetics of H+ fluxresponses; (D) steady-state H+ flux values. Mean ± SE (n = 14–46). Within each treatment different low case letters indicate significant differences byDuncan’s multiple range test at P < 0.05.
treatment (Fig. 7A), suggesting that rapid changes incell volume were not the reasons behind observed theNaCl-induced K+ efflux. These results are consistentwith our previous observation on both root (Shabala andLew 2002, Chen et al. 2005) and mesophyll (Shabala2000) tissues showing high specificity of observed ionflux responses. Moreover, no significant (at P < 0.05)genotypic difference in osmotic adjustment was foundbetween cultivars, with all genotypes increasing their netK+ uptake by approximately 200 nmol m–2 s–1 (Fig. 7B).Thus, the observed effects are indeed related to specificionic but not osmotic component of salt stress.
Pharmacology data suggests involvement ofvoltage-gated K+-permeable channels in K+retention in leaves
The magnitude of the peak NaCl-induced K+ fluxfrom leaf mesophyll was strongly (approximately 70%)inhibited by TEA, a known blocker of K+-permeableplasma membrane channels (Shabala et al. 2007)(Fig. 8A, B). Leaf pre-treatment with 1 mM vanadate(a known blocker of P-type H+-ATPase) exacerbateddetrimental effects of salinity on K+ retention in leafmesophyll. Both peak- and steady-state K+ efflux valueswere significantly higher (Fig. 8A, B), suggesting thatMP maintenance provided by H+-ATPase plays an
–200
–100
0
100
200
300
0 5 10 15 20 25 30
CM 72GairdnerTX 9425Naso Nijo
Time (min)
170 mannitol
0
50
100
150
200
250
300
A
B
TX9425 CM72 Naso Nijo Gairdner
Net
K+ fl
ux (
nmol
m–2
s–1
)
Mag
nitu
de o
f pea
k K
+ fl
uxre
spon
se (
nmol
m–2
s–1
)
Fig. 7. Net K+ fluxes measured from the mesophyll tissue of fourbarley cultivars contrasting in salinity tolerance (CM72 and TX9425,tolerant; NN and Gairdner, sensitive) in response to 170 mM mannitoltreatment (isotonic to 100 mM NaCl). (A) Kinetics of K+ flux responses;(B) the magnitude of K+ flux response [a difference between steady-state(before stress) and peak K+ flux values]. Mean ± SE (n = 6).
522 Physiol. Plant. 149, 2013

–1600
–1400
–1200
–1000
–800
–600
–400
–200
0
200
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Time (min)
Control
20 mM TEA
1 mM vanadate–250
–150
–50
50
150
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Time (min)
Control
20 mM TEA
1 mM vanadate
CA
100 NaCl 100 NaCl
–300
–200
–100
0
100
200Control 20 mM TEA 1 mM Vanadate
initial flux
flux after adding salt
steady state flux
average flux–1700
–1500
–1300
–1100
–900
–700
–500
–300
–100
100Control 20 mM TEA 1 mM Vanadate
initial flux
peak flux
steady flux
average fluxDB
b
b
b
b
c
cb c
a
a
a a
a
b
c b
bb
b
a
a
a
a
a
Net
K+ fl
ux (
nmol
m–2
s–1
)
Net
H+ fl
ux (
nmol
m–2
s–1
)
Fig. 8. Ion fluxes measured from mesophyll after cutting barley leaf (Gairdner). (A) Transient net K+ fluxes, (B) calculation of K+ fluxes, (C) transientnet H+ fluxes and (D) calculation of H+ fluxes. Mean ± SE (n = 24–30). Within each treatment different low case letters indicate significant differencesby Duncan’s multiple range test at P < 0.05.
important role in controlling K+ fluxes across the mes-ophyll plasma membrane. Consistent with this notion,vanadate treatment significantly reduced the magnitudeof NaCl-induced H+ efflux from mesophyll tissue (Fig.8C, D). Interestingly, however, leaf pre-treatment withTEA also reduced the magnitude of NaCl-induced H+
efflux, suggesting a possibility of a feedback controlover H+-ATPase activity by cytosolic K+.
