
COLOR DIFFERENCES AMONG CLOSELY RELATED SPECIES OF RED-BREASTED MEADOWLARKS (STURNELLA
Upload
independentCategory
view
4download
0
Journal of Invertebrate Pathology 88 (2005) 100–107
www.elsevier.com/locate/yjipa
European Leucoma salicis NPV is closely related to North American Orgyia pseudotsugata MNPV
Agata Jakubowska a,b, Monique M. van Oers a, Jenny S. Cory a, Jadwiga Ziemnicka b, Just M. Vlak a,¤
a Laboratory of Virology, Wageningen University, Binnenhaven 11, Wageningen 6709 PD, The Netherlandsb Department of Biological Control and Quarantine, Institute of Plant Protection, Miczurina 20, Poznan 60-318, Poland
Received 12 October 2004; accepted 24 December 2004
Abstract
The satin moth Leucoma salicis L. (Lepidoptera, Lymantriidae) is a frequent defoliator of poplar trees (Populus spp.) in Europeand Asia (China, Japan). Around 1920 the insect was introduced into the USA and Canada. In this paper, a multicapsid nucleopoly-hedrovirus isolated from L. salicis larvae in Poland (LesaNPV) was characterized and appeared to be a variant of Orgyia pseudotsug-ata (Op) MNPV. O. pseudotsugata, the Douglas Wr tussock moth (Lepidoptera, Lymantriidae), occurs exclusively in North America.Sequences of three conserved baculovirus genes, polyhedrin, lef-8, and pif-2, were ampliWed in polymerase chain reactions usingdegenerate primer sets, and revealed a high degree of homology to OpMNPV. Restriction enzyme analysis conWrmed the closerelationship between LesaNPV and OpMNPV, although a number of restriction fragment length polymorphisms were observed.The lef-7 gene, encoding late expression factor 7, and the ctl-2 gene, encoding a conotoxin-like protein, were chosen as putativemolecular determinants of the respective viruses. The ctl-2 region appeared suitable for unequivocal identiWcation of either virus asLesaNPV lacked a dUTPase gene in this region. Our observations may suggest that LesaNPV, along with L. salicis, was introducedinto O. pseudotsugata after introduction of the former insect into North America in the 1920s. 2005 Elsevier Inc. All rights reserved.
Keywords: Leucoma salicis; Orgyia pseudotsugata; Lymantriidae; Nucleopolyhedrovirus; LesaNPV; OpMNPV; Taxonomy
1. Introduction
The satin moth Leucoma salicis L. (Lepidoptera,Lymantriidae), previously known as Stilpnotia salicis, isa serious defoliator occuring throughout Europe andAsia (Lipa and Ziemnicka, 1996). In the 1920s, the insectwas introduced into North America, where it was Wrstdetected near Boston, Massachusetts. Currently, it is dis-tributed over New England in the northern UnitedStates and British Columbia in Canada (Langor, 1995).Satin moth larvae feed on all species of poplar and
¤ Corresponding author. Fax: +31 317 484820.E-mail address: [email protected] (J.M. Vlak).
0022-2011/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.jip.2004.12.002
willow (Populus spp.), but also on oak and crabapple. Onboth continents they are most common on eastern cot-tonwood, white and black poplar, bigtooth, and trem-bling aspen, in both planted trees and natural stands.Usually there is only one generation of the insect peryear, although up to three generations may occur in thewarmer climate zones. Larvae diapause in the L2 stage,but hibernation as eggs has also been reported (Ziemni-cka, 2000). In Europe, the Wrst signs of tree damageappear in late May when larvae resume feeding. Aftermid-June the late instar larvae are capable of massive,complete defoliation of trees. Severe feeding damageresults in reduced growth of stems and Wnally tree mor-tality (Langor, 1995).

A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107 101
Leucoma salicis has been shown to be susceptible to anumber of entomopathogens, including viruses, bacteria,spiroplasma, fungi, and microsporidia (Lipa and Ziemni-cka, 1996). The occurrence of a baculovirus infecting satinmoth larvae was Wrst reported by Weiser et al. (1954) andlater Skatulla (1985). This virus, L. salicis (Lesa) NPV(also known as SsMNPV), was shown to play a majorrole in regulating the size of Leucoma salicis populations(Ziemnicka, 1982). The biological activity of LesaNPVagainst satin moth larvae has been evaluated (Lameriset al., 1985) and a sevenfold diVerence in virulencebetween LesaNPV from Poland and former Yugoslaviahas been noted. The genome of LesaNPV has been esti-mated at 128–134 kb in size, based on restriction enzymeanalysis of four Polish isolates (Strokovskaya et al., 1996)but its phylogenetic status has not been investigated.
Baculoviruses comprise a family of double strandedDNA viruses infecting primarily insects from the ordersLepidoptera, Diptera, and Hymenoptera. The family Bac-uloviridae is divided into two genera, Nucleopolyhedrovi-rus (NPV) and Granulovirus (GV) (Blissard et al., 2000)based on occlusion body morphology. The lepidopteran-speciWc NPVs are further divided into two groups, group INPV and group II NPV based on single gene phylogeny(Bulach et al., 1999), and conWrmed by whole genomephylogenies (Herniou et al., 2001). Up until now, morethan 700 baculoviruses have been recorded, and many ofthese have been characterized biologically and/or bio-chemically (Moscardi, 1999). More than 23 baculovirusgenomes have been fully sequenced and characterized(Lange et al., 2004). Most phylogenetic analyses so farhave been based on single-gene sequences from lepi-dopteran baculoviruses, which often led to conXictingphylogenies when diVerent genes were used.
