
Regulation of ammonium ion assimilation enzymes inNeurospora crassa nit-2 andms-5 mutant strains
15
Biochemical Genetics, Vol.31, Nos. 9/10, 1993 Regulation of Ammonium Ion Assimilation Enzymes in Neurospora crassa nit-2 and ms-5 Mutant Strains Jorge Calder6n 1,3 and Luz Maria Martinez 2 Received 26 Feb. 1993--Final13Aug. 1993 In Neurospora crassa the nit-2 and nmr-1 (ms-5) loci represent the major control genes encoding regulatory proteins that allow the coordinated expression of various systems involved with the utilization of a secondary nitrogen source. In this paper we examine the effect of the nit-2 and ms-5 (nmr-1 locus) mutations on the regulation of the ammonium assimilation enzymes, glutamine synthetase and glutamate dehydrogenase, which are regulated by the products of these genes; however, glutamate synthase is not so regulated. Glutamine synthetase and glutamate dehydrogenase levels are also regulated by the amino nitrogen content. We present evidence that the ms-5 and gln r strains, which behave very similarly in their resistance to glutamine repression, are different and map in different loci. KEY WORDS: Neurospora; glutamine synthetase; glutamate dehydrogenase; glutamate synthase; regulation. INTRODUCTION The enzymes that participate in the assimilation of ammonium ion in Neurospora crassa are glutamine synthetase (GS; EC 6.3.1.2), NADP- dependent glutamate dehydrogenase (GDH; EC 1.4.1.4), and NADH- dependent glutamate synthase (GOGAT; EC 1.4.7.1) (Hummelt and Mora, This work was supported by DGAPA Grant IN203193 from the Universidad Nacional Au- tdnoma de M6xicoand by CONACYT. 1Departamento de EcologfaMolecular, Centro de Investigacidnsobre Fijacidn de Nitrdgeno, UNAM Cuernavaca,Apartado Postal 565-A, Mor., Mdxico. 2Departamento de Biotecnologfa,Instituto de InvestigacionesBiom6dicas,Universidad Nacio- nal Autdnoma de M6xico,Apartado Postal 70228, M6xicoD.F., C.P. 4510. 3To whomcorrespondenceshould be addressed. 425 0006-2928/93/1000-0425507.00/0 © 1993 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Regulation of ammonium ion assimilation enzymes inNeurospora crassa nit-2 andms-5 mutant strains

Biochemical Genetics, Vol. 31, Nos. 9/10, 1993
Regulation of Ammonium Ion Assimilation Enzymes in Neurospora crassa nit-2 and ms-5 Mutant Strains
Jorge Calder6n 1,3 and Luz Maria Mart inez 2
Received 26 Feb. 1993--Final 13Aug. 1993
In Neurospora crassa the nit-2 and nmr-1 (ms-5) loci represent the major control genes encoding regulatory proteins that allow the coordinated expression of various systems involved with the utilization of a secondary nitrogen source. In this paper we examine the effect of the nit-2 and ms-5 (nmr-1 locus) mutations on the regulation of the ammonium assimilation enzymes, glutamine synthetase and glutamate dehydrogenase, which are regulated by the products of these genes; however, glutamate synthase is not so regulated. Glutamine synthetase and glutamate dehydrogenase levels are also regulated by the amino nitrogen content. We present evidence that the ms-5 and gln r strains, which behave very similarly in their resistance to glutamine repression, are different and map in different loci.
KEY WORDS: Neurospora; glutamine synthetase; glutamate dehydrogenase; glutamate synthase; regulation.
INTRODUCTION
The enzymes that participate in the assimilation of ammonium ion in Neurospora crassa are glutamine synthetase (GS; EC 6.3.1.2), NADP- dependent glutamate dehydrogenase (GDH; EC 1.4.1.4), and NADH- dependent glutamate synthase (GOGAT; EC 1.4.7.1) (Hummelt and Mora,
This work was supported by DGAPA Grant IN203193 from the Universidad Nacional Au- tdnoma de M6xico and by CONACYT.
1 Departamento de Ecologfa Molecular, Centro de Investigacidn sobre Fijacidn de Nitrdgeno, UNAM Cuernavaca, Apartado Postal 565-A, Mor., Mdxico.
2 Departamento de Biotecnologfa, Instituto de Investigaciones Biom6dicas, Universidad Nacio- nal Autdnoma de M6xico, Apartado Postal 70228, M6xico D.F., C.P. 4510.
3 To whom correspondence should be addressed.
425 0006-2928/93/1000-0425507.00/0 © 1993 Plenum Publishing Corporation

426 Calder6n and Martinez
1980a,b). GS is composed of two polypeptides (Calder6n et al., 1990; Dfivila et aL, 1980; Sfinchez et al., 1980) and glutamine regulates the level of specific enzyme synthesis (Vichido et aL, 1978), which correlates with the amount of mRNA (Sfinchez et aL, 1978; Lara et aL, 1982). The carbon source also regulates GS (Mora et al., 1980). GDH is a hexamer, comprising six identical subunits, which has been physicochemically and genetically characterized; multiple mutants with altered GDH structure have been obtained, its amino acid sequence has been established, and its structural gene has been cloned (Brett et aL, 1976; Fincham, 1950; Fincham and Baron, 1977; Holder et aL, 1975; Kinnaird et aL, 1982; Scale et al., 1977; Sidding et al., 1980). GDH is regulated by the nitrogen source at the level of de novo enzyme synthesis (Hernfindez et aL, 1983), and its activity is also regulated by the carbon source (Dantzing et aL, 1979). GOGAT is a monomer with a molecular weight of 200,000, and its activity is repressed by glutamate (Hummelt and Mora, 1980b; Lomnitz et al., 1987). GOGAT participates in a glutamine cycle in which glutamate and glutamine are continually resynthesized (Cal- der6n et aL, 1985, 1989; Calder6n and Mora, 1985, 1989; Mora et al., 1988).
