
Assessing unmodified 70-mer oligonucleotide probe performance on glass-slide microarrays

wwwelseviercomlocateclinchim
Clinica Chimica Acta 36
Review
Highly parallel microbial diagnostics using oligonucleotide microarrays
Alexander Loy a1 Levente Bodrossy b
a Department of Microbial Ecology University of Vienna A-1090 Vienna Austriab Department of BioresourcesMicrobiology ARC Seibersdorf research GmbH A-2444 Seibersdorf Austria
Received 3 April 2005 accepted 5 May 2005
Available online 26 August 2005
Abstract
Oligonucleotide microarrays are highly parallel hybridization platforms allowing rapid and simultaneous identification of many
different microorganisms and viruses in a single assay In the past few years researchers have been confronted with a dramatic increase in
the number of studies reporting development andor improvement of oligonucleotide microarrays for microbial diagnostics but use of the
technology in routine diagnostics is still constrained by a variety of factors Careful development of microarray essentials (such as
oligonucleotide probes protocols for target preparation and hybridization etc) combined with extensive performance testing are thus
mandatory requirements for the maturation of diagnostic microarrays from fancy technological gimmicks to robust and routinely
applicable tools
D 2005 Elsevier BV All rights reserved
Keywords Microbial diagnostics Microarray Hybridization Oligonucleotide Marker gene
Contents
106
110
110
110
111
114
114
114
1 Basic concept
2 Microarray hybridization formats
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
32 Development and optimisation of microarray probe sets specificity sensitivity and uniformity
33 Further selected strategies to increase specificity andor sensitivity
4 Data analysis and quantification potential
5 Diagnostic applications
6 Conclusions and outlook
115
Acknowledgements 116
References 116
1 Basic concept
DNA microarray (microchip biochip gene chip) tech-
nology allows the parallel analysis of highly complex gene
0009-8981$ - see front matter D 2005 Elsevier BV All rights reserved
doi101016jcccn200505041
Corresponding author Tel +43 50550 3548 fax +43 50550 3444
E-mail addresses loymicrobial-ecologynet (A Loy)
LeventeBodrossyarcsacat (L Bodrossy)
URLs wwwmicrobioal-ecologynet (A Loy) wwwarcsacatuub
microbiology (L Bodrossy)1 Tel +43 1 4277 54207 fax +43 1 4277 54389
mixtures in a single assay and thus symbolises as no other
method the (post-)genomic era of high-throughput science
While microarrays initially emerged as tools for genome-
wide expression analysis and are nowadays routinely used
for this purpose they are also increasingly being developed
for diagnostic applications [1] Microbial diagnostic micro-
arrays (MDMs) consist of nucleic acid probe sets with each
probe being specific for a given strain subspecies species
genus or higher taxon [1] A characteristic MDM experi-
ment is depicted in Fig 1 MDMs fall into two distinct
3 (2006) 106 ndash 119
Fig 1 Typical steps of a diagnostic microarray experiment
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 107
categories according to their intended use Environmental
MDMs [23] are primarily applied in environmental and
industrial microbiology to obtain a picture of the structure of
the microbial community being analysed Requirements for
this class of MDMs are the parallel detection of many
microorganisms at the level of species genus or a higher
taxon and the potential for some level of quantification
Detectionidentification MDMs typically applied in clin-
ical veterinary food and biodefense microbiology [45]
must usually enable the reliable detection andor identifica-
tion at the speciessubspeciesstrain level of one or a few
microbes out of many that may be present in a sample It
Table 1
Selected applications of oligonucleotide microarrays for microbial diagnostics
Platform Target organisms Marker gene(s) Highest
phylogenetic
resolution
L 1) No of probes Field 2) No of
samples
analysed
Sample type Reference
Planar glass slide Entamoeba histolytica
E dispar Giardia lamblia
Cryptosporidium parvum
Various Species subtypes 20ndash30 88 CF ndash ndash [79]
Planar glass slide Cryptosporidium hsp70 Isolates 15 68 E ndash ndash [94]
Affymetrix Bacteria 16S rRNA Higher level
bacterial taxa
20 31179 E 1 Air filtrate [17]
Planar glass slide Bacteria involved in
nitrification denitrification
nitrogen fixation methane
oxidation and sulfite reduction
nirS nirK amoA nifH
pmoA dsrAB
Species 50 763 E 1 Marine sediment [47]
Planar glass slide 3) Bacteria causing abortion and
sterility in mares
23S rRNA Species subspecies 24ndash32 32 C 21 Cervical swabs [21]
Planar glass slide Bacterial fish pathogens 16S rRNA Species 22ndash31 18 CF ndash ndash [18]
Planar glass slide 4) Bacterial fish pathogens cyt rpoN gyrB toxR ureC
dly vapA fatA A plassal
Subspecies 25 9 EF ndash ndash [95]
Planar glass slide Campylobacter jejuni C coli
C lari C upsaliensis
fur glyA cdtABC ceuBC fliY Species 17ndash35 74 C 16+6 Isolates and
mixed cultures
[76]
Planar glass slide 4) Campylobacter spp 16S rRNA 16Sndash23S
intergenic spacer
Campylobacter-specific genes
Species 27ndash35 5 CF 10+65 Chicken cloacal
swabs
[9697]
Planar glass slide 5) Cyanobacteria 16S rRNA Above genus 20ndash29 19 E 1 Lake water [98]
Planar glass slide Enterococcus 16S and 23S rRNA Species 41 18 CF 2 Milk [99]
Planar glass slide Escherichia coli Shigella Sal-
monella
gyrB Species 15ndash19 10 C ndash ndash [27]
Planar glass slide Selected taxa of marine bacter-
ioplankton
16S rRNA Higher taxa 15ndash20 21 E 1 Sea water [58]
Planar glass slide Listeria iap hly inlB plcA plcB clpE Species 17ndash33 132 C ndash ndash [36]
Planar glass slide Listeria spp Campylobacter
spp Staphylococcus aureus
Clostridium perfringens
Various Species 17ndash35 178 BF ndash ndash [100]
Planar glass slide Marine bacterioplankton 16S rRNA Species 15ndash20 21 E 1 Sea water [101]
Planar glass slide Methanotrophs pmoA Species subspecies 17ndash27 61 E gt100 Landfill cover soil [139]
Planar glass slide Mycobacterium spp gyrB Species 13ndash15 28 C 40 Human sputum [102]
Planar glass slide 6) Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash16 11) 18 C ndash ndash [103]
3D-surface Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash23 43 C 31 Clinical samples [90]
Planar glass slide 4) Pathogenic Vibrio spp vvh viuB ompU toxR tcpI
hlyA tlh tdh trh etc
Species 30ndash32 13 CF 30 Oyster [82]
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
108
Planar glass slide 7) Quinolone-resistant
Escherichia coli
gyrA Genotype 19 42 C ndash ndash [104]
Planar glass slide Respiratory bacterial pathogens gyrB parE Species 20ndash24 27 C 94 Middle ear fluid
throat swabs
[105]
Planar glass slide Rhodocyclales 16S rRNA Species 18 79 E 1 Activated sludge [56]
3D-surface Some Bacillus spp 16S rRNA Species 15ndash23 30 BC ndash ndash [64]
Planar glass slide 7) Some Bacillus spp 16Sndash23S intergenic spacer Species 18ndash22 42 BC ndash ndash [22]
Planar glass slide Sulphate-reducing prokaryotes 16S rRNA Species 18 132 CE 6+8 Hypersaline
cyanobacterial
lake mat periodontal
tooth pocket fen soil
[15106]
3D-surface Thermophilic anaerobic
Archaea and Bacteria
16S rRNA Genus 17ndash20 17 E 3 Oil reservoir
formation waters
[77]
Affymetrix 8) Major potential biowarfare agents (bacteria viruses
eukaryotes)
Various Species 20 53660 BC 2 Air filtrate [107]
Planar glass slide Acute respiratory
disease-associated
Adenoviruses
E1A fibre hexon Serotypes 60ndash72 36 C 19 Throat swabs nasal
wash samples
[80]
Planar glass slide Human group A rotaviruses VP7 Genotype 18ndash26 50 C ndash ndash [108]
Planar glass slide Human papillomaviruses E1 Types 20 51 C 130 Clinical samples [109]
Planar glass slide 5) Human papillomaviruses Various Types 14ndash28 10 C 100 Clinical samples [110]
Planar glass slide Human papillomaviruses Various Types 30 27 C 73 Clinical samples
(tonsillar cancer)
[111]
Planar glass slide Influenza viruses Various Subtype 17ndash29 476 C ndash ndash [112]
3D-surface 9) Influenza viruses Various (ao hemagglutinin
and neuraminidase genes)
Subtypes 45ndash65 29 C ndash ndash [113]
Planar glass slide Orthopoxviruses C23LB29R gene ORF 62
(varicella-zoster)
Types 13ndash21 57 BC ndash ndash [114]
3D-surface Orthopoxviruses crmB Species 12ndash16 15 C ndash ndash [115]
Planar glass slide 10) Viruses Various Serotype 70 1600 C 6 Clinical samples [50]
1) Length of probe [nt]2) B biodefense C clinical microbiology F food microbiology E environmental microbiology3) On-Chip PCR4) Multiplex PCR6) DNA ligation detection reaction5) SOLAC-short oligo ligation assay on chip8) Perfect match and single mismatch probes used7) 3ndash10 diagnostic regions were identified per microorganism and each one of them was covered by 100ndash300 probes9) Flow-Thru Chip10) 70mer discriminatory oligonucleotides were designed based on all published viral genome sequences11) Probes comprised of 5 specific nucleotides (participating in specific ligation)+10ndash11 spacer T residues
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
109
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119110
should be noted however that there are also medical
microbiology contexts where (as described for environ-
mental MDMs) the microbial community structure has to be
addressed eg the composition of the human gastro-
intestinal tract microbiota
Although recent studies (Table 1) undoubtedly demon-
strate that MDMs are valuable tools for identification of
microorganisms and viruses in a highly parallel fashion
their use for routine diagnostics is still hampered by a lack
of standardisation (regarding factors such as probes target
genes hybridization platforms protocols and data analy-
sis) and insufficient evaluation of newly developed
MDMs
In this review we summarise recent progress in the
MDM field Many (seemingly conflicting) parameters and
steps have to be integrated during microarray design and
application in order to fulfil the potential of MDMs as
high-throughput screening tools for routine diagnostic
purposes We thus highlight crucial points related to
development and evaluation of MDMs The key to a
validated MDM and consequently to reliable results is
rigorous in silico and in vitro performance testing
Furthermore we exemplarily show recent technological
advances which should further improve the use of MDMs
in the near future
2 Microarray hybridization formats
Although many diverse microarray platforms have
become available over the past few years [6] only a limited
number of solid supports are currently used for MDMs For
Affymetrix microarrays (wwwaffymetrixcom) oligonu-
cleotide probes are synthesised directly onto the microarray
surface by employing specific masks and the photolithog-
raphy method This approach enables a very high probe
density (well over 100000 probes per microarray) The
high price low flexibility and lack of a suitably high
number of validated oligonucleotide probes currently limit
the wide application of Affymetrix GeneChips in microbial
diagnostics In contrast the NimbleGen technology
(wwwnimblegencom) uses digital micromirrors instead of
physical masks to guide on-chip probe synthesis allowing a
more flexible design of custom-made high-density micro-
arrays Specific three-dimensional microarray formats such
as the gel-pad platform (wwwbiochipruen) [78] or the
PamGene system (wwwpamgenecom) [910] are coupled
with appropriate hybridization and detection devices that
offer the option to record hybridization and dissociation
events in real-time Melting curves for all probes on a
microarray can thus be rapidly established making the
development of validated probe sets significantly easier
[1112] However these systems are currently available
only in a few laboratories
The pioneer microarray format and still the most widely
used miniaturised solid support for the covalent immobili-
sation of probes are planar 13 in glass slides Oligonu-
cleotides are in most cases tethered via their 5V ends to
reactive groups on the coating layer of the glass surface The
establishment of microarray core facilities (including micro-
array spotting and detection devices) in many laboratories
and the general utility flexibility and moderate price of
planar glass microarrays are mainly responsible for the
success of this standard format
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
Two main parameters affect the resolution of a diagnostic
microarray assay (i) the degree of conservation of the
marker gene and (ii) the length of the oligonucleotide used