The latest analyses based on complete genomesequences enable the selection of genes suitable for singlegene phylogenetic studies (Herniou et al., 2004): lef-8and pif-2 (Ac22). The polyhedrin gene sequence is themost widely used gene for phylogenetic analyses, how-ever, phylogenies derived for polyhedrin are usually atvariance to those of other core genes (Harrison and Bon-ning, 2003; Herniou et al., 2001), and it has recently beenshown that this is in part due to the mosaic structure ofthis gene in Autographa californica MNPV (Lange et al.,2004). Polyhedrin, lef-8, and pif-2 are conserved in alllepidopteran baculoviruses analyzed thus far. Polyhe-drin is the major protein of occlusion bodies (OBs). Lef-8 encodes a late expression factor which is required fortranscription of late baculovirus genes and forms,together with lef-4, lef-9, and p47, the baculovirusencoded RNA polymerase (Titterington et al., 2003). Pif-2 is essential for oral infectivity, but is not required forvirus replication in cultured insect cells (Pijlman et al.,2003).
The aim of the current study was to characterize Les-aNPV on a molecular basis and to evaluate its taxo-
nomic status using sequences of the conservedbaculovirus genes, lef-8, pif-2, and polyhedrin.
2. Material and methods
2.1. Insects and viruses
Satin moth larvae were reared in the Department ofBiocontrol and Quarantine, Institute of Plant Protectionin Poznan, Poland. Larvae at the stage of L3 and L4were collected from poplar trees in the years 1998–2002and reared in the laboratory on fresh poplar (Populusnigra) leaves during the season. Second instar larvaewere kept at 4 °C over winter. The larvae were reared inlarge glass vessels at 20–25 °C, 60–70% relative humidityand 18:6 h photoperiod up to pupation. They were fedwith poplar leaves changed at least every 2 days. Emerg-ing adults were transferred to new vessels to lay eggs onpaper. Egg masses were placed in plastic or glass boxeswith fresh leaves.
LesaNPV was isolated in Poland (Kutno) in 1984from a number of infected larvae feeding on poplar treesand stored at ¡20 °C. The original virus isolate wasfreshly ampliWed in fourth instar larvae of Leucoma sali-cis reared in the laboratory in 2004. Larvae were infectedindividually by feeding with poplar leaf discs contami-nated with 10�l of virus suspension (107 occlusionbodies/ml). Occlusion bodies (OBs) were puriWed fromdead larvae as described by Muñoz et al. (1997).
OpMNPV (TM Biocontrol-1) was kindly obtainedfrom Dr. Imre Otvos, PaciWc Forestry Centre, Victoria,Canada.
2.2. DNA extraction and restriction enzyme analysis
DNA was isolated according to Reed et al. (2003)with the modiWcation of using dialysis following phe-nol:chloroform:isoamyl alcohol extraction. DNA solu-tion was dialyzed against three changes of TE buVer(1 mM Tris–HCl, 0.1 mM EDTA, pH 8.0) at 4 °C for 24–48 h. For restriction enzyme analyses 1 �g of DNA wasdigested for 3.5 h at 37 °C with HindIII, NotI or PstI,electrophoresed in 0.7% TAE [40 mM Tris–acetate,1 mM EDTA (pH 8.0)], separated in 0.6% agarose gels at15 mA for 18 h and analyzed under UV light after stain-ing the DNA with ethidium bromide.
2.3. PCR ampliWcation and cloning
PuriWed DNA was used as a template for PCR. Thedegenerate primer set for the polh gene was previouslydescribed by de Moraes and Maruniak (1997), and for thelef-8 and pif-2 genes by Herniou et al. (2004). The primerset used for the lef-7 region was 5�-gtaaaacgacggccagtCACAATTCGTTACACGCG-3� (forward) and 5�-aacag

102 A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107
ctatgaccatgGAGGGGCGACTTGATTTC-3� (reverse),and for the ctl-2 region, 5�-gtaaaacgacggccagtCGTGCAGCCGTTGCTGGTGT-3� (forward) and 5�-aacagctatgaccatgGCAGGTGGAGGTGTATGAG-3� (reverse).The nucleotides in lower case represent recognition sitesfor primers used in subsequent sequence analysis. Foramplifying the dUTPase region the forward primer 5�-GTGCATGACAGGTGCGTGATTG-3� and thereverse primer 5�-CAATGCAGTGCTCGTCGACAAG-3� were used.
Each 25�l PCR reaction mixture contained 30–50 ngviral DNA, 400 nM of each primer (Table 1), 0.2 mM ofeach dATP, dCTP, dGTP, and dTTP, 0.5 U Taq DNApolymerase (Promega) and 1.5 mM MgCl2 and 2.5 �l10£ reaction buVer (Promega). Reactions were carriedout in a Bio-Rad thermocycler using the followingparameters: 95 °C denaturation (5 min), 10 cycles of94 °C (60 s), 45 °C (45 s), 72 °C (60 s), followed by 25cycles of 94 °C (45 s), 50 °C (30 s), 72 °C (60 s), and Wnally72 °C (5 min) for polh, ctl-2, and lef-7 gene-speciWcprimer sets, and 95 °C denaturation (5 min), 10 cycles of94 °C (60 s), 42 °C (45 s), 72 °C (60 s), followed by 25cycles of 94 °C (45 s), 60 °C (30 s), 72 °C (60 s), and Wnally72 °C (5 min) for lef-8 and pif-2 speciWc primer sets. PCRproducts that did not contain universal M13 (-20) andM13 R primers at the 5�end and 3�end, respectively, werecloned into pGEM-T easy (Promega). The PCR prod-ucts were either directly sequenced (primer sets with5�extensions of universal M13 (-20) and M13 R primers)or after cloning into pGEM-T easy. The nucleotidesequences of the PCR products were determined byautomated sequencing (BaseClear, The Netherlands).The sequences obtained for polyhedrin, lef-8, and pif-2
were deposited in GenBank under Accession Nos.AY729808, AY729809, and AY729810, respectively.
