
Highly parallel microbial diagnostics using oligonucleotide microarrays
Upload
khangminh22Category
view
3download
0
SUMMARY
Globe artichoke (Cynara scolymus) is a vegetable cropaffected by many viruses causing significant economiclosses. Virus detection and identification is important tofarmers, plant pathologists and others involved in plantprotection activities of quarantine and certification pro-grams. To this aim, the potentiality of an oligonucleotidediagnostic array based on the Combimatrix platform forthe detection and identification of 14 artichoke-infectingviruses was examined. More than 190 oligonucleotidevirus probes (40-mer) were selected and used in the mi-croarray diagnostic technique developed in this investi-gation. Most of the probes were highly specific for de-tection, identification and discrimination of target virus-es. Further, for the first time the Combimatrix platformwas applied directly to evaluate the sanitary status of ar-tichoke samples collected during field surveys carriedout in 6 different Italian regions. The repeatability, speci-ficity and sensitivity of the oligonucleotide diagnostic ar-ray developed in this investigation demonstrated its ap-plicability for routine diagnostic use in artichokegermplasm as it detected simultaneously 14 viruses inone single hybridization event.
Key words: microarray, simultaneous multiple detec-tion, artichoke viruses, diagnosis, 40-mer oligonucleotideartichoke virus probes.
INTRODUCTION
Globe artichoke (Cynara scolymus) is a vegetablecrop grown mainly in countries bordering the Mediter-ranean basin, Italy, Spain, and Egypt accounting forabout 80% of its global production. This crop is affect-ed by many diseases that may cause severe economiclosses, i.e, a significant decrease in the quantity andquality of the yield.
Corresponding author: M. BarbaFax: +39.06.82070243E-mail: [email protected]
More than 22 viruses infect artichoke causing seri-ous concern to the artichoke industry. Some of theseviruses are very damaging and are in the OEPP/EPPOlist of quarantine organisms for the European Union,i.e., Tomato spotted wilt virus (TSWV).
The risk of introduction of undesirable microorgan-isms into a country through imported germplasm is highas disease symptoms may be absent in young propaga-tive material and their visual inspection may not revealthe presence of systemic pathogens such as viruses. Inview of the above it is crucial to assess the presence of vi-ral pathogens especially when plantlets used for the es-tablishment of new artichoke crops are produced by invitro propagation. Given the international rules issued toregulate the commercialization and sanitary requirementsfor the safe world-wide movement of germplasm, thepresence/absence of damaging systemic pathogens mustbe ascertained in transported plant material.
Pathogen diagnosis and management strategies in-creasingly require fast and accurate methods for the de-tection and identification of multiple microorganismsfrom complex samples. However, before disease controlmeasures can be undertaken, accurate diagnosis is cru-cial to confirm the presence of a virus and determine itsidentity (Li et al., 2004). Existing methods for screeninga broad range of viruses are restricted to detecting oneor few viruses per assay. There is a need to improve di-agnostic techniques that identify, in a single experi-ment, more than one or few viruses.
Microarray detection method provides the great capa-bility for parallel yet specific testing and can be usedto detect genera, species and/or strains of multiple orindividual viruses (Hadidi et al., 2004; Hadidi and Bar-ba, 2008; Barba and Hadidi, 2007, 2011a, 2011b). Differ-ent microarray platforms, using different solid support(surfaces or beads) and oligonucleotide probe synthesis,have been developed for plant virus detection (Tiberi-ni and Barba, 2012; Abdullahi et al., 2005, 2011; Nico-laisen, 2011; Engel et al., 2010; Tiberini et al., 2010;Zhang et al., 2010; Wei et al., 2009; Pasquini et al.,2008, 2010; Lenz et al., 2008; Gambino and Gribaudo,2006; Bystricka et al., 2003, 2005; Deyong et al., 2005;Boonham et al., 2003; Lee et al., 2003).
Among the different available microarray technolog-
Journal of Plant Pathology (2013), 95 (1), 145-154 Edizioni ETS Pisa, 2013 145
DEVELOPMENT OF A OLIGONUCLEOTIDE-BASED MICROARRAYFOR DETECTION OF MULTIPLE ARTICHOKE VIRUSES
A. Tiberini and M. Barba
Consiglio per la Ricerca e la Sperimentazione in Agricoltura-Centro di Ricerca per la Patologia Vegetale,Via C.G. Bertero 22, 00156 Rome, Italy
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 145
146 Microarray for artichoke virus detection Journal of Plant Pathology (2013), 95 (1), 145-154
ical platforms developed until now, the Customarray®by Combimatrix is considered highly suitable for si-multaneous and multiple detection of viruses in veg-etable crops (Tiberini et al., 2010; Tiberini and Barba,2012).
Based on the previous positive experience (Tiberiniet al., 2010; Tiberini and Barba, 2012), a dedicated Cus-tomarray® (4x2K) chip was developed including thedesign and synthesis of 40-mer oligonucleotide probesspecific for the detection of each of fourteen virusesknown to induce severe damage in artichoke. Theseviruses are included in the Italian national certificationprograms for the production of pathogen-free propaga-tive artichoke germplasm.
The oligonucleotide diagnostic microarray-based de-tection method developed in this investigation was suc-cessfully used in monitoring the sanitary status of arti-choke plantations in 6 Italian Regions where this crop iscultivated.