as a probe to target it [13]
The most widely employed target molecule for the
detection and phylogenetic analysis of microorganisms is
the small-subunit ribosomal RNA (SSU rRNA) and its gene
[714ndash18] The popularity of the SSU rRNA is reflected in
the existence of large and regularly updated sequence [19
20] and probe databases (httpwwwmicrobial-ecologynet
probebase) [16] for this target molecule The main
limitation of using the SSU rRNA (gene) as a marker in
microarray assays is that resolution below the species level is
generally not possible owing to high overall sequence
conservation Because differentiation of strains is often
essential in clinical diagnostics in order to initiate appro-
priate treatment of an infection less-conserved target
molecules are needed Potential probe targets which offer
strain-level resolution include (i) the large-subunit ribosomal
RNA (LSU rRNA) [21] (ii) the SSU-LSU rRNA intergenic
spacer region [22] (iii) house-keeping genes (eg rpoB
[23ndash25] gyrA [26] gyrB [27] recA [28] tuf [2930] groEL
[31] atpD [30] ompA gapA pgi [32] tmRNA [33]) (iv)
virulence genes [34ndash37] (v) antibiotic resistance genes
[938] (vi) functional genes encoding enzymes responsible
for specific metabolic traits [139ndash42] etc (for a detailed
list please refer to wwwarcsacatuubmicrobiology)
However individual sequence databases for these alternative
markers if they exist at all currently contain considerably
fewer entries than the SSU rRNA databases constraining the
development and evaluation of encompassing probe sets for
microarrays [43]
The hybridization properties of long oligonucleotide
probes (typically 50ndash100mer) which show pronounced
hysteresis (higher temperatures for dissociation than for
association) are fundamentally different from those of short
probes (typically 15ndash30mer) [4445] While in principle
short oligonucleotides allow the discrimination of single
nucleotide differences under optimal conditions (see below
for further details) this does not hold true for long
oligonucleotides Their threshold for differentiation is
approximately 75ndash87 sequence similarity [414647]
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

Fig 1 Typical steps of a diagnostic microarray experiment
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 107
categories according to their intended use Environmental
MDMs [23] are primarily applied in environmental and
industrial microbiology to obtain a picture of the structure of
the microbial community being analysed Requirements for
this class of MDMs are the parallel detection of many
microorganisms at the level of species genus or a higher
taxon and the potential for some level of quantification
Detectionidentification MDMs typically applied in clin-
ical veterinary food and biodefense microbiology [45]
must usually enable the reliable detection andor identifica-
tion at the speciessubspeciesstrain level of one or a few
microbes out of many that may be present in a sample It
Table 1
Selected applications of oligonucleotide microarrays for microbial diagnostics
Platform Target organisms Marker gene(s) Highest
phylogenetic
resolution
L 1) No of probes Field 2) No of
samples
analysed
Sample type Reference
Planar glass slide Entamoeba histolytica
E dispar Giardia lamblia
Cryptosporidium parvum
Various Species subtypes 20ndash30 88 CF ndash ndash [79]
Planar glass slide Cryptosporidium hsp70 Isolates 15 68 E ndash ndash [94]
Affymetrix Bacteria 16S rRNA Higher level
bacterial taxa
20 31179 E 1 Air filtrate [17]
Planar glass slide Bacteria involved in
nitrification denitrification
nitrogen fixation methane
oxidation and sulfite reduction
nirS nirK amoA nifH
pmoA dsrAB
Species 50 763 E 1 Marine sediment [47]
Planar glass slide 3) Bacteria causing abortion and
sterility in mares
23S rRNA Species subspecies 24ndash32 32 C 21 Cervical swabs [21]
Planar glass slide Bacterial fish pathogens 16S rRNA Species 22ndash31 18 CF ndash ndash [18]
Planar glass slide 4) Bacterial fish pathogens cyt rpoN gyrB toxR ureC
dly vapA fatA A plassal
Subspecies 25 9 EF ndash ndash [95]
Planar glass slide Campylobacter jejuni C coli
C lari C upsaliensis
fur glyA cdtABC ceuBC fliY Species 17ndash35 74 C 16+6 Isolates and
mixed cultures
[76]
Planar glass slide 4) Campylobacter spp 16S rRNA 16Sndash23S
intergenic spacer
Campylobacter-specific genes
Species 27ndash35 5 CF 10+65 Chicken cloacal
swabs
[9697]
Planar glass slide 5) Cyanobacteria 16S rRNA Above genus 20ndash29 19 E 1 Lake water [98]
Planar glass slide Enterococcus 16S and 23S rRNA Species 41 18 CF 2 Milk [99]
Planar glass slide Escherichia coli Shigella Sal-
monella
gyrB Species 15ndash19 10 C ndash ndash [27]
Planar glass slide Selected taxa of marine bacter-
ioplankton
16S rRNA Higher taxa 15ndash20 21 E 1 Sea water [58]
Planar glass slide Listeria iap hly inlB plcA plcB clpE Species 17ndash33 132 C ndash ndash [36]
Planar glass slide Listeria spp Campylobacter
spp Staphylococcus aureus
Clostridium perfringens
Various Species 17ndash35 178 BF ndash ndash [100]
Planar glass slide Marine bacterioplankton 16S rRNA Species 15ndash20 21 E 1 Sea water [101]
Planar glass slide Methanotrophs pmoA Species subspecies 17ndash27 61 E gt100 Landfill cover soil [139]
Planar glass slide Mycobacterium spp gyrB Species 13ndash15 28 C 40 Human sputum [102]
Planar glass slide 6) Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash16 11) 18 C ndash ndash [103]
3D-surface Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash23 43 C 31 Clinical samples [90]
Planar glass slide 4) Pathogenic Vibrio spp vvh viuB ompU toxR tcpI
hlyA tlh tdh trh etc
Species 30ndash32 13 CF 30 Oyster [82]
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
108
Planar glass slide 7) Quinolone-resistant
Escherichia coli
gyrA Genotype 19 42 C ndash ndash [104]
Planar glass slide Respiratory bacterial pathogens gyrB parE Species 20ndash24 27 C 94 Middle ear fluid
throat swabs
[105]
Planar glass slide Rhodocyclales 16S rRNA Species 18 79 E 1 Activated sludge [56]
3D-surface Some Bacillus spp 16S rRNA Species 15ndash23 30 BC ndash ndash [64]
Planar glass slide 7) Some Bacillus spp 16Sndash23S intergenic spacer Species 18ndash22 42 BC ndash ndash [22]
Planar glass slide Sulphate-reducing prokaryotes 16S rRNA Species 18 132 CE 6+8 Hypersaline
cyanobacterial
lake mat periodontal
tooth pocket fen soil
[15106]
3D-surface Thermophilic anaerobic
Archaea and Bacteria
16S rRNA Genus 17ndash20 17 E 3 Oil reservoir
formation waters
[77]
Affymetrix 8) Major potential biowarfare agents (bacteria viruses
eukaryotes)
Various Species 20 53660 BC 2 Air filtrate [107]
Planar glass slide Acute respiratory
disease-associated
Adenoviruses
E1A fibre hexon Serotypes 60ndash72 36 C 19 Throat swabs nasal
wash samples
[80]
Planar glass slide Human group A rotaviruses VP7 Genotype 18ndash26 50 C ndash ndash [108]
Planar glass slide Human papillomaviruses E1 Types 20 51 C 130 Clinical samples [109]
Planar glass slide 5) Human papillomaviruses Various Types 14ndash28 10 C 100 Clinical samples [110]
Planar glass slide Human papillomaviruses Various Types 30 27 C 73 Clinical samples
(tonsillar cancer)
[111]
Planar glass slide Influenza viruses Various Subtype 17ndash29 476 C ndash ndash [112]
3D-surface 9) Influenza viruses Various (ao hemagglutinin
and neuraminidase genes)
Subtypes 45ndash65 29 C ndash ndash [113]
Planar glass slide Orthopoxviruses C23LB29R gene ORF 62
(varicella-zoster)
Types 13ndash21 57 BC ndash ndash [114]
3D-surface Orthopoxviruses crmB Species 12ndash16 15 C ndash ndash [115]
Planar glass slide 10) Viruses Various Serotype 70 1600 C 6 Clinical samples [50]
1) Length of probe [nt]2) B biodefense C clinical microbiology F food microbiology E environmental microbiology3) On-Chip PCR4) Multiplex PCR6) DNA ligation detection reaction5) SOLAC-short oligo ligation assay on chip8) Perfect match and single mismatch probes used7) 3ndash10 diagnostic regions were identified per microorganism and each one of them was covered by 100ndash300 probes9) Flow-Thru Chip10) 70mer discriminatory oligonucleotides were designed based on all published viral genome sequences11) Probes comprised of 5 specific nucleotides (participating in specific ligation)+10ndash11 spacer T residues
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
109
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119110
should be noted however that there are also medical
microbiology contexts where (as described for environ-
mental MDMs) the microbial community structure has to be
addressed eg the composition of the human gastro-
intestinal tract microbiota
Although recent studies (Table 1) undoubtedly demon-
strate that MDMs are valuable tools for identification of
microorganisms and viruses in a highly parallel fashion
their use for routine diagnostics is still hampered by a lack
of standardisation (regarding factors such as probes target
genes hybridization platforms protocols and data analy-
sis) and insufficient evaluation of newly developed
MDMs
In this review we summarise recent progress in the
MDM field Many (seemingly conflicting) parameters and
steps have to be integrated during microarray design and
application in order to fulfil the potential of MDMs as
high-throughput screening tools for routine diagnostic
purposes We thus highlight crucial points related to
development and evaluation of MDMs The key to a
validated MDM and consequently to reliable results is
rigorous in silico and in vitro performance testing
Furthermore we exemplarily show recent technological
advances which should further improve the use of MDMs
in the near future
2 Microarray hybridization formats
Although many diverse microarray platforms have
become available over the past few years [6] only a limited
number of solid supports are currently used for MDMs For
Affymetrix microarrays (wwwaffymetrixcom) oligonu-
cleotide probes are synthesised directly onto the microarray
surface by employing specific masks and the photolithog-
raphy method This approach enables a very high probe
density (well over 100000 probes per microarray) The
high price low flexibility and lack of a suitably high
number of validated oligonucleotide probes currently limit
the wide application of Affymetrix GeneChips in microbial
diagnostics In contrast the NimbleGen technology
(wwwnimblegencom) uses digital micromirrors instead of
physical masks to guide on-chip probe synthesis allowing a
more flexible design of custom-made high-density micro-
arrays Specific three-dimensional microarray formats such
as the gel-pad platform (wwwbiochipruen) [78] or the
PamGene system (wwwpamgenecom) [910] are coupled
with appropriate hybridization and detection devices that
offer the option to record hybridization and dissociation
events in real-time Melting curves for all probes on a
microarray can thus be rapidly established making the
development of validated probe sets significantly easier
[1112] However these systems are currently available
only in a few laboratories
The pioneer microarray format and still the most widely
used miniaturised solid support for the covalent immobili-
sation of probes are planar 13 in glass slides Oligonu-
cleotides are in most cases tethered via their 5V ends to
reactive groups on the coating layer of the glass surface The
establishment of microarray core facilities (including micro-
array spotting and detection devices) in many laboratories
and the general utility flexibility and moderate price of
planar glass microarrays are mainly responsible for the
success of this standard format
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
Two main parameters affect the resolution of a diagnostic
microarray assay (i) the degree of conservation of the
marker gene and (ii) the length of the oligonucleotide used
as a probe to target it [13]
The most widely employed target molecule for the
detection and phylogenetic analysis of microorganisms is
the small-subunit ribosomal RNA (SSU rRNA) and its gene
[714ndash18] The popularity of the SSU rRNA is reflected in
the existence of large and regularly updated sequence [19
20] and probe databases (httpwwwmicrobial-ecologynet
probebase) [16] for this target molecule The main
limitation of using the SSU rRNA (gene) as a marker in
microarray assays is that resolution below the species level is
generally not possible owing to high overall sequence
conservation Because differentiation of strains is often
essential in clinical diagnostics in order to initiate appro-
priate treatment of an infection less-conserved target
molecules are needed Potential probe targets which offer
