
collision-induced dissociation of hypersolvated lanthanide ...

Double Dissociation Between the Contributions of the Septaland Temporal Hippocampus to Spatial Learning:
The Role of Prior Experience
Livia de Hoz* and Stephen J. Martin*
ABSTRACT: The mammalian hippocampus is anatomically heteroge-neous along its longitudinal axis, and there is evidence that distinctfunctions are executed by different septotemporal subregions. The bestdocumented example is the dependency of spatial learning on the sep-tal, but not the temporal, hippocampus. Here, we carried out a water-maze memory task in rats with partial lesions of the septal or temporalhippocampus made either before or after training. We then studiedmemory retention, reversal, and new spatial learning in a novel environ-ment. This resulted in the surprising finding that spatial learning in anew environment is dependent on the temporal hippocampus in ratswith preoperative experience of a different pool. Rats with septal hippo-campal lesions made after learning not only retained the focused searchstrategy that was acquired during preoperative training, but were alsocapable of rapid spatial learning in a second pool. This demonstratesthat once spatial information has been acquired in one context, relatednew learning in a different context can be mediated by the temporalhippocampus, a result that challenges the widely held view that spatialmemory is an exclusive function of the septal hippocampus. VC 2014Wiley Periodicals, Inc.
KEY WORDS: dorsal; ventral; pretraining; procedural; water maze
INTRODUCTION
There is mounting evidence for a division of labor along the longitu-dinal (septotemporal or dorsoventral) axis of the hippocampus. Anatom-ical heterogeneities along this axis have long been recognized (Amaraland Witter, 1989; Poppenk et al., 2013): the septal hippocampusreceives its input from the dorsolateral band of the entorhinal cortex,itself a target primarily for neocortical visual and spatial information;the temporal hippocampus is innervated by the ventromedial entorhinal
cortex, a region that receives less visuospatial informa-tion, but acquires substantial inputs from, for exam-ple, the insular and piriform cortices (Kerr et al.,2007) and the amygdala (Van Groen and Wyss,1990). Consistent with this pattern of connectivity,unit-recording studies reveal a dorsoventral gradient inthe scale of spatial representation within both entorhi-nal cortex and hippocampus: entorhinal grid cellsexhibit an increasing expansion of spatial scale fromdorsolateral to ventromedial locations (Brun et al.,2008), hippocampal place fields become progressivelylarger along the septotemporal axis of the structure(Jung et al., 1994; Kjelstrup et al., 2008), and ventralCA1 pyramidal cells are more excitable (Doughertyet al., 2012). As well as differences in afferent connec-tivity, efferents from septal and temporal regions ofthe hippocampus target distinct downstream struc-tures. For example, there are extensive projectionsfrom the temporal—but not the septal—hippocampusto the prefrontal cortex and ventral striatum, regionsthat play key roles in emotion, motivation, and goal-directed action; for review, see Bannerman et al.(2004); Bast (2007) and Bast (2011); van der Meerand Redish (2011b).
Preliminary evidence for a corresponding differentia-tion of function along the longitudinal axis of the hip-pocampus was presented by Nadel (1968). The ideathat septal and temporal hippocampus might be func-tionally distinct was later explored in detail by Moseret al. (1993, 1995) who found that the septal, but notthe temporal, hippocampus is necessary for spatiallearning in a watermaze reference-memory task (seeMoser and Moser, 1998a, for review); however, thetemporal hippocampus may play a role in spatial learn-ing under certain circumstances (de Hoz et al., 2003;Ferbinteanu et al., 2003; Broadbent et al., 2004; Lour-eiro et al., 2011). In contrast, the temporal hippocam-pus has been consistently implicated in affective andmotivational processes, consistent with its anatomicalconnectivity (see Barkus et al., 2010; Fanselow andDong, 2010, for review). At face value, these findingssupport a double dissociation of function within thehippocampus. However, rather than playing mutuallyexclusive roles in different forms of memory, the twopoles of the hippocampus may also work in conjunctionin many situations, performing distinct operationswithin a single cognitive domain—cf. Small (2002).
Centre for Cognitive and Neural Systems (CCNS), University of Edin-burgh, Edinburgh, Scotland, United KingdomAdditional Supporting Information may be found in the online version ofthis article.Grant sponsor: MRC Research Fellowship held by LdH; Grant number:G81/478; Grant sponsor: MRC Programme Grant held by RGM Morris;Grant number: G9200370.*Correspondence to: Livia de Hoz; Department of Neurogenetics, MaxPlanck Institute for Experimental Medicine, Hermann-Rein-Str. 3, 37075G€ottingen, Germany. E-mail: [email protected] and Stephen J. Martin;Division of Neuroscience, Medical Research Institute, Ninewells Hospitaland Medical School, University of Dundee, Dundee, DD1 9SY, Scotland,UK. E-mail: [email protected] for publication 9 April 2014.DOI 10.1002/hipo.22285Published online 7 May 2014 in Wiley Online Library(wileyonlinelibrary.com).
VC 2014 WILEY PERIODICALS, INC.
HIPPOCAMPUS 24:990–1005 (2014)

In a previous study of memory consolidation involving hip-pocampal lesions (Martin et al., 2005), we gained the impres-sion that prior watermaze experience might have a differentialeffect on the performance of rats with septal and temporal hip-pocampal lesions. Prior experience can, after all, have profoundeffects on the impact of a variety of manipulations (Bannermanet al., 1995; Saucier and Cain, 1995; Moser and Moser,1998b; Otnaess et al., 1999; Wang et al., 2012a; Inglis et al.,2013). In the present study, we further explore the influence ofprior spatial training on the ability of rats with septal or tem-poral hippocampal lesions to acquire new spatial information.
MATERIALS AND METHODS
Subjects
Adult male Lister-hooded rats (n 5 99) obtained from acommercial supplier (Charles River, UK) were used as subjects.They were pair-housed with ad libitum access to food andwater, and maintained on a 12-h light/12-h dark cycle. Theanimals’ care and maintenance and all experimental procedureswere carried out in accordance with the Animals (ScientificProcedures) Act 1986, and subject to the European Commun-ities Council Directive of 24 November 1986 (86/609/EEC)and local ethical review at the University of Edinburgh.
Apparatus
Behavioral testing was carried out in two watermazes. The“upstairs” pool was used for training and retention testing andthe “downstairs” pool was used to assess new learning in anovel environment. Both “upstairs” and “downstairs” water-mazes were circular pools, approximately 2 m in diameter, 0.6m deep, and filled with water at 25 6 1�C to a depth of 30cm. Escape from the water was provided by an “Atlantisplatform” (Spooner et al., 1994) which became available by ris-ing from the bottom of the pool only if the animals swamwithin a specified “dwell radius” of the correct location for apredetermined “dwell time.” When raised, the top surface ofthe platform remained 1.5 cm below the water surface. Inorder to render the hidden platform invisible, the water wasmade opaque by the addition of 200 ml of latex liquid. Eachpool was located in the center of a different, well-lit room con-taining prominent and distinctive combinations of extra-mazecues, such as wall posters, metal racks, cupboards, etc. Arecessed video camera was mounted on the ceiling to monitorswim paths, and the video signal was relayed to a videorecorder and an image analyzer located in an adjacent room.Coordinates were sampled using an on-line data acquisitionsystem (“WaterMaze”; Actimetrics, Wilmette, IL) that recordsthe path taken by an animal and calculates a number of per-formance measures such as escape latency, swim speed, andpercentage time in each quadrant of the pool. The experi-menter was blind to the rats’ lesion groups while running thebehavior.
Cued pretraining
One day of visually cued non-spatial pretraining was con-ducted in the “upstairs” watermaze, with curtains drawn aboutthe pool to occlude extramaze cues. Each rat received 10spaced trials; the location of the platform was indicated by avisible cue suspended approximately 25 cm above the watersurface. Trials were carried out in two blocks of five trials each(inter-trial interval � 20 min; inter-block interval � 3 h). Theplatform was moved every two trials to one of four possiblelocations, according to a quasi-random schedule (examplesequence: NW, NW, SE, SE, NE for the first five-trial block;NE, NE, SW, SW, NW for the second block); the dwell radiuswas set at 20 cm and the dwell time was always 1 s.
Training
Spatial reference memory training began 3 days later inthe same “upstairs” watermaze (see Fig. 1). This phase com-prised 10 trials per day, in two sessions of 5 consecutive trialseach (inter-session interval � 2 h), for 4 days. Rats weregiven up to 120 s to find an escape platform located in eitherthe NE or SW quadrant of the pool (fixed for a given rat,and counterbalanced across groups). The dwell radius wasalways 20 cm, but the dwell time was gradually increasedover days as follows: day 1: 1.0 s; day 2: 1.5 s; day 3: 2.0 s;day 4: 2.5 s. This schedule was intended to promote highlypersistent and accurate searching over the platform location(see Riedel et al., 1999; Micheau et al., 2004). Rats wereplaced into the pool facing the side-wall; after locating theescape platform, they remained on the platform for 30 sbefore being removed by the experimenter. On the rare trialsin which a rat failed to escape within 2 min, the experi-menter placed a hand above the correct location in order toguide the animal to the platform. For each animal, the plat-form position (NE or SW) remained constant throughouttraining, but start locations (N, S, E, or W) were variedquasi-randomly across trials.
Surgery
Surgery was carried out either 2 weeks prior to the start ofcued pretraining (“pre-training surgery” groups; Fig. 1A) or 1–2 days after the end of training (and 2 weeks prior to the startof retention testing; “post-training surgery” groups; Fig. 1B).Rats were assigned to either sham, septal hippocampal, or tem-poral hippocampal lesion groups based on escape latencies dur-ing the final day of training, such that the different lesiongroups had equivalent mean performance. Temporal hippocam-pal lesions targeted the temporal two thirds of the hippocam-pus, sparing the septal third of the structure, whereas septalhippocampal lesions targeted the septal two thirds of the struc-ture. The terms “septal” and “temporal” are used throughoutin preference to “dorsal” and “ventral” owing to the large sizeof our lesions—a septal lesion for example might have involvedthe removal of some ventral, as well as dorsal, hippocampaltissue.