2.4. Sequence analysis and phylogeny
Baculovirus polyhedrin, lef-8, and pif-2 gene sequenceswere obtained from GenBank (see Table 1) to be com-pared with LesaNPV. The BLAST program (Altschulet al., 1990) at the National Center for BiotechnologyInformation (NCBI) was used for nucleotide and pre-dicted amino acid sequence homology searches. Align-ments of 498, 390, and 315 nt long fragments for polh, lef-8, and pif-2 genes, respectively, were performed. The maxi-mum parsimony alignments of the nucleotide sequenceswere performed using PAUP 4.0 (SwoVord, 1998). Theanalysis of lef-8 and pif-2 was combined as their tree struc-tures were found to be congruent in phylogenetic analyses(Herniou et al., 2004). The polyhedrin gene was analyzedseparately. The alignment for lef-8 + pif-2 contained eightNPV sequences, and for polyhedrin 18 baculovirussequences. The alignments were bootstrapped 1000 timesand dendrograms were drawn using Tree View.
2.5. Cross infectivity
Two separate bioassays were performed to check thecross infectivity of LesaNPV and OpMNPV for O. pseu-dotsugata and L. salicis larvae, respectively. The bioassayon O. pseudotsugata was performed by Andrea Schiller inthe laboratory of Dr. Imre Otvos, Canadian Forest Ser-vice. O. pseudotsugata larvae were reared on artiWcial diet(Thompson and Peterson, 1978). Newly molted L3 lar-vae were exposed to six concentrations of LesaNPV:
Table 1Baculovirus sequences used for phylogenetic analyses
Virus name Abbreviation Polh Pif-2 Lef-8 References
Autographa californica MNPV AcMNPV L22858 L22858 L22858 Ayres et al. (1994)Amsacta albistriga NPV AmalNPV AF118850 — — —Anticarsia gemmatalis MNPV AgMNPV Y17753 — — Zanotto et al. (1992)Anagrapha falcifera NPV AnafaNPV U64896 — — Federici and Hice (1997)Archips cerasivoranus NPV ArceNPV U40834 — — —Attacus riccini NPV AtruNPV S68462 — — Hu et al. (1993)Bombyx mori MNPV BmMNPV U75359 — — Gomi et al. (1999)Choristoneura fumiferana MNPV CfMNPV AF512031 AF512031 AF512031 Lee et al. (1992)Choristoneura rosaceana MNPV ChroMNPV U91940 — — Lucarotti and Morin (1997)Epiphyas postvittana NPV EppoNPV AY043265 AY043265 AY043265 Hyink et al. (2002)Heliconius erato NPV HeerNPV — AY449792 AY449771 Herniou et al. (2004)Helicoverpa armigera SNPV HaSNPV AF271059 AF271059 AF271059 Chen et al. (1997)Leucoma salicis MNPV LesaNPV AY729808 AY729809 AY729810 This studyLonomia obliqua MNPV LoobMNPV AF232690 — — WolV et al. (2002)Lymantria dispar MNPV LdMNPV AF081810 AF081810 AF081810 Kuzio et al. (1999)Orgyia pseudotsugata MNPV OpMNPV NC001875 NC001875 NC001875 Ahrens et al. (1997)Perina nuda NPV PnNPV U22824 — — Chou et al. (1996)Rachplusia ou MNPV RoMNPV AF068270 — — Harrison and Bonning (1999)Spodoptera exigua MNPV SeMNPV AF169823 — — Van Strien et al. (1992)Thysanoplusia orichalcea NPV ThorNPV AF169480 — — Cheng et al. (1998)Xestia c-nigrum GV XecnGV U70069 — — Hayakawa et al. (1999)

A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107 103
101–106 OBs per larvae. Larvae were starved for 24 h andthen fed a diet plug contaminated with one of the sixvirus concentrations and reared separately for 21 days.After they consumed the entire contaminated diet plugthey were given fresh diet and the diet was changedweekly. The larvae were incubated at 25°C with a relativehumidity of 60–70% and a 16:8 h day:night photoperiod.Twenty larvae were used per dose, with no replicates.Twenty larvae inoculated with distilled water served ascontrols. Mortality was recorded daily for 21 days postinoculation. There was no mortality in controls.
OpMNPV was tested on Wrst instar (3 days old)L. salicis larvae. Six virus concentrations were used: 103–108 OBs per 12 larvae. Larvae were starved for 6 h andthen fed with leaf discs contaminated with virus suspen-sion. After they consumed the contaminated leaves theywere given fresh leaves. Tested larvae were incubated inglass vessels (Ø 10 cm) on the lab bench, at 20 °C, with50% relative humidity and a 14:10 h day:night photope-riod. Twenty-four larvae per dose were tested and therewere three replicates. In each replicate, 24 larvae servedas a control. Mortality was recorded for 15 days post ini-tial exposure. There was no mortality in controls.
3. Results
3.1. Polh, lef-8, and pif-2 gene sequencing and phylogenetic analysis
The purpose of this study was to determine the taxo-nomic position of LesaNPV. To this aim, a 600-nt frag-
ment of the polyhedrin (polh) sequence, a 800-ntfragment of the lef-8 gene, and a 450 nt fragment of thepif-2 gene were ampliWed by PCR using degenerate prim-ers. Alignment of polh, lef-8, and pif-2 sequences indi-cates the closest relationship of LesaNPV with group Ibaculoviruses, and in particular with OpMNPV. Thephylogenetic analysis of the polh sequence of group Ibaculoviruses showed a strongly supported group com-prising LesaNPV, OpMNPV and Perina nuda MNPV(Fig. 1A). The combined lef-8 and pif-2 analysis localizedLesaNPV together with OpMNPV and CfMNPV, andthis was supported by high bootstrap scores (Fig. 1B). Inboth cases (polh and lef8 + pif-2) LesaNPV is mostclosely related to OpMNPV. The degree of homologybetween polh, lef-8 and pif-2 gene fragments of LesaNPVand OpMNPV was 99, 98, and 97%, respectively, at thenucleotide level. The sequence of the polyhedrin proteinswas identical for these two viruses. The LesaNPV andOpMNPV Lef-8 protein sequence varied in two aminoacids and the Pif-2 proteins revealed diVerences in Wveamino acid positions.
