
Reproductive strategies in two closely related stony corals (Agaricia, Scleractinia)

Published Ahead of Print 7 August 2013. 2013, 87(20):11135. DOI: 10.1128/JVI.01711-13. J. Virol.
Martinez and Christopher S. SullivanChun Jung Chen, Jennifer E. Cox, Rodney P. Kincaid, Angel PolyomavirusRelated Circulating Strains of a Divergent MicroRNA Targetomes of Closely
http://jvi.asm.org/content/87/20/11135Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://jvi.asm.org/content/87/20/11135#ref-list-1at:
This article cites 90 articles, 38 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

Divergent MicroRNA Targetomes of Closely Related CirculatingStrains of a Polyomavirus
Chun Jung Chen,a Jennifer E. Cox,a Rodney P. Kincaid,a Angel Martinez,b Christopher S. Sullivana
The University of Texas at Austin, Molecular Genetics & Microbiology, Austin, Texas, USAa; American Chemical Society Project SEED Summer Internship Program, JamesBowie High School, Austin, Texas, USAb
Hundreds of virus-encoded microRNAs (miRNAs) have been uncovered, but an in-depth functional understanding is lacking formost. A major challenge for the field is separating those miRNA targets that are biologically relevant from those that are not ad-vantageous to the virus. Here, we show that miRNAs from related variants of the polyomavirus simian vacuolating virus 40(SV40) have differing host target repertoires (targetomes) while their direct autoregulatory activity on virus-encoded early geneproducts is completely preserved. These results underscore the importance of miRNA-mediated viral gene autoregulation insome polyomavirus life cycles. More broadly, these findings imply that some host targets of virus-encoded miRNAs are likely tobe of little selective advantage to the virus, and our approach provides a strategy for prioritizing relevant targets.
MicroRNAs (miRNAs) are a class of eukaryotic small RNAmolecules that play a regulatory role in several biological
processes relevant to virus infection, including the immuneresponse, apoptosis, and tumorigenesis (1). Virus-encodedmiRNAs identified from several different families, including theherpes-, retro-, and polyomavirus families, have generated muchinterest as potential effectors of pathogenesis. Over 300 viralmiRNAs have been identified, yet only a small fraction have well-understood functions (2–5). Unlike host miRNAs, most viralmiRNAs are not well conserved, and only �25% or fewer arelikely to serve as mimics or “analogs” of host miRNAs (1, 2). Assuch, identifying the most relevant targets of viral miRNAs is notstraightforward. A valuable approach toward understanding thefunctions of viral miRNAs relies on high-throughput target tran-script identification (6–13). However, it is unlikely that all of theseidentified targets are relevant to the virus infectious cycle, thuslimiting the utility of such approaches as stand-alone platformsfor determining viral miRNA function. Here, we take advantage ofnatural variations in miRNA gene products from closely relatedvirus strains, with the assumption that important miRNA targettranscripts will be preserved throughout evolution.
miRNAs are �22 nucleotides long (reviewed in reference 14)and are derived from primary transcripts (pri-miRNA) contain-ing hairpin precursor molecules (pre-miRNA) (15, 16). The pri-miRNA is cleaved by the double-stranded RNA-specific endonu-clease Drosha to liberate the pre-miRNA (17–19) that is thenexported to the cytoplasm (20–22). There, Dicer further cleavesthe pre-miRNA (23, 24), and typically a single-stranded 22-mer,enriched from one arm of the hairpin, is more abundantly re-tained in the RNA-induced silencing complex (RISC). The other,less-abundant strand is sometimes referred to as the star strand orpassenger strand (14). The 5= end of the 22-mer, referred to as theseed region (nucleotides 2 to 8), is especially important for mRNAtarget binding and typically binds with perfect complementarityto the 3= untranslated region (UTR) of the target transcript (25).Most miRNA-targeted transcripts display impaired translationfollowed by subsequent increased turnover (26), which can man-ifest as an overall decreased steady-state level of the targets (25,27). In addition, although rare for most animal miRNAs, someplant and viral miRNAs can bind with perfect complementarity
(all �22 nucleotides) to their targets and direct small interferingRNA (siRNA)-like cleavage, resulting in robust decreases in thesteady-state levels of the targeted transcripts.
The polyomaviruses are a family of small viruses with circular,double-stranded DNA genomes. Most polyomaviruses arethought to take up lifelong infections of their hosts, although themechanisms for how this occurs are poorly understood. In addi-tion, polyomaviruses can undergo robust lytic infection. There arecurrently 12 known human polyomaviruses, of which at leastfour—Merkel cell polyomavirus (MCPyV), trichodysplasiaspinulosa polyomavirus (TSPyV), BK virus (BKPyV), and JC virus(JCPyV)—are associated with serious disease in immunosup-pressed humans (28–32). Simian vacuolating virus 40 (SV40), aprototypic polyomavirus, undergoes lytic infection in culturedAfrican green monkey cells and as such has been a valuable labo-ratory model for polyomavirus infection (33). We have previouslydemonstrated that several members of the Polyomaviridae (SV40,BK virus, JC virus, simian agent 12 [SA12], murine polyomavirus[muPyV], and MCPyV) express miRNAs that lie antisense to theearly transcripts and possess the ability to cleave these transcriptsvia an siRNA-like mechanism (34–38). The conserved nature ofthis mode of autoregulation among divergent polyomaviruses im-plies importance. However, at least three observations could sug-gest otherwise. First, the degree of regulation imparted by themiRNA is partial. That is, at least in the laboratory models of lyticinfection (35, 37), a high fraction (�50%) of intact early tran-scripts remains uncleaved by the viral miRNA RISC (35, 37). Sec-ond, a host target has been reported for the JC virus star strandmiRNA (39). Third, our data suggest that at least some strains of
Received 25 June 2013 Accepted 30 July 2013
Published ahead of print 7 August 2013
Address correspondence to Christopher S. Sullivan,[email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JVI.01711-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01711-13
October 2013 Volume 87 Number 20 Journal of Virology p. 11135–11147 jvi.asm.org 11135
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

polyomavirus likely do not encode miRNAs (J. E. Cox and C. S.Sullivan, unpublished data). Therefore, it remains to be deter-mined if autoregulation of the antisense early transcripts is trulyimportant in the polyomaviral life cycle.
Here, we address the question of whether polyomaviralmiRNA-mediated autoregulation of the early transcripts is a rele-vant activity or, rather, is a consequence of the genomic location ofthe polyomavirus miRNAs (antisense and therefore necessarilyperfectly complementary to the early transcripts). We screened all63 deposited fully sequenced isolates of SV40 for possible varia-tions in their pre-miRNAs and derivative miRNAs. We uncovered17 different classes of pre-miRNA primary sequence variants,some of which result in different miRNA products. We identifieda naturally circulating variant virus (the RI257 strain) that gener-ates derivative miRNAs, all possessing different seeds than themiRNA derivatives from the majority of SV40 isolates. We showthat, as would be predicted from the altered seed repertoires, thereference strain 776 miRNAs target a different repertoire of hosttranscripts than do those of the RI257 strain. However, strikingly,the RI257 strain miRNAs efficiently autoregulate early transcriptlevels to a similar degree as those of strain 776. These results un-derscore the likely importance of SV40 miRNA-mediated auto-regulation of viral gene expression. Furthermore, this work dem-onstrates that highly similar viruses can tolerate substantialvariability in their miRNA targetomes.
MATERIALS AND METHODSSV40 sequence analysis and alignment. Sixty-three unique SV40 com-plete genome sequences were aligned based on an �120-bp region en-compassing the pre-miRNA, by using Geneious software (Biomatters,New Zealand).
Cell culture and RNA isolation. Human embryonic kidney (HEK)cells (293 and 293T cells), African green monkey kidney epithelial cells(BSC-40 cells), and African green monkey kidney fibroblast cells (COS-7cells) were obtained from the American Type Culture Collection (Manas-sas, VA). All cells were maintained in Dulbecco’s modified Eagle’s me-dium (DMEM) supplemented with 10% fetal bovine serum (FBS; LifeTechnologies, NY). Total RNA was harvested using an in-house phenol,isoamyl alcohol, guanidinium isothiocyanate, and beta-mercaptoethanol(PIG-B) solution as described previously (36, 40–42).
Vector construction, transfection, and high-resolution Northernblot analysis. All DNA vector constructs were confirmed by sequenceanalysis through the Institute of Cellular and Molecular Biology Sequenc-ing Facility at the University of Texas at Austin. For a list of the primersused in the construction of the 17 representative SV40 microRNA expres-sion vectors, see Table S1 in the supplemental material. Briefly, the prim-ers were annealed and filled in by using Phusion high-fidelity DNA poly-merase (New England BioLabs, MA) according to the manufacturer’sprotocol. The PCR products were cloned into the KpnI/XhoI sites of thepcDNA3.1neo expression vector.
293T cells were plated in 6-well plates and transfected using Lipo-fectamine 2000 transfection reagent (Life Technologies) according to themanufacturer’s instruction. 293T cells were also transfected with theempty pcDNA3.1neo vector as a negative control. Total RNA was har-vested at 48 h posttransfection and subjected to a modified version ofhigh-resolution Northern blot analysis (43). Briefly, 30 �g of total RNAwas separated on a Tris-borate-EDTA-urea-15% polyacrylamide gel. Thebromophenol blue marker was allowed to migrate 30 cm along the lengthof the gel. The RNA was transferred into an Amersham Hybond N� mem-brane (GE Healthcare, PA) and probed for miRNA as previously de-scribed (44). Quantification of the band signals was performed usingQuantity One software (Bio-Rad, CA) The probe sequences used were asfollows: 776-5p probe (5p and 3p refer to the 5= and 3= arm, respectively,
of the miRNA stem-loop precursor), CAAGGCTCATTTCAGGC;RI257-5p probe, CAACGCACATTTCAGTC; MC-028846B-5p probe,CAAAGCTCATTTCAGGC; SV40-3p probe, CTCAGGGCATGAAACAGGC.
Construction of the RI257-MIR virus. To generate the RI257-MIRchimeric virus, overlapping PCR was used to generate two fragmentsjoined together via a linker, by using Phusion high-fidelity DNA polymer-ase (New England BioLabs). The resulting fragment was inserted into theNheI/BstXI sites of the pSVB3 vector to generate pRI257-MIR. TheRI257-MIR chimeric virus was produced as described previously (45).Briefly, pRI257-MIR was digested with BamHI, followed by intramolec-ular ligation of the viral DNA. The ligation reaction was transfected intoBSC-40 cells and amplified. The primers used were as follows: linerprimer, GACTGTGAAGATTCGGGGGACTGAAATGTGCGTTGTGATTGTGAGTCAACGCCTGTTTCATGCCCTGAGTCTTCTAGGTTGTC;fragment 1 forward primer, TATCGTCCATTCCGACAG; fragment 1 re-verse primer, CACATTTCAGTCCCCCGAATCTTCACAGTCTGTTTATGATCACAATCAACCATATCACATTTGTAAAGGTTTTACTTGCTTTAAAAAACC; fragment 2 forward primer, TGTTTCATGCCCTGAGTCTTCTAGGTTGTCATTCCCCGCCTGTTTTTCCTGCACATTTTCCTCCTCAGCATCATCATCACTGTTTCTTAG; fragment 2 reverse primer,ACTGCAAACAATGGCCTG.