MATERIALS AND METHODS
Plant and virus sources. Positive- and negative-sensesingle-stranded RNA viruses were used in this study(Table 1).
The characterized virus isolates are maintained atthe Plant Pathology Research Center - CRAPAV collec-tion (COLMIA collection) as lyophilized leaves of natu-rally or artificially infected plants of Nicotiana benthami-ana and/or Gomphrena globosa.
All virus isolates were transferred to herbaceoushosts and maintained in a green house under controlledconditions. The presence of viruses was ascertainedand confirmed by ELISA or RT-PCR.
Samples collected from more than 20 artichoke va-rieties during field surveys in 6 Italian Regions arepresented in Table 2. These samples were analyzed bymicroarray hybridization. Healthy artichoke plantswere used as internal negative controls in each test.
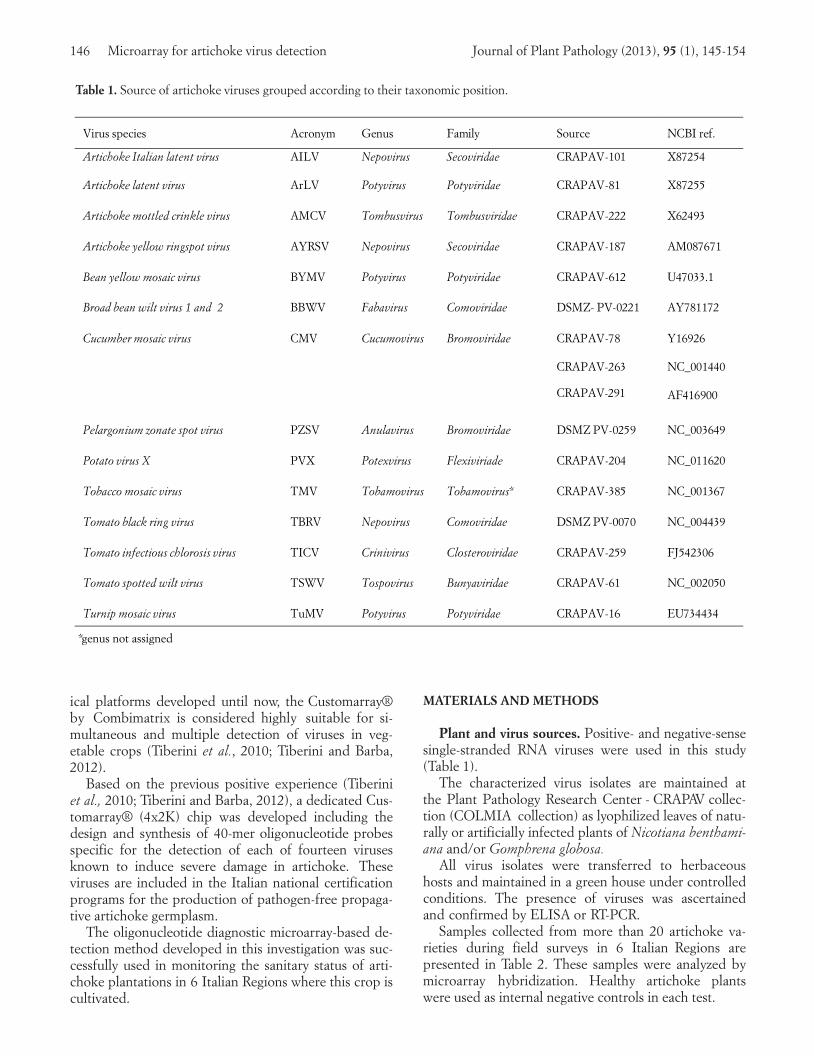
Table 1. Source of artichoke viruses grouped according to their taxonomic position.
Virus s pecies Acr o ny m G en us Fa m ily So urce NC B I r ef .
Ar tich o ke I ta lia n la te n t v ir u s A I L V N ep o virus S ec o viri da e C R A P A V - 101 X872 54
Ar tich o ke la ten t v ir u s A r L V Po tyviru s Po tyvirida e C R A P A V - 81 X872 55
Ar tich o ke mo ttled crinkle virus A M C V To mbu s virus To mbu s viri da e C R A P A V - 222 X624 93
Ar tich o ke ye llo w r in g s p o t v ir u s A Y R SV N ep o virus S ec o viri da e C R A P A V - 187 A M0876 71
Bea n ye llo w mo s a ic virus B YMV Po tyviru s Po tyvirida e C R A P A V - 612 U470 33.1
Br o a d bea n w ilt viru s 1 a n d 2 B B W V Fa ba virus C o mo viri da e DSM Z - P V- 022 1 A Y781 172
C u cu mber mo s a ic virus C MV C u cu mo virus Br o mo viri da e C R A P A V -78
C R A P A V - 2 63
C R A P A V -291
Y169 26
NC _00 1440
AF416 900
Pela r g o n iu m z o n a te s p o t v ir u s P Z SV An u la virus Br o mo viri da e DSM Z P V- 025 9 NC _00 3649
Po ta to virus X P VX Po texv ir u s Flexiv ir ia de C R A P A V - 204 NC _01 1620
To ba cc o mo s a ic virus T MV To ba mo virus To ba mo virus * C R A P A V - 385 NC _00 1367
To ma to bla ck r in g virus TB R V N ep o virus C o mo viri da e DSM Z P V- 007 0 NC _00 4439
To ma to in fectio u s ch lo r o s is virus T I C V C r in ivi r u s C lo s terovi r ida e C R A P A V - 259 FJ 54 2306
To ma to s p o tted w ilt viru s T SW V To s p o virus Bu n ya viri da e C R A P A V - 61 NC _00 2050
Tu r n ip mo s a ic virus T u MV Po tyviru s Po tyvirida e C R A P A V - 16 EU734 434
*genus not assigned
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 146

Journal of Plant Pathology (2013), 95 (1), 145-154 Tiberini and Barba 147
Probe Design. The nucleotide sequences of viruseswere retrieved from NCBI GenBank (see Table 3). Thesequences were aligned using VectorNTI 10 (Invitro-gen, Carlsbad, CA, USA) and used to design specific40-mer oligonucleotides for each virus isolate withEXIQON “Design oligonucleotides for expression ar-rays” software (http://oligo.lnatools.com) and Oligoar-ray 2.1 Software (Rouillard et al., 2003).
