
Integumental amino acid uptake in a carnivorous predator mollusc ( Sepia officinalis, Cephalopoda

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolog
Dietary selectivity for the toxic cyanobacterium Lyngbya majuscula
and resultant growth rates in two species of opisthobranch mollusc
Angela Capper a,*, Ian R. Tibbetts a, Judith M. O’Neil a,b, Glendon R. Shaw c
a Centre for Marine Studies, School of Life Sciences, The University of Queensland, Brisbane, Queensland 4072, Australiab University of Maryland, Centre for Environmental Science, Cambridge, MD 21673, USA
c National Centre for Environmental Toxicology (EnTox), The University of Queensland, Kessels Road,
Coopers Plains, Queensland 4108, Australia
Received 29 November 2004; received in revised form 18 July 2005; accepted 16 October 2005
Abstract
Trophodynamics of blooms of the toxic marine cyanobacterium Lyngbya majuscula were investigated to determine dietary
specificity in two putative grazers: the opisthobranch molluscs, Stylocheilus striatus and Bursatella leachii. S. striatus is
associated with L. majuscula blooms and is known to sequester L. majuscula metabolites. The dietary specificity and
toxicodynamics of B. leachii in relation to L. majuscula is less well documented. In this study we found diet history had no
significant effect upon dietary selectivity of S. striatus when offered a range of plant species. However, L. majuscula chemotype
may alter S. striatus’ selectivity for this cyanobacterium. Daily biomass increases between small and large size groups of both
species were recorded in no-choice consumption trials using L. majuscula. Both S. striatus and B. leachii preferentially
consumed L. majuscula containing lyngbyatoxin-a. Increase in mass over a 10-day period in B. leachii (915%) was significantly
greater than S. striatus (150%), yet S. striatus consumed greater quantities of L. majuscula (g day�1) and thus had a lower
conversion efficiency (0.038) than B. leachii (0.081) based on sea hare weight per gram of L. majuscula consumed day�1. Our
findings suggest that growth rates and conversion efficiencies may be influenced by sea hare maximum growth potential,
acquisition of secondary metabolites or diet type.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Bursatella leachii; Feeding preference and deterrence; Lyngbya majuscula toxins; Sea hare; Stylocheilus striatus (formerly longicauda)
1. Introduction
Opisthobranch molluscs are known to sequester die-
tary-derived secondary metabolites from chemically
defended foods (Faulkner, 1984; Hay and Fenical,
1988; Avila, 1995; Rogers et al., 1995; Faulkner, 1997;
Yamada and Kigoshi, 1997; Pennings et al., 2001,
0022-0981/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.jembe.2005.10.009
* Corresponding author. Smithsonian Marine Station, 701 Seaway
Drive, Fort Pierce, FL 34949, USA. Tel.: +1 772 465 6630x106; fax:
+1 772 461 8154.
E-mail address: [email protected] (A. Capper).
Masuda et al., 2002). Sequestered compounds can accu-
mulate in the skin or mantle (Ginsburg and Paul, 2001),
be incorporated as exudants (such as ink, opaline secre-
tions and mucous) (Paul and Pennings, 1991; Rogers et
al., 2000) or stored in the digestive gland (Winkler, 1969;
Watson, 1973; Pennings et al., 1993, Pennings and Paul,
1993a; Pennings and Carefoot, 1995; Pennings et al.,
1999). Retention of these compounds is thought to en-
hance defense strategies (Paul and Pennings, 1991;
Rogers et al., 1995, 2000; Ginsburg and Paul, 2001),
however the location of storage sites are not optimally
situated for defensive purposes (Pennings and Paul,
y and Ecology 331 (2006) 133–144

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144134
1993a,b) and may be present as the result of either a
passive or active mechanism of degradation (Avila,
1995) or detoxification (Pennings et al., 1996, 1999).
Whilst there is some debate regarding the function of
dietary sequestration, association with a chemically
defended host can at least provide the oligophagous
consumer with a source of food and a refuge from
predation (Hay, 1992; Rogers et al., 2000; Ginsburg
and Paul, 2001; Cruz-Rivera and Paul, 2002).
Stylocheilus striatus Quoy and Gaimard, 1832 (for-
merly Stylocheilus longicauda Quoy and Gaimard,
1824 (Rudman, 1999)) is a herbivorous grazer that
often exhibits a narrow diet breadth (Paul and Pennings,
1991) and sequesters secondary metabolites from its
algal host (Pennings and Paul, 1993a,b). Blooms of
the toxic cyanobacterium Lyngbya majuscula often
yield dense aggregations of S. striatus (Paul and Pen-
nings, 1991; Pennings and Paul, 1993b) although they
were also observed in large numbers in areas devoid of
L. majuscula growth where other food sources probably
supported their growth (personal observation; Turtle-
Trax, 2000).
Bursatella leachii de Blainville 1817 is a generalist
grazer of benthic cyanobacteria (Ramos et al., 1995)
and macroalgae (Paige, 1988), and may periodically
dominate marine communities. Little is known of se-
questration and secondary metabolite uptake mechan-
isms in B. leachii. Like S. striatus, B. leachii uses
chemical cues from L. majuscula to induce settlement
and rapid growth of larvae (Switzer-Dunlap and Had-
field, 1977; Paige, 1988). A high temporal and spatial
variability in such cues has been observed (Cardellina
et al., 1979; Hay and Fenical, 1988; Paul and Pennings,
1991; Nagle and Paul, 1999; Thacker and Paul, 2001).
Nagle et al., (1998) suggested that different chemotypes
of L. majuscula (which differ in composition or con-
centration of secondary metabolites) should differ dra-
matically in acceptability among potential grazers. L.
majuscula blooms in Moreton Bay, Australia often
differ in toxic metabolite expression (SEQRWQMS,
2001): Deception Bay predominantly produces debro-
moaplsyiatoxin (DAT); Eastern Banks produces lyng-
byatoxin-a (LTA); and Adams Beach produces DAT
and LTA in the early stages of bloom growth, followed
by LTA only during later growth phases (N.T. Osborne,
personal communication; Capper et al., unpublished
data). These blooms are genetically similar (T. Salmon,
personal communication) and differences in secondary
metabolite production may have a physico-environmen-
tal origin.
Large aggregations of both S. striatus and B. leachii
were observed residing in L. majuscula at the Adams
Beach site over a period of 4 months (personal obser-
vation). Both LTA and DAT were isolated from S.
striatus (Kato and Scheuer, 1975a; Mynderse et al.,
1977; Cardellina et al., 1979; Capper et al., 2005),
however the dynamics of algal toxin sequestration by
B. leachii has yet to be examined.
In this study we evaluate the dietary selectivity of S.
striatus with regard to chemotype using L. majuscula
collected from three study sites. To ascertain whether
diet history affected feeding choice, the feeding prefer-
ence of S. striatus previously exposed to L. majuscula
were compared to dnaı̈veT S. striatus, which had been
reared on a microalgal diet in a site devoid of L.
majuscula or macroalgal growth. To compare interspe-
cific variation in response to different diets we exam-
ined the feeding preferences of both S. striatus and B.
leachii using L. majuscula from the Adams Beach site
which contained two secondary metabolites (LTA and
DAT) in preliminary trials. Conversion of L. majuscula
to actual body mass in these two species was also
investigated.