Glutamine, the final product of ammonium ion assimilation, plays a central role in microorganisms. It is required for protein synthesis and as a nitrogen donor for a number of biosynthetic reactions, and it has been proposed as the metabolite responsible for nitrogen catabolic repression in several microorganisms, including N. crassa (Vaca and Mora, 1977; Marzluf, 1981). Nitrogen catabolic repression is the process by which favored nitro- gen sources such as ammonium ion and L-glutamine (primary nitrogen sources in N. crassa) are utilized preferentially through repressing the synthesis of many permeases and enzymes necessary for utilization of other nitrogen sources (secondary nitrogen source), such as nitrate, nitrite, pu- fine, and amino acids (Marzluf, 1981).
In Neurospora crassa, the product of the nit-2 gene is required for expression of genes involved with the utilization of secondary nitrogen sources (Coddington, 1976; Facklan and Marzluf, 1978; Fu and Marzluf, 1987, 1988, 1990a; Marzluf, 1981; Reinert and Marzluf, 1975). The nit-2 gene appears to encode a regulatory protein that acts in a positive fashion to activate the expression of a large family of unlinked nitrogen structural genes (Marzluf, 1981). The nit-2 gene has been cloned (Stewart and Vollmer, 1986) and characterized (Fu and Marzluf, 1987, 1990a): it encodes a protein of 1036 amino acid residues with a single zinc finger domain that functions as a site-specific DNA-binding protein (Fu and Marzluf, 1987, 1990b).
In N. crassa, two allelic mutant strains have been isolated (nmr-1 and ms-5) which fail to respond to nitrogen catabolic repression (Dunn-Coleman et al., 1981; Premakumar et al., 1980). nmr is unlinked to nit-2 and appears to have a regulatory function, acting in a negative manner to repress the

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 427
synthesis of nitrate reductase and other nitrogen catabolic enzymes, such as L-amino acid oxidase and histidase (DeBusk and Ogilvie, 1984; Dunn- Coleman et al., 1981; Premakumar et al., 1980). Mutations in the nmr locus relieve glutamine-mediated repression, so that high levels of ammonia or glutamine fail to repress the synthesis of nitrate reductase (DeBusk and Ogilvie, 1984; Dunn-Coleman et al., 1981; Premakumar et al., 1980). The nmr gene product does not directly repress the expression of the nit-2 gene (Fu and Marzluf, 1987). The nmr gene has been cloned and shown to be expressed constitutively (Fu et al., 1988). The ms-5 mutant strain contains a single base change which results in the substitution of aspartate for glycine at residue 386 of the nrnr gene (Jarai and Marzluf, 1990). Also, a N. crassa mutant has been isolated (glnr), whose arginine catabolism is resistant to glutamine repression. Arginase and glutamine synthetase are less sensitive to repression by glutamine in this mutant which accumulates glutamine, arginine, and other amino acids during exponential growth (Gonzfilez et al., 1983).
The ammonium ion assimilation enzymes are regulated by the nitrogen source, and the nit-2 and nmr loci represent major control genes which coordinate expression of the various systems involved with the utilization of secondary nitrogen sources. In this paper we examine the effect of the nit-2 and ms-5 (nmr-1 locus) mutations on the regulation of the ammonium ion assimilation enzymes. Also, evidence is presented that shows that the ms-5 andgln r mutations are in different loci.
M A T E R I A L S A N D M E T H O D S
Strains. All the strains came either from the collection of J. Mora Centro de Investigaci6n sobre Fijaci6n de Nitr6geno Cuernavaca Morelos, M6xico, or from the Fungal Genetic Stock Center, Kansas City, Kansas. The basic stocks were the 74-A wild-type strain; the nit-2, ms-5, and gln r regulatory mutant strains (Coddington, 1976; Gonzfilez et aL, 1983; Premakumar et aL, 1980); the gln- lbR8 strain, which lacks the GS[3 monomer (D[41d] aavila et al., 1983); and the nit-2;gln-lbR8 strain.
Growth Conditions. N. crassa was grown on the liquid minimal medium of Vogel (1964) supplemented with 1.5% (w/v) sucrose. In some experi- ments the carbon and nitrogen sources were substituted as stated in the text. Batch-fed, ammonium ion-limited cultures of iV. crassa were achieved by pumping NH4CI at a dilution rate of 0.03 Ixmol m1-1 hr -1 into an aerated reactor vessel held at 30°C containing Vogel medium lacking the nitrogen source (Limon-Lason et aL, 1977). Conidia were harvested from slants of Vogel medium supplemented with 1.5% (w/v) sucrose, which had been

428 Calder6n and Martlnez
grown in the dark for 3 days at 29°C, followed by 2 days under incandescent light at 25°C.