3.2. Restriction proWles of LesaNPV in comparison to OpMNPV
Since sequencing of three conserved baculovirusesgenes suggested a very high level of similarity betweenLesaNPV and OpMNPV their restriction endonucleasedigestion proWles were compared (Fig. 2). Restrictionpatterns using HindIII, NotI or PstI, indicated that Les-aNPV and OpMNPV, although closely related, showedvery clear restriction fragment length polymorphisms
Fig. 1. Baculovirus group I phylogenies. (A) polyhedrin tree, (B) combined lef-8 and pif-2 tree. The phylogeny trees were obtained by maximum parsi-mony analyses of partial DNA sequence data, numbers indicate bootstrap scores.

104 A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107
(RFLPs). The most similar were the PstI digestion pro-Wles, although clear diVerences were observed in the 5–9 kbp area. Comparison of the NotI and PstI proWlesshowed common bands as well as bands speciWc to eithervirus. The presence of submolar bands in the LesaNPVdigests suggests genetic heterogeneity.
3.3. Analysis of the lef-7 and ctl-2 regions
To locate diVerences between LesaNPV andOpMNPV, two genome fragments were selected thatcould have the potential to serve as molecular determi-nants for either virus, the lef-7 region and the conotoxin
Fig. 2. Comparison of restriction enzyme digestion proWles of Les-aNPV and OpMNPV. Viral DNA was isolated from polyhedra frominfected satin moth larvae (LesaNPV) and from powder formulatedTM Biocontrol-1 (OpMNPV). LesaNPV and OpMNPV DNA weredigested with HindIII (lanes 1 and 2), PstI (lanes 3 and 4) and NotI(lanes 5 and 6) and analysed by elecrophoresis in an ethidium bromidestained 0.7% agarose gel. M-Lambda EcoRI/BamHI/HindIII frag-ments. Asterisks (*) indicate minor molar bands.
like-2 (ctl-2) region, both present in OpMNPV. Lef-7encodes a late expression factor found to be necessaryfor late promoter-driven reporter gene expression in aSpodoptera frugiperda cell line, but not in a Trichoplu-sia ni cell line. Substitution in this gene may modulatethe ability of NPVs to replicate in diVerent cell lines(Lu and Miller, 1995). Lef-7 was also found to undergopositive selection in alternate hosts (Harrison and Bon-ning, 2004). Ctl-2 gene was selected after analysis ofdiVerences between the LesaNPV and OpMNPVrestriction enzyme (HindIII) proWles, using the wholegenome sequence of OpMNPV (Ahrens et al., 1997) tolocate variable regions. A single ctl gene is present inseveral baculovirus genomes (Harrison and Bonning,2003), whereas OpMNPV encodes two ORFs withconotoxin-like domains, ctl-1 and ctl-2. As the ctl-2gene appears to be unique for OpMNPV, this regionhad the potential to show diVerences between the twoviruses. Primers speciWc for the lef-7 and ctl-2 regionswere designed using the published OpMNPV sequence(Ahrens et al., 1997). The targeted regions also con-tained homologous repeats (hrs), sites of frequentrecombination and rearrangement in baculovirusgenomes: the ctl-2 region harbors hr2; the lef-7 regionhr4.
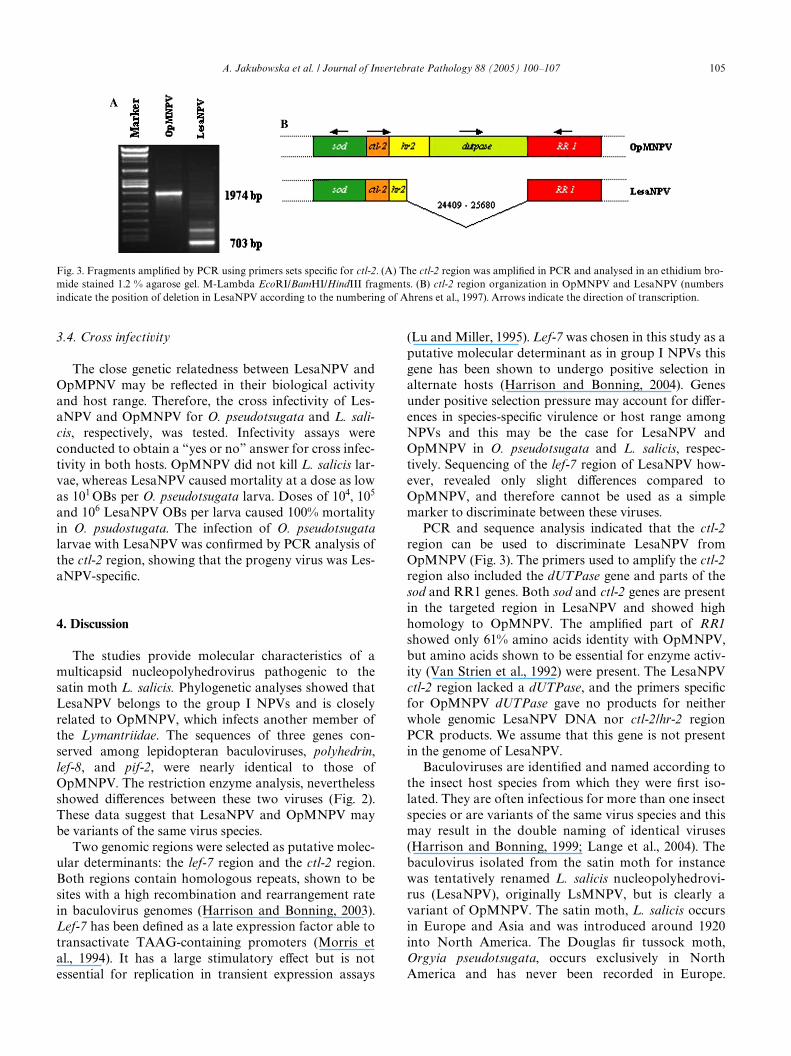
With primers speciWc for the lef-7/hr4 region, PCRproducts of a similar, expected size were obtained forboth viruses. Only minor diVerences (2%) were revealedat the sequence level (data not shown). PCR ampliWca-tion of the ctl-2/hr2 region resulted in a fragment of theexpected size for OpMNPV (1974 bp), whereas for Les-aNPV several shorter fragments were obtained(Fig. 3A). The ampliWed ctl-2 region of OpMNPV con-tained part of the superoxide dismutase (sod) gene(5�end), ctl-2, hr-2, dUTPase, and part of the ribonucleo-tide reductase large subunit (RR1) gene (3�end). Onlyone of the major PCR products obtained with Les-aNPV, a 703 bp product, was ampliWed from the regionof interest. This LesaNPV product lacked nucleotides24,409–25,680 (Fig. 3B) when compared to theOpMNPV genome (numbering of Ahrens et al., 1997).In OpMNPV, the fragment of 1271 nt which is absent inthe LesaNPV