2. Materials and methods
2.1. Study sites
Two areas were chosen as persistent dnuisanceTbloom sites (Fig. 1): Eastern Banks (EB) situated in
Eastern Moreton Bay (27826VS, 153822VE) in clear
oceanic waters; and Deception Bay (DB), situated
near the Western shore of Moreton Bay (27805VS,153809VE) where it often receives pulse run-off events
from terrestrial and riverine sources (Albert et al.,
2005). Adams Beach (AB) was chosen as the third
site (Fig. 1). It is situated in a sheltered bay (27851VS,153841VE) and is subjected to localised run-off during
storm events from North Stradbroke Island (personal
observation) and turbid run-off from Southern More-
ton Bay during periods of elevated rainfall. Blooms at
all sites produce either LTA or DAT or a combination
of both. All sites have extensive seagrass beds with
macroalgae locally dominating in places (Abal et al.,
2001).
2.2. Study organisms
S. striatus were collected from blooms (dbloomTStylocheilus) at the aforementioned sites throughout
the year. A second group of S. striatus was obtained
from a prawn pond at Bribie Island Aquaculture Re-
search Centre (BIARC). Sea hares had recruited in the
pond as larvae and as there was no apparent L. majus-

Fig. 1. Map of Lyngbya majuscula bloom sites in Moreton Bay with
blooms superimposed. Circle size provides approximate estimation of
bloom size between sites.
A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144 135
cula growth, it is unlikely they were exposed to
L. majuscula (dnon-bloomT Stylocheilus). In the ab-
sence of L. majuscula or macroalgae it is assumed
this group of S. striatus was feeding on microalgae.
In the laboratory the dnon-bloomT group of S. striatus
was maintained on a diet of blanched lettuce prior to
feeding assays. B. leachii were collected from a bloom
at Adams Beach. Sea hares were maintained, and
experiments conducted, in separate 50-l tanks in a
closed circuit aquarium system at Moreton Bay Re-
search Station (MBRS), located in close proximity to
the Adams Beach site. Salinities were kept between 34
and 36 ppt and temperature consistent with ambient (24
8C) with 12:12 h light and dark cycles.
L. majuscula was collected when available at the
three different locations and maintained in aquaria at
MBRS. Macroalgae and seagrasses were collected lo-
cally from the shore at Dunwich, North Stradbroke
Island. A representative range of locally abundant rho-
dophytes, phaeophytes and chlorophytes were collected
on a daily basis to ensure only healthy robust samples
were used in feeding trials. Algal availability differed
seasonally leading to different choices being presented
in some experiments. Plant species tested included the
seagrasses Zostera capricorni and Halophila ovalis;
red alga Acanthophora spicifera; green alga Caulerpa
taxifolia; brown algae Dictyota dichotoma, Sargassum
flavicans and Lobophora variegata; and the cyanobac-
terium L. majuscula.
2.3. Chemical extraction
To determine concentrations of L. majuscula sec-
ondary metabolites, voucher samples from all assays
were frozen at �20 8C and then freeze-dried. Plant
material was then extracted three times in acetone,
sonicated and filtered under vacuum. Three replicates
were pooled and dried by rotary evaporation. Concen-
trations of secondary metabolites (mg kg�1) were de-
termined by HPLC-MS/MS using a PE/Sciex API 300
mass spectrometer equipped with a high flow electro-
spray interface (TurboIonspray) coupled to a Perkin
Elmer series 200 HPLC system. Separation was
achieved using a 150�4.6 mm Altima C18 column
(Alltech) run at 35 8C, with a mobile phase consisting
of 80:20 acetonitrile/hi-pure water containing 0.1%
formic acid and 2 mM ammonium formate at a flow
rate of 0.8 ml min�1. The flow was split post column
such that the flow to the mass spectrometer interface
was 250 Al min�1. Under these conditions, the reten-
tion times were 11.72 min for LTA and 9.3 min for
DAT. The mass spectrometer was operated in the pos-
itive ion, multiple ion monitoring mode. Ions monitored
with dwell times of 300 ms were 410.3 and 438.3
(M+H)+ for LTA and 543.3 for DAT. Quantification
was achieved by comparison to standards of DAT
(kindly provided by Dr. R.E. Moore, Department of
Chemistry, University of Hawaii at Manoa) and LTA
(Calbiochem) run under the same conditions. Using a
20-Al injection the detection limit for both toxins is
typically 0.01 mg l�1 in the extracted solution.
2.4. Multiple choice feeding assays (intra- and inter-
specific dietary selectivity)
Dietary preferences of S. striatus was determined by
conducting multiple choice feeding assays using
L. majuscula from the three different locations in Mor-
eton Bay, i.e. Adams Beach, Eastern Banks and Decep-
tion Bay (Table 1).
