
Regulation of ammonium ion assimilation enzymes inNeurospora crassa nit-2 andms-5 mutant strains

Agric. Biol. Chem., 45 (ll), 2417-2423, 1981 2417
Derepressed Syntheses of Sporulation Marker Enzymes
in a Bacillus Species Mutant
Akira Yokota and Ken-ichi Sasajima
Institute for Fermentation, Osaka, Juso-honmachi 2-chome, Yodogawa-ku, Osaka 532, Japan Received January 23, 1981
Sporulation marker enzymes, D-glucose dehydrogenase and L-alanine dehydrogenase weresynthesized derepressively by vegetative cells of a Bacillus species mutant which was isolated as animproved D-ribose producer. It was also elucidated by electron microscopy that no morphologicalchange concerning sporulation took place during the course of the enzyme syntheses in the mutantstrain. The presence of Mn2+ and Ca2+ in the medium was necessary for the morphologicaldevelopment of sporulation even in the mutant strain. The mechanism of derepressed enzymesyntheses is discussed in relation to regulation of sporulation.
The characteristics of GLD* (EC 1.1.1.47)of a Bacillus species mutant BG1722, whichwas isolated as an improved D-ribose pro-
ducer, were described in a previous paper.1}GLD was synthesized derepressively at thelate logarithmic growth phase in the mutant
strain.GLD is one of spore-specific enzymes in
bacilli. Its synthesis is one of the markerscharacterizing the sequential biochemicalchanges of sporulation.2~22) Properties of
GLDof B. cereus,13~16) B. megaterium11~20)
and B. subtilis8ai) have been described here-tofore. Fujita et al.21) reported recently that
GLD was not present in the mother cellcompartment but only in the forespore com-
partment. Piggot and Coote22) also reported
that GLD was not synthesized in the mutantstrains which were blocked before the stageIII (engulfment). This study was undertakento characterize the mutation of strain BG1722; relationship between the spore-specificbiochemical changes and the morphologicalchanges during sporulation.
* Abbreviations: GLD, D-glucose dehydrogenase;AlaD, L-alanine dehydrogenase; 8AGr, 8-azaguanine re-
sistance; ade, adenine requirement; xan, xanthine require-ment; tkt, transketolase deficiency; PGA, 3-phosphogly-
ceric acid.
MATERIALS AND METHODS
Bacteria. Bacillus species No. 102 (will type)23) and animproved D-ribose producer, mutant strain BG1722
(8AGr ade+ xan+ tkt) which was derived from a transke-tolase mutant BG1107 (8AGr ade xan tktfA) were used.
Media andgrowth conditions. Penassay broth containing1.75% Difco antibiotic medium No. 3, 4% sorbitol and0.01% each of L-tyrosine, L-tryptophan and l-
phenylalanine, or a modified Mangalo's medium (Mmedium)25) containing 0.5% sorbitol, 0.65% sodium l-
glutamate, 1.25mM KH2PO4, 0.1% Na2SO4, 0.01%
MgSO4-7H2O, 10mg of FeSO4 per liter, 4mg of biotinper liter, 3mg of thiamineà"HC1 per liter and 0.5 mMeachof L-tyrosine, L-tryptophan and L-phenylalanine was usedforcultureof bacteria. Where stated, M medium was
supplemented with 5/^m MnCl2 and/or 0.5mM CaCl2. A
loopful of the cells cultured on a nutrient agar slant wasinoculated into 20ml of Penassay broth in a 200-ml
Erlenmeyer flask and the cultures were incubated 24 hr at37°C on a rotary shaker. The cells were centrifuged andsuspended in an equal volume of M medium. This suspen-sion was used to inoculate 200-ml batches of M medium in1-liter Erlenmeyer flasks to an absorbancy of0.2 (650 nm).The flasks were incubated at 37°C on a rotary shaker andsingle flasks harvested at intervals of2 hr. Time courses ofgrowth and sporulation were followed by measurementsof absorbance at 650nm with a Gil ford spectrophotom-
eter, Model 240, and of refractile spores by phase-contrast microscopy, respectively.