genome, harbors the dUTPase gene(953 nt), a large part of hr-2 (144 nt), two intragenicregions (52 and 102 nt) and 20 nt from the 3�end of theRR1 gene (Fig. 3B). The nature of the minor LesaNPVPCR products was not analyzed. An additional set ofprimers was designed to amplify the dUTPase gene.These primers were used for a nested PCR on the ctl-2/hr2 products of both viruses. A product of the expectedsize was obtained for OpMNPV and no products werefound for LesaNPV, suggesting that dUTPase wasindeed absent from this part of the genome of the lattervirus. When these dUTPase primers were used for aPCR with the LesaNPV genome as template no ampliW-
cation product was observed.
A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107 105
3.4. Cross infectivity
The close genetic relatedness between LesaNPV andOpMPNV may be reXected in their biological activityand host range. Therefore, the cross infectivity of Les-aNPV and OpMNPV for O. pseudotsugata and L. sali-cis, respectively, was tested. Infectivity assays wereconducted to obtain a “yes or no” answer for cross infec-tivity in both hosts. OpMNPV did not kill L. salicis lar-vae, whereas LesaNPV caused mortality at a dose as lowas 101 OBs per O. pseudotsugata larva. Doses of 104, 105
and 106 LesaNPV OBs per larva caused 100% mortalityin O. psudostugata. The infection of O. pseudotsugatalarvae with LesaNPV was conWrmed by PCR analysis ofthe ctl-2 region, showing that the progeny virus was Les-aNPV-speciWc.
4. Discussion
The studies provide molecular characteristics of amulticapsid nucleopolyhedrovirus pathogenic to thesatin moth L. salicis. Phylogenetic analyses showed thatLesaNPV belongs to the group I NPVs and is closelyrelated to OpMNPV, which infects another member ofthe Lymantriidae. The sequences of three genes con-served among lepidopteran baculoviruses, polyhedrin,lef-8, and pif-2, were nearly identical to those ofOpMNPV. The restriction enzyme analysis, neverthelessshowed diVerences between these two viruses (Fig. 2).These data suggest that LesaNPV and OpMNPV maybe variants of the same virus species.
Two genomic regions were selected as putative molec-ular determinants: the lef-7 region and the ctl-2 region.Both regions contain homologous repeats, shown to besites with a high recombination and rearrangement ratein baculovirus genomes (Harrison and Bonning, 2003).Lef-7 has been deWned as a late expression factor able totransactivate TAAG-containing promoters (Morris etal., 1994). It has a large stimulatory eVect but is notessential for replication in transient expression assays
(Lu and Miller, 1995). Lef-7 was chosen in this study as aputative molecular determinant as in group I NPVs thisgene has been shown to undergo positive selection inalternate hosts (Harrison and Bonning, 2004). Genesunder positive selection pressure may account for diVer-ences in species-speciWc virulence or host range amongNPVs and this may be the case for LesaNPV andOpMNPV in O. pseudotsugata and L. salicis, respec-tively. Sequencing of the lef-7 region of LesaNPV how-ever, revealed only slight diVerences compared toOpMNPV, and therefore cannot be used as a simplemarker to discriminate between these viruses.
PCR and sequence analysis indicated that the ctl-2region can be used to discriminate LesaNPV fromOpMNPV (Fig. 3). The primers used to amplify the ctl-2region also included the dUTPase gene and parts of thesod and RR1 genes. Both sod and ctl-2 genes are presentin the targeted region in LesaNPV and showed highhomology to OpMNPV. The ampliWed part of RR1showed only 61% amino acids identity with OpMNPV,but amino acids shown to be essential for enzyme activ-ity (Van Strien et al., 1992) were present. The LesaNPVctl-2 region lacked a dUTPase, and the primers speciWcfor OpMNPV dUTPase gave no products for neitherwhole genomic LesaNPV DNA nor ctl-2/hr-2 regionPCR products. We assume that this gene is not presentin the genome of LesaNPV.