Sea hares were acclimatized for 48 h in either four
or five replicate (depending upon availability of sea
hares at time of testing) 50 l aquaria that were masked
to eliminate behavioural interactions. Sea hares were
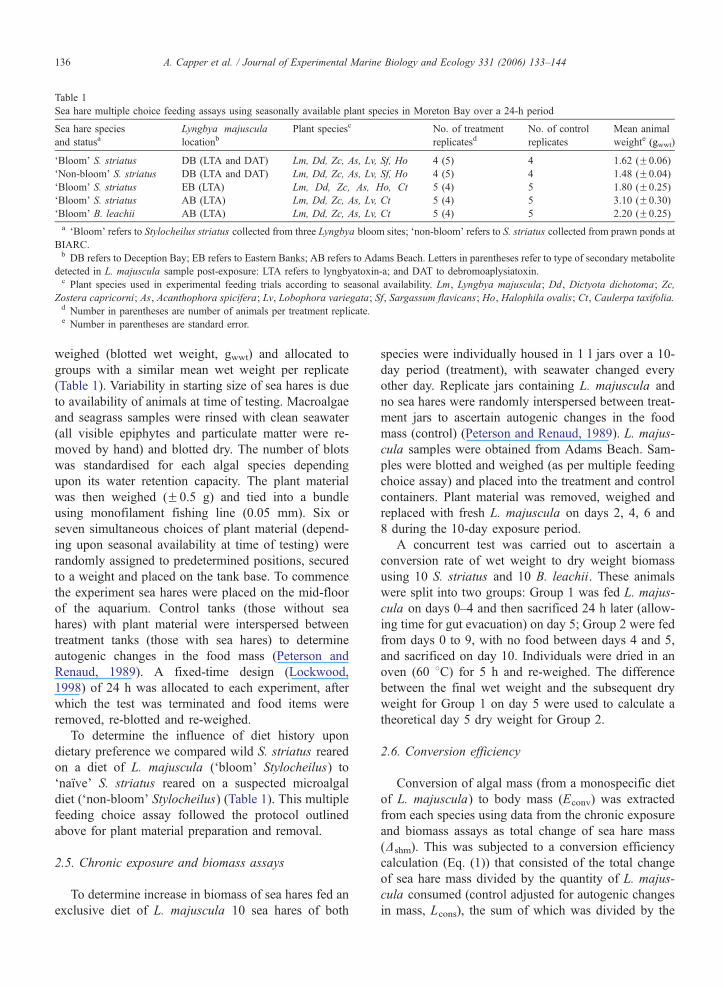
Table 1
Sea hare multiple choice feeding assays using seasonally available plant species in Moreton Bay over a 24-h period
Sea hare species
and statusaLyngbya majuscula
locationbPlant speciesc No. of treatment
replicatesdNo. of control
replicates
Mean animal
weighte (gwwt)
dBloomT S. striatus DB (LTA and DAT) Lm, Dd, Zc, As, Lv, Sf, Ho 4 (5) 4 1.62 (F0.06)
dNon-bloomT S. striatus DB (LTA and DAT) Lm, Dd, Zc, As, Lv, Sf, Ho 4 (5) 4 1.48 (F0.04)
dBloomT S. striatus EB (LTA) Lm, Dd, Zc, As, Ho, Ct 5 (4) 5 1.80 (F0.25)
dBloomT S. striatus AB (LTA) Lm, Dd, Zc, As, Lv, Ct 5 (4) 5 3.10 (F0.30)
dBloomT B. leachii AB (LTA) Lm, Dd, Zc, As, Lv, Ct 5 (4) 5 2.20 (F0.25)
a dBloomT refers to Stylocheilus striatus collected from three Lyngbya bloom sites; dnon-bloomT refers to S. striatus collected from prawn ponds at
BIARC.b DB refers to Deception Bay; EB refers to Eastern Banks; AB refers to Adams Beach. Letters in parentheses refer to type of secondary metabolite
detected in L. majuscula sample post-exposure: LTA refers to lyngbyatoxin-a; and DAT to debromoaplysiatoxin.c Plant species used in experimental feeding trials according to seasonal availability. Lm, Lyngbya majuscula; Dd, Dictyota dichotoma; Zc,
Zostera capricorni; As, Acanthophora spicifera; Lv, Lobophora variegata; Sf, Sargassum flavicans; Ho, Halophila ovalis; Ct, Caulerpa taxifolia.d Number in parentheses are number of animals per treatment replicate.e Number in parentheses are standard error.
A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144136
weighed (blotted wet weight, gwwt) and allocated to
groups with a similar mean wet weight per replicate
(Table 1). Variability in starting size of sea hares is due
to availability of animals at time of testing. Macroalgae
and seagrass samples were rinsed with clean seawater
(all visible epiphytes and particulate matter were re-
moved by hand) and blotted dry. The number of blots
was standardised for each algal species depending
upon its water retention capacity. The plant material
was then weighed (F0.5 g) and tied into a bundle
using monofilament fishing line (0.05 mm). Six or
seven simultaneous choices of plant material (depend-
ing upon seasonal availability at time of testing) were
randomly assigned to predetermined positions, secured
to a weight and placed on the tank base. To commence
the experiment sea hares were placed on the mid-floor
of the aquarium. Control tanks (those without sea
hares) with plant material were interspersed between
treatment tanks (those with sea hares) to determine
autogenic changes in the food mass (Peterson and
Renaud, 1989). A fixed-time design (Lockwood,
1998) of 24 h was allocated to each experiment, after
which the test was terminated and food items were
removed, re-blotted and re-weighed.
To determine the influence of diet history upon
dietary preference we compared wild S. striatus reared
on a diet of L. majuscula (dbloomT Stylocheilus) to
dnaı̈veT S. striatus reared on a suspected microalgal
diet (dnon-bloomT Stylocheilus) (Table 1). This multiple
feeding choice assay followed the protocol outlined
above for plant material preparation and removal.
2.5. Chronic exposure and biomass assays
To determine increase in biomass of sea hares fed an
exclusive diet of L. majuscula 10 sea hares of both
species were individually housed in 1 l jars over a 10-
day period (treatment), with seawater changed every
other day. Replicate jars containing L. majuscula and
no sea hares were randomly interspersed between treat-
ment jars to ascertain autogenic changes in the food
mass (control) (Peterson and Renaud, 1989). L. majus-
cula samples were obtained from Adams Beach. Sam-
ples were blotted and weighed (as per multiple feeding
choice assay) and placed into the treatment and control
containers. Plant material was removed, weighed and
replaced with fresh L. majuscula on days 2, 4, 6 and
8 during the 10-day exposure period.
A concurrent test was carried out to ascertain a
conversion rate of wet weight to dry weight biomass
using 10 S. striatus and 10 B. leachii. These animals
were split into two groups: Group 1 was fed L. majus-
cula on days 0–4 and then sacrificed 24 h later (allow-
ing time for gut evacuation) on day 5; Group 2 were fed
from days 0 to 9, with no food between days 4 and 5,
and sacrificed on day 10. Individuals were dried in an
oven (60 8C) for 5 h and re-weighed. The difference
between the final wet weight and the subsequent dry
weight for Group 1 on day 5 were used to calculate a
theoretical day 5 dry weight for Group 2.
2.6. Conversion efficiency
Conversion of algal mass (from a monospecific diet
of L. majuscula) to body mass (Econv) was extracted
from each species using data from the chronic exposure
and biomass assays as total change of sea hare mass
(Dshm). This was subjected to a conversion efficiency
calculation (Eq. (1)) that consisted of the total change
of sea hare mass divided by the quantity of L. majus-
cula consumed (control adjusted for autogenic changes
in mass, Lcons), the sum of which was divided by the

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144 137
experimental duration (t =10 days) (Rogers et al.,
1995).
Econv ¼Dshm=Lconsð Þ
t: ð1Þ
The effect of sea hare size was also assessed. Smal-
ler animals have a rapid growth rate and thus conver-
sion efficiency is expected to be greater. Mean initial
size of species were: (1) large B. leachii 1.077 gwwt(F0.080 SE); (2) small B. leachii 0.251 gwwt (F0.024
SE); (3) large S. striatus 3.246 gwwt (F0.058 SE); (4)
small S. striatus 0.112 gwwt (F0.005 SE).
2.7. Statistical analysis
Data were analysed using StatisticaR software.