Preparation of cell-free extracts. Individual flasks wereharvestedat intervals of 2 hr, and sample (10 ml) removed
for measurement of the culture absorbance at 650 nm andcounting refractile spores. The remained culture (190 ml)
2418 A. Yokota and K. Sasajima
was centrifuged at 13,000xg for 10 min and cells werewashedwith 0.05 m Tris-HCl buffer (pH 7.5) containing
lmM MgSO4, lmM CaCl2 and 0.1him ZnSO4, and sus-
pended in the same buffer. Subsequently, the cells were
d
isrupted with Kubota sonic oscillator at 160W for 10min under circulation of ice water. The cell debris were
removed by centrifugation and the supernatant solutionwas used for the determination of GLD activity. To
determine AlaD activity, lactate dehydrogenase should beinactivated by heating the supernatant at 65°C for 5 minand be removed by centrifugation at 12,000 x g for 10 min.Protein was determined by the method of Lowry et al.26)
Enzyme assays: The method of determination of GLDactivity was described in a previous paper.1} AlaD activitywas assayed at 30°C by the method ofWiame et al.21) In
all assays, 1 unit of the enzyme activity is defined as achange of absorbance of 0.001 per min at the wavelengthof 340 nm under the conditions described.
Measurement of refractility. Refractility was shown asthe percentage of the cells containing refractile spores
which were counted with a phase-contrast microscope.
Electron microscopy. The cells were dipped in 2.5%glutaraldehyde dissolved in acetate-Veronal buffer of
Kellenberger et al.28) for 1 hr at 4°C. The fixed cells were
washed in the same buffer and treated with 1% osmium inTryptone medium overnight at 4°C. After several wash-
ings with the buffer, the samples were suspended in 3%agar solution and allowed to solidify. The agar blocks
werethen treated with 0.5% uranyl acetate in acetate-Veronalbuffer for 2hr at 4°C. These were dehydrated
through an ethanol series (50, 70, 80, 90, 95 and 100%)and propylene oxide and embedded in Epon 812. Thinsections were obtained by cutting the embedded agar
blocks with a glass knife on a LKB 4800A ultramicro-
tome, mounted on collodion-coated 200-mesh coppergrids and stained with uranyl acetate and Reynold's lead
citrate.29) The specimens were examined and photo-
graphed in an electron microscope, Model JEM-T7S(Japan Electron Optics Laboratory, Co.) with an accel-
erating voltage of 60kV. The original plate magnifi-cations ranged from 5x 103 to 15x 103.
RESULTS
D
erepressed syntheses of sporulation marker
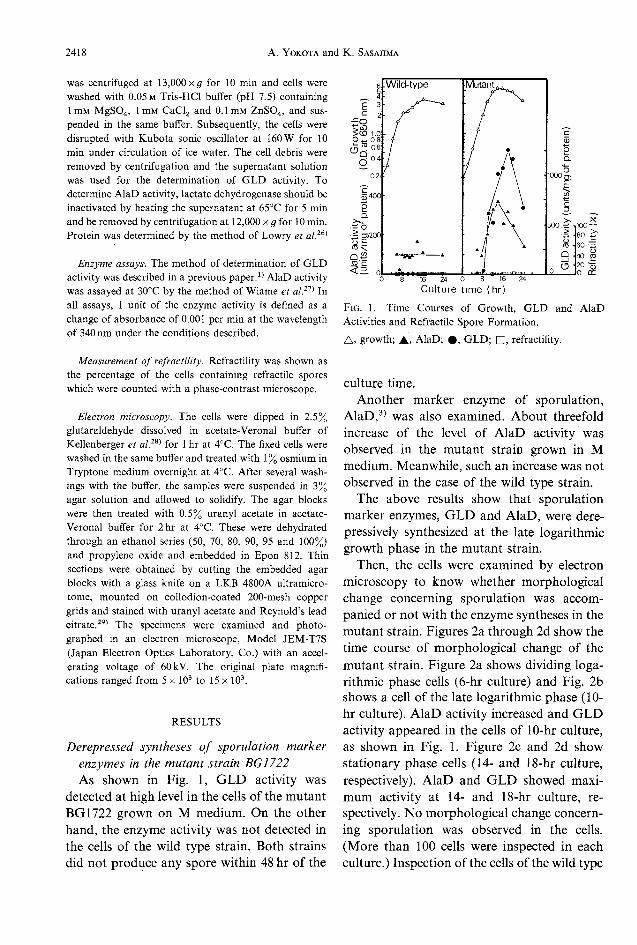
enzymes in the mutant strain BG1722As shown in Fig. 1, GLD activity was
detected at high level in the cells of the mutantBG1722 grown on M medium. On the otherhand, the enzyme activity was not detected inthe cells of the wild type strain. Both strainsdid not produce any spore within 48 hr of the
Wi ld-type[à"Mutant ^
Q.