The 40-mer oligonucleotides were designed with thefollowing parameters: i) melting temperature range: 80-85°C; ii) secondary structure temperature threshold:70°C; iii) cross hybridization temperature threshold:65°C (15-80); iv) GC content: 40-60% (Chou el al.,2008).
To confirm the specificity of the probes designed byboth softwares, each probe was assessed by the nu-
cleotide identity score after BLASTN alignment. Inparticular, the oligonucleotide sequence was blastedagainst all known sequences of plant viruses to selectonly the ones showing no significant, or possibly thelowest, homology to other viruses except the specifictargets.
CustomArray® synthesis. A sectored oligonu-cleotide microarray chip containing four sets of 2,000features (4x2K) was synthesized at the Plant FunctionalGenomics Centre of University of Verona, Italy using aCustomArray® Synthesizer and following the manufac-turer instructions.
The oligonucleotide microarray included, in additionto the selected newly synthesized probes and negativeand positive controls provided by the manufacturer, a
Table 2. Artichoke samples collected from 6 Italian Regions during field surveys.
Sa m ple na m e Art icho ke Va riet y L o ca t io n Reg io nP o n t- zia Bianco Pertosa Pontecagnano Campania*P o n t- p etr a Petrelcina Pontecagnano Campania*P o n t- io r Capuanella Nera Pontecagnano Campania*P o n t- cio Capuanella Olivastra Pontecagnano Campania*P o n t- car Capuanella Carbone Pontecagnano Campania*P o n t- ca s t Castel San Lorenzo Pontecagnano Campania*P o n t - g e n n ar o Castellamare Pontecagnano Campania*P o n t- is o r Tondo di Paestum Pontecagnano Campania*P o n t- m o n t Montoro Pontecagnano Campania*P o n t- p asc Pascaiola Pontecagnano Campania*P is a - GR 1 Graleo Pisa Toscana**P is a - GR 2 Graleo Pisa Toscana**P is a - T EMA 1 Tema Pisa Toscana**P is a - T EMA 2 Tema Pisa Toscana**Mo n s - 1 Jesino Monsampolo Marche**Mo n s - 6 Ascolano Monsampolo Marche**Mo n s - 11 Urbisaglia Monsampolo Marche**Mo n s - 21 Castorano Monsampolo Marche**Mo n s - 26 Tardivo di Pesaro Monsampolo Marche**Mo n s - 37 Montelupone Monsampolo Marche**Mo n s - 41 Mazzaferrata Monsampolo Marche**T ar - B 1 C3 Tarquinia Lazio**T ar - AP 3 Apollo Tarquinia Lazio**T ar - C 5 Opal Tarquinia Lazio**T ar - 20- 1c Apollo Tarquinia Lazio**T ar - 1 C3 Tarquinia Lazio**T ar - 2 C3 Tarquinia Lazio**T ar - 3 C3 Tarquinia Lazio**T ar - 4 C3 Tarquinia Lazio**T ar - 5 C3 Tarquinia Lazio**T ar - 6 C3 Tarquinia Lazio**T o r - OS 6 S4 Torreinpietra Lazio**T o r - OS 8 C3 Torreinpietra Lazio**T o r - OS A 1 VT Torreinpietra Lazio**VP 16 Violetto di Provenza Bari Puglia*VP 18 Violetto di Provenza Bari Puglia*Sar 1 Spinoso Sardo Cagliari Sardinia**Sar 2 Spinoso Sardo Cagliari Sardinia*** R e g i o n s i t u a t e d i n Sou t h pa r t of I t a l y * * Re g i o n si t u a te d i n C e n t r a l p a r t of I ta l y
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 147

148 Microarray for artichoke virus detection Journal of Plant Pathology (2013), 95 (1), 145-154
selection of probes derived from our previous studies(Tiberini et al., 2010; Tiberini and Barba, 2012) con-sisting of: 27 for Cucumber mosaic virus (CMV), 20for Tobacco mosaic virus (TMV), 10 for TSWV, 8 forTomato infectious chlorosis virus (TICV), 10 for Potatovirus X (PVX), 10 for Pelargonium zonate virus(PZSV), 10 for Broad bean wilt virus (BBWV) and 10
for Tomato black ringspot virus (TBRV) (see Table 1).The oligonucleotide probes included in each of the foursub-arrays provided in the chip, were spatially organ-ized by virus, as reported in Fig. 1. In such a way, theprobes targeting the same virus, repeated up to 4times, are gathered in clusters spaced in each sub-array.