5= RACE analysis to map the cleavage site of early transcripts.BSC-40 cells were seeded in a T25 tissue culture flask and infected withstrain 776 or the RI257-MIR virus at a multiplicity of infection (MOI) of10 as described in the SV40 protocol book (45, 46). Total RNA was har-vested at 60 h postinfection (hpi) as described above. The total RNA wasfurther purified using an Oligotex mRNA Mini Kit (Qiagen, CA) accord-ing to the manufacturer’s protocol. 5= rapid amplification of cDNA ends(RACE) was performed using a FirstChoice RNA ligase-mediated rapidamplification of cDNA ends (RLM-RACE) kit (Life Technologies) ac-cording to the manufacturer’s protocol. The reverse transcription reac-tion was performed using SuperScript III (Life Technologies) according tothe manufacturer’s protocol. The reverse-transcribed cDNA was sub-jected to RNaseH (Life Technologies) treatment at 37°C for 20 min. Onemicroliter of the reverse transcription product was used as the templatefor the first round of nested PCR using Taq DNA polymerase (New Eng-land BioLabs). One microliter of the first-round PCR was then used in thesecond round of nested PCR using Taq DNA polymerase. The PCR prod-ucts from both rounds were TA cloned into pCR2.1-TOPO using a TOPOTaq polymerase-amplified (TA) cloning kit (Life Technologies) accordingto the manufacturer’s protocol. A combined 15 clones from each infectionsample were sequence analyzed through the Institute of Cellular and Mo-lecular Biology Sequencing Facility at the University of Texas at Austin.The primers used were as follows: reverse transcription primer, GCTTTATTTGTAACCATTATAAGCTGCAATAAACAAGT; 5= RACE first-round PCR forward primer, GCTGATGGCGATGAATGAACACTG; 5=RACE first-round reverse primer, TAACAACAACAATTGCATTCATTTTATGTTTCAGGTTC; 5= RACE second-round PCR forward primer, CGCGGATCCGAACACTGCGTTTGCTGGCTTTGATG; 5= RACE second-round PCR reverse primer, AGGGGGAGGTGTGGGAGGTTTTTTAAAGCAAGTAAAAC.
Luciferase assays. The cellular 3= UTR reporters were constructed bycloning fragments of the corresponding genomic regions of the targetsfrom 293T genomic DNA using the primers listed in Table S2 in thesupplemental material. Briefly, the PCR products are generated usingKOD Hot Start DNA polymerase (EMD Millipore, MA) according to themanufacturer’s protocol and cloned into the pcDNA3.1dsRluc vector,which expressed a destabilized version of Renilla luciferase. The restric-tion sites used are listed in Table S2. The ARCN1 seed mutant reportercontains engineered point mutations at the fourth and fifth nucleotidescomplementary to the 5= end of the 776-3p long (776-3pL) miRNA. TheC9orf140 seed mutant reporter contains engineered point mutations atthe second and third nucleotides complementary to the 5= end of theRI257-5p miRNA. The seed mutant reporters were constructed with the
Chen et al.
11136 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

corresponding wild-type reporter as the template for site-directed mu-tagenesis using PfuUltra II fusion HS DNA polymerase (Agilent Technol-ogies, CA), according to the manufacturer’s protocol. The primers usedfor constructing the seed mutant reporters are listed in Table S2. 293Tcells were plated in 24-well plates and transfected using TurboFect trans-fection reagent (Thermo Scientific, PA) according to the manufacturer’sprotocol. Cells were transfected with the reporter and the miRNA expres-sion vector. 293T cells were also transfected with the empty reporter as anegative control. The pcDNA3.1Luc2CP vector was also cotransfected tonormalize for transfection efficiency. Cells were collected 24 h posttrans-fection and analyzed with a dual-luciferase reporter assay system (Pro-mega, WI) according to the manufacturer’s instructions. The luciferasereadings were collected using a Luminoskan Ascent microplate luminom-eter (Thermo Scientific). Results from the Renilla luciferase were normal-ized to the firefly luciferase readings, and the ratios to results with theempty vector control were plotted as a bar graph.
The 776/RI257-5p miRNA perfect-match reporter contains twomiRNA binding sites that are perfectly complementary to the 776-5pmiRNA and two miRNA binding sites that are perfectly complementaryto the RI257-5p miRNA. The 776/RI257-3p miRNA perfect-match re-porter is likewise, except with a perfect match to the 3p miRNAs. Thereporters were generated by first annealing synthesized oligonucleotidescarrying the miRNA sites, followed by PCR amplification and addition ofrestriction sites at both ends, using Phusion high-fidelity DNA polymer-ase (New England BioLabs). The PCR products were cloned into thepcDNA3.1dsRluc vector via the XhoI/XbaI sites. The dual-luciferase assaywas performed with 293 cells as described above. 293 cells were also trans-fected with the reporters along with the Merkel cell polyomavirus (MCV)miRNA expression vector as a negative control. As a second negativecontrol, the MCV miRNA reporter containing binding sites that are per-fectly complementary to the MCV miRNA was transfected as well. Theprimers used were as follows: 5p reporter forward oligonucleotide, CCCAAGGCTCATTTCAGGCCCCTCAGTCCTGTCCCCAAGGCTCATTTCAGGCCCCTCAGTCCTGTGATCACAACGC; 5p reporter reverse oli-gonucleotide, GATTCGGGGGACTGAAATGTGCGTTGTGATGAGGATTCGGGGGACTGAAATGTGCGTTGTGATCACAGGACTGTGG; 5preporter forward primer, ATCGCTCGAGCCCAAGGCTCATTTCAGGCCCCTCAGTCCTGTC; 5p reporter reverse primer, ATCGTCTAGAGATTCGGGGGACTGAAATGTGCGTTGTGATGAG; 3p reporter forwardoligonucleotide, GGAAGACTCAGGGCATGAAACAGGCATTGAGTCGGAAGACTCAGGGCATGAAACAGGCATTGAGTGGAAGACTCAG;3p reporter reverse oligonucleotide, GTCAACGCCTGTTTCATGCCCTGAGTCTTCGAGGTCAACGCCTGTTTCATGCCCTGAGTCTTCCACTCAATGCCTG; 3p reporter forward primer, ATCGCTCGCGGGAAGACTCAGGGCATGAAACAGGCATTGAGTC, 3p reporter reverse primer,ATCGTCTAGAGTCAACGCCTGTTTCATGCCCTGAGTCTTCGAG.
SV40 infections. BSC-40 cells were seeded in 6-well plates. The cellswere infected with either reference strain 776, the miRNA null mutant SM(35), or the RI257-MIR virus at an MOI of 10 when the cells were freshlyconfluent. The media from the plates were aspirated, and 500 �l of virusinoculum was used per well. The plates were rocked back and forth every15 min for 2 h at 37°C (46). The virus inoculum was replaced with DMEMwith 2% FBS. The titers of all three viruses are �1.0 � 108 PFU/ml. Virustiters were determined by using a modified version of the protocol de-scribed in Drayman et al. 47. Briefly, BSC-40 cells were seeded in 6-wellplates and infected with serially diluted strain 776, mutant strain SM, orthe RI257-MIR virus as described above. The infected cells were collectedby trypsinization and fixed in 4% paraformaldehyde in phosphate-buff-ered saline (PBS) for 20 min at 37°C. The fixed cells were then permeab-ilized using 3% bovine serum albumin (BSA) in PBS with 0.1% TritonX-100 at room temperature for 10 min. The cells were washed with PBSand stained for large T antigen using pAb416 antibodies (kindly providedby Jim Pipas, University of Pittsburgh) for 1 h at room temperature. Thecells were then washed 3 times with PBS, followed by secondary antibodyincubation with Alexa Fluor 488 goat anti-mouse antibody (Life Technol-
ogies) for 1 h at room temperature. The cells were washed 3 times withPBS and analyzed using a BD LSRFortessa cell analyzer (BD Biosciences,CA).
Small RNA library generation and computational analysis of se-quencing reads. BSC-40 cells were seeded in a T75 tissue culture flask andinfected with either strain 776 or the RI257-MIR virus at an MOI of 10 asdescribed above. Total RNA from strain 776- and RI257-MIR virus-in-fected BSC-40 cells was harvested at 40 hpi. Two hundred micrograms oftotal RNA was gel fractionated to isolate small RNAs. The small RNAsfrom strain 776 infection was subjected to SOLiD sequencing as previ-ously described (40, 41). The small RNAs from RI257-MIR virus infectionwere subjected to Illumina sequencing as described previously (48).
FACS and microarray analysis. 293T cells were seeded in 10-cmdishes and transfected with either empty pcDNA3.1neo vector, BPCV1miRNA expression vector, strain 776 miRNA expression vector, or strainRI257 miRNA expression vector using X-tremeGENE 9 transfection re-agent (Roche, IN). pEGFP vector that expresses an enhanced version ofthe green fluorescent protein (GFP) was cotransfected as a transfectioncontrol. Cells were trypsinized at 48 h posttransfection and sorted basedon the GFP signal. Cell sorting was performed by the Institute of Cellularand Molecular Biology fluorescence-activated cell sorting (FACS) facilityat the University of Texas at Austin. Total RNA was harvested from theGFP-positive sorted fraction. Ten micrograms of the total RNA wastreated with DNase (Qiagen) followed by purification using an RNeasyMinElute cleanup kit (Qiagen) according to the manufacturer’s instruc-tions. RNA integrity was verified with 1.0% denaturing morpholinepro-panesulfonic acid (MOPS)-formaldehyde-agarose gel electrophoresis.The purified RNA was used as a template to make biotin-labeled cRNAusing an Illumina TotalPrep RNA amplification kit (Ambion) accordingto the manufacturer’s guidelines. Labeled cRNA was precipitated over-night with isopropanol and sodium acetate. Biotinylated cRNA is hybrid-ized to Illumina HumanHT-12 v4.0 microArray chips at the KeckBiotechnology Resource Laboratory (Yale University) according to Illu-mina’s protocols. Quality control and data analysis were carried out ac-cording to the instructions provided by Illumina.
BSC-40 cells were seeded in a T25 tissue culture flask and infected withstrain 776 or the RI257-MIR virus at an MOI of 10 as described above.Total RNA was harvested at 44 hpi. RNA purification and microarrayanalysis were performed as described above. DAVID functional annota-tion clustering analysis was performed at the highest stringency settingusing online-based software (DAVID Bioinformatics Resources 6.7 [49,50]).
Northern blot analysis of cleavage fragments of early transcripts.BSC-40 cells were seeded in a T25 tissue culture flask and infected withstrain 776 or the RI257-MIR virus at an MOI of 10 as described above.Total RNA was harvested at 12, 36, and 60 hpi as described above. Twomicrograms of total RNA was subjected to 1.25% denaturing MOPS-formaldehyde-agarose gel electrophoresis as described previously (95).The RNA was transferred onto a Nytran SPC nylon transfer membrane(Whatman, NJ) using a TurboBlotter System (Whatman) according tothe manufacturer’s instructions. The membrane was probed for the cleav-age fragment of viral early transcripts in ExpressHyb hybridization solu-tion (Clontech, CA) at 45°C. Quantification of the band signals was per-formed using Quantity One software (Bio-Rad). The probe sequencesused were as follows: SV40 early 3p cleavage probe 1, GAAAAAAATGCTTTATTTGTGAAATTTGTGATGCTATT; probe 2, GCTTTATTTGTAACCATTATAAGCTGCAATAAACAAGT; probe 3, TAACAACAACAATTGCATTCATTTTATGTTTCAGGTTC; probe 4, AGGGGGAGGTGTGGGAGGTTTTTTAAAGCAAGTAAAAC; probe 5, CTCTACAAATGTGGTATGGCTGATTATGATCATGAACA.
RESULTSIdentification of 17 classes of sequence variants in the SV40 pre-miRNA genomic region. Polyomaviruses are common humanpathogens that can be associated with cancer and other serious
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11137
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

diseases in immunosuppressed patients (28–32). We have previ-ously identified several different animal and human polyomavi-ruses that produce miRNAs capable of autoregulating early viralgene expression (34–38). Whether this function is beneficial topolyomavirus biology remains unknown. The fact that this modeof gene regulation is conserved among diverse polyomavirusesimplies importance. However, it is also possible that targeting nu-merous host transcripts is a key function of these miRNAs. Teas-ing out the relevant importance of these two non-mutually exclu-sive models is complicated by the fact that no variations in themiRNA seed sequences have been reported for different strains ofthe same virus. Such variants could allow prioritization of themost relevant targets, since even single nucleotide changes in theseed can direct a different efficiency of activity and spectrum oftarget transcripts (51–54). Presumably, only important targets,host or viral, will be preserved among the variant viruses. There-fore, we first set out to identify isolates of the same virus speciesthat give rise to variant derivative miRNAs.