^oo.tTI
3
CM
>e,100 -^-
å I60 -1
'£8 * 16 24 0 8 16 24
Culture time (hr)
Fig.1. Time Courses of Growth, GLD and AlaDActivities and Refractile Spore Formation.
A, growth; A, AlaD; #, GLD; Q, retractility.
culture time.Another marker enzyme of sporulation,
AlaD,3) was also examined. About threefoldincrease of the level of AlaD activity was
observed in the mutant strain grown in Mmedium.Meanwhile, such an increase was not
observed in the case of the wild type strain.
The above results show that sporulationmarker enzymes, GLD and AlaD, were dere-
pressively synthesized at the late logarithmicgrowth phase in the mutant strain.Then, the cells were examined by electronmicroscopy to know whether morphological
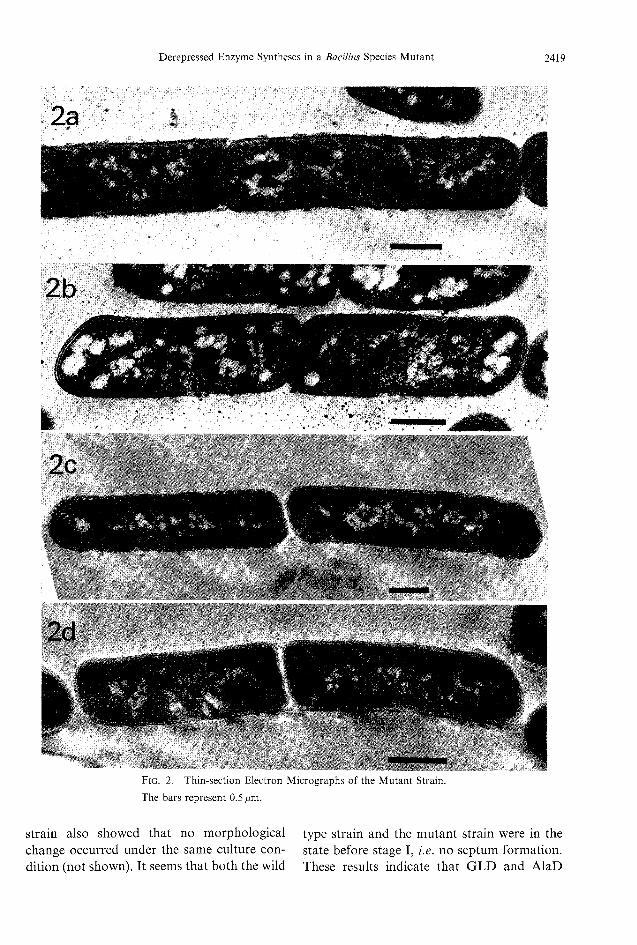
change concerning sporulation was accom-panied or not with the enzyme syntheses in themutant strain. Figures 2a through 2d show thetime course of morphological change of themutant strain. Figure 2a shows dividing loga-rithmic phase cells (6-hr culture) and Fig. 2bshows a cell of the late logarithmic phase (10-hr culture). AlaD activity increased and GLDactivity appeared in the cells of 10-hr culture,as shown in Fig. 1. Figure 2c and 2d showstationary phase cells (14- and 18-hr culture,
respectively). AlaD and GLD showed maxi-
mumactivity at 14- and 18-hr culture, re-spectively. No morphological change concern-
ing sporulation was observed in the cells.(More than 100 cells were inspected in each
culture.) Inspection of the cells of the wild type
Derepressed EnzymeSyntheses in a Bacillus Species Mutant 2419
2c
2d
Fig. 2. Thin-section Electron Micrographs of the Mutant Strain.The bars represent 0.5 /im.