Table 3. Virus probes selected for good hybridization signal intensity given during the hybridization events. Name ofeach virus probe, its sequence, melting temperature, and genome position (start and end) are presented.
Pro b e Sequ ence T m Sta rt E nd
A I L V 8 T A AT AC GG AGGC AGG C AAAA A T GC AAGG G AG A T TT C GC AA 92 122 4 126 4
AI L V 9 C C AAC C GC AGT GC A G AC GGC C C AAA T T C C A C AC A T T GA C A 92 783 823
Ar L V 3 AA T GGGCT GT T GAA C AC GAGG C A T A T C TTT GA AG GAG A G A 90 265 305
Ar L V 5 C TT G TT C A T AAGG G A G C G C GT T A AGG C C T GGGA A T TT G T A 92 8 48
Ar L V 6 AACG AA T GGGT T GA T C C TGT T G T GGA G AA GT A T G AGG A C A 91 334 374
Ar L V 7 AC GT T GC C C T AAA GT C A T C T GT A GT T C A GT T A GT T C GAG A 90 461 501
Ar L V 8 T A T GA GG AC AA C T A C A T GC C AAG C GC AC T GAAC C GT GAA G 91 364 404
Ar L V 9 AC C C G AG A T GC A T GGT C AC AACT C T GC AC GC A T T AA G AAC 91 47 87
AYRSV 4 GC AA T GGC A G GG AA G A T GG AA GG AAGG C GC AAAG AAC C A T 90 742 782
AYRSV 10 T A T A T GC T C C AC T C G A T AAAA C G GC AA T C T AC T C TTC A T T 90 417 2 421 2
BYMV 3 GAAC AG A C G AGG A G AAC AC A G AG A G AC A C AC AGC A G G A G A 91 929 4 933 4
BYMV 4 C AA GGT G AA T GGAC AA T GA T GG A T GGT GA GG A G C AAGT GA 91 896 0 900 0
BYMV 7 C AA GG AAAG AAC AAG A G GAC AAGG C AG AA G C T AAGA T TT A 90 569 3 573 3
BYMV 10 AAG AAA GG AAAG GT G AAA G GT A C T AC A GT T GGCAT GGGG A 92 581 6 585 6
AM CV 1 GT A C AAAG C AGT GAA G AA G C G AGG A GGT AAG AAAC A G C A G 90 282 8 286 8
AM CV 2 T A GA T GGG A G A T GG A T TT G TTT GGA G A GG A G GGT GT T GA C 91 255 8 259 8
AM CV 4 T A AA T GGG A GT GGT T C A T GAAG A G AC GG C A G AG A G GT GA T 90 386 426
AM CV 12 GT G T GG TT C T GA G A T A C AT GA GG A C AGGG AAAA T AGAG T G 90 289 329
AM CV 14 C AA GG AAAG C T GGGG TTT C GGGA AAGT T G T A TTT A AG AG A 91 407 9 413 9
T u M V 4 GAG A C AAC AAG A T GGGT A G GG A GGT GT A T GGA G AT GA C G A 90 594 8 598 8
T u M V 8 GC A G C AC AA G AAG A GGG C G AACGT GAC AA T GAAG T A AACG 92 888 7 892 7
T u M V 9 T A T GGAA G A GT GGG AC AT A GGGG AA GT AA T GC T AA G AAAC 92 801 9 805 9
T u M V 12 GG AA T GT GGG T GA T GA T GG A C G G A G A T GA T C A GG T GGAA T 90 920 8 924 8
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 148

Journal of Plant Pathology (2013), 95 (1), 145-154 Tiberini and Barba 149
RNA extraction, cDNA synthesis of total plantRNA and fluorescent labeling of cDNA with cyanine 3or cyanine 5. Total RNA from infected and healthyplants, used in each hybridization event, was extractedusing the RNeasy Plant Mini kit (Qiagen, Hilden, Ger-many) according to the manufacturer’s instructions.
First-strand cDNA was prepared in reverse transcrip-tion (RT) reactions (final volume 30 µl) were preparedusing 2 µg of total RNA, 2 µM random hexamers and 6µM oligo dT, as described (Pasquini et al., 2008).
The cDNA was cleaned using an Amicon YM-100column (Millipore Corporation, Bedford MA, USA)dried and suspended in 50 µl of 0.1 M Na(HCO3)2buffer, pH 9, and transferred in a vial with an aliquotsof dried Cy3 or Cy5 dye (GE Healthcare, Little Chal-font U.K) previously suspended in dimethyl sulfoxide,divided into 10 aliquots and then dried. The cDNAand labeling mixture were incubated for 2 h at roomtemperature, and the unincorporated dyes were deacti-vated by adding 15 µl of 4 M hydroxylamine. The la-beled cDNA was cleaned using Cyscribe cleaning kit(GE Healthcare).