Results of multiple choice feeding assays were analysed
using paired-sample t-tests on individual plants. t-tests
were chosen as they are very robust to departures from
normality. If significant differences occurred between
means for replicate treatment arenas and control arenas
then differential consumption and a preference was
Fig. 2. Stylocheilus striatus and Bursatella leachii multiple choice feeding a
(A) dbloomT (7) vs. dnon-bloomT(X) S. striatus from Deception Bay (cont
control (5), n =5; (C) S. striatus from Adams Beach (n) vs. control (5), n
Paired sample t-tests were used to analyse data. Asterisk (*) denotes a signific
data sets using Levene’s test. Lyngbyatoxin-a (LTA) and debromoaplysiatox
bars. n =number of replicates.
interpreted (Peterson and Renaud, 1989). Whilst their
use may be inappropriate for checking differences be-
tween feeding preferences within the same aquaria, the
technique is not inappropriate for testing for differences
in the same food between aquaria, since observations
from different tanks are independent (W. Venables,
personal communication). Levene’s homogeneity of
variance was used to ascertain normality of data.
Prior to analyses, data that deviated from normal dis-
tribution were transformed using either square root or
logarithmic transformations. To assess variance be-
tween L. majuscula chemotypes, factorial ANOVA
was used on individual (and thus independent) plant
species. To determine the influence of diet history
(dbloomT vs. dnon-bloomT) upon dietary preference t-
tests were used on control adjusted proportions of
individual plants. Biomass changes and consumption
of L. majuscula results were subjected to repeated
measures ANOVA and corresponding post-hoc analy-
sis, which allow comparisons among multiple non-in-
dependent treatments based on ranks (Conover, 1980).
Fisher’s LSD pair-wise comparison tests were used to
ssays. Data are mean proportion of plants consumed (blotted gwwt) by:
rol adjustedFS.E, n =4); (B) S. striatus from Eastern Banks (n) vs.
=5; and (D) B. leachii from Adams Beach (n) vs. control (5), n =5.
ant difference ( p b0.05). Homogeneity of variance was verified for all
in (DAT) concentration in L. majuscula is shown in mg kg�1 above

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144138
compare the means. A comparison of the gain in dry
weight between the two species from days 5 and 10 was
carried out using single factor ANOVA. Conversion
efficiency data was square-root transformed to meet
parametric criterion and subjected to factorial ANOVA.
3. Results
3.1. Multiple choice feeding assays
Two populations of S. striatus were examined and
compared: those collected from a L. majuscula bloom
(dbloomT Stylocheilus) and those unlikely to have been
exposed to L. majuscula (dnon-bloomT Stylocheilus).
Paired t-tests were used to determine differences in
individual plant consumption (control adjusted) be-
tween populations. Diet history had no significant effect
on dietary selectivity (t-test, p N0.01 for all alga, Fig.
2A). L. majuscula used in this assay contained both
LTA (0.24 mg kg�1) and DAT (0.02 mg kg�1).
In S. striatus and B. leachii multiple choice feeding
assays, L. majuscula was the most preferred food (t-
test, p b0.0001, Fig. 2B, C, D) compared to all other
6
5
4
3
2
1
0
7S. striatus weight(A)
Mea
n bl
otte
d w
et w
eigh
t (g)
L. majuscula consumed by S. striatus (B)
DayDay 0 Day 2 Day 4
6
5
4
3
2
1
0
7
n=10
*14.32
0.69
p=0.1
p=0.00
Fig. 3. Small Stylocheilus striatus and Bursatella leachii chronic exposure
striatus (0.11F0.01 SE) and B. leachii (0.25F0.02 SE) over a period of 10
by S. striatus and B. leachii over a period of 10 days (control adjustedFSE)
indicate a significant difference ( p b0.05) using Fisher’s LSD pair-wise co
toxin-a in mg kg�1 (n/d=not detected).
plants tested at Eastern Banks and Adams Beach.
Whilst S. striatus from Eastern Banks also consumed
small quantities of C. taxifolia (t-test p =0.035; Fig.
2B) and H. ovalis (t-test, p=0.036, Fig. 2B), both S.
striatus and B. leachii from Adams Beach consumed
only L. majuscula ( p b0.0001, Fig. 2C, D) suggesting
there were no interspecific differences in choice of
preferred food.
Voucher samples of L. majuscula used in S. striatus
multiple choice feeding assays yielded different concen-
trations of LTA for each location: Deception Bay (0.24
mg kg�1, Fig. 2A); Eastern Banks (56.98 mg kg�1, Fig.
2B); and Adams Beach (28.37 mg kg�1, Fig. 2C,D).
DAT was also detected in Deception Bay L. majuscula
(0.02 mg kg�1, Fig. 2A). Intraspecific differences were
observed in L. majuscula consumption from the three
locations (Deception Bay*Eastern Banks*Adams Beach
ANOVA F73.08, df2, p b0.0001, Fig. 2A, B, C).
3.2. Chronic exposure and biomass assays
All sea hares increased in biomass during laboratory
trials between days 0 and 10 (Fig. 3A and Fig. 4A).
B. leachii weight
*
L. majuscula consumed by B. leachii
of testDay 6 Day 8 Day 10
*
n=9
*n/d
0.33
0.16
19
02
and biomass assays. (A) Mean blotted wet weight (gwwt) of small S.
days. (B) Mean blotted wet weight (gwwt) of L. majuscula consumed
. Repeated measures ANOVAwere used to analyse data. Asterisks (*)
mparisons. Numbers above bars represent concentrations of lyngbya-

Day of test
Mea
n bl
otte
d w
et w
eigh
t (g)
Day 0 Day 2 Day 4 Day 6 Day 8 Day 10
10
6543210
789
S. striatus weight B. leachii weight(A)
L. majuscula consumed by S. striatus L. majuscula consumed by B. leachii
10
6543210
789
(B)
*
**
*
*
*
*n=10n=9
n/d
14.32
0.69
0.33
0.16
p<0.001
p<0.001
Fig. 4. Large Stylocheilus striatus and Bursatella leachii chronic exposure and biomass assays. (A) Mean blotted wet weight (gwwt) of large S.
striatus (3.25F0.06 g SE) and B. leachii (1.08F0.08 g SE) over a period of 10 days. (B) Mean blotted wet weight (gwwt) of L. majuscula
consumed by S. striatus and B. leachii over a period of 10 days (control adjustedFSE). Repeated measures ANOVA was used to analyse data.
Filled symbols represent significant differences ( p b0.05) between species increase in biomass using Fisher’s LSD pair-wise comparisons. Asterisks
(*) indicate a significant difference ( p b0.05) between rate of daily biomass increase between species using Fisher’s LSD pair-wise comparisons.
Numbers above bars represent concentrations of lyngbyatoxin-a in mg kg�1 (n/d=not detected).
A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144 139
Whilst small B. leachii (0.25F0.02 g SE) and S.
striatus (0.11F0.005 g SE) differed in initial wet
weights at the start of the experiment (ANOVA,
F28.33, df1, p b0.0001), their final weights were not sig-
nificantly different (ANOVA, F3.703, df1, p =0.0713).
Likewise, the daily rate of mass change� dx�ymass
t
�was not
significantly different between species over the trial
period (ANOVA, F2.69, df1, p =0.119) (Fig. 3A). There-
fore, we can say that the average rate of growth between
the species in this size range was approximately the
same over the trial period. However, a significant dif-
ference in this variable was noted between days 8 and
10 using Fisher’s LSD post-hoc analysis (Fig. 3A).