Our strategy to identify miRNA seed variants of the same spe-cies is outlined in Fig. 1A. SV40 is one of the best-studied polyo-maviruses, with the full genomes of 63 different isolates depositedin GenBank. Therefore, we focused on SV40. We first identified alldeposited strains that possess nucleotide variation in the generalpre-miRNA region of the genome. Because flanking regions canaffect the processing of pre-miRNAs (19), we defined our regionof interest as containing the predicted hairpin stem-loop structureplus an additional 10 nucleotides on either side of the hairpin
(nucleotides 2764 to 2881 in the 776 reference strain). From the 63isolates examined, we identified 17 different classes that contain atleast a single nucleotide change in the pre-miRNA and/or nearbyflanking regions (Fig. 1B). Most classes contained only one or twonucleotide substitutions or had larger duplications or insertionsthat were not predicted to dramatically alter the secondary struc-ture of the pre-miRNAs. One notable exception was strain RI257,which contains 22 individual nucleotide changes (Fig. 1B) butnonetheless preserves a high-scoring predicted pre-miRNA struc-ture (VmiR analysis [44]; data not shown). Thus, we identified 17different classes of SV40 strains that could possibly give rise toaltered miRNA derivatives.
Strain RI257: a variant SV40 strain with altered pre-miRNAprocessing and a different seed repertoire. As an initial screen foridentifying derivative miRNA variants, we conducted high-reso-lution Northern blot analysis (43) of the 17 different classes ofSV40 pre-miRNA and miRNA derivatives. We synthesized thegenomic regions encompassing each pre-miRNA sequence vari-ant and engineered expression vectors for a single strain that wasrepresentative of each of the 17 classes of sequence variants (Fig.1). We transfected cells, harvested total RNA, and conductedhigh-resolution denaturing polyacrylamide gel electrophoresis.Blots were then probed with radioactive oligonucleotides. Theseresults showed that 14 of the 17 representative strains displayed aband pattern similar to that of reference strain 776. Consistentwith previous low-resolution Northern analysis (35), multiplemiRNA derivatives are observed for the strain 776-like miRNAs.
FIG 1 Sequence alignment of 63 deposited SV40 genome sequences reveals 17 classes of variants in the pre-miRNA genomic region. (A) Experimental workflowfor identification of unique pre-miRNA variants of SV40. Sixty-three fully sequenced SV40 genomes were aligned, and this resulted in 17 classes of variants in thepre-miRNA genomic region. The 17 pre-miRNA genomic regions were synthesized and cloned into pcDNA3.1neo. 293T cells were transfected with theexpression vectors. RNA was harvested for high-resolution Northern blot analysis and next-generation sequencing. CMV, human cytomegalovirus promoter/enhancer in the pcDNA3.1neo vector. (B) Sequence alignment of the 17 classes of variants in the pre-miRNA genomic region. Each class was numbered from 1to 17, and the strain names that were mentioned in this study are indicated. Polymorphic bases are highlighted: A is in red, T in green, C in purple, and G in yellow.The pre-miRNA genomic region of 776 was used as the reference strain in the alignment process, and the corresponding 776 pre-miRNA genomic location isnumbered at the 5= and 3= end of the alignment. The 5p (black) and 3p (blue) miRNA derivatives are underlined and indicated by the arrows.
Chen et al.
11138 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
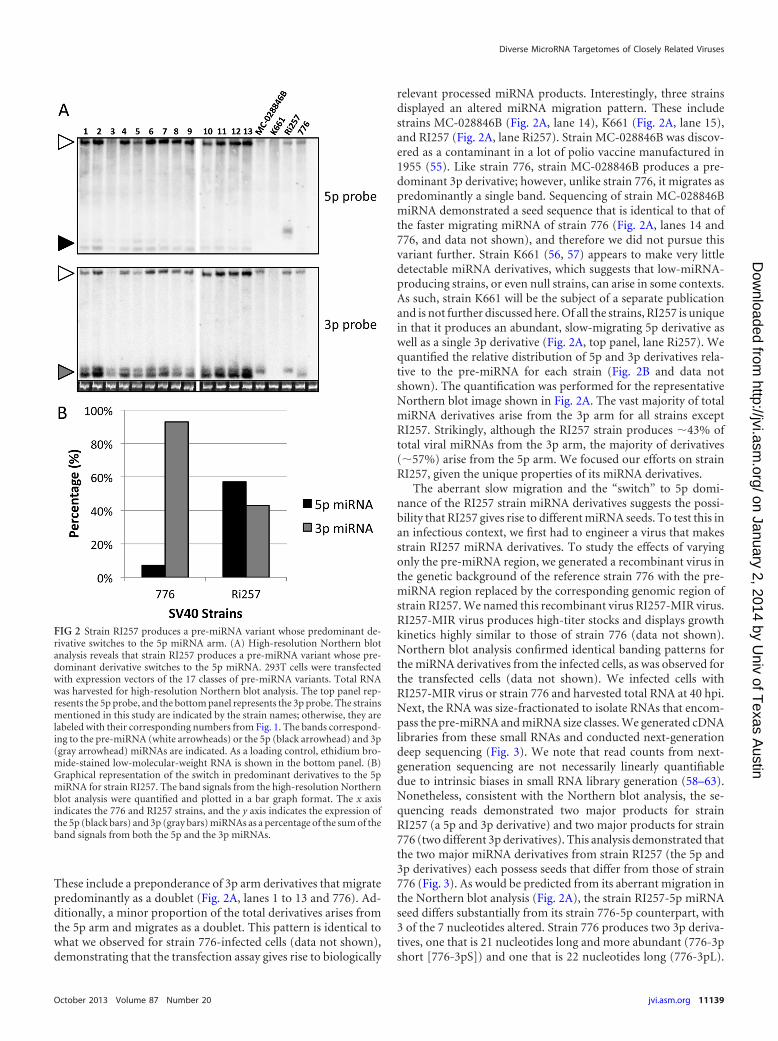
These include a preponderance of 3p arm derivatives that migratepredominantly as a doublet (Fig. 2A, lanes 1 to 13 and 776). Ad-ditionally, a minor proportion of the total derivatives arises fromthe 5p arm and migrates as a doublet. This pattern is identical towhat we observed for strain 776-infected cells (data not shown),demonstrating that the transfection assay gives rise to biologically
relevant processed miRNA products. Interestingly, three strainsdisplayed an altered miRNA migration pattern. These includestrains MC-028846B (Fig. 2A, lane 14), K661 (Fig. 2A, lane 15),and RI257 (Fig. 2A, lane Ri257). Strain MC-028846B was discov-ered as a contaminant in a lot of polio vaccine manufactured in1955 (55). Like strain 776, strain MC-028846B produces a pre-dominant 3p derivative; however, unlike strain 776, it migrates aspredominantly a single band. Sequencing of strain MC-028846BmiRNA demonstrated a seed sequence that is identical to that ofthe faster migrating miRNA of strain 776 (Fig. 2A, lanes 14 and776, and data not shown), and therefore we did not pursue thisvariant further. Strain K661 (56, 57) appears to make very littledetectable miRNA derivatives, which suggests that low-miRNA-producing strains, or even null strains, can arise in some contexts.As such, strain K661 will be the subject of a separate publicationand is not further discussed here. Of all the strains, RI257 is uniquein that it produces an abundant, slow-migrating 5p derivative aswell as a single 3p derivative (Fig. 2A, top panel, lane Ri257). Wequantified the relative distribution of 5p and 3p derivatives rela-tive to the pre-miRNA for each strain (Fig. 2B and data notshown). The quantification was performed for the representativeNorthern blot image shown in Fig. 2A. The vast majority of totalmiRNA derivatives arise from the 3p arm for all strains exceptRI257. Strikingly, although the RI257 strain produces �43% oftotal viral miRNAs from the 3p arm, the majority of derivatives(�57%) arise from the 5p arm. We focused our efforts on strainRI257, given the unique properties of its miRNA derivatives.
The aberrant slow migration and the “switch” to 5p domi-nance of the RI257 strain miRNA derivatives suggests the possi-bility that RI257 gives rise to different miRNA seeds. To test this inan infectious context, we first had to engineer a virus that makesstrain RI257 miRNA derivatives. To study the effects of varyingonly the pre-miRNA region, we generated a recombinant virus inthe genetic background of the reference strain 776 with the pre-miRNA region replaced by the corresponding genomic region ofstrain RI257. We named this recombinant virus RI257-MIR virus.RI257-MIR virus produces high-titer stocks and displays growthkinetics highly similar to those of strain 776 (data not shown).Northern blot analysis confirmed identical banding patterns forthe miRNA derivatives from the infected cells, as was observed forthe transfected cells (data not shown). We infected cells withRI257-MIR virus or strain 776 and harvested total RNA at 40 hpi.Next, the RNA was size-fractionated to isolate RNAs that encom-pass the pre-miRNA and miRNA size classes. We generated cDNAlibraries from these small RNAs and conducted next-generationdeep sequencing (Fig. 3). We note that read counts from next-generation sequencing are not necessarily linearly quantifiabledue to intrinsic biases in small RNA library generation (58–63).Nonetheless, consistent with the Northern blot analysis, the se-quencing reads demonstrated two major products for strainRI257 (a 5p and 3p derivative) and two major products for strain776 (two different 3p derivatives). This analysis demonstrated thatthe two major miRNA derivatives from strain RI257 (the 5p and3p derivatives) each possess seeds that differ from those of strain776 (Fig. 3). As would be predicted from its aberrant migration inthe Northern blot analysis (Fig. 2A), the strain RI257-5p miRNAseed differs substantially from its strain 776-5p counterpart, with3 of the 7 nucleotides altered. Strain 776 produces two 3p deriva-tives, one that is 21 nucleotides long and more abundant (776-3pshort [776-3pS]) and one that is 22 nucleotides long (776-3pL).
FIG 2 Strain RI257 produces a pre-miRNA variant whose predominant de-rivative switches to the 5p miRNA arm. (A) High-resolution Northern blotanalysis reveals that strain RI257 produces a pre-miRNA variant whose pre-dominant derivative switches to the 5p miRNA. 293T cells were transfectedwith expression vectors of the 17 classes of pre-miRNA variants. Total RNAwas harvested for high-resolution Northern blot analysis. The top panel rep-resents the 5p probe, and the bottom panel represents the 3p probe. The strainsmentioned in this study are indicated by the strain names; otherwise, they arelabeled with their corresponding numbers from Fig. 1. The bands correspond-ing to the pre-miRNA (white arrowheads) or the 5p (black arrowhead) and 3p(gray arrowhead) miRNAs are indicated. As a loading control, ethidium bro-mide-stained low-molecular-weight RNA is shown in the bottom panel. (B)Graphical representation of the switch in predominant derivatives to the 5pmiRNA for strain RI257. The band signals from the high-resolution Northernblot analysis were quantified and plotted in a bar graph format. The x axisindicates the 776 and RI257 strains, and the y axis indicates the expression ofthe 5p (black bars) and 3p (gray bars) miRNAs as a percentage of the sum of theband signals from both the 5p and the 3p miRNAs.
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11139
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
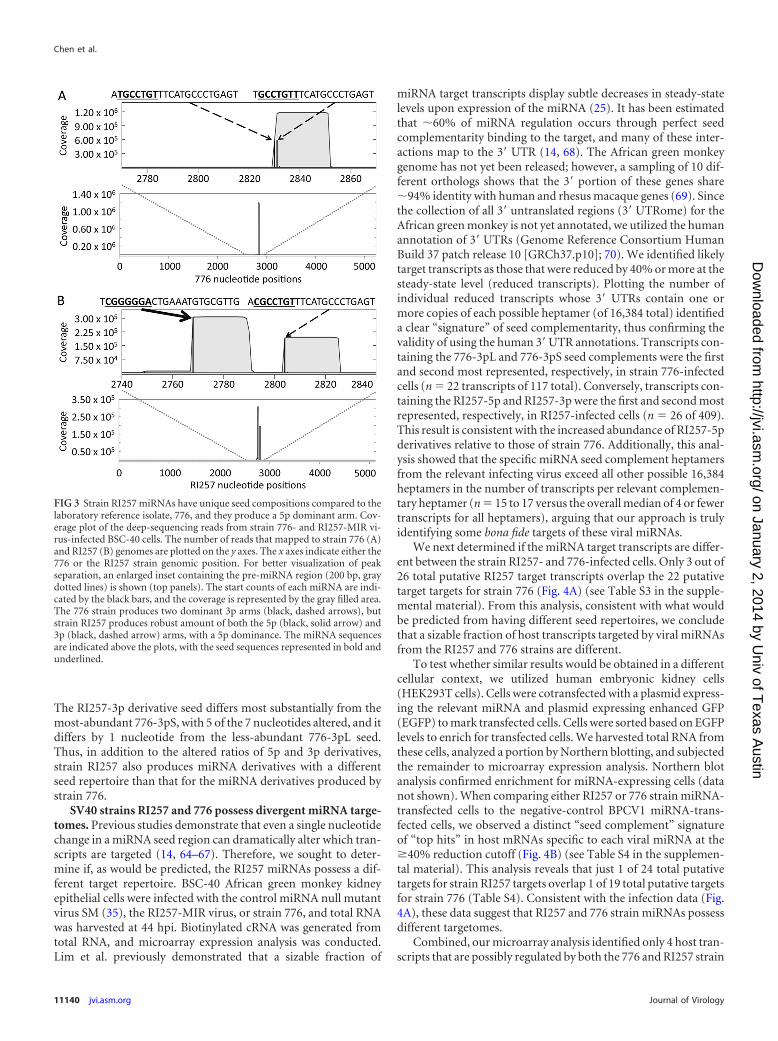
The RI257-3p derivative seed differs most substantially from themost-abundant 776-3pS, with 5 of the 7 nucleotides altered, and itdiffers by 1 nucleotide from the less-abundant 776-3pL seed.Thus, in addition to the altered ratios of 5p and 3p derivatives,strain RI257 also produces miRNA derivatives with a differentseed repertoire than that for the miRNA derivatives produced bystrain 776.