strain-level resolution include (i) the large-subunit ribosomal
RNA (LSU rRNA) [21] (ii) the SSU-LSU rRNA intergenic
spacer region [22] (iii) house-keeping genes (eg rpoB
[23ndash25] gyrA [26] gyrB [27] recA [28] tuf [2930] groEL
[31] atpD [30] ompA gapA pgi [32] tmRNA [33]) (iv)
virulence genes [34ndash37] (v) antibiotic resistance genes
[938] (vi) functional genes encoding enzymes responsible
for specific metabolic traits [139ndash42] etc (for a detailed
list please refer to wwwarcsacatuubmicrobiology)
However individual sequence databases for these alternative
markers if they exist at all currently contain considerably
fewer entries than the SSU rRNA databases constraining the
development and evaluation of encompassing probe sets for
microarrays [43]
The hybridization properties of long oligonucleotide
probes (typically 50ndash100mer) which show pronounced
hysteresis (higher temperatures for dissociation than for
association) are fundamentally different from those of short
probes (typically 15ndash30mer) [4445] While in principle
short oligonucleotides allow the discrimination of single
nucleotide differences under optimal conditions (see below
for further details) this does not hold true for long
oligonucleotides Their threshold for differentiation is
approximately 75ndash87 sequence similarity [414647]
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004
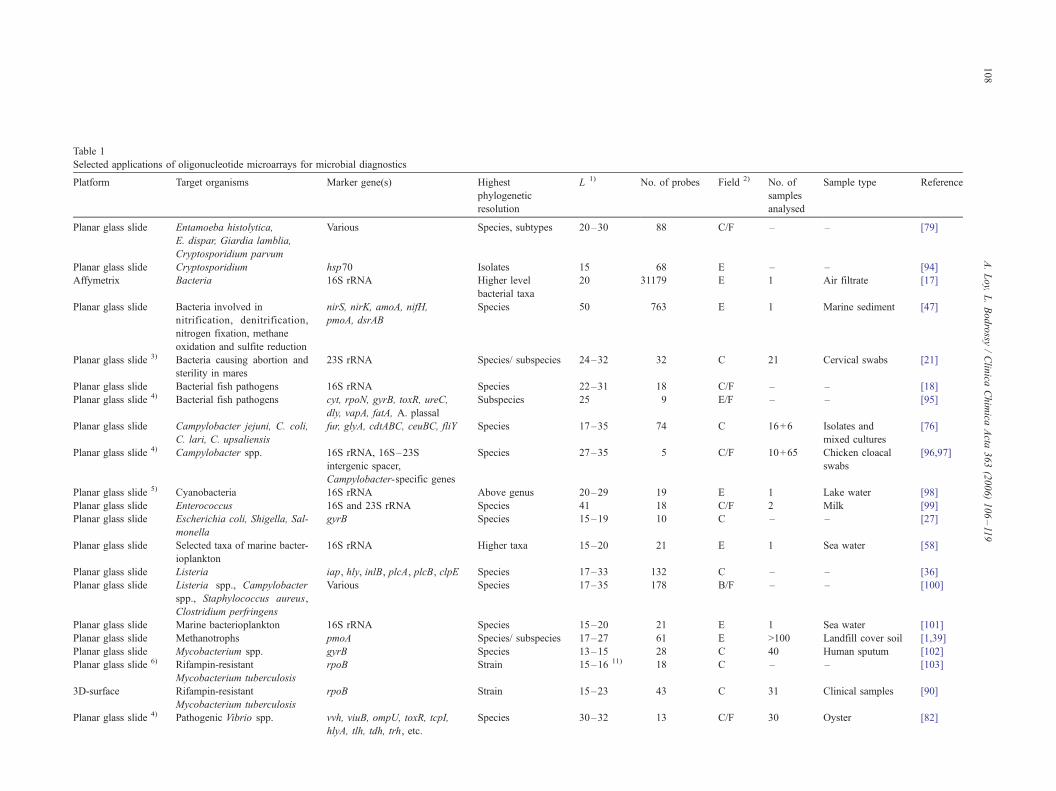
Table 1
Selected applications of oligonucleotide microarrays for microbial diagnostics
Platform Target organisms Marker gene(s) Highest
phylogenetic
resolution
L 1) No of probes Field 2) No of
samples
analysed
Sample type Reference
Planar glass slide Entamoeba histolytica
E dispar Giardia lamblia
Cryptosporidium parvum
Various Species subtypes 20ndash30 88 CF ndash ndash [79]
Planar glass slide Cryptosporidium hsp70 Isolates 15 68 E ndash ndash [94]
Affymetrix Bacteria 16S rRNA Higher level
bacterial taxa
20 31179 E 1 Air filtrate [17]
Planar glass slide Bacteria involved in
nitrification denitrification
nitrogen fixation methane
oxidation and sulfite reduction
nirS nirK amoA nifH
pmoA dsrAB
Species 50 763 E 1 Marine sediment [47]
Planar glass slide 3) Bacteria causing abortion and
sterility in mares
23S rRNA Species subspecies 24ndash32 32 C 21 Cervical swabs [21]
Planar glass slide Bacterial fish pathogens 16S rRNA Species 22ndash31 18 CF ndash ndash [18]
Planar glass slide 4) Bacterial fish pathogens cyt rpoN gyrB toxR ureC
dly vapA fatA A plassal
Subspecies 25 9 EF ndash ndash [95]
Planar glass slide Campylobacter jejuni C coli
C lari C upsaliensis
fur glyA cdtABC ceuBC fliY Species 17ndash35 74 C 16+6 Isolates and
mixed cultures
[76]
Planar glass slide 4) Campylobacter spp 16S rRNA 16Sndash23S
intergenic spacer
Campylobacter-specific genes
Species 27ndash35 5 CF 10+65 Chicken cloacal
swabs
[9697]
Planar glass slide 5) Cyanobacteria 16S rRNA Above genus 20ndash29 19 E 1 Lake water [98]
Planar glass slide Enterococcus 16S and 23S rRNA Species 41 18 CF 2 Milk [99]
Planar glass slide Escherichia coli Shigella Sal-
monella
gyrB Species 15ndash19 10 C ndash ndash [27]
Planar glass slide Selected taxa of marine bacter-
ioplankton
16S rRNA Higher taxa 15ndash20 21 E 1 Sea water [58]
Planar glass slide Listeria iap hly inlB plcA plcB clpE Species 17ndash33 132 C ndash ndash [36]
Planar glass slide Listeria spp Campylobacter
spp Staphylococcus aureus
Clostridium perfringens
Various Species 17ndash35 178 BF ndash ndash [100]
Planar glass slide Marine bacterioplankton 16S rRNA Species 15ndash20 21 E 1 Sea water [101]
Planar glass slide Methanotrophs pmoA Species subspecies 17ndash27 61 E gt100 Landfill cover soil [139]
Planar glass slide Mycobacterium spp gyrB Species 13ndash15 28 C 40 Human sputum [102]
Planar glass slide 6) Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash16 11) 18 C ndash ndash [103]
3D-surface Rifampin-resistant
Mycobacterium tuberculosis
rpoB Strain 15ndash23 43 C 31 Clinical samples [90]
Planar glass slide 4) Pathogenic Vibrio spp vvh viuB ompU toxR tcpI
hlyA tlh tdh trh etc
Species 30ndash32 13 CF 30 Oyster [82]
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
108
Planar glass slide 7) Quinolone-resistant
Escherichia coli
gyrA Genotype 19 42 C ndash ndash [104]
Planar glass slide Respiratory bacterial pathogens gyrB parE Species 20ndash24 27 C 94 Middle ear fluid
throat swabs
[105]
Planar glass slide Rhodocyclales 16S rRNA Species 18 79 E 1 Activated sludge [56]
3D-surface Some Bacillus spp 16S rRNA Species 15ndash23 30 BC ndash ndash [64]
Planar glass slide 7) Some Bacillus spp 16Sndash23S intergenic spacer Species 18ndash22 42 BC ndash ndash [22]
Planar glass slide Sulphate-reducing prokaryotes 16S rRNA Species 18 132 CE 6+8 Hypersaline
cyanobacterial
lake mat periodontal
tooth pocket fen soil
[15106]
3D-surface Thermophilic anaerobic
Archaea and Bacteria
16S rRNA Genus 17ndash20 17 E 3 Oil reservoir
formation waters
[77]
Affymetrix 8) Major potential biowarfare agents (bacteria viruses
eukaryotes)
Various Species 20 53660 BC 2 Air filtrate [107]
Planar glass slide Acute respiratory
disease-associated
Adenoviruses
E1A fibre hexon Serotypes 60ndash72 36 C 19 Throat swabs nasal
wash samples
[80]
Planar glass slide Human group A rotaviruses VP7 Genotype 18ndash26 50 C ndash ndash [108]
Planar glass slide Human papillomaviruses E1 Types 20 51 C 130 Clinical samples [109]
Planar glass slide 5) Human papillomaviruses Various Types 14ndash28 10 C 100 Clinical samples [110]
Planar glass slide Human papillomaviruses Various Types 30 27 C 73 Clinical samples
(tonsillar cancer)
[111]
Planar glass slide Influenza viruses Various Subtype 17ndash29 476 C ndash ndash [112]
3D-surface 9) Influenza viruses Various (ao hemagglutinin
and neuraminidase genes)
Subtypes 45ndash65 29 C ndash ndash [113]
Planar glass slide Orthopoxviruses C23LB29R gene ORF 62
(varicella-zoster)
Types 13ndash21 57 BC ndash ndash [114]
3D-surface Orthopoxviruses crmB Species 12ndash16 15 C ndash ndash [115]
Planar glass slide 10) Viruses Various Serotype 70 1600 C 6 Clinical samples [50]
1) Length of probe [nt]2) B biodefense C clinical microbiology F food microbiology E environmental microbiology3) On-Chip PCR4) Multiplex PCR6) DNA ligation detection reaction5) SOLAC-short oligo ligation assay on chip8) Perfect match and single mismatch probes used7) 3ndash10 diagnostic regions were identified per microorganism and each one of them was covered by 100ndash300 probes9) Flow-Thru Chip10) 70mer discriminatory oligonucleotides were designed based on all published viral genome sequences11) Probes comprised of 5 specific nucleotides (participating in specific ligation)+10ndash11 spacer T residues
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
109
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119110
should be noted however that there are also medical
microbiology contexts where (as described for environ-
mental MDMs) the microbial community structure has to be
addressed eg the composition of the human gastro-
intestinal tract microbiota
Although recent studies (Table 1) undoubtedly demon-
strate that MDMs are valuable tools for identification of
microorganisms and viruses in a highly parallel fashion
their use for routine diagnostics is still hampered by a lack
of standardisation (regarding factors such as probes target
genes hybridization platforms protocols and data analy-
sis) and insufficient evaluation of newly developed
MDMs
In this review we summarise recent progress in the
MDM field Many (seemingly conflicting) parameters and
steps have to be integrated during microarray design and
application in order to fulfil the potential of MDMs as
high-throughput screening tools for routine diagnostic
purposes We thus highlight crucial points related to
development and evaluation of MDMs The key to a
validated MDM and consequently to reliable results is
rigorous in silico and in vitro performance testing
Furthermore we exemplarily show recent technological
advances which should further improve the use of MDMs
in the near future
2 Microarray hybridization formats
Although many diverse microarray platforms have
become available over the past few years [6] only a limited
number of solid supports are currently used for MDMs For
Affymetrix microarrays (wwwaffymetrixcom) oligonu-
cleotide probes are synthesised directly onto the microarray
surface by employing specific masks and the photolithog-
raphy method This approach enables a very high probe
density (well over 100000 probes per microarray) The
high price low flexibility and lack of a suitably high
number of validated oligonucleotide probes currently limit
the wide application of Affymetrix GeneChips in microbial
diagnostics In contrast the NimbleGen technology
(wwwnimblegencom) uses digital micromirrors instead of
physical masks to guide on-chip probe synthesis allowing a
more flexible design of custom-made high-density micro-
arrays Specific three-dimensional microarray formats such
as the gel-pad platform (wwwbiochipruen) [78] or the
PamGene system (wwwpamgenecom) [910] are coupled
with appropriate hybridization and detection devices that
offer the option to record hybridization and dissociation
events in real-time Melting curves for all probes on a
microarray can thus be rapidly established making the
development of validated probe sets significantly easier
[1112] However these systems are currently available
only in a few laboratories
The pioneer microarray format and still the most widely
used miniaturised solid support for the covalent immobili-
sation of probes are planar 13 in glass slides Oligonu-
cleotides are in most cases tethered via their 5V ends to
reactive groups on the coating layer of the glass surface The
establishment of microarray core facilities (including micro-
array spotting and detection devices) in many laboratories
and the general utility flexibility and moderate price of
planar glass microarrays are mainly responsible for the
success of this standard format
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
Two main parameters affect the resolution of a diagnostic
microarray assay (i) the degree of conservation of the
marker gene and (ii) the length of the oligonucleotide used
as a probe to target it [13]
The most widely employed target molecule for the
detection and phylogenetic analysis of microorganisms is
the small-subunit ribosomal RNA (SSU rRNA) and its gene
[714ndash18] The popularity of the SSU rRNA is reflected in
the existence of large and regularly updated sequence [19
20] and probe databases (httpwwwmicrobial-ecologynet
probebase) [16] for this target molecule The main
limitation of using the SSU rRNA (gene) as a marker in
microarray assays is that resolution below the species level is
generally not possible owing to high overall sequence
conservation Because differentiation of strains is often
essential in clinical diagnostics in order to initiate appro-
priate treatment of an infection less-conserved target
molecules are needed Potential probe targets which offer
strain-level resolution include (i) the large-subunit ribosomal
RNA (LSU rRNA) [21] (ii) the SSU-LSU rRNA intergenic
spacer region [22] (iii) house-keeping genes (eg rpoB
[23ndash25] gyrA [26] gyrB [27] recA [28] tuf [2930] groEL
[31] atpD [30] ompA gapA pgi [32] tmRNA [33]) (iv)
virulence genes [34ndash37] (v) antibiotic resistance genes
[938] (vi) functional genes encoding enzymes responsible
for specific metabolic traits [139ndash42] etc (for a detailed
list please refer to wwwarcsacatuubmicrobiology)
However individual sequence databases for these alternative
markers if they exist at all currently contain considerably
fewer entries than the SSU rRNA databases constraining the
development and evaluation of encompassing probe sets for
microarrays [43]
The hybridization properties of long oligonucleotide
probes (typically 50ndash100mer) which show pronounced
hysteresis (higher temperatures for dissociation than for
association) are fundamentally different from those of short
probes (typically 15ndash30mer) [4445] While in principle
short oligonucleotides allow the discrimination of single
nucleotide differences under optimal conditions (see below
for further details) this does not hold true for long
oligonucleotides Their threshold for differentiation is
approximately 75ndash87 sequence similarity [414647]
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

Planar glass slide 7) Quinolone-resistant
Escherichia coli
gyrA Genotype 19 42 C ndash ndash [104]