The evaluation of labeling efficiency was based onthe formula reported previously (Tiberini et al., 2010;Tiberini and Barba, 2012) by calculating the total num-ber of dye molecules incorporated per 1,000 nu-cleotides.
Hybridization, scanning and slide stripping. Thechip to be hybridized was inserted using a special hy-bridization clamp in a four time sectored chamberwhich enabled four independent and simultaneous hy-
bridization events to be performed. The hybridiza-tions, including scanning and slide stripping, were car-ried out according to Tiberini et al. (2010) followingthe manufacturer’s instructions by increasing the hy-bridization temperature from 42 to 50°C.
Taking into consideration the dye swapping, atleast two independent hybridization events were per-formed using all the targets considered in this study tobalance any interference due to the fluorophore used.
Scanning was performed using an Axon 4200A arrayscanner (Axon Instruments Ltd., Aberdeen, Scotland,UK), equipped with a 532 nm laser for Cy3 and a 635nm laser for Cy5 fluorescence measurements. Laserpower was fixed at 50% of its potential for bothlasers, while photomultiplier tube power ranged from45 to 65%, depending on signal intensity.
The stripping procedure was based on denaturationof DNA:RNA and/or DNA:DNA hybrids. This denatu-ration resulted in labeled targets being removed fromthe oligonucleotide probes and subsequently washed offthe CustomArray® while the original probes remainedon the slide.
Data analysis. Data were analyzed using the Gen-Pix 4.0 software and the background was subtractedas specified in the program by applying the followingdefault local subtraction method: the background for afeature is calculated from a region near the feature(three times around and excluding the neighboringspots) and its intensity is subtracted from the medianintensity of the feature analyzed. Initially, all signalsbelow 300 RU were considered negative.
Fig. 1. Schematic disposition and organization of all probes included in each sub-array. Probes are organ-ized virus per virus; each probe is repeated at least 5 times
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 149

The threshold to consider a reaction positive wasset at 3X (the median spot intensity plus the standarddeviation of the signal from all probes initially consid-ered negative).
Validation of the array. To assess the performanceand to validate the protocol and the array, eight morenon target samples were added to the 14 viruses tar-get samples. The non-targeted samples included:Pepino mosaic virus (PepMV), Plum pox virus (PPV),Prunus necrotic ringspot virus (PNRSV), Watermelonmosaic virus (WMV), Asparagus virus 1 (AV1), andthree healthy controls (artichoke, N. benthamiana,Chenopodium quinoa) were assayed in more than 30 ex-periments.
In each experiment two samples, previously labeledwith the two different fluorochromes Cy3 and Cy5,were combined in the hybridization mixture, mixing al-ternatively target and non-target were combined in thehybridization mixture, mixing alternatively target andnon-target samples.
The performance criteria included the following pa-rameters: diagnostic sensitivity; diagnostic specificityand diagnostic accuracy.
The diagnostic sensitivity is the proportion ofknown infected reference samples that test positive inthe assay; diagnostic specificity is the proportion of un-infected reference samples that test negative in the as-say; and diagnostic accuracy is the average of diagnos-tic sensitivity and diagnostic specificity which gives acomplete evaluation of the protocol parameters. The di-agnostic accuracy was calculated as the average valuesof 30 experiments. Results were classified as true posi-tive (TP) or true negative (TN) if they were in agree-ment with the oligonucleotide specificity determined inthe set-up of the chip. Alternately, they were classifiedas false positive (FP) or false negative (FN) if theydisagreed with the previously determined oligonu-cleotides specificity.
The performance criteria were calculated as follows:diagnostic sensitivity (D-SN) represents the ratio ofTP and the sum of TP and FN [TP/(TP + FN)];diagnostic specificity (D-SP) represents the ratio ofTN and the sum FP and TN [TN/(FP + TN)]; and di-agnostic accuracy (D- AC) is the average of D-SN andD-SP, i.e. [(TP + TN)/(TP + TN + FP + FN)].
The performance criteria were estimated for the besttwo probes of each virus included in the array. The val-ues obtained were averaged per single virus for the en-tire array.
RESULTS
Probe design and array synthesis. A total of 137virus oligonucleotide probes were selected, from 1,000
designed probes. The average percentage identity thateach selected probe shared with its own target was sig-nificantly high, reaching more than 98%.
The best 87 out of 137 virus probes were included onthe chip. Fifteen probes for each of the following virus-es: AILV, AYRSV, BYMV, AMCV, TuMV; 12 probesfor ArLV. Each sub-array contained the above men-tioned probes plus 115 probes tested and validated inour previous studies (Tiberini et al., 2010, Tiberini andBarba, 2012) as indicated in Materials and Methods.
The chip, divided into 4 sub-arrays, each with theoligonucleotide set of 193 virus probes, as reported inTable 3, spatially ordered according to the scheme re-ported in Fig. 1, enabled four different, independentand simultaneous hybridization events to be performedsuccessfully.
RNA extraction and cDNA labeling. Each total RNAextract obtained from infected plants was suitable to usein terms of concentration and molecular reactivity.