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 991
Hippocampus
Lesions were made with ibotenic acid [Biotechnology, CA;dissolved in phosphate-buffered saline (pH 7.4) at 10 mg/mL]following the protocol of Jarrard (1989). Rats were anesthetizedwith 2% tribromoethanol (10 mL/kg i.p.) and placed in aKopf (Tujunga, CA) stereotaxic frame such that bregma andlambda were on a horizontal plane. Several injections of ibo-tenic acid (10 for septal lesions; 9 for temporal lesions) weremade at different rostrocaudal and dorsoventral levels via a 1lL microsyringe securely attached to the frame by a Kopf ste-
reotaxic arm. Ibotenic acid (0.05 lL, 0.08 lL or 0.1 lL perinjection) was injected at a rate of 0.1 lL/min, beginning 30 safter the needle was lowered. The needle was removed veryslowly 90 s after the injection. A total of 0.6 lL per hemi-sphere was necessary for both types of lesion. The co-ordinateswere modified from Jarrard (1989) to suit the slightly differentbrain size of Lister-hooded rats; see de Hoz et al. (2003) for afull account of the technique used. Sham lesions were made inthe same way, but injections were replaced with piercing of the
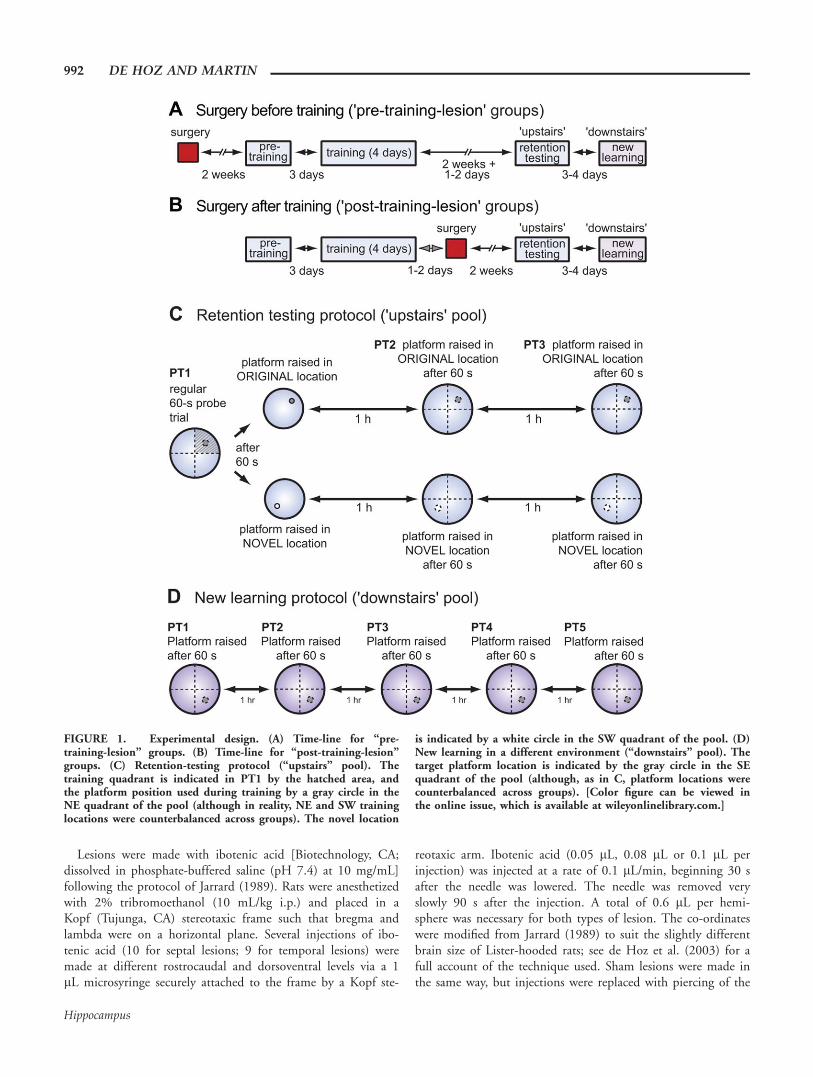
FIGURE 1. Experimental design. (A) Time-line for “pre-training-lesion” groups. (B) Time-line for “post-training-lesion”groups. (C) Retention-testing protocol (“upstairs” pool). Thetraining quadrant is indicated in PT1 by the hatched area, andthe platform position used during training by a gray circle in theNE quadrant of the pool (although in reality, NE and SW traininglocations were counterbalanced across groups). The novel location
is indicated by a white circle in the SW quadrant of the pool. (D)New learning in a different environment (“downstairs” pool). Thetarget platform location is indicated by the gray circle in the SEquadrant of the pool (although, as in C, platform locations werecounterbalanced across groups). [Color figure can be viewed inthe online issue, which is available at wileyonlinelibrary.com.]
992 DE HOZ AND MARTIN
Hippocampus
dura, a procedure intended to cause comparable neocorticaldamage.
Retention Testing (“Upstairs” Pool)
This phase began 15 days after the last training day. Eachanimal received three probe trials (PT1–3) at 1-h intervals.
The first probe, a 60-s swim with the platform unavailable,provided a conventional measure of memory retention. After60 s, the platform was raised in the original training locationfor some rats (the “original” sub-groups), providing an oppor-tunity for reminding or re-learning; or in a novel location inthe opposite quadrant of the pool for the others (the “novel”sub-groups), providing an opportunity for new learning (Fig.1C). The idea behind this testing protocol is that chance per-formance on PT1 is not always the result of a lack of memory.Reminding an animal of the correct location by allowing it tofind the platform can sometimes lead to a rapid reinstatementof an apparently lost memory (de Hoz et al., 2004). To controlfor new leaning, as distinct from reminding, it is necessary touse an additional sub-group of rats that are exposed to a novellocation to provide a measure of new learning—or reversallearning if the animals retain a memory of the original locationand experience proactive interference. Using this protocol, bet-ter performance in the sub-group exposed to the original loca-tion, relative to animals exposed to a new location, isinterpreted as memory or reminding of the location learnedduring training. After the platform was raised, rats wereallowed a further 60 s to locate and climb onto it, and allowedto remain there for 30 s; note that although risen, the platformwas still hidden below the water surface. If the platform wasnot found within the allotted time, its location was indicatedby the experimenter. Rats were placed into the pool in eitherthe adjacent left (AL) or adjacent right (AR) quadrants withrespect to the training location. Start positions were counterbal-anced both across probe trials and across rats. PT2 and PT3were carried out in exactly the same way, one hour after eachother.
New Learning (“Downstairs” Pool)
This was assessed 3 days later in a novel environment (the“downstairs” watermaze). As before, platform locations andstart positions were counterbalanced across trials and acrossanimals. As all rats were initially unfamiliar with the targetplatform location, as well as the testing room, rats assigned to“novel” and “original” sub-groups for “upstairs” retention test-ing were considered as a single group in the analysis of“downstairs” new learning. The protocol was identical to thatof retention testing except that five rewarded probe trials werecarried out (Fig. 1D).
Analysis of “Focused Searching”
Search patterns during probe trials were typically character-ized by periods of wide, sweeping exploration covering largeareas of the pool, punctuated by episodes of tight, focusedsearching in a single location, sometimes around the platformlocation, sometimes elsewhere; see Riedel et al. (1999);Micheau et al., (2004). In order to quantify this behavior dur-ing the first upstairs and downstairs probe trials, we sampledthe x-y coordinates of the rats’ swim paths at 10 Hz over thefirst 60 s (i.e., a total of 600 coordinates), and conducted ananalysis of focused searching on successive rolling 5-s windows
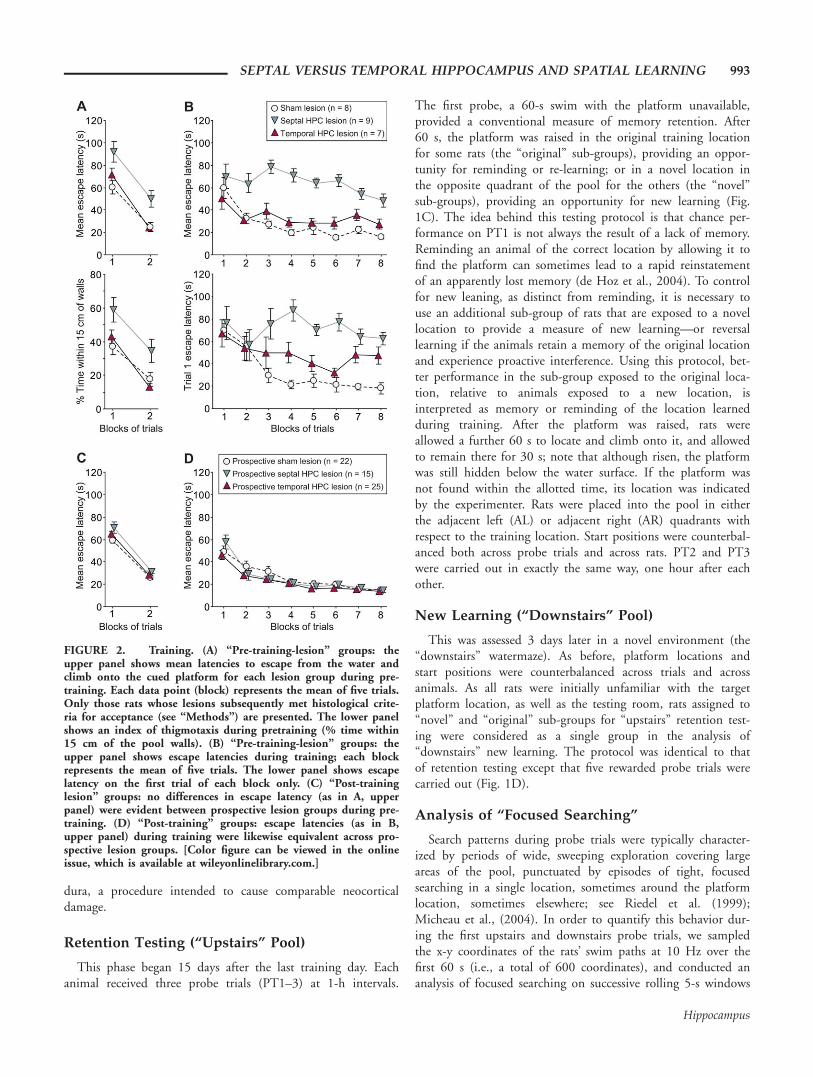
FIGURE 2. Training. (A) “Pre-training-lesion” groups: theupper panel shows mean latencies to escape from the water andclimb onto the cued platform for each lesion group during pre-training. Each data point (block) represents the mean of five trials.Only those rats whose lesions subsequently met histological crite-ria for acceptance (see “Methods”) are presented. The lower panelshows an index of thigmotaxis during pretraining (% time within15 cm of the pool walls). (B) “Pre-training-lesion” groups: theupper panel shows escape latencies during training; each blockrepresents the mean of five trials. The lower panel shows escapelatency on the first trial of each block only. (C) “Post-traininglesion” groups: no differences in escape latency (as in A, upperpanel) were evident between prospective lesion groups during pre-training. (D) “Post-training” groups: escape latencies (as in B,upper panel) during training were likewise equivalent across pro-spective lesion groups. [Color figure can be viewed in the onlineissue, which is available at wileyonlinelibrary.com.]