Whilst daily increases in mass were not dissimilar,
small S. striatus actually consumed significantly larger
amounts of L. majuscula throughout the trial period
(ANOVA F6.23, df4, p =0.0002) (Fig. 3B). No correla-
tion was observed between consumption rates and LTA
concentrations for small S. striatus (r2=0.067,
p =0.674) or small B. leachii (r2=0.200, p =0.450).
Large B. leachii increased more rapidly in biomass
on a daily basis when fed a mono-specific diet of L.
majuscula compared to large S. striatus (ANOVA
F14.29, df4, p b0.0001) (Fig. 4A). However, large S.
striatus consumed significantly more L. majuscula
than large B. leachii (ANOVA F79.87, df4, p b0.0001)
until day 8 when biomass increase reached a steady
state, apparently due to cessation of feeding (Fig. 4B).
The final weights of both species were not significantly
different (ANOVA, F0.29, df1, p =0.596). A single mor-
tality occurred in both small B. leachii and large S.
striatus during experimental trials; however, it is not
clear if this can be attributed to this particular diet or to
the levels of secondary metabolites contained within L.
majuscula. No correlation between consumption rates
and LTA concentrations were observed for large S.
striatus (r2=0.008, p =0.890) or large B. leachii
(r2=0.485, p =0.192).
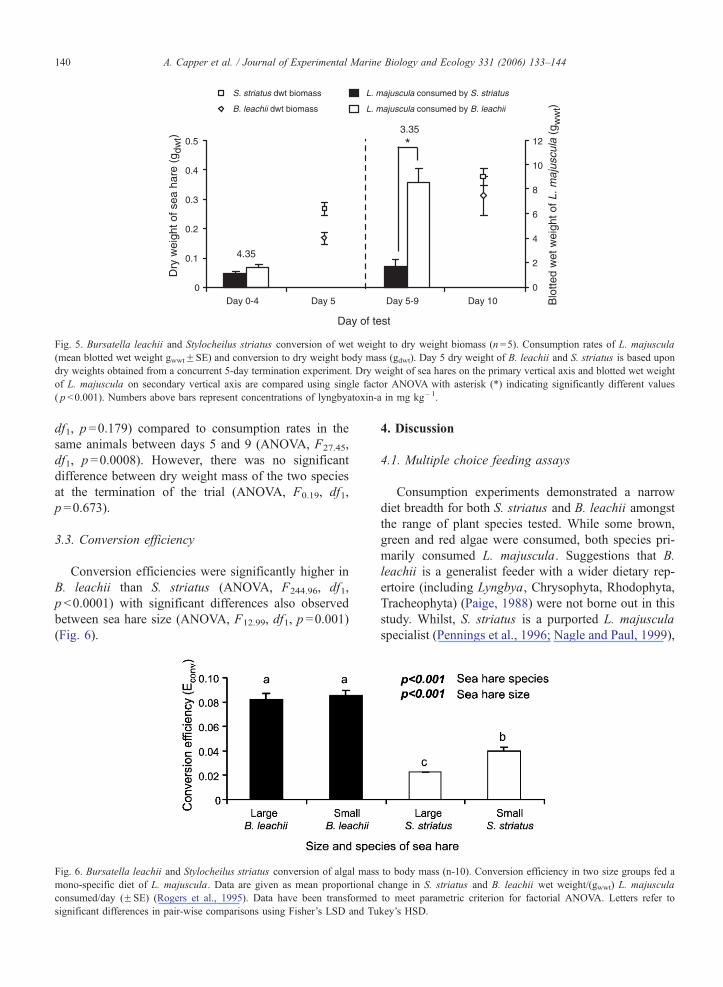
A concurrent trial to show conversion to actual body
mass (wet weight to dry weight, gdwt) of sea hares fed
L. majuscula from the Adams Beach site was carried
out (Fig. 5). S. striatus and B. leacheii were sacrificed
at day 10. Similar amounts of L. majuscula were con-
sumed between species on days 0–4 (ANOVA, F2.17,
0.5
0.4
0.3
0.2
0.1
0
12
10
8
6
4
2
0
Day 5 Day 10
L. majuscula consumed by S. striatus
L. majuscula consumed by B. leachii
Dry
wei
ght o
f sea
har
e (g
dwt)
Blo
tted
wet
wei
ght o
f L. m
ajus
cula
(g w
wt)
Day of test
S. striatus dwt biomass
B. leachii dwt biomass
*
Day 0-4 Day 5-9
4.35
3.35
Fig. 5. Bursatella leachii and Stylocheilus striatus conversion of wet weight to dry weight biomass (n =5). Consumption rates of L. majuscula
(mean blotted wet weight gwwtFSE) and conversion to dry weight body mass (gdwt). Day 5 dry weight of B. leachii and S. striatus is based upon
dry weights obtained from a concurrent 5-day termination experiment. Dry weight of sea hares on the primary vertical axis and blotted wet weight
of L. majuscula on secondary vertical axis are compared using single factor ANOVA with asterisk (*) indicating significantly different values
( p b0.001). Numbers above bars represent concentrations of lyngbyatoxin-a in mg kg�1.
A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144140
df1, p =0.179) compared to consumption rates in the
same animals between days 5 and 9 (ANOVA, F27.45,
df1, p =0.0008). However, there was no significant
difference between dry weight mass of the two species
at the termination of the trial (ANOVA, F0.19, df1,
p =0.673).
3.3. Conversion efficiency
Conversion efficiencies were significantly higher in
B. leachii than S. striatus (ANOVA, F244.96, df1,
p b0.0001) with significant differences also observed
between sea hare size (ANOVA, F12.99, df1, p =0.001)
(Fig. 6).
Fig. 6. Bursatella leachii and Stylocheilus striatus conversion of algal mass
mono-specific diet of L. majuscula. Data are given as mean proportional
consumed/day (FSE) (Rogers et al., 1995). Data have been transformed
significant differences in pair-wise comparisons using Fisher’s LSD and Tu
4. Discussion
4.1. Multiple choice feeding assays
Consumption experiments demonstrated a narrow
diet breadth for both S. striatus and B. leachii amongst
the range of plant species tested. While some brown,
green and red algae were consumed, both species pri-
marily consumed L. majuscula. Suggestions that B.
leachii is a generalist feeder with a wider dietary rep-
ertoire (including Lyngbya, Chrysophyta, Rhodophyta,
Tracheophyta) (Paige, 1988) were not borne out in this
study. Whilst, S. striatus is a purported L. majuscula
specialist (Pennings et al., 1996; Nagle and Paul, 1999),
to body mass (n-10). Conversion efficiency in two size groups fed a
change in S. striatus and B. leachii wet weight/(gwwt) L. majuscula
to meet parametric criterion for factorial ANOVA. Letters refer to
key’s HSD.