The cDNA indirect protocol was good and showed adye rate incorporation of 38-45 dye molecules/1000 nt,a value in line with the expected rate of 30-60 dye mole-cules per 1000 nt (Tiberini et al., 2010; Tiberini andBarba, 2012).
Virus detection: identification of good virus probesand quality of hybridization. In each hybridizationevent, the standard amount of 2 µg total RNA was suffi-cient for a good and clear positive hybridization signal.
All viruses were detected by microarray hybridizationwith a wide spectrum of signal intensity proportional tothe titer of viral RNA in the tested sample.
Each hybridization event identified several probesthat hybridized to their respective targets with good hy-bridization signal intensities, without cross-hybridiza-tion reactions and a low level of background (Table 3;Figg. 2-4).
The hybridization quality and intensity was not af-fected by the fluorochromes Cy3 and Cy5, incorporatedin cDNAs of plant samples infected with 14 viruses. Fig.2 shows microarray hybridization images obtained fromeach single cDNA synthesized from the 14 referenceisolates. In the images it is possible to observe the gen-eral aspect of the arrays where the samples, virus pervirus, were hybridized.
RNA from healthy control did not give any signifi-cant hybridization signal with viral probes. It hybridizedonly with plant ribosomal probes.
Virus detection of artichoke samples collected infield surveys. As mentioned above, 2 µg total RNA weresufficient for good and clear positive signals in each hy-bridization event.
Good hybridization signal intensities with low back-ground level were obtained from all virus-positive sam-
150 Microarray for artichoke virus detection Journal of Plant Pathology (2013), 95 (1), 145-154
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 150

Journal of Plant Pathology (2013), 95 (1), 145-154 Tiberini and Barba 151
Fig. 2. Microarray hybridization of cDNAs, labeled with Cy5 (red) and/or Cy3 (green) of all the 14 reference artichoke virus iso-lates included. The images represent the detection of the respective artichoke viruses using the designed virus-specific 40-meroligonucleotide probes. Panel a: the summarized images obtained in all the hybridization events; panel b: the same imageswith overlapped identification grid as reported in Fig. 1.
Fig. 3. Microarray hybridization of cDNAs labeled with Cy5 (red) and/or Cy3 (green) of four samples collected from field surveysin different Italian Regions. The images show detection of four artichoke viruses using virus-specific 40-mer oligonucleotideprobes. Panel a): sample VP 16 infected by AILV (Cy5-labeled); panel b): sample Tar-20-1c infected by AMCV (Cy5 labeled);panel c): sample Mons-6 infected by ArLV (Cy3-labeled); panel d): sample Tar-1 infected by TSWV (Cy5-labeled).
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 151

152 Microarray for artichoke virus detection Journal of Plant Pathology (2013), 95 (1), 145-154
Fig. 4. Microarray hybridization of cDNAs of two samples collected from filed surveys in different Italian Region. cDNAswere labeled with Cy5 (red) and/or Cy3 (green). Images represent the detection of artichoke viruses using the designed virus-specific 40-mer oligonucleotide probes. Panel a): summary image of the two samples which show the samples are infected byAILV, ArLV in mixed infection (Cy5 and Cy3- labeled); panel b): sample Tor- OS6 infected by AILV and ArLV in mixed infection(Cy5-labeled); panel c): sample Tor-OSA1 infected by ArLV in single infection (Cy3-labeled).
Table 4. Validation parameters (%) of each artichoke virus probe utilized.
496
497
499
Virus % Dia g nostic -S e nsit ivi t y
% Dia g nostic-S pe c ific it y
% Diagnostic-Accurancy
A ILV 96 99 97
Ar LV 98 98 99*
AMCV 99 98 99
AY RS V 97 99 98
B W YV 97 98 97
B BW V 98 98 98
TuMV 98 98 98
CMV 100 100 100
P ZS V 92 90 91
P VX 100 100 100
TMV 98 100 99
TSW V 98 100 99
TB RV 97 98 97
T I CV 100 100 100
Total 94.7± 5.8 95.2± 5.7 94.8± 5.6* Gr e y r ec ta n g le s i n d icate th e h ig h e s t dia g n o s tic ac cu r ac y D - AC
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 152

ples collected in field surveys. It was possible to identifyin several samples a single and/or mixed infection. ArLVhad the highest incidence of infection as compared toAILV, AMCV and TSWV. The latter virus has been re-ported in new outbreaks in Lazio Region, in particular.
In Fig. 3, panels a, b, c and d show images of microar-ray hybridization patterns obtained with four field sam-ples (samples 1, 2, 3 and 4), infected respectively by Ar-LV, AMCV, AILV, and TSWV in single infection for eachvirus. Samples were collected from the Tarquinia area(Lazio Region), where all the four viruses were detectedin the field. Two samples (5 and 6), collected in theabove mentioned area, were either singly- infected (sam-ple 5) or doubly-infected (sample 6) as shown in Fig. 4,panels a, b and c. Panel a presents summary image oftwo samples that are infected by AILV, ArLV in mixedinfection (cDNA labeled with Cy5 and Cy3). Panel bshows sample 6, whose cDNA was labeled with Cy5, in-fected with ArLV and AILV. Panel c shows sample 5,whose cDNA was labeled with Cy3, infected by ArLV.