Discussion
Potassium retention in mesophyll is essential forsalinity tolerance
The previous work in our laboratory has emphasized theimportance of K+ retention in roots of a range of plantspecies, including barley (Chen et al. 2005, 2007b,2007c) and wheat (Cuin et al. 2008, 2012). Here wereport that K+ retention in leaf mesophyll is also essentialto confer salinity stress tolerance in these species (Figs5 and 6). At the cellular level, K+ catalyzes manymetabolic reactions and there are at least 80 enzymesknown that require K+ (Evans and Sorger 1966, Suelter1970); most of these are present in the leaf mesophyll.Thus, inability of mesophyll cells to retain K+ will resultin substantial decline in leaf photochemistry, ultimatelyaffecting plant growth and biomass accumulation. Inaddition to activation of enzymes, K+ is used as a majorcounter ion for cytoplasmic polyanions, control of
photoassimilate loading into phloem, and is essential forthe formation and maintenance of plasma MP and elec-trogenic process in chloroplast membranes (Hedrich andSchroeder 1989, Fang et al. 1995, Marschner et al. 1996,Talbott and Zeiger 1996). Potassium also plays a criticalrole in determining cell fate and transition to PCD, bycontrolling caspase (in animal systems; Hughes andCidlowski 1999) or protease and endonuclease (in plantsystems; Demidchik et al. 2010) activity. It was shownearlier that Arabidopsis gork mutants lacking outwardrectifying depolarization-activated K+ (KOR) channelsand, thus capable to retain more K+ under saline con-ditions, had less than 1/3 of terminal deoxynucleotidyltransferase dUTP nick end labeling (TUNEL) stainednuclei compared with wild-type plants (Demidchik et al.2010). We believe that a similar scenario is applicablein the case of leaf mesophyll in wheat and barley, andthat salt-induced damage to leaf lamina observed asnecrotic spots on leaf tips in wheat and barley may bea result of mesophyll cells undergoing PCD as a resultof K+ loss. Direct quantification of PCD-related events(such as cytochrome C release; nuclei TUNEL stainingor DNA laddering) is needed to support this hypothesis.
Downstream targets
Several types of K+-permeable channels at the mesophyllplasma membrane may mediate K+ efflux uponsalinity stress (Shabala 2003).This includes Shaker-like
Physiol. Plant. 149, 2013 523

outward-rectifying K+-selective KOR channels andnon-selective cation channels (NSCC). KOR channels(GORK in Arabidopsis) are activated by membranedepolarization at potentials more positive than Ek (Hosyet al. 2003, Very and Sentenac 2003). NSCCs appearto form a large, heterogeneous group of channels, with40 putative NSCCs revealed in the Arabidopsis genomesequence (Maser et al. 2001, Demidchik et al. 2002).Most of these are only weakly voltage-dependent butmay be activated by other factors such as mechanicaltension, calcium, cyclic nucleotides and glutamate(Demidchik et al. 2002, Demidchik and Maathuis2007). NSCCs discriminate poorly between K+ and Na+
(K+/Na+ selectivity ratios between 0.3 and 3, Demidchiket al. 2002) and, thus, could mediate both Na+ uptakeand K+ efflux under saline conditions.
It was shown that acute NaCl treatment resultsin substantial depolarization of plasma membrane inleaf mesophyll (e.g. beans, Shabala 2000; Arabidopsis,Shabala et al. 2006; tobacco, Shabala et al. 2007). Thus,accumulation of substantial amounts of NaCl in theleaf apoplast may activate KOR channels and result inK+ leakage from the mesophyll. Our pharmacologicaldata reported here suggests that this scenario is alsotrue for barley mesophyll. Indeed, K+ leakage wassubstantially suppressed by using TEA, a known blockerof KOR channels (Fig. 8) but enhanced in leavespre-treated with orthovanadate, a known inhibitorof P2B type of H+-ATPase. H+-ATPase is primarilyresponsible for maintenance of plasma MP (Sze et al.1999), and suppression of its activity prevents plantability to restore (otherwise depolarized) MP, resultingin larger K+ leakage via depolarization-activatedKOR channels.