Planar glass slide Respiratory bacterial pathogens gyrB parE Species 20ndash24 27 C 94 Middle ear fluid
throat swabs
[105]
Planar glass slide Rhodocyclales 16S rRNA Species 18 79 E 1 Activated sludge [56]
3D-surface Some Bacillus spp 16S rRNA Species 15ndash23 30 BC ndash ndash [64]
Planar glass slide 7) Some Bacillus spp 16Sndash23S intergenic spacer Species 18ndash22 42 BC ndash ndash [22]
Planar glass slide Sulphate-reducing prokaryotes 16S rRNA Species 18 132 CE 6+8 Hypersaline
cyanobacterial
lake mat periodontal
tooth pocket fen soil
[15106]
3D-surface Thermophilic anaerobic
Archaea and Bacteria
16S rRNA Genus 17ndash20 17 E 3 Oil reservoir
formation waters
[77]
Affymetrix 8) Major potential biowarfare agents (bacteria viruses
eukaryotes)
Various Species 20 53660 BC 2 Air filtrate [107]
Planar glass slide Acute respiratory
disease-associated
Adenoviruses
E1A fibre hexon Serotypes 60ndash72 36 C 19 Throat swabs nasal
wash samples
[80]
Planar glass slide Human group A rotaviruses VP7 Genotype 18ndash26 50 C ndash ndash [108]
Planar glass slide Human papillomaviruses E1 Types 20 51 C 130 Clinical samples [109]
Planar glass slide 5) Human papillomaviruses Various Types 14ndash28 10 C 100 Clinical samples [110]
Planar glass slide Human papillomaviruses Various Types 30 27 C 73 Clinical samples
(tonsillar cancer)
[111]
Planar glass slide Influenza viruses Various Subtype 17ndash29 476 C ndash ndash [112]
3D-surface 9) Influenza viruses Various (ao hemagglutinin
and neuraminidase genes)
Subtypes 45ndash65 29 C ndash ndash [113]
Planar glass slide Orthopoxviruses C23LB29R gene ORF 62
(varicella-zoster)
Types 13ndash21 57 BC ndash ndash [114]
3D-surface Orthopoxviruses crmB Species 12ndash16 15 C ndash ndash [115]
Planar glass slide 10) Viruses Various Serotype 70 1600 C 6 Clinical samples [50]
1) Length of probe [nt]2) B biodefense C clinical microbiology F food microbiology E environmental microbiology3) On-Chip PCR4) Multiplex PCR6) DNA ligation detection reaction5) SOLAC-short oligo ligation assay on chip8) Perfect match and single mismatch probes used7) 3ndash10 diagnostic regions were identified per microorganism and each one of them was covered by 100ndash300 probes9) Flow-Thru Chip10) 70mer discriminatory oligonucleotides were designed based on all published viral genome sequences11) Probes comprised of 5 specific nucleotides (participating in specific ligation)+10ndash11 spacer T residues
ALoyLBodrossy
Clin
icaChimica
Acta
363(2006)106ndash119
109
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119110
should be noted however that there are also medical
microbiology contexts where (as described for environ-
mental MDMs) the microbial community structure has to be
addressed eg the composition of the human gastro-
intestinal tract microbiota
Although recent studies (Table 1) undoubtedly demon-
strate that MDMs are valuable tools for identification of
microorganisms and viruses in a highly parallel fashion
their use for routine diagnostics is still hampered by a lack
of standardisation (regarding factors such as probes target
genes hybridization platforms protocols and data analy-
sis) and insufficient evaluation of newly developed
MDMs
In this review we summarise recent progress in the
MDM field Many (seemingly conflicting) parameters and
steps have to be integrated during microarray design and
application in order to fulfil the potential of MDMs as
high-throughput screening tools for routine diagnostic
purposes We thus highlight crucial points related to
development and evaluation of MDMs The key to a
validated MDM and consequently to reliable results is
rigorous in silico and in vitro performance testing
Furthermore we exemplarily show recent technological
advances which should further improve the use of MDMs
in the near future
2 Microarray hybridization formats
Although many diverse microarray platforms have
become available over the past few years [6] only a limited
number of solid supports are currently used for MDMs For
Affymetrix microarrays (wwwaffymetrixcom) oligonu-
cleotide probes are synthesised directly onto the microarray
surface by employing specific masks and the photolithog-
raphy method This approach enables a very high probe
density (well over 100000 probes per microarray) The
high price low flexibility and lack of a suitably high
number of validated oligonucleotide probes currently limit
the wide application of Affymetrix GeneChips in microbial
diagnostics In contrast the NimbleGen technology
(wwwnimblegencom) uses digital micromirrors instead of
physical masks to guide on-chip probe synthesis allowing a
more flexible design of custom-made high-density micro-
arrays Specific three-dimensional microarray formats such
as the gel-pad platform (wwwbiochipruen) [78] or the
PamGene system (wwwpamgenecom) [910] are coupled
with appropriate hybridization and detection devices that
offer the option to record hybridization and dissociation
events in real-time Melting curves for all probes on a
microarray can thus be rapidly established making the
development of validated probe sets significantly easier
[1112] However these systems are currently available
only in a few laboratories
The pioneer microarray format and still the most widely
used miniaturised solid support for the covalent immobili-
sation of probes are planar 13 in glass slides Oligonu-
cleotides are in most cases tethered via their 5V ends to
reactive groups on the coating layer of the glass surface The
establishment of microarray core facilities (including micro-
array spotting and detection devices) in many laboratories
and the general utility flexibility and moderate price of
planar glass microarrays are mainly responsible for the
success of this standard format
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
Two main parameters affect the resolution of a diagnostic
microarray assay (i) the degree of conservation of the
marker gene and (ii) the length of the oligonucleotide used
as a probe to target it [13]
The most widely employed target molecule for the
detection and phylogenetic analysis of microorganisms is
the small-subunit ribosomal RNA (SSU rRNA) and its gene
[714ndash18] The popularity of the SSU rRNA is reflected in
the existence of large and regularly updated sequence [19
20] and probe databases (httpwwwmicrobial-ecologynet
probebase) [16] for this target molecule The main
limitation of using the SSU rRNA (gene) as a marker in
microarray assays is that resolution below the species level is
generally not possible owing to high overall sequence
conservation Because differentiation of strains is often
essential in clinical diagnostics in order to initiate appro-
priate treatment of an infection less-conserved target
molecules are needed Potential probe targets which offer
strain-level resolution include (i) the large-subunit ribosomal
RNA (LSU rRNA) [21] (ii) the SSU-LSU rRNA intergenic
spacer region [22] (iii) house-keeping genes (eg rpoB
[23ndash25] gyrA [26] gyrB [27] recA [28] tuf [2930] groEL
[31] atpD [30] ompA gapA pgi [32] tmRNA [33]) (iv)
virulence genes [34ndash37] (v) antibiotic resistance genes
[938] (vi) functional genes encoding enzymes responsible
for specific metabolic traits [139ndash42] etc (for a detailed
list please refer to wwwarcsacatuubmicrobiology)
However individual sequence databases for these alternative
markers if they exist at all currently contain considerably
fewer entries than the SSU rRNA databases constraining the
development and evaluation of encompassing probe sets for
microarrays [43]
The hybridization properties of long oligonucleotide
probes (typically 50ndash100mer) which show pronounced
hysteresis (higher temperatures for dissociation than for
association) are fundamentally different from those of short
probes (typically 15ndash30mer) [4445] While in principle
short oligonucleotides allow the discrimination of single
nucleotide differences under optimal conditions (see below
for further details) this does not hold true for long
oligonucleotides Their threshold for differentiation is
approximately 75ndash87 sequence similarity [414647]
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119110
should be noted however that there are also medical
microbiology contexts where (as described for environ-
mental MDMs) the microbial community structure has to be
addressed eg the composition of the human gastro-
intestinal tract microbiota
Although recent studies (Table 1) undoubtedly demon-
strate that MDMs are valuable tools for identification of
microorganisms and viruses in a highly parallel fashion
their use for routine diagnostics is still hampered by a lack
of standardisation (regarding factors such as probes target
genes hybridization platforms protocols and data analy-
sis) and insufficient evaluation of newly developed
MDMs
In this review we summarise recent progress in the
MDM field Many (seemingly conflicting) parameters and
steps have to be integrated during microarray design and
application in order to fulfil the potential of MDMs as
high-throughput screening tools for routine diagnostic
purposes We thus highlight crucial points related to
development and evaluation of MDMs The key to a
validated MDM and consequently to reliable results is
rigorous in silico and in vitro performance testing
Furthermore we exemplarily show recent technological
advances which should further improve the use of MDMs
in the near future
2 Microarray hybridization formats
Although many diverse microarray platforms have
become available over the past few years [6] only a limited
number of solid supports are currently used for MDMs For
Affymetrix microarrays (wwwaffymetrixcom) oligonu-
cleotide probes are synthesised directly onto the microarray
surface by employing specific masks and the photolithog-
raphy method This approach enables a very high probe
density (well over 100000 probes per microarray) The
high price low flexibility and lack of a suitably high
number of validated oligonucleotide probes currently limit
the wide application of Affymetrix GeneChips in microbial
diagnostics In contrast the NimbleGen technology
(wwwnimblegencom) uses digital micromirrors instead of
physical masks to guide on-chip probe synthesis allowing a
more flexible design of custom-made high-density micro-
arrays Specific three-dimensional microarray formats such
as the gel-pad platform (wwwbiochipruen) [78] or the
PamGene system (wwwpamgenecom) [910] are coupled
with appropriate hybridization and detection devices that
offer the option to record hybridization and dissociation
events in real-time Melting curves for all probes on a
microarray can thus be rapidly established making the
development of validated probe sets significantly easier
[1112] However these systems are currently available
only in a few laboratories
The pioneer microarray format and still the most widely
used miniaturised solid support for the covalent immobili-
sation of probes are planar 13 in glass slides Oligonu-
cleotides are in most cases tethered via their 5V ends to
reactive groups on the coating layer of the glass surface The
establishment of microarray core facilities (including micro-
array spotting and detection devices) in many laboratories
and the general utility flexibility and moderate price of
planar glass microarrays are mainly responsible for the
success of this standard format
3 Development and analytical performance
31 Resolution choice of marker genes and probe lengths
Two main parameters affect the resolution of a diagnostic
microarray assay (i) the degree of conservation of the
marker gene and (ii) the length of the oligonucleotide used
as a probe to target it [13]
The most widely employed target molecule for the
detection and phylogenetic analysis of microorganisms is
the small-subunit ribosomal RNA (SSU rRNA) and its gene
[714ndash18] The popularity of the SSU rRNA is reflected in
the existence of large and regularly updated sequence [19
20] and probe databases (httpwwwmicrobial-ecologynet
probebase) [16] for this target molecule The main
limitation of using the SSU rRNA (gene) as a marker in
microarray assays is that resolution below the species level is
generally not possible owing to high overall sequence
conservation Because differentiation of strains is often
essential in clinical diagnostics in order to initiate appro-
priate treatment of an infection less-conserved target
molecules are needed Potential probe targets which offer
strain-level resolution include (i) the large-subunit ribosomal
RNA (LSU rRNA) [21] (ii) the SSU-LSU rRNA intergenic
spacer region [22] (iii) house-keeping genes (eg rpoB