Cultures were started by inoculating conidia into Erlenmeyer flasks containing various media. The initial optical density of each culture at 540 nm was 0.05. Incubation was carried out for 12 hr at 30°C with shaking at 250 rpm.
Crosses were made on Westergaard synthetic crossing medium (West- ergaard and Mitchell, 1947) as described (Davis and de Serres, 1970).
Determination of Enzyme Activities. Cell-free extracts of N. crassa were prepared as described previously (D~tvila et al., 1978). GS activity was measured by its synthetase activity as described by Ferguson and Sims (1974). NADP-GDH was measured as described by Fincham (1957), and GOGAT as by Hummelt and Mora (1980a). Nitrate reductase was assayed as described elsewhere (Sorger et aL, 1974).
Determination of Amino Acid Pools. Samples were prepared by homog- enizing mycelium with 80% (v/v) ethanol. The homogenates were boiled for 10 min, cooled, and filtered through membrane filters (Type RAWP047; Millipore Corp., Bedford, Massachusetts). The filtrates were lyophilized and the samples were resuspended in lithium citrate buffer (pH 2.88). The amino acids were separated with an Aminco Amino Acid Analyzer and quantified in an Aminco Ratio Fluorimeter after coupling with o-phthalalde- hyde. Arginine was extracted from filtered mycelium with 2 ml of 5% (w/v) trichloroacetic acid. After centrifugation, arginine was determined in the supernatant fluid by the method of Van Pilsum et al. (1956).
Immunoprecipitation of GS Activity. GS was purified to homogeneity by DEAE-cellulose chromatography, followed by affinity chromatography on anthranilate-bound Sepharose (Palacios, 1976). Rabbits were immunized as described previously (Palacios et al., 1977). Immunoprecipitation was done by using N. crassa extracts containing a constant amount of protein. The extracts were incubated for 90 min at 4°C in the presence of synthetase buffer (50 mM K2804, 50 mm imidazole, 50 mM 2-mercaptoethanol, pH 7.2) with anti-GS serum; the final volume was 0.5 ml. After incubation the enzyme was measured in the supernatant. Sufficient anti-GS (200 Ixl) was added to precipitate all GS activity from the supernatant; the immunopre- cipitates were used for gel electrophoresis.
Electrophoresis. The immunoprecipitates were subjected to polyacryl- amide slab gel electrophoresis in the presence of sodium dodecyl sialfate and 7 M urea (Sfinchez et aL, 1980). The gels were stained with Coomassie blue and scanned with a transmittance/reflactence scanning densitometer (Hoefer Scientific Instruments).
Protein Determination. Samples of mycelium were collected on mem- brane filters (Type HA, 0.45 Ixm; Millipore Corp.) and washed with 2 volumes of distilled water. The suspended samples were then precipitated

with 2 ml of 5% (w/v) trichloracetic and centrifuged for 5 rain at 2000 rpm, and the pellets were suspended in 1.0 N NaOH. Protein was determined by the method of Lowry et al. (1951).
RESULTS
1 O0
Growth of the Wild-Type, nit-2, and ms-5 Strains Under Different Nitrogen and Carbon Conditions. The nit-2 strain showed a growth pattern similar to that of the wild-type strain on excess ammonium ion or glutamine plus sucrose as the nitrogen and carbon source, respectively, or when glycerol was used as the carbon source, instead of sucrose, with excess ammonium ion. However, it grew more slowly than the wild-type strain on glutamate, or on ammonium ion batch-fed limited cultures, plus sucrose as the nitrogen and carbon source, respectively (Figs. 1 and 2). To observe a more drastic effect of the nit-2 mutation on N. crassa growth, we obtained a double mutant strain of the nit-2 strain with the gln-lbR8 strain that expresses only GSc~ and has a lower GS activity than the wild-type strain. The nit-2;gln-lbR8 strain almost failed to grow on ammonium ion-limited medium, while thegIn-lbR8, after a lag phase, reached a yield similar to that of the wild-type strain (Fig. 2). The ms-5 strain grew slightly more slowly than the wild-type strain under all
1 2
E
o
10
A
/
t i i I I
6 12 3 6 12 6 12
Time (h)
Regulation ofGS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 429
Fig. 1. Growth curves of the wild-type (O), nit-2 (D), and ms-5 (~) strains from cultures grown on 25 mM NH4NO3 (A), 5 mM glutamine (B), 5 mM glutamate (C), and 25 mM NH4NO3 with 2% glycerol instead of 2% sucrose (D).