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 993
Hippocampus

(550 in total). For each window, if the standard deviation ofposition in both the x and y directions was less than 15 cm,then the rat was deemed to be searching in a dense, focusedmanner. The threshold value of 15 cm was chosen because itresulted in a measure that accorded very well with subjectiveestimates of the occurrence of focused searching. Finally, thepercentage of 5-s windows in which focused searching occurredwas calculated.
Perfusion, Histology, and Analysis of Lesions
At the end of testing, rats were perfused intracardially withsaline followed by 10% formalin under terminal pentobarbi-tone anesthesia (Euthatal; 1 mL). After formalin perfusion,rats’ brains were removed and stored in 10% formalin for 24 hbefore being blocked and embedded in egg yolk: the brain wasplaced in a plastic cube filled with fresh egg yolk, incubated at5�C in a tray of formalin for 24 h, then left in formalin for afurther 24 h at room temperature. The block of hardened eggyolk containing the brain was then removed and left in forma-lin for an additional 24-h period. Coronal 30-mm sectionsthrough the hippocampus and other structures were cut usinga cryostat: 1-in-5 sections was mounted on a slide and stainedwith cresyl violet. The volumes of spared hippocampal andsubicular tissue in each brain were calculated using NIH Image1.63 (NIH, USA; de Hoz et al., 2003). Partial lesions wererequired to leave an intact tissue volume of 25–50% of thehippocampus, starting from the non-lesioned pole.
The relative volume of spared hippocampal tissue was cal-culated by measuring the area of the combined dentate gyrusand CA fields spared in each section of a particular brainaccording to the following protocol: Each coronal sectioncontaining hippocampal tissue was placed under a photoma-croscope (Wild, Heerbrugg, Switzerland), and the imagetaken by a mounted video camera was imported into NIHImage 1.63 (NIH, USA). The area of spared hippocampaltissue in each section was then outlined and calculated auto-matically. Surrounding fibers such as the fimbria wereexcluded on the grounds that they would not be consideredin a section were all the hippocampal cells dead. The sectionswere spaced 150 mm apart, yielding up to 32 sections in asham-lesioned animal, and fewer in animals with acceptablepartial lesions. For each rat, the total hippocampal “volume”was calculated by adding the area of hippocampal tissuespared in each successive section. The proportion of hippo-campus spared at the septal and temporal poles of the struc-ture in each animal was expressed as a percentage of themean sham hippocampal “volume.” Values for the left andright hippocampi were initially calculated separately and thenaveraged. Sparing of subicular tissue was calculated in exactlythe same way. Strict criteria for acceptance of a lesion wereused. The lesion had to be confined to the hippocampus inall cases, and leave intact tissue volumes of 25–50% startingfrom the non-lesioned pole, with minimal sparing (<10%unilaterally) elsewhere in the structure, and little or no dam-age to the neocortex.
Data Analysis
Several measures of performance were assessed, includingescape latency, search density, swim speed, and time spent ineach of the four quadrants of the pool during probe trials.Unless otherwise stated, statistical analysis comprised an analy-sis of variance (ANOVA) followed by appropriate post hoc com-parisons (Fisher’s protected LSD). Numerical data are reportedas mean 6 standard error (S.E.M.) throughout.
RESULTS
For brevity, “hippocampus” and “hippocampal” are hereafterabbreviated to “HPC” when referring to specific lesion groups.The final group sizes after histological assessment were as fol-lows: in the pre-training lesion groups there were 8 sham, 9septal, and 7 temporal-HPC-lesioned rats; in the post-traininglesion groups there were 22 sham, 15 septal, and 25 temporal-HPC-lesioned rats. See Supporting Information Results andFig. S1 for a quantitative description of the histological assess-ment of the lesions.
Initial “Upstairs” Learning
When surgery was carried out before the start of watermazetesting, differences between the lesion groups were already evi-dent during the pre-training phase of task acquisition, settingthe flavor for the rest of the study, throughout which we founda marked effect of search strategy on spatial performance. Onearly trials, rats tended to swim around the pool walls(“thigmotactic” swimming), before exploring the interior of thepool and locating the platform; this tendency was most pro-nounced in rats with septal HPC lesions made before training.Figure 2A (upper panel) shows mean escape latencies over thetwo blocks of five pre-training trials. There was a highly signifi-cant overall improvement in latency across trials[F(9,189) 5 35.1; P< 0.0005], and a main effect of group[F(2,21) 5 7.72; P< 0.005], with the septal-HPC-lesionedgroup taking significantly longer to reach the platform than thesham [P< 0.005] and temporal-HPC-lesioned [P< 0.01]groups. This latency difference was paralleled by a higher levelof thigmotactic swimming in septal-HPC-lesioned rats, com-pared to sham and temporal-HPC-lesioned animals [P< 0.02in both cases; main effect of group: F(9,189) 5 47.6;P< 0.0005] (Fig. 2A; lower panel).
During spatial training, with the exception of the septal-HPC-lesioned group, rats trained after surgery rapidly learnedto locate and raise the hidden platform, as indicated by theoverall fall in escape latencies across trials [F(31,819) 5 5.23;P< 0.0005] (Fig. 2B). As training progressed and dwell timesincreased, rats developed a characteristic strategy of focusedand persistent searching above the submerged platform. How-ever, the overall effect of lesion type was significant[F(2,21) 5 28.9; P< 0.0005], with septal-HPC-lesioned rats
994 DE HOZ AND MARTIN
Hippocampus

exhibiting markedly longer latencies than those with sham[P< 0.0005] or temporal HPC lesions [P< 0.0005]; the lattergroups did not differ [P> 0.2]. Temporal-HPC-lesioned ratsshowed slightly more forgetting than shams between blocks oftrials, both within and between days (corresponding to inter-block intervals of approximately 3 h and 24 h, respectively).For this reason, an analysis of escape latencies on the first trialof each block again revealed a significant overall effect of group[F(2,21) 5 18.3; P< 0.0005], but this time shams performedsignificantly better than both temporal and septal-HPC-lesioned groups [P< 0.005 in both cases], although temporal-HPC-lesioned rats performed significantly better than septal-HPC-lesioned animals [P< 0.05]. Importantly, no significantgroup differences in thigmotaxis [F(2,21)5 2.95; P> 0.05] orswim speed [F(2,21) 5 2.26; P> 0.1] were observed duringtraining. During the final block of five trials, all groups spentan average of <10% of their time within 15 cm of the poolwalls (sham 5 9.16 6 0.95%; temporal HPC lesion 5 7.33 6
1.54; septal HPC lesion 5 9.81 6 2.12%; F(2,21) 5 0.59;P> 0.5).
The performance of rats scheduled to receive post-trainingsurgery was similar to that of the future sham-lesioned groups.No differences between the prospective groups were observedduring pretraining [F(2,59) 5 2.63; P> 0.05] (Fig. 2C) ortraining [F< 1] (Fig. 2D).
“Upstairs” Memory Retention Testing
Two weeks after acquisition training all rats received a mem-ory probe in the upstairs watermaze. The % time spent in allquadrants of the pool during this 60-s probe trial is shown inFigure 3. Rats with pre-training sham and temporal HPClesions (Fig. 3A) exhibited very good retention of the platformlocation reflected in a strong bias for the training quadrant.Little memory was evident in the septal-HPC-lesioned group.An ANOVA revealed a significant interaction of lesion typeand quadrant [F(4,42) 5 5.67; P 5 0.001]. An ANOVA of per-
centage time in the training quadrant only revealed a maineffect of lesion type [F(2,21) 5 13.2; P< 0.0005], with theseptal-HPC-lesioned group performing significantly worse thanboth shams [P< 0.0005] and temporal-HPC-lesioned rats[P< 0.002]; the latter two groups did not differ. Whereas bothsham and temporal-HPC-lesioned rats spent significantly moretime in the target quadrant than predicted by chance, this wasnot true of septal-HPC-lesioned rats [sham: t(7) 5 5.65;P 5 0.001; temporal HPC lesion: t(6) 5 16.5; P< 0.0002; sep-tal HPC lesion: t(8) 5 0.88; P> 0.4]. However, the lattergroup exhibited significantly below-chance search performancein the opposite quadrant [t(8) 5 4.79; P 5 0.001], indicating aweak bias toward the general area of the pool in which theplatform had formerly been located.
In rats with post-training lesions, good retention was evidentonly in the sham-lesioned group (Fig. 3B), with the partiallesion groups showing a weak bias for the training location. AnANOVA revealed a significant interaction of lesion group andquadrant [F(2,118) 5 5.43; P< 0.0005], and an ANOVA oftime in the training quadrant only revealed a significant effectof lesion type [F(2,59) 5 11.5; P< 0.0005], with sham-lesionedrats performing better than both septal [P< 0.002] andtemporal-HPC-lesioned [P< 0.0005] groups. Only shams spentsignificantly more time in the training quadrant than predictedby chance [sham: t(21) 5 5.37; P< 0.0005; septal HPC lesion:t(14) 5 0.91; P> 0.3; temporal HPC lesion: t(24) 5 0.92;P> 0.3], although both septal and temporal-HPC-lesionedgroups searched at below-chance levels in the opposite quadrantof the pool, indicating a weak bias toward the correct generalarea of the pool [septal HPC lesion: t(14) 5 3.30; P 5 0.005;temporal HPC lesion: t(24) 5 2.97; P< 0.01]. These results arenot the result of small differences in hippocampal or subicularsparing across groups (see Supplementary Results).
Reminding and Reversal Learning
At the end of the first probe trial, consisting of a 60-s freeswim, the platform was raised in either the original location
FIGURE 3. Retention testing—PT1 only (“upstairs watermaze”). Percentage time spent ineach quadrant of the pool during the first upstairs probe trial (PT1; Tr 5 training;Opp 5 opposite; AL 5 adjacent left; AR 5 adjacent right) in rats with pre-training surgery (A),and post-training surgery (B). [Color figure can be viewed in the online issue, which is avail-able at wileyonlinelibrary.com.]
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 995
Hippocampus
(for half the animals) or a novel location, to test remindingand reversal/novel learning, respectively (see de Hoz et al.,2004). This procedure, a 60-s free-swim followed by 30 s onthe platform, was repeated three times. As a result, the first 60s of PT1 are identical for all animals, but subsequent PTs mustbe treated differently for rats that were reminded of the origi-nal training location and for those that were “reminded” of anovel location.
Figure 4 shows time spent in the target quadrant (whetheroriginal or novel) across all three upstairs probes. As analysis isalways centered on the current location of the platform, a dif-ference between novel and original groups at any stage duringretention testing must reflect memory for the original traininglocation. In the following analysis, we focus first on the overalldifference between novel and original sub-groups, as an indexof memory retention, followed by an analysis of the improve-ment in performance in the novel sub-groups only—a measureof new learning or reversal learning, in contrast to the re-learning or reminding that occurs when rats are re-exposed tothe original location.