The overall amount of K+ leaked from wheatmesophyll was substantially smaller compared withappropriate amount leaked from barley (Figs 5 and6 for comparison). This is somewhat counterintuitive,given the fact that barley is always classified as moretolerant to salinity compared with wheat (Greenwayand Munns 1980). However, these findings are in goodagreement with our early observations on wheat andbarley roots. Indeed, the magnitude of NaCl-inducedK+ loss from wheat roots was only approximately 10%of that measured in barley under similar conditions(Cuin et al. 2008). Moreover, while K+ retention inbarley roots showed a strong positive correlation withactivity of the plasma membrane H+-ATPase and itsability to maintain more negative MP (Chen et al.2007b), no such correlation was found in wheat (Cuinet al. 2008). It may be suggested, therefore, that thepathway of K+ leakage from both root and leaf tissuesdiffer substantially between wheat and barley. In barley,
voltage-gated KOR channels are obviously the dominantmechanisms (Chen et al. 2007b) and these are activatedimmediately upon acute NaCl treatment as discussedabove. In wheat, the major bulk of K+ leakage may occurthrough NSCC. These channels show only weak voltagedependence (Demidchik and Maathuis 2007) and assuch, are not expected to be dramatically activated byNa+ entering the cell. However, NSCC channels areknown to be directly activated by various forms of ROS(Demidchik et al. 2003, 2007), and ROS accumulationin both plant root and leaf tissues (Cavalcanti et al.2007) is widely reported under saline conditions.Moreover, ROS accumulation is a much slower processcompared with the almost instantaneous membranedepolarization upon acute saline stress. As such, ROS-induced activation of NSCC will be more gradualand take much longer then instantaneous activation ofvoltage-gated KOR channels. This may explain larger K+
leakage from barley compared with wheat tissues underconditions of acute salt stress in our experiments. Underlong-term stress conditions, however, ROS-induced K+
leak may become more essential, which may explaingenerally higher salt sensitivity of wheat compared withbarley (Munns and Tester 2008).
Suitability for screening
Our finding of the importance of K+ retention in leafmesophyll for salinity tolerance in cereals may be ofsignificant importance to plant breeders. To be usefulin MAS-based breeding programs, however, this traitshould be reliably estimated using some relatively inex-pensive and user-friendly high-throughput technology.The non-invasive MIFE technique shows great potentialof development as such a high-throughput tool.
In this work, ion flux responses were measuredfrom both epidermal and mesophyll tissues, withconsistent results. For obvious reasons, measuring NaCl-induced K+ efflux in mesophyll is easier to conduct,as these measurements eliminate a requirement foradditional labor- and time-consuming procedures ofremoving epicuticular waxes. Cutting a leaf segmentand exposing mesophyll tissue takes only a few seconds;immobilizing these segments in prefabricated Perspexholders is also a matter of seconds and requires nospecial skills.
Net K+ fluxes were measured in this study for upto 30 min upon NaCl addition. However, for screeningpurposes, much shorter (e.g. 2–3 min) measurementswill suffice. Indeed, peak K+ flux responses occurred atapproximately 1.5 min after NaCl treatment (Fig. 6B), andpeak values were proven to be a sensitive physiologicalindex for tissue tolerance in barley (Fig. 6). Thus,
524 Physiol. Plant. 149, 2013

even allowing several minutes for electrode positioningand manipulation in the chamber, one completerecord will require no longer then 6–7 min (includingimmobilization time). The current MIFE configurationallows parallel recording of up to three specimens; thisamounts into 25–30 specimens per hour, or 160–200specimens per standard business day. Moreover, existingMIFE electronics has an eight-channel amplifier, and theabove limitation of three channels is caused merely bythe type of manipulator used for electrode positioning.For a relatively modest cost, the latter can be automated(e.g. using robotic arm) enabling eight specimens to beprocessed simultaneously. Altogether, it will increasethe screening capacity to over 500 specimens perday, which is large enough to be termed as highthroughput.
Acknowledgements – This work was supported by the GrainResearch and Development Corporation grant to S. S. andM. Z. and by the Australian Research Council Discoverygrant to S. S. We acknowledge Associate Professor NoelDavies for performing the GC-MS of foliar waxes andDr Karsten Goemann for assistance with ESEM (both atthe Central Science Laboratory, University of Tasmania).Associate Professor John Ross is thanked for his assistancewith sample derivatization.