In all hybridization experiments, including those offield samples, the hybridization background level waslow which indicated a high quality of hybridization. Nohybridizations were obtained for negative controls rep-resented by random oligonucleotides and other internalcontrols provided by the manufacture.
Validation of the array. Good hybridization signalswere obtained in all experiments with clear back-grounds. The diagnostic sensitivity values and the diag-nostic specificity were generally high, more than 97%and up to 100% (Table 4). Thirteen probes showed adiagnostic accuracy of 95% to 99%, whereas that ofPZSV probes was 91%. This low value was due to thelower value of diagnostic sensitivity because occasional-ly, the oligonucleotide probes did not hybridize to someinfected reference samples. The oligonucleotide probesfor ArLV, AMCV, CMV, PVX, TICV, TMV and TSWVshowed the highest diagnostic accuracy values (Table 4).
DISCUSSION
The multiple and simultaneous detection and identi-fication of the most economically important viruses af-fecting artichoke have been achieved by using oligonu-cleotide microarray-based technique that exploits theCombimatrix platform system as shown in this study.
The Combimatrix platform was particularly useful inthe initial screening of the most suitable probes amongthe 1,000 designed and evaluated in preliminary experi-ments. The selected oligonucleotide probes were highlyspecific as they identified and discriminated all arti-choke viruses utilized in this study. The high quality ofhybridization was confirmed by the following parame-ters: i) the background level was low, ii) no hybridiza-
tion occurred with negative controls and other controlsprovided by the manufacture, iii) good quality hy-bridization signal was observed between probes includ-ed by manufacture as internal controls and their targets.
All the selected artichoke virus species which belongto different genera and families, were specifically andclearly detected following the described methodologyno matter their genomic organization, as RNA positive(i.e. ArlV) and/or RNA negative strand (i.e. TSWV).
The array was successfully applied to the detection ofviruses affecting fresh artichoke samples collected dur-ing routine field surveys indicating its potential use formass scale diagnosis. This aspect is of particular impor-tance as artichoke is a vegetative tissue rich in oxidativechemicals which in some cases interfere with diagnosticprotocol based on the use of serological kits and/or de-crease the quality of total RNA to be tested in RT-PCR.
In addition, the short protocol set-up which was usedfor testing field samples yielded results in no more than2 days which is the time generally needed to get an assayresult from ELISA and/or RT-PCR.
In particular, the overlapping with ELISA in terms ofsensitivity (Tiberini and Barba, 2012) and 2 workingdays, highlighted that the use of the Combimatrix plat-form for artichoke viruses detection and identification islimited only by the spectrum of probes present on thearray. In combination with previous studies (Tiberini etal., 2010; Tiberini and Barba, 2012) and the presentstudy which showed its routine use for the detection of14 virus species in artichoke germplasm, the Combima-trix platform as a microarray diagnostic method hasbeen ascertained because of its validation parameters, inparticular by the good diagnostic repeatability, sensitivi-ty and specificity values. These data confirmed the use-fulness of this technique, Combimatrix platform-based,in the diagnosis of viruses, belonging to different generaand families.
The Combimatrix microarray platform was suitablein multiple and simultaneous detection of the major ar-tichoke viruses present in single and mixed infection, al-lowing to simplify the sample preparation and to reducethe cost of the method. The use of an array divided into4 identical sectors and the re-use of the same chip forfurther hybridizations, including the use of both fluo-rochromes in the same array, exploit the possibility ofre-using the same chip at least 8 times and testing 64samples. In view of above, this array showed the poten-tial to fulfill the requirements of accurate disease diag-nosis as it has the following advantages: i) a great con-trol over the probe design as oligonucleotide probescould be designed using all available sequence informa-tion; ii) the array can be constructed with higher densityand complexity, because of its synthesis in situ; iii) theease of in silico design and the high specificity ofoligonucleotides enable the discrimination of the mixedinfection by different pathogens and even similar
Journal of Plant Pathology (2013), 95 (1), 145-154 Tiberini and Barba 153
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 153

strains; iv) the possibility to use the array with field sam-ples and complex matrices.
In conclusion, the array analyzed in our study may beused in many applications, such as certification andquarantine programs, epidemiological studies, transmis-sion and host range and disease management and con-trol with direct effect on security of the food supply.
REFERENCES
Abdullahi A., Gryshan Y., Rott M., 2011. Single-channeloligonucleotide microarray for amplification-free detectionof grapevine viruses. Journal of Virological Methods 178:1-15.
Abdullahi I., Koebler M., Stachewicz H., Winter S., 2005.The 18s rDNA sequence of Synchyntium endobioticumand its utility in microarrays for the simultaneous detectionof fungal and viral pathogens of potato. Applied Microbiol-ogy and Biotechnology 68: 368-375.
Barba M., Hadidi A., 2007. DNA microarrays: technology, ap-plications, and potential applications for the detection ofplant viruses and virus-like pathogens. In: Rao G.P.,Valverdi R.A., Dovas C.I. (eds). Techniques in Diagnosis ofPlant Viruses, pp. 227-247. Studium Press LLC, HoustonTX, USA.
Barba M., Hadidi A., 2011a. DNA microarrays and other fu-ture trends in detection and typing of viruses, viroids andphytoplasmas. In: Hadidi A., Barba M., Candresse T., Jelk-mann W. (eds). Virus and Virus-Like Diseases of Pomeand Stone Fruits, pp. 363-372. The American Phytopatho-logical Society Press, St. Paul, MN, USA.