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144 141
it can occasionally be found on other plant species once
its host supply is depleted (Pennings and Paul, 1993a).
The ability to actively sequester secondary metabolites
from L. majuscula and other host plant species (Pen-
nings and Paul, 1993a,b) may confer some ecological
advantages to S. striatus. Whilst some L. majuscula
secondary metabolites can actively stimulate feeding
(Nagle et al., 1998; Cruz-Rivera and Paul, 2002), at
higher concentrations these compounds may deter feed-
ing (Pennings and Paul, 1993b). An ability to detect
changes in secondary metabolite concentrations could
potentially reduce negative post-ingestive conse-
quences for the consumer (Freeland and Janzen, 1974).
Intraspecific differences were observed in feeding
behaviour from S. striatus collected at sites Adams
Beach, Deception Bay and Eastern Banks in relation
to their L. majuscula chemotypes (i.e. those containing
different secondary metabolites). L. majuscula from
Eastern Banks was consumed in higher quantities
than either Deception Bay or Adams Beach, yet it
contained the highest concentrations of LTA, leading
us to infer LTA did not act as a feeding deterrent to S.
striatus collected from this site. L. majuscula from
Deception Bay, which contained the lowest concentra-
tions of LTA, but also contained DAT, was consumed in
much lower quantities during the 24-h test period and
ranked second in preference to the brown algae D.
dichotoma by both dbloomT and dnon-bloomT S. stria-tus. The cumulative effect of the two secondary meta-
bolites or the presence of DAT itself (or other
compounds not tested for) may have deterred feeding
in S. striatus. However, this seems unlikely as Nagle et
al. (1998) found that DAT had no deterrent effect upon
S. striatus at natural concentrations. S. striatus is capa-
ble of detoxifying both DAT and LTA into less harmful
acetates (Kato and Scheuer, 1975a,b; Pennings et al.,
1996). The capability to store or detoxify compounds
allows it to continually consume L. majuscula with no
apparent deleterious effects allowing this species to
capitalise on an ephemerally abundant food source.
Interestingly, both dbloomT and dnon-bloomT S. striatusfollowed the same feeding pattern, suggesting recent
diet history does not affect preference. High consump-
tion levels of D. dichotoma highlight the ability of this
herbivore to switch to other host species if its preferred
host becomes unavailable or unpalatable due to intol-
erable concentrations of active compounds or synergis-
tic effects between multiple secondary metabolites.
Host switching could also be a strategy to maximize
nutritional intake. Whilst the broader dietary spectrum
reported for B. leachii may result in a more nutritionally
rich diet, it may also be a means of preventing harmful
additive accumulations of secondary metabolites. When
fed a monospecific diet of L. majuscula, B. leachii
transfers LTA from body tissue to body secretions,
such as ink and faecal matter (Capper et al., 2005).
Excretion rather than storage of secondary metabolites
may help prevent autotoxicity in this species.
4.2. Chronic consumption, biomass increase and
conversion efficiency
The nutritional value of L. majuscula to S. striatus
and B. leachii was measured using growth (wet
weight, gwwt; and dry weight, gdwt) and conversion
efficiencies as indicators of performance. It has been
suggested that S. striatus is a specialist feeder of L.
majuscula (Pennings et al., 1996; Nagle and Paul,
1999). If this is the case, one might then predict that
S. striatus would exhibit better growth rates and con-
version efficiencies than a grazer with a wider dietary
repertoire, e.g. B. leachii (Paige, 1988). Differences in
growth rates were indeed observed between size
groups (small vs. large) of S. striatus and B. leachii.
Whilst small animals of both species maintained an
equivalent growth rate, consumption of L. majuscula
by S. striatus was much greater. Large S. striatus also
consumed significantly greater amounts of L. majus-
cula on days 0–8 compared to large B. leachii, yet B.
leachii increased more rapidly on a daily basis in wet
weight mass (gwwt) between days 0 and 8. Therefore, it
appears that S. striatus needs to consume a greater
biomass of L. majuscula to gain an equivalent mass
increase to B. leachii. The reason that no L. majuscula
was consumed by large S. striatus on days 8–10 is
unknown. A satiation point may have been reached or
body mass may have already increased to maximum
size for this organism. A corresponding decrease in
biomass was observed between days 8 and 10 con-
firming that high levels of L. majuscula consumption
are required to maintain overall body mass in S. stria-
tus. There was no correlation between this lack of
consumption and LTA concentrations. A new batch
of L. majuscula was collected for this trial on day
8 from the same site and whilst it contained equivalent
concentrations of LTA to previous samples, it is pos-
sible that it may have contained other unidentified
secondary metabolites, to which S. striatus may have
been sensitive. Malyngamides produced by L. majus-
cula in Guam have resulted in suppressed growth rates
in S. longicauda (Pennings and Paul, 1993b). Whilst
this study was limited to identifying the two predom-
inant toxins produced by L. majuscula in Moreton
Bay, other compounds may have been present which

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144142
deterred feeding in S. striatus (with a resultant de-
crease in mass), but not B. leachii.
In a test to determine conversion of L. majuscula to
actual body tissue (i.e. dry mass, gdwt) we found no
significant difference in dry weights between species
on day 5 or day 10 even though a significant variation
was observed between levels of L. majuscula consump-
tion in the later stages of the trial. Large B. leachii
increased in blotted wet weight (gwwt) body mass by
915% between days 0 and 9 (mean startingwwt1.09F0.07 g SE; mean finalwwt 9.98F0.02 g SE,
whereas large S. striatus only showed a 150% increase
(mean startingwwt 2.61F0.13 g SE; mean finalwwt3.92F0.87 g SE). The actual dry mass of both animals
at the end of 10 days (i.e., conversion of known quantity
of L. majuscula to actual tissue mass) was 9.5% of the
final wet weight for S. striatus (meandwt 0.372F0.03 g
SE) and 3.13% for B. leachii (meandwt 0.312F0.07 g
SE). Thus, we infer that whilst B. leachii consumed a
greater quantity of L. majuscula, this was not converted
to tissue mass and might be associated with increased
tissue fluid levels, mucous production or egg production.
Using the conversion efficiency equation suggested
by Rogers et al. (1995) Econv ¼ Dshm=Lconsð Þt
, all sea hares
had positive conversion efficiencies in the laboratory.
Both small and large B. leachii have similar conver-
sion efficiency levels (prior to statistical transforma-
tion) of 0.081 and 0.080, respectively, whereas small
S. striatus and large S. striatus were much lower at
0.038 and 0.022, respectively. Therefore, it appears
that whilst large and small S. striatus may consume
greater quantities of L. majuscula in general, they
have lower growth rates and conversion efficiencies
than B. leachii when wet weight mass is used. This
may then lead us to question why S. striatus is the
dspecialistT in this case. This may be related to the size
of the sea hare. Whilst body size was similar for small
S. striatus and B. leachii during the test period, S.
striatus may have reached its maximum size during
the test period. B. leachii on the other hand do not
reach sexual maturity until 2–3 months post-hatching
(Paige, 1986, 1988) and may still have been in a
dgrowing phaseT, resulting in higher conversion effi-
ciencies to sustain maximum growth potential. Mea-
suring conversion efficiencies for both species until a
growth plateau is reached may assist in the interpre-
tation of these results.