References
Agrawal PB, Nierstrasz VA, Klug-Santner BG, Gubitz GM,Lenting HBM, Warmoeskerken MMCG (2007) Waxremoval for accelerated cotton scouring with alkalinepectinase. Biotechnol J 2: 306–315
Bewick TA, Shilling DG, Querns R (1993) Evaluation ofepicuticular wax removal from whole leaves withchloroform. Weed Technol 7: 706–716
Blumwald E, Aharon GS, Apse MP (2000) Sodium transportin plant cells. Biochim Biophys Acta 1465: 140–151
Cavalcanti FR, Lima JPMS, Ferreira-Silva SL, Viegas RA,Silveria JAG (2007) Roots and leaves display contrastingoxidative response during salt stress and recovery incowpea. J Plant Physiol 164: 591–600
Chen Z, Cuin TA, Zhou M, Twomey A, Naidu BP, ShabalaS (2007a) Compatible solute accumulation andstress-mitigating effects in barley genotype contrasting intheir genotype. J Exp Bot 58: 4245–4255
Chen Z, Pottosin II, Cuin TA, Fuglsang AT, Tester M, Jha D,Zepeda-Jazo I, Zhou MX, Palmgren MG, Newman IA,Shabala S (2007b) Root plasma membrane transporterscontrolling K+/Na+ homeostasis in salt-stressed barley.Plant Physiol 145: 1714–1725
Chen Z, Newman I, Zhou M, Mendham N, Zhang G,Shabala S (2005) Screening plants for salt tolerance bymeasuring K+ flux: a case study for barley. Plant CellEnviron 28: 1230–1246
Chen Z, Zhou M, Newman IA, Mendham NJ, Zhang GP,Shabala S (2007c) Potassium and sodium relations insalinised barley tissues as a basis of differential salttolerance. Funct Plant Biol 34: 150–162
Cuin TA, Betts SA, Chalmandrier R, Shabala S (2008) Aroot’s ability to retain K+ correlates with salt tolerance inwheat. J Exp Bot 59: 2697–2706
Cuin TA, Bose J, Stefano G, Jha D, Tester M, Mancuso S,Shabala S (2011) Assessing of root plasma membraneand tonoplast Na+/H+ exchangers in salinity tolerancein wheat: in planta quantification methods. Plant CellEnviron 34: 947–961
Cuin TA, Tian Y, Betts SA, Chalmandrier R, Shabala S(2009) Ionic relations and osmotic adjustment in durumand bread wheat under saline conditions. Funct PlantBiol 36: 1110–1119
Cuin TA, Zhou M, Parsons D, Shabala S (2012) Geneticbehaviour of physiological traits conferring K+/Na+
homeostasis in wheat. Plant Biol 14: 438–446Davies MJ, Poole RJ, Rea PA, Sanders D (1992) Potassium
transport into plant vacuoles energized directly by aproton-pumping inorganic pyrophosphatase. Proc NatlAcad Sci USA 89: 11701–11705
Demidchik V, Cuin TA, Svistunenko D, Smith SJ, Miller AJ,Shabala S, Sokolik A, Yurin V (2010) Arabidopsis rootK+-efflux conductance activated by hydroxyl radicals:signal-channel properties, genetic basis and involvementin stress-induced cell death. J Cell Sci 123: 1468–1479
Demidchik V, Davenport RJ, Tester M (2002) Nonselectivecation channels in plants. Annu Rev Plant Biol 53:67–107
Demidchik V, Shabala SN, Coutts KB, Tester MA, DaviesJM (2003) Free oxygen radicals regulate plasmamembrane Ca2+- and K+-permeable channels in plantroot cells. J Cell Sci 116: 81–88
Demidchik V, Shabala SN, Davies JM (2007) Spatialvariation in H2O2 response of Arabidopsis thaliana rootepidermal Ca2+ flux and plasma membrane Ca2+
channels. Plant J 49: 377–386Demidchik V, Maathuis JM (2007) Physiological roles of
nonselective cation channels in plant: from salt stress tosignalling and development. New Phytol 175: 387–404
Dreyer I, Uozum N (2011) Potassium channels in plantcells. FEBS J 278: 4293–4303