strain also showed that no morphologicalchange occurred under the sameculture con-dition (not shown). It seems that both the wild
type strain and the mutant strain were in thestate before stage I, i.e. no septum formation.These results indicate that GLDand AlaD
2420 A. Yokota and K. Sasajima
£
24
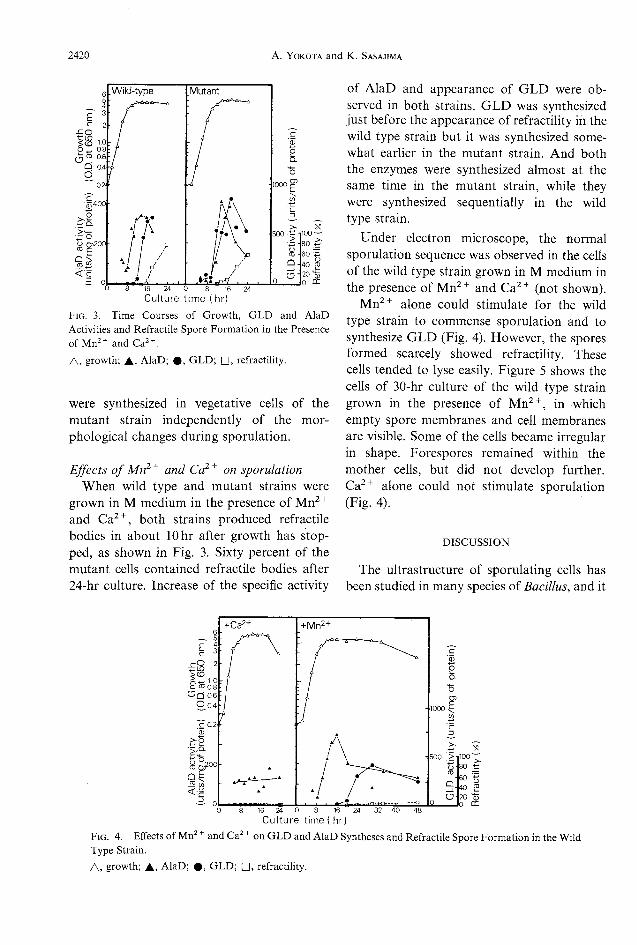
Culture time (hr)Fig. 3. Time Courses of Growth, GLDand AlaDActivities and Refractile Spore Formation in the PresenceofMn2+ and Ca2+.A, growth; A, AlaD; *. GLD; Q, retractility.
were synthesized in vegetative cells of themutant strain independently of the mor-phological changes during sporulation.
Effects ofMn2+and Ca2+ on sporulationWhenwild type and mutant strains weregrown in M medium in the presence of Mn2+and Ca2+, both strains produced refractilebodies in about lOhr after growth has stop-ped, as shown in Fig. 3. Sixty percent of themutant cells contained refractile bodies after24-hr culture. Increase of the specific activity
of AlaD and appearance of GLDwere ob-served in both strains. GLDwas synthesizedjust before the appearance of refractility in thewild type strain but it was synthesized some-what earlier in the mutant strain. And boththe enzymes were synthesized almost at thesame time in the mutant strain, while theywere synthesized sequentially in the wildtype strain.
Under electron microscope, the normalsporulation sequence was observed in the cellsof the wild type strain grown in Mmediuminthe presence of Mn2+and Ca2+ (not shown).