Baculoviruses are identiWed and named according tothe insect host species from which they were Wrst iso-lated. They are often infectious for more than one insectspecies or are variants of the same virus species and thismay result in the double naming of identical viruses(Harrison and Bonning, 1999; Lange et al., 2004). Thebaculovirus isolated from the satin moth for instancewas tentatively renamed L. salicis nucleopolyhedrovi-rus (LesaNPV), originally LsMNPV, but is clearly avariant of OpMNPV. The satin moth, L. salicis occursin Europe and Asia and was introduced around 1920into North America. The Douglas Wr tussock moth,Orgyia pseudotsugata, occurs exclusively in NorthAmerica and has never been recorded in Europe.
Fig. 3. Fragments ampliWed by PCR using primers sets speciWc for ctl-2. (A) The ctl-2 region was ampliWed in PCR and analysed in an ethidium bro-mide stained 1.2 % agarose gel. M-Lambda EcoRI/BamHI/HindIII fragments. (B) ctl-2 region organization in OpMNPV and LesaNPV (numbersindicate the position of deletion in LesaNPV according to the numbering of Ahrens et al., 1997). Arrows indicate the direction of transcription.

106 A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107
The insects are taxonomically related, both belong tothe family Lymantriidae, subfamily Orgyiinae, but todiVerent tribes, Lymantriini (L. salicis) and Orgyiini (O.pseudotsugata). The high level of similarity in severalcore genes indicates that the NPVs isolated from bothinsects are closely related.
A nucleopolyhedrovirus infecting O. pseudotsugatawas Wrst documented by Eveden and Jost (1947). Latertwo diVerent types of occlusion bodies were isolatedfrom O. pseudotsugata larvae (Hughes and Addison,1970), OpMNPV and OpSNPV. In the satin moth fromEurope, only an MNPV type has been described, andpreliminary results using OpSNPV-speciWc primers in aPCR reaction with LesaNPV as a template suggest thatOpSNPV is absent. These results suggest two possiblescenarios: LesaNPV may have migrated together withthe host insect L. salicis to North America and may bethe ancestor of OpMNPV, or alternatively that Les-aNPV and OpMNPV have a common ancestor with abroader geographic distribution.
A correlation between baculovirus phylogenies andthe taxonomic and ecological relationships of theirhosts has been observed and coevolution betweeninteracting insect and virus species has been postulated(Herniou et al., 2003). Therefore, when a baculovirusinvades a new host species (e.g., O. pseudotsugata) it islikely that this host would be closely related to its cur-rent host. Adaptation to the new host may eventuallylead to the isolation of a diVerent genotype (Herniou etal., 2004). This may be the case for the satin moth andthe Douglas Wr tussock moth and their NPVs.Although the host plants of these two forest species arenot the same, their geographical ranges overlap in thePaciWc northwest. In addition, it is well established thatbaculoviruses can be dispersed rapidly by a variety ofmeans, including birds (Entwistle et al., 1993), whichwill enable virus to spread over wide areas and encoun-ter a variety of potential host species. Additionally,studies on the geographic distribution of OrygiaMNPVs and SNPVs show that the MNPV only occursin the area around British Columbia, Washington, andIdaho, whereas the SNPV is distributed throughout therange of the Orgyia species (Hughes, 1976), providingindirect support for an introduction in that area. Les-aNPV was infectious for O. pseudostugata, whereasOpMNPV was not infectious for L. salicis. This sug-gests that LesaNPV could have infected O. pseudotsug-ata and has since evolved away from an ancestral virusinto OpMNPV. This would also imply that OpMNPVhas acquired a dUTPase gene recently. This is notimpossible and is in line with the phylogenetic observa-tion that the dUTPase gene is lost and gained by bac-uloviruses (Herniou et al., 2003). It would requirefurther sequencing and analysis of the LesaNPVgenome to conWrm that a dUTPase gene is indeedabsent.
Several other baculoviruses isolated from Orgyia spe-cies are singly enveloped NPVs (Hughes, 1976; Richardset al., 1999; Sohi et al., 1984), and more importantly,phylogenetic evidence has shown that both Old and Newworld Orgyia NPV species, except OpMNPV but includ-ing OpSNPV, cluster together within the group II NPVs(Herniou et al., 2003). Thus there is little evidence forMNPVs being found in other Orgyia species and thedata imply a close relation between other Orgyia NPVs.Further information is needed to support the hypothesisof host switching, in particular, analysis of archival sam-ples of OpMNPV or NPVs from L. salicis. Both virusesand their insect hosts provide an excellent system forstudying baculovirus host range and evolution.
Acknowledgments
This research was partly supported by a scholarshipfrom the European Union (Functional Biodiversity andCrop Protection), contract HPMT-CT-2000-00199. Theauthors are grateful to Dr. Imre Otvos and AndreaSchiller for providing OpMNPV and for carrying outinfectivity assays on O. pseudotsugata.
References
Ahrens, C.H., Russell, R.L., Funk, C.J., Evans, J.T., Harwood, S.H., Rohr-mann, G.F., 1997. The sequence of the Orgyia pseudotsugata multinu-cleocapsid nuclear polyhedrosis virus genome. Virology 229, 381–399.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990.Basic local alignment search tool. J. Mol. Biol. 215, 403–410.
Ayres, M.D., Howard, S.C., Kuzio, J., Lopez-Ferber, M., Possee, R.D.,1994. The complete DNA sequence of Autographa californicanuclear polyhedrosis virus. Virology 202, 586–605.
Blissard, G.W., Black, B., Crook, N., Keddie, B.A., Possee, R., Rohr-man, G.F., Theilman, D.A., Volkman, L., 2000. Family Baculoviri-dae. In: M.H.V. Van Regenmortel et al. (Eds.), Virus Taxonomy:Seventh Report of the International Committee on Taxonomy ofViruses. Academic Press, San Diego, California.