[23ndash25] gyrA [26] gyrB [27] recA [28] tuf [2930] groEL
[31] atpD [30] ompA gapA pgi [32] tmRNA [33]) (iv)
virulence genes [34ndash37] (v) antibiotic resistance genes
[938] (vi) functional genes encoding enzymes responsible
for specific metabolic traits [139ndash42] etc (for a detailed
list please refer to wwwarcsacatuubmicrobiology)
However individual sequence databases for these alternative
markers if they exist at all currently contain considerably
fewer entries than the SSU rRNA databases constraining the
development and evaluation of encompassing probe sets for
microarrays [43]
The hybridization properties of long oligonucleotide
probes (typically 50ndash100mer) which show pronounced
hysteresis (higher temperatures for dissociation than for
association) are fundamentally different from those of short
probes (typically 15ndash30mer) [4445] While in principle
short oligonucleotides allow the discrimination of single
nucleotide differences under optimal conditions (see below
for further details) this does not hold true for long
oligonucleotides Their threshold for differentiation is
approximately 75ndash87 sequence similarity [414647]
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 111
However probes of increased length display orders of
magnitude higher target binding capacities and hence the
use of long oligonucleotides should generally improve the
detection sensitivity of a microarray [48] Typically long
oligonucleotide MDMs are used in combination with
universal (not PCR based low bias) amplification strategies
or without any amplification [474950] In these approaches
probes target various non-conserved genes which are
specific to the microorganisms targeted Thus the low
differentiation power of long oligonucleotide probes is
compensated for by the host specificity of the genes they are
designed against With or without universal amplification
the resulting target represents the entire gene pool present in
the investigated sample without any reduction in its
complexity Higher binding capacities of the long oligop-
robes and higher target complexity result in similar relative
detection thresholds to that of short oligoprobes in
conjunction with PCR amplification (approximately 5)
[14751]
There is no universal answer to the question of which
probe-target combination is the best for a diagnostic
microarray as this will depend strongly on the intended
application In principle the highly parallel nature of
microarrays allows various probes of different length and
targeting different genes to be applied simultaneously
although homogenous hybridization behaviour of complex
probetarget combinations remains a problem (see below)
32 Development and optimisation of microarray probe
sets specificity sensitivity and uniformity
A crucial and challenging first step in the development of
oligonucleotide microarrays is the design of a suitable set of
probes and this is thus presented in more detail in the
following section
The following criteria set the quality standard for a
microarray probe set All probes on a microarray should
(i) be highly specific for their target genes ie not cross-
hybridize with non-target sequences (specificity) (ii) bind
efficiently to target sequences to allow the detection of
low abundance targets in complex mixtures (sensitivity)
and (iii) display a similar hybridization behaviour ie
similar thermodynamic characteristics under the same
experimental conditions (homogeneity uniformity)
Unfortunately these ideals represent conflicting goals in
practice and thus efforts to fulfil these criteria need to be
carefully balanced during probe design and experimental
procedures
Design of microarray probes in silico usually entails the
use of specific software tools in conjunction with an
underlying sequence database Numerous software tools
have been developed and widely applied for the parallel
design of oligonucleotide probes for whole genome
microarrays [5253] but not so for diagnostic microarrays
Although only one probe can be designed at a time
making the design of multiple probes laborious the probe
software tool of the ARB program package [19] is
commonly used for the design of 10ndash100mer oligonucleo-
tide probes for diverse hybridization formats [155455]
including diagnostic microarrays [56ndash58] The first step in
the design of a diagnostic oligonucleotide is selection of
the target group either arbitrarily or based on sequence
analysis (eg a phylogenetically coherent group of
organisms) (Fig 2) Second depending on user-defined
settings the ARB probe search algorithm identifies unique
sequence stretches which could serve as probe target sites
and subsequently returns a ranked list of potential
oligonucleotides Third the suggested probes can be
matched against all (usually aligned) sequences in the
database This probe match option of ARB is highly
beneficial because the user has the opportunity to evaluate
the specificity of a given probe in silico by checking for
the number types and positions of mismatches to non-
target sequences It is vital to stress that the value of this
evaluation step and thus the quality of a developed probe
set will strongly correlate with the completeness and up-to-
dateness of the underlying sequence database Regarding
the position of mismatches to non-target organisms a rule
of thumb for short oligonucleotides is that mismatches
located in the middle of the probe-binding site destabilize
more strongly the formation of the probe-target duplex In
contrast an even distribution of mismatches in the probe-
binding site is required to achieve optimal discrimination
for long oligonucleotides [4648] Please note that single
mismatches located at the terminal or next-to-terminal
position can hardly be resolved in microarray hybridization
[114857] In a further step the local alignments of
perfectly-matched and mismatched target sites can be used
as input for other freely-available software such as
CalcOligo (wwwarcsacatuubmicrobiology) Mfold
[59] or HyTher [60] to calculate the theoretical thermo-
dynamic properties of these potential hybridization events
However these nearest-neighbour algorithms for the
calculation of free energies (DG) and melting temperatures
(Tm) have been developed for hybridizations in solution
involving known concentrations of probe and target
molecules factors which are not readily known or do
not apply for hybridization of immobilized probes on
microarrays Data obtained in the laboratory must therefore
accurately verify the use of these algorithms to predict
potential cross-hybridization events of a microarray probe
[1153961]
For single probes it is in most cases possible to adjust
experimental conditions in such a way that no cross-
hybridization occurs However the most widely used
microarray hybridization formats only allow hybridization
andor washing at a single stringency making it impossible
to provide optimal hybridization conditions for all probes
on a microarray Thus promiscuous binding of some
probes to non-target sequences (false-positive signals) is a
frequently encountered problem [115] Nonetheless sev-
eral approaches still guarantee the reliability of identifica-
Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

Creationupdate ofsequence database
Probe design
Specificity sensitivity homogeneity
Secondary structures (∆G Tm)
Predicted hybridization behavior
Thorough experimental evaluation(Specificity sensitivity homogeneity)
Pre-selected probe set
Refined probe set
Elimination of ldquobadrdquo probes
Suitable software
Identification of target groupeg by phylogenetic analysis
Fully evaluatedfinal microarray
Re-design
Preliminary microarray
In silico
In vitro
Type number position of mismatch(es)
Re-evaluation
12
34
ARB
ARB
ARBPrimRose
ARBPrimRoseOligoCheckCalcOligoMfoldHyTher
CalcOligoMfoldHyTher
Fig 2 Schematic representation of the procedures involved in the development and evaluation of a microarray probe set Types of molecular interactions of the
probe and target molecules 1 oligo-oligo dimer 2 oligo hairpin 3 secondary structure of the target 4 oligo-target duplex Free energies and melting
temperatures of the different types of interactions of probe and target molecules can be calculated to estimate their influence on duplex yield
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119112
tion by microarrays One approach involves the design of
multiple probes having identical specificities for the same
target sequence or group of sequences Therefore all
probes in a set of probes perfectly matching the target
organism must show positive hybridization signals to
minimize the risk of false-positive identification
[346263] This multiple probe concept is further extend-
able by probes having hierarchical (nested) specificities for
the target sequences (Fig 3) which also enables the
detection of novel members of known groups [54] Nested
probes can be best exploited if rRNA or its gene serves as
the target molecule [15545664] In contrast the highly
variable third codon (lsquolsquowobblersquorsquo) position in protein-coding
target genes hampers the design of probes having a broader
specificity and spanning more distantly related sequences
In addition to or instead of the use of multiple probes for
one marker gene it is possible to use more than one marker
gene of a target organism as a probe target during
hybridization This multiple probe-multiple target strategy
additionally increases redundancy and hence the confi-
12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

12345678910111213141516171819202122232425262728
A1 A2 A3
B1 B2C1
D1
E1
A4
Phylogeny Sequence alignment
Fig 3 Illustration of the nested multiple probe concept A schematic sequence alignment is displayed in dashed lines Colored boxes show perfectly matched
probe target sites As an example sequence number one is unambiguously rated as present if each probe in the set of probes E1YD1YC1YB1 B2YA1
A2 A3 perfectly matching this sequence at identical and hierarchical levels of specificity shows a positive hybridization signal
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 113
dence in a positive call [6263] Furthermore careful
selection of multiple marker genes may also allow
genotyping of the detected microbes Information on
virulence factors antibiotic resistance etc can thereby
be obtained simultaneously in a single diagnostic assay
Another prominent way (also applied in the Affymetrix
GeneChips protocols [17]) to ensure specificity is the
inclusion of so-called mismatch-control probes on the
microarray [6566] Comparison of signal intensities from
perfectly matched and mismatched probes allows cross-
hybridization to be identified and its extent estimated
In summary one set of experimental conditions is not
suitable to ensure absolute specificity of all probes during
simultaneous hybridization on a microarray However
microarrays offer possibilities to compensate for lack of
specificity on the single probe level by inclusion of a
multiplicity of redundant probes an option which is not
available for traditional hybridization formats
Sensitivity is (i) commonly defined as the lowest
absolute andor relative abundance of the target group
which is detectable and (ii) dependent on the duplex yields
of the individual probes on a microarray To avoid the
worst-case scenario low sensitivity of a given probe
leading to a false-negative result a variety of parameters
can be adjusted during the design of the actual microarray
and the experimental set-up An important strategy for
increasing duplex yield is to reduce steric hindrance during
microarray hybridization Lifting the oligonucleotide probe
physically away from the surface of the microarray by the
introduction of simple spacer elements enhances its
accessibility by several orders of magnitude [6768]
Furthermore the tendency of a probe and its target to form
stable secondary self-structures must be minimized
[16970] Tm and DG of secondary structures can be
calculated by using the software tools mentioned above but
should be treated cautiously due to the limited knowledge
on the thermodynamics of microarray hybridization (see
above) If RNA is the target molecule complexity of the
target is typically reduced by physical or chemical
fragmentation [14571] A popular method to create short
DNA fragments of varying length is by random prime
labelling [49] Partial digestion with DNase I [49] or use of
hydroxyl radical-producing reagents are other options to
fragment DNA the latter non-enzymatic treatment being
more reproducible [72] So-called helper or chaperone