When analysed in this way, pre-training sham-lesioned ratsmaintained very good memory for the training location acrossPTs, as well as rapid reversal learning in the novel sub-group,evident in the fact that by PT3 this group had developed abias toward the novel quadrant (Fig. 4A). Septal-HPC-lesionedrats exhibited a slight overall difference between novel andoriginal sub-groups, consistent with a limited sparing of mem-ory for the original location. However, little new-learning abil-ity was evident in the novel sub-group (Fig. 4B). Rats withpre-training temporal-HPC lesions, like shams, maintainedvery good memory for the training location, but little newlearning occurred in the novel sub-group (Fig. 4C). AnANOVA of performance on PT1-3 in which pre-training lesiongroup and novel versus original sub-group were entered asbetween-subjects factors, and probe trial was entered as awithin-subjects factor revealed a highly significant interactionof lesion type with novel versus original sub-group[F(2,18) 5 6.08; P 5 0.01]. Follow-up ANOVAs of perform-ance in each lesion group revealed significantly better perform-ance in original compared to novel sub-groups (and hence
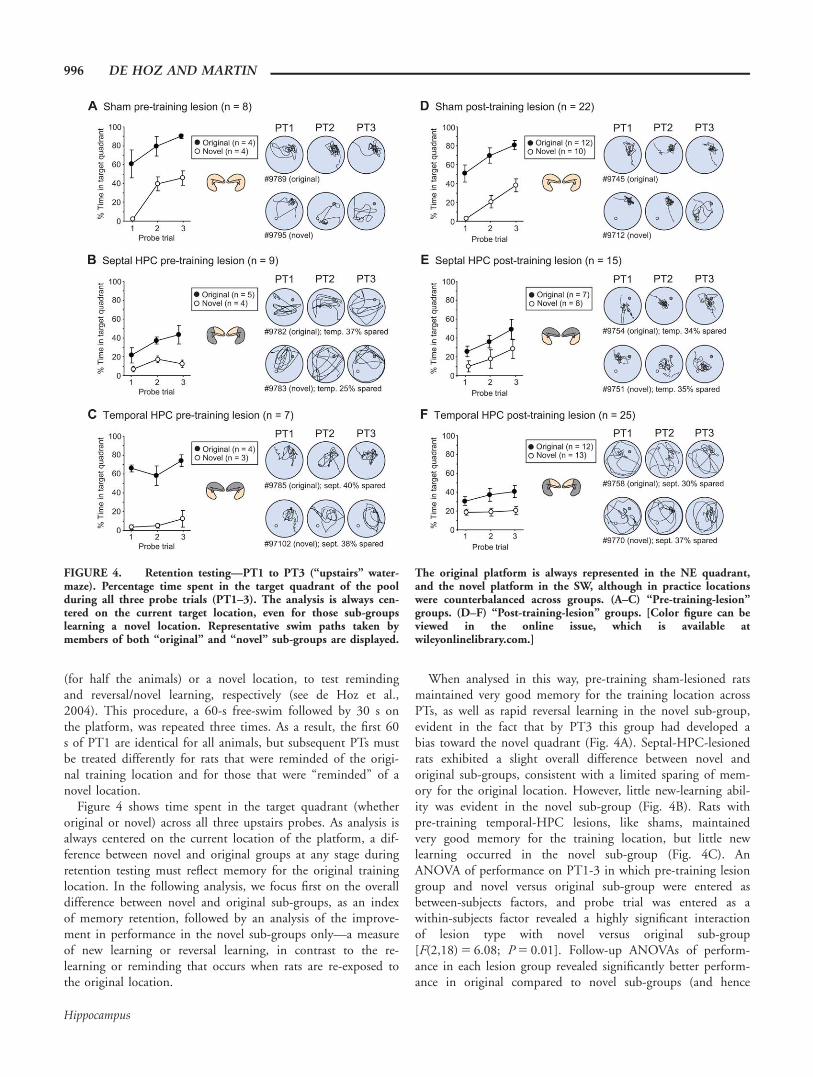
FIGURE 4. Retention testing—PT1 to PT3 (“upstairs” water-maze). Percentage time spent in the target quadrant of the poolduring all three probe trials (PT1–3). The analysis is always cen-tered on the current target location, even for those sub-groupslearning a novel location. Representative swim paths taken bymembers of both “original” and “novel” sub-groups are displayed.
The original platform is always represented in the NE quadrant,and the novel platform in the SW, although in practice locationswere counterbalanced across groups. (A–C) “Pre-training-lesion”groups. (D–F) “Post-training-lesion” groups. [Color figure can beviewed in the online issue, which is available atwileyonlinelibrary.com.]
996 DE HOZ AND MARTIN
Hippocampus

significant memory) in all cases [sham: F(1,6) 5 22.6;P< 0.005; septal HPC lesion: F(1,7) 5 10.9; P< 0.02; tempo-ral HPC lesion: F(1,5) 5 86.9; P< 0.0005]. However, analysisof the original sub-groups only revealed a significant overalleffect of lesion type [F(2,10) 5 12.4; P< 0.002], with septal-HPC-lesioned rats performing more poorly than shams andtemporal-HPC-lesioned animals [P< 0.005 in both cases]. AnANOVA of the improvement between PT1–3 in novel sub-groups revealed significantly greater new learning in shamscompared to septal and temporal-HPC-lesioned rats [P< 0.005in both cases; sham improvement 5 44.0 6 7.1 percentagepoints; septal-HPC-lesion improvement 5 5.3 6 4.9 percentagepoints; temporal-HPC-lesion improvement 5 9.2 6 6.3 per-centage points; main effect of lesion type: F(2,8) 5 12.5;P< 0.005].
Owing to the small sample sizes involved in the analysis ofnovel and original sub-groups with pre-training surgery, weconducted a parallel non-parametric analysis of time in the tar-get quadrant (as a percentage of total time in the tar-get 1 opposite quadrants) on “upstairs” probe trials 1–3; SeeSupplementary Information.
As expected, rats given post-training sham surgery also main-tained good memory for the training location across PTs, withfurther improvements in search accuracy across probes. Thisgroup also showed rapid reversal learning in the novel sub-group (Fig. 4D). Septal-HPC-lesioned animals showed a mod-est sparing of memory, indicated by a difference between origi-nal and novel sub-groups, as well as an overall increase in timespent in the target quadrant across probes (Fig. 4E). Rats withtemporal-HPC lesions made after training also showed a mod-est sparing of memory indicated by a difference between noveland original sub-groups, but there was little overall improve-ment on time spent in the target quadrant across probes (Fig.4F; see Supporting Information Fig. S2 for further analysis).An overall ANOVA of performance on PT1–3 in which post-training lesion group and novel versus original sub-group wereentered as between-subjects factors, and probe trial was enteredas a within-subjects factor revealed a highly significant interac-tion of lesion type with novel versus original sub-group[F(2,56) 5 5.03; P 5 0.01]. Follow-up ANOVAs of perform-ance in each lesion group revealed significantly better perform-ance in original compared to novel sub-groups (and hencememory) in sham [F(1,20) 5 37.2; P< 0.0005] and temporal-HPC lesioned groups [F(1,23) 5 7.51; P< 0.02], and a trendtoward a difference in septal-HPC-lesioned rats [F(1,13) 5
3.18; P< 0.1]. An analysis of performance in original sub-groups only revealed a significant main effect of lesion type[F(2,28) 5 9.56; P 5 0.001], with significant differencesbetween sham and both septal [P 5 0.002] and temporal-HPC-lesioned groups [P< 0.0005]. The latter two partial-lesion groups did not differ [P> 0.9]. Considering both noveland original sub-groups together, both shams and septal-HPC-lesioned rats showed an overall improvement in the time spentin the target quadrant across probe trials [sham: F(2,40) 5
19.1; P< 0.0005; septal HPC lesion: F(2,26) 5 7.83;P 5 0.002]. No overall improvement was seen in the temporal-
HPC-lesioned group [F(2,46) 5 2.14; P> 0.1; but see Sup-porting Information Fig. S2]. An ANOVA of the improvementbetween PT1–3 in novel sub-groups only [shamimprovement 5 35.3 6 6.6 percentage points; septal-HPC-lesion improvement 5 18.4 6 8.4 percentage points; temporal-HPC-lesion improvement 5 1.9 6 3.2 percentage points; maineffect of lesion type: F(2,28) 5 9.36; P 5 0.001] revealed sig-nificantly poorer new learning in temporal-HPC-lesioned ratscompared to shams [P< 0.0005]. There was also a trendtoward a difference between temporal and septal-HPC-lesionedgroups [P< 0.1], and between sham and septal-HPC-lesionedgroups [P< 0.1].
In de Hoz et al. (2004), we found that a single presentationof the platform at the end of PT1, acted as a reminder of theoriginal platform location in the group with temporal hippo-campal lesions; this reminder was independent of whether theplatform was raised in the original platform location or a in anovel location. Here we replicated this pattern of results, asillustrated in detail in Fig. S2 and described in the SupportingInformation Results.
New Learning (“Downstairs” Watermaze)
Animals from a given lesion group were pooled together forthe downstairs training, independently of whether they hadundergone reminding or reversal learning “upstairs,” since therewas no significant difference in performance between novel andoriginal sub-groups [pre-training surgery: F(1,18) 5 0.22;P> 0.6; post-training surgery: F(1,56) 5 0.03; P> 0.8].
Despite the challenging training protocol consisting of suc-cessive rewarded probe trials separated by 1-h intervals, sham-lesioned rats quickly learned to search persistently in the train-ing location until the platform became available after 60 s; per-formance typically reached an asymptote by PT3, with littlefurther improvement over the final two probes. Althoughpoorer than shams, rats given pre-training temporal HPClesions showed a steady increase in the bias toward the trainingquadrant (Fig. 5A). Little learning was evident in septal-HPC-lesioned animals. An ANOVA of performance in each lesiongroup over the five probe trials revealed a significant overallimprovement across probe trials [F(4,84) 5 23.0; P< 0.0005],a significant effect of lesion type [F(2,21) 5 18.5; P< 0.005],and a significant interaction between lesion and probe trial[F(8,84) 5 3.99; P< 0.0005]. Analysis of mean performanceover the final three probe trials (Fig. 5B) revealed a similar pat-tern of results, with a main effect of lesion type[F(2,21) 5 23.1; P< 0.0005], and superior performance intemporal-HPC-lesioned with respect to septal-HPC-lesionedrats [P< 0.005]. Shams performed significantly better thanboth lesion groups [P< 0.005 and P< 0.0005, respectively].Representative swim paths are shown in Figure 5C.