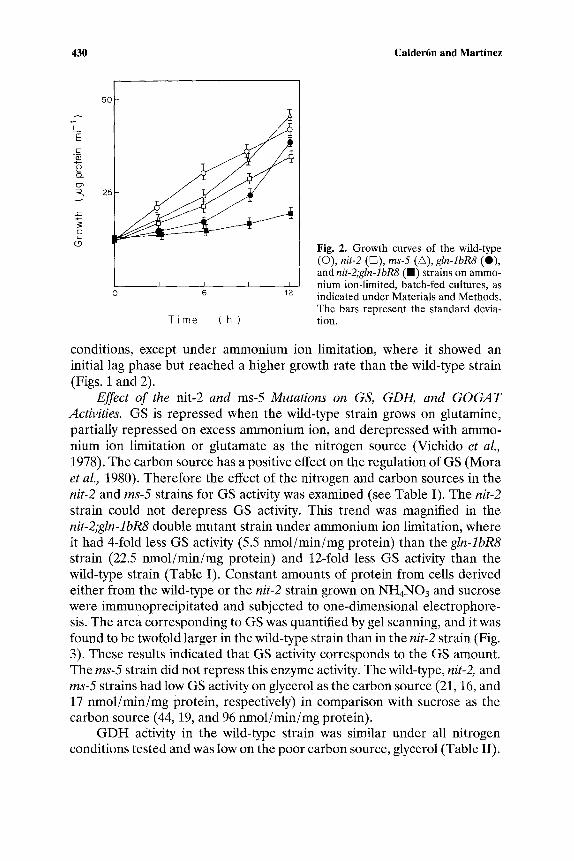
430 Calder6n and Martinez
L E c::
o
o
(._9
5 0
25
I I I I 0 6 1£
T i m e ( h )
Fig. 2. Growth curves of the wild-type (0), nit-2 ([:5), ms-5 (A), gln-lbR8 (0), and nit-2;gln-lbR8 ( I ) strains on ammo- nium ion-limited, batch-fed cultures, as indicated under Materials and Methods. The bars represent the standard devia- tion.
conditions, except under ammonium ion limitation, where it showed an initial lag phase but reached a higher growth rate than the wild-type strain (Figs. 1 and 2).
Effect of the nit-2 and ms-5 Mutations on GS, GDH, and G O G A T Activities. GS is repressed when the wild-type strain grows on glutamine, partially repressed on excess ammonium ion, and derepressed with ammo- nium ion limitation or glutamate as the nitrogen source (Vichido et al., 1978). The carbon source has a positive effect on the regulation of GS (Mora et al., 1980). Therefore the effect of the nitrogen and carbon sources in the nit-2 and ms-5 strains for GS activity was examined (see Table I). The nit-2 strain could not derepress GS activity. This trend was magnified in the nit-2;gln-lbR8 double mutant strain under ammonium ion limitation, where it had 4-fold less GS activity (5.5 nmol/min/mg protein) than the gln-lbR8 strain (22.5 nmol/min/mg protein) and 12-fold less GS activity than the wild-type strain (Table I). Constant amounts of protein from cells derived either from the wild-type or the nit-2 strain grown on NH4NO 3 and sucrose were immunoprecipitated and subjected to one-dimensional electrophore- sis. The area corresponding to GS was quantified by gel scanning, and it was found to be twofold larger in the wild-type strain than in the nit-2 strain (Fig. 3). These results indicated that GS activity corresponds to the GS amount. The ms-5 strain did not repress this enzyme activity. The wild-type, nit-2, and ms-5 strains had low GS activity on glycerol as the carbon source (21, 16, and 17 nmol/min/mg protein, respectively) in comparison with sucrose as the carbon source (44, 19, and 96 nmol/min/mg protein).
GDH ac'tivity in the wild-type strain was similar under all nitrogen conditions tested and was low on the poor carbon source, glycerol (Table II).

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 431
Table 1. Glutamine Synthetase Activity in the Wild Type, nit-2, and ms-5 Strains Grown Under Different Conditions a
i
Strain
Condition Wild type nit-2 ms-5
NH4NO3 + sucrose b 44 (4) 19 (4) 96 (8) Glutamine + sucrose b 13 (2) 9 (1) 68 (5) Glutamate + sucrose b 113 (8) 40 (3) 127 (9) Ammonium-limited + sucrose c 68 (5) 20 (4) 53 (5) NH4NO3 + glycerol d 21 (2) 16 (2) 17 (3)
aGlutamine synthetase activity was determined after 12 hr of growth and is expressed as nano- moles of c~-glutamyl hydroxamate produced per minute per milligram of protein. The numbers in parentheses represent the standard deviations.
b25 ~ NH4NO3, 5 mM glutamate, and 5 mM glutamine plus 1.5% sucrose were used as the nitrogen and carbon sources, respectively.
CBatch-fed, ammonium ion-limited cultures (as indicated under Materials and Methods) plus 1.5% sucrose.
d25 mM Ntl4NO 3 and 2% glycerol were used as the nitrogen and carbon sources.
The nit-2 strain had lower G D H activity than the wild-type strain on glutamine, being 3.5-fold lower. The ms-5 strain had a slightly higher G D H activity than the wild-type strain, except on glutamate, where G D H activity was similar to that in the wild-type strain (Table II).
G O G A T activity in the wild-type strain was induced under nitrogen- limited conditions and repressed by glutamate as reported previously (Hum- melt and Mora 1980b; Lomnitz et al., 1987) (Table III) . In contrast with G D H and GS, the nit-2 strain did not exhibit reduced G O G A T activity and, instead, showed higher G O G A T activity under all conditions tested than the wild-type strain (Table III) . The ms-5 strain had G O G A T activity similar to that of the n#-2 strain, except on ammonium ion limitation, where this activity was even lower than in the wild-type strain (Table III) . However, glutamate repressed G O G A T activity in all strains (Table III) .