probes target a sequence region adjacent to a probe target
site and have been demonstrated to increase target
accessibility by relieving secondary structures [5873]
However the helper probe approach is less practical for
high-density microarrays as each probe on the microarray
requires its own specific helper counterpart
Unique to formats involving the simultaneous hybrid-
ization of many probes is that the probes have to be fine-
tuned for uniform thermodynamic behaviour One way to
achieve this is by using probes that are identical in length
and adding tertiary amine salts such as tetramethylammo-
nium chloride to the hybridization andor washing buffers
[1556] Thereby differences in the G+C contents and
thus duplex stabilities among the probes are attenuated
[7475] Another strategy to equalize melting properties of
different oligonucleotide probes is to manipulate their
length [139]
One must realize that current in silico approaches for
predicting the hybridization behaviour of microarray
probes are limited due to the aforementioned reasons At
best they will lead to a pre-filtered set of candidate probes
whose true experimental performance will only be uncov-
ered after extensive empirical testing (Fig 2) In practice a
suitable set of test targets should contain at least one
perfectly matched target for each probe on the microarray
After challenging the microarrayrsquos performance by indi-
vidual hybridizations with each test target lsquolsquobadrsquorsquo probes
showing low sensitivity andor specificity are removed or
replaced Subsequently concentration series of targets
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119114
perfectly matching those probes that have displayed the
highest and lowest duplex yield should be hybridized to
the microarray giving an impression of the range of
sensitivities achievable for the individual probes In
conclusion the key to a reliable diagnostic microarray
lies within its concerted evaluation integrating in silico
predictions adjustment of hybridization conditions and
thorough testing in the laboratory
33 Further selected strategies to increase specificity and
or sensitivity
The most common way to enhance detectability of a target
gene is the use of PCR [1151776ndash78] PCR is applied to
focus the labelling to the target genes and thus to enrich them
relative to non-targeted background DNA In case of a single
marker gene highly selective PCR primers may achieve
optimal enrichment of target sequences from organisms of
interest [56] However a high number of selective PCR
primers is needed to cover all organisms targeted by a
microarray Running multiple PCR reactions considerably
increases the hands-on-time of the microarray assay and thus
simultaneous target amplification via multiplex PCR is
desirable [3579ndash82] Multiplex PCR has the inherent
drawback that potential biases increase with increased
complexity of the reactants (primerstarget genes) [the
number of successfully applied primer pairs within a multi-
plex PCR is normally low up to 12 [83]] Consequently
comparable to the actual microarray hybridization the
implementation of a pre-hybridization amplification step
based on multiplex PCR requires extensive careful testing
and validation
A promising approach to increase the sensitivity of a
microarray assay is tyramide signal amplification (TSA)
[51] Upon hybridization this method relies on enzymatic
amplification of the signal by employing horse radish
peroxidase-mediated deposition of fluorochrome-labelled
tyramides at the location of the probe
The ultimate specificity of microarray technology
depends on the discrimination between a fully complemen-
tary target and a non-target differing in only a single
nucleotide Recently various enzyme-assisted hybridization
strategies (also used in single nucleotide polymorphism and
resequencing assays [8485]) have gained attention because
of their promise in strongly discriminating single mis-
matches located near the 3V end of microarray probes
[2186ndash90]
4 Data analysis and quantification potential
For microarrays used in transcriptome analysis signal
intensity ratios indicating relative levels of expression are
obtained upon competitive hybridization of a control
sample versus the sample of interest (two color experi-
ment) [91] In contrast diagnostic microarray experiments
usually involve the hybridization of a single sample per
microarray (single color experiment) Fluorescence signals
for the individual probes are typically rated as present or
absent (Fig 1) a decision made depending on either
simple visual inspection of the scanned microarray image
or an arbitrary signal intensity threshold [11521
225880] Subsequently complex hybridization patterns
are in most cases still translated manually into inventory
lists of organisms present in the analyzed samples This is
a tedious procedure which becomes more problematic with
increasing numbers of probes per microarray and increas-
ing numbers of samples to be analyzed and thus
constitutes a major bottleneck of microarray-based diag-
nostics In an attempt to bridge the gap between data
collection and analysis a simple command line-based
program was recently developed for diagnostic microarrays
[15] [an advanced graphical user interface-based version
ChipAnalyser is under development (Harald Meier
unpublished data)]
Although systematic and stochastic errors associated
with the different steps during microarray fabrication and
target preparation greatly hamper quantification the funda-
mental potential of microarrays to provide quantitative data
on the abundance of the detected target in a sample is widely
acknowledged Adopting the two color hybridization
approach from transcriptome microarray analysis relative
abundance of targets in a sample (labelled in color one) can
be measured by competitive hybridization with target
mixtures of known concentrations (labelled in color two)
on the microarray [192] For single color hybridizations a
linear correlation between the signal intensity of a probe and
the concentration of the respective target sequence has been
observed for a certain range of target concentrations
[414793] However the slope of this linear relationship
will vary among the different probes immobilized on a
microarray because as mentioned previously different
probes display different affinities to their targets For a
reliable quantitative interpretation of a microarray hybrid-
ization a standard curve is thus needed for each probe on
the microarray This tedious evaluation procedure has not
been accomplished for microarrays published to date and
therefore the ability to correctly quantify across all probes
on a microarray is yet to be demonstrated A semi-
quantitative comparison of similar community structures
is however possible enabling the researcher to detect
spatial and temporal changes in microbial community
composition [39]
5 Diagnostic applications
MDMs are increasingly used in clinical environmental
and food microbiology as well as in biowarfare agent
detection [43] (Table 1) While some clinical applications
are limited to genotyping of isolated strains there are more
and more publications on cultivation-independent rapid
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 115
detection and identification (in some cases also genotyping)
of pathogens from clinical specimens Microarrays used
merely for genotyping of isolates are outside the scope of
our review and are thus not listed in Table 1
6 Conclusions and outlook
Technologies used for MDMs are rather diverse PCR
amplification followed by conventional fluorescence label-
ling is used in most cases Specificity is then defined by the
hybridization step Depending on the marker genes applied
current MDMs can provide resolution at various taxonomic
levels down to species or even strain level Ribosomal RNA
genes are often used as marker genes but less so in clinical
microbiology due to their limited phylogenetic resolution
Alternative higher resolution phylogenetic markers are thus
beginning to be applied with short oligonucleotide probes
used in a hierarchical and parallel manner being the
preferred choice
Reliable analysis of 10ndash30 samples can be achieved
within 24 h (in most cases within a working day) by one
researcher The costs of consumables are approximately
30ndash100 EUR per analysis depending on the exact
methodology used Although this may seem expensive it
must be remembered that a single analysis provides parallel
detection and identification of tens to hundreds of micro-
organisms potentially present In addition basic genotyping
information can be obtained as well as semi-quantitative
information on the relative abundance of the detected
microorganisms The latter aspect is especially useful when
comparing similar samples for changes in the microbial
community structure (eg gut microbiota) Absolute
quantification is troubled by biases associated with for
example PCR amplification nucleic acid recovery and
different numbers of (marker) gene copies per genome The
sensitivity of MDMs is normally limited by the relative
abundance of the microbial population within the targeted
community with reported detection limits being 1ndash5
The hybridization potential of the probes is predicted based
on the nearest neighbour model and a number of empirical
microarray-specific factors
MDM technology lends itself to automation DNARNA
purification amplification labelling can all be carried out
by existing laboratory robots Moreover there are proof-of-
concept experiments where all these tasks are carried out in
a single lab-on-a-chip device [116] The subsequent hybrid-
ization wash and scan steps can also already be carried out
by various instruments in an automated manner (for details
please refer to httpgenomicshomecom) Automated
result interpretation is primarily a bioinformatics challenge
In general MDM technology is analogous to the applica-
tion of microarrays in gene expression profiling a high-
throughput screening tool with limited quantification
potential MDM results should thus be confirmed via
established lower-throughput but more sensitive andor
truly quantitative alternative methods such as fluorescence
in situ hybridization quantitative PCR or immunological
detection techniques
Even though MDMs are already widely applied in most
fields of microbiology anticipated improvements (listed and
discussed below) should overcome some of the aforemen-
tioned limitations
The prediction of the hybridization behaviour of the
probes is still rudimentary and requires improvement This
can be achieved via input from bioinformatics and the
establishment of datasets on array-specific effects such as
immobilisation and steric hindrance
Large databases comparable in size to that of the 16S
rRNA databases are critically needed for high resolution
phylogenetic markers Such markers will drastically
improve the applicability of MDMs for clinical and food
microbiology epidemiology and related fields If in such
databases sequence information is linked to clinical traits
(eg pathogenicity host specificity antibiotic resistance
geographic origin etc) MDM-based detection will serve
the additional purpose of providing a prediction of these
functions at least on some of them at a given level of
certainty
Some enzyme-based labelling methods [86ndash88117]
have demonstrated the potential for an improved detection
sensitivity (down to about 01 of the total community
targeted) although at the cost of losing all quantification
potential
Current hybridization platforms predominantly use
fluorescence-based optical detection Both the fluorescent
label and the detection devices confer high costs to this
technology seriously limiting its spread across diagnostic
laboratories With a few exceptions hybridization results
are read after irreversible processing steps thus the result
reflects hybridization under a selected suboptimal hybrid-
ization condition Alternative detection platforms are thus
being developed and elaborated which if successful could
have two major advantages One is reduced costs the other
is on-line detection of hybridization events Electronic and
mechanical detection methods [116118ndash121] require
cheaper or no labelling and simpler cheaper readout
devices Label-free detection may also help in cutting the
time-to-result a critical issue in many medical applica-
tions On-line detection of the hybridization event may be
possible via a number of techniques [8ndash10116118ndash125]
Such an improvement will drastically increase the infor-