In groups with post-training surgery, septal-HPC-lesionedrats exhibited facilitated new learning relative to temporal-HPC-lesioned animals (Fig. 5D). An ANOVA of performancein each lesion group over the five probe trials revealed a signifi-cant overall improvement across trials [F(4,236) 5 42.7;
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 997
Hippocampus
P< 0.0005], a significant effect of lesion type [F(2,59) 5 15.7;P< 0.0005], and a significant lesion group x probe trial inter-action [F(8,236) 5 28.4; P< 0.0005]. An analysis of mean per-
formance over the final three probe trials (Fig. 5E) likewiserevealed a main effect of lesion type [F(2,59) 5 24.3;P< 0.0005], with poorer memory in temporal-than septal-HPC-lesioned rats [P 5 0.01], and both groups significantlybelow shams [P< 0.0005 and P< 0.002, respectively]. Repre-sentative swim paths are shown in Figure 5F.
Although the sample sizes used in the post-training surgerygroups were higher than those used in the pre-training surgerygroups, the difference between septal- and temporal-HPC-lesioned groups in the “downstairs” pool was large in bothcases, though opposite in direction. For example, the effect size(Cohen’s d) for the superiority of septal- versus temporal-HPC-lesioned rats over the final three probe trials was 0.94; basedon the observed means and standard deviations, group sizes ofn 5 10 would have been sufficient to yield a significant groupdifference during this phase of testing.
Search Strategy
What follows is an analysis of the degree of focused search-ing within a small area of the pool in different groups on PT1of retention testing “upstairs” and PT1 of new learning“downstairs.” Swim speed was negatively correlated with searchdensity such that speed was high when an animal searched dif-fusely, and low when an animal searched densely in one loca-tion (Supporting Information Fig. S3).
Focused searching during PT1 upstairs (retention test) inrats with pre-training lesions (Fig. 6A) was similar acrossgroups. An ANOVA did not reveal a significant effect of lesiontype [F(2,21) 5 2.52; P> 0.1], although septal-HPC-lesionedrats exhibited slightly lower scores than sham and temporal-HPC-lesioned groups (see example swim paths in Figs. 4A–C).In rats with post-training surgery (Fig. 6B), the effect of lesiontype was significant [F(2,59) 5 40.7; P< 0.0005], but therewas no difference between sham and septal-HPC-lesionedgroups [P> 0.1], despite the large difference in memorybetween these groups; scores in both groups were higher thanthat of temporal-HPC-lesioned rats [P< 0.0005 in both cases].These differences in search strategy were maintained across thesubsequent two probe trials (see paths in Figs. 4D–F).
In PT1 downstairs (new learning), all groups with pre-training lesions exhibited a diffuse search pattern ranging overlarge areas of the pool; no group differences in focused
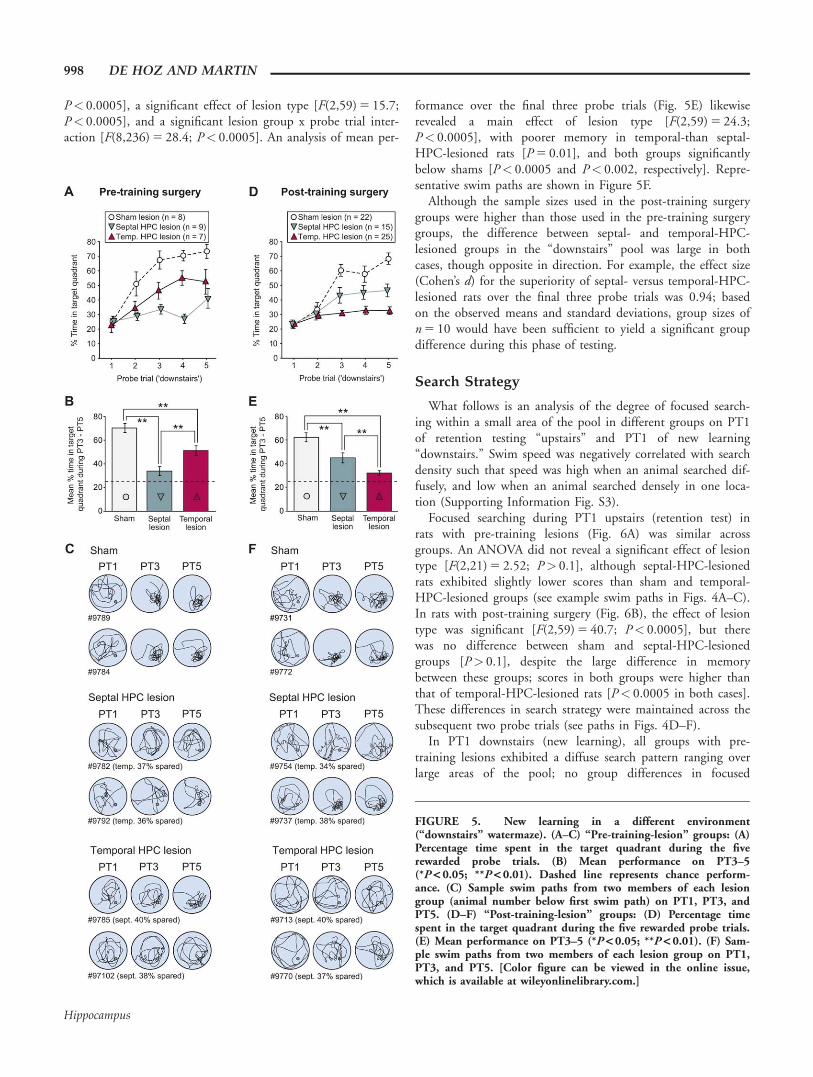
FIGURE 5. New learning in a different environment(“downstairs” watermaze). (A–C) “Pre-training-lesion” groups: (A)Percentage time spent in the target quadrant during the fiverewarded probe trials. (B) Mean performance on PT3–5(*P < 0.05; **P < 0.01). Dashed line represents chance perform-ance. (C) Sample swim paths from two members of each lesiongroup (animal number below first swim path) on PT1, PT3, andPT5. (D–F) “Post-training-lesion” groups: (D) Percentage timespent in the target quadrant during the five rewarded probe trials.(E) Mean performance on PT3–5 (*P < 0.05; **P < 0.01). (F) Sam-ple swim paths from two members of each lesion group on PT1,PT3, and PT5. [Color figure can be viewed in the online issue,which is available at wileyonlinelibrary.com.]
998 DE HOZ AND MARTIN
Hippocampus
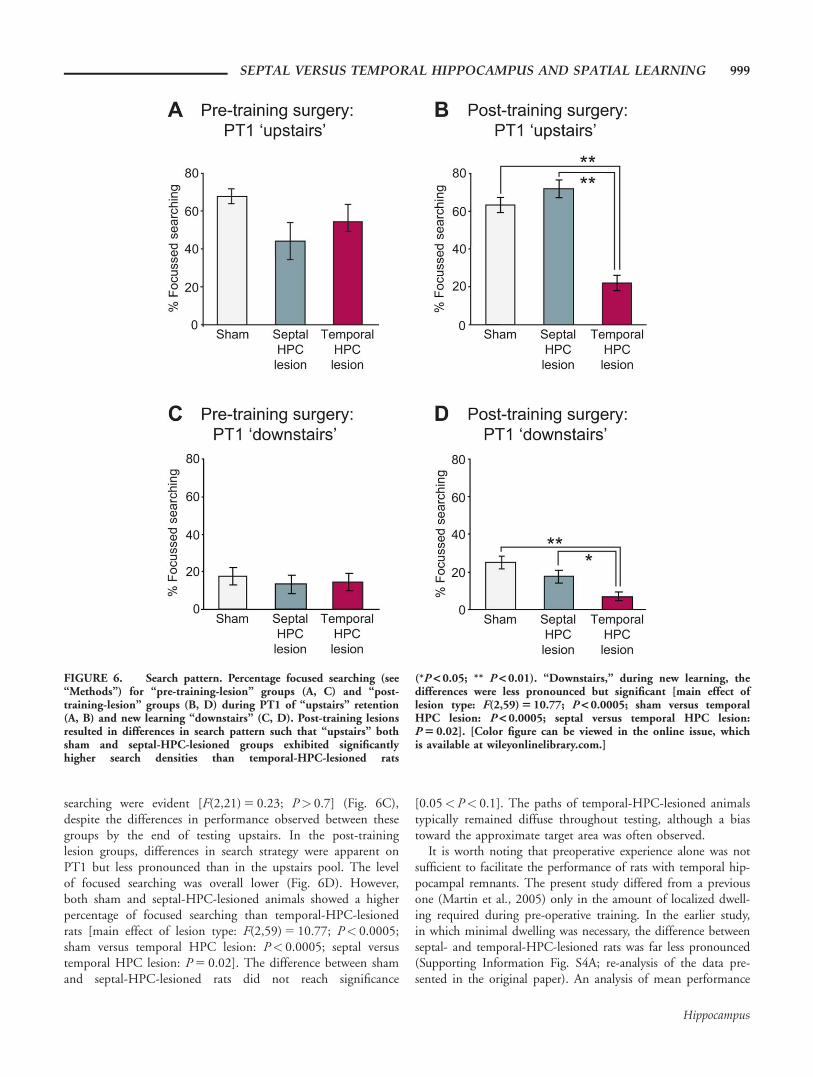
searching were evident [F(2,21) 5 0.23; P> 0.7] (Fig. 6C),despite the differences in performance observed between thesegroups by the end of testing upstairs. In the post-traininglesion groups, differences in search strategy were apparent onPT1 but less pronounced than in the upstairs pool. The levelof focused searching was overall lower (Fig. 6D). However,both sham and septal-HPC-lesioned animals showed a higherpercentage of focused searching than temporal-HPC-lesionedrats [main effect of lesion type: F(2,59) 5 10.77; P< 0.0005;sham versus temporal HPC lesion: P< 0.0005; septal versustemporal HPC lesion: P 5 0.02]. The difference between shamand septal-HPC-lesioned rats did not reach significance
[0.05<P< 0.1]. The paths of temporal-HPC-lesioned animalstypically remained diffuse throughout testing, although a biastoward the approximate target area was often observed.
It is worth noting that preoperative experience alone was notsufficient to facilitate the performance of rats with temporal hip-pocampal remnants. The present study differed from a previousone (Martin et al., 2005) only in the amount of localized dwell-ing required during pre-operative training. In the earlier study,in which minimal dwelling was necessary, the difference betweenseptal- and temporal-HPC-lesioned rats was far less pronounced(Supporting Information Fig. S4A; re-analysis of the data pre-sented in the original paper). An analysis of mean performance
FIGURE 6. Search pattern. Percentage focused searching (see“Methods”) for “pre-training-lesion” groups (A, C) and “post-training-lesion” groups (B, D) during PT1 of “upstairs” retention(A, B) and new learning “downstairs” (C, D). Post-training lesionsresulted in differences in search pattern such that “upstairs” bothsham and septal-HPC-lesioned groups exhibited significantlyhigher search densities than temporal-HPC-lesioned rats
(*P < 0.05; ** P < 0.01). “Downstairs,” during new learning, thedifferences were less pronounced but significant [main effect oflesion type: F(2,59) 5 10.77; P < 0.0005; sham versus temporalHPC lesion: P < 0.0005; septal versus temporal HPC lesion:P 5 0.02]. [Color figure can be viewed in the online issue, whichis available at wileyonlinelibrary.com.]