Glutamine, Glutamate, and Alanine Content in the Wild-Type, nit-2, and ms-5 Strains Grown Under Different Nitrogen and Carbon Conditions. In comparison with the wild-type strain, the nit-2 strain had a very low glutamine content under all conditions and was 20-fold less when grown in NH4NOa and glutamine. In contrast, the ms-5 strain had a very high glutamine content, 15-fold higher than in the wild type on NH4NO 3 and glutamine (Table IV).
The nit-2 strain had a higher glutamate content on glutamine and ammonium ion limitation (2.5-fold) than the wild-type strain, and the ms-5 strain had a glutamate content similar to that of the wild-type strairo, except
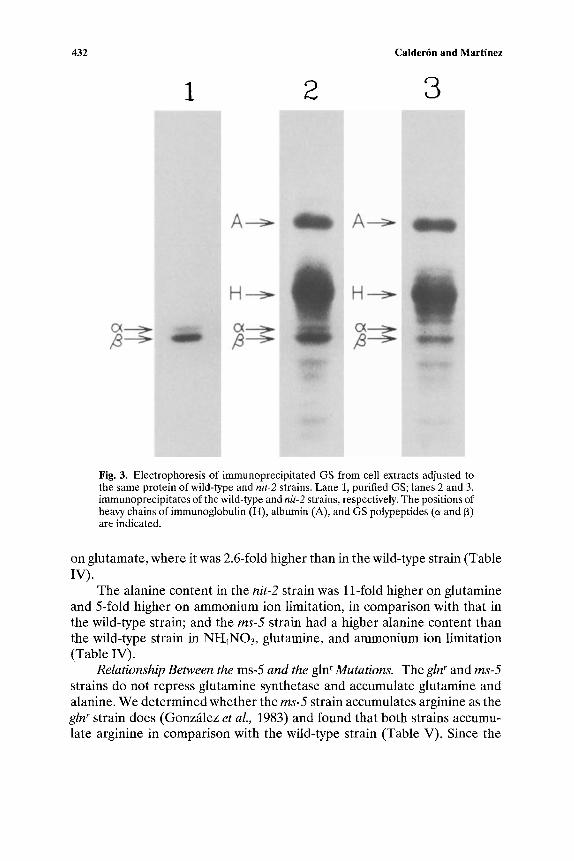
432 Calder6n and Martlnez
1 2 3 ii!i~i iiiiii~i!i! ii i~ ~
i'ii~i')i/~ ' ii ~,i'i~i~?,iiiil ' ~i ii i~' ,i ~,~, ; ~' !!ii :ii~:: ii~i!i i!~i~:i!%?~i ~
i'i/iii~! !!i!ili~ ~ ~ii~ii~i!!~ % .......... ; i ;!~ii ii:~
H -----~ [1 -~
:~:~ ~!:1%%i/i~!! !ii~i i! i ~ ~ ~!!i~!~ ~(, ~ .....
Fig. 3. Electrophoresis of immunoprecipitated GS from cell extracts adjusted to the same protein of wild-type and nit-2 strains. Lane 1, purified GS; lanes 2 and 3, immunoprecipitates of the wild-type and nit-2 strains, respectively. The positions of heavy chains of immunoglobulin (H), albumin (A), and GS polypeptides (a and [3) are indicated.
on glutamate, where it was 2.6-fold higher than in the wild-type strain (Table IV).
The alanine content in the nit-2 strata was 1 l-fold higher on glutamine and 5-fold higher on ammonium ion limitation, in comparison with that in the wild-type strain; and the ms-5 strain had a higher alanine content than the wild-type strain in NHaNO3, glutamine, and ammonium ion limitation (Table W).
Relationship Between the ms-5 and the gln r Mutations. The gln r and ms-5 strains do not repress glutamine synthetase and accumulate glutamine and alanine. We determined whether the ms-5 strain accumulates arginine as the gln r strain does (Gonzfilez et al., 1983) and found that both strains accumu- late arginine in comparison with the wild-type strain (Table V). Since the

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms.5 Mutant Strains 433
Table II. Glutamate Dehydrogenase Activity in the Wild-Type, nit-2, and ms-5 Strains Grown Under Different Conditions a
Strain
Condition Wild type nit-2 ms-5
NH4NO3 + sucrose b 1.2 (0.2) 0.5 (0.1) 1.6 (0.3) Glutamine + sucrose b 1.4 (0.3) 0.4 (0.1) 2.8 (0.3) Glutamate + sucrose b 1.2 (0.3) 1.0 (0.1) 1.1 (0.2) Ammonium-limited + sucrose c 1.4 (0.3) 0.8 (0.1) 1.7 (0.3)
NH4NO 3 + glyceroF 0.7 (0.2) 0,3 (0.1) 1.0 (0.2)
aGlutamate dehydrogenase activity was determined after 12 hr of growth and is expressed as micromoles of NADPH oxidized per minute per milligram of protein. The numbers in paren- theses represent the standard deviations.
b25 mM NH4NO3, 5 mM glutamate, and 5 mM glutamine plus 1.5% sucrose were used as the nitrogen and carbon sources, respectively.