mation obtainable from any hybridization event Associa-
tion and dissociation kinetics and maximum hybridization
signal at the optimal hybridization stringency for each
individual probe can all be obtained By considering these
extra information the reliability of MDM-based diagnos-
tics can be further improved On-line hybridization
detection is also another means of cutting the time-to-
result A label-free on-line hybridization detection
approach could make it possible to obtain results within
less than 2 h
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119116
Multiple marker genes providing reliable information on
the phylogeny and identity of the detected microbe and its
clinical traits should be used on a single MDM in many
cases to provide all the relevant information to the diagnostic
laboratory As discussed earlier multiplex amplification is
still a serious limitation to this approach A true on-chip PCR
where also the amplification primers are immobilised would
enable unlimited multiplexing solving this problem and
opening new horizons in the field
Acknowledgements
Related work at Seibersdorf research (LB) was funded by
the Fonds zur Forderung der wissenschaftlichen Forschung
Austria (P15044) and through the EU 5th Framework
Quality of Life and Management of Living Resources Grant
QLK-3 CT-2000-01528 Research of AL was funded by a
Marie Curie Intra-European Fellowship within the 6th
European Community Framework Programme Funding
from the EU towards COST Action 853 contributed by
enhancing exchange of ideas The authors thank Michael
Taylor for critical revision of the manuscript AL is greatly
indebted to Michael Wagner for ongoing support LB is
indebted to Fodor Szilvia for her support and fresh
independent views and ideas Helpful comments and
discussions from many active members of the Yahoo
discussion groups on microarrays and microbial diagnos-
ticsmicrobial ecology (httpgroupsyahoocomgroup
microarray and httpgroupsyahoocomgroupMDME)
are acknowledged
References
[1] Bodrossy L Stralis-Pavese N Murrell JC Radajewski S Weilhar-
ter A Sessitsch A Development and validation of a diagnostic
microbial microarray for methanotrophs Environ Microbiol 20035
566ndash82
[2] Zhou J Microarrays for bacterial detection and microbial community
analysis Curr Opin Microbiol 20036288ndash94
[3] Letowski J Brousseau R Masson L DNA microarray applications in
environmental microbiology Anal Lett 2003363165ndash84
[4] Stenger D Andreadis J Voraa G Pancrazio J Potential applications
of DNA microarrays in biodefense-related diagnostics Curr Opin
Biotechnol 200213208ndash12
[5] Clewley JP A role for arrays in clinical virology fact or fiction J
Clin Virol 2004292ndash12
[6] Gershon D Microarrays go mainstream Nature Methods 20041
263ndash70
[7] Guschin DY Mobarry BK Proudnikov D Stahl DA Rittmann BE
Mirzabekov AD Oligonucleotide microchips as genosensors for
determinative and environmental studies in microbiology Appl
Environ Microbiol 1997632397ndash402
[8] Fotin AV Drobyshev AL Proudnikov DY Perov AN Mirzabekov
AD Parallel thermodynamic analysis of duplexes on oligodeoxyr-
ibonucleotide microchips Nucleic Acids Res 1998261515ndash21
[9] Anthony RM Schuitema ARJ Oskam L Klatser PR Direct
detection of Staphylococcus aureus mRNA using a flow through
microarray J Microbiol Methods 20056047ndash54
[10] Wu Y de Kievit P Vahlkamp L et al Quantitative assessment of a
novel flow-through porous microarray for the rapid analysis of gene
expression profiles Nucleic Acids Res 200432e123
[11] Urakawa H Noble PA El Fantroussi S Kelly JJ Stahl DA Single-
base-pair discrimination of terminal mismatches by using oligonu-
cleotide microarrays and neural network analyses Appl Environ
Microbiol 200268235ndash44
[12] Urakawa H El Fantroussi S Smidt H et al Optimization of single-
base-pair mismatch discrimination in oligonucleotide microarrays
Appl Environ Microbiol 2003692848ndash56
[13] Bodrossy L Sessitsch A Oligonucleotide microarrays in microbial
diagnostics Curr Opin Microbiol 20047245ndash54
[14] Ludwig W Schleifer KH Phylogeny of bacteria beyond the 16S
rRNA standard ASM News 1999651515ndash21
[15] Loy A Lehner A Lee N et al Oligonucleotide microarray for 16S
rRNA gene-based detection of all recognized lineages of sulfate-
reducing prokaryotes in the environment Appl Environ Microbiol
2002685064ndash81
[16] Loy A Horn M Wagner M probeBase an online resource for
rRNA-targeted oligonucleotide probes Nucleic Acids Res 200331
514ndash6
[17] Wilson KH Wilson WJ Radosevich JL et al High-density micro-
array of small-subunit ribosomal DNA probes Appl Environ Micro-
biol 2002682535ndash41
[18] Warsen AE Krug MJ LaFrentz S Stanek DR Loge FJ Call DR
Simultaneous discrimination between 15 fish pathogens by using 16S
ribosomal DNA PCR and DNA microarrays Appl Environ Micro-
biol 2004704216ndash21
[19] Ludwig W Strunk O Westram R et al ARB a software environ-
ment for sequence data Nucleic Acids Res 2004321363ndash71
[20] Cole JR Chai B Marsh TL et al The ribosomal database project
(RDP-II) previewing a new autoaligner that allows regular updates
and the new prokaryotic taxonomy Nucleic Acids Res 200331
442ndash3
[21] Mitterer G Huber M Leidinger E et al Microarray-based
identification of bacteria in clinical samples by solid-phase PCR
amplification of 23S ribosomal DNA sequences J Clin Microbiol
2004421048ndash57
[22] Nubel U Schmidt PM Reiss E Bier F Beyer W Naumann D
Oligonucleotide microarrays for identification of Bacillus anthracis
based on intergenic transcribed spacers in ribosomal DNA FEMS
Microbiol Lett 2004240215ndash23
[23] Dahllof I Baillie H Kjelleberg S rpoB-based microbial community
analysis avoids limitations inherent in 16S rRNA gene intraspecies
heterogeneity Appl Environ Microbiol 2000663376ndash80
[24] Drancourt M Roux V Fournier PE Raoult D rpoB gene sequence-
based identification of aerobic Gram-positive cocci of the genera
Streptococcus Enterococcus Gemella Abiotrophia and Granulica-
tella J Clin Microbiol 200442497ndash504
[25] Mollet C Drancourt M Raoult D rpoB sequence analysis as a
novel basis for bacterial identification Mol Microbiol 199726
1005ndash1011
[26] Brisse S Verhoef J Phylogenetic diversity of Klebsiella pneumo-
niae and Klebsiella oxytoca clinical isolates revealed by randomly
amplified polymorphic DNA gyrA and parC genes sequencing
and automated ribotyping Int J Syst Evol Microbiol 200151
915ndash24
[27] Kakinuma K Fukushima M Kawaguchi R Detection and identi-
fication of Escherichia coli Shigella and Salmonella by microarrays
using the gyrB gene Biotechnol Bioeng 200383721ndash8
[28] Lloyd AT Sharp PM Evolution of the recA gene and the molecular
phylogeny of bacteria J Mol Evol 199337399ndash407
[29] Baldauf SL Palmer JD Doolittle WF The root of the universal tree
and the origin of eukaryotes based on elongation factor phylogeny
Proc Natl Acad Sci U S A 1996937749ndash54
[30] Ludwig W Neumaier J Klugbauer N et al Phylogenetic relation-
ships of Bacteria based on comparative sequence analysis of
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 117
elongation factor Tu and ATP-synthase beta-subunit genes Antonie
Van Leeuwenhoek 199364285ndash305
[31] Wong RSY Chow AW Identification of enteric pathogens by heat
shock protein 60 kDa (HSP60) gene sequences FEMS Microbiol
Lett 2002206107ndash13
[32] Wertz JE Goldstone C Gordon DM Riley MA A molecular
phylogeny of enteric bacteria and implications for a bacterial species
concept J Evol Biol 2003161236ndash48
[33] Zwieb C Gorodkin J Knudsen B Burks J Wower J tmRDB
(tmRNA database) Nucleic Acids Res 200331446ndash7
[34] Sergeev N Volokhov D Chizhikov V Rasooly A Simultaneous
analysis of multiple staphylococcal enterotoxin genes by an
oligonucleotide microarray assay J Clin Microbiol 200442
2134ndash43
[35] Chizhikov V Rasooly A Chumakov K Levy DD Microarray
analysis of microbial virulence factors Appl Environ Microbiol
2001673258ndash63
[36] Volokhov D Rasooly A Chumakov K Chizhikov V Identification
of Listeria species by microarray-based assay J Clin Microbiol 2002
404720ndash8
[37] Saunders NA Underwood A Kearns AM Hallas G A virulence-
associated gene microarray a tool for investigation of the evolution
and pathogenic potential of Staphylococcus aureus Microbiology
20041503763ndash71
[38] Grimm V Ezaki S Susa M Knabbe C Schmid RD Bachmann
TT Use of DNA microarrays for rapid genotyping of TEM beta-
lactamases that confer resistance J Clin Microbiol 200442
3766ndash3774
[39] Stralis-Pavese N Sessitsch A Weilharter A et al Optimisation of
diagnostic microarray for application in analysing landfill methano-
troph communities under different plant covers Environ Microbiol
20046347ndash63
[40] Rhee SK Liu X Wu L Chong SC Wan X Zhou J Detection of
genes involved in biodegradation and biotransformation in microbial
communities by using 50-mer oligonucleotide microarrays Appl
Environ Microbiol 2004704303ndash17
[41] Taroncher-Oldenburg G Griner EM Francis CA Ward BB
Oligonucleotide microarray for the study of functional gene diversity
in the nitrogen cycle in the environment Appl Environ Microbiol
2003691159ndash71
[42] Wu L Thompson DK Li G Hurt RA Tiedje JM Zhou J
Development and evaluation of functional gene arrays for detection
of selected genes in the environment Appl Environ Microbiol 2001
675780ndash90
[43] Hashsham SA Wick LM Rouillard JM Gulari E Tiedje JM
Potential of DNA microarrays for developing parallel detection tools
(PDTs) for microorganisms relevant to biodefense and related
research needs Biosens Bioelectron 200420668ndash83
[44] Wetmur JG DNA probes applications of the principles of
nucleic acid hybridization Crit Rev Biochem Mol Biol 199126
227ndash259
[45] Hughes TR Mao M Jones AR et al Expression profiling using
microarrays fabricated by an ink-jet oligonucleotide synthesizer Nat
Biotechnol 200119342ndash7
[46] Kane MD Jatkoe TA Stumpf CR Lu J Thomas JD Madore SJ
Assessment of the sensitivity and specificity of oligonucleotide
(50mer) microarrays Nucleic Acids Res 2000284552ndash7
[47] Tiquia SM Wu L Chong SC et al Evaluation of 50-mer
oligonucleotide arrays for detecting microbial populations in envi-
ronmental samples BioTechniques 200436664ndash5
[48] Letowski J Brousseau R Masson L Designing better probes effect
of probe size mismatch position and number on hybridization in
DNA oligonucleotide microarrays J Microbiol Methods 200457
269ndash78
[49] Vora GJ Meador CE Stenger DA Andreadis JD Nucleic acid
amplification strategies for DNA microarray-based pathogen detec-
tion Appl Environ Microbiol 2004703047ndash54
[50] Wang D Coscoy L Zylberberg M et al Microarray-based detection
and genotyping of viral pathogens Proc Natl Acad Sci U S A 2002
9915687ndash92
[51] Denef VJ Park J Rodrigues JL Tsoi TV Hashsham SA Tiedje JM
Validation of a more sensitive method for using spotted oligonucleo-
tide DNA microarrays for functional genomics studies on bacterial
communities Environ Microbiol 20035933ndash43
[52] Rouillard JM Zuker M Gulari E OligoArray 20 design of
oligonucleotide probes for DNA microarrays using a thermodynamic
approach Nucleic Acids Res 2003313057ndash62
[53] Gordon PM Sensen CW Osprey a comprehensive tool employing
novel methods for the design of oligonucleotides for DNA sequenc-
ing and microarrays Nucleic Acids Res 200432e133
[54] Behr T Koob C Schedl M et al A nested array of rRNA targeted
probes for the detection and identification of enterococci by reverse
hybridization Syst Appl Microbiol 200023563ndash72
[55] Wagner M Horn M Daims H Fluorescence in situ hybridization for
the identification and characterisation of prokaryotes Curr Opin
Microbiol 20036302ndash9
[56] Loy A Schulz C Lucker S et al 16S rRNA gene-based
oligonucleotide microarray for environmental monitoring of the
betaproteobacterial order lsquolsquoRhodocyclalesrsquorsquo Appl Environ Microbiol
200571(3)1373ndash86
[57] Bodrossy L Diagnostic oligonucleotide microarrays for microbiol-
ogy In Blalock E editor A beginnerrsquos guide to microarrays 1 ed
New Yorkrsquo Kluwer Academic Publishers 2003 p 43ndash92
[58] Peplies J Lau SC Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S
rRNA-based analysis of marine bacterioplankton Environ Microbiol
20046638ndash45
[59] Zuker M Mfold web server for nucleic acid folding and hybrid-
ization prediction Nucleic Acids Res 2003313406ndash15
[60] SantaLucia Jr J A unified view of polymer dumbbell and
oligonucleotide DNA nearest-neighbor thermodynamics Proc Natl
Acad Sci U S A 1998951460ndash5
[61] Pfunder M Frey JE Dissociation analysis in PCR- and 1xSSC-