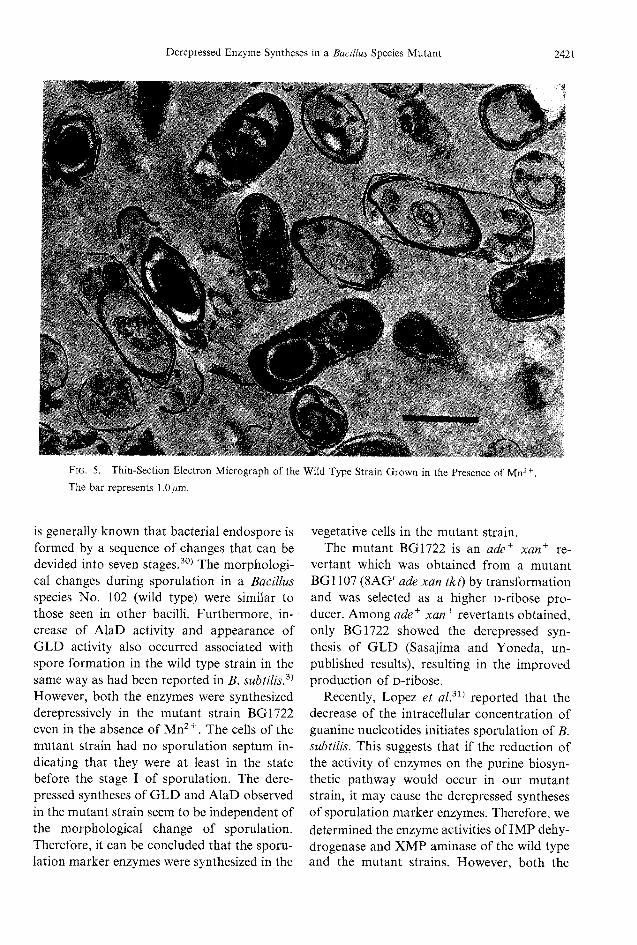
Mn2+alone could stimulate for the wildtype strain to commense sporulation and tosynthesize GLD(Fig. 4). However, the sporesformed scarcely showed refractility. Thesecells tended to lyse easily. Figure 5 shows thecells of 30-hr culture of the wild type straingrown in the presence of Mn2+, in whichempty spore membranes and cell membranesare visible. Some of the cells became irregularin shape. Forespores remained within themother cells, but did not develop further.Ca2+ alone could not stimulate sporulation(Fig. 4).
DISCUSSION
The ultrastructure of sporulating cells hasbeen studied in many species of Bacillus, and it
16 24 32 40 48
Culture time(hr)
Fig. 4. Effects ofMn2+ and Ca2+ on GLDand AlaD Syntheses and Refractile Spore Formation in the WildType Strain.
A, growth; A, AlaD; #, GLD; D, retractility.
Derepressed EnzymeSyntheses in a Bacillus Species Mutant 242 1
fcl<
Fig. 5. Thin-Section Electron Micrograph of the Wild Type Strain Grown in the Presence of Mn2+.The bar represents 1.0/urn.
is generally knownthat bacterial endospore isformed by a sequence of changes that can bedevided into seven stages.30) The morphologi-cal changes during sporulation in a Bacillusspecies No. 102 (wild type) were similar tothose seen in other bacilli. Furthermore, in-
crease of AlaD activity and appearance ofGLDactivity also occurred associated withspore formation in the wild type strain in thesame way as had been reported in B. subtilis.3}However, both the enzymeswere synthesizedderepressively in the mutant strain BG1722even in the absence of Mn2+.The cells of themutant strain had no sporulation septum in-dicating that they were at least in the statebefore the stage I of sporulation. The dere-pressed syntheses of GLDand AlaD observedin the mutant strain seem to be independent ofthe morphological change of sporulation.Therefore, it can be concluded that the sporu-lation marker enzymeswere synthesized in the
vegetative cells in the mutant strain.The mutant BG1722is an ade+ xan+ re-vertant which was obtained from a mutantBG1 107 (8AGr ade xan tkt) by transformationand was selected as a higher D-ribose pro-
ducer. Amongade+ xan+ revertants obtained,only BG1722 showed the derepressed syn-
thesis of GLD (Sasajima and Yoneda, un-published results), resulting in the improvedproduction of D-ribose.