The acquisition of secondary metabolites from a host
plant may come at a cost. Storage and/or detoxification
mechanisms require a high energy expenditure that can
greatly reduce net energy gained from nutrition (Porter
and Orcutt, 1980) and potentially limit S. striatus’
ability to grow (Pennings and Carefoot, 1995). It is
widely known that S. striatus is able to actively seques-
ter many secondary metabolites from L. majuscula
(Paul and Pennings, 1991; Pennings and Paul,
1993a,b; Nagle et al., 1998; see reviews by Faulkner,
1984; Avila, 1995; Nagle and Paul, 1999) including
DAT and LTA (Kato and Scheuer, 1975a,b; Capper et
al., 2005), which are often compartmentalized in the
digestive gland (Watson, 1973; Pennings et al.,
1993a,b; Pennings and Carefoot, 1995; Pennings et
al., 1999). The cost/benefit ratios of different accumu-
lated compounds are likely to vary significantly be-
tween species (Rogers et al., 1995) and it may be less
energetically expensive for B. leachii to excrete or egest
sequestered metabolites than to detoxify them (Pen-
nings and Paul, 1993a; Capper et al., 2005). A further
compounding factor may be food type. Whilst S. stria-
tus exhibited low conversion efficiencies for L. majus-
cula, it may be even lower for other plant species.
Conversion efficiencies for the range of plants tested
both species for is not known.
Whilst the high degree of variability in LTA and
DAT recorded during these experiments may have af-
fected the results presented, we regarded it as important
to expose both species of sea hares to levels that are
reflected in the environment, i.e. secondary metabolites
can vary dramatically both temporally and spatially in
L. majuscula blooms in South East Queensland, Aus-
tralia (personal observation; N.T. Osborne, personal
communication). Therefore, the findings of this study
have allowed us to gauge the impacts of daily changes
in the two secondary metabolites investigated (LTA and
DAT) upon sea hare feeding patterns. Whilst S. striatus
did show significant intraspecific differences between
L. majuscula chemotypes, no correlation was found
between LTA concentration and consumption patterns
in either species during testing.
In summary, this study demonstrates that while other
macrophytes may be consumed, L. majuscula is a
preferred food choice for the two species of sea hare
tested. Both species preferentially consumed and grew
well on an exclusive diet of L. majuscula. Whilst, B.
leachii exhibited a greater wet weight daily mass in-
crease, this may be associated with either fluid retention
or mucous production as tissue dry weights were lower
than S. striatus. Growth rates and conversion efficien-
cies may be influenced by sea hare age and maximum
growth potential, acquisition of secondary metabolites
resulting in higher energetic costs or be related to
specific food type and its nutritional content. Further
studies of purported dspecialistT and dgeneralT L. majus-cula grazers are required that incorporate not only

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144 143
dietary preference and secondary metabolite acquisi-
tion, but also growth rates at different life stages on
monospecific and alternative diets to fully understand
grazer bloom dynamics.
Acknowledgements
The authors wish to thank W. Knibb and D. Willett
for supply dnaı̈veT Stylocheilus striatus from prawn
ponds at Bribie Island Aquaculture Research Centre
(BIARC); K. and K. Townsend, S. Litherland for help
at Moreton Bay Research Station (MBRS); G. Eagle-
sham and N. Osborne for help with toxicology work at
National Research Centre for Environmental Toxicol-
ogy (EnTox); J. Phillips (CMM, University of Queens-
land) for identifying marine plant species used in trials;
and N. Khan (University of Queensland) and W. Ven-
ables (CSIRO, Queensland) for help and guidance with
statistics. This project has been supported by: Centre
for Marine Studies Moreton Bay Research Station
Scholarship; the South East Queensland Regional
Water Quality Management Strategy (SEQRWQMS)
award of $25,000 for biotic interactions work; research
grants and International Post-Graduate Scholarships
from the Department of Zoology and Entomology
and the Centre for Marine Studies, School of Life
Sciences, University of Queensland, Brisbane, Austra-
lia; and R. Ritson-Williams for feedback and valuable
critique of the manuscript. [SS]
References
Abal, E.G., Dennison, W.C., Greenfield, P.F., 2001. Managing the
Brisbane River and Moreton Bay: an integrated research/manage-
ment program to reduce the impacts on an Australian estuary.
Water Sci. Technol. 43, 57–70.
Albert, S., O’Neil, J.M., Udy, J.W., Ahern, K.S., O’Sullivan, C.M.,
Dennison, W.C., 2005. Blooms of the cyanobacteria Lyngbya
majuscula in coastal Queensland, Australia: disparate sites, com-
mon factors. Mar. Pollut. Bull. 51, 428–437.
Avila, C., 1995. Natural products of optisthobranch molluscs: a
biological review. Oceanogr. Mar. Biol., Ann. Rev. 33, 487–559.
Capper, A., Tibbetts, I.R., O’Neil, J.M., Shaw, G.R., 2005. The fate of
Lyngbya majuscula toxins in three potential consumers. J. Chem.
Ecol. 31 (7), 1595–1604.
Cardellina, J.H., Marner, F., Moore, R.E., 1979. Seaweed dermatitis:
structure of Lyngbyatoxin-A. Science 204, 193–195.
Conover, W.J., 1980. Practical Nonparametric Statistics, 2nd ed. John
Wiley and Sons, New York. 493 pp.
Cruz-Rivera, E., Paul, V.J., 2002. Coral reef benthic cyanobacteria as
food and refuge: diversity, chemistry and complex interactions. Proc.
9th Int. Coral Reef Symposium, Bali, Indonesia, pp. 515–520.
Faulkner, D.J., 1984. Marine natural products: metabolites of ma-
rine algae and herbivorous marine molluscs. Nat. Prod. Rep. 1,
251–280.
Faulkner, D.J., 1997. Marine natural products. Nat. Prod. Rep. 14,
259–302.
Freeland, W.J., Janzen, D.H., 1974. Strategies in herbivory by
mammals: the role of plant secondary compounds. Am. Nat.
108, 269–289.
Ginsburg, D.W., Paul, V.J., 2001. Chemical defenses in the sea hare
Aplysia parvula: importance of diet and sequestration of algal
secondary metabolites. Mar. Ecol. Prog. Ser. 215, 261–274.
Hay, M.E., 1992. The role of seaweed chemical defenses in the
evolution of feeding specialization and in the mediation of com-
plex interactions. In: Paul, V.J. (Ed.), Ecological Roles of Marine
Natural Products. Comstock Publ. Assoc.
Hay, M.E., Fenical, W., 1988. Marine plant–herbivore interactions:
the ecology of chemical defense. Ann. Rev. Ecolog. Syst. 19,
114–145.