buffer as a prerequisite for selection of 13mer microarray probe
sets with uniform hybiridzation behavior Mol Biotechnol 2005
29(1)1ndash10
[62] Al Khaldi SF Myers KM Rasooly A Chizhikov V Genotyping of
Clostridium perfringens toxins using multiple oligonucleotide micro-
array hybridization Mol Cell Probes 200418359ndash67
[63] Volokhov D Pomerantsev A Kivovich V Rasooly A Chizhikov V
Identification of Bacillus anthracis by multiprobe microarray hybrid-
ization Diagn Microbiol Infect Dis 200449163ndash71
[64] Liu WT Mirzabekov AD Stahl DA Optimization of an oligonu-
cleotide microchip for microbial identification studies a non-
equilibrium dissociation approach Environ Microbiol 20013
619ndash29
[65] Huang S Wang Y Chen P Qian HR Yeo A Bemis K Sum a new
way to incorporate mismatch probe measurements Genomics 2004
84767ndash77
[66] Lee I Dombkowski AA Athey BD Guidelines for incorporating
non-perfectly matched oligonucleotides into target-specific hybrid-
ization probes for a DNA microarray Nucleic Acids Res 2004
32681ndash90
[67] Shchepinov MS Case-Green SC Southern EM Steric factors
influencing hybridization of nucleic acids to oligonucleotide arrays
Nucleic Acids Res 1997251155ndash61
[68] Anthony RM Brown TJ French GL Rapid diagnosis of bacteremia
by universal amplification of 23S ribosomal DNA followed by
hybridization to an oligonucleotide array J Clin Microbiol 2000
38781ndash8
[69] Matveeva OV Shabalina SA Nemtsov VA Tsodikov AD Geste-
land RF Atkins JF Thermodynamic calculations and statistical
correlations for oligo-probes design Nucleic Acids Res 200331
4211ndash7
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119118
[70] Lane S Evermann J Loge F Call DR Amplicon secondary structure
prevents target hybridization to oligonucleotide microarrays Biosens
Bioelectron 200420728ndash35
[71] Southern EM DNA fingerprinting by hybridization to oligonucleo-
tide arrays Electrophoresis 1995161539ndash42
[72] Zhang XX Kosier B Priefer UB Genetic diversity of indigenous
Rhizobium leguminosarum bv viciae isolates nodulating two differ-
ent host plants during soil restoration with alfalfa Mol Ecol 2001
102297ndash305
[73] Chandler DP Newton GJ Small JA Daly DS Sequence versus
structure for the direct detection of 16S rRNA on planar
oligonucleotide microarrays Appl Environ Microbiol 200369
2950ndash8
[74] Maskos U Southern EM Parallel analysis of oligodeoxyribonucleo-
tide (oligonucleotide) interactions I Analysis of factors influencing
oligonucleotide duplex formation Nucleic Acids Res 199220
1675ndash8
[75] Jacobs KA Rudersdorf R Neill SD Dougherty JP Brown EL
Fritsch EF The thermal stability of oligonucleotide duplexes is
sequence independent in tetraalkylammonium salt solutions appli-
cation to identifying recombinant DNA clones Nucleic Acids Res
1988164637ndash50
[76] Volokhov D Chizhikov V Chumakov K Rasooly A Microarray-
based identification of thermophilic Campylobacter jejuni C coli C
lari and C upsaliensis J Clin Microbiol 2003414071ndash80
[77] Bonch-Osmolovskaya EA Miroshnichenko ML Lebedinsky AV et
al Radioisotopic culture-based and oligonucleotide microchip
analyses of thermophilic microbial communities in a continental
high-temperature petroleum reservoir Appl Environ Microbiol 2003
696143ndash51
[78] Koizumi Y Kelly JJ Nakagawa T et al Parallel characterization of
anaerobic toluene-and ethylbenzene-degrading microbial consortia
by PCR-denaturing gradient gel electrophoresis RNA-DNA mem-
brane hybridization and DNA microarray technology Appl Environ
Microbiol 2002683215ndash25
[79] Wang Z Vora GJ Stenger DA Detection and genotyping of
Entamoeba histolytica Entamoeba dispar Giardia lamblia and
Cryptosporidium parvum by oligonucleotide microarray J Clin
Microbiol 2004423262ndash71
[80] Lin B Vora GJ Thach D et al Use of oligonucleotide microarrays
for rapid detection and serotyping of acute respiratory disease-
associated adenoviruses J Clin Microbiol 2004423232ndash9
[81] Boriskin YS Rice PS Stabler RA et al DNA microarrays for virus
detection in cases of central nervous system infection J Clin
Microbiol 2004425811ndash8
[82] Panicker G Call DR Krug MJ Bej AK Detection of pathogenic
vibrio spp in shellfish by using multiplex PCR and DNA micro-
arrays Appl Environ Microbiol 2004707436ndash44
[83] Rudi K Rud I Holck A A novel multiplex quantitative DNA array
based PCR (MQDA-PCR) for quantification of transgenic maize in
food and feed Nucleic Acids Res 200331e62
[84] Ericsson O Sivertsson A Lundeberg J Ahmadian A Microarray-
based resequencing by apyrase-mediated allele-specific extension
Electrophoresis 2003243330ndash8
[85] Lindroos K Sigurdsson S Johansson K Ronnblom L Syvanen AC
Multiplex SNP genotyping in pooled DNA samples by a four-color
microarray system Nucleic Acids Res 200230e70
[86] Rudi K Treimo J Nissen H Vegarud G Protocols for 16S rDNA
array analyses of microbial communities by sequence-specific label-
ing of DNA probes Sci World J 20033578ndash84
[87] Busti E Bordoni R Castiglioni B et al Bacterial discrimination by
means of a universal array approach mediated by LDR (ligase
detection reaction) BMC Microbiol 2002227
[88] Gharizadeh B Kaller M Nyren P et al Viral and microbial
genotyping by a combination of multiplex competitive hybridization
and specific extension followed by hybridization to generic tag
arrays Nucleic Acids Res 200331e146
[89] Lovmar L Fock C Espinoza F Bucardo F Syvanen AC Bondeson
K Microarrays for genotyping human group a rotavirus by multiplex
capture and type-specific primer extension J Clin Microbiol 2003
415153ndash8
[90] Mikhailovich V Lapa S Gryadunov D et al Identification of
rifampicin-resistant Mycobacterium tuberculosis strains by hybrid-
ization PCR and ligase detection reaction on oligonucleotide
microchips J Clin Microbiol 2001392531ndash40
[91] Schena M Microarray analysis Hoboken Canadarsquo John Wiley and
Sons Inc 2003
[92] Rudi K Treimo J Moen B Rud I Vegarud G Internal controls for
normalizing DNA arrays BioTechniques 33 (2002) 496 498 500
[93] Cho JC Tiedje JM Quantitative detection of microbial genes
by using DNA microarrays Appl Environ Microbiol 200268
1425ndash30
[94] Straub TM Daly DS Wunshel S Rochelle PA DeLeon R Chandler
DP Genotyping Cryptosporidium parvum with an hsp70 single-
nucleotide polymorphism microarray Appl Environ Microbiol 2002
681817ndash26
[95] Gonzalez SF Krug MJ Nielsen ME Santos Y Call DR Simulta-
neous detection of marine fish pathogens by using multiplex PCR
and a DNA microarray J Clin Microbiol 2004421414ndash9
[96] Keramas G Bang DD Lund M et al Use of culture PCR analysis
and DNA microarrays for detection of Campylobacter jejuni and
Campylobacter coli from chicken feces J Clin Microbiol 200442
3985ndash91
[97] Keramas G Bang DD Lund M et al Development of a sensitive
DNA microarray suitable for rapid detection of Campylobacter spp
Mol Cell Probes 200317187ndash96
[98] Castiglioni B Rizzi E Frosini A et al Development of a universal
microarray based on the ligation detection reaction and 16S rRNA
gene polymorphism to target diversity of cyanobacteria Appl
Environ Microbiol 2004707161ndash72
[99] Lehner A Loy A Behr T Gaenge H Schleifer K-H Wagner M
Oligonucleotide microarray for simultaneous identification and
differentiation of Enterococcus species in food samples FEMS
Microbiol Lett 2005246(1)133ndash42
[100] Sergeev N Distler M Courtney S et al Multipathogen oligonucleo-
tide microarray for environmental and biodefense applications
Biosens Bioelectron 200420684ndash98
[101] Peplies J Lau SCK Pernthaler J Amann R Glockner FO
Application and validation of DNA microarrays for the 16S rRNA-
based analysis of marine bacterioplankton Appl Environ Microbiol
200571(5)2723ndash31
[102] Fukushima M Kakinuma K Hayashi H Nagai H Ito K Kawaguchi
R Detection and identification of Mycobacterium species isolates by
DNA microarray J Clin Microbiol 2003412605ndash15
[103] Deng JY Zhang XE Lu HB et al Multiplex detection of mutations
in clinical isolates of rifampin-resistant Mycobacterium tuberculosis
by short oligonucleotide ligation assay on DNA chips J Clin
Microbiol 2004424850ndash2
[104] Yu X Susa M Knabbe C Schmid RD Bachmann TT Development
and validation of a diagnostic DNA microarray to detect quinolone-
resistant Escherichia coli among clinical isolates J Clin Microbiol
2004424083ndash91
[105] Roth SB Jalava J Ruuskanen O Ruohola A Nikkari S Use of an
oligonucleotide array for laboratory diagnosis of bacteria responsible
for acute upper respiratory infections J Clin Microbiol 200442
4268ndash74
[106] Loy A Kusel K Lehner A Drake HL Wagner M Microarray and
functional gene analyses of sulfate-reducing prokaryotes in low-
sulfate acidic fens reveal cooccurrence of recognized genera and
novel lineages Appl Environ Microbiol 2004706998ndash7009
[107] Wilson WJ Strout CL DeSantis TZ Stilwell JL Carrano AV
Andersen GL Sequence-specific identification of 18 pathogenic
microorganisms using microarray technology Mol Cell Probes 2002
16119ndash27
A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004

A Loy L Bodrossy Clinica Chimica Acta 363 (2006) 106ndash119 119
[108] Chizhikov V Wagner M Ivshina A Hoshino Y Kapikian AZ
Chumakov K Detection and genotyping of human group a
rotaviruses by oligonucleotide microarray hybridization J Clin
Microbiol 2002402398ndash407
[109] Klaassen CH Prinsen CF de Valk HA Horrevorts AM Jeunink
MA Thunnissen FB DNA microarray format for detection and
subtyping of human papillomavirus J Clin Microbiol 200442
2152ndash60
[110] Delrio-Lafreniere SA Browning MK McGlennen RC Low-density
addressable array for the detection and typing of the human
papillomavirus Diagn Microbiol Infect Dis 20044823ndash31
[111] Oh TJ Kim CJ Woo SK et al Development and clinical
evaluation of a highly sensitive DNA microarray for detection and
genotyping of human papillomaviruses J Clin Microbiol 200442
3272ndash80
[112] Sengupta S Onodera K Lai A Melcher U Molecular detection and
identification of influenza viruses by oligonucleotide microarray
hybridization J Clin Microbiol 2003414542ndash50
[113] Kessler N Ferraris O Palmer K Marsh W Steel A Use of the DNA
flow-thru chip a three-dimensional biochip for typing and subtyping
of influenza viruses J Clin Microbiol 2004422173ndash85
[114] Laassri M Chizhikov V Mikheev M Shchelkunov S Chumakov K
Detection and discrimination of orthopoxviruses using microarrays of
immobilized oligonucleotides J Virol Methods 200311267ndash78
[115] Lapa S Mikheev M Shchelkunov S et al Species-level identifica-
tion of orthopoxviruses with an oligonucleotide microchip J Clin
Microbiol 200240753ndash7
[116] Liu RH Yang J Lenigk R Bonanno J Grodzinski P Self-contained
fully integrated biochip for sample preparation polymerase chain
reaction amplification and DNA microarray detection Anal Chem
2004761824ndash31
[117] Baner J Isaksson A Waldenstrom E Jarvius J Landegren U Nilsson
M Parallel gene analysis with allele-specific padlock probes and tag
microarrays Nucleic Acids Res 200331e103
[118] McKendry R Zhang J Arntz Y et al Multiple label-free biodetection
and quantitative DNA-binding assays on a nanomechanical cantilever
array Proc Natl Acad Sci U S A 2002999783ndash8
[119] Gabig-Ciminska M Los M Holmgren A et al Detection of
bacteriophage infection and prophage induction in bacterial cultures
by means of electric DNA chips Anal Biochem 200432484ndash91
[120] Zimmermann K Eiter T Scheiflinger F Consecutive analysis of
bacterial PCR samples on a single electronic microarray J Microbiol
Methods 200355471ndash4
[121] Westin L Miller C Vollmer D et al Antimicrobial resistance and
bacterial identification utilizing a microelectronic chip array J Clin
Microbiol 2001391097ndash104
[122] Meunier-Prest R Raveau S Finot E Legay G Cherkaoui-Malki M
Latruffe N Direct measurement of the melting temperature of
supported DNA by electrochemical method Nucleic Acids Res
200331e150
[123] Stimpson DI Hoijer JV Hsieh WT et al Real-time detection of
DNA hybridization and melting on oligonucleotide arrays by using
optical wave guides Proc Natl Acad Sci U S A 1995926379ndash83
[124] Nelson BP Liles MR Frederick KB Corn RM Goodman RM
Label-free detection of 16S ribosomal RNA hybridization on
reusable DNA arrays using surface plasmon resonance imaging
Environ Microbiol 20024735ndash43
[125] Podsadlowski V Peter C Cammann K Brunen-Nieweler C
Borchers T Rapid species identification in food samples with an
evanescent-field-DNA-biosensor-chip 2004 Statusseminar Chip
Technologies Microarrays Hygiene and Health Frankfurt a Main
26-2712004
Copyright © 2022 FDOKUMEN




