Recently, Lopez et al?X) reported that thedecrease of the intracellular concentration ofguanine nucleotides initiates sporulation of B.subtilis. This suggests that if the reduction ofthe activity of enzymes on the purine biosyn-thetic pathway would occur in our mutantstrain, it maycause the derepressed synthesesof sporulation marker enzymes. Therefore, wedetermined the enzyme activities of IMPdehy-drogenase and XMPaminase of the wild typeand the mutant strains. However, both the

2422 A.Yokota and K. Sasajima
enzyme activities of the mutant strain weresimilar to those of the wild type strain (datanot shown). Furthermore, even when adenineand guanosine were added to the medium,
GLD was also synthesized derepressively inthe mutant strain (data not shown). Theseresults indicate that the derepressed enzymesyntheses was not caused by the decrease
of purine nucleotides.Keilman et al}1] and Beaman et al.32) alsoreported that many sporulation marker en-zymes including GLD were synthesized in thecells of stage 0 or I in B. subtilis even when
sporulation was inhibited by an antibioticnetropsin. This phenomenon is very similar tothe derepressed syntheses of sporulation mark-
er enzymes in our mutant strain describedhere.However, it was not confirmed whether
or not GLD activity detected in this case wasdue to inositol dehydrogenase (R. H. Doi,
personal communication), which appears dur-ing logarithmic growth phase as reported by
F
ujita et al.21)
Manganese ion is known to be an essentialelement for the sporulation of bacilli.33~35)
Several mechanisms of Mn2 + effects were elu-
cidated.36 ~40) Vasantha and Freese39) reported
that the strict Mn2+ requirement for sporu-
lationin B. subtilis results exclusively from theMn2+ requirement of PGA mutase. Theirresults suggests that the absence of Mn2+
probably cause the repression of sporulationin our wild type strain by a similar mechanism,but in the mutant strain the derepressed syn-theses of GLD and AlaD may occur overcom-ing the repression of sporulation by accumu-lation of PGA.
The results described in this paper mayindicate that a mutation related to the reg-
ulation of expression of sporulation-specificgenes has occurred in the mutant strain and
that GLD synthesis and increase of AlaDactivity which are accompanied by sporulation
in the wild type strain occur in the vegetativecells in the mutant strain derepressively andindependently of the morphological changes
concerning sporulation.It is a further problem to study the mech-
anism of the derepressed syntheses of thesesporulation marker enzymes in the mutant
strain to know some aspects of the regulatorymechanism of sporulation.
Acknowledgments. We wish to thank Dr. T. Iijima,Director of this Institute, for his encouragement and
valuable discussions. Grateful acknowledgment is alsomade to Mr. I. Miura, Central Research Division of
Takeda Chemical Industries, Ltd., for his advice onelectron microscopy. We also thank Mrs. Shinko
Sugimoto for her skillful assistance.
REFERENCES
1) A. Yokota, K. Sasajima and M. Yoneda, Agric. Biol.Chem., 43, 271 (1979).
2) K. F. Bott and R. Davidoff-Abelson, /. BacterioL,92, 229 (1966).
3) S. C. Warren, Biochem. J., 109, 811 (1968).4) D. Kay and S. C. Warren, Biochem. J., 109, 819
(1968).5) W. M.Waites,D. Kay,I. W. Dawes,D. A.Wood,S.
C.Warrenand J. Madelstam, Biochem. /., 118, 667(1970).
6) R. F. Ramaley and L. Burden, /. BacterioL, 101,1 (1970).
7) K. F. Bott, /. BacterioL, 108, 720 (1971).
8) Y. Hachisuka and K. Tochikubo, /. BacterioL, 107,442 (1971).