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 999
Hippocampus

over the final three probe trials revealed a main effect of lesiontype [F(2,63) 5 69.5; P< 0.0005], but the trend toward betterperformance in septal versus temporal-HPC-lesioned rats didnot reach significance [0.05<P< 0.1; Fig. S4B].
In Martin et al. (2005), despite the fact that lesions wereperformed after training, focused searching was very limitedand showed similar levels in the different lesion groups, prob-ably because focused searching was not required during train-ing itself (Supporting Information Fig. S4C, left-hand panel;see Fig. S4C, right-hand panel, and D-F for swim-speed dataand illustrative swim paths, respectively). Only when persistentdwelling was required during training in the present studywere rats with temporal HPC lesions, able to learn a platformlocation post-operatively.
DISCUSSION
The key result of these experiments is that the presence orlack of experience prior to a partial lesion of the hippocampus
determines the capacity of animals with septal or temporal hip-pocampal lesions to learn the position of a platform in a novelenvironment, revealing a double dissociation of functionbetween these two poles of the hippocampus, summarized inFigure 7. When partial hippocampal lesions were made beforeany experience in the watermaze, rats with temporal hippocam-pal lesions (i.e., septal sparing), but not rats with septal hippo-campal lesions, could learn the position of a platform, both inthe original pool, and in a new environment. When the lesionswere made after initial training, however, the reverse patternwas true—only rats with lesions of the septal hippocampus(i.e., temporal sparing) could learn the platform position in thenew environment.
This result was dependent on “procedural” aspects of thetraining protocol—such as the pre-operative acquisition of afocused search strategy, and was independent of the rats’ famili-arity with the specific environment in which new learning wasassessed. Conversely, when lesions were made before task acqui-sition, and the animals had no prior experience in the water-maze, we observed superior learning abilities in rats with intactseptal hippocampi, albeit coupled with an impairment in rever-sal learning in the same spatial context. The significance ofthese findings is twofold: first, the former result appears tochallenge the widely accepted idea that the septal hippocampusis the primary substrate for spatial memory, and raises impor-tant issues regarding functional differentiation along the longi-tudinal axis of the structure. Second, the phenomenon issensitive to “procedural” elements of task performance, andmight be explained by the connectivity of the temporal hippo-campus with output structures necessary for the expression ofhippocampus-dependent information—cf. Mogenson and Yang(1991). Our main findings, obtained through measures oflearning, memory, reminding, and subsequent new learning ina novel environment, can be summarized as follows:
First, when lesions were made before training, septal-HPClesions severely impaired the learning of a watermaze spatial-reference-memory task, whereas temporal HPC lesions hadminimal effects. Consequently, rats with septal HPC lesionsexhibited limited memory in a probe trial conducted 15–16days after training, whereas temporal-HPC-lesioned rats per-formed as well as controls. These findings are consistent withprevious evidence that the septal (dorsal) hippocampus is criti-cal for spatial learning and memory under these circumstances(see Moser and Moser, 1998a; Fanselow and Dong, 2010, forreview), and may reflect the poverty of visuospatial informationreaching the hippocampus after lesions of this type. Althoughin the present study, rats with septal HPC lesions exhibitedincreased thigmotactic swimming, and consequently increasedescape latency during cued pretraining (perhaps a consequenceof using an “Atlantis” platform whose location is indicated bya hanging cue, but is not directly visible or available; as well asa relatively long “dwell” time of 1 s) the difference in thigmo-taxis had disappeared by the end of training, and cannotexplain the subsequent performance of this group.
Second, after the first retention probe, rats received eitherfurther training with the platform in its original location, or
FIGURE 7. Overview. Index of new learning ability in the sec-ond (“downstairs”) pool: mean percentage time spent in the train-ing quadrant during the last three probe trials expressed as apercentage of sham performance. An ANOVA revealed a significantinteraction between hippocampal lesion type (septal or temporal)and the time of lesioning (pre- or post-training) [F(1,52) 5 15.2;P < 0.0005], confirming the existence of a double dissociationbetween the performance of animals with temporal and septal hip-pocampal lesions depending on the time (i.e., before or after ini-tial training) at which the lesions are made.
1000 DE HOZ AND MARTIN
Hippocampus

were faced with a platform in a novel location in the samepool: although both sham and temporal-HPC-lesioned ratsremembered the original location, only shams exhibited suc-cessful reversal learning when presented with a novel platformposition in the same pool, quickly learning to search in thenew location. This result highlights the lack of behavioral flexi-bility in rats with temporal HPC lesions—while learning andmemory at first appeared to be near-normal, performance wasseverely disrupted when task demands changed. Third, consist-ent with their performance during training, rats with temporal-HPC lesions performed well in a test of new learning in anunfamiliar environment, and significantly better than rats withseptal-HPC lesions. Their search was not as focused of that ofshams but it was clearly biased toward the training quadrant,indicating that, under these circumstances, rats with temporalHPC lesions retain a capacity to associate a spatial locationwith reward.
When the lesions were made after the acquisition phase, allgroups performed equivalently during pre-operative training,but rats with either septal or temporal lesions were severely,and equally, impaired in the initial retention test conductedafter surgery. This suggests that although the temporal hippo-campus is not necessary for the encoding and storage of mem-ory in this task, it does play a role under normal circumstancesin which learning occurs in an intact animal and memories areencoded in a distributed fashion throughout the hippocampus,a result that is consistent with the findings of Moser and Moser(1998b), and more recently Loureiro et al. (2011). However,septal-HPC-lesioned rats did remember the focused and persis-tent search strategy acquired during training, although theywere unable to apply this strategy in the correct location; incontrast, temporal-HPC-lesioned rats did not retain this strat-egy, and searched widely throughout the pool. Second,although both septal and temporal lesions disrupted initialmemory retention, rats with septal HPC lesions showed betterreversal (reflected in their overall improvement across probe tri-als) relative to those with temporal HPC lesions. In fact, thelatter group was paradoxically reminded of the original locationwhen presented with a platform in a novel location, a resultthat echoes the lack of flexibility observed in rats withtemporal HPC lesions made before training, and replicates ourprevious observations (Supporting Information Fig. S2; de Hozet al., 2004). Third, finally, and most surprisingly, septal-HPC-lesioned rats were substantially better than those with temporalHPC lesions in subsequent new learning in a differentenvironment.
To assess new learning we used a protocol that paralleled thatused during reminding/reversal and consisted of successive probetrials. This protocol was designed for animals with prior knowl-edge of the watermaze “principle” and is a test of rapid newlearning. Its efficacy is demonstrated in that the sham groupdeveloped a strong spatial bias for the platform location, typicallyafter only one or two trials, even in a completely new environ-ment. Using this protocol, we replicate previous findings of supe-rior new learning in rats with temporal HPC lesions over thosewith septal HPC lesions, when the lesions are made before the
rats have had any watermaze experience (Moser et al. 1993 and1995). Conversely, in rats with temporal HPC lesions made afterinitial training, there is little improvement over the five probe tri-als in the second, downstairs, pool. Nonetheless, animals withseptal HPC lesions (and temporal sparing) were capable of rapidnew learning, spending an average of more than 40% of theirtime in the target quadrant after only two exposures to its loca-tion. Of course, although rapid new learning in a new environ-ment is dramatically impaired in the former group, performancemight have improved with continued training. For example, ratswith small temporal hippocampal lesions made after initial train-ing and sparing two thirds of the hippocampus have previouslybeen found to be unimpaired in subsequent new learningassessed using eight conventional (spaced) trials, followed by aprobe (Moser and Moser, 1998b). In general, when studying thecapacity for new learning of animals with considerable experiencein the watermaze, as is the case here, the initial stages of learningare probably more informative than the effect of extensive train-ing. Rats without a hippocampus can eventually learn a water-maze task by recruiting systems other than the hippocampuswhich can take over some aspects of performance (Morris et al.,1990).
This summary highlights major differences in the effects ofseptal and temporal HPC lesions on flexibility, search strategy,and the impact of prior experience on new-learning ability in adifferent environment. These differences are consistent with theconnectivity and functional interactions of the hippocampuswith other brain areas involved in watermaze performance; seeBast, 2011, for review. For example, the hippocampal projec-tion to the medial prefrontal cortex (mPFC) arises from thetemporal two-thirds of the hippocampus in rats (Swanson,1981; Jay and Witter, 1991)—the very portion that was absentin our temporal-HPC-lesioned group. Several rodent studies ofspatial memory reveal functional interactions between the ven-tral CA1/subicular region and the mPFC—e.g., Floresco et al.(1997); Seamans et al. (1998). Ventral hippocampal activitymodulates prefrontal dopaminergic transmission (Grace et al.,2007; Taepavarapruk et al., 2008), and unit-recording studiesreveal coordinated firing in both prefrontal and temporal-hippocampal neurons during behavior—e.g. Hyman et al.(2005); Jones and Wilson (2005). Moreover, the PFC becomesimportant for the retrieval of hippocampus-dependent memo-ries soon after learning (Takashima et al., 2009; Tse et al.,2011; Wang et al., 2012b), and a single episode of hippocam-pal LTP induction causes a marked facilitation in the transmis-sion of hippocampal activity to the mPFC (Canals et al.,2009). The prefrontal cortex is also necessary for rapid reversallearning in spatial-memory tasks (de Bruin et al., 1994; Lacroixet al., 2002). This might explain why rats with temporal HPClesions, who have not developed the use of an alternativeretrieval route during acquisition, find themselves with spatialinformation stored in an area of the hippocampus that is nolonger connected to the retrieval route; see Sparks et al.(2011). The limited reversal-learning abilities of rats withseptal HPC sparing are therefore consistent with the lack ofconnectivity of this part of the hippocampus with the PFC. In
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 1001
Hippocampus

addition, mPFC is believed to speed up the acquisition ofmemories when the information to be learned is consistentwith prior knowledge (Tse et al., 2007; van Kesteren et al.,2010; Tse et al., 2011; Wang et al., 2012b), which is the casein the downstairs pool.