CBatch-fed, ammonium ion-limited cultures (as indicated under Materials and Methods) plus 1.5% sucrose.
a25 mM NH4NO3 and 2% glycerol were used as the nitrogen and carbon sources.
two strains behave similarly, we decided to measure nitrate reductase activity, which has been shown not to be repressed in the ms-5 strain (Premakumar et al., 1980). We found that in the gin r strain nitrate reductase was repressed, in contrast with the ms-5 strain (Table V).
In accordance with the nitrogen repression of nitrate reductase in the
Table III. Glutamate Synthase Activity in the Wild-Type, nit-2, and ms-5 Strains Grown Under Different Conditions a
Strain
Condition Wild type nit-2 ms-5
NH4NO 3 + sucrose b 6 (1) 9 (1) 11 (2) Glutamine + sucrose b 7 (t) 10 (1) 12 (2) Glutamate + sucrose b 3 ( l ) 4 (1) 5 (1) Ammonium-limited + sucrose c 17 (2) 22 (2) 13 (2) NH4NO3 + glycerol a 8 (1) 10 (1) 10 (2)
aGlutamate synthase activity was determined after 12 hr of growth and is expressed as nano- moles of NADH oxidized per minute per milligram of protein. The numbers in parentheses represent the standard deviations.
b25 mM NH4NO3, 5 mM glutamate, and 5 mM glutamine plus 1.5% sucrose were used as the nitrogen and carbon sources, respectively.
CBatch-fed, ammonium ion-limited cultures (as indicated under Materials and Methods) plus 1.5% sucrose.
a25 irlM NH4NO3 and 2% glycerol were used as the nitrogen and carbon sources.

434 Calder6n and Martlnez
Table IV. Glutamine, Glutamate, and Alanine Pools in the Wild-Type nit-2, and ms-5 Strains Grown Under Different Conditions a
Strain
Wild type nit-2 ms-5
Condition Gin Glu Ala Gin Glu Ala Gin Glu Ala
NH4NO3 b 0.09 0.47 0.09 0.004 0.34 0.18 1 . 3 1 0.34 0.45 Glutamine b 0.10 0 . 3 3 0.08 0 .005 0.86 0 . 9 1 1 . 5 6 0.42 0.20 Glutamate b 0.06 0.46 0.26 ND e 0 . 3 1 0 . 1 1 0.32 1 . 2 2 0.30 Ammonium ion-limited c ND 0 . 0 5 0.01 ND 0.13 0 . 0 5 0 . 0 1 0 . 0 5 0.20 NH4NO3 a 0.02 0.16 0.12 0 .008 0 . 1 3 0.10 0.14 0 . 1 5 0.18
i i i i
aThe amino acid determinations were made after 12 hr of growth and they are micromoles per milligram of protein,
expressed as
b25 mM NH4NO3, 5 mM glutamate, and 5 mM glutamine plus 1.5% sucrose were used as the nitrogen and carbon sources, respectively,
CBatch-fed, ammonium ion-limited cultures (as indicated under Materials and Methods) plus 1.5% sucrose.
~25 mM NH4NO3 and 2% glycerol were used as the nitrogen and carbon sources. eNot detected.
gi f t r strain, we found that this strain was resistant to chlorate in the presence of ammonium ion or glutamine (data not shown) (Cove, 1976, 1979). The gln r mutation confers resistance to L-methionine sulfoximine, an inhibitor of glutamine synthetase (Gonzfilez et aL, 1983; Ronzio and Meister, 1968). We found that the ms-5 strain was also resistant to L-methionine sulfoximine. To determine the linkage relationship of the ms-5 and gl f l r mutations, we followed the differences already mentioned, in order to identify the progeny of the cross between these two strains. The result obtained, that 23% of the progeny had a wild-type phenotype, indicated that these mutations are not allelic.
Table V. Nitrate Reductase Activity and Arginine Pool in the Wild Type, ms-5, and gin'* i i i i
Strain Arginine b Nitrate reductase c
Wild type 0.1 28.4 ms-5 0.3 146.9 gin r 0.2 16.3
aNitrate reductase activity and arginine pool were determined after 12 hr of growth on 25 mM NH4NO3 and 1.5% sucrose as the nitrogen and carbon sources.
bThe arginine pool is expressed as micromoles per milligram of protein. CNitrate reductase activity is expressed as nanomoles of NADPH per minute per milligram of protein.