Eleuch L, Jilal A, Grando S, Ceccarelli S, von Korff SM,Tsujimoto H, Hajer A, Daaloul A, Baum M (2008)Genetic diversity and association analysis for salinitytolerance, heading date and plant height of barleygermplasm using simple sequence repeat markers. JIntegr Plant Biol 50: 1004–1014
Evans HJ, Sorger GJ (1966) Role of mineral elements withemphasis on the univalent cations. Annu Rev PlantPhysiol 17: 47–76
Physiol. Plant. 149, 2013 525

Fang Z, Mi F, Berkowitz GA (1995) Molecular andphysiological analysis of a thylakoid K+ channelprotein. Plant Physiol 108: 1725–1734
Flowers TJ (2004) Improving crop salt tolerance. J Exp Bot55: 307–319
Greenway H, Munns R (1980) Mechanisms of salttolerance in nonhalophytes. Annu Rev Plant Physiol 31:149–190
Hedrich R, Schroeder JI (1989) The physiology of ionchannels and electrogenic pumps in higher plants. AnnuRev Plant Physiol 40: 539–569
Horie T, Schroeder JI (2004) Sodium transporters in plants.Diverse genes and physiological functions. Plant Physiol136: 2457–2462
Hosy E, Vavasseur A, Mouline K, Dreyer I, Gaymard F,Poree F, Boucherez J, Lebaudy A, Bouchez D, Very A,Simonneau T, Thibaud J, Sentenac H (2003) TheArabidopsis outward K+ channel GORK is involved inregulation of stomatal movements and planttranspiration. Proc Natl Acad Sci USA 100: 5549–5554
Hughes FM, Cidlowski JA (1999) Potassium is a criticalregulator of apoptotic enzymes in vitro and in vivo. AdvEnzyme Regul 39: 157–171
James RA, Rivelli AR, Munns R, von Caemmerer S (2002)Factors affecting CO2 assimilation, leaf injury andgrowth in salt-stressed durum wheat. Funct Plant Biol29: 1393–1403
Koch K, Barthlott W, Koch S, Hommes A, Wandelt K,Mamdouh W, De-Feyter S, Broekmann P (2006)Structural analysis of wheat wax (Triticum aestivum, c.v.‘Naturastar’ L.): from the molecular level to threedimensional crystals. Planta 223: 258–270
Maser P, Thomine S, Schroeder JI, Ward JM, Hirschi K, SzeH, Talke IN, Amtmann A, Maathuis FJM, Sanders D,Harper JF, Tchieu J, Gribskov M, Persans MW, Salt DE,Kim SA, Guerinot ML (2001) Phylogenetic relationshipswithin cation transporter families of Arabidopsis. PlantPhysiol 126: 1646–1667
Maathuis F, Amtmann A (1999) K+ nutrition and Na+
toxicity: the basis of cellular K+/Na+ ratios. Ann Bot 84:123–133
Mano Y, Takeda K (1997) Mapping quantitative trait locifor salt tolerance at germination and the seedling stagein barley (Hordeum vulgare L.). Euphytica 94: 263–272
Marschner H, Kirkby EA, Cakmak I (1996) Effect of mineralnutritional status on shoot-root partitioning ofphotoassimilates and cycling of mineral nutrients. J ExpBot 47: 1255–1263
Munns R, James RA, Lauchli A (2006) Approaches toincreasing the salt tolerance of wheat and other cereals.J Exp Bot 57: 1025–1043
Munns R, Tester M (2008) Mechanism of salinitytolerance. Annu Rev Plant Biol 59: 651–681
Newman IA (2001) Ion transport in plants: measurement offluxes using ion-selective microelectrodes to
characterize transporter function. Plant Cell Environ 24:1–14
Osaki M, Shinano T, Tadano T (1993) Effect of nitrogen,phosphorus, or potassium deficiency on theaccumulation of ribulose-1,5-bisphosphatecarboxylase/oxygenase and chlorophyll in several fieldcrops. Soil Sci Plant Nutr 39: 417–425
Plett D, Safwat G, Gilliham M, Møller IS, Roy S, Shirley N,Jacobs A, Johnson A, Tester M (2010) Improved salinitytolerance of rice through cell type-specific expression ofAtHKT1;1. PLoS One 5: e12571
