
Common Arterial Trunk Associated with Functionally ... - MDPI
Upload
independentCategory
view
2download
0
Two Zebrafish eIF4E Family Members Are DifferentiallyExpressed and Functionally Divergent*
Received for publication, December 15, 2003Published, JBC Papers in Press, December 29, 2003, DOI 10.1074/jbc.M313688200
Javier Robalino‡§¶, Bhavesh Joshi‡, Scott C. Fahrenkrug‡�, and Rosemary Jagus‡**‡‡
From the ‡Center of Marine Biotechnology, Baltimore, Maryland 21202, and §Escuela Superior Politecnica del Litoral,Km 30.5 Vıa Prosperina, Guayaquil, Ecuador, and **Program in Oncology, Greenebaum Cancer Center,Baltimore, Maryland 21201
Eukaryotic translation initiation factor 4E (eIF4E) isan essential component of the translational machinerythat binds m7GTP and mediates the recruitment ofcapped mRNAs by the small ribosomal subunit. Re-cently, a number of proteins with homology to eIF4Ehave been reported in plants, invertebrates, and mam-mals. Together with the prototypical translation factor,these constitute a new family of structurally relatedproteins. To distinguish the prototypical translationfactor eIF4E from other family members, it hasbeen termed eIF4E-1 (Keiper, B. D., Lamphear, B. J.,Deshpande, A. M., Jankowska-Anyszka, M., Aamodt,E. J., Blumenthal, T., and Rhoads, R. E. (2000) J. Biol.Chem. 275, 10590–10596). We describe the characteriza-tion of two eIF4E family members in the zebrafish Daniorerio. Based on their relative identities with humaneIF4E-1, these zebrafish proteins are termed eIF4E-1A(82%) and eIF4E-1B (66%). eIF4E-1B, originally termedeIF4E(L), has been reported previously as the zebrafisheIF4E-1 counterpart (Fahrenkrug, S. C., Dahlquist,M. O., Clark, K., and Hackett, P. B. (1999) Differentiation65, 191–201; Fahrenkrug, S. C., Joshi, B., Hackett, P. B.,and Jagus, R. (2000) Differentiation 66, 15–22). Sequencecomparisons suggest that the two genes probablyevolved from a duplication event that occurred duringvertebrate evolution. eIF4E-1A is expressed ubiqui-tously in zebrafish, whereas expression of eIF4E-1B isrestricted to early embryonic development and to go-nads and muscle of the tissues investigated. The abilityof these two zebrafish proteins to bind m7GTP, eIF4G,and 4E-BP, as well as to complement yeast conditionallydeficient in functional eIF4E, show that eIF4E-1A is afunctional equivalent of human eIF4E-1. Surprisingly,although eIF4E-1B possesses all known residuesthought to be required for interaction with the capstructure, eIF4G, and 4E-BPs, it fails to interact withany of these components, suggesting that this proteinserves a role other than that assigned to eIF4E.
Eukaryotic mRNAs are modified post-transcriptionally bythe addition of a 7-methylguanosine linked by a 5�–5�-triphos-phate bridge to the first transcribed residue (reviewed in Refs.4 and 5). This modification, known as a “cap” structure, isrequired for recruitment of mRNA by the translational machin-ery and is recognized by eukaryotic initiation factor 4E(eIF4E)1 (reviewed in Refs. 6 and 7). Recruitment of mRNAs tothe ribosome by eIF4E is accomplished through its interactionwith the factor eIF4G (reviewed in Ref. 8). eIF4G interacts witheIF4E, eIF4A (an ATP-dependent RNA-helicase (9, 10)), andribosome-bound eIF3 to place the small ribosomal subunit atthe 5� end of mRNA in close proximity to the cap (11–14). eIF4Ehas been characterized not only as a general translation factorbut as a protein that can modulate both the overall rates oftranslation and the mRNA selectivity of the translational ap-paratus (6, 7). The importance of the function of eIF4E isillustrated by the lethality of eif4e gene disruption in Saccha-romyces cerevisiae (15). Both the activity and the structure ofeIF4E are highly conserved throughout evolution; the humanand yeast homologues are 32% identical at the amino acid level,and the mammalian factor can rescue eif4e gene disruption inS. cerevisiae (15).
Sequence comparisons and mutational analyses of eIF4Ehave revealed a phylogenetically conserved �170-amino acidcore with eight conserved tryptophans, which appears to besufficient for cap binding (16–18). The three-dimensionalstructure of the mouse and yeast eIF4E core sequences boundto cap analogues have been solved by x-ray crystallography andNMR, respectively (16, 19). The core protein shows a cupped-hand shape, consisting of a curved, eight-stranded, antiparallel�-sheet backed by three �-helices (16, 19). The cap analoguebinds in a narrow slot within the concave surface (16, 19). Thebinding of the nucleotide residue of the cap structure resultsfrom the stacking of the alkylated base between two conservedTrp residues and the formation of hydrogen bonds between thenucleotide base, the polypeptide backbone, and a conserved Gluresidue (Trp-56, Trp-102, and Glu-103 for mouse eIF4E) (16,18, 20, 21). The presence of the N7-methyl moiety of the cap isdetected by another Trp residue (Trp-166 in mouse eIF4E) (16,19). Crystallographic studies have also revealed that residuesHis-37, Pro-38, Val-69, Trp-73, Leu-131, Glu-132, and Leu-135of mouse eIF4E make contacts with the eIF4E-binding domainsof eIF4Gs and 4E-BPs (22), although mutational analyses haveso far only identified Trp-73 (or its equivalent) as being re-
* This work was supported in part by National Science FoundationGrants MCB-9808401 and MCB-0134013 (to R. J. and B. J.). The costsof publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked “advertise-ment” in accordance with 18 U.S.C. Section 1734 solely to indicate thisfact.
¶ Supported by scholarships from Fundacion para la Ciencia y Tec-nologıa, Ecuador. Present address: Medical University of South Caro-lina, 173 Ashley Ave., Charleston, SC 29425.
� Present address: Dept. of Animal Science, University of Minnesota,495B Animal Science/Veterinary Medicine, 1988 Fitch Ave., St. Paul,MN 55108.
‡‡ To whom correspondence should be addressed: Center of MarineBiotechnology, Suite 236 Columbus Center, 701 E. Pratt St., Baltimore,MD 21202 Tel.: 410-234-8822; Fax: 410-234-8896; E-mail: [email protected].
1 The abbreviations used are: eIF4E, eukaryotic translation initiationfactor 4E; 4E-BPs, eIF4E-binding proteins; RT, reverse transcription;UTR, untranslated region; VSIEF, vertical slab isoelectric focusing;PVDF, polyvinylidene difluoride; ESTs, expressed sequence tags;CHAPS, 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonicacid.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 11, Issue of March 12, pp. 10532–10541, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org10532
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

quired for these interactions (19, 22). In contrast to the con-served cap binding core, the N and C termini of eIF4Es vary inlength, showing little or no conservation across the phyla. Mostof the residues in these regions have been shown to be dispen-sable for cap binding and complementation in yeast (17).
Both the level and activity of eIF4E are regulated (reviewedin Refs. 23 and 24). Higher levels of eIF4E generally correlatewith higher rates of protein synthesis and cell growth (re-viewed in Ref. 8). eIF4E undergoes regulated phosphorylationby Mnk1 and -2 (25, 26) at the residue equivalent to Ser-209 inhuman eIF4E-1 (27–29) (reviewed in Refs. 24 and 30). Therelationship between eIF4E phosphorylation and eIF4E activ-ity is not fully understood, although phosphorylation has beenshown to be important for normal growth (31). The positivecorrelation between phosphorylation and overall protein syn-thetic rate initially led to the supposition that phosphorylationincreases the ability of eIF4E to bind cap structures. However,subsequent kinetic analysis has revealed that phosphorylationdiminishes the ability of eIF4E to bind cap by increasing theoff-rate of eIF4E, suggesting that phosphorylation may serve tofacilitate release of eIF4E from the 5� cap structure during theinitiation cycle (32). Recruitment of mRNA by eIF4E can alsobe regulated by its association with a family of small eIF4E-binding proteins (4E-BPs), the members of which share homol-ogy with eIF4G and act as competitive inhibitors of the eIF4E/eIF4G interaction (22, 24, 33, 34).
Recently, a number of proteins with homology to eIF4E havebeen reported in plants, invertebrates, and mammals. Togetherwith the translation factor, these constitute a new family ofstructurally related proteins. This multiplicity of eIF4Es couldreflect simple functional redundancy or that the eIF4E-relatedproteins support alternate roles. These issues have been ad-dressed to a limited extent in Caenorhabditis elegans, in whichthe existence of several eIF4E-like factors can be partiallyexplained by the necessity of recruiting mRNAs with both7-methyl- and 2,2,7-methylguanosine caps to the initiationcomplex. In most systems, however, the biological significanceof multiple eIF4E-like genes remains to be explored.
We describe here the characterization of two eIF4E familymembers in the zebrafish Danio rerio. Based on their relativeidentity with human eIF4E-1, these zebrafish proteins aretermed eIF4E-1A and eIF4E-1B. The similarity of structure inthe core regions of eIF4E-1A and eIF4E-1B to human eIF4E-1suggests that both proteins could function as bona fide trans-lation factors. However, we demonstrate that only eIF4E-1Apossesses eIF4E-like activities and that this factor, unlikeeIF4E-1B, is able to complement eIF4E deficiency in S. cerevi-siae. Thus, by the analyses described, zebrafish eIF4E-1A is afunctional equivalent of human eIF4E-1, whereas the role ofeIF4E-1B remains to be determined.