The temporal hippocampus also exhibits the strongest ana-tomical and functional connectivity with the ventral striatum(Groenewegen et al., 1987; Mogenson and Yang, 1991; Blahaet al., 1997; Naber and Witter, 1998; Johnson et al., 2007;Goto and Grace, 2008; Ito et al., 2008). This pathway haslong been implicated in conveying spatial information tomotor effector sites (Mogenson and Yang, 1991; Groenewegenet al., 1996; Floresco et al., 1997), and unit-recording studiesreveal functional interactions between the two structures thatmay be critical in coupling spatial representations to informa-tion about reward (van der Meer and Redish, 2011a). Theseobservations may be relevant to the preservation of a densesearch strategy, albeit one that is dissociated from its targetlocation, in rats with temporal HPC sparing. Whereas pre-operatively-acquired spatial information seems to be degradedby septal HPC lesions, “procedural” learning mediated bytemporal HPC interactions with the striatum seems to remainunaffected by the lesion. Our data suggest that the retention ofa dense search strategy is dependent not only on the integrityof downstream structures as Micheau et al. (2004) suggest, butalso on their connectivity with the temporal hippocampus.Interactions between the HPC and mPFC may also be rele-vant. Unit-recording data reveal the existence of cells in bothstructures that respond in anticipation of the delayed deliveryof food in an unmarked goal location—essentially a dry-landanalog of our watermaze-training procedure (Hok et al., 2005,2007); this activity is abolished by temporal HPC lesions, afterwhich rats become less persistent in waiting for reward in theappropriate location (Burton et al., 2009).
These anatomical considerations offer a plausible explanationfor the superior performance of rats with post-training septal-HPC lesions on new learning in a different environment. Intask-experienced septal-HPC-lesioned rats, the encoding of asingle goal location may not require very extensive hippocam-pal resources. Analogous findings have recently been presentedby Wang et al. (2012a) in a contextual fear-conditioning task:septal hippocampal inactivation impaired the initial acquisitionof contextual fear, but subsequent learning in a second contextwas unaffected, suggesting that prior experience renders thetemporal hippocampus sufficient for “second learning.”
In our study, the limited visuospatial sensory input to thetemporal hippocampus might be sufficient for rapid spatiallearning owing to the ability of the mPFC and ventral striatumto link available information with appropriate goal-directedaction and the formation of new reward-location associations(van Kesteren et al., 2010). Without this ability, animals withtemporal HPC lesions (septal sparing) might be able to processthe rich visuospatial input that the septal region receives inorder to form a post-operative spatial memory, or to reactivatea cortico-hippocampal memory trace formed pre-operatively,despite impairments in behavioral flexibility and the capacity
for reversal learning resulting from the loss of connections withthe mPFC. This argument also implies that the isolated septalhippocampus can bypass the absent outputs to executive struc-tures usually involved in memory retrieval, when all aspects ofthe task are learned postoperatively. However, early in thecourse of spatial learning the demands placed upon sensoryinformation are likely to be paramount. For this reason, theisolated temporal hippocampus may simply be incapable ofsupporting accurate spatial learning under the more frequently-studied conditions in which lesions are made before the startof training. This view is consistent with a detailed behavioralanalysis of watermaze learning in mice with septal-HPC lesions(Ruediger et al., 2012); despite progressing successfully throughmany of the search-strategy phases identified in the study, theseanimals were unable to acquire a final phase of accurate,focused searching in the correct location (cf. Fig. 5C in thepresent study, swim paths). It is also consistent with the find-ings of Nadel et al. (2013) according to which the human pos-terior (septal) hippocampus is important in detailed spatialinformation, while the anterior (temporal) hippocampus isimportant in context differentiation. In the training-before-surgery condition, the fact that we allowed our septal HPClesioned rats to acquire this spatial strategy before lesioningmay explain their subsequent success. In short, the presentfindings and those of de Hoz et al. (2003) do not contradictprevious reports of septal HPC “superiority”; we have simplyuncovered situations in which the contribution of the temporalhippocampus is revealed.
Animals in this study were trained with the Atlantis platformusing a relatively high dwelling time (see Materials and Meth-ods). It is worth discussing when and how learning this densesearch strategy affected spatial memory. Persistent, focusedswimming is not always the result of training with high dwell-ing times. For example, when animals remember the positionof the platform, as is the case with sham-lesioned rats, andthose with temporal HPC lesions made before training, thesearch density during a probe trial is high, independently ofwhether training involves a high dwelling time (the presentstudy) or a short dwelling time (de Hoz et al., 2003; Martinet al., 2005). Conversely, the degree of focused searching dur-ing an initial probe trial is not a good predictor of subsequentnew learning. For example, while rats with sham and septalHPC lesions made before “upstairs” training have similarsearch densities during new learning in the “downstairs” pool(Fig. 6C), only shams show rapid new learning (Fig. 5A). Also,in a short-dwell-time study (Martin et al., 2005; Fig. S4), littlefocused searching was evident in any group, yet shams per-formed at levels comparable to those in the present study. Inbrief, when performance is close to ceiling-levels, as in sham-operated rats, the use of long dwell-times does not improveperformance. However, when some capacity for new learningremains, and the circumstances of learning are challenging (asingle exposure followed an hour later by a 60-s probe trial),having learned to be persistent—and to perform a focusedsearch in order to obtain the platform—improves post-operative performance. This lifts the performance of partially
1002 DE HOZ AND MARTIN
Hippocampus

lesioned rats away from chance levels, and reveals the superiorabilities of animals with temporal hippocampal sparing underthese circumstances (compare Figs. 5D; S4A). The ability tocarry out a focused search might endow the rats with a way ofdisplaying that they “remember” that the platform can befound in a given position. The distinction between“remembering” (recollection) and “knowing” (familiarity) hasbecome instrumental in redefining the role of the human hip-pocampus in memory (Tulving, 1985). The confounding effectof the strength of memory, usually stronger in the“remembering” condition, has been addressed more recently(Smith et al., 2011; Yu et al., 2012). Here, the tendency fordense and focused searching can be dissociated from thestrength of the memory.
Although we have focused exclusively on dissociationsbetween the septal and temporal hippocampus, anatomical andgenomic data support a division into 3 longitudinal zones,with an “intermediate” region lying between septal and tempo-ral regions; see Fanselow and Dong (2010) for review. Recentevidence suggests that rats with selective sparing of the interme-diate hippocampus perform better than those with either septalor temporal sparing in a test of one-trial place memory, per-haps because, as the authors suggest, this region—like the sep-tal hippocampus—receives strong visuospatial input, but also—like the temporal pole—sends outputs to structures involved inbehavioral control (Bast et al., 2009). In view of this finding, itwould be interesting to test animals with lesions sparing theintermediate hippocampus using the procedures detailed in thepresent study.
In summary, a “unitary-function” hypothesis is often con-trasted with the idea that the hippocampus contains distinct,albeit interconnected, functional units, possibly subserving dif-ferent forms of memory. But the notion that the hippocampusis functionally differentiated along its septotemporal axis doesnot imply that different areas of the structure always mediatedistinct forms of memory. In humans, for instance, a numberof imaging studies suggest that each pole of the hippocampusis activated by the same type of mnemonic information but atdifferent stages of processing—e.g., Lepage et al. (1998). Ourresults are likewise consistent with the view that septal andtemporal tissue can play interdependent roles in a single formof hippocampal memory. Moreover, in contrast to the notionthat the involvement of the temporal hippocampus in spatialmemory is somewhat secondary, our data suggest that inappropriate circumstances this role can be more importantthan that of the septal hippocampus.
Acknowledgments
We are very grateful to Ann Paterson for assistance withbehavioral testing and data analysis, Jane Tulloch and KateShires for assistance with histology and histological analysis,and Richard Morris, Tobias Bast and other members of theCCNS for helpful discussion. This work was supported by anMRC Research Fellowship held by LdH, and an MRC Pro-gram Grant held by RGMM.
REFERENCES
Amaral DG, Witter MP. 1989. The three-dimensional organization ofthe hippocampal formation: A review of anatomical data. Neuro-science 31:571–591.
Bannerman DM, Good MA, Butcher SP, Ramsay M, Morris RGM.1995. Distinct components of spatial learning revealed by priortraining and NMDA receptor blockade. Nature 378:182–186.
Bannerman DM, Rawlins JN, McHugh SB, Deacon RM, Yee BK,Bast T, Zhang WN, Pothuizen HH, Feldon J. 2004. Regional dis-sociations within the hippocampus—memory and anxiety. Neuro-sci Biobehav Rev 28:273–283.
Barkus C, McHugh SB, Sprengel R, Seeburg PH, Rawlins JN,Bannerman DM. 2010. Hippocampal NMDA receptors and anxi-ety: At the interface between cognition and emotion. Eur J Phar-macol 626:49–56.
Bast T. 2007. Toward an integrative perspective on hippocampal func-tion: From the rapid encoding of experience to adaptive behavior.Rev Neurosci 18:253–281.
Bast T. 2011. The hippocampal learning-behavior translation and thefunctional significance of hippocampal dysfunction in schizophre-nia. Curr Opin Neurobiol 21:492–501.
Bast T, Wilson IA, Witter MP, Morris RGM. 2009. From rapid placelearning to behavioral performance: A key role for the intermediatehippocampus. PLoS Biol 7:e1000089.
Blaha CD, Yang CR, Floresco SB, Barr AM, Phillips AG. 1997. Stim-ulation of the ventral subiculum of the hippocampus evokes gluta-mate receptor-mediated changes in dopamine efflux in the ratnucleus accumbens. Eur J Neurosci 9:902–911.
Broadbent NJ, Squire LR, Clark RE. 2004. Spatial memory, recogni-tion memory, and the hippocampus. Proc Natl Acad Sci USA 101:14515–14520.
Brun VH, Solstad T, Kjelstrup KB, Fyhn M, Witter MP, MoserEI, Moser MB. 2008. Progressive increase in grid scale fromdorsal to ventral medial entorhinal cortex. Hippocampus 18:1200–1212.
Burton BG, Hok V, Save E, Poucet B. 2009. Lesion of the ventral andintermediate hippocampus abolishes anticipatory activity in themedial prefrontal cortex of the rat. Behav Brain Res 199:222–234.
Canals S, Beyerlein M, Merkle H, Logothetis NK. 2009. FunctionalMRI evidence for LTP-induced neural network reorganization.Curr Biol 19:398–403.
de Bruin JP, Sanchez-Santed F, Heinsbroek RP, Donker A, Postmes P.1994. A behavioural analysis of rats with damage to the medialprefrontal cortex using the Morris water maze: Evidence for behav-ioural flexibility, but not for impaired spatial navigation. Brain Res652:323–333.
de Hoz L, Knox J, Morris RGM. 2003. Longitudinal axis of the hip-pocampus: Both septal and temporal poles of the hippocampussupport water maze spatial learning depending on the training pro-tocol. Hippocampus 13:587–603.
de Hoz L, Martin SJ, Morris RGM. 2004. Forgetting, reminding, andremembering: The retrieval of lost spatial memory. PLoS Biol 2:E225.
Dougherty KA, Islam T, Johnston D. 2012. Intrinsic excitability ofCA1 pyramidal neurones from the rat dorsal and ventral hippo-campus. J Physiol 590:5707–5722.
Fanselow MS, Dong HW. 2010. Are the dorsal and ventral hippocam-pus functionally distinct structures? Neuron 65:7–19.
Ferbinteanu J, Ray C, McDonald RJ. 2003. Both dorsal and ventralhippocampus contribute to spatial learning in Long-Evans rats.Neurosci Lett 345:131–135.