SV40 strains RI257 and 776 possess divergent miRNA targe-tomes. Previous studies demonstrate that even a single nucleotidechange in a miRNA seed region can dramatically alter which tran-scripts are targeted (14, 64–67). Therefore, we sought to deter-mine if, as would be predicted, the RI257 miRNAs possess a dif-ferent target repertoire. BSC-40 African green monkey kidneyepithelial cells were infected with the control miRNA null mutantvirus SM (35), the RI257-MIR virus, or strain 776, and total RNAwas harvested at 44 hpi. Biotinylated cRNA was generated fromtotal RNA, and microarray expression analysis was conducted.Lim et al. previously demonstrated that a sizable fraction of
miRNA target transcripts display subtle decreases in steady-statelevels upon expression of the miRNA (25). It has been estimatedthat �60% of miRNA regulation occurs through perfect seedcomplementarity binding to the target, and many of these inter-actions map to the 3= UTR (14, 68). The African green monkeygenome has not yet been released; however, a sampling of 10 dif-ferent orthologs shows that the 3= portion of these genes share�94% identity with human and rhesus macaque genes (69). Sincethe collection of all 3= untranslated regions (3= UTRome) for theAfrican green monkey is not yet annotated, we utilized the humanannotation of 3= UTRs (Genome Reference Consortium HumanBuild 37 patch release 10 [GRCh37.p10]; 70). We identified likelytarget transcripts as those that were reduced by 40% or more at thesteady-state level (reduced transcripts). Plotting the number ofindividual reduced transcripts whose 3= UTRs contain one ormore copies of each possible heptamer (of 16,384 total) identifieda clear “signature” of seed complementarity, thus confirming thevalidity of using the human 3= UTR annotations. Transcripts con-taining the 776-3pL and 776-3pS seed complements were the firstand second most represented, respectively, in strain 776-infectedcells (n � 22 transcripts of 117 total). Conversely, transcripts con-taining the RI257-5p and RI257-3p were the first and second mostrepresented, respectively, in RI257-infected cells (n � 26 of 409).This result is consistent with the increased abundance of RI257-5pderivatives relative to those of strain 776. Additionally, this anal-ysis showed that the specific miRNA seed complement heptamersfrom the relevant infecting virus exceed all other possible 16,384heptamers in the number of transcripts per relevant complemen-tary heptamer (n � 15 to 17 versus the overall median of 4 or fewertranscripts for all heptamers), arguing that our approach is trulyidentifying some bona fide targets of these viral miRNAs.
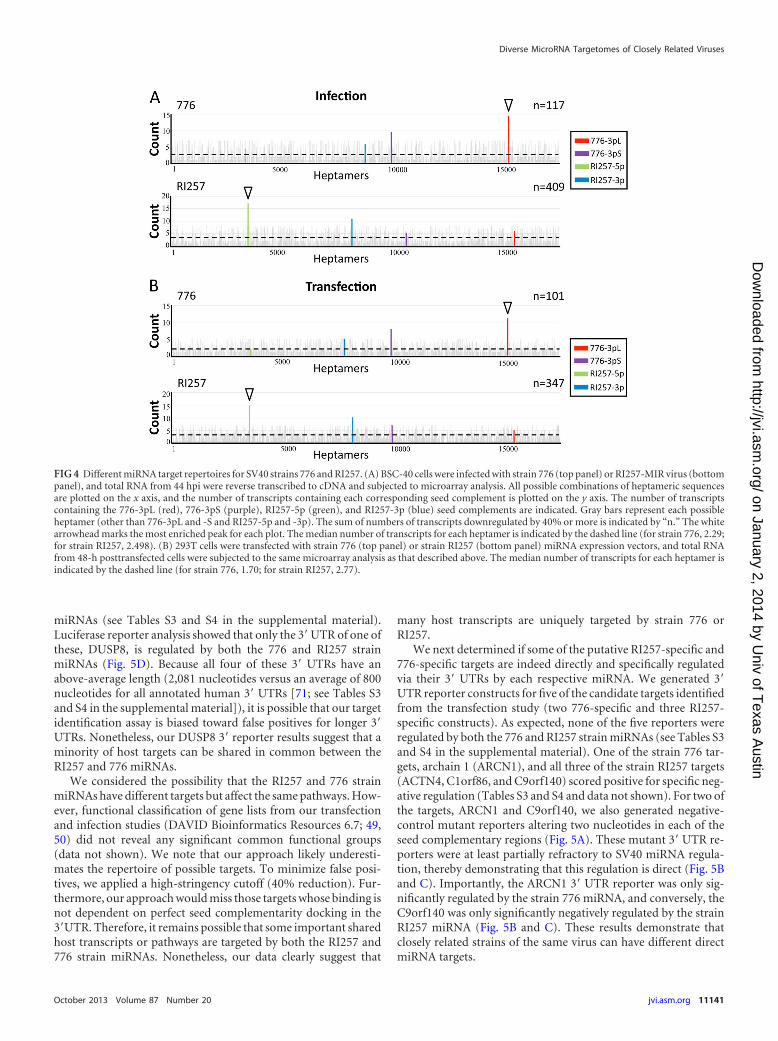
We next determined if the miRNA target transcripts are differ-ent between the strain RI257- and 776-infected cells. Only 3 out of26 total putative RI257 target transcripts overlap the 22 putativetarget targets for strain 776 (Fig. 4A) (see Table S3 in the supple-mental material). From this analysis, consistent with what wouldbe predicted from having different seed repertoires, we concludethat a sizable fraction of host transcripts targeted by viral miRNAsfrom the RI257 and 776 strains are different.
To test whether similar results would be obtained in a differentcellular context, we utilized human embryonic kidney cells(HEK293T cells). Cells were cotransfected with a plasmid express-ing the relevant miRNA and plasmid expressing enhanced GFP(EGFP) to mark transfected cells. Cells were sorted based on EGFPlevels to enrich for transfected cells. We harvested total RNA fromthese cells, analyzed a portion by Northern blotting, and subjectedthe remainder to microarray expression analysis. Northern blotanalysis confirmed enrichment for miRNA-expressing cells (datanot shown). When comparing either RI257 or 776 strain miRNA-transfected cells to the negative-control BPCV1 miRNA-trans-fected cells, we observed a distinct “seed complement” signatureof “top hits” in host mRNAs specific to each viral miRNA at the�40% reduction cutoff (Fig. 4B) (see Table S4 in the supplemen-tal material). This analysis reveals that just 1 of 24 total putativetargets for strain RI257 targets overlap 1 of 19 total putative targetsfor strain 776 (Table S4). Consistent with the infection data (Fig.4A), these data suggest that RI257 and 776 strain miRNAs possessdifferent targetomes.
Combined, our microarray analysis identified only 4 host tran-scripts that are possibly regulated by both the 776 and RI257 strain
FIG 3 Strain RI257 miRNAs have unique seed compositions compared to thelaboratory reference isolate, 776, and they produce a 5p dominant arm. Cov-erage plot of the deep-sequencing reads from strain 776- and RI257-MIR vi-rus-infected BSC-40 cells. The number of reads that mapped to strain 776 (A)and RI257 (B) genomes are plotted on the y axes. The x axes indicate either the776 or the RI257 strain genomic position. For better visualization of peakseparation, an enlarged inset containing the pre-miRNA region (200 bp, graydotted lines) is shown (top panels). The start counts of each miRNA are indi-cated by the black bars, and the coverage is represented by the gray filled area.The 776 strain produces two dominant 3p arms (black, dashed arrows), butstrain RI257 produces robust amount of both the 5p (black, solid arrow) and3p (black, dashed arrow) arms, with a 5p dominance. The miRNA sequencesare indicated above the plots, with the seed sequences represented in bold andunderlined.
Chen et al.
11140 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
miRNAs (see Tables S3 and S4 in the supplemental material).Luciferase reporter analysis showed that only the 3= UTR of one ofthese, DUSP8, is regulated by both the 776 and RI257 strainmiRNAs (Fig. 5D). Because all four of these 3= UTRs have anabove-average length (2,081 nucleotides versus an average of 800nucleotides for all annotated human 3= UTRs [71; see Tables S3and S4 in the supplemental material]), it is possible that our targetidentification assay is biased toward false positives for longer 3=UTRs. Nonetheless, our DUSP8 3= reporter results suggest that aminority of host targets can be shared in common between theRI257 and 776 miRNAs.
We considered the possibility that the RI257 and 776 strainmiRNAs have different targets but affect the same pathways. How-ever, functional classification of gene lists from our transfectionand infection studies (DAVID Bioinformatics Resources 6.7; 49,50) did not reveal any significant common functional groups(data not shown). We note that our approach likely underesti-mates the repertoire of possible targets. To minimize false posi-tives, we applied a high-stringency cutoff (40% reduction). Fur-thermore, our approach would miss those targets whose binding isnot dependent on perfect seed complementarity docking in the3=UTR. Therefore, it remains possible that some important sharedhost transcripts or pathways are targeted by both the RI257 and776 strain miRNAs. Nonetheless, our data clearly suggest that
many host transcripts are uniquely targeted by strain 776 orRI257.