EXPERIMENTAL PROCEDURES
Rearing and Spawning Zebrafish—Adult Fish were purchased fromAquarium Center (Baltimore, MD) and kept at 28.5 °C in a constantflow-through system. Embryos were obtained by spontaneous spawn-ing, maintained at 28.5 °C, and staged as described (35). Staged em-bryos were immediately snap-frozen and stored at �80 °C for futureuse.
Culture of Zebrafish Cell Lines, zfl—Cells were grown at 28 °C inL-15 medium supplemented with 10% fetal calf serum but withoutsodium bicarbonate.
Reverse Transcription and PCR Amplification Reactions—One vol-ume of fresh tissue, embryos, or harvested cells was homogenized in 8volumes of Trizol (Invitrogen), essentially as described (36). For PCR, 1�g of total RNA was reverse-transcribed using Superscript II reversetranscriptase (Invitrogen) and oligo(dT)18 in accordance with the man-ufacturer’s instructions. Amplification reactions (primers as indicated)were performed under standard conditions for either Pfu (Stratagene)or Taq (Fisher) DNA polymerases with 5 cycles of annealing at 58 °C,
followed by 35 cycles of annealing at 68 °C. Extension for all cycles wasallowed for 1 min at 72 °C. Whenever purified PCR products weretransferred into plasmids, insertions were sequenced in both orienta-tions to ensure that no errors had been introduced due to amplification.
For expression studies, DNA-contamination controls included reac-tions treated identically but lacking reverse transcriptase, as well asreactions lacking template. The products were resolved by TAE-agaroseelectrophoresis and recorded in a fluorimager (Amersham Biosciences).
Generation of cDNAs Encoding Zebrafish eIF4E Family MemberseIF4E-1A and eIF4E-1B—A partial cDNA (EST AA542678) cloned intothe vector pSPORT1 (Invitrogen) was purchased from Genome SystemsInc. and found to contain residues equivalent to all but the initiationcodon and 5�-UTR of eIF4E-1A mRNA. From this partial cDNA andoverlapping sequence information from the EST data base, a cDNAfragment encoding the complete eIF4E-1A protein was generated byPCR amplification using Pfu polymerase, the sense primerz4E1ANcoI5–23(F), named for the corresponding nucleotides followingthe ATG of eIF4E-1A, and the antisense primer z4E1ABamHI927–906(R), named by the same convention. Similarly, a cDNA fragmentencoding eIF4E-1B was generated from the plasmid pBKSII(�)-zeIF4E-L (2), using the sense primer z4E1BNcoI5–42(F) and the anti-sense primer z4E1BBamHI653–632(R). In either case, the purifiedPCR product was digested with NcoI and BamHI and transferred intothe vector pCITE4a(�) (Novagen).
Generation of cDNA Encoding Zebrafish 4E-BP—A DNA fragmentencoding zebrafish 4E-BP was isolated following amplification by RT-PCR using Taq DNA polymerase. Primers for amplification were basedon the nucleotide alignment of multiple partial sequences encodingzebrafish 4E-BP in the EST data bank, forward primer z4EBPNdeI3–24(F) and reverse primer z4EBPBamHI339–319(R). PCR productswere transferred into the vector pGEM-T-easy (Promega) generatingpGEMTEz4EBP. Based upon consensus sequences from alignment ofmultiple ESTs, no errors had been introduced during amplification. Theinsert was transferred into pCITE4a(�) following digestion with NdeIand BamHI generating pCITE4az4EBP.
Generation of cDNA Encoding the eIF4E-binding Domain of HumaneIF4GI—The 1366-bp ApaI/BamHI fragment from plasmidpBSKII(�)HFC1 (a gift from the laboratory of Dr. R. E. Rhoads) wasmodified by addition of the sequence CATATGTCCATGG, 5� of theintact ApaI site such that the ATG of both the NdeI and NcoI sitesintroduced were in-frame with the eIF4GI coding sequence. Followingthe modification, the NdeI/BamHI fragment was transferred into the NdeIand BamHI sites within pCITE4a(�), generating pCITE4ah4GI4EBDHis.This encodes a protein containing amino acids 159–614 of eIF4GI fused toa C-terminal His tag and containing the eIF4E-binding domain.
Use of eIF4E-deficient Yeast Strain (JOS003)—Generation of a G418-resistant S. cerevisiae strain, JOS003, lacking an endogenous func-tional yeast eif4e gene is described elsewhere (37). In essence, theendogenous yeast eif4e gene was replaced by homologous recombinationwith a linear DNA fragment containing the G418 resistance cassetteKanMX4 (37, 38). This strain is able to survive due to the expression ofLeu-selectable, extrachromosomal, human eIF4E-1, driven by the glu-cose-repressible, galactose-dependent GAL1 promoter. Medium lackinggalactose and containing glucose suppresses growth of this strain, al-lowing complementation tests to be performed. All yeast cells weregrown at 30 °C in either YP medium (1% yeast extract, 2% bactopep-tone) or synthetic complete (SC) medium lacking only leucine or bothleucine and uracil, as necessary. Medium was supplemented with either2% D-galactose or 2% D-glucose (dextrose) as carbon source and 200�g/ml G418 for continuous selection of cells lacking the yeast eif4e gene.
Construction of cDNAs Encoding Variants of eIF4E-1A and eIF4E-1BLacking the Regions N-terminal to the Conserved Cores—cDNAs encod-ing variants of eIF4E-1A and eIF4E-1B, which lack the regions N-terminal to the conserved cores (eIF4E-1A-(�1–33), and eIF4E-1B-(�1–32), respectively), were generated by PCR amplification using theprimers z4E1ANcoI98–118(F) and z4E1ABamHI927–906(R) orz4E1BNcoI95–118(F) and z4E1BamHI653–632(R), respectively. NcoI-BamHI fragments were subsequently transferred into the plasmidpCITE4a.
Construction of cDNAs Encoding Variants of eIF4E-1A-(�1–33) andeIF4E-1B-(�1–32) Fused to the N-terminal Residues of eIF4E-1B andeIF4E-1A, Respectively—DNA fragments encoding for the N-terminalregion of eIF4E-1B-(�33–214) or sequence lacking the potential to codefor this region (�1–32) were generated by PCR from eIF4E-1B cDNAusing the primers z4E1BNcoI5–23(F) and z4E1BPstI105–84(R) andz4E1BPstI112–132(F) and z4E1BBamHI653–632(R), respectively. ThePCR product encoding the N-terminal region of eIF4E-1B (eIF4E-1B-(�33–214)) was transferred into NcoI-PstI digested pCITE4aeIF4E-1A
Two eIF4Es in Zebrafish 10533
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

to generate a cDNA encoding a protein corresponding to the conservedcore of eIF4E-1A fused to the N-terminal region of eIF4E-1B, eIF4E-1B-(�33–214)-eIF4E-1A-(�1–33). The PCR product lacking the poten-tial to code for the N-terminal region of eIF4E-1B (eIF4E-1B-(�1–32))was transferred into PstI-BamHI digested pCITE4aeIF4E-1A in orderto generate a cDNA corresponding to the conserved core of eIF4E-1Bfused to the N-terminal region of eIF4E-1A, eIF4E-1A-(�34–215)-eIF4E-1B-(�1–32).
eIF4E Complementation Assay in S. cerevisiae Conditionally LackingFunctional eIF4E—Initially, NcoI-BamHI fragments frompCITE4aeIF4E constructs were transferred into the Escherichia coliexpression vector pET11d. XbaI-BamHI fragments derived from theresulting pET11deIF4E constructs were subsequently transferred intothe Ura-selectable yeast expression vector pRS416GPD (39), whichallows expression under the control of the constitutive yeast glyceral-dehyde-3-phosphate dehydrogenase promoter. JOS003 transfected withpRS416GPDeIF4E constructs were selected on SCGal medium contain-ing G418 and lacking Leu and Ura. Complementation tests were per-formed either by streaking or plating �1,000 cells from a log-phaseculture on YP-medium containing G418 and either galactose or dex-trose. Plates were incubated at 30 °C for 3 days, and growth wasassessed visually by colony formation. Growth on plates containingdextrose indicates the ability of an ectopic eif4e gene to complementeIF4E deficiency.
Preparation of Protein Extracts from Cultured Cells or Tissues—Ingeneral, cultured cells or tissues were homogenized in 10 volumes ofice-cold buffer containing 25 mM Tris-HCl, pH 7.4, 1 mM EDTA, 2 mM
EGTA, 1 mM dithiothreitol, 100 mM KCl, 0.5% Elugent, and CompleteTM
Protease Inhibitors (Roche Applied Science). Homogenates were clari-fied by centrifugation (10,000 � g, 4 °C, for 10 min). Supernatants (2–5mg/ml protein) were frozen in liquid N2 and stored at �80 °Cuntil used.
Preparation of Recombinant eIF4E and 4E-BP from E. coli—Therelevant pET11deIF4E constructs were transfected into BL21(DE3)-pLysS cells and expressed as described (40). 500-ml cultures, derivedfrom the relevant cells grown in LB medium, were induced at 0.5–0.6A600 with 1 mM isopropyl-1-thio-�-D-galactopyranoside for 30 min, fol-lowed by further 1.5 h of induction in the presence of 0.2 �g/ml rifam-picin. The recombinant proteins were recovered from inclusion bodies,essentially by the method of Rudolph and co-workers (41, 42) usingsolubilization in guanidine-HCl, followed by staged dialyses. His-tagged4E-BP was purified from E. coli cell extracts by affinity chromatographyusing HisBind (Novagen).
In Vitro Expression and Protein/Protein Interaction Assays—Forproduction of 35S-radiolabeled proteins translated in vitro, the relevantpCITE4a constructs were used as templates in the rabbit reticulocyte-coupled transcription-translation system, containing [35S]Met (0.25mCi/ml), essentially as described previously (43). For interaction as-says, the fragment of human eIF4GI containing the eIF4E-bindingdomain from pCITE4ah4GI4EBDHis or zebrafish 4E-BP was co-trans-lated with either S-tagged eIF4E-1A or eIF4E-1B in 50-�l reactions for60 min at 30 °C. Reactions were diluted with 10 volumes of S-binding/washing buffer (Novagen) and incubated with 15 �l of S-protein-agarose (Novagen) for 45 min at 20 °C. S-protein-agarose beads wererecovered by centrifugation and washed 10 times with buffer (1 mleach), prior to elution with SDS-PAGE sample buffer. Samples of frac-tions, equivalent to 2 �l of the initial translation reactions, were ana-lyzed by SDS-PAGE and autoradiography.
For the interaction of zebrafish ovary eIF4Es with His-tagged 4E-BP,anti-His/protein G-Sepharose beads were prepared by binding anti-His5
antibody (Qiagen) to protein G-Sepharose beads in binding buffer con-taining 300 mM NaCl, 25 mM KCl, 20 mM HEPES-KOH, pH 7.2, 2 mM
EGTA, 10% glycerol, 1 mg/ml soybean trypsin inhibitor, 200 �g/mlbovine serum albumin, at 4 °C, with shaking, overnight. The beadswere washed 6 times in binding buffer and incubated with His-tagged4E-BP for 2 h at 4 °C. Binding of His-tagged 4E-BP was confirmed byWestern analysis of bound material, using INDIA-HisProbe-HRP(Pierce) and ECL for visualization of bead-bound His-tagged protein.Recombinant eIF4E-1A and eIF4E-1B or ovary extracts (pooled from 10fish) were incubated with His-tagged human 4E-BP bound to anti-His/protein G-Sepharose beads for 2 h at 4 °C. After 3 washes in bindingbuffer and 3 washes in low salt binding buffer containing 25 mM KCl, 20mM HEPES-KOH, pH 7.2, 2 mM EGTA, 10% glycerol, 200 �g/ml bovineserum albumin, bound proteins were eluted in 100 mM glycine, pH 2.5,and analyzed for the presence of eIF4E-1A and -1B by high-Tris SDS-PAGE and immunoblotting.
m7GTP-affinity Chromatography—Ovary extracts (from one fish) orin vitro translation reactions (50 �l) were bound to 50 �l of m7GTP-
Sepharose equilibrated with column buffer containing 25 mM HEPES-KOH, pH 7.2, 10% glycerol, 100 mM KCl, 0.1 mM EDTA, and 1 mM
dithiothreitol. The resin was washed extensively with 50 volumes of thesame buffer containing 200 �M GTP, prior to elution with buffer con-taining 200 �M m7GTP. Equivalent volumes of fractions representingeither 10 �l of ovary extract or 2 �l of translation reaction were used forfurther analyses. For chromatography of proteins produced from E. coli,50 ml of buffer containing 1.6 �g/ml each of recombinant eIF4E-1A andeIF4E-1B and 100 �g/ml total E. coli proteins was applied to 0.5 ml ofm7GTP-Sepharose resin. The resin was washed with 100 volumes ofbuffer prior to the collection of 0.5-ml fractions from progressive elu-tions with 5 ml of the buffers as indicated. Samples from fractions,equivalent to 3 �l of the applied material, were used for SDS-PAGE andimmunoblotting analyses.
SDS-PAGE—For separation of different eIF4Es, samples were elec-trophoresed on 17.5% polyacrylamide/high-Tris for 1600 V-h for in-creased resolution. All buffers and solutions were essentially asdescribed (44).
VSIEF—The isoelectric focusing method of O’Farrell (45), modifiedfor use in vertical slab gels, was used essentially as described (46) butwith modifications to the Pharmalyte composition and buffers used togive optimal resolution of eIF4E-1A and eIF4E-1B. For eIF4E-1A, sam-ples were focused through pH gradients of 5 to 8.5, using Pharmalytes3–10 in the presence of 9 M urea, 5% v/v �-mercaptoethanol, 2% w/vCHAPS (Fluka) with 25 mM NaOH at the anode and 25 mM asparticacid at the cathode. For eIF4E-1B, samples were focused through pHgradients of 8–10, using a mixture of Pharmalytes 3-10 and 9-11 (4:1),with 25 mM NaOH at the anode and 25 mM glutamic acid at the cathode.Samples were electrophoresed at 2 mA/gel for 20 h prior to immuno-blotting. pH strips were used to verify the pH gradient of the gels aftereach run.
Immunoblotting—Proteins were electrotransferred from gels toPVDF membranes and probed with rabbit anti-TATKSGSTTKNRFVV(gift from Dr. Simon Morley) at 60 V for 1.25 h for SDS-PAGE and 1 hfor VSIEF gels, prior to visualization by enhanced chemiluminescence,as described (47).
RESULTS
Cloning and Sequence Characterization of a cDNAEncoding a New eIF4E-related Protein from Zebrafish
An EST clone (GenBankTM accession number AA542678),generated by the Washington University zebrafish genome pro-ject, was identified by BLAST search (48) for clones encodingproteins similar to human eIF4E-1. This clone was purchasedfrom Genome Systems Inc. and represents a cDNA with codingpotential for a protein 82% identical to human eIF4E-1 and84% identical to Xenopus eIF4E-1. From this partial cDNA andoverlapping sequence information from the NCBI data bases, acDNA fragment containing a complete eIF4E-1-related proteincoding sequence was generated by PCR amplification.2 Se-quence comparison of this cDNA (GenBankTM accession num-ber AF257519) with that of the cDNA encoding eIF4E(L) (Gen-BankTM accession number AF176317) (3), showed that the twoare distinct, having considerable differences in the length andsequence of their 3�- UTR (data not shown). The close sequencerelationship between these cDNAs suggests that the two genesencoding these zebrafish eIF4E homologues share a commonorigin. Based on the closer sequence similarity of AF257519 tothe human and Xenopus eIF4E-1 cDNAs, we termed the pre-dicted protein encoded by AF257519 as eIF4E-1A, and werenamed the less similar predicted protein, formerly termedeIF4E(L), eIF4E-1B.
The deduced amino acid sequence of zebrafish eIF4E-1A isaligned with those of zebrafish eIF4E-1B, human eIF4E-1, andXenopus eIF4E-1 in Fig. 1, with predicted biochemical charac-teristics given in Table I. Zebrafish eIF4E-1A and eIF4E-1Bare �80% identical to each other in their core sequences (Fig. 1)and share extensive homology over the core sequences with
2 J. Robalino, B. Joshi, and R. Jagus, GenBankTM accessionnumber AF257519.
Two eIF4Es in Zebrafish10534
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

human and Xenopus eIF4E-1. Over the entire protein,eIF4E-1A shows 82% identity and 91% similarity to humaneIF4E-1, whereas eIF4E-1B shows 66% identity and 77% sim-ilarity. Higher sequence identities are seen in the conservedcore (equivalent to residues 36–200 in human eIF4E-1), withgreater sequence diversity in the N termini. Over the conservedcore, eIF4E-1A shows 89% identity and 96% similarity to hu-man eIF4E-1, whereas eIF4E-1B shows 75% identity and 87%similarity. Both proteins contain the eight tryptophan residuesconserved in all eIF4E-1 sequences known. In addition, all theresidues recognized as essential for cap binding activity (16,19), those equivalent to Trp-56, Trp-102, and Trp-166 of humaneIF4E-1 and the conserved Glu at a position equivalent toGlu-103, are strictly conserved in both zebrafish eIF4Es (high-lighted in red in Fig. 1), suggesting that both are good candi-dates to function as eIF4E. The tryptophan residue equivalentto Trp-73 in human eIF4E-1, which is required for interactionwith eIF4G and 4E-BP (22), is found at the correspondingpositions in both zebrafish eIF4Es within the conserved regioninvolved in this interaction (highlighted in dark purple in Fig.1). Similarly, all residues known to make contacts with theeIF4E-binding domains of eIF4Gs and 4E-BPs, those equiva-lent to His-37, Pro-38, Val-69, Trp-73, Leu-131, Glu-132, andLeu-135 in mouse eIF4E-1 (22), are present in both proteins.Like all other eIF4E sequences characterized so far, the Ntermini of the zebrafish eIF4Es are quite divergent, both fromeach other and from eIF4Es from other species. Like prototyp-ical eIF4E-1s, zebrafish eIF4E-1A has an acidic N-terminalregion, contributing to its predicted acidic isoelectric point (5.5)(Table I). The molecular weight of eIF4E-1A (24.8) is also ingood agreement with those of human and Xenopus eIF4Es(50–52). The predicted molecular weight eIF4E-1B is 24.7.However, eIF4E-1B is unusual in having a lysine-rich region(PKKKVEKKK) N-terminal to the conserved core, which con-tributes to its overall basic predicted isoelectric point (9.39).Short sequences rich in basic amino acids are characteristic ofnuclear localization sequences, such as that for SV40 T-anti-gen, although specific consensus sequences have not been for-mulated (53, 54). This opens up the possibility that the N-terminal region of zebrafish eIF4E-1B could function inthis capacity.
Analysis of the Expression Patterns ofeIF4E-1A and eIF4E-1B
Expression in Different Tissues—To assess whethereIF4E-1A and eIF4E-1B represent eIF4E family members thatare actually expressed in zebrafish, we looked for the tran-scripts of each in a variety of tissues using RT-PCR (Fig. 2). Arepresentative experiment in which equal amounts of RNAfrom each tissue were subjected to oligo(dT)-primed reversetranscription, followed by PCR with gene-specific primers, isshown in Fig. 2A. mRNA for eIF4E-1A was detected in alltissues examined, whereas eIF4E-1B mRNA was detectableonly in ovary and muscle.
The suggestion of cell type-specific expression of eIF4E-1Aand -1B observed by RT-PCR analysis was re-evaluated in avariety of tissues at the protein level. Although resolution ofpurified eIF4E-1A and eIF4E-1B could be achieved by high-Tris SDS-PAGE, for the analysis of crude extracts we foundsuperior separation of the two forms by taking advantage oftheir differing isoelectric points. Following vertical slab isoelec-tric focusing (VSIEF), proteins from adult tissues were electro-transferred to a membrane and subjected to immunoblotting byusing an antibody raised against a peptide corresponding to theC-terminal sequence of human eIF4E-1, TATKSGSTTKNR-FVV (Fig. 2B). The C terminus of eIF4E-1A differs by oneresidue from this sequence, whereas eIF4E-1B differs at threepositions from the human-derived peptide. Nevertheless, both
TABLE IPhysical characteristics of zebrafish eIF4Es
FIG. 1. Multiple alignment of predicted amino acid sequences of zebrafish eIF4Es, human eIF4E-1, and Xenopus eIF4E-1. Thepredicted amino acid sequences of eIF4E-1A and eIF4E-1B were aligned with those of human eIF4E-1 and X. laevis eIF4E-1. The period indicatesidentity at that position, relative to human eIF4E-1. A dash represents a gap in the sequence. The residues within the core region are indicatedin black. Variations relative to human eIF4E-1 are identified by the symbol of the respective residue at each position. Residues in red are essentialfor cap binding activity. Residues in blue represent the region N-terminal to the conserved core. Residues in purple represent the conserved regionsurrounding the tryptophan (Trp-73 in human, highlighted in dark purple) that is essential for interaction with eIF4G and 4E-BPs (22). Residuesin green define the sequence of the peptide used to raise the anti-eIF4E antibody used in this investigation. Highlighted in dark green is Ser-209from human eIF4E-1, the site of phosphorylation by cellular kinases (27).
Two eIF4Es in Zebrafish 10535
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

recombinant eIF4E-1A and eIF4E-1B interact with the anti-body. As expected, recombinant eIF4E-1A focused at a pHbetween 5 and 7, whereas recombinant eIF4E-1B focused be-tween pH 9 and pH 10. eIF4E-1A was detected in all tissuesexamined. eIF4E-1B on the other hand, was detected only inovary, testes, and muscle, consistent with the RNA expressiondata. Ovary cell fractionation studies indicate that botheIF4E-1A and -1B are found in both nucleus and cytoplasm(results not shown).
Developmental Expression—We further analyzed the expres-sion characteristics of both zebrafish eIF4Es, by assaying RNAfrom embryos at different developmental stages (Fig. 3). RT-PCR was performed under conditions similar to those used forthe tissue distribution assay. eIF4E-1A mRNA was detected atall stages of zebrafish development examined, whereaseIF4E-1B mRNA was detected only in stages that precedePrim6 (24 h post-fertilization). The analyses of the expressionpatterns of the eIF4E family members suggest that eIF4E-1Ais ubiquitously expressed, as expected for an essential transla-tion factor. However, eIF4E-1B appears non-essential for cellviability, given that neither its mRNA nor its protein accumu-late in many cell types, including an actively replicating ze-brafish liver cell line, zfl (see Fig. 2).
eIF4E-1A, but Not eIF4E-1B, Is Functionally Equivalentto Human eIF4E-1
Assessment of Functionality of eIF4E-1A and eIF4E-1B byComplementation in S. cerevisiae Conditionally Deficient ineIF4E—Although there is considerable sequence divergencebetween human eIF4E-1 and S. cerevisiae eIF4E (31% iden-
FIG. 2. Expression of zebrafish eIF4E-1A and eIF4E-1B in different tissues. A, expression of RNA. Total RNA (1 �g) from samplesof the indicated tissue was reverse-transcribed and used as template for PCR, with primers specific for either eIF4E-1A or eIF4E-1B cDNAs.Amplicons were resolved in TAE-agarose gels and stained with ethidium bromide. The size of the expected amplicons obtained by usingeIF4E-1A or eIF4E-1B cDNAs as templates is indicated to the right of each panel. The target of each pair of primers used for the reactionsis indicated to the left of each panel. B, analysis of eIF4E-1A and eIF4E-1B expression in different cell types by isoelectric focusing/immunoblotting analysis. Recombinant eIF4E-1A and eIF4E-1B, and extracts from adult zebrafish tissues and zfl cells, were subjected tovertical slab gel isoelectric focusing and immunoblotting, essentially as described (46). Samples were focused in the presence of 9 M urea, 5%v/v �-mercaptoethanol, 2% w/v CHAPS (Fluka). In the top panel, a pH range of 5–8.5, using ampholytes 3–10 with 50 mM NaOH at the anodeand 20 mM aspartic acid at the cathode, was used. In the bottom panel, samples were focused through a pH range of 8–10, using a mixtureof ampholytes 3–10 and 9–11 (4:1), with 25 mM NaOH at the anode and 25 mM glutamic acid at the cathode. Proteins were electrophoresedat 2 mA/gel for 20 h. Proteins were transferred to PVDF membranes and probed with anti-TATKSGSTTKNRFVV. Recombinant humaneIF4E-1 was included as a marker of pI 6.1 (only seen in top panel).
FIG. 3. RT-PCR analysis of zebrafish eIF4E-1A and eIF4E-1BmRNAs during development. Total RNA (1 �g) from samples of theindicated embryonic stage was reverse-transcribed and used as tem-plate for PCR, with primers specific for either eIF4E-1A or eIF4E-1B.Amplicons were resolved in TAE-agarose gels and stained withethidium bromide. The size of the expected amplicons obtained by usingeIF4E-1A or eIF4E-1B cDNAs as templates is indicated to the right ofeach panel. The target of each pair of primers used for the reactions isindicated to the left of each panel.
Two eIF4Es in Zebrafish10536
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from
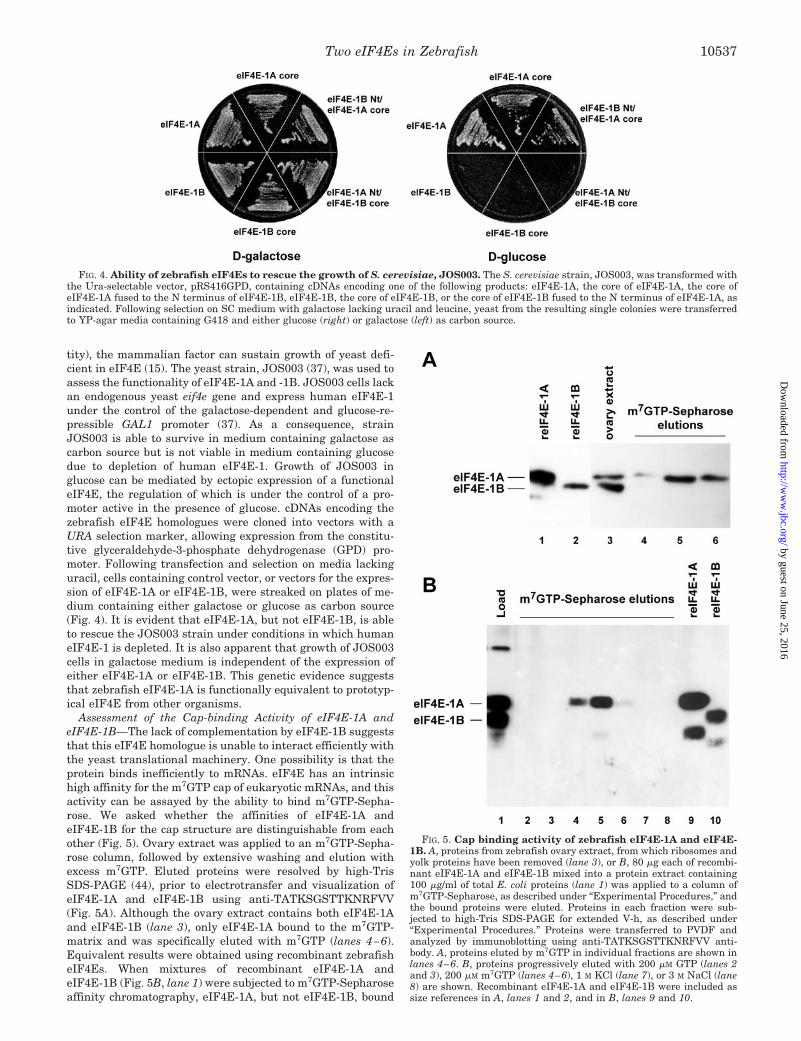
tity), the mammalian factor can sustain growth of yeast defi-cient in eIF4E (15). The yeast strain, JOS003 (37), was used toassess the functionality of eIF4E-1A and -1B. JOS003 cells lackan endogenous yeast eif4e gene and express human eIF4E-1under the control of the galactose-dependent and glucose-re-pressible GAL1 promoter (37). As a consequence, strainJOS003 is able to survive in medium containing galactose ascarbon source but is not viable in medium containing glucosedue to depletion of human eIF4E-1. Growth of JOS003 inglucose can be mediated by ectopic expression of a functionaleIF4E, the regulation of which is under the control of a pro-moter active in the presence of glucose. cDNAs encoding thezebrafish eIF4E homologues were cloned into vectors with aURA selection marker, allowing expression from the constitu-tive glyceraldehyde-3-phosphate dehydrogenase (GPD) pro-moter. Following transfection and selection on media lackinguracil, cells containing control vector, or vectors for the expres-sion of eIF4E-1A or eIF4E-1B, were streaked on plates of me-dium containing either galactose or glucose as carbon source(Fig. 4). It is evident that eIF4E-1A, but not eIF4E-1B, is ableto rescue the JOS003 strain under conditions in which humaneIF4E-1 is depleted. It is also apparent that growth of JOS003cells in galactose medium is independent of the expression ofeither eIF4E-1A or eIF4E-1B. This genetic evidence suggeststhat zebrafish eIF4E-1A is functionally equivalent to prototyp-ical eIF4E from other organisms.
Assessment of the Cap-binding Activity of eIF4E-1A andeIF4E-1B—The lack of complementation by eIF4E-1B suggeststhat this eIF4E homologue is unable to interact efficiently withthe yeast translational machinery. One possibility is that theprotein binds inefficiently to mRNAs. eIF4E has an intrinsichigh affinity for the m7GTP cap of eukaryotic mRNAs, and thisactivity can be assayed by the ability to bind m7GTP-Sepha-rose. We asked whether the affinities of eIF4E-1A andeIF4E-1B for the cap structure are distinguishable from eachother (Fig. 5). Ovary extract was applied to an m7GTP-Sepha-rose column, followed by extensive washing and elution withexcess m7GTP. Eluted proteins were resolved by high-TrisSDS-PAGE (44), prior to electrotransfer and visualization ofeIF4E-1A and eIF4E-1B using anti-TATKSGSTTKNRFVV(Fig. 5A). Although the ovary extract contains both eIF4E-1Aand eIF4E-1B (lane 3), only eIF4E-1A bound to the m7GTP-matrix and was specifically eluted with m7GTP (lanes 4–6).Equivalent results were obtained using recombinant zebrafisheIF4Es. When mixtures of recombinant eIF4E-1A andeIF4E-1B (Fig. 5B, lane 1) were subjected to m7GTP-Sepharoseaffinity chromatography, eIF4E-1A, but not eIF4E-1B, bound
FIG. 4. Ability of zebrafish eIF4Es to rescue the growth of S. cerevisiae, JOS003. The S. cerevisiae strain, JOS003, was transformed withthe Ura-selectable vector, pRS416GPD, containing cDNAs encoding one of the following products: eIF4E-1A, the core of eIF4E-1A, the core ofeIF4E-1A fused to the N terminus of eIF4E-1B, eIF4E-1B, the core of eIF4E-1B, or the core of eIF4E-1B fused to the N terminus of eIF4E-1A, asindicated. Following selection on SC medium with galactose lacking uracil and leucine, yeast from the resulting single colonies were transferredto YP-agar media containing G418 and either glucose (right) or galactose (left) as carbon source.
FIG. 5. Cap binding activity of zebrafish eIF4E-1A and eIF4E-1B. A, proteins from zebrafish ovary extract, from which ribosomes andyolk proteins have been removed (lane 3), or B, 80 �g each of recombi-nant eIF4E-1A and eIF4E-1B mixed into a protein extract containing100 �g/ml of total E. coli proteins (lane 1) was applied to a column ofm7GTP-Sepharose, as described under “Experimental Procedures,” andthe bound proteins were eluted. Proteins in each fraction were sub-jected to high-Tris SDS-PAGE for extended V-h, as described under“Experimental Procedures.” Proteins were transferred to PVDF andanalyzed by immunoblotting using anti-TATKSGSTTKNRFVV anti-body. A, proteins eluted by m7GTP in individual fractions are shown inlanes 4–6. B, proteins progressively eluted with 200 �M GTP (lanes 2and 3), 200 �M m7GTP (lanes 4–6), 1 M KCl (lane 7), or 3 M NaCl (lane8) are shown. Recombinant eIF4E-1A and eIF4E-1B were included assize references in A, lanes 1 and 2, and in B, lanes 9 and 10.
Two eIF4Es in Zebrafish 10537
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

to the column and was specifically eluted with m7GTP (lanes4–6). Analyses using in vitro transcribed and translated 35S-eIF4E-1A and 35S-eIF4E1B gave similar results (data notshown). These data demonstrate that eIF4E-1A is a bona fidecap-binding protein and further suggest that eIF4E-1B is notfunctionally equivalent to human eIF4E-1.
eIF4E-1A, but Not eIF4E-1B, Interacts in Vitro with HumaneIF4GI—The finding that eIF4E-1B is deficient in cap bindingactivity prompted us to consider that a possible function ofeIF4E-1B is as a tissue-specific translational repressor. Wehypothesized that eIF4E-1B could compete in vivo witheIF4E-1A for binding to eIF4GI. An interaction betweeneIF4E-1B and eIF4GI would preclude eIF4E-1A from incorpo-rating into functional eIF4F complex, thus inhibiting cap-de-pendent translation. A mechanism analogous to this has beencharacterized in mammals; the 4E-BP family of translationalrepressors compete with cellular eIF4G for a common bindingsite on eIF4E-1 (22, 24, 33, 34). eIF4E-1B could therefore beinvolved in a novel mechanism for control of translation bymimicking the eIF4G binding activity of eIF4E. We tested thishypothesis by asking if eIF4E-1B could interact with theeIF4E-1-binding domain of a vertebrate eIF4G homologue.
A polypeptide corresponding to residues 159–614 of humaneIF4GI (molecular mass �45 kDa), which contained theeIF4E-1 interaction domain, was co-translated with S-taggedvariants of either eIF4E-1A or eIF4E-1B in a reticulocyte cell-free translation system in the presence of [35S]Met (Fig. 6,lanes 1 and 5). After synthesis, reactions were incubated withS-protein-agarose. Following extensive washing, proteinsbound to the matrix were eluted with SDS-PAGE sample buffer(lanes 3 and 4 and lanes 7 and 8). Fractions were resolved by
high-Tris SDS-PAGE and analyzed by autoradiography.Whereas the eIF4GI polypeptide co-purified with eIF4E-1A,eIF4E-1B failed to interact with the same polypeptide at de-tectable levels.
These data, coupled with the yeast complementation data,demonstrate that like human eIF4E-1, zebrafish eIF4E-1A isable to interact with human eIF4GI in vitro and yeast eIF4G invivo. In contrast, eIF4E-1B has a low affinity for eIF4GI andthus is unlikely to function as an efficient in vivo competitor ofeIF4E-1A. This was corroborated by the finding that neitherthe addition of eIF4E-1A nor of eIF4E-1B at concentrations upto 5-fold that of the endogenous rabbit eIF4E-1 protein pro-duced any significant effect on the kinetics of protein synthesisin the reticulocyte translation system (data not shown).
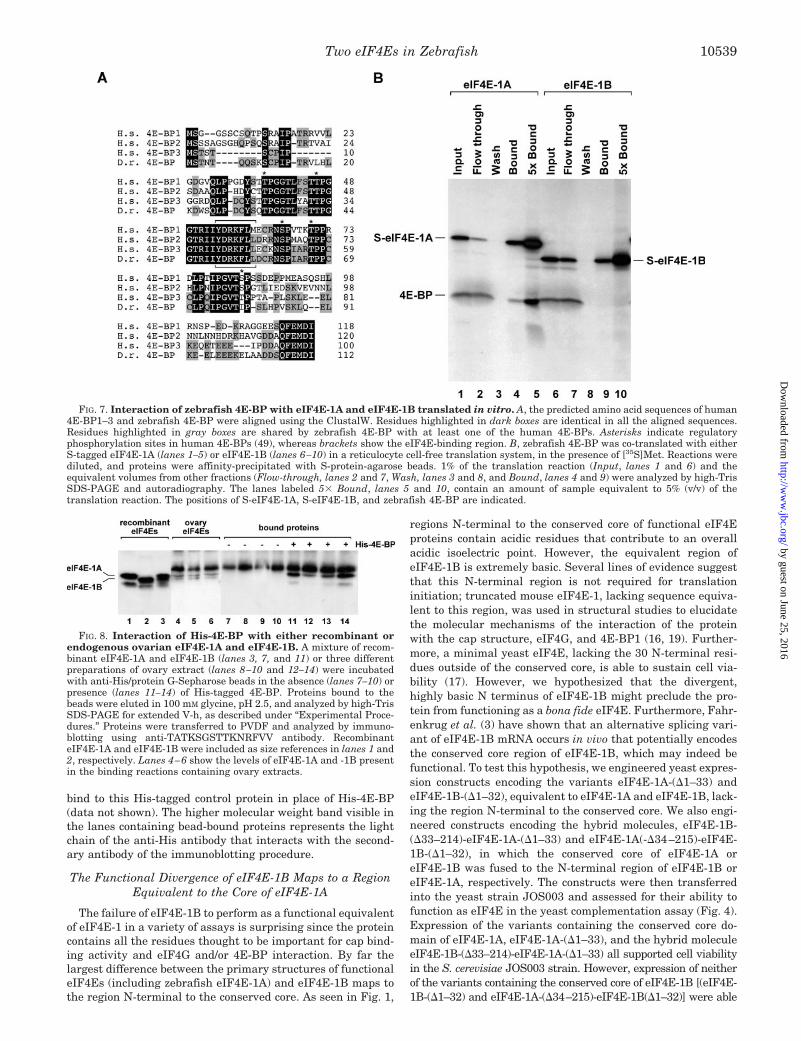
eIF4E-1A, but Not eIF4E-1B, Interacts with Zebrafish 4E-BP—Since eIF4Gs and 4E-BPs bind to eIF4E through commonmotifs, we anticipated that zebrafish eIF4E-1A, but not eIF4E-1B, would be targeted by the 4E-BP repressors. However, thereremained the possibility that a homologue of eIF4E that isdeficient in both cap binding activity and eIF4G interactioncould potentially bind to 4E-BPs and work as a translationalde-repressor.
By using RT-PCR, programmed with primers based on ze-brafish ESTs encoding a 4E-BP, we amplified and cloned azebrafish cDNA encoding a protein of 117 amino acids. Analignment of the predicted protein encoded by the zebrafishcDNA with human 4E-BP1–3, is presented in Fig. 7A. The closesequence similarity and conservation of the eIF4E-binding sitestrongly suggests that this cDNA (GenBankTM accession num-ber AF332983) encodes a member of the 4E-BP family that is68% identical to human 4E-BP3 (55)
To assess the functionality of the cloned zebrafish 4E-BP,and to explore the possibility that eIF4E-1B is a de-repressor of4E-BP-mediated inhibition of translation, we performed invitro interaction assays with S-tagged variants of eIF4E-1Aand eIF4E-1B in a manner analogous to the eIF4E/eIF4G in-teraction assay described above. After synthesis, reactionswere incubated with S-protein-agarose. Following extensivewashing, proteins bound to the matrix were eluted with SDS-PAGE sample buffer (Fig. 7B, lanes 4 and 5 and lanes 9 and 10).The data showed that zebrafish 4E-BP was enriched in thefraction of bound proteins in the presence of eIF4E-1A but noteIF4E-1B. These data demonstrate that the 4E-BP-mediatedtranslational repression pathway is conserved in zebrafish.However, eIF4E-1B does not bind to zebrafish 4E-BP with anaffinity that would be consistent with a role as a de-repressor of4E-BP-mediated inhibition of translation.
The failure of eIF4E-1B to interact with eIF4G or 4E-BPcould reflect the fact that the recombinant protein is simplymisfolded and therefore cannot interact with eIF4G or 4E-BP.To address this possibility, a co-purification approach usingHis-tagged 4E-BP was implemented to look at the interactionof endogenous eIF4E-1B from zebrafish ovary with 4E-BP. Amixture of recombinant eIF4E-1A and -1B or zebrafish ovaryextracts were incubated with His-tagged 4E-BP bound to anti-His/protein G-Sepharose beads (Fig. 8). The ovary extractscontained similar levels of eIF4E-1A and -1B (Fig. 8, lanes4–6). eIF4E-1A associated with the beads in a 4E-BP-depend-ent manner (Fig. 8, lanes 7–14). In contrast to eIF4E-1A, nei-ther recombinant eIF4E-1B nor eIF4E-1B from zebrafish ovarybound to His-tagged 4E-BP (Fig. 8, lanes 11–14), consistentwith the inability of in vitro translated eIF4E-1B to interactwith 4E-BP (Fig. 7). To ensure that eIF4E-1A was binding to4E-BP and not to the His tag in this assay, recombinant His-tagged growth hormone was used as a substitute for His-4E-BP1 in control experiments. As expected, eIF4E-1A did not
FIG. 6. Interaction of zebrafish eIF4E-1A and eIF4E-1B witheIF4G. A peptide derived from human eIF4GI (residues 159–614) wasco-translated with either S-tagged eIF4E-1A (lanes 1–4) or eIF4E-1B(lanes 5–8) in a reticulocyte cell-free translation system, in the presenceof [35S]Met. Reactions were diluted, and proteins were affinity-precip-itated with S-protein-agarose beads. 1% (v/v) of the translation reaction(Input, lanes 1 and 5) and equivalent volumes from the other fractions(Flow-through, lanes 2 and 6, and Bound, lanes 3 and 7) were analyzedby high-Tris SDS-PAGE and autoradiography. The lanes labeled 5�Bound (lanes 4 and 8) contain an amount of sample equivalent to 5%(v/v) of the translation reaction. The positions of S-eIF4E-1A, S-eIF4E-1B, and the human eIF4GI peptide are indicated.
Two eIF4Es in Zebrafish10538
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from
bind to this His-tagged control protein in place of His-4E-BP(data not shown). The higher molecular weight band visible inthe lanes containing bead-bound proteins represents the lightchain of the anti-His antibody that interacts with the second-ary antibody of the immunoblotting procedure.
The Functional Divergence of eIF4E-1B Maps to a RegionEquivalent to the Core of eIF4E-1A
The failure of eIF4E-1B to perform as a functional equivalentof eIF4E-1 in a variety of assays is surprising since the proteincontains all the residues thought to be important for cap bind-ing activity and eIF4G and/or 4E-BP interaction. By far thelargest difference between the primary structures of functionaleIF4Es (including zebrafish eIF4E-1A) and eIF4E-1B maps tothe region N-terminal to the conserved core. As seen in Fig. 1,
regions N-terminal to the conserved core of functional eIF4Eproteins contain acidic residues that contribute to an overallacidic isoelectric point. However, the equivalent region ofeIF4E-1B is extremely basic. Several lines of evidence suggestthat this N-terminal region is not required for translationinitiation; truncated mouse eIF4E-1, lacking sequence equiva-lent to this region, was used in structural studies to elucidatethe molecular mechanisms of the interaction of the proteinwith the cap structure, eIF4G, and 4E-BP1 (16, 19). Further-more, a minimal yeast eIF4E, lacking the 30 N-terminal resi-dues outside of the conserved core, is able to sustain cell via-bility (17). However, we hypothesized that the divergent,highly basic N terminus of eIF4E-1B might preclude the pro-tein from functioning as a bona fide eIF4E. Furthermore, Fahr-enkrug et al. (3) have shown that an alternative splicing vari-ant of eIF4E-1B mRNA occurs in vivo that potentially encodesthe conserved core region of eIF4E-1B, which may indeed befunctional. To test this hypothesis, we engineered yeast expres-sion constructs encoding the variants eIF4E-1A-(�1–33) andeIF4E-1B-(�1–32), equivalent to eIF4E-1A and eIF4E-1B, lack-ing the region N-terminal to the conserved core. We also engi-neered constructs encoding the hybrid molecules, eIF4E-1B-(�33–214)-eIF4E-1A-(�1–33) and eIF4E-1A(-�34–215)-eIF4E-1B-(�1–32), in which the conserved core of eIF4E-1A oreIF4E-1B was fused to the N-terminal region of eIF4E-1B oreIF4E-1A, respectively. The constructs were then transferredinto the yeast strain JOS003 and assessed for their ability tofunction as eIF4E in the yeast complementation assay (Fig. 4).Expression of the variants containing the conserved core do-main of eIF4E-1A, eIF4E-1A-(�1–33), and the hybrid moleculeeIF4E-1B-(�33–214)-eIF4E-1A-(�1–33) all supported cell viabilityin the S. cerevisiae JOS003 strain. However, expression of neitherof the variants containing the conserved core of eIF4E-1B [(eIF4E-1B-(�1–32) and eIF4E-1A-(�34–215)-eIF4E-1B(�1–32)] were able
FIG. 7. Interaction of zebrafish 4E-BP with eIF4E-1A and eIF4E-1B translated in vitro. A, the predicted amino acid sequences of human4E-BP1–3 and zebrafish 4E-BP were aligned using the ClustalW. Residues highlighted in dark boxes are identical in all the aligned sequences.Residues highlighted in gray boxes are shared by zebrafish 4E-BP with at least one of the human 4E-BPs. Asterisks indicate regulatoryphosphorylation sites in human 4E-BPs (49), whereas brackets show the eIF4E-binding region. B, zebrafish 4E-BP was co-translated with eitherS-tagged eIF4E-1A (lanes 1–5) or eIF4E-1B (lanes 6–10) in a reticulocyte cell-free translation system, in the presence of [35S]Met. Reactions werediluted, and proteins were affinity-precipitated with S-protein-agarose beads. 1% of the translation reaction (Input, lanes 1 and 6) and theequivalent volumes from other fractions (Flow-through, lanes 2 and 7, Wash, lanes 3 and 8, and Bound, lanes 4 and 9) were analyzed by high-TrisSDS-PAGE and autoradiography. The lanes labeled 5� Bound, lanes 5 and 10, contain an amount of sample equivalent to 5% (v/v) of thetranslation reaction. The positions of S-eIF4E-1A, S-eIF4E-1B, and zebrafish 4E-BP are indicated.
FIG. 8. Interaction of His-4E-BP with either recombinant orendogenous ovarian eIF4E-1A and eIF4E-1B. A mixture of recom-binant eIF4E-1A and eIF4E-1B (lanes 3, 7, and 11) or three differentpreparations of ovary extract (lanes 8–10 and 12–14) were incubatedwith anti-His/protein G-Sepharose beads in the absence (lanes 7–10) orpresence (lanes 11–14) of His-tagged 4E-BP. Proteins bound to thebeads were eluted in 100 mM glycine, pH 2.5, and analyzed by high-TrisSDS-PAGE for extended V-h, as described under “Experimental Proce-dures.” Proteins were transferred to PVDF and analyzed by immuno-blotting using anti-TATKSGSTTKNRFVV antibody. RecombinanteIF4E-1A and eIF4E-1B were included as size references in lanes 1 and2, respectively. Lanes 4–6 show the levels of eIF4E-1A and -1B presentin the binding reactions containing ovary extracts.
Two eIF4Es in Zebrafish 10539
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

to complement the lack of functional eIF4E, suggesting that sub-stitutions of residues within the conserved core region of eIF4E-1Brelative to that of eIF4E-1A are responsible for the failure ofeIF4E-1B to function. The ability of eIF4E-1A-(�1–33) to supportthe growth of yeast lacking a functional eif4e gene demonstratesthat 33 amino acids at the N terminus, representing the entireregion N-terminal to the conserved core, are not essential for func-tion. This is the most extensive N-terminal truncation product ofeIF4E-1 to date that shows biological activity.
DISCUSSION
Our studies have shown that zebrafish eIF4E-1A appears torepresent the functional equivalent of human eIF4E-1. In con-trast to this, a related form, eIF4E-1B, constitutes a previouslyunknown type of vertebrate eIF4E-1 with unexpected proper-ties and a restricted tissue distribution. The translation factor,eIF4E, functions to recruit mRNA to the ribosome by bindingsimultaneously to the 5� cap structure and to the scaffoldtranslation factor eIF4G. Prototypical eIF4E is an essentialtranslation factor; thus it is expressed in every cell type and isrequired for cell viability and cell cycle progression. With thisin mind, we propose that zebrafish eIF4E-1A is a suitablecandidate for eIF4E in this organism. Such an assignment isconsistent with the expression data, the functional assays, andthe ability of eIF4E-1A to support growth of the yeast strain,JOS003, conditionally deficient in eIF4E, in medium contain-ing glucose. In contrast to eIF4E-1A, the data presented hereshow that neither recombinant nor endogenous eIF4E-1B ex-hibits any detectable activity in standard assays of eIF4E func-tion. This is despite the fact that eIF4E-1B contains extensivesimilarity to known vertebrate eIF4Es and contains all aminoacid residues shown to be required for binding the mRNA-capstructure, eIF4G, and 4E-BPs. Furthermore, data from expres-sion analyses in zebrafish tissues, cultured cells, and at differ-ent stages of development suggest that the presence ofeIF4E-1B is not essential for survival.
The position and phase of the intron/exon junctions ineIF4E-1B closely resemble those described for human eIF4E-1(3, 56, 57). This, along with the high identity of the cDNAsequence of eIF4E-1B to eIF4E-1A, suggests that eIF4E-1B hasarisen via a gene duplication event that occurred during ver-tebrate evolution. Interestingly, the 5� end of zebrafisheIF4E-1B and mammalian eIF4E-1 transcripts are quite dis-tinct, and comparison of the mammalian eIF4E-1 promotersequences with zebrafish eIF4E-1B promoter reveals no obvi-ous conservation (2). It is possible that the gene duplicationevent that gave rise to eIF4E-1B excluded the 5� region of theancestral eif4e gene or that selective pressure is responsible forthe divergence in this region.
Although we have shown here that eIF4E-1B lacks three ofthe most prominent activities of eIF4E, cap binding, eIF4Gbinding, and 4E-BP binding, it remains to be addressedwhether eIF4E-1B can interact with other eIF4E-binding pro-teins (e.g. 4E-T, maskin) (58, 59). However, since all knownproteins that associate with eIF4E seem to share a commoneIF4E-binding domain, it seems unlikely that eIF4E-1B willinteract with them. The novel N terminus of eIF4E-1B, with itspotential nuclear localization sequence, may be indicative of itsability to undertake an alternate function such as RNA/ribo-nucleoprotein transport, RNA splicing, or localization ofmRNAs during specific developmental stages or differentiatedstates. We have observed, however, that in normal fish ovaryeIF4E-1B seems to be associated with both cytoplasmic andnuclear compartments, as assayed by cell fractionation (datanot shown). Future studies will be required to clarify the in-tracellular distribution of eIF4E-1A and eIF4E-1B in intactcells. The expression patterns of eIF4E1-B in fish embryos
suggest it may fulfill a specialized function in zebrafish germcells or during development/differentiation.
Recently, cDNAs or genes encoding proteins that exhibit dif-ferent degrees of identity to known prototypical eIF4E counter-parts have been identified from a number of organisms (1, 60, 62,63).3,4 Many possible examples of eIF4E-related proteins are stillrepresented only by cDNA sequences from ESTs, or as genomicsequences, and their possible functions have not been reported orinvestigated. Several of these eIF4E-related factors have beenfunctionally characterized, including the Schizosaccharomycespombe, eIF4E2 (65), the eIF4E family members in C. elegans thatinteract with trimethylated cap structures (1), and the eIF4E-like protein 4EHP/nCBP/IFE-4 (1, 60, 61). Some of these studiesdemonstrate that the multiplicity of proteins related to prototyp-ical eIF4E cannot simply be explained by functional redundancy.Although S. pombe eIF4E1 and eIF4E2 show almost identicalcap binding characteristics, eIF4E2 alone cannot support viabil-ity (65). Likewise, in C. elegans, RNA interference experimentshave shown that depletion of IFE-3 alone is lethal, suggestingthat the other four forms cannot fulfill the same function or donot exhibit the same expression pattern (1). Other studies haveshown that eIF4E-related proteins can fulfill specialized func-tions. Recognition of trimethylated cap structures by IFE-1, -2and -5 in C. elegans is to date the best example of specializedfunction for eIF4E-related proteins (1, 62, 64). This particularfunction is likely to reflect the specialized nature of nematodemRNAs, the majority of which have trimethylated cap structures(65). A role for vertebrate eIF4Es (including eIF4E-1B) in therecruitment of mRNAs with trimethylated cap structures seemsunlikely, since vertebrate mRNAs are not known to possess suchstructures. This possibility, however, has not been formallyexcluded.
Even though it is possible that the presence of multipleproteins related to eIF4E is the result of unproductive geneduplications giving rise to non-functional genes, the apparentconservation of at least some of these eIF4E-related factorssuggests they serve important functions. One such example is4EHP, conserved over vast evolutionary stretches from C. el-egans, Drosophila, plants, and mammals and represented inEST clones across the eukaryotic phyla. Likewise, ESTs encod-ing eIF4E-1B-like proteins have been identified in Oncorhyn-chus mykiss, Xenopus laevis, Silurana tropicalis, and Susscrofa (GenBankTM accession numbers BX303305, BQ398016,AL867687, and BF080360, respectively), and evidence for itspresence in humans can be found in the human genome se-quence. These observations, together with the data presentedhere in zebrafish, suggest that eIF4E-1B may have a role inall vertebrates.
The recognition of eIF4E-1A as the prototypical eIF4E inzebrafish will allow studies on the role of this regulatory trans-lation factor during vertebrate development more easily thancan be achieved in mouse. Similarly, the recognition ofeIF4E-1B as a form not functional in cap binding or eIF4G/4E-BP binding will provide unique opportunities for investigat-ing the relationship between structure and function in eIF4Efamily members. Furthermore, further studies of eIF4E-1Bshould allow the uncovering of additional roles for eIF4E fam-ily members.
Acknowledgments—We thank Dr. Simon Morley for the anti-TATKSGSTTKNRFVV. We also thank Joseph Garner and Eric Witzelfor help with reagents used and lively conversation.
3 B. Joshi, K. K. Norris, and R. Jagus, GenBankTM accession numberAF068117.
4 B. Joshi and R. Jagus, GenBankTM accession number AF068116.
Two eIF4Es in Zebrafish10540
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. Keiper, B. D., Lamphear, B. J., Deshpande, A. M., Jankowska-Anyszka, M.,Aamodt, E. J., Blumenthal, T., and Rhoads, R. E. (2000) J. Biol. Chem. 275,10590–10596
2. Fahrenkrug, S. C., Dahlquist, M. O., Clark, K., and Hackett, P. B. (1999)Differentiation 65, 191–201
3. Fahrenkrug, S. C., Joshi, B., Hackett, P. B., and Jagus, R. (2000) Differentia-tion 66, 15–22
4. Shuman, S. (2000) Prog. Nucleic Acids Res. Mol. Biol. 66, 1–405. Furuichi, Y., and Shatkin, A. J. (2000) Adv. Virus Res. 55, 135–1846. Gingras, A. C., Raught, B., and Sonenberg, N. (1999) Annu. Rev. Biochem. 8,
913–9637. McKendrick, L., Pain, V. M., and Morley, S. J. (1999) Int. J. Biochem. Cell Biol.
31, 31–358. Hershey, J. W. B., and Merrick, W. C. (2000) in Translational Control of Gene
Expression (Sonenberg, N., Hershey, J. W. B., and Mathews, M. B., eds) pp.33–88, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
9. Abramson, R. D., Dever, T. E., Lawson, T. G., Ray, B. K., Thach, R. E., andMerrick, W. C. (1987) J. Biol. Chem. 262, 3826–3834
10. Lorsch, J. R., and Herschlag, D. (1998) Biochemistry 37, 2194–2120611. Lamphear, B. J., Kirchweger, R., Skern, T., and Rhoads, R. E. (1995) J. Biol.