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 435
DISCUSSION
The regulation by nitrogen and carbon sources of the N. crassa enzymes that participate in the assimilation of ammonium ion (GS, GDH, and GOGAT) is well documented (Calder6n et al., 1989, 1990; Calderdn and Mora, 1985, 1989; Dantzing et al., 1979; Hernfindez et al., 1983; Hummelt and Mora, 1980b; Lara et al., 1982; Lomnitz et al., 1987; Mora et al., 1980; Sfinchez et al., 1978; Vichido et al., 1978). Nevertheless, the involvement of the regulatory system responsible for nitrogen catabolite repression on the regulation of the ammonium ion assimilation enzymes has been scarcely studied (Dantz- ing et al., 1979; Dunn-Coleman et al., 1981; Premakumar et al., 1980). In N. crassa the nit-2 and nmr-1 ( m s - 5 ) loci represent the major control genes: the nit-2 gene is required for expression of genes involved with the utilization of secondary nitrogen sources and the product of the m s - 5 gene acts in a negative manner to repress the synthesis of nitrogen catabolite enzymes (Coddington, 1976; DeBusk and Ogilvie, 1984; Dunn-Coleman et al., 1981; Facklan and Marzluf, 1978; Fu and Marzluf, 1987, 1988; Marzluf, 1981; Premakumar et al., 1980; Reinert and Marzluf, 1975). A N. crassa mutant has also been isolated (glnr), whose arginine catabolism and GS are less sensitive to glutamine repression (Gonzfilez et al., 1983). In this work we have examined the effect of the nit-2 and m s - 5 mutations on the regulation of ammonium ion assimilation enzymes, and the allelism of the m s - 5 and gln ~ mutations in N. crassa was also determined.
In the wild-type strain, GS activity is repressed by ammonium ion, by glutamine, or when a poor carbon source is utilized as previously reported (Lara et al., 1982; Mora et al., 1980; Sfinchez et al., 1978; Vichido et al., 1978). In the nit-2 mutant, GS activity is severely reduced under all nitrogen conditions tested. The gel scanning of the GS immunoprecipitates of the wild-type and nit-2 strains indicate that the GS activity corresponds to the GS amount so the low GS activity found in the nit-2 strain is not the result of inhibition of the enzyme. The nit-2 strain has lower GDH activity than the wild-type strain, something that was more apparent on ammonium ion and glutamine. Dantzing et al. (1979) also found low levels of GDH activity in nit-2 strains in excess ammonium ion. The low GS and GDH activity in the nit-2 strain is reflected by the low glutamine content and the low growth rate observed under nitrogen-limited conditions such as glutamate and ammo- nium ion-limited cultures. These results indicate that the GS and GDH are regulated by the nit-2 product. The similar growth of the nit-2 and wild-type strains in excess ammonium ion indicates that although the nit-2 strain has a reduced capacity to assimilate ammonium ion, it is enough to provide the nitrogen necessary for growth; therefore in this strain, glutamate and

436 Calder6n and Martlnez
glutamine must be cycling at lower rates than the wild-type strain (Calder6n et al., 1989).
In the ms-5 strain the GS activity is not repressed by ammonium ion or glutamine. Our results indicate that GS is regulated by the ms-5 product. A similar low GS activity was found in the wild-type, nit-2, and ms-5 strains on glycerol as the carbon source, something that may be due to another regulatory mechanism. The ms-5 strain has higher levels of GDH activity than the wild-type strain under all tested conditions, except on glutamate. The similar levels of GDH on glutamate in the ms-5 strain could be explained by the high glutamate pool in this strain that may repress the GDH (Hernfindez et al., 1983). The high GDH and GS activities in the ms-5
strain is reflected by the high glutamine content. These results indicate that GDH is regulated by the ms-5 locus, by the carbon source, and by another mechanism that may sense the amino nitrogen content. The slow growth rate of the ms-5 strain could be explained by the high expenditure of ATP due to increased glutamine synthesis and its cycling (Calderdn et al., 1985, 1989, 1990; Calder6n and Mora, 1985, 1989; Mora et al., 1988).
GOGAT activity is not repressed in the nit-2 strain, where it was found to be higher. These results indicate that GOGAT activity is not regulated by the product of the nit-2 gene. The high GOGAT activity in the nit-2 may compensate for the reduced synthesis of glutamate by GDH. In the ms-5
strain the increased GOGAT activity may compensate higher glutamate utilization for glutamine synthesis, except in ammonium ion-limited cul- tures, where ammonium ion limitation may restrict glutamine synthesis. These results are in accordance with the proposal that GOGAT functions to adjust the glutamate/glutamine ratio (Calder6n et al., 1990; Calder6n and Mora, 1985; Lomnitz et al., 1987).
The gln r strain behaves similarly to the ms-5 strain as shown by the lower GS repression by glutamine, the resistance to L-methionine sulfoximine, and the accumulation of amino acids; however, the ms-5 strain does not induce nitrate reductase on NH4NO3. Furthermore, the result of the cross between these two strains showed segregation at a high frequency, indicating that these mutations are not allelic. These results indicate the complex regula- tion of the GS that is regulated by the nit-2, ms-5, and gln r loci.
The results presented here show that the N. crassa GS and GDH encoding genes are coordinately transcriptionally regulated together with the genes encoding proteins capable of catabolizing secondary nitrogen sources by the regulatory proteins encoded by the nit-2 and nmr-1 genes. This kind of regulation may ensure optimal ammonium ion assimilation under conditions of nitrogen limitation.

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 437
ACKNOWLEDGMENTS
We are grateful to Jaime Mora for his encouragement and discussion throughout this work, to Yolanda Mora and Isabel P6rez Montfort for reviewing the manuscript, to Gisela Du Pont and Leticia Olvera for techni- cal assistance, to Dalia Terrones for amino acid determinations, to Esther Wolburg for preparation of the manuscript, and to David Link for computer support.