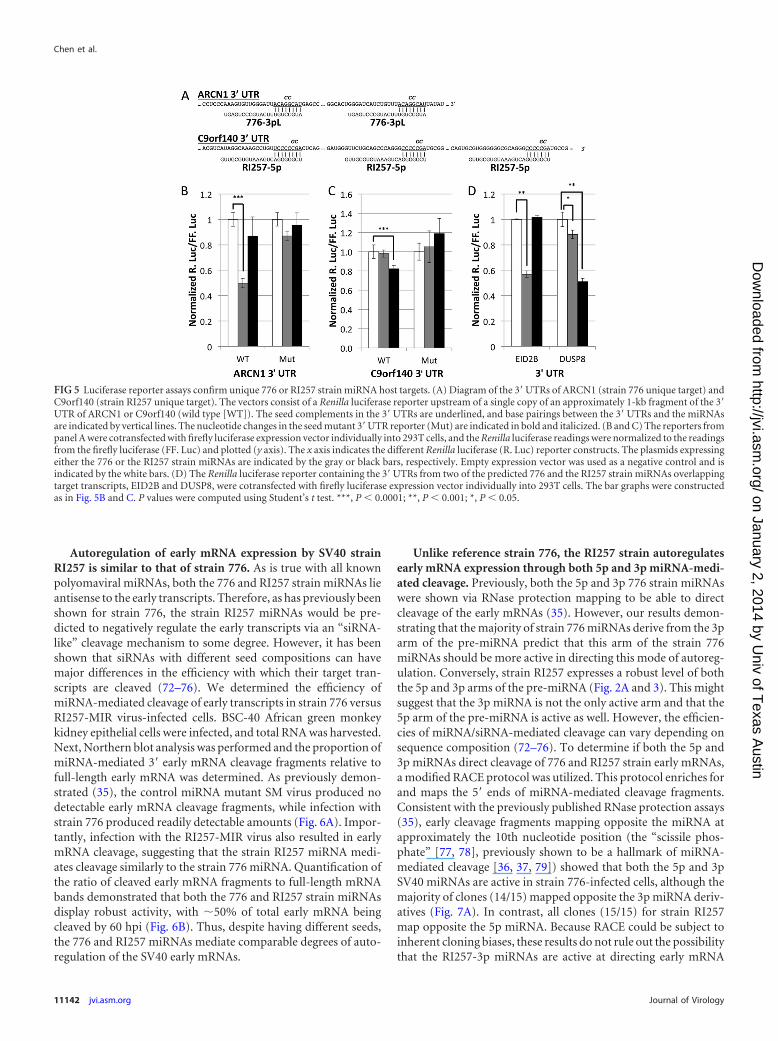
We next determined if some of the putative RI257-specific and776-specific targets are indeed directly and specifically regulatedvia their 3= UTRs by each respective miRNA. We generated 3=UTR reporter constructs for five of the candidate targets identifiedfrom the transfection study (two 776-specific and three RI257-specific constructs). As expected, none of the five reporters wereregulated by both the 776 and RI257 strain miRNAs (see Tables S3and S4 in the supplemental material). One of the strain 776 tar-gets, archain 1 (ARCN1), and all three of the strain RI257 targets(ACTN4, C1orf86, and C9orf140) scored positive for specific neg-ative regulation (Tables S3 and S4 and data not shown). For two ofthe targets, ARCN1 and C9orf140, we also generated negative-control mutant reporters altering two nucleotides in each of theseed complementary regions (Fig. 5A). These mutant 3= UTR re-porters were at least partially refractory to SV40 miRNA regula-tion, thereby demonstrating that this regulation is direct (Fig. 5Band C). Importantly, the ARCN1 3= UTR reporter was only sig-nificantly regulated by the strain 776 miRNA, and conversely, theC9orf140 was only significantly negatively regulated by the strainRI257 miRNA (Fig. 5B and C). These results demonstrate thatclosely related strains of the same virus can have different directmiRNA targets.
FIG 4 Different miRNA target repertoires for SV40 strains 776 and RI257. (A) BSC-40 cells were infected with strain 776 (top panel) or RI257-MIR virus (bottompanel), and total RNA from 44 hpi were reverse transcribed to cDNA and subjected to microarray analysis. All possible combinations of heptameric sequencesare plotted on the x axis, and the number of transcripts containing each corresponding seed complement is plotted on the y axis. The number of transcriptscontaining the 776-3pL (red), 776-3pS (purple), RI257-5p (green), and RI257-3p (blue) seed complements are indicated. Gray bars represent each possibleheptamer (other than 776-3pL and -S and RI257-5p and -3p). The sum of numbers of transcripts downregulated by 40% or more is indicated by “n.” The whitearrowhead marks the most enriched peak for each plot. The median number of transcripts for each heptamer is indicated by the dashed line (for strain 776, 2.29;for strain RI257, 2.498). (B) 293T cells were transfected with strain 776 (top panel) or strain RI257 (bottom panel) miRNA expression vectors, and total RNAfrom 48-h posttransfected cells were subjected to the same microarray analysis as that described above. The median number of transcripts for each heptamer isindicated by the dashed line (for strain 776, 1.70; for strain RI257, 2.77).
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11141
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
Autoregulation of early mRNA expression by SV40 strainRI257 is similar to that of strain 776. As is true with all knownpolyomaviral miRNAs, both the 776 and RI257 strain miRNAs lieantisense to the early transcripts. Therefore, as has previously beenshown for strain 776, the strain RI257 miRNAs would be pre-dicted to negatively regulate the early transcripts via an “siRNA-like” cleavage mechanism to some degree. However, it has beenshown that siRNAs with different seed compositions can havemajor differences in the efficiency with which their target tran-scripts are cleaved (72–76). We determined the efficiency ofmiRNA-mediated cleavage of early transcripts in strain 776 versusRI257-MIR virus-infected cells. BSC-40 African green monkeykidney epithelial cells were infected, and total RNA was harvested.Next, Northern blot analysis was performed and the proportion ofmiRNA-mediated 3= early mRNA cleavage fragments relative tofull-length early mRNA was determined. As previously demon-strated (35), the control miRNA mutant SM virus produced nodetectable early mRNA cleavage fragments, while infection withstrain 776 produced readily detectable amounts (Fig. 6A). Impor-tantly, infection with the RI257-MIR virus also resulted in earlymRNA cleavage, suggesting that the strain RI257 miRNA medi-ates cleavage similarly to the strain 776 miRNA. Quantification ofthe ratio of cleaved early mRNA fragments to full-length mRNAbands demonstrated that both the 776 and RI257 strain miRNAsdisplay robust activity, with �50% of total early mRNA beingcleaved by 60 hpi (Fig. 6B). Thus, despite having different seeds,the 776 and RI257 miRNAs mediate comparable degrees of auto-regulation of the SV40 early mRNAs.
Unlike reference strain 776, the RI257 strain autoregulatesearly mRNA expression through both 5p and 3p miRNA-medi-ated cleavage. Previously, both the 5p and 3p 776 strain miRNAswere shown via RNase protection mapping to be able to directcleavage of the early mRNAs (35). However, our results demon-strating that the majority of strain 776 miRNAs derive from the 3parm of the pre-miRNA predict that this arm of the strain 776miRNAs should be more active in directing this mode of autoreg-ulation. Conversely, strain RI257 expresses a robust level of boththe 5p and 3p arms of the pre-miRNA (Fig. 2A and 3). This mightsuggest that the 3p miRNA is not the only active arm and that the5p arm of the pre-miRNA is active as well. However, the efficien-cies of miRNA/siRNA-mediated cleavage can vary depending onsequence composition (72–76). To determine if both the 5p and3p miRNAs direct cleavage of 776 and RI257 strain early mRNAs,a modified RACE protocol was utilized. This protocol enriches forand maps the 5= ends of miRNA-mediated cleavage fragments.Consistent with the previously published RNase protection assays(35), early cleavage fragments mapping opposite the miRNA atapproximately the 10th nucleotide position (the “scissile phos-phate” [77, 78], previously shown to be a hallmark of miRNA-mediated cleavage [36, 37, 79]) showed that both the 5p and 3pSV40 miRNAs are active in strain 776-infected cells, although themajority of clones (14/15) mapped opposite the 3p miRNA deriv-atives (Fig. 7A). In contrast, all clones (15/15) for strain RI257map opposite the 5p miRNA. Because RACE could be subject toinherent cloning biases, these results do not rule out the possibilitythat the RI257-3p miRNAs are active at directing early mRNA
FIG 5 Luciferase reporter assays confirm unique 776 or RI257 strain miRNA host targets. (A) Diagram of the 3= UTRs of ARCN1 (strain 776 unique target) andC9orf140 (strain RI257 unique target). The vectors consist of a Renilla luciferase reporter upstream of a single copy of an approximately 1-kb fragment of the 3=UTR of ARCN1 or C9orf140 (wild type [WT]). The seed complements in the 3= UTRs are underlined, and base pairings between the 3= UTRs and the miRNAsare indicated by vertical lines. The nucleotide changes in the seed mutant 3= UTR reporter (Mut) are indicated in bold and italicized. (B and C) The reporters frompanel A were cotransfected with firefly luciferase expression vector individually into 293T cells, and the Renilla luciferase readings were normalized to the readingsfrom the firefly luciferase (FF. Luc) and plotted (y axis). The x axis indicates the different Renilla luciferase (R. Luc) reporter constructs. The plasmids expressingeither the 776 or the RI257 strain miRNAs are indicated by the gray or black bars, respectively. Empty expression vector was used as a negative control and isindicated by the white bars. (D) The Renilla luciferase reporter containing the 3= UTRs from two of the predicted 776 and the RI257 strain miRNAs overlappingtarget transcripts, EID2B and DUSP8, were cotransfected with firefly luciferase expression vector individually into 293T cells. The bar graphs were constructedas in Fig. 5B and C. P values were computed using Student’s t test. ***, P � 0.0001; **, P � 0.001; *, P � 0.05.
Chen et al.
11142 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
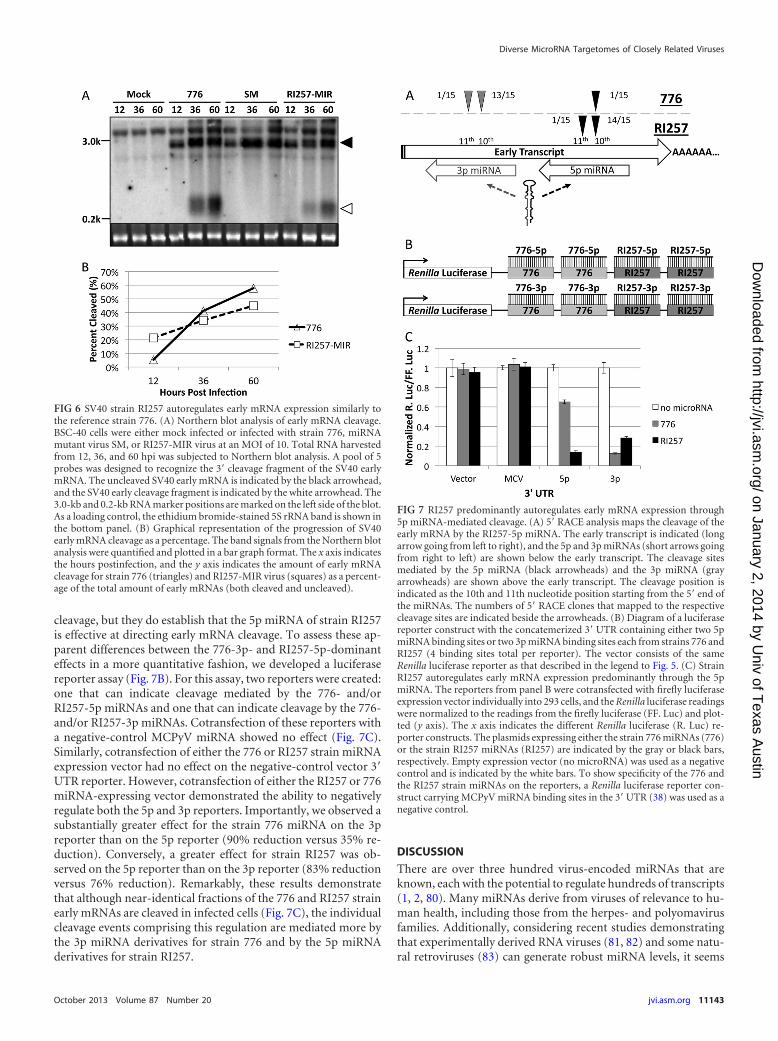
cleavage, but they do establish that the 5p miRNA of strain RI257is effective at directing early mRNA cleavage. To assess these ap-parent differences between the 776-3p- and RI257-5p-dominanteffects in a more quantitative fashion, we developed a luciferasereporter assay (Fig. 7B). For this assay, two reporters were created:one that can indicate cleavage mediated by the 776- and/orRI257-5p miRNAs and one that can indicate cleavage by the 776-and/or RI257-3p miRNAs. Cotransfection of these reporters witha negative-control MCPyV miRNA showed no effect (Fig. 7C).Similarly, cotransfection of either the 776 or RI257 strain miRNAexpression vector had no effect on the negative-control vector 3=UTR reporter. However, cotransfection of either the RI257 or 776miRNA-expressing vector demonstrated the ability to negativelyregulate both the 5p and 3p reporters. Importantly, we observed asubstantially greater effect for the strain 776 miRNA on the 3preporter than on the 5p reporter (90% reduction versus 35% re-duction). Conversely, a greater effect for strain RI257 was ob-served on the 5p reporter than on the 3p reporter (83% reductionversus 76% reduction). Remarkably, these results demonstratethat although near-identical fractions of the 776 and RI257 strainearly mRNAs are cleaved in infected cells (Fig. 7C), the individualcleavage events comprising this regulation are mediated more bythe 3p miRNA derivatives for strain 776 and by the 5p miRNAderivatives for strain RI257.