9) C. Prasad, M. Diesterhaft and E. Freese, /.
BacterioL, 110, 321 (1972).10) J. G. Coote, /. Gen. MicrobioL, 71, 1 (1972).
ll) G. R. Keilman, K. Burtis, B. Tanimoto and R. H.Doi, /. BacterioL, 128, 80 (1976).
12) M. Young, /. BacterioL, 126, 928 (1976).
1
3) R. Doi, H. Halvorson and B. Church, /. BacterioL,
ll, 43 (1959).14) J. A. Bach and H. L. Sadoff, /. BacterioL, 83, 699
(1962).
1
5) H. L. Sadoff, J. A. Bach andJ. W. Kools, "Spores,"Vol. 3, ed. by L.L. Campbell and H. O. Halvorson,
Am. Soc. MicrobioL, Washington, D. C, 1965, p. 97.
1
6) H. L. Sadoff, "Methods in Enzymology," Vol. 9, ed.
by W. A. Wood, Academic Press Inc., New York,1966, p. 103.
1
7) R. Gavard and C. Combre, Comptes Rendus Acad.Set, 249, 2243 (1959).
1
8) D. Banauch, W. Brummer, W. Ebeling, H. Metz, H.Rindfrey, H. Lang, H.Leybold and W. Rick,Z. Klin.
Chem. Klin. Biochem., 13, 101 (1975).19) H. E. Pauly and G. Pfleiderer, Hoppe-Seyler's
Physiol. Chem., 356, 1613 (1975).20) H. E. Pauly and G. Pfleiderer, Biochemistry, 16, 4599
(1977).
21) Y. Fujita, R. Ramaley and E. Freese, J. BacterioL,

Derepressed EnzymeSyntheses in a Bacillus Species Mutant 2423
132, 282 (1977).
P. J. Piggot and J. G. Coote, Bacteriol Rev., 40, 908(1976).
I. Nogami, M. Kida, T. Iijima and M. Yoneda,Agric. Biol. Chem., 32, 144 (1968).
K. Sasajima and M. Yoneda, Agric. Biol. Chem., 35,509 (1971).
R. Mangalo and J. T. Wachsman, J. Bacteriol., 83,27 (1962).
O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J.Randall, J. Biol. Chem., 193, 265 (1951).J. M. Wiame, A. Pierard and F. Ramos, "Methods inEnzymology," Vol. 5, ed. by S. P. Colowick and N.O. Kaplan, Academic Press Inc., New York, 1962, p.
673.
E. Kellenberger, A. Ryter and J. Sechaud, /.Biophysic. Biochem. Cytol, 4, 671 (1958).
E. S. Reynolds, J. Cell Biol, 17, 208 (1963).W. G. Murrell, Advan. Microbial Physiol, 1, 133
(1967).
31) J. M. Lopez, C. L. Marks and E. Freese, Biochim.Biophys. Ada, 587, 238 (1979).
32) B. L. Beaman, K. C. Burtis, R. H. Doi, J. P. Yeggy
and D. P. Stahly, Can. J. Microbiol, 26, 420 (1980).33) J. Charney, W. P. Fisher and C. P. Hegarty, /.
Bacterial, 62, 145 (1951).34) W. H. Crosby, R. A. Greene and R. A. Slepecky,
"Spore Research," ed. by A. N. Barker, G. W. Gouldand J. Wolf, Academic Press Inc., New York, 1971,
p.143.
35) E. D. Weinberg, Appl Microbiol, 12, 436 (1964).36) Y. K. Oh and E. Freese, /. BacterioL, 127, 739
(1976).
37) A. Akrigg and J. Mandelstam, Biochem. /., 172, 63(1978).
38) A. Akrigg, Biochem. J., Ill, 69 (1978).
39) N. Vasantha and E. Freese, J. Gen. Microbiol, 111,329 (1979).
40) R. P. Singh and P. Setlow, J. Bacterial, 139, 889
(1979).
Copyright © 2022 FDOKUMEN




