Barba M., Hadidi A., 2011b. DNA microarray-based detec-tion and identification of plant viruses and otherpathogens. Current Topics in Plant Biology 12: 203-215.
Boonham N., Walsh K., Smith P., Madagan K., Graham I.,Barker I., 2003. Detection of potato viruses using microar-ray technology: towards a generic method for plant viral dis-ease diagnosis. Journal of Virological Methods 108: 181-187.
Bystrická D., Lenz O., Mráz I., Dedic P., Síp M., 2003. DNAmicroarray: parallel detection of potato viruses. Acta Vi-rologia 47: 41-44.
Bystricka D., Lenz O., Mraz I., Piherova L., Kmoch S., SipM., 2005. Oligonucleotide-based microarray: a new im-provement in microarray detection of plant viruses. Journalof Virological Methods 128: 176-182.
Deyong Z., Willingmann P., Heinze C., Adam G., Pfunder M.,Frey B., Frey J.E., 2005. Differetiation of Cucumber mosaicvirus isolates by hybridization to oligonucleotides in a mi-croarray format. Journal of Virological Methods 123: 101-108.
Engel E.A., Escobar P.F, Rojas L.A., Rivera P.A., Fiore N.,2010. A diagnostic oligonucleotide microarray for simulta-neous detection of grapevine viruses. Journal of VirologicalMethods 163: 445-451.
Gambino G., Gribaudo I., 2006. Simultaneous detection ofnine grapevine viruses by multiplex reverse transcription-polymerase chain reaction with coamplification of a plantRNA as internal control. Phytopathology 96: 1223-1229.
Hadidi A., Czosnek H., Barba M., 2004. DNA microarraysand their potential applications for the detection of plantviruses, viroids, and phytoplasmas. Journal of Plant Pathol-ogy 86: 97-104.
Hadidi A., Barba, M., 2008. DNA microarrays: 21st centurypathogen detection. Acta Horticulturae 781: 331-339.
Lee G., Min B.E., Kim C.S., Choi S.H., Harn C.H., Kim S.U.,Ryu K.H., 2003. Plant virus cDNA chip hybridization fordetection and differentiation of four cucurbit-infection To-bamoviruses. Journal of Virological Methods 110: 19-24.
Lenz O., Petrzik K., Spak J., 2008. Investigating the sensitivityof a fluorescence-based microarray for the detection offruit-tree virus. Journal of Virological Methods 148: 96-105.
Li Y., Li T., Liu S., Qiu M., Han Z., Jiang Z., Li R., Ying K.,Xie Y., Mao Y., 2004. Systematic comparison of the fidelityof aRNA, mRNA and T-RNA on gene expression profilingusing cDNA microarray. Journal of Biotechnology 107: 19-27.
Nicolaisen M., 2011. An oligonucleotide-based microarray fordetection of plant RNA viruses. Journal of VirologicalMethods 173: 134-143.
Pasquini G., Barba M., Hadidi A., Faggioli F., Negri R., SobolI., Tiberini A., Caglayan K., Mazyad H., Anfoka G., Ghan-im M., Zeidan M., Czosnek H., 2008. Oligonucleotide mi-croarray-based detection and genotyping of Plum poxvirus. Journal of Virological Methods 147: 118-126.
Pasquini G., Faggioli F., Luigi M., Gentili A., Hadidi A.,Canini I, Gabriele L., Czosnek H., Tiberini A., CaglayanK., Mazyad H., Anfoka K., Barba M., 2010. Validation of amicroarray protocol for detection and genotyping isolatesof Plum pox virus. Julius-Kuhn-Archiv 427: 56-60.
Rouillard J.M., Zuker M., Gularil E., 2003. OligoArray 2.0:design of oligonucleotide probes for DNA microarrays us-ing a thermodynamic approach. Nucleic Acids Research 31:3057-3062.
Tiberini A., Tomassoli L., Barba M., Hadidi A., 2010.Oligonucleotide microarray-based and identification of tenmajor tomato viruses. Journal of Virological Methods 168:133-140.
Tiberini A., Barba M., 2012. Optimization and improvementof oligonucleotide microarray-based detection of tomatoviruses and pospiviroids. Journal of Virological Methods185: 43-51.
Wei T., Pearson M.N., Blohm D., Nolte M., Armstrong K.,2009. Development of a short oligonucleotide microarrayfor the detection and identification of multiple potyviruses.Journal of Virological Methods 162: 109-118.
Zhang Y., Yin J., Li G., Li M., Huang X., Chen H., WenjunZhao W., Zhu S.F., 2010. Oligonucleotide microarray witha minimal number of probes for the detection and identifi-cation of thirteen genera of plant viruses. Journal of Viro-logical Methods 167: 53-60.
154 Microarray for artichoke virus detection Journal of Plant Pathology (2013), 95 (1), 145-154
Received July 12, 2012Accepted August 22, 2012
016_JPP1337RP(Tiberini)_145col 26-03-2013 14:59 Pagina 154
Copyright © 2022 FDOKUMEN




