DISCUSSION
There are over three hundred virus-encoded miRNAs that areknown, each with the potential to regulate hundreds of transcripts(1, 2, 80). Many miRNAs derive from viruses of relevance to hu-man health, including those from the herpes- and polyomavirusfamilies. Additionally, considering recent studies demonstratingthat experimentally derived RNA viruses (81, 82) and some natu-ral retroviruses (83) can generate robust miRNA levels, it seems
FIG 6 SV40 strain RI257 autoregulates early mRNA expression similarly tothe reference strain 776. (A) Northern blot analysis of early mRNA cleavage.BSC-40 cells were either mock infected or infected with strain 776, miRNAmutant virus SM, or RI257-MIR virus at an MOI of 10. Total RNA harvestedfrom 12, 36, and 60 hpi was subjected to Northern blot analysis. A pool of 5probes was designed to recognize the 3= cleavage fragment of the SV40 earlymRNA. The uncleaved SV40 early mRNA is indicated by the black arrowhead,and the SV40 early cleavage fragment is indicated by the white arrowhead. The3.0-kb and 0.2-kb RNA marker positions are marked on the left side of the blot.As a loading control, the ethidium bromide-stained 5S rRNA band is shown inthe bottom panel. (B) Graphical representation of the progression of SV40early mRNA cleavage as a percentage. The band signals from the Northern blotanalysis were quantified and plotted in a bar graph format. The x axis indicatesthe hours postinfection, and the y axis indicates the amount of early mRNAcleavage for strain 776 (triangles) and RI257-MIR virus (squares) as a percent-age of the total amount of early mRNAs (both cleaved and uncleaved).
FIG 7 RI257 predominantly autoregulates early mRNA expression through5p miRNA-mediated cleavage. (A) 5= RACE analysis maps the cleavage of theearly mRNA by the RI257-5p miRNA. The early transcript is indicated (longarrow going from left to right), and the 5p and 3p miRNAs (short arrows goingfrom right to left) are shown below the early transcript. The cleavage sitesmediated by the 5p miRNA (black arrowheads) and the 3p miRNA (grayarrowheads) are shown above the early transcript. The cleavage position isindicated as the 10th and 11th nucleotide position starting from the 5= end ofthe miRNAs. The numbers of 5= RACE clones that mapped to the respectivecleavage sites are indicated beside the arrowheads. (B) Diagram of a luciferasereporter construct with the concatemerized 3= UTR containing either two 5pmiRNA binding sites or two 3p miRNA binding sites each from strains 776 andRI257 (4 binding sites total per reporter). The vector consists of the sameRenilla luciferase reporter as that described in the legend to Fig. 5. (C) StrainRI257 autoregulates early mRNA expression predominantly through the 5pmiRNA. The reporters from panel B were cotransfected with firefly luciferaseexpression vector individually into 293 cells, and the Renilla luciferase readingswere normalized to the readings from the firefly luciferase (FF. Luc) and plot-ted (y axis). The x axis indicates the different Renilla luciferase (R. Luc) re-porter constructs. The plasmids expressing either the strain 776 miRNAs (776)or the strain RI257 miRNAs (RI257) are indicated by the gray or black bars,respectively. Empty expression vector (no microRNA) was used as a negativecontrol and is indicated by the white bars. To show specificity of the 776 andthe RI257 strain miRNAs on the reporters, a Renilla luciferase reporter con-struct carrying MCPyV miRNA binding sites in the 3= UTR (38) was used as anegative control.
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11143
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
likely that numerous additional viral miRNAs await discovery.Despite much progress in identifying new viral miRNAs, few havewell-understood functions. A major advance in understandingmiRNA function occurred with the observation that some miRNAtargets display subtle decreases in steady-state levels upon miRNAbinding (25, 27, 84), thereby making target identification amena-ble to high-throughput detection methods. In addition, recenthigh-throughput positive enrichment strategies either with (6, 8,10, 12) or without (9) cross-linking have shown much success inidentifying viral miRNA target transcripts associated with proteincomponents of the silencing machinery. However, it remains un-clear what fraction of these targets is important during the infec-tious cycle.
Here, we present data that further emphasize the importanceof viral miRNA-mediated autoregulation of polyomavirus earlytranscripts. Our approach is to identify closely related strains ofviruses that possess different seed repertoires. Since the seed re-gion plays a major role in dictating target interactions, variantswith different seed repertoires would be expected to have differentmiRNA targets. This approach assumes that targets (host or viral)preserved throughout evolution will be important, while the less-relevant, or possibly “niche-specific,” targets will be unique toeach strain. Using this approach for SV40, we identify a class ofcirculating variants, typified by the strain RI257 miRNAs, whichdiffer substantially from other classes of SV40 miRNAs. Unlikestrain 776, in which the vast majority of miRNAs are derived fromthe 3p arm of the pre-miRNA, strain RI257 gives rise to abundantmiRNAs detectable from both arms. In addition, both the 5p and3p RI257 strain miRNA derivatives possess different seed se-quences than those of strain 776. We show that, as would be pre-dicted by the different seeds, 776 and RI257 miRNAs have sometargets unique to each respective virus. While this clearly demon-strates that some miRNA targets are unique to each strain, it re-mains to be addressed if these unique targets play any role in thepolyomavirus life cycle.
As strain RI257 has been isolated independently in differentgeographical regions (85), we conclude that strain RI257-like vi-ruses are circulating in the wild. Thus, the different repertoires ofhost targets for 776 and RI257 strain miRNAs demonstrate that
wild circulating strains of SV40 can tolerate different “targe-tomes” (Fig. 4). These data suggest that some viral miRNA targetsmay be of little selective advantage to the virus. This is consistentwith the notion that some viral miRNA targets could be neutral oreven disadvantageous to the virus, as long as the sum total ofregulation imparted by the targets is of sufficient benefit (1). Here,we present data only for some targets of SV40 in cultured cells, andit remains to be seen if this applies in vivo or to viruses other thanmembers of the Polyomaviridae. However, in light of our findings,a note of caution is warranted as to the meaningfulness of viralmiRNA targets identified via high-throughput studies in the ab-sence of secondary criteria.
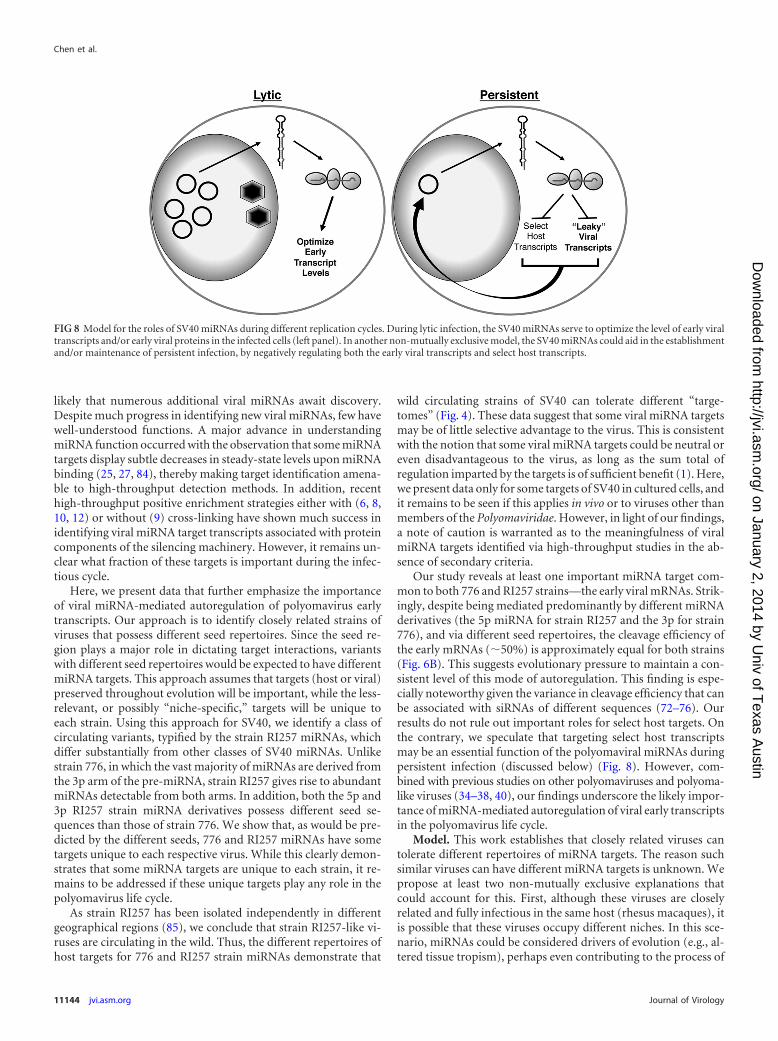
Our study reveals at least one important miRNA target com-mon to both 776 and RI257 strains—the early viral mRNAs. Strik-ingly, despite being mediated predominantly by different miRNAderivatives (the 5p miRNA for strain RI257 and the 3p for strain776), and via different seed repertoires, the cleavage efficiency ofthe early mRNAs (�50%) is approximately equal for both strains(Fig. 6B). This suggests evolutionary pressure to maintain a con-sistent level of this mode of autoregulation. This finding is espe-cially noteworthy given the variance in cleavage efficiency that canbe associated with siRNAs of different sequences (72–76). Ourresults do not rule out important roles for select host targets. Onthe contrary, we speculate that targeting select host transcriptsmay be an essential function of the polyomaviral miRNAs duringpersistent infection (discussed below) (Fig. 8). However, com-bined with previous studies on other polyomaviruses and polyoma-like viruses (34–38, 40), our findings underscore the likely impor-tance of miRNA-mediated autoregulation of viral early transcriptsin the polyomavirus life cycle.
Model. This work establishes that closely related viruses cantolerate different repertoires of miRNA targets. The reason suchsimilar viruses can have different miRNA targets is unknown. Wepropose at least two non-mutually exclusive explanations thatcould account for this. First, although these viruses are closelyrelated and fully infectious in the same host (rhesus macaques), itis possible that these viruses occupy different niches. In this sce-nario, miRNAs could be considered drivers of evolution (e.g., al-tered tissue tropism), perhaps even contributing to the process of
FIG 8 Model for the roles of SV40 miRNAs during different replication cycles. During lytic infection, the SV40 miRNAs serve to optimize the level of early viraltranscripts and/or early viral proteins in the infected cells (left panel). In another non-mutually exclusive model, the SV40 miRNAs could aid in the establishmentand/or maintenance of persistent infection, by negatively regulating both the early viral transcripts and select host transcripts.
Chen et al.
11144 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

viral speciation (1). Second, any particular individual miRNA tar-get could be of no selective advantage to the virus, as long as thesum total of regulation imparted by the miRNA on other tran-scripts is advantageous (1). Both of these models may be relevantto other virus families in addition to the Polyomaviridae.
Combined with previous published studies (34–38, 40), thefindings from this study are consistent with the following modelfor polyomaviruses. First, the striking consistency between strainRI257 and 776 in the fraction of early transcripts that are subject tomiRNA-mediated cleavage implies that maintaining an optimaldegree of this regulation provides a selective advantage. Sincethese results were obtained during lytic infection, we propose thatone function of the SV40 miRNA could be to optimize the abun-dance of early mRNAs and/or early proteins during lytic infection(or, at minimum, to avoid excessive cleavage of the early tran-scripts). Second, a non-mutually exclusive model is for a role ofthe miRNA during persistent infection (1, 2, 86). Although themechanisms are poorly defined, SV40 and other polyomavirusesestablish long-term persistent infections in vivo. Clearly, nega-tively regulating early gene transcripts could be a way to enforcepersistence, akin to the role that has been proposed for some her-pesviral miRNAs (1, 87, 88). Indeed, a recent report fromBrokema and Imperiale on the BK human polyomavirus is con-sistent with this notion (86). Furthermore, considering the robustchanges in host gene expression associated with lytic infection (thepresent work and references 76 to 80) compared to the generallymore subtle degree of regulation imparted by miRNAs, it seemslikely that viral miRNA regulation of select host targets may bemost relevant during persistent infection. In this model, viralmiRNA targeting of the early transcripts and select host tran-scripts plays a role in promoting and/or reinforcing persistentinfection. This could occur analogous to some herpesviralmiRNAs, in which host targets are associated with increased cellviability, immune evasion, and the indirect negative regulation ofviral lytic genes (1, 87–94). Clearly, such models await future test-ing in relevant persistent models of polyomavirus infection.