Floresco SB, Seamans JK, Phillips AG. 1997. Selective roles forhippocampal, prefrontal cortical, and ventral striatal circuits inradial-arm maze tasks with or without a delay. J Neurosci 17:1880–1890.
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 1003
Hippocampus

Goto Y, Grace AA. 2008. Dopamine modulation of hippocampal-prefrontal cortical interaction drives memory-guided behavior.Cereb Cortex 18:1407–1414.
Grace AA, Floresco SB, Goto Y, Lodge DJ. 2007. Regulation of firingof dopaminergic neurons and control of goal-directed behaviors.Trends Neurosci 30:220–227.
Groenewegen HJ, Vermeulen-Van der Zee E, te Kortschot A, WitterMP. 1987. Organization of the projections from the subiculum tothe ventral striatum in the rat. A study using anterograde trans-port of Phaseolus vulgaris leucoagglutinin. Neuroscience 23:103–120.
Groenewegen HJ, Wright CI, Beijer AV. 1996. The nucleus accum-bens: Gateway for limbic structures to reach the motor system?Prog Brain Res 107:485–511.
Hok V, Save E, Lenck-Santini PP, Poucet B. 2005. Coding for spatialgoals in the prelimbic/infralimbic area of the rat frontal cortex.Proc Natl Acad Sci USA 102:4602–4607.
Hok V, Lenck-Santini PP, Roux S, Save E, Muller RU, Poucet B.2007. Goal-related activity in hippocampal place cells. J Neurosci27:472–482.
Hyman JM, Zilli EA, Paley AM, Hasselmo ME. 2005. Medial pre-frontal cortex cells show dynamic modulation with the hippocam-pal theta rhythm dependent on behavior. Hippocampus 15:739–749.
Inglis J, Martin SJ, Morris RGM. 2013. Upstairs/downstairs revisited:spatial pretraining-induced rescue of normal spatial learning duringselective blockade of hippocampal N-methyl-D-aspartate receptors.Eur J Neurosci 37:718–727.
Ito R, Robbins TW, Pennartz CM, Everitt BJ. 2008. Functional inter-action between the hippocampus and nucleus accumbens shell isnecessary for the acquisition of appetitive spatial context condition-ing. J Neurosci 28:6950–6959.
Jarrard LE. 1989. On the use of ibotenic acid to lesion selectively dif-ferent components of the hippocampal formation. J NeurosciMethods 29:251–259.
Jay TM, Witter MP. 1991. Distribution of hippocampal CA1 andsubicular efferents in the prefrontal cortex of the rat studied bymeans of anterograde transport of Phaseolus vulgaris-leucoaggluti-nin. J Comp Neurol 313:574–586.
Johnson A, van der Meer MA, Redish AD. 2007. Integrating hippo-campus and striatum in decision-making. Curr Opin Neurobiol17:692–697.
Jones MW, Wilson MA. 2005. Theta rhythms coordinatehippocampal-prefrontal interactions in a spatial memory task.PLoS Biol 3:e402.
Jung MW, Wiener SI, McNaughton BL. 1994. Comparison of spatialfiring characteristics of units in dorsal and ventral hippocampus ofthe rat. J Neurosci 14:7347–7356.
Kerr KM, Agster KL, Furtak SC, Burwell RD. 2007. Functional neu-roanatomy of the parahippocampal region: The lateral and medialentorhinal areas. Hippocampus 17:697–708.
Kjelstrup KB, Solstad T, Brun VH, Hafting T, Leutgeb S, Witter MP,Moser EI, Moser MB. 2008. Finite scale of spatial representationin the hippocampus. Science 321:140–143.
Lacroix L, White I, Feldon J. 2002. Effect of excitotoxic lesions of ratmedial prefrontal cortex on spatial memory. Behav Brain Res 133:69–81.
Lepage M, Habib R, Tulving E. 1998. Hippocampal PET activationsof memory encoding and retrieval: The HIPER model. Hippocam-pus 8:313–322.
Loureiro M, Lecourtier L, Engeln M, Lopez J, Cosquer B, Geiger K,Kelche C, Cassel JC, Pereira de Vasconcelos A. 2011. The ventralhippocampus is necessary for expressing a spatial memory. BrainStruct Funct 217:93–106.
Martin SJ, de Hoz L, Morris RGM. 2005. Retrograde amnesia: Nei-ther partial nor complete hippocampal lesions in rats result in pref-
erential sparing of remote spatial memory, even after reminding.Neuropsychologia 43:609–624.
Micheau J, Riedel G, Roloff EL, Inglis J, Morris RGM. 2004.Reversible hippocampal inactivation partially dissociates how andwhere to search in the water maze. Behav Neurosci 118:1022–1032.
Mogenson GJ, Yang CR. 1991. The contribution of basal forebrain tolimbic-motor integration and the mediation of motivation toaction. Adv Exp Med Biol 295:267–290.
Moser E, Moser M-B, Andersen P. 1993. Spatial learning impair-ment parallels the magnitude of dorsal hippocampal lesions, butis hardly present following ventral lesions. J Neurosci 13:3916–3925.
Moser M-B, Moser EI, Forrest E, Andersen P, Morris RGM. 1995.Spatial learning with a minislab in the dorsal hippocampus. ProcNatl Acad Sci USA 92:9697–9701.
Moser M-B, Moser EI. 1998a. Functional differentiation in the hippo-campus. Hippocampus 8:608–619.
Moser M-B, Moser EI. 1998b. Distributed encoding and retrieval ofspatial memory in the hippocampus. J Neurosci 18:7535–7542.
Morris RG, Shenk F, Tweedie F, Jarrard LE. 1990. Ibotenate lesions ofhippocampus and/or subiculum: dissociating components of allo-centric spatial learning. Eur J Neurosci 2:1016–1028.
Naber PA, Witter MP. 1998. Subicular efferents are organized mostlyas parallel projections: A double-labeling, retrograde-tracing studyin the rat. J Comp Neurol 393:284–297.
Nadel L. 1968. Dorsal and ventral hippocampal lesions and behavior.Physiol Behav 3:891–900.
Nadel L, Hoscheidt S, Ryan LR. 2013. Spatial cognition and thehippocampus: The anterior-posterior axis. J Cog Neurosci 25:22–28.
Otnaess MK, Brun VH, Moser M-B, Moser EI. 1999. Pretrainingprevents spatial learning impairment after saturation of hippocam-pal long-term potentiation. J Neurosci 19:RC49.
Poppenk J, Evensmoen HR, Moscovitch M, Nadel L. 2013. Long-axisspecialization of the human hippocampus. Trends Cogn Sci 17:230–240.
Riedel G, Micheau J, Lam AG, Roloff EL, Martin SJ, Bridge H, deHoz L, Poeschel B, McCulloch J, Morris RGM. 1999. Reversibleneural inactivation reveals hippocampal participation in severalmemory processes. Nat Neurosci 2:898–905.
Ruediger S, Spirig D, Donato F, Caroni P. 2012. Goal-orientedsearching mediated by ventral hippocampus early in trial-and-errorlearning. Nat Neurosci 15:1563–1571.
Saucier D, Cain DP. 1995. Spatial learning without NMDA receptor-dependent long-term potentiation. Nature 378:186–189.
Seamans JK, Floresco SB, Phillips AG. 1998. D1 receptor modula-tion of hippocampal-prefrontal cortical circuits integrating spatialmemory with executive functions in the rat. J Neurosci 18:1613–1621.
Small SA. 2002. The longitudinal axis of the hippocampal formation:Its anatomy, circuitry, and role in cognitive function. Rev Neurosci13:183–194.
Smith CN, Wixted JT, Squire LR. 2011. The hippocampus supportsboth recollection and familiarity when memories are strong. J Neu-rosci 31:15693–15702.
Sparks FT, Lehmann H, Sutherland RJ. 2011. Between-systems mem-ory interference during retrieval. Eur J Neurosci 34:780–786.
Spooner RI, Thomson A, Hall J, Morris RG, Salter SH. 1994. TheAtlantis platform: A new design and further developments of Bure-sova’s on-demand platform for the water maze. Learn Mem 1:203–211.
Swanson LW. 1981. A direct projection from Ammon’s horn to pre-frontal cortex in the rat. Brain Res 217:150–154.
Taepavarapruk P, Howland JG, Ahn S, Phillips AG. 2008. Neural cir-cuits engaged in ventral hippocampal modulation of dopamine
1004 DE HOZ AND MARTIN
Hippocampus

function in medial prefrontal cortex and ventral striatum. BrainStruct Funct 213:183–195.
Takashima A, Nieuwenhuis IL, Jensen O, Talamini LM, Rijpkema M,Fernandez G. 2009. Shift from hippocampal to neocortical cen-tered retrieval network with consolidation. J Neurosci 29:10087–10093.
Tse D, Langston RF, Kakeyama M, Bethus I, Spooner PA, Wood ER,Witter MP, Morris RGM. 2007. Schemas and memory consolida-tion. Science 316:76–82.
Tse D, Takeuchi T, Kakeyama M, Kajii Y, Okuno H, Tohyama C,Bito H, Morris RGM. 2011. Schema-dependent gene activa-tion and memory encoding in neocortex. Science 333:891–895.
Tulving E. 1985. How many memory systems are there? Am Psychol40:385–398.
van der Meer MA, Redish AD. 2011a. Theta phase precession in ratventral striatum links place and reward information. J Neurosci31:2843–2854.
van der Meer MA, Redish AD. 2011b. Ventral striatum: A criticallook at models of learning and evaluation. Curr Opin Neurobiol21:387–392.
van Groen T, Wyss JM. 1990. Extrinsic projections from area CA1 ofthe rat hippocampus: olfactory, cortical, subcortical, and bilateral hip-pocampal formation projections. J. Comp.Neurology 302:515–528.
van Kesteren MT, Fernandez G, Norris DG, Hermans EJ. 2010. Per-sistent schema-dependent hippocampal-neocortical connectivityduring memory encoding and postencoding rest in humans. ProcNatl Acad Sci USA 107:7550–7555.
Wang SH, Finnie PS, Hardt O, Nader K. 2012a. Dorsal hippocampusis necessary for novel learning but sufficient for subsequent similarlearning. Hippocampus 22:2157–2170.
Wang SH, Tse D, Morris RGM. 2012b. Anterior cingulate cortex inschema assimilation and expression. Learn Mem 19:315–318.
Yu SS, Johnson JD, Rugg MD. 2012. Hippocampal activity duringrecognition memory co-varies with the accuracy and confidence ofsource memory judgments. Hippocampus 22:1429–1437.
SEPTAL VERSUS TEMPORAL HIPPOCAMPUS AND SPATIAL LEARNING 1005
Hippocampus
Copyright © 2022 FDOKUMEN




