Rengasamy P (2010) Soil processes affecting cropproduction in salt-affected soils. Funct Plant Biol 37:613–620
Rhee Y, Hlousek-Radojcic A, Ponsamuel J, Liu D,Post-Beittenmiller D (1998) Epicuticular waxaccumulation and fatty acid elongation activities areinduced during leaf development of leeks. Plant Physiol116: 901–911
Shabala S (2000) Ionic and osmotic components of saltstress specifically modulate net ion fluxes from beanmesophyll. Plant Cell Environ 23: 825–837
Shabala S (2003) Regulation of potassium transport inleaves: from molecular to tissue level. Ann Bot 92:627–634
Shabala S, Cuin TA (2008) Potassium transport and plantsalt tolerance. Physiol Plant 133: 651–669
Shabala S, Cuin TA, Prismall L, Nemchinov LG (2007)Expression of animal CED-9 anti-apoptotic gene intobacco modifies plasma membrane ion fluxes inresponse to salinity and oxidative stress. Planta 227:189–197
Shabala S, Demidchik V, Shabala L, Cuin TA, Smith SJ,Miller AJ, Davies JM, Newman IA (2006) ExtracellularCa2+ ameliorate NaCl induced K+ loss from Arabidopsisroot and leaf cells by controlling plasma membraneK+-permeable channels. Plant Physiol 141: 1653–1665
Shabala S, Lew RR (2002) Turgor regulation in osmoticallystressed Arabidopsis epidermal root cells. Direct supportfor the role of inorganic ion uptake as revealed byconcurrent flux and cell turgor measurements. PlantPhysiol 129: 290–299
Shabala S, Newman I (1999) Light-induced changes inhydrogen, calcium, potassium, and chloride ion fluxesand concentrations from mesophyll and epidermaltissues of bean leaves. Understanding the ionic basis oflight-induced bioelectrogenesis. Plant Physiol 119:1115–1124
Shabala S, Shabala L (2002) Kinetics of net H+, Ca2+, K+,Na+, NH4
+, and Cl– fluxes associated with post-chillingrecovery of plasma membrane transporters in Zea maysleaf and root tissues. Physiol Plant 114: 47–56
Smethurst CF, Shabala S (2003) Screening methods forwaterlogging tolerance in lucerne: comparative analysisof waterlogging effects on chlorophyll fluorescence,
526 Physiol. Plant. 149, 2013

photosynthesis, biomass and chlorophyll content. FunctPlant Biol 30: 335–343
Suelter CH (1970) Enzymes activated by monovalentcations. Science 168: 789–795
Sze H, Li X, Palmgren MG (1999) Energization of plant cellmembranes by H+-pumping ATPase: regulation andbiosynthesis. Plant Cell 11: 677–689
Talbott LD, Zeiger E (1996) Central roles for potassium andsucrose in guard-cell osmoregulation. Plant Physiol 111:1051–1057
Tegg R, Melian L, Wilson CR, Shabala S (2005) Plant cellgrowth and ion flux responses to the streptomycetephytotoxin thaxtomin A: calcium and hydrogen fluxpatterns revealed by the non-invasive MIFE technique.Plant Cell Physiol 46: 638–648
Very A, Sentenac H (2003) Molecular mechanisms andregulation of K+ transport in higher plants. Annu RevPlant Biol 54: 575–603
Xu R, Wang J, Li C, Johnson P, Lu C, Zhou M (2012) Asingle locus is responsible for salinity tolerance in aChinese landrace barley (Hordeum vulgare L.). PLoSOne 7: e43079
Zhou G, Johnson P, Ryan PR, Delhaize E, Zhou M (2012)Quantitative trait loci for salinity tolerance in barley(Hordeum vulgare L.). Mol Breed 29: 427–436
Zivanovic BD, Pang J, Shabala S (2005) Light-inducedtransient ion flux response from maize leaves and theirassociation with leaf growth and photosynthesis. PlantCell Environ 28: 340–352
Edited by J. K. Schjørring
Physiol. Plant. 149, 2013 527
Copyright © 2022 FDOKUMEN




