Kato, Y., Scheuer, P.J., 1975a. Aplysiatoxin and debromoaplysiatoxin,
constituents of the marine mollusk Stylocheilus longicauda (Quoy
and Gaimard, 1824). J. Am. Chem. Soc. 96, 2245–2246.
Kato, Y., Scheuer, P.J., 1975b. The aplysiatoxins. Pure Appl. Chem.
41, 1–14.
Lockwood, J.R., 1998. On the statistical analysis of multiple-choice
feeding preference experiments. Oecologia 116, 475–481.
Masuda, M., Kawaguchi, S., Abe, T., Kawamoto, T., Suzuki, M.,
2002. Additional analysis of chemical diversity of the red algal
genus Laurencia (Rhodomelaceae) from Jpn. Phycol. Res. 50,
135.
Mynderse, J.S., Moore, R.E., Kashiwagi, M., Norton, T.R., 1977.
Antileukemia activity in Oscillatoriaceae: isolation of debromoa-
plysiatoxin from Lyngbya. Science 196, 538–539.
Nagle, D.G., Paul, V.J., 1999. Production of secondary metabolites by
filamentous tropical marine cyanobacteria: ecological functions of
the compounds. J. Phycol. 35, 1412–1421.
Nagle, D.G., Camacho, F.T., Paul, V.J., 1998. Dietary preferences of
the opisthobranch mollusc Stylocheilus longicauda for secondary
metabolites produced by the tropical cyanobacterium Lyngbya
majuscula. Mar. Biol. 132, 267–273.
Paige, J.A., 1986. The laboratory culture of two aplysiids, Aplysia
brasiliana (Rang), 1828, and Bursatella leachii plei (Rang, 1828)
(Gastropoda: Opistobranchia) in artificial seawater. Veliger 29,
64–69.
Paige, J.A., 1988. Biology, metamophosis and postlarval development
of Bursatella leachii plei Rang (Gastropoda: Opistobranchia).
Bull. Mar. Sci. 42, 65–75.
Paul, V.J., Pennings, S.C., 1991. Diet-derived chemical defenses in
the sea hare Stylocheilus longicauda (Quoy et Gaimard 1824). J.
Exp. Mar. Biol. 151, 227–243.
Pennings, S.C., Carefoot, T.H., 1995. Post-ingestive consequences of
consuming secondary metabolites in sea hares (Gastropoda:
Opisthobranchia). Comp. Biochem. Physiol. 111, 249–256.
Pennings, S.C., Paul, V.J., 1993a. Sequestration of dietary secondary
metabolites by three species of sea hares: location, specificity and
dynamics. Mar. Biol. 117, 535–546.
Pennings, S.C., Paul, V.J., 1993b. Secondary chemistry does not limit
dietary range of the specialist sea hare Stylocheilus longicauda
(Quoy et Gaimard 1824). J. Exp. Mar. Biol. Ecol. 174, 97–113.
Pennings, S.C., Nadeau, M.T., Paul, V.J., 1993. Selectivity and
growth of the generalist herbivore Dolabella auricularia feeding
upon complementary resources. Ecology 74, 879–890.
Pennings, S.C., Weiss, A.M., Paul, V.J., 1996. Secondary metabolites
of the cyanobacterium Microcoleus lyngbyaceus and the sea hare
Stylocheilus longicauda: palatability and toxicity. Mar. Biol. 126,
735–743.

A. Capper et al. / Journal of Experimental Marine Biology and Ecology 331 (2006) 133–144144
Pennings, S.C., Paul, V.J., Dunbar, D.C., Hamann, M.T., Lumbang,
W.A., Novack, B., Jacobs, R.S., 1999. Unpalatable compounds in
the marine gastropod Dolabella auricularia: distribution and
effect of diet. J. Chem. Ecol. 25, 735–755.
Pennings, S.C., Nastisch, S., Paul, V.J., 2001. Vulnerability of sea
hares to fish predators: importance of diet and fish species. Coral
Reefs 20, 320–324.
Peterson, C.H., Renaud, P.E., 1989. Analysis of feeding preference
experiments. Oecologia 80, 82–86.
Porter, K.G., Orcutt, J.D.J., 1980. Nutritional adequacy, manageabil-
ity, and toxicity as factors that determine food quality of green and
blue-green algae for Daphnia. In: Kerfoot, W.C. (Ed.), Evolution
and Ecology of Zooplankton Communities. University Press of
New England, Hanover, NH, USA.
Ramos, L.J., Rocafort, J.L.L., Miller, M.W., 1995. Behavior pat-
terns of the Aplysiid gastropod Bursatella leachii in its natural
habitat and in the laboratory. Neurobiol. Learn. Mem. 63,
246–259.
Rogers, C.N., Steinberg, P.D., de Nys, R., 1995. Factors associated
with oligophagy in two species of sea hares (Mollusca: Anaspi-
dea). J. Exp. Mar. Biol. Ecol. 192, 47–73.
Rogers, C.N., de Nys, R., Charlton, T.S., Steinberg, P.D., 2000.
Dynamics of algal secondary metabolites in two species of sea
hare. J. Chem. Ecol. 26, 721–744.
All in-text references underlined in blue are linked to publications o
Rudman,W.B., 1999. Stylocheilus longicauda and Stylocheilus citrina
Nomenclature discussion [In] Sea Slug Forum. http://seaslug.info/
display.cfm?id=2517 date accessed 5 October 2005.
SEQRWQMS, 2001. Lyngbya Update Newsletter, http://www.epa.
qld.gov.au/environmental_management/coast_and_oceans/marine_
habitats/lyngbya_updates/science_update/ date accessed 5 Oct 2005.
Switzer-Dunlap, M., Hadfield, M.G., 1977. Observations on devel-
opmental, larval growth and metamorphosis of four species of
Aplysiidae (Gastropoda: Opisthobranchia) in laboratory culture.
J. Exp. Mar. Biol. Ecol. 29, 245–261.
Thacker, R.W., Paul, V.J., 2001. Are benthic cyanobacteria indicators
of nutrient enrichment? Relationships between cyanobacterial
abundance and environmental factors on the reef flats of Guam.
Bull. Mar. Sci. 69, 497–508.
TurtleTrax, 2000. The Algae of Honokowai. http://www.turtles.org/
limu/limu.htm date accessed 5 October 2005.
Watson, M., 1973. Midgut gland toxins of Hawaiin sea hares: I.
Isolation and preliminary toxicological observations. Toxicon
11, 259–267.
Winkler, L.R., 1969. Distribution of organic bromide compounds in
Aplysia californica Cooper 1863. Veliger 11, 268–271.
Yamada, K., Kigoshi, H., 1997. Bioactive compounds from the sea
hares of two genera: Aplysia and Dolabella. Chem. Soc. Jpn. 70,
1479–1489.
n ResearchGate, letting you access and read them immediately.
Copyright © 2022 FDOKUMEN




