Bulach, D.M., Kumar, C.A., Zaia, A., Liang, B., Tribe, D.E., 1999.Group II nucleopolyhedrovirus subgroups revealed by phyloge-netic analysis of polyhedrin and DNA polymerase gene sequences.J. Invertebr. Pathol. 73, 59–73.
Chen, X., Hu, Z., Vlak, J.M., 1997. Sequence analysis of the Heliothisarmigera single-nucleocapsid nucleopolyhedrovirus polyhedrin.Zhongguo Bingduxue 12, 346–353.
Cheng, X.W., Carner, G.R., Fescemyer, H.W., 1998. Polyhedrinsequence determines the tetrahedral shape of occlusion bodies inThysanoplusia orichalcea single-nucleocapsid nucleopolyhedrovi-rus. J. Gen. Virol. 79, 2549–2556.
Chou, C.M., Huang, C.J., Lo, C.F., Kou, G.H., Wang, C.H., 1996. Char-acterization ofPerina nuda nucleopolyhedrovirus (PenuNPV) poly-hedrin gene. J. Invertebr. Pathol. 67, 259–266.
de Moraes, R.R., Maruniak, J.E., 1997. Detection and identiWcation ofmultiple baculoviruses using the polymerase chain reaction (PCR)and restriction endonuclease analysis. J. Virol. Methods 63, 209–217.
Entwistle, P.F., Forkner, A.C., Green, B.M., Cory, J.S., 1993. Avian dis-persal of nuclear polyhedrosis virus after induced epizootics in the

A. Jakubowska et al. / Journal of Invertebrate Pathology 88 (2005) 100–107 107
pine beauty moth, Panolis Xammea, (Lepidoptera: Noctuidae). Biol.Control 3, 61–69.
Eveden, J., Jost, E.J., 1947. A report of the tussock moth control, NorthIdaho, 1947. U.S. Dep. Agr., Forest Serv., Idaho State Forest. Dep.51pp.
Federici, B.A., Hice, R.H., 1997. Organization and molecular character-ization of genes in the polyhedrin region of the Anagrapha falciferamultinucleocapsid NPV. Arch. Virol. 142, 333–348.
Gomi, S., Majima, K., Maeda, S., 1999. Sequence analysis of the genomeof Bombyx mori nucleopolyhedrovirus. J. Gen. Virol. 80, 323–337.
Harrison, R.L., Bonning, B.C., 1999. The nucleopolyhedroviruses ofRachiplusia ou and Anagrapha falcifera are isolates of the samevirus. J. Gen. Virol. 80, 2793–2798.
Harrison, R.L., Bonning, B.C., 2003. Comparative analysis of thegenomes of Rachiplusia ou and Autographa californica multiplenucleopolyhedroviruses. J. Gen. Virol. 84, 1827–1842.
Harrison, R.L., Bonning, B.C., 2004. Application of maximum-likeli-hood models to selection pressure analysis of group I nucleopoly-hedrovirus genes. J. Gen. Virol. 85, 197–210.
Hayakawa, T., Ko, R., Okano, K., Seong, S.I., Goto, C., Maeda, S.,1999. Sequence analysis of the Xestia c-nigrum granulovirusgenome. Virology 262, 277–297.
Herniou, E.A., Luque, T., Chen, X., Vlak, J.M., Winstanley, D., Cory, J.,O’Reilly, D.R., 2001. Use of whole genome sequence data to inferbaculovirus phylogeny. J. Virol. 75, 8117–8126.
Herniou, E.A., Olszewski, J.A., Cory, J.S., O’Reilly, D.R., 2003. Thegenome sequence and evolution of baculoviruses. Annu. Rev. Ento-mol. 48, 211–234 Review.
Herniou, E.A., Olszewski, J.A., O’Reilly, D.R., Cory, J.S., 2004. Ancientcoevolution of baculoviruses and their insect hosts. J. Virol. 78,3244–3251.
Hu, J., Ding, H., Wu, X., 1993. Cloning and sequencing of Attacus ricininuclear polyhedrosis virus polyhedrin gene. Yi Chuan Xue Bao 20,300–304.
Hughes, K.M., 1976. Notes on the nuclear polyhedrosis viruses of tussockmoths of the genus Orgyia (Lepidoptera). Can. Ent. 108, 479–484.
Hughes, K.M., Addison, R.B., 1970. Two nuclear polyhedrosis virusesof the Douglas-Wr tussock moth. J. Invert. Pathol. 16, 196–204.
Hyink, O., Dellow, R.A., Olsen, M.J., Caradoc-Davies, K.M., Drake,K., Herniou, E.A., Cory, J.S., O’Reilly, D.R., Ward, V.K., 2002.Whole genome analysis of the Epiphyas postvittana nucleopolyhe-drovirus. J. Gen. Virol 83, 957–971.
Kuzio, J., Pearson, M.N., Harwood, S.H., Funk, C.J., Evans, J.T., Sla-vicek, J.M., Rohrmann, G.F., 1999. Sequence and analysis of thegenome of a baculovirus pathogenic for Lymantria dispar. Virology253, 17–34.
Lameris, A.M.C., Ziemnicka, J., Peters, D., Grijpma, P., Vlak, J.M.,1985. Potential of baculoviruses for control of the satin moth, Leu-coma salicis L. (Lepidoptera: Lymantriidae). Med. Fac Land-bouww. Rijksuniv. Gent. 50, 431–439.
Lange, M., Wang, H., Zhihong, H., Jehle, J.A., 2004. Towards a molec-ular identiWcation and classiWcation system of lepidopteran-speciWcbaculoviruses. Virology 325, 36–47.