In conclusion, a surprising degree of plasticity in miRNA tar-gets can be tolerated by closely related viruses, yet at the same time,the capacity to regulate common viral transcripts is maintained.This work not only advances our understanding of the polyoma-viruses but also may be applicable to other type of viruses thatutilize miRNAs in their infectious cycles.
ACKNOWLEDGMENTS
We thank the members of the Sullivan laboratory for useful discussionsand comments regarding the manuscript, Scott Hunicke-Smith, the Uni-versity of Texas at Austin Genomic Sequencing and Analysis Facility(GSAF), for useful input on next-generation sequencing, and MariannaGrenadier for the illustrations. We also thank Marvin Whiteley and Ste-phen Trent for use of equipment.
This work was supported by grant RO1AI077746 from the NationalInstitutes of Health and an Investigators in Pathogenesis of InfectiousDisease Award from Burroughs Wellcome to C.S.S. and a Project SEEDscholarship to A.M. from the American Chemical Society.
C.J.C., J.E.C., and C.S.S. conceived the project, C.J.C., J.E.C., and A.M.performed the experiments, C.J.C., J.E.C., R.P.K., and C.S.S. analyzed thedata, and C.J.C., J.E.C., and C.S.S. wrote the manuscript.
REFERENCES1. Kincaid RP, Sullivan CS. 2012. Virus-encoded microRNAs: an overview
and a look to the future. PLoS Pathog. 8:e1003018. doi:10.1371/journal.ppat.1003018.
2. Grundhoff A, Sullivan CS. 2011. Virus-encoded microRNAs. Virology411:325–343.
3. Gottwein E, Cullen BR. 2008. Viral and cellular microRNAs as determi-nants of viral pathogenesis and immunity. Cell Host Microbe 3:375–387.
4. Boss IW, Plaisance KB, Renne R. 2009. Role of virus-encoded microRNAs inherpesvirus biology. Trends Microbiol. 17:544–553.
5. Tuddenham L, Pfeffer S. 2011. Roles and regulation of microRNAs incytomegalovirus infection. Biochim. Biophys. Acta 1809:613– 622.
6. Gottwein E, Corcoran DL, Mukherjee N, Skalsky RL, Hafner M, Nus-baum JD, Shamulailatpam P, Love CL, Dave SS, Tuschl T, Ohler U,Cullen BR. 2011. Viral microRNA targetome of KSHV-infected primaryeffusion lymphoma cell lines. Cell Host Microbe 10:515–526.
7. Grey F, Tirabassi R, Meyers H, Wu G, McWeeney S, Hook L, NelsonJA. 2010. A viral microRNA down-regulates multiple cell cycle genesthrough mRNA 5=UTRs. PLoS Pathog. 6:e1000967. doi:10.1371/journal.ppat.1000967.
8. Skalsky RL, Corcoran DL, Gottwein E, Frank CL, Kang D, Hafner M,Nusbaum JD, Feederle R, Delecluse H-J, Luftig MA, Tuschl T, Ohler U,Cullen BR. 2012. The viral and cellular microRNA targetome in lympho-blastoid cell lines. PLoS Pathog. 8:e1002484. doi:10.1371/journal.ppat.1002484.
9. Dölken L, Malterer G, Erhard F, Kothe S, Friedel CC, Suffert G,Marcinowski L, Motsch N, Barth S, Beitzinger M. 2010. Systematicanalysis of viral and cellular microRNA targets in cells latently infectedwith human �-herpesviruses by RISC immunoprecipitation assay. CellHost Microbe 7:324 –334.
10. Riley KJ, Rabinowitz GS, Yario TA, Luna JM, Darnell RB, Steitz JA.2012. EBV and human microRNAs co-target oncogenic and apoptoticviral and human genes during latency. EMBO J. 31:2207–2221.
11. Plaisance-Bonstaff K, Renne R. 2011. Viral miRNAs, p 43– 66. In van RijRP (ed), Antiviral RNAi. Humana Press, Totowa, NJ.
12. Haecker I, Gay LA, Yang Y, Hu J, Morse AM, McIntyre LM, Renne R.2012. Ago HITS-CLIP expands understanding of Kaposi’s sarcoma-associated herpesvirus miRNA function in primary effusion lymphomas.PLoS Pathog. 8:e1002884. doi:10.1371/journal.ppat.1002884.
13. Ziegelbauer JM, Sullivan CS, Ganem D. 2009. Tandem array-based expres-sion screens identify host mRNA targets of virus-encoded microRNAs. Nat.Genet. 41:130–134.
14. Bartel DP. 2009. MicroRNAs: target recognition and regulatory func-tions. Cell 136:215–233.
15. Carthew RW, Sontheimer EJ. 2009. Origins and mechanisms of miRNAsand siRNAs. Cell 136:642– 655.
16. Kim VN, Han J, Siomi MC. 2009. Biogenesis of small RNAs in animals.Nat. Rev. Mol. Cell Biol. 10:126 –139.
17. Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, Lee J, Provost P, RådmarkO, Kim S. 2003. The nuclear RNase III Drosha initiates microRNA pro-cessing. Nature 425:415– 419.
18. Zeng Y, Yi R, Cullen BR. 2005. Recognition and cleavage of primarymicroRNA precursors by the nuclear processing enzyme Drosha. EMBO J.24:138 –148.
19. Zeng Y, Cullen BR. 2005. Efficient processing of primary microRNAhairpins by Drosha requires flanking nonstructured RNA sequences. J.Biol. Chem. 280:27595–27603.
20. Yi R, Qin Y, Macara IG, Cullen BR. 2003. Exportin-5 mediates thenuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev.17:3011–3016.
21. Lund E, Güttinger S, Calado A, Dahlberg JE, Kutay U. 2004. Nuclearexport of microRNA precursors. Science 303:95–98.
22. Bohnsack MT. 2004. Exportin 5 is a RanGTP-dependent dsRNA-bindingprotein that mediates nuclear export of pre-miRNAs. RNA 10:185–191.
23. Bernstein E, Caudy AA, Hammond SM, Hannon GJ. 2001. Role for abidentate ribonuclease in the initiation step of RNA interference. Nature409:363–366.
24. Hutvágner G, McLachlan J, Pasquinelli AE, Balint E, Tuschl T, ZamorePD. 2001. A cellular function for the RNA-interference enzyme Dicer inthe maturation of the let-7 small temporal RNA. Science 293:834 – 838.
25. Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J,Bartel DP, Linsley PS, Johnson JM. 2005. Microarray analysis shows thatsome microRNAs downregulate large numbers of target mRNAs. Nature433:769 –773.
26. Bazzini AA, Lee MT, Giraldez AJ. 2012. Ribosome profiling shows thatmiR-430 reduces translation before causing mRNA decay in zebrafish.Science 336:233–237.
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11145
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

27. Samols MA, Skalsky RL, Maldonado AM, Riva A, Lopez MC, Baker HV,Renne R. 2007. Identification of cellular genes targeted by KSHV-encodedmicroRNAs. PLoS Pathog. 3:e65. doi:10.1371/journal.ppat.0030065.
28. Spurgeon ME, Lambert PF. 2013. Merkel cell polyomavirus: a newlydiscovered human virus with oncogenic potential. Virology 435:118 –130.
29. Dalianis T, Hirsch HH. 2013. Human polyomaviruses in disease andcancer. Virology 437:63–72.
30. Feltkamp MCW, Kazem S, van der Meijden E, Lauber C, GorbalenyaAE. 2013. From Stockholm to Malawi: recent developments in studyinghuman polyomaviruses. J. Gen. Virol. 94:482– 496.
31. DeCaprio JA, Garcea RL. 2013. A cornucopia of human polyomaviruses.Nat. Rev. Microbiol. 11:264 –276.
32. Kazem S, van der Meijden E, Feltkamp MCW. 2013. The trichodysplasiaspinulosa-associated polyomavirus; virological background and clinicalimplications. APMIS 121:770 –782.
33. Cole CN. 1996. Polyomaviridae: the viruses and their replication, p 1997–2043. In Fields BN, Knipe DM, Howley PM, Chanock RM, Monath TP,Melnick JL, Roizman B, Straus SE (ed), Fields virology, 3rd ed. Lippincott-Raven Press, Philadelphia, PA.
34. Cantalupo P, Doering A, Sullivan CS, Pal A, Peden KWC, Lewis AM,Pipas JM. 2005. Complete nucleotide sequence of polyomavirus SA12. J.Virol. 79:13094 –13104.
35. Sullivan CS, Grundhoff AT, Tevethia S, Pipas JM, Ganem D. 2005.SV40-encoded microRNAs regulate viral gene expression and reduce sus-ceptibility to cytotoxic T cells. Nature 435:682– 686.
36. Seo GJ, Fink LHL, O’Hara B, Atwood WJ, Sullivan CS. 2008. Evolu-tionarily conserved function of a viral microRNA. J. Virol. 82:9823–9828.
37. Sullivan CS, Sung CK, Pack CD, Grundhoff A, Lukacher AE, BenjaminTL, Ganem D. 2009. Murine polyomavirus encodes a microRNA thatcleaves early RNA transcripts but is not essential for experimental infec-tion. Virology 387:157–167.
38. Seo GJ, Chen CJ, Sullivan CS. 2009. Merkel cell polyomavirus encodes amicroRNA with the ability to autoregulate viral gene expression. Virology383:183–187.
39. Bauman Y, Nachmani D, Vitenshtein A, Tsukerman P, Drayman N,Stern-Ginossar N, Lankry D, Gruda R, Mandelboim O. 2011. An iden-tical miRNA of the human JC and BK polyoma viruses targets the stress-induced ligand ULBP3 to escape immune elimination. Cell Host Microbe9:93–102.
40. Chen CJ, Kincaid RP, Seo GJ, Bennett MD, Sullivan CS. 2011. Insightsinto Polyomaviridae microRNA function derived from study of the bandi-coot papillomatosis carcinomatosis viruses. J. Virol. 85:4487– 4500.
41. Lin Y-T, Kincaid RP, Arasappan D, Dowd SE, Hunicke-Smith SP,Sullivan CS. 2010. Small RNA profiling reveals antisense transcriptionthroughout the KSHV genome and novel small RNAs. RNA 16:1540 –1558.
42. Weber K, Bolander ME, Sarkar G. 1998. PIG-B: a homemade monopha-sic cocktail for the extraction of RNA. Mol. Biotechnol. 9:73–77.
43. Koscianska E, Starega-Roslan J, Czubala K, Krzyzosiak WJ. 2011. High-resolution Northern blot for a reliable analysis of microRNAs and theirprecursors. ScientificWorldJournal 11:102–117.
44. Grundhoff A. 2006. A combined computational and microarray-basedapproach identifies novel microRNAs encoded by human gamma-herpesviruses. RNA 12:733–750.
45. Kraus RJ, Mertz JE. 2001. Quantitation and structural analysis of SV40RNAs, p 87–101. In Raptis L (ed), SV40 protocols. Humana Press, NewYork, NY.
46. Tremblay JD, Sachsenmeier KF, Pipas JM. 2001. Propagation of wild-type and mutant SV40, p 1–7. In Raptis L (ed), SV40 protocols. HumanaPress, New York, NY.
47. Drayman N, Kler S, Ben-nun-Shaul O, Oppenheim A. 2010. Rapidmethod for SV40 titration. J. Virol. Methods 164:145–147.
48. Burgos KL, Javaherian A, Bomprezzi R, Ghaffari L, Rhodes S,Courtright A, Tembe W, Kim S, Metpally R, Keuren-Jensen KV. 2013.Identification of extracellular miRNA in human cerebrospinal fluid bynext-generation sequencing. RNA 19:712–722.
49. Huang DW, Sherman BT, Lempicki RA. 2009. Systematic and integra-tive analysis of large gene lists using DAVID bioinformatics resources.Nat. Protoc. 4:44 –57.
50. Huang DW, Sherman BT, Lempicki RA. 2009. Bioinformatics enrich-ment tools: paths toward the comprehensive functional analysis of largegene lists. Nucleic Acids Res. 37:1–13.