REFERENCES
Brett, M., Chambers, G. K., Holder, A. A., Fincham, J. R. S., and Wootton, J. C. (1976). Mutational amino acid replacements in Neurospora crassa NADP-specific glutamate dehydrogenase. J. MoL Biol. 106:1.
Calder6n, J., and Mora, J. (1985). Glutamine cycling in Neurospora crassa. J. Gen. Microbiol. 131:3237.
Calder6n, J., and Mora, J. (1989). Glutamine assimilation pathways in Neurospora crassa growing on glutamine as sole nitrogen and carbon source. J. Gen. Microbiol. 135:2699.
Calder6n, J., Morett, E., and Mora, J. (1985). c0-Amidase pathway in the degradation of glutamine in Neurospora crassa. Z Bacteriol. 161:807.
Calder6n, J., Cooper, A. J. L., Gelbard, A. S., and Mora, J. (1989). I3N Isotope studies of glutamine assimilation pathways in Neurospora crassa. J. Bacteriol. 171:1772.
Calder6n, J., Martinez, L. M., and Mora, J. (1990). Isolation and Characterization of a Neurospora crassa mutant altered in the e~ polypeptide of glutamine synthetase. J. BacterioI. 172:4996.
Coddington, A. (1976). Biochemical studies on the nit mutants ofNeurospora crassa. Mol. Gen. Genet. 145:195.
Cove, D. J. (1976). Chlorate toxicity inAspergillus nidulans studies of mutants altered in nitrate assimilation. Mol. Gen. Genet. 146:147.
Cove, D. J. (1979). Genetic studies of nitrate assimilation in Aspergillus nidulans. Biol. Rev. 54:291.
Dantzing, A. A., Weighman, F. L., Jr., and Mason, A. (1979). Regulation of glutamate dehydrogenase in nit-2 and am mutants of Neurospora crassa. J. Bacteriol. 137:1333.
D~ivilla, G., S~inchez, F., Palacios, R., and Mora, J. (1978). Genetics and physiology of Neurospora crassa glutamine auxotrophs. J. BacterioL 134:693.
D~vila, G., Lara, M., Guzm~n, J., and Mora, J. (1980). Relation between structure and function of Neurospora crassa glutamine synthetase. Biochem. Biophys. Res. Commun. 92:134.
D~ivila, G., Brom, S., Mora, Y, Palacios, R., and Mora, J. (1983). Genetic and biochemical characterization of glutamine synthetase from Neurospora crassa auxotrophs and their revertants. J. Bacteriol. 156:993.
Davis, R. H., and De Serres, F. J. (1970). Genetic and microbiological research technique for Neurospora crassa. Methods Enzymol. 17:79.
DeBusk, R. M., and Ogilvie, S. (1984). Regulation of amino acid utilization in Neurospora crassa: effect of nmr-1 and ms-5 mutations. J. Bacteriol. 160:656.
Dunn-Coleman, N. S., Tomsett, A. B., and Garrett, R. H. (1981). The regulation of nitrate assimilation in Neurospora crassa: Biochemical analysis of the nmr-1 mutants. Mol. Gen. Genet. 182:234.
Facklan, T. J., and Marzluf, G. A. (1978). Nitrogen regulation of amino acid catabolism in Neurospora crassa. Biochem. Genet. 16:343.
Ferguson, A. R., and Sims, A. P. (1974). The regulation of glutamine metabolism in Candida utilis: The role of glutamine in the control of glutamine synthetase. J. Gen. Microbiol. 80:159.

438 Caiderdn and Martlnez
Fincham, J. R. S. (1950). Mutant strains on Neurospora deficient in aminating ability. J. Biol. Chem. 181:61.
Fincham, J. R. S. (1957). A modified glutamic acid dehydrogenase as a result of a gene mutation in Neurospora crassa. Biochem. J. 65:721.
Fincham, J. R. S., and Baron, A. J. (197-7). The molecular basis of an osmotically reparable mutant of Neurospora crassa producing unstable glutamate dehydrogenase. J. Mol. Biol. 110:627.
Fu, Y. H., and Marzluf, G. A. (1987). Characterization of nit-2, the major regulatory gene of Neurospora crassa. Mol. Cell. Biol. 7:1691.
Fu, Y. H., and Marzluf, G. A. (1988). Metabolic control and autogenous regulation of nit-3, the nitrate reductase structural gene ofNeurospora crassa. J. Bacteriol. 170:657.
Fu, Y. H., and Marzluf, G. A. (1990a). The major nitrogen regulatory gene ofNeurospora crassa encodes a protein with putative zinc finger DNA-binding domain. MoL Cell. BioL 10:1056.
Fu, Y. H., and Marzluf, G. A. (1990b). nit-2, the major positive acting nitrogen regulatory gene of Neurospora crassa, encode a sequence-specific DNA-binding protein. Proc. Natl. Acad. Sci. USA 87:5331.
Fu, Y. H., Young, J. L., and Marzluf, G, A. (1988). Molecular cloning and characterization of a negative-acting nitrogen regulatory gene of Neurospora crassa. Mol. Gen. Genet. 214:74.
Gonzfilez, A., Tenorio, M., Vaca, G., and Mora, J. (1983). Neurospora crassa mutant impaