
Differential expression of B7 co-stimulatory molecules by astrocytes correlates with T cell...
11
International Immunology, Vol. 11, No. 7, pp. 1169–1179 © 1999 The Japanese Society for Immunology Differential expression of B7 co-stimulatory molecules by astrocytes correlates with T cell activation and cytokine production Jeanne M. Soos, Timothy A. Ashley, Jennifer Morrow, Juan Carlos Patarroyo 2 , Brian E. Szente 1 and Scott S. Zamvil 2 Center for Neurologic Diseases, Harvard Institutes of Medicine, and 1 Vascular Research Division, Department of Pathology, Brigham and Women’s Hospital, Boston, MA 02115, USA 2 Department of Neurology, C-440, University of California, San Francisco, 521 Parnassus Avenue, San Francisco, CA 94143-0114, USA Keywords: antigen presentation, astrocytes, costimulation, experimental allergic encephalomyelitis, MHC class II, multiple sclerosis, T cells Abstract Whether astrocytes utilize B7:CD28 co-stimulation to activate T cells mediating CNS inflammatory disease is controversial. In this report, primary astrocytes and murine astrocyte lines, generated by immortalization at two different times, day 7 or 45 of culture, were examined for their capability to express B7 co-stimulatory molecules and to participate in B7:CD28 co-stimulation. Following exposure to IFN-γ, primary astrocytes and astrocyte lines up-regulated MHC class II and B7-2 (CD86) molecules. However, B7-1 (CD80) expression was not inducible on primary astrocytes examined after IFN-γ stimulation beginning on day 7 or on astrocyte lines immortalized on day 7. B7-1 expression was inducible on primary astrocytes examined later and could be up-regulated on astrocyte lines immortalized later. Unlike B7-1, temporal discordant expression of other co-stimulatory/adhesion molecules was not observed. Both B7-1 – /B7-2 F and B7-1 F /B7-2 F astrocyte lines were capable of stimulating proliferation of encephalitogenic T h 1 cells, utilizing B7-2 for B7:CD28 co-stimulation. However, lines derived from immortalization later (B7-1 F /B7-2 F ) were more effective in stimulating proliferation of naive myelin basic protein-specific CD4 F T cells. Astrocyte lines that expressed both B7-1 and B7-2 also stimulated T h p cells to secrete proinflammatory T h 1 cytokines, whereas lines that expressed B7-2 only stimulated T h p cells to produce a T h 2 cytokine pattern. Thus, we demonstrate for the first time that individual astrocytes can differentially express B7-1 molecules, which may correlate with their ability to stimulate proinflammatory and regulatory patterns of cytokine production. These results suggest that astrocytes have potential for both promoting and down-regulating T cell responses, and that temporal differences in expression of B7 molecules should be considered when evaluating immune regulation by astrocytes. Introduction Whether astrocytes, the most abundant glial cell population within the CNS, participate in the activation of T cells causing CNS inflammatory disease is not clear. It has been reported that astrocytes within inflammatory lesions of multiple sclerosis (MS) and experimental allergic encephalomyelitis (EAE) express MHC class II molecules, suggesting that they may serve as antigen-presenting cells (APC) for the activation of CD4 1 T cells in vivo (1,2). In vitro studies have shown that astrocytes exposed to IFN-γ up-regulate class II molecules Correspondence to: S. S. Zamvil as above; Email: [email protected] Transmitting editor: L. Steinman Received 18 December 1998, accepted 1 April 1999 and can present antigen to T cells (3–8). It has recently been reported that IFN-γ-activated astrocytes can also express elements of the endocytic pathway (8,9) and are capable of processing native CNS autoantigen (9), a function that could be necessary for initiating activation of autopathogenic T cells in vivo. However, certain reports indicate that astrocytes are less competent APC (5,10–12), and suggest that astrocytes may serve to down-regulate CNS inflammatory responses (13,14). It is also not clear whether astrocytes are capable of
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Differential expression of B7 co-stimulatory molecules by astrocytes correlates with T cell...

International Immunology, Vol. 11, No. 7, pp. 1169–1179 © 1999 The Japanese Society for Immunology
Differential expression of B7 co-stimulatorymolecules by astrocytes correlates withT cell activation and cytokine production
Jeanne M. Soos, Timothy A. Ashley, Jennifer Morrow, Juan Carlos Patarroyo 2,Brian E. Szente 1 and Scott S. Zamvil 2
Center for Neurologic Diseases, Harvard Institutes of Medicine, and 1Vascular Research Division,Department of Pathology, Brigham and Women’s Hospital, Boston, MA 02115, USA2Department of Neurology, C-440, University of California, San Francisco, 521 Parnassus Avenue, SanFrancisco, CA 94143-0114, USA
Keywords: antigen presentation, astrocytes, costimulation, experimental allergic encephalomyelitis, MHCclass II, multiple sclerosis, T cells
Abstract
Whether astrocytes utilize B7:CD28 co-stimulation to activate T cells mediating CNS inflammatorydisease is controversial. In this report, primary astrocytes and murine astrocyte lines, generated byimmortalization at two different times, day 7 or 45 of culture, were examined for their capability toexpress B7 co-stimulatory molecules and to participate in B7:CD28 co-stimulation. Followingexposure to IFN- γ, primary astrocytes and astrocyte lines up-regulated MHC class II and B7-2(CD86) molecules. However, B7-1 (CD80) expression was not inducible on primary astrocytesexamined after IFN- γ stimulation beginning on day 7 or on astrocyte lines immortalized on day 7.B7-1 expression was inducible on primary astrocytes examined later and could be up-regulatedon astrocyte lines immortalized later. Unlike B7-1, temporal discordant expression of otherco-stimulatory/adhesion molecules was not observed. Both B7-1 –/B7-2F and B7-1 F/B7-2F astrocytelines were capable of stimulating proliferation of encephalitogenic T h1 cells, utilizing B7-2 forB7:CD28 co-stimulation. However, lines derived from immortalization later (B7-1 F/B7-2F) were moreeffective in stimulating proliferation of naive myelin basic protein-specific CD4 F T cells. Astrocytelines that expressed both B7-1 and B7-2 also stimulated T hp cells to secrete proinflammatory T h1cytokines, whereas lines that expressed B7-2 only stimulated T hp cells to produce a T h2 cytokinepattern. Thus, we demonstrate for the first time that individual astrocytes can differentially expressB7-1 molecules, which may correlate with their ability to stimulate proinflammatory and regulatorypatterns of cytokine production. These results suggest that astrocytes have potential for bothpromoting and down-regulating T cell responses, and that temporal differences in expression of B7molecules should be considered when evaluating immune regulation by astrocytes.
Introduction
Whether astrocytes, the most abundant glial cell populationwithin the CNS, participate in the activation of T cells causingCNS inflammatory disease is not clear. It has been reportedthat astrocytes within inflammatory lesions of multiple sclerosis(MS) and experimental allergic encephalomyelitis (EAE)express MHC class II molecules, suggesting that they mayserve as antigen-presenting cells (APC) for the activation ofCD41 T cells in vivo (1,2). In vitro studies have shown thatastrocytes exposed to IFN-γ up-regulate class II molecules
Correspondence to: S. S. Zamvil as above; Email: [email protected]
Transmitting editor: L. Steinman Received 18 December 1998, accepted 1 April 1999
and can present antigen to T cells (3–8). It has recently beenreported that IFN-γ-activated astrocytes can also expresselements of the endocytic pathway (8,9) and are capable ofprocessing native CNS autoantigen (9), a function that couldbe necessary for initiating activation of autopathogenic T cellsin vivo. However, certain reports indicate that astrocytes areless competent APC (5,10–12), and suggest that astrocytesmay serve to down-regulate CNS inflammatory responses(13,14). It is also not clear whether astrocytes are capable of

1170 B7 co-stimulation by astrocytes
activating unprimed T cells (5), which could be relevant tothe recruitment of T cells involved in the later phases of CNSinflammatory responses (15).
B7-1 (CD80) and B7-2 (CD86) are cell surface proteins onAPC that provide potent co-stimulatory signals for T activationand differentiation through their interaction with the T cellcounter-receptor, CD28 (16,17). Results from several investi-gations in EAE (18–21), as well as other T cell-mediatedinflammatory and autoimmune conditions (22–26), have indi-cated that B7-1 and B7-2 have distinct effects in the differenti-ation of precursor T cells (Thp) into proinflammatory (Th1) andregulatory (Th2) cells. In vivo studies of EAE indicate that B7-1 promotes differentiation of encephalitogenic Th1 cells (18–20) and some studies also suggest that B7-2 promotes Th2differentiation (18,20). Although the role of B7:CD28 co-stimulation in T cell activation and regulation by individualresident CNS accessory cell subpopulations has been investi-gated in both MS and EAE (8,27–29), the relative contributionof B7-1 and B7-2 expression to T cell activation within theCNS has not yet been clarified. In examination of MS brain,some investigators reported that B7-1 molecules wereexpressed primarily on microglia (27,28) and not astrocytes(28). However, in one recent study of relapsing EAE, B7-2co-stimulatory molecules were observed on both astrocytesand microglia, while B7-1 expression on astrocytes occurredlater (29). Reports regarding B7 expression and B7:CD28 co-stimulation by astrocytes examined in vitro have also conflicted(7,8,11,12,28,30). Some investigators have reported thatIFN-γ-stimulated astrocytes express both B7-1 and B7-2molecules, and participate in B7:CD28 co-stimulation throughthe engagement of B7-2, but not B7-1, molecules with CD28expressed on T cells (7). In contrast, other investigators haveobserved that astrocytes do not express B7-1 or B7-2 co-stimulatory molecules and do not utilize the B7:CD28co-stimulatory pathway (11,12,30).
In the present investigation we have examined whetherastrocytes can differentially express B7-1 and B7-2 co-stimulatory molecules for B7:CD28 co-stimulation, a possibilitythat could influence their capability to serve as APC inT cell activation and regulation, and could also provide anexplanation for the conflicting results reported regarding T cellactivation by astrocytes (7,8,11,12,28,30). In order to examinethis possibility, immortalized and primary astrocytes wereexamined at two different time periods. Immortalization cancapture cells at different stages of development, allowingone to examine whether there are temporal changes duringdifferentiation. In addition, immortalization allows one to selectpure astrocyte lines, which can be examined in the absenceof potential contaminating microglia, a concern raised byother investigators who have studied B7 expression andB7:CD28 co-stimulation by astrocytes (7,12). We haveobserved that astrocytes immortalized earlier express B7-2,but not B7-1, molecules even when activated by IFN-γ, whileastrocytes immortalized later in culture up-regulated bothB7-1 and B7-2 molecules following IFN-γ-stimulation. Similarto the immortalized astrocytes, expression of B7-1 moleculeson primary astrocytes was not observed earlier, but wasinducible by IFN-γ stimulation later. We observed that B7-2expression by astrocytes is sufficient for the activation ofmature encephalitogenic T cells, suggesting that B7-2 expres-
sion by astrocytes could contribute to the activation of ence-phalitogenic T cells in the acute phase of EAE. AlthoughB7-1–/B7-21 astrocytes did not support proliferation of Thp,they induced Thp to produce a Th2 cytokine pattern. Incontrast, expression of B7-1 by astrocytes correlated bothwith more effective activation of Thp cells and the secretionof Th1 cytokines, suggesting that some astrocytes havepotential to activate naive T cells to differentiate into pro-inflammatory Th1 cells. Thus, the potential for temporalchanges in B7-1 expression should be considered wheninvestigating T cell activation by primary astrocytes. Differen-tial expression of B7-1 co-stimulatory molecules may influencethe ability of astrocytes to serve as functional APC for thestimulation of T cells in vivo.
Methods
Development of primary astrocyte cultures
Primary glial cultures were prepared from the cerebral hemi-spheres of newborn B10.PL (H-2u) mice using establishedmethods (31) as described previously (9). After removal ofthe meninges under a dissecting microscope, tissue from thecerebral lobes was dissociated and passed through a sterile70 µm nylon cell strainer. This cell suspension was centrifuged,then treated with trypsin–EDTA for 5 min at 37°C. Cellsseparated in this manner were then washed and plated incomplete RPMI containing 10% heat-inactivated FCS,L-glutamine and penicillin/streptomycin. Cells were initiallyplated at 53104 cells/cm2 for enrichment of astrocyte growthor grown in 75 cm2 flasks that had been treated for 1 h withpoly-D-lysine (10 mg/ml) and washed 3 times with sterilewater. Cells plated in flasks were cultured for 7 days. In orderto remove non-adherent cells, including oligodendrocytesand microglia, flasks were agitated for 8 h at 200 r.p.m. onan orbital shaker. Media, containing non-adherent cells, wasremoved. Media was replaced and agitation was continuedfor an additional 18 h at 200 r.p.m. Again, non-adherent cellswere removed and media replaced. Ninety-seven percent ofthe remaining cells were GFAP1 and ,1% stained positivefor Mac-1, a cell surface molecule expressed on microglia,monocytes and macrophages.
Immortalization of primary astrocytes
The SV40-406 cell line produces a replication defectiveretrovirus encoding the SV40 large T antigen in the pZip-NeoSV(X)1 construct (32,33) containing neomycin phospho-transferase (Neo). Immortalization by this method can ‘fix’cells in differentiation, but does not confer tumorigenicity (32).SV40-406 cells were plated at 53105 cells/75 mm2 plate in10 ml of RPMI containing 10% heat-inactivated FCS andgrown for 24 h (9). The media was changed, then collected24–48 h later and stored at –80°C. Astrocytes cultured in vitrowere exposed to SV40-406 culture supernatants for 2 h in thepresence of polybrene (4 µg/ml). After 1 day or more oftreatment, cells were exposed to G418 (400 µg/ml). Astrocytelines were obtained from immortalization of three separateprimary cultures. Three colonies were recovered from cellsimmortalized after 7 days, providing lines 3.1, 3.2 and 3.3.Two colonies were obtained from cells immortalized after 45
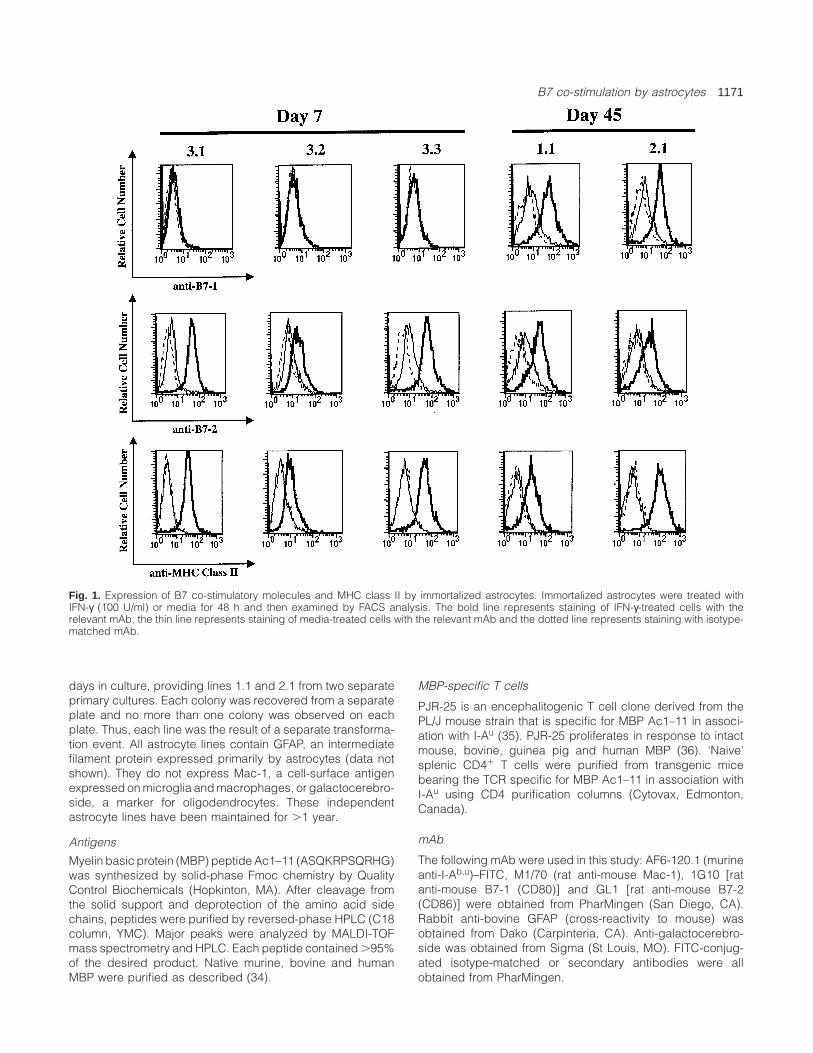
B7 co-stimulation by astrocytes 1171
Fig. 1. Expression of B7 co-stimulatory molecules and MHC class II by immortalized astrocytes. Immortalized astrocytes were treated withIFN-γ (100 U/ml) or media for 48 h and then examined by FACS analysis. The bold line represents staining of IFN-γ-treated cells with therelevant mAb, the thin line represents staining of media-treated cells with the relevant mAb and the dotted line represents staining with isotype-matched mAb.
days in culture, providing lines 1.1 and 2.1 from two separateprimary cultures. Each colony was recovered from a separateplate and no more than one colony was observed on eachplate. Thus, each line was the result of a separate transforma-tion event. All astrocyte lines contain GFAP, an intermediatefilament protein expressed primarily by astrocytes (data notshown). They do not express Mac-1, a cell-surface antigenexpressed on microglia and macrophages, or galactocerebro-side, a marker for oligodendrocytes. These independentastrocyte lines have been maintained for .1 year.
Antigens
Myelin basic protein (MBP) peptide Ac1–11 (ASQKRPSQRHG)was synthesized by solid-phase Fmoc chemistry by QualityControl Biochemicals (Hopkinton, MA). After cleavage fromthe solid support and deprotection of the amino acid sidechains, peptides were purified by reversed-phase HPLC (C18column, YMC). Major peaks were analyzed by MALDI-TOFmass spectrometry and HPLC. Each peptide contained .95%of the desired product. Native murine, bovine and humanMBP were purified as described (34).
MBP-specific T cells
PJR-25 is an encephalitogenic T cell clone derived from thePL/J mouse strain that is specific for MBP Ac1–11 in associ-ation with I-Au (35). PJR-25 proliferates in response to intactmouse, bovine, guinea pig and human MBP (36). ‘Naive’splenic CD41 T cells were purified from transgenic micebearing the TCR specific for MBP Ac1–11 in association withI-Au using CD4 purification columns (Cytovax, Edmonton,Canada).
mAb
The following mAb were used in this study: AF6-120.1 (murineanti-I-Ab,u)–FITC, M1/70 (rat anti-mouse Mac-1), 1G10 [ratanti-mouse B7-1 (CD80)] and GL1 [rat anti-mouse B7-2(CD86)] were obtained from PharMingen (San Diego, CA).Rabbit anti-bovine GFAP (cross-reactivity to mouse) wasobtained from Dako (Carpinteria, CA). Anti-galactocerebro-side was obtained from Sigma (St Louis, MO). FITC-conjug-ated isotype-matched or secondary antibodies were allobtained from PharMingen.

1172 B7 co-stimulation by astrocytes
Proliferation assaysTo measure antigen-specific T cell proliferation, APC (astro-cytes and splenocytes) were first treated with mitomycin C(60 µg/ml/106 APC) for 1 h at 37°C and washed 3 times withmedia. T cells (104/well), either the T cell clone PJR-25 orCD41 purified T cells from MBP-TCR transgenic mice, werecultured in 96-well plates with the MBP peptide Ac1–11 atvarious concentrations and APC (43104/well). Media usedfor the proliferation assays was RPMI supplemented with5% heat-inactivated FCS and L-glutamine. In all proliferationexperiments, cultures were incubated for 48 h at 37°C.[3H]Thymidine (1 µCi/ml) was added and cultured for 18 h priorto harvesting and counting. The mean c.p.m. of [3H]thymidineincorporation were calculated for triplicate cultures.
Flow cytometryFor staining cell surface molecules, astrocytes cultured aloneor treated with IFN-γ (100 U/ml) for 48 h were removed fromflasks by treatment with PBS/EDTA, counted, then aliquotedat 106 cells/tube and washed with FACS buffer (PBS containing0.5% BSA and 10 mM sodium azide). Cells were stained witheither primary unlabeled mAb or FITC–mAb for 45 min atroom temperature, then washed. In all cases, isotype-matchedcontrol unlabeled mAb or FITC–mAb were included as con-trols. When the primary mAb was unlabeled, cells were thenincubated for 45 min with a FITC–antibody directed againstthe primary mAb, then washed. Samples were analyzed ona FACSort (Becton Dickinson, Mountain View, CA) using10,000 events per sample.
Staining of intracellular proteins (GFAP and galactocerobro-side) was performed as described (37). After washing cells inFACS buffer they were treated with a 2.5% paraformaldehydesolution for 10 min at room temperature, washed in saponinbuffer (PBS containing 0.1% saponin and 10 mM sodiumazide), then incubated with primary mAb for 1 h at roomtemperature. Cells were washed with saponin buffer andincubated for 1 h with a FITC–antibody directed against theprimary mAb. Again, cells were washed with saponin buffer,then resuspended in FACS buffer and analyzed by FACS asdescribed above.
Cytokine ELISASupernatants from proliferation assays were obtained 24–48 h after initiation of culture. Ninety-six-well plates werecoated overnight in carbonate buffer containing anti-cytokinecapture antibody (1 µg/ml). The plates were washed andblocked for 2 h with 10% blocking solution (Kirkegaard& Perry, Gaithersburg, MD). The plates were washed andsupernatants from proliferation assays and the appropriatestandards were added and incubated overnight at 4°C. Theplates were washed again and the appropriate biotinylatedanti-cytokine detecting mAb (1 µg/ml) was added and incub-ated for 1 h at room temperature. The plates were washed andfurther developed by addition of avidin–peroxidase (Sigma, StLouis, MO) and substrate. Absorbance was measured at450 nm. The amount of cytokine in each supernatant wasextrapolated from the standard curve. The standards usedwere recombinant cytokine curves generated using 1:2 dilu-tions of which the initial concentration used for IL-4 was1600 pg/ml, IL-2 was 3200 pg/ml and IFN-γ was 6400 pg/ml.
Results
Time-dependent B7 co-stimulatory molecule expression byimmortalized astrocyte lines and primary astrocytes
Individual immortalized astrocyte lines were generated fromprimary B10.PL (H-2u) astrocyte cultures using a replication-defective retrovirus containing SV40 large T antigen construct(32,33) (see Methods). Immortalization of murine astrocytesby this method does not confer tumorigenicity (32). Primaryastrocytes were immortalized either after 7 or 45 days inculture. Astrocyte lines generated by immortalizationexpressed GFAP, but not Mac-1 (data not shown). Lowlevels of constitutive B7-2 expression were observed onall astrocyte lines, which increased significantly after IFN-γstimulation (Fig. 1). Following IFN-γ stimulation astrocyte linesimmortalized at the later time point (1.1 and 2.1) alsoexpressed B7-1. In contrast, when astrocytes immortalizedearlier in culture (3.1, 3.2, and 3.3) were examined, neitherconstitutive nor inducible B7-1 expression was observed.Lack of B7-1 expression was also observed when theseastrocytes (3.1, 3.2 and 3.3) were stained with a separateanti-B7-1 mAb (16-10A1), although this second anti-B7-1mAb, like the one used for B7-1 staining shown in Fig. 1,stained positively for B7-1 when staining lines 1.1 and 2.1(data not shown). Similar to primary astrocytes, these astrocytelines did not express cell surface class II without stimulationwith IFN-γ although class II (I-Au) expression was induced byIFN-γ stimulation (Fig. 1) In several separate experimentsclass II expression by line 3.2 was consistently less thanother astrocyte lines. In some experiments (Fig. 1) less B7-2expression was also observed on line 3.2, although in otherexperiments (data not shown) the levels of B7-2 up-regulationby this line were similar to the other lines when tested at thesame time.
Astrocyte lines were also examined for expression of otherco-stimulatory/adhesion molecules. Similar to previous obser-vations of primary astrocytes (7,12), constitutive ICAM andVCAM expression was observed on all of the astrocyte lines,regardless of the timing of immortalization (see Fig. 2). Up-regulation of ICAM was observed on all astrocyte lines. Nosignificant constitutive or inducible CD40 expression wasobserved on any of these lines. Thus, in contrast with theappearance of IFN-γ-inducible B7-1 expression on astrocytesimmortalized later, temporally discordant expression of theseother co-stimulatory/adhesion molecules was not observed.
In order to further evaluate whether the differentialB7-1 expression observed on immortalized lines reflected atemporal change in B7 expression in a larger astrocytepopulation, B7-1 and B7-2 expression was examined onprimary astrocyte cultures at two different time points (Fig. 3).Unactivated astrocytes and astrocytes activated with IFN-γfor 48 hours beginning on day 7 and examined on day 9showed expression of B7-2 while B7-1 was not detectable.However, when unactivated astrocytes and astrocytes activ-ated with IFN-γ for 48 hours beginning on day 28 wereexamined on day 30, expression of both B7-1 and B7-2was observed. Similarly, when examined with a second anti-B7-1 (16-10A1), B7-1 expression was not detected onunstimulated or IFN-γ-stimulated primary astrocytes examined
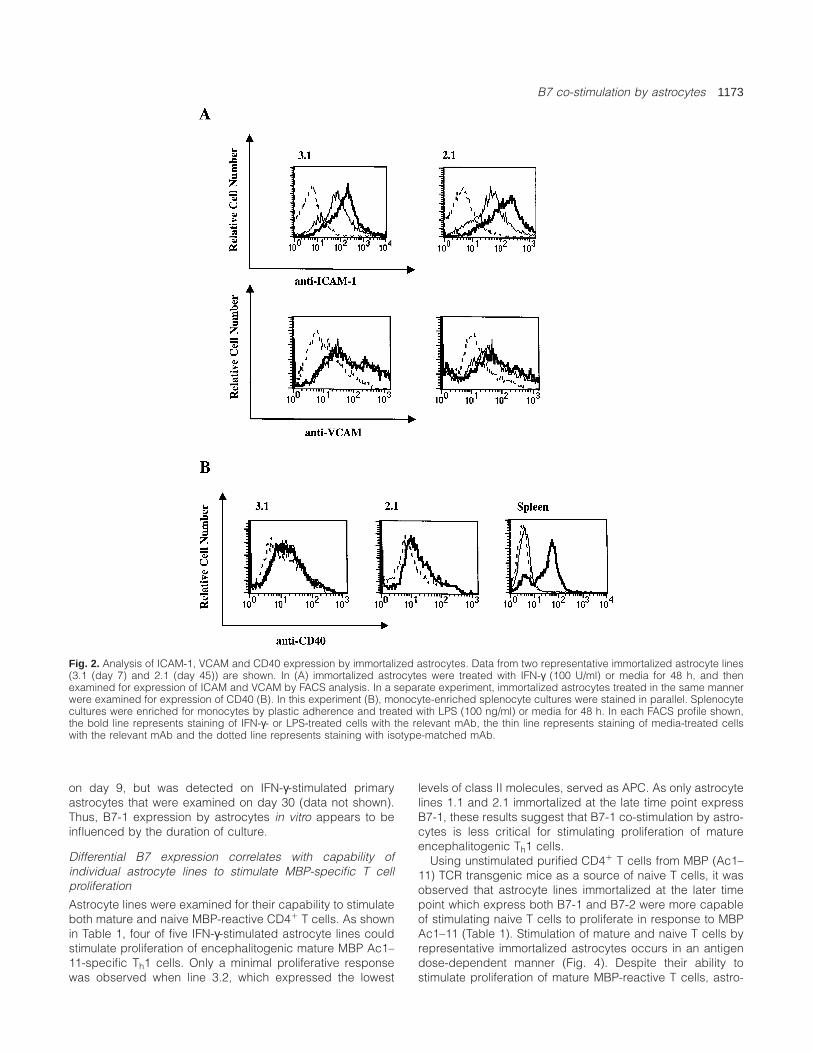
B7 co-stimulation by astrocytes 1173
Fig. 2. Analysis of ICAM-1, VCAM and CD40 expression by immortalized astrocytes. Data from two representative immortalized astrocyte lines(3.1 (day 7) and 2.1 (day 45)) are shown. In (A) immortalized astrocytes were treated with IFN-γ (100 U/ml) or media for 48 h, and thenexamined for expression of ICAM and VCAM by FACS analysis. In a separate experiment, immortalized astrocytes treated in the same mannerwere examined for expression of CD40 (B). In this experiment (B), monocyte-enriched splenocyte cultures were stained in parallel. Splenocytecultures were enriched for monocytes by plastic adherence and treated with LPS (100 ng/ml) or media for 48 h. In each FACS profile shown,the bold line represents staining of IFN-γ- or LPS-treated cells with the relevant mAb, the thin line represents staining of media-treated cellswith the relevant mAb and the dotted line represents staining with isotype-matched mAb.
on day 9, but was detected on IFN-γ-stimulated primaryastrocytes that were examined on day 30 (data not shown).Thus, B7-1 expression by astrocytes in vitro appears to beinfluenced by the duration of culture.
Differential B7 expression correlates with capability ofindividual astrocyte lines to stimulate MBP-specific T cellproliferation
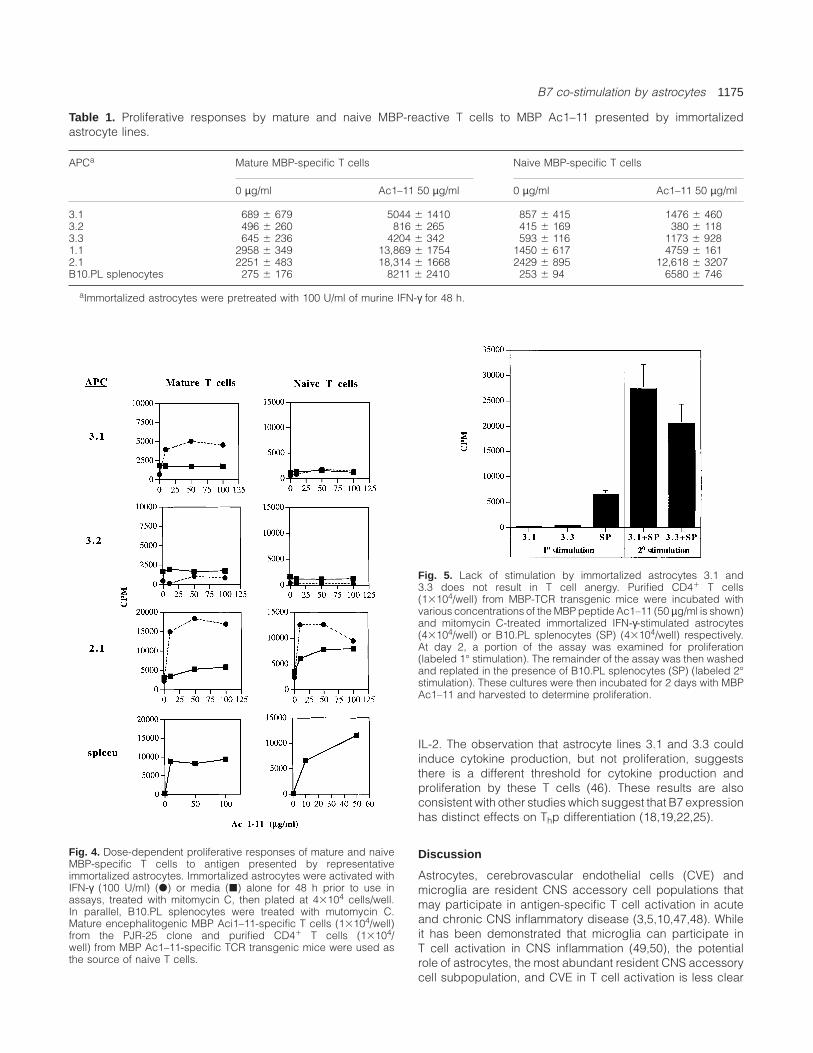
Astrocyte lines were examined for their capability to stimulateboth mature and naive MBP-reactive CD41 T cells. As shownin Table 1, four of five IFN-γ-stimulated astrocyte lines couldstimulate proliferation of encephalitogenic mature MBP Ac1–11-specific Th1 cells. Only a minimal proliferative responsewas observed when line 3.2, which expressed the lowest
levels of class II molecules, served as APC. As only astrocytelines 1.1 and 2.1 immortalized at the late time point expressB7-1, these results suggest that B7-1 co-stimulation by astro-cytes is less critical for stimulating proliferation of matureencephalitogenic Th1 cells.
Using unstimulated purified CD41 T cells from MBP (Ac1–11) TCR transgenic mice as a source of naive T cells, it wasobserved that astrocyte lines immortalized at the later timepoint which express both B7-1 and B7-2 were more capableof stimulating naive T cells to proliferate in response to MBPAc1–11 (Table 1). Stimulation of mature and naive T cells byrepresentative immortalized astrocytes occurs in an antigendose-dependent manner (Fig. 4). Despite their ability tostimulate proliferation of mature MBP-reactive T cells, astro-
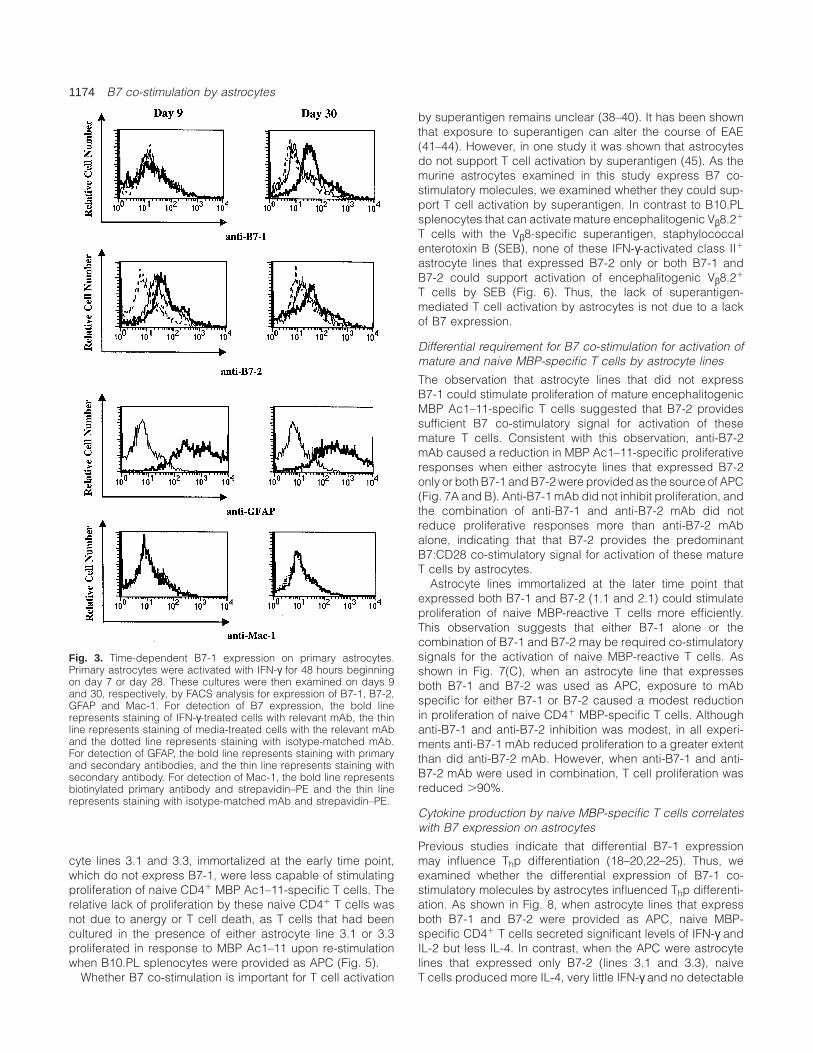
1174 B7 co-stimulation by astrocytes
Fig. 3. Time-dependent B7-1 expression on primary astrocytes.Primary astrocytes were activated with IFN-γ for 48 hours beginningon day 7 or day 28. These cultures were then examined on days 9and 30, respectively, by FACS analysis for expression of B7-1, B7-2,GFAP and Mac-1. For detection of B7 expression, the bold linerepresents staining of IFN-γ-treated cells with relevant mAb, the thinline represents staining of media-treated cells with the relevant mAband the dotted line represents staining with isotype-matched mAb.For detection of GFAP, the bold line represents staining with primaryand secondary antibodies, and the thin line represents staining withsecondary antibody. For detection of Mac-1, the bold line representsbiotinylated primary antibody and strepavidin–PE and the thin linerepresents staining with isotype-matched mAb and strepavidin–PE.
cyte lines 3.1 and 3.3, immortalized at the early time point,which do not express B7-1, were less capable of stimulatingproliferation of naive CD41 MBP Ac1–11-specific T cells. Therelative lack of proliferation by these naive CD41 T cells wasnot due to anergy or T cell death, as T cells that had beencultured in the presence of either astrocyte line 3.1 or 3.3proliferated in response to MBP Ac1–11 upon re-stimulationwhen B10.PL splenocytes were provided as APC (Fig. 5).
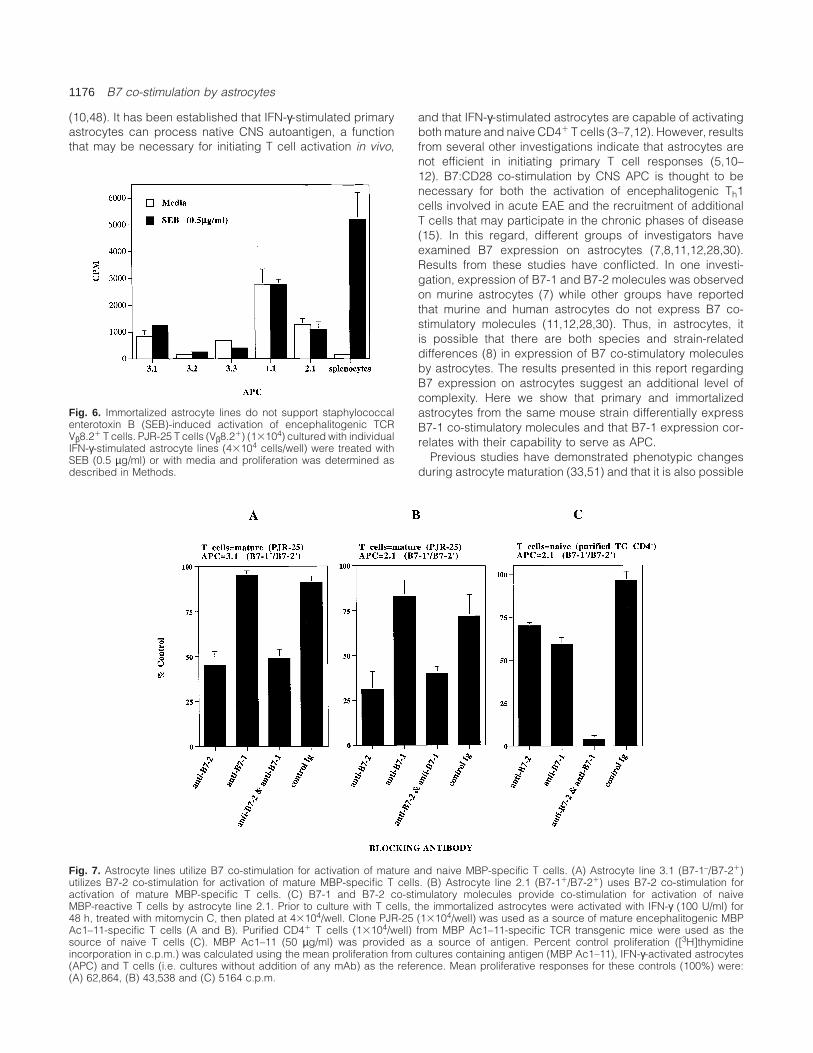
Whether B7 co-stimulation is important for T cell activation
by superantigen remains unclear (38–40). It has been shownthat exposure to superantigen can alter the course of EAE(41–44). However, in one study it was shown that astrocytesdo not support T cell activation by superantigen (45). As themurine astrocytes examined in this study express B7 co-stimulatory molecules, we examined whether they could sup-port T cell activation by superantigen. In contrast to B10.PLsplenocytes that can activate mature encephalitogenic Vβ8.21
T cells with the Vβ8-specific superantigen, staphylococcalenterotoxin B (SEB), none of these IFN-γ-activated class II1
astrocyte lines that expressed B7-2 only or both B7-1 andB7-2 could support activation of encephalitogenic Vβ8.21
T cells by SEB (Fig. 6). Thus, the lack of superantigen-mediated T cell activation by astrocytes is not due to a lackof B7 expression.
Differential requirement for B7 co-stimulation for activation ofmature and naive MBP-specific T cells by astrocyte lines
The observation that astrocyte lines that did not expressB7-1 could stimulate proliferation of mature encephalitogenicMBP Ac1–11-specific T cells suggested that B7-2 providessufficient B7 co-stimulatory signal for activation of thesemature T cells. Consistent with this observation, anti-B7-2mAb caused a reduction in MBP Ac1–11-specific proliferativeresponses when either astrocyte lines that expressed B7-2only or both B7-1 and B7-2 were provided as the source of APC(Fig. 7A and B). Anti-B7-1 mAb did not inhibit proliferation, andthe combination of anti-B7-1 and anti-B7-2 mAb did notreduce proliferative responses more than anti-B7-2 mAbalone, indicating that that B7-2 provides the predominantB7:CD28 co-stimulatory signal for activation of these matureT cells by astrocytes.
Astrocyte lines immortalized at the later time point thatexpressed both B7-1 and B7-2 (1.1 and 2.1) could stimulateproliferation of naive MBP-reactive T cells more efficiently.This observation suggests that either B7-1 alone or thecombination of B7-1 and B7-2 may be required co-stimulatorysignals for the activation of naive MBP-reactive T cells. Asshown in Fig. 7(C), when an astrocyte line that expressesboth B7-1 and B7-2 was used as APC, exposure to mAbspecific for either B7-1 or B7-2 caused a modest reductionin proliferation of naive CD41 MBP-specific T cells. Althoughanti-B7-1 and anti-B7-2 inhibition was modest, in all experi-ments anti-B7-1 mAb reduced proliferation to a greater extentthan did anti-B7-2 mAb. However, when anti-B7-1 and anti-B7-2 mAb were used in combination, T cell proliferation wasreduced .90%.
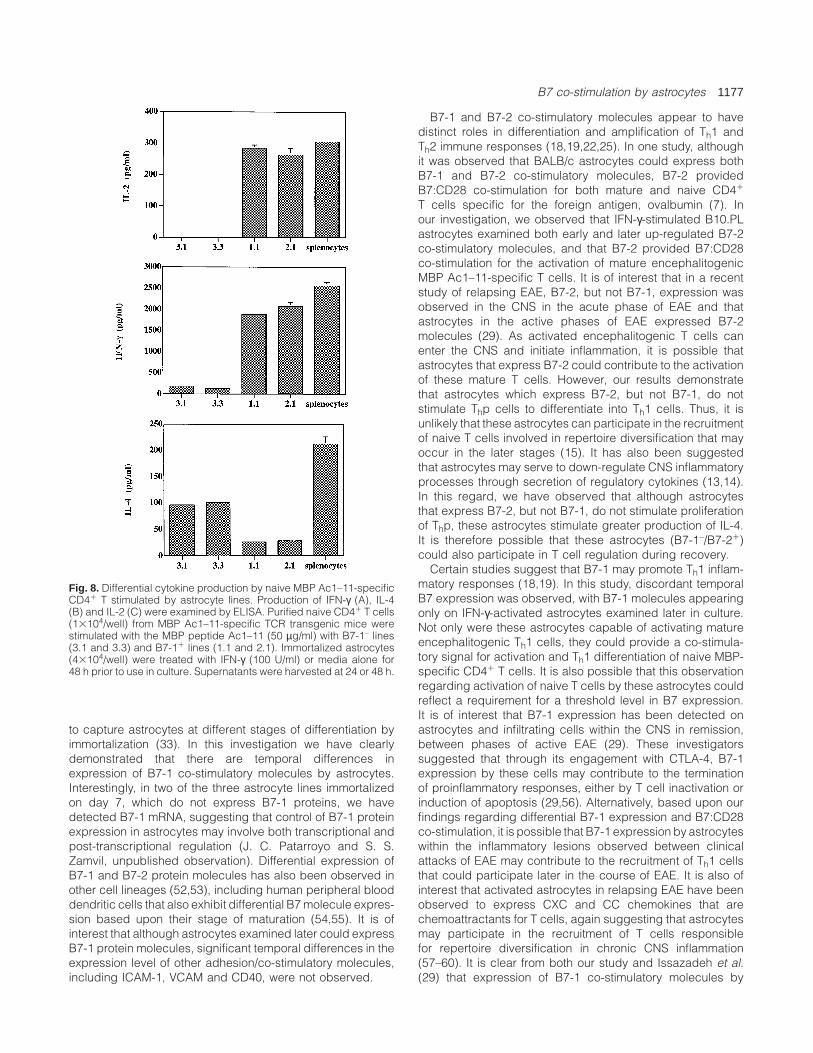
Cytokine production by naive MBP-specific T cells correlateswith B7 expression on astrocytes
Previous studies indicate that differential B7-1 expressionmay influence Thp differentiation (18–20,22–25). Thus, weexamined whether the differential expression of B7-1 co-stimulatory molecules by astrocytes influenced Thp differenti-ation. As shown in Fig. 8, when astrocyte lines that expressboth B7-1 and B7-2 were provided as APC, naive MBP-specific CD41 T cells secreted significant levels of IFN-γ andIL-2 but less IL-4. In contrast, when the APC were astrocytelines that expressed only B7-2 (lines 3.1 and 3.3), naiveT cells produced more IL-4, very little IFN-γ and no detectable
B7 co-stimulation by astrocytes 1175
Table 1. Proliferative responses by mature and naive MBP-reactive T cells to MBP Ac1–11 presented by immortalizedastrocyte lines.
APCa Mature MBP-specific T cells Naive MBP-specific T cells
0 µg/ml Ac1–11 50 µg/ml 0 µg/ml Ac1–11 50 µg/ml
3.1 689 6 679 5044 6 1410 857 6 415 1476 6 4603.2 496 6 260 816 6 265 415 6 169 380 6 1183.3 645 6 236 4204 6 342 593 6 116 1173 6 9281.1 2958 6 349 13,869 6 1754 1450 6 617 4759 6 1612.1 2251 6 483 18,314 6 1668 2429 6 895 12,618 6 3207B10.PL splenocytes 275 6 176 8211 6 2410 253 6 94 6580 6 746
aImmortalized astrocytes were pretreated with 100 U/ml of murine IFN-γ for 48 h.
Fig. 4. Dose-dependent proliferative responses of mature and naiveMBP-specific T cells to antigen presented by representativeimmortalized astrocytes. Immortalized astrocytes were activated withIFN-γ (100 U/ml) (d) or media (j) alone for 48 h prior to use inassays, treated with mitomycin C, then plated at 43104 cells/well.In parallel, B10.PL splenocytes were treated with mutomycin C.Mature encephalitogenic MBP Aci1–11-specific T cells (13104/well)from the PJR-25 clone and purified CD41 T cells (13104/well) from MBP Ac1–11-specific TCR transgenic mice were used asthe source of naive T cells.
Fig. 5. Lack of stimulation by immortalized astrocytes 3.1 and3.3 does not result in T cell anergy. Purified CD41 T cells(13104/well) from MBP-TCR transgenic mice were incubated withvarious concentrations of the MBP peptide Ac1–11 (50 µg/ml is shown)and mitomycin C-treated immortalized IFN-γ-stimulated astrocytes(43104/well) or B10.PL splenocytes (SP) (43104/well) respectively.At day 2, a portion of the assay was examined for proliferation(labeled 1° stimulation). The remainder of the assay was then washedand replated in the presence of B10.PL splenocytes (SP) (labeled 2°stimulation). These cultures were then incubated for 2 days with MBPAc1–11 and harvested to determine proliferation.
IL-2. The observation that astrocyte lines 3.1 and 3.3 couldinduce cytokine production, but not proliferation, suggeststhere is a different threshold for cytokine production andproliferation by these T cells (46). These results are alsoconsistent with other studies which suggest that B7 expressionhas distinct effects on Thp differentiation (18,19,22,25).
Discussion
Astrocytes, cerebrovascular endothelial cells (CVE) andmicroglia are resident CNS accessory cell populations thatmay participate in antigen-specific T cell activation in acuteand chronic CNS inflammatory disease (3,5,10,47,48). Whileit has been demonstrated that microglia can participate inT cell activation in CNS inflammation (49,50), the potentialrole of astrocytes, the most abundant resident CNS accessorycell subpopulation, and CVE in T cell activation is less clear
1176 B7 co-stimulation by astrocytes
(10,48). It has been established that IFN-γ-stimulated primaryastrocytes can process native CNS autoantigen, a functionthat may be necessary for initiating T cell activation in vivo,
Fig. 6. Immortalized astrocyte lines do not support staphylococcalenterotoxin B (SEB)-induced activation of encephalitogenic TCRVβ8.21 T cells. PJR-25 T cells (Vβ8.21) (13104) cultured with individualIFN-γ-stimulated astrocyte lines (43104 cells/well) were treated withSEB (0.5 µg/ml) or with media and proliferation was determined asdescribed in Methods.
Fig. 7. Astrocyte lines utilize B7 co-stimulation for activation of mature and naive MBP-specific T cells. (A) Astrocyte line 3.1 (B7-1–/B7-21)utilizes B7-2 co-stimulation for activation of mature MBP-specific T cells. (B) Astrocyte line 2.1 (B7-11/B7-21) uses B7-2 co-stimulation foractivation of mature MBP-specific T cells. (C) B7-1 and B7-2 co-stimulatory molecules provide co-stimulation for activation of naiveMBP-reactive T cells by astrocyte line 2.1. Prior to culture with T cells, the immortalized astrocytes were activated with IFN-γ (100 U/ml) for48 h, treated with mitomycin C, then plated at 43104/well. Clone PJR-25 (13104/well) was used as a source of mature encephalitogenic MBPAc1–11-specific T cells (A and B). Purified CD41 T cells (13104/well) from MBP Ac1–11-specific TCR transgenic mice were used as thesource of naive T cells (C). MBP Ac1–11 (50 µg/ml) was provided as a source of antigen. Percent control proliferation ([3H]thymidineincorporation in c.p.m.) was calculated using the mean proliferation from cultures containing antigen (MBP Ac1–11), IFN-γ-activated astrocytes(APC) and T cells (i.e. cultures without addition of any mAb) as the reference. Mean proliferative responses for these controls (100%) were:(A) 62,864, (B) 43,538 and (C) 5164 c.p.m.
and that IFN-γ-stimulated astrocytes are capable of activatingboth mature and naive CD41 T cells (3–7,12). However, resultsfrom several other investigations indicate that astrocytes arenot efficient in initiating primary T cell responses (5,10–12). B7:CD28 co-stimulation by CNS APC is thought to benecessary for both the activation of encephalitogenic Th1cells involved in acute EAE and the recruitment of additionalT cells that may participate in the chronic phases of disease(15). In this regard, different groups of investigators haveexamined B7 expression on astrocytes (7,8,11,12,28,30).Results from these studies have conflicted. In one investi-gation, expression of B7-1 and B7-2 molecules was observedon murine astrocytes (7) while other groups have reportedthat murine and human astrocytes do not express B7 co-stimulatory molecules (11,12,28,30). Thus, in astrocytes, itis possible that there are both species and strain-relateddifferences (8) in expression of B7 co-stimulatory moleculesby astrocytes. The results presented in this report regardingB7 expression on astrocytes suggest an additional level ofcomplexity. Here we show that primary and immortalizedastrocytes from the same mouse strain differentially expressB7-1 co-stimulatory molecules and that B7-1 expression cor-relates with their capability to serve as APC.
Previous studies have demonstrated phenotypic changesduring astrocyte maturation (33,51) and that it is also possible
B7 co-stimulation by astrocytes 1177
Fig. 8. Differential cytokine production by naive MBP Ac1–11-specificCD41 T stimulated by astrocyte lines. Production of IFN-γ (A), IL-4(B) and IL-2 (C) were examined by ELISA. Purified naive CD41 T cells(13104/well) from MBP Ac1–11-specific TCR transgenic mice werestimulated with the MBP peptide Ac1–11 (50 µg/ml) with B7-1– lines(3.1 and 3.3) and B7-11 lines (1.1 and 2.1). Immortalized astrocytes(43104/well) were treated with IFN-γ (100 U/ml) or media alone for48 h prior to use in culture. Supernatants were harvested at 24 or 48 h.
to capture astrocytes at different stages of differentiation byimmortalization (33). In this investigation we have clearlydemonstrated that there are temporal differences inexpression of B7-1 co-stimulatory molecules by astrocytes.Interestingly, in two of the three astrocyte lines immortalizedon day 7, which do not express B7-1 proteins, we havedetected B7-1 mRNA, suggesting that control of B7-1 proteinexpression in astrocytes may involve both transcriptional andpost-transcriptional regulation (J. C. Patarroyo and S. S.Zamvil, unpublished observation). Differential expression ofB7-1 and B7-2 protein molecules has also been observed inother cell lineages (52,53), including human peripheral blooddendritic cells that also exhibit differential B7 molecule expres-sion based upon their stage of maturation (54,55). It is ofinterest that although astrocytes examined later could expressB7-1 protein molecules, significant temporal differences in theexpression level of other adhesion/co-stimulatory molecules,including ICAM-1, VCAM and CD40, were not observed.
B7-1 and B7-2 co-stimulatory molecules appear to havedistinct roles in differentiation and amplification of Th1 andTh2 immune responses (18,19,22,25). In one study, althoughit was observed that BALB/c astrocytes could express bothB7-1 and B7-2 co-stimulatory molecules, B7-2 providedB7:CD28 co-stimulation for both mature and naive CD41
T cells specific for the foreign antigen, ovalbumin (7). Inour investigation, we observed that IFN-γ-stimulated B10.PLastrocytes examined both early and later up-regulated B7-2co-stimulatory molecules, and that B7-2 provided B7:CD28co-stimulation for the activation of mature encephalitogenicMBP Ac1–11-specific T cells. It is of interest that in a recentstudy of relapsing EAE, B7-2, but not B7-1, expression wasobserved in the CNS in the acute phase of EAE and thatastrocytes in the active phases of EAE expressed B7-2molecules (29). As activated encephalitogenic T cells canenter the CNS and initiate inflammation, it is possible thatastrocytes that express B7-2 could contribute to the activationof these mature T cells. However, our results demonstratethat astrocytes which express B7-2, but not B7-1, do notstimulate Thp cells to differentiate into Th1 cells. Thus, it isunlikely that these astrocytes can participate in the recruitmentof naive T cells involved in repertoire diversification that mayoccur in the later stages (15). It has also been suggestedthat astrocytes may serve to down-regulate CNS inflammatoryprocesses through secretion of regulatory cytokines (13,14).In this regard, we have observed that although astrocytesthat express B7-2, but not B7-1, do not stimulate proliferationof Thp, these astrocytes stimulate greater production of IL-4.It is therefore possible that these astrocytes (B7-1–/B7-21)could also participate in T cell regulation during recovery.
Certain studies suggest that B7-1 may promote Th1 inflam-matory responses (18,19). In this study, discordant temporalB7 expression was observed, with B7-1 molecules appearingonly on IFN-γ-activated astrocytes examined later in culture.Not only were these astrocytes capable of activating matureencephalitogenic Th1 cells, they could provide a co-stimula-tory signal for activation and Th1 differentiation of naive MBP-specific CD41 T cells. It is also possible that this observationregarding activation of naive T cells by these astrocytes couldreflect a requirement for a threshold level in B7 expression.It is of interest that B7-1 expression has been detected onastrocytes and infiltrating cells within the CNS in remission,between phases of active EAE (29). These investigatorssuggested that through its engagement with CTLA-4, B7-1expression by these cells may contribute to the terminationof proinflammatory responses, either by T cell inactivation orinduction of apoptosis (29,56). Alternatively, based upon ourfindings regarding differential B7-1 expression and B7:CD28co-stimulation, it is possible that B7-1 expression by astrocyteswithin the inflammatory lesions observed between clinicalattacks of EAE may contribute to the recruitment of Th1 cellsthat could participate later in the course of EAE. It is also ofinterest that activated astrocytes in relapsing EAE have beenobserved to express CXC and CC chemokines that arechemoattractants for T cells, again suggesting that astrocytesmay participate in the recruitment of T cells responsiblefor repertoire diversification in chronic CNS inflammation(57–60). It is clear from both our study and Issazadeh et al.(29) that expression of B7-1 co-stimulatory molecules by

1178 B7 co-stimulation by astrocytes
astrocytes is not static. Although we did not observe temporaldiscordant expression of ICAM, VCAM and CD40, it is possiblethat in further analysis of these astrocytes we may discovertemporal changes in the IFN-γ-stimulated expression of otheradhesion/co-stimulatory molecules that may have alsocontributed to differences in Thp activation and differentiationinto Th1 and Th2 cytokine-producing cells that we haveobserved.
In vitro analyses, such as this one, serve as an importantsupplement for examining the potential function of individualcell populations in vivo (3–9,12). Results from this study,which demonstrate both temporal changes in B7-1 expressionand differential activation of CD41 T cells by separate astro-cyte populations, suggest that astrocytes can have bothproinflammatory and regulatory influences on CD41 T cellactivation. However, it is also recognized that propagation ofastrocytes in vitro may introduce variables not encounteredin vivo, which may influence their capability to regulateT cell activation. In further studies, examination of astrocytesisolated from the CNS at different stages of EAE and analysisof mice in which B7 expression has been targeted to astro-cytes, may permit clarification of the role of B7 expression byastrocytes in vivo. Nevertheless, the results reported in thisstudy underscore the importance for considering potentialtemporal changes in B7 expression when examining antigenpresentation and T cell activation by astrocytes. Differentialexpression of B7-1 co-stimulatory molecules may be onemechanism involved in determining whether astrocytes parti-cipate in regulation or T cell activation in CNS inflammatoryconditions in vivo.
Acknowledgements
We thank Drs Vijay Kuchroo, Arlene Sharpe, Patricia Nelson, AnthonySlavin and Byron Waksman for insightful discussion and critical reviewof the manuscript. We are grateful to Drs J. Jacobberger andP. S. Jat for the SV40-406 cell line. J. M. S. is supported by apostdoctoral fellowship from the National Multiple Sclerosis Society.S. S. Z. is a Harry Weaver Neuroscience Scholar of the NationalMultiple Sclerosis Society and is also supported by grantK08-NS01771 from the National Institutes of Health.
Abbreviations
APC antigen-presenting cellCVE cerebrovascular endothelial cellEAE experimental allergic encephalomyelitisMBP myelin basic proteinMS multiple sclerosisSEB staphylococcal enterotoxin B
References
1 Sakai, K., Tabira, T., Endoh, M. and Steinman, L. 1986. Iaexpression in chronic relapsing experimental allergicencephalomyelitis induced by long-term cultured T cell lines inmice. Lab. Invest. 54:345.
2 Traugott, U. 1989. Detailed analysis of early immunopathologicevents during lesion formation in acute experimental autoimmuneencephalomyelitis. Cell. Immunol. 119:114.
3 Fontana, A., Fierz, W. and Wekerle, H. 1984. Astrocytes presentmyelin basic protein to encephalitogenic T-cell lines. Nature307:273.
4 Takiguchi, M. and Frelinger, J. A. 1986. Induction of antigen
presentation ability in purified cultures of astroglia by interferon-γ. J. Mol. Cell. Immunol. 2:269.
5 Sedgwick, J. D., Mossner, R., Schwender, S. and ter Meulen, V.1991. Major histocompatibility complex-expressing nonhemato-poietic astroglial cells prime only CD81 T lymphocytes: astroglialcells as perpetuators but not initiators of CD41 T cell responsesin the central nervous system. J. Exp. Med. 173:1235.
6 Frei, K., Lins, H., Schwerdel, C. and Fontana, A. 1994. Antigenpresentation in the central nervous system. The inhibitory effectof IL-10 on MHC class II expression and production of cytokinesdepends on the inducing signals and the type of cell analyzed.J. Immunol. 152:2720.
7 Nikcevich, K. M., Gordon, K. B., Tan, L., Hurst, S. D., Kroepfl, J. F.,Gardiner, M., Barrett, T. A. and Miller, S. D. 1997. IFN-γ activatedprimary murine astrocytes express B7 co-stimulatory moleculesand prime naive antigen-specific T cells. J. Immunol. 158:614.
8 Tan, L., Gordon, K. B., Mueller, J. P., Matis, L. A. and Miller, S. D.1998. Presentation of proteolipid protein epitopes andB7-dependent activation of encephalitogenic T cells by IFN-γ-activated SJL/J astrocytes. J. Immunol. 160:4271.
9 Soos, J. M., Morrow, J., Ashley, T. A., Szente, B. E., Bikoff, E. K.and Zamvil, S. S. 1998. Astrocytes express elements of theclass II endocytic pathway and process central nervous systemautoantigen for presentation to encephalitogenic T cells.J. Immunol. 161:5959.
10 Matsumoto, Y., Ohmori, K. and Fujiwara, M. 1992. Immuneregulation by brain cells in the central nervous system: microgliabut not astrocytes present myelin basic protein toencephalitogenic T cells under in vivo-mimicking conditions.Immunology 76:209.
11 Williams, K., Ulvestad, E. and Antel, J. P. 1994. B7/BB-1 antigenexpression on adult human microglia studied in vitro and in situ.Eur. J. Immunol. 24:3031.
12 Aloisi, F., Ria, F., Penna, G. and Adorini, L. 1998. Microglia aremore efficient that astrocytes in antigen processing and in Th1but not Th2 cell activation. J. Immunol. 160:4671.
13 Weber, F., Meinl, E., Aloisi, F., Nevinny-Stickel, C., Albert, E.,Wekerle, H. and Holfeld, R. 1994. Human astrocytes are onlypartially competent antigen presenting cells. Possible implicationsfor lesion development in multiple sclerosis. Brain 117:59.
14 Meinl, E., Aloisi, F., Ertl, B., Weber, F., de Waal Malefyt, R.,Wekerle, H. and Holfeld, R. 1994. Multiple sclerosis.Immunomodulatory effects of human astrocytes on T cells.Brain 117:1323.
15 Lehmann, P., Forsthuber, T., Miller, A. and Sercarz, E. 1992.Spreading of T cell autoimmunity to cryptic determinants of anautoantigen. Nature 358:155.
16 Lindsley, P. S. and Ledbetter, J. A. 1993. The role of the CD28receptor during T cell responses. Annu. Rev. Immunol. 11:191.
17 Croft, M., Bradley, L. M. and Swain, S. L. 1994. Naive versusmemory CD4 T cell response to antigen. Memory cells are lessdependent on accessory cell co-stimulation and can respond tomany antigen-presenting cell types including resting B cells.J. Immunol. 152:2675.
18 Kuchroo, V. K., Prabhu Das, M., Brown, J. A., Ranger, A. M.,Zamvil, S. S., Sobel, R. A., Weiner, H. L., Nabavi, N. and Glimcher,L. H. 1995. B7-1 and B7-2 co-stimulatory molecules activatedifferentially the Th1/Th2 developmental pathways: application toautoimmune disease therapy. Cell 80:707.
19 Miller, S. D., Vanderlugt, C. L., Lenschow, D. J., Pope, J. G.,Karandikar, N. J., Del Canto, M. C. and Bluestone, J. A. 1995.Blockade of CD28/B7-1 interaction prevents epitope spreadingand clinical relapses of murine EAE. Immunity 3:739.
20 Racke, M. K., Scott, D. E., Quigley, L., Gray, G. S., Abe, R.,June, C. H. and Perrin, P. J. 1995. Distinct roles for B7-1 (CD-80) and B7-2 (CD-86) in the initiation of experimental allergicencephalomyelitis. J. Clin. Invest. 96:2195.
21 Cross, A. H., Girard, T. J., Giacoletto, K. S., Evans, R. J., Keeling,R. M., Lin, R. F., Trotter, J. L. and Karr, R. W. 1995. Long-terminhibition of murine experimental autoimmune encephalomyelitisusing CTLA-4-Fc supports a key role for CD28 co-stimulation[see comments]. J. Clin. Invest. 95:2783.
22 Xu, H., Heeger, P. S. and Fairchild, R. L. 1997. Distinct roles for

B7 co-stimulation by astrocytes 1179
B7-1 and B7-2 determinants during priming of effector CD81
Tc1 and regulatory CD41 Th2 cells for contact hypersensitivty.J. Immunol. 159:4217.
23 Harris, N., Peach, R., Naemura, J., Linsley, P. S., Le Gros, G. andRonchese, F. 1997. CD80 stimulation is essential for the inductionof airway eosinophilia. J. Exp. Med. 185:177.
24 Tsuyuki, S., Tsuyuki, J., Einsle, K., Kopf, M. and Coyle, A. J. 1997.co-stimulation through B7-2 (CD86) is required for the inductionof a lung mucosal T helper cell 2 (TH2) immune response andaltered airway responsiveness. J. Exp. Med. 185:1671.
25 Murphy, M. L., Engwerda, C. R., Gorak, P. M. A. and Kaye, P. M.1997. B7-2 blockade enhances T cell responses to Leishmaniadonovani. J. Immunol. 159:4460.
26 Subramanian, G., Kazura, J. W., Pearlman, E., Jia, X., Malhotra,I. and King, C. L. 1997. B7-2 requirment for helminth-inducedgranuloma formation and CD4 type 2 T helper cell cytokineexpression. J. Immunol. 158:5914.
27 Windhagen, A., Newcombe, J., Dangond, F., Strand, C.,Woodroofe, M. N., Cuzner, M. L. and Hafler, D. A. 1995. Expressionof co-stimulatory molecules B7-1 (CD80), B7-2 (CD86), andinterleukin 12 cytokine in multiple sclerosis lesions. J. Exp. Med.182:1985.
28 De Simone, R., Giampaolo, A., Giometto, B., Gallo, P., Levi, G.,Peschle, C. and Aloisi, F. 1995. The co-stimulatory molecule B7is expressed on human microglia in culture and in multiplesclerosis acute lesions. J. Neuropathol. Exp. Neurol. 54:175.
29 Issazadeh, S., Navikas, V., Schaub, M., Sayegh, M. and Khoury,S. 1998. Kinetics of expression of co-stimulatory molecules andtheir ligands in murine relapsing experimental autoimmuneencephalomyelitis in vivo. J. Immunol. 161:1104.
30 Satoh, J., Lee, Y. B. and Kim, S. U. 1995. T-cell co-stimulatorymolecules B7-1 (CD80) and B7-2 (CD86) are expressed in humanmicroglia but not in astrocytes in culture. Brain Res. 704:92.
31 McCarthy, K. D. and de Vellis, J. 1980. Preparation of separateastroglial and oligodendroglial cell cultures from rat cerebraltissue. J. Cell Biol. 85:890.
32 Jat, P. S., Cepko, C. L., Mulligan, R. C. and Sharp, P. A.1986. Recombinant retroviruses encoding simian virus 40 largeT antigen and polyomavirus large and middle T antigens. Mol.Cell. Biol. 6:1204.
33 Frisa, P. S., Goodman, M. N., Smith, G. M., Silver, J. andJaccobberger, J. W. 1994. Immortalization of immature and matureastrocytes with SV40 T antigen. J. Neurosci. Res. 39:47.
34 Smith, M. E. 1969. An in vitro system for the study of myelinsysnthesis. J. Neurochem. 16:83.
35 Zamvil, S. S., Mitchell, D. J., Moore, A. C., Kitamura, K., Steinman,L. and Rothbard, J. B. 1986. T cell epitope of the autoantigenmyelin basic protein that induces encephalomyelitis. Nature324:258.
36 Zamvil, S. S. and Steinman, L. 1990. The T lymphocyte inexperimental allergic encephalomyelitis. Annu. Rev. Immunol.8:579.
37 Bikoff, E. K. 1992. Formation of complexes between self-peptidesand MHC class II molecules in cells defective for presentation ofexogenous protein antigens. J. Immunol. 149:1.
38 Damle, N. K., Klussman, K., Leytze, G. and Linsley, P. S. 1993.Proliferation of human T lymphocytes induced with superantigensis not dependent on co-stimulation by the CD28 counter-receptorB7. J. Immunol. 150:726.
39 Nestle, F. O., Thompson, C., Shimizu, Y., Turka, L. A. andNickoloff, B. J. 1994. Co-stimulation of superantigen-activatedT lymphocytes by autologous dendritic cells is dependent on B7.Cell. Immunol. 156:220.
40 Vella, A. T., Mitchell, T., Groth, B., Linsley, P. S., Green, J. M.,Thompson, C. B., Kappler, J. W. and Marrack, P. 1997. CD28engagement and proinflammatory cytokines contribute to T cellexpansion and long-term survival in vivo. J. Immunol. 158:4714.
41 Soos, J. M., Schiffenbauer, J. and Johnson, H. M. 1993. Treatmentof PL/J mice with the superantigen, staphycoccal enterotoxin B,
prevents development of experimental allergic encephalomyelitis.J. Neuroimmunol. 43:39.
42 Schiffenbauer, J., Johnson, H. M., Butfiloski, E., Wegrzyn, L.and Soos, J. M. 1993. Staphylococcal enterotoxins reactivateexperimental allergic encephalomyelitis. Proc. Natl Acad. Sci.USA 90:8543.
43 Brocke, S., Gaur, A., Piercy, C., Gautam, A., Gijbels, K., Fathman,C. G. and Steinman, L. 1993. Induction of relapsing paralysis inexperimental autoimmune encephalomyelitis by bacterialsuperantigen. Nature 365:642.
44 Soos, J. M., Hobeika, A. C., Butfiloski, E. J., Schiffenbauer, J. andJohnson, H. M. 1995. Accelerated induction of experimentalallergic encephalomyelitis in PL/J mice by a non-Vβ8-specificsuperantigen. Proc. Natl Acad. Sci. USA 92:6082.
45 Rott, O., Tontsch, U. and Fleischer, B. 1993. Dissociation ofantigen-presenting capacity of astrocytes for peptide-antigensversus superantigens. J. Immunol. 150:87.
46 Brown, D. R., Swier, K., Moskowitz, N. H., Naujokas, M. F.,Locksley, R. M. and Reiner, S. L. 1997. T helper subsetdifferentiation in the absence of invariant chain. J. Exp. Med.185:31.
47 Hickey, W. F. and Kimura, H. 1988. Perivascular microglial cellsof the CNS are bone marrow-derived and present antigen in vivo.Science 239:290.
48 McCarron, R. M., Spatz, M., Kempski, O., Hogan, R. N., Muehl,L. and McFarlin, D. E. 1986. Interaction between myelin basicprotein-sensitized T lymphocytes and murine cerebral vascularendothelial cells. J. Immunol. 137:3428.
49 Cash, E. and Rott, O. 1994. Microglia cells qualify as thestimulators of unprimed CD41 and CD81 T lymphocytes in thecentral nervous system. Clin. Exp. Immunol. 98:313.
50 Ford, A. L., Foulcher, E., Lemckert, F. A. and Sedgwick, J. D.1996. Microglia induce CD4 T lymphocyte final effector functionand death. J. Exp. Med. 184:1737.
51 Goodman, M. N., Silver, J. and Jacobberger, J. W. 1993.Establishment and neurite outgrowth properties of neonatal andadult rat olfactory bulb glial cell lines. Brain Res. 619:199.
52 Freeman, G. J., Borriello, F., Hodes, R. J., Reiser, H., Gribben,J. G., Ng, J. W., Kim, J., Goldberg, J. M., Hathcock, K., Laszlo,G., Lombard, L. A., Wang, S., Gray, G. S., Nadler, L. M. andSharpe, A. H. 1993. Murine B7-2, an alternative CTLA4 counter-receptor that co-stimulates T cell proliferation and interleukin 2production. J. Exp. Med. 178:2185.
53 Prabhu Das, M. R., Zamvil, S. S., Borriello, F., Weiner, H. L.,Sharpe, A. H. and Kuchroo, V. K. 1995. Reciprocal expression ofco-stimulatory molecules, B7-1 and B7-2, on murine T cellsfollowing activation. Eur. J. Immunol. 25:207.
54 Thomas, R. and Lipsky, P. E. 1994. Human peripheral blooddendritic cell subsets. Isolation and characterization of precursorand mature antigen-presenting cells. J. Immunol. 153:4016.
55 Banchereau, J. and Steinman, R. M. 1998. Dentritic cells and thecontrol of immunity. Nature 392:245.
56 Gold, R., Schmied, M., Tontsch, U., Hartung, H. P., Wekerle, H.,Toyka, K. V. and Lassmann, H. 1996. Antigen presentation byastrocytes primes rat T lymphocytes for apoptotic cell death. Amodel for T-cell apoptosis in vivo. Brain 119:651.
57 Glabinski, A. R., Tani, M., Strieter, R. M., Tuohy, V. K. andRansohoff, R. M. 1997. Synchronous synthesis of alpha- andbeta-chemokines by cells of diverse lineage in the central nervoussystem of mice with relapses of chronic experimental autoimmuneencephalomyelitis. Am. J. Pathol. 150:617.
58 Karpus, W. J. and Ransohoff, R. M. 1998. Chemokine regulationof experimental autoimmune encephalomyelitis: temporal andspatial expression patterns govern disease pathogenesis.J. Immunol. 161:2667.
59 Majumder, S., Zhou, L. Z., Chaturvedi, P., Babcock, G., Aras, S.and Ransohoff, R. M. 1998. Regulation of human IP-10 geneexpression in astrocytoma cells by inflammatory cytokines.J. Neurosci. Res. 54:169.
60 Ransohoff, R. M. and Tani, M. 1998. Do chemokines mediateleukocyte recruitment in post-traumatic CNS inflammation? TrendsNeurosci. 21:154.




