Chem. 270, 21975–2198312. Imataka, H., and Sonenberg, N. (1997) Mol. Cell. Biol. 17, 6940–694713. Hentze, M. W. (1997) Science 275, 500–50114. Keiper, B. D., Gan, W., and Rhoads, R. E. (1999) Int. J. Biochem. Cell Biol. 31,
37–4115. Altmann, M., Muller, P. P., Pelletier, J., Sonenberg, N., and Trachsel, H.
(1989) J. Biol. Chem. 264, 12145–1214716. Marcotrigiano, J., Gingras, A. C., Sonenberg, N., and Burley, S. K. (1997) Cell
89, 951–96117. Vasilescu, S., Ptushkina, M., Linz, B., Muller, P. P., and McCarthy, J. E. (1996)
J. Biol. Chem. 271, 7030–703718. Morino, S., Hazama, H., Ozaki, M., Teraoka, Y., Shibata, S., Doi, M., Ueda, H.,
Ishida, T., and Uesugi, S. (1996) Eur. J. Biochem. 239, 597–60119. Matsuo, H., Li, H., McGuire, A. M., Fletcher, C. M., Gingras, A. C., Sonenberg,
N., and Wagner, G. (1997) Nat. Struct. Biol. 4, 717–72420. Ueda, H., Iyo, H., Doi, M., Inoue, M., and Ishida, T. (1991) Biochim. Biophys.
Acta 1075, 181–18621. Stolarski, R., Sitek, A., Stepinski, J., Jankowska, M., Oksman, P., Temeriusz,
A., Darzynkiewicz, E., Lonnberg, H., and Shugar, D. (1996) Biochim. Bio-phys. Acta 1293, 97–105
22. Marcotrigiano, J., Gingras, A. C., Sonenberg, N., and Burley, S. K. (1999) Mol.Cell 3, 707–716
23. Raught, B., and Gingras, A. C. (1999) Int. J. Biochem. Cell Biol. 31, 43–5724. Raught, B., Gingras, A. C., and Sonenberg, N. (2000) in Translational Control
of Gene Expression (Sonenberg, N., Hershey, J. W. B., and Mathews, M. B.,eds) pp. 245–294, Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, NY
25. Pyronnet, S., Imataka, H., Gingras, A. C., Fukunaga, R., Hunter, T., andSonenberg, N. (1999) EMBO J. 18, 270–279
26. Waskiewicz, A. J., Johnson, J. C., Penn, B., Mahalingam, M., Kimball, S. R.,and Cooper, J. A. (1999) Mol. Cell. Biol. 19, 1871–1880
27. Joshi, B., Cai, A. L., Keiper, B. D., Minich, W. B., Mendez, R., Beach, C. M.,Stepinski, J., Stolarski, R., Darzynkiewicz, E., and Rhoads, R. E. (1995)J. Biol. Chem. 270, 14597–14603
28. Flynn, A., and Proud, C. G. (1995) J. Biol. Chem. 270, 21684–2168829. Makkinje, A., Xiong, H., Li, M., and Damuni, Z. (1995) J. Biol. Chem. 270,
14824–1482830. Pyronnet, S. (2000) Biochem. Pharmacol. 60, 1237–124331. Lachance, P. E., Miron, M., Raught, B., Sonenberg, N., and Lasko, P. (2002)
Mol. Cell. Biol. 22, 1656–166332. Scheper, G. C., van Kollenburg, B., Hu, J., Luo, Y., Goss, D. J., and Proud, C. G.
(2002) J. Biol. Chem. 277, 3303–3309
33. Haghighat, A., Mader, S., Pause, A., and Sonenberg, N. (1995) EMBO J. 14,5701–5709
34. Fletcher, C. M., McGuire, A. M., Gingras, A. C., Li, H., Matsuo, H., Sonenberg,N., and Wagner, G. (1998) Biochemistry 37, 9–15
35. Westerfield, M. (1995) The Zebrafish Book, 3 Ed., University of Oregon Press,Eugene, OR
36. Chomczynski, P. (1993) BioTechniques 15, 532–534, 536–53737. Joshi, B., Robalino, J., Schott, E. J., and Jagus, R. (2002) BioTechniques 33,
392–40138. McKendrick, L., Morley, S. J., Pain, V. M., Jagus, R., and Joshi, B. (2001) Eur.
J. Biochem. 268, 5375–538539. Mumberg, D., Muller, R., and Funk, M. (1995) Gene (Amst.) 156, 119–12240. Studier, F. W., Rosenberg, A. H., and Dunn, J. J. (1990) Methods Enzymol. 185,
49–5641. Rudolph, R., Bohm, G., Lilie, H., and Jaenicke, R. (1997) in Protein Function:
A Practical Approach (Creighton, T. E., ed) pp. 57–100, Oxford UniversityPress, Oxford
42. Lilie, H., Schwarz, E., and Rudolph, R. (1998) Curr. Opin. Biotechnol. 9,497–501
43. Jagus, R., Joshi, B., Miyamoto, S., and Beckler, G. (1998) in Current Protocolsin Cell Biology (Bonifacino, J. S., Dasso, M., Harford, J. B., Lippincott-Schwartz, J., and Yamada, K. M., eds) pp. 11.12.11–11.12.26, John Wiley &Sons, Inc., New York
44. Fling, S. P., and Gregerson, D. S. (1986) Anal. Biochem. 155, 83–8845. O’Farrell, P. H. (1975) J. Biol. Chem. 250, 4007–402146. Jagus, R., Huang, W.-I., Hiremath, L. S., Stern, B. D., and Rhoads, R. E. (1993)
Dev. Genet. 14, 412–42347. Savinova, O., and Jagus, R. (1997) Methods 11, 419–42548. Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990) J.
Mol. Biol. 215, 403–41049. Gingras, A. C., Raught, B., Gygi, S. P., Niedzwiecka, A., Miron, M., Burley,
S. K., Polakiewicz, R. D., Wyslouch-Cieszynska, A., Aebersold, R., andSonenberg, N. (2001) Genes Dev. 15, 2852–2864
50. Sonenberg, N., Morgan, M. A., Merrick, W. C., and Shatkin, A. J. (1978) Proc.Natl. Acad. Sci. U. S. A. 75, 4843–4847
51. Sonenberg, N., Rupprecht, K., Hecht, S. M., and Shatkin, A. J. (1979) Proc.Natl. Acad. Sci. U. S. A.76, 4345–4349
52. Wakiyama, M., Suzuki, A., Saigoh, M., Saka, I. N., Miyoshi, H., Kojima, S., andMiura, K. (2001) Biosci. Biotechnol. Biochem. 65, 232–235
53. Silver, P. A. (1991) Cell 64, 489–49754. Boulikas, T. (1993) Crit. Rev. Eukaryotic Gene Expression 3, 193–22755. Ptushkina, M., von der Haar, T., Vasilescu, S., Frank, R., Birkenhager, R., and
McCarthy, J. E. (1998) EMBO J. 17, 14798–1480856. Fahrenkrug, S. C. (1997) Organization and Expression of Zebrafish elF4E.
Ph.D. thesis, University of Minnesota, Minneapolis-St. Paul, MN57. Gao, M., Rychlik, W., and Rhoads, R. E. (1998) J. Biol. Chem. 273, 4622–462858. Dostie, J., Ferraiuolo, M., Pause, A., Adam, S. A., and Sonenberg, N. (2000)
EMBO J. 19, 3142–315659. Stebbins-Boaz, B., Cao, Q., de Moor, C., Mendez, R., and Richter, J. D. (1999)
Mol. Cell 4, 1017–102760. Rom, E., Kim, H. C., Gingras, A. C., Marcotrigiano, J., Favre, D., Olsen, H.,
Burley, S. K., and Sonenberg, N. (1998) J. Biol. Chem. 273, 13104–1310961. Ruud, K. A., Kuhlow, C., Goss, D. J., and Browning, K. S. (1998) J. Biol. Chem.
24, 10325–1033062. Jankowska-Anyszka, M., Lamphear, B. J., Aamodt, E. J., Harrington, T.,
Darzynkiewicz, E., Stolarski, R., and Rhoads, R. E. (1998) J. Biol. Chem.273, 10538–10542
63. Ptushkina, M., Berthelot, K., von der Haar, T., Geffers, L., Warwicker, J., andMcCarthy, J. E. (2001) Nucleic Acids Res. 29, 4561–4569
64. Miyoshi, H., Dwyer, D. S., Keiper, B. D., Jankowska-Anyszka, M., Darzynk-iewicz, E., and Rhoads, R. E. (2002) EMBO J. 21, 4680–4690
65. Liou, R. F., and Blumenthal, T. (1990) Mol. Cell. Biol. 10, 1764–1768
Two eIF4Es in Zebrafish 10541
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from

Javier Robalino, Bhavesh Joshi, Scott C. Fahrenkrug and Rosemary JagusFunctionally Divergent
Two Zebrafish eIF4E Family Members Are Differentially Expressed and
doi: 10.1074/jbc.M313688200 originally published online December 29, 20032004, 279:10532-10541.J. Biol. Chem.
10.1074/jbc.M313688200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/279/11/10532.full.html#ref-list-1
This article cites 59 references, 25 of which can be accessed free at
by guest on June 25, 2016http://w
ww
.jbc.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