51. Fedorov Y. 2006. Off-target effects by siRNA can induce toxic phenotype.RNA 12:1188 –1196.
52. Jackson AL. 2006. Widespread siRNA “off-target” transcript silencingmediated by seed region sequence complementarity. RNA 12:1179 –1187.
53. Grimson A, Farh KK-H, Johnston WK, Garrett-Engele P, Lim LP,Bartel DP. 2007. MicroRNA targeting specificity in mammals: determi-nants beyond seed pairing. Mol. Cell 27:91–105.
54. van Dongen S, Abreu-Goodger C, Enright AJ. 2008. DetectingmicroRNA binding and siRNA off-target effects from expression data.Nat. Methods 5:1023–1025.
55. Rizzo P, Di Resta I, Powers A, Ratner H, Carbone M. 1999. Uniquestrains of SV40 in commercial poliovaccines from 1955 not readily iden-tifiable with current testing for SV40 infection. Cancer Res. 59:6103– 6108.
56. Lednicky JA, Arrington AS, Stewart AR, Dai XM, Wong C, Jafar S,Murphey-Corb M, Butel JS. 1998. Natural isolates of simian virus 40from immunocompromised monkeys display extensive genetic heteroge-neity: new implications for polyomavirus disease. J. Virol. 72:3980 –3990.
57. Forsman ZH, Lednicky JA, Fox GE, Willson RC, White ZS, HalvorsonSJ, Wong C, Lewis AM, Butel JS. 2004. Phylogenetic analysis of polyo-mavirus simian virus 40 from monkeys and humans reveals genetic vari-ation. J. Virol. 78:9306 –9316.
58. Hafner M, Renwick N, Brown M, Mihailovic A, Holoch D, Lin C, PenaJTG, Nusbaum JD, Morozov P, Ludwig J, Ojo T, Luo S, Schroth G,Tuschl T. 2011. RNA-ligase-dependent biases in miRNA representationin deep-sequenced small RNA cDNA libraries. RNA 17:1697–1712.
59. Jayaprakash AD, Jabado O, Brown BD, Sachidanandam R. 2011. Iden-tification and remediation of biases in the activity of RNA ligases in small-RNA deep sequencing. Nucleic Acids Res. 39:e141– e141.
60. Linsen SE, de Wit E, Janssens G, Heater S, Chapman L, Parkin RK, FritzB, Wyman SK, de Bruijn E, Voest EE, Kuersten S, Tewari M, CuppenE. 2009. Limitations and possibilities of small RNA digital gene expressionprofiling. Nat. Methods 6:474 – 476.
61. Sorefan K, Pais H, Hall AE, Kozomara A, Griffiths-Jones S, Moulton V,Dalmay T. 2012. Reducing ligation bias of small RNAs in libraries for nextgeneration sequencing. Silence 3:4. doi:10.1186/1758-907X-3-4.
62. Sun G, Wu X, Wang J, Li H, Li X, Gao H, Rossi J, Yen Y. 2011. Abias-reducing strategy in profiling small RNAs using Solexa. RNA 17:2256 –2262.
63. Willenbrock H, Salomon J, Sokilde R, Barken KB, Hansen TN, NielsenFC, Moller S, Litman T. 2009. Quantitative miRNA expression analysis:comparing microarrays with next-generation sequencing. RNA 15:2028 –2034.
64. Lewis BP, Burge CB, Bartel DP. 2005. Conserved seed pairing, oftenflanked by adenosines, indicates that thousands of human genes aremicroRNA targets. Cell 120:15–20.
65. Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. 2008. Theimpact of microRNAs on protein output. Nature 455:64 –71.
66. Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, RajewskyN. 2008. Widespread changes in protein synthesis induced by microRNAs.Nature 455:58–63.
67. Hafner M, Landthaler M, Burger L, Khorshid M, Hausser J, BerningerP, Rothballer A, Ascano M, Jungkamp A-C, Munschauer M, Ulrich A,Wardle GS, Dewell S, Zavolan M, Tuschl T. 2010. Transcriptome-wideidentification of RNA-binding protein and microRNA target sites byPAR-CLIP. Cell 141:129 –141.
68. Zisoulis DG, Lovci MT, Wilbert ML, Hutt KR, Liang TY, PasquinelliAE, Yeo GW. 2010. Comprehensive discovery of endogenous Argonautebinding sites in Caenorhabditis elegans. Nat. Struct. Mol. Biol. 17:173–179.
69. Spindel E, Pauley M, Jia Y, Gravett C, Thompson S, Boyle N, Ojeda S,Norgren R. 2005. Leveraging human genomic information to identifynonhuman primate sequences for expression array development. BMCGenomics 6:160.
70. Flicek P, Ahmed I, Amode MR, Barrell D, Beal K, Brent S, Carvalho-Silva D, Clapham P, Coates G, Fairley S, Fitzgerald S, Gil L, Garcia-Giron C, Gordon L, Hourlier T, Hunt S, Juettemann T, Kahari AK,Keenan S, Komorowska M, Kulesha E, Longden I, Maurel T, McLarenWM, Muffato M, Nag R, Overduin B, Pignatelli M, Pritchard B,Pritchard E, Riat HS, Ritchie GRS, Ruffier M, Schuster M, Sheppard D,Sobral D, Taylor K, Thormann A, Trevanion S, White S, Wilder SP,Aken BL, Birney E, Cunningham F, Dunham I, Harrow J, Herrero J,Hubbard TJP, Johnson N, Kinsella R, Parker A, Spudich G, Yates A,
Chen et al.
11146 jvi.asm.org Journal of Virology
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from

Zadissa A, Searle SMJ. 2013. Ensembl 2013. Nucleic Acids Res. 41:D48 –D55.
71. Mignone F, Gissi C, Liuni S, Pesole G. 2002. Untranslated regions ofmRNAs. Genome Biol. 3:reviews0004.1–reviews0004.10. doi:10.1186/gb-2002-3-3-reviews0004.
72. Harborth J, Elbashir SM, Vandenburgh K, Manninga H, Scaringe SA,Weber K, Tuschl T. 2003. Sequence, chemical, and structural variation ofsmall interfering RNAs and short hairpin RNAs and the effect on mam-malian gene silencing. Antisense Nucleic Acid Drug Dev. 13:83–105.
73. Reynolds A, Leake D, Boese Q, Scaringe S, Marshall WS, Khvorova A.2004. Rational siRNA design for RNA interference. Nat. Biotechnol. 22:326 –330.
74. Schwarz DS, Ding H, Kennington L, Moore JT, Schelter J, Burchard J,Linsley PS, Aronin N, Xu Z, Zamore PD. 2006. Designing siRNA thatdistinguish between genes that differ by a single nucleotide. PLoS Genet.2:e140. doi:10.1371/journal.pgen.0020140.
75. Holen T, Amarzguioui M, Wiiger MT, Babaie E, Prydz H. 2002.Positional effects of short interfering RNAs targeting the human coagula-tion trigger tissue factor. Nucleic Acids Res. 30:1757–1766.
76. Haley B, Zamore PD. 2004. Kinetic analysis of the RNAi enzyme com-plex. Nat. Struct. Mol. Biol. 11:599 – 606.
77. Yekta S, Shih I, Bartel DP. 2004. MicroRNA-directed cleavage of HOXB8mRNA. Science 304:594 –596.
78. Hornstein E, Mansfield JH, Yekta S, Hu JK-H, Harfe BD, McManusMT, Baskerville S, Bartel DP, Tabin CJ. 2005. The microRNA miR-196acts upstream of Hoxb8 and Shh in limb development. Nature 438:671–674.
79. Barth S, Pfuhl T, Mamiani A, Ehses C, Roemer K, Kremmer E, Jaker C,Hock J, Meister G, Grasser FA. 2008. Epstein-Barr virus-encodedmicroRNA miR-BART2 down-regulates the viral DNA polymeraseBALF5. Nucleic Acids Res. 36:666 – 675.
80. Cullen BR. 2011. Viruses and microRNAs: RISCy interactions with seri-ous consequences. Genes Dev. 25:1881–1894.
81. Varble A, Chua MA, Perez JT, Manicassamy B, Garcia-Sastre A, tenOever BR. 2010. Engineered RNA viral synthesis of microRNAs. Proc.Natl. Acad. Sci. U. S. A. 107:11519 –11524.
82. Langlois RA, Shapiro JS, Pham AM, ten Oever BR. 2012. In vivo deliveryof cytoplasmic RNA virus-derived miRNAs. Mol. Ther. 20:367–375.
83. Kincaid RP, Burke JM, Sullivan CS. 2012. From the cover: RNA virusmicroRNA that mimics a B-cell oncomiR. Proc. Natl. Acad. Sci. U. S. A.109:3077–3082.
84. Gottwein E, Mukherjee N, Sachse C, Frenzel C, Majoros WH, Chi J-TA,Braich R, Manoharan M, Soutschek J, Ohler U, Cullen BR. 2007. A viral
microRNA functions as an orthologue of cellular miR-155. Nature 450:1096 –1099.
85. Fagrouch Z, Karremans K, Deuzing I, van Gessel S, Niphuis H, BogersW, Verschoor EJ. 2011. Molecular analysis of a novel simian virus 40(SV40) type in rhesus macaques and evidence for double infections withthe classical SV40 type. J. Clin. Microbiol. 49:1280 –1286.
86. Broekema NM, Imperiale MJ. 2013. miRNA regulation of BK polyoma-virus replication during early infection. Proc. Natl. Acad. Sci. U. S. A.110:8200 – 8205.
87. Murphy E, Vanícek J, Robins H, Shenk T, Levine AJ. 2008. Suppres-sion of immediate-early viral gene expression by herpesvirus-codedmicroRNAs: implications for latency. Proc. Natl. Acad. Sci. U. S. A.105:5453–5458.
88. Dölken L, Krmpotic A, Kothe S, Tuddenham L, Tanguy M, Marci-nowski L, Ruzsics Z, Elefant N, Altuvia Y, Margalit H, Koszinowski UH,Jonjic S, Pfeffer S. 2010. Cytomegalovirus microRNAs facilitate persistentvirus infection in salivary glands. PLoS Pathog. 6:e1001150. doi:10.1371/journal.ppat.1001150.
89. Poole LJ, Yu Y, Kim PS, Zheng Q-Z, Pevsner J, Hayward GS. 2002.Altered patterns of cellular gene expression in dermal microvascular en-dothelial cells infected with Kaposi’s sarcoma-associated herpesvirus. J.Virol. 76:3395–3420.
90. Ebrahimi B, Dutia BM, Roberts KL, Garcia-Ramirez JJ, Dickinson P,Stewart JP, Ghazal P, Roy DJ, Nash AA. 2003. Transcriptome profile ofmurine gammaherpesvirus-68 lytic infection. J. Gen. Virol. 84:99 –109.
91. Chandriani S, Ganem D. 2007. Host transcript accumulation during lyticKSHV infection reveals several classes of host responses. PLoS One 2:e811.doi:10.1371/journal.pone.0000811.
92. Marquitz AR, Mathur A, Shair KHY, Raab-Traub N. 2012. Infection ofEpstein-Barr virus in a gastric carcinoma cell line induces anchorage in-dependence and global changes in gene expression. Proc. Natl. Acad. Sci.U. S. A. 109:9593–9598.
93. Marcinowski L, Lidschreiber M, Windhager L, Rieder M, Bosse JB,Rädle B, Bonfert T, Györy I, de Graaf M, da Costa OP, Rosenstiel P,Friedel CC, Zimmer R, Ruzsics Z, Dölken L. 2012. Real-time transcrip-tional profiling of cellular and viral gene expression during lytic cytomeg-alovirus infection. PLoS Pathog. 8:e1002908. doi:10.1371/journal.ppat.1002908.
94. Lei X, Bai Z, Ye F, Xie J, Kim C-G, Huang Y, GaoS-J. 2010. Regulationof NF-�B inhibitor I�B and viral replication by a KSHV microRNA. Nat.Cell Biol. 12:193–199.
95. Sambrook J, Russell DW. 2001. Molecular cloning: a laboratory manual,3rd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Diverse MicroRNA Targetomes of Closely Related Viruses
October 2013 Volume 87 Number 20 jvi.asm.org 11147
on January 2, 2014 by Univ of T
exas Austin
http://jvi.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