Langor, D.W., 1995. Satin moth. Nat. Resour. Can., Can. For. Serv.,North. For. Cent., Edmonton, Alberta. For. LeaX. 35.
Lee, H.Y., Arif, B., Dobos, P., Krell, P., 1992. IdentiWcation of bentDNA and ARS fragments in the genome of Choristoneura fumifer-ana nuclear polyhedrosis virus. Virus Res. 24, 249–264.
Lipa, J.J., Ziemnicka, J., 1996. Spiroplasma and microsporidian infec-tions in populations of Leucoma salicis L. (Lepidoptera: Lymantrii-dae) in Poland. In: Proceedings of International Conference“Integrated Management of Forest Lymantriidae”, Warsaw-Sêko-cin (Poland) pp. 27–29.
Lu, A., Miller, L.K., 1995. The roles of eighteen baculovirus late expres-sion factor genes in transcription and DNA replication. J. Virol. 69,975–982.
Lucarotti, C.J., Morin, B., 1997. A nuclear polyhedrosis virus from theobliquebanded leafroller, Choristoneura rosaceana (Harris) (Lepi-doptera: Tortricidae). J. Invertebr. Pathol. 70, 121–126.
Morris, T.D., Todd, J.W., Fisher, B., Miller, L.K., 1994. IdentiWcationof lef-7: a baculovirus gene aVecting late gene expression. Virology200, 360–369.
Moscardi, F., 1999. Assessment of the application of baculoviruses forcontrol of Lepidoptera. Annu. Rev. Entomol. 44, 57–89.
Muñoz, D., Vlak, J.M., Caballero, P., 1997. In vivo recombinationbetween two strains of the genus nucleopolyhedrovirus in its natu-ral host, Spodoptera exigua. Appl. Environ. Microbiol. 63, 3025–3031.
Pijlman, G.P., Pruijssers, A.J., Vlak, J.M., 2003. IdentiWcation of pif-2, athird conserved baculovirus gene required for per os infection ofinsects. J. Gen. Virol. 84, 2041–2049.
Reed, C., Otvos, I.S., Reardon, R., Ragenovich, I., Williams, H.L., 2003.EVects of long-term storage on the stability of OpMNPV DNAcontained in TM Biocontrol-1. J. Invertebr. Pathol. 84, 104–113.
Richards, A., Speight, M., Cory, J., 1999. Characterization of a nucleo-polyhedrovirus from the vapourer moth, Orgyia antiqua (Lepidop-tera, Lymantriidae). J. Invertebr. Pathol. 74, 137–142.
Skatulla, U., 1985. Untersuchungen zur Wirkung eines Kernpolyeder-virus aus Leucoma salicis (Lep., Lymantriidae) auf einige Lymantri-iden-Arten. Anz. für Schändlings. PXanzenschutz Umweltschutz.58, 41–47.
Sohi, S.S., Percy, M., Cunningham, J.C., 1984. Replication and serialpassage of a singly enveloped baculovirus from Orgyia leucostigmain homologous cell lines. Intervirology 21, 50–60.
Strokovskaya, L., Ziemnicka, J., Michalik, J., 1996. Genetic variabil-ity of four natural isolates of the Stilpnotia salicis multiple-envel-oped nuclear polyhedrosis virus. Acta Biochim. Polonica 43, 633–638.
SwoVord, D.L., 1998. PAUP*. Phylogenetic analysis using parsimony(* and other methods). Version 4. Sinauer Associates, Sunderland,MA.
Thompson, C.G., and Peterson, L.J., 1978. Rearing the Douglas-Wr tus-sock moth. USDA Agriculture Handbook No. 520, Combined For-estry Pest Research and Development Program, Washington, DC.20pp.
Titterington, J.S., Nun, T.K., Passarelli, A.L., 2003. Functional dissec-tion of the baculovirus late expression factor-8 gene: sequencerequirements for late gene promoter activation. J. Gen. Virol. 84,1817–1826.
Van Strien, E.A., Zuidema, D., Goldbach, R.W., Vlak, J.M., 1992.Nucleotide sequence and transcriptional analysis of the polyhedringene of Spodoptera exigua nuclear polyhedrosis virus. J. Gen. Virol.73, 2813–2821.
Weiser, J., Ludvik, J., Veber, J., 1954. Polyedrie bekyne vbrove (Stilpno-tia salicis L., Lepidoptera) (in Czechish) Zool. Entomol. Listy 4,238–242.
WolV, J.L.C., Moraes, R.H.P., Kitajima, E., de Souza Leal, E., Zanotto,P.M., 2002. IdentiWcation and characterization of a baculovirusfrom Lonomia obliqua (Lepidoptera: Saturniidae). J. Invertebr.Pathol. 79, 137–145.
Zanotto, P.M., Sampaio, M.J., Johnson, D.W., Rocha, T.L., Maruniak,J.E., 1992. The Anticarsia gemmatalis nuclear polyhedrosis viruspolyhedrin gene region: sequence analysis, gene product and struc-tural comparisons. J. Gen. Virol. 73, 1049–1056.
Ziemnicka, J., 1982. Studies on nuclear and cytoplasmic polyhe-drosis viruses of the satin moth (Stilpnotia salicis L.) (Lepi-doptera, Lymantriidae). Prace Nauk. Inst. Ochr. Roslin 23, 75–142.
Ziemnicka, J., 2000. Rola bakulowirusa SsMNPV w regulacji liczebno-sci populacji bialki wierzbowki Stilpnotia salicis L. (Lepidoptera:Lymantriidae). (in Polish) Rozprawy Naukowe Instytutu OchronyRoslin, Zeszyt 6, 85.
Copyright © 2022 FDOKUMEN




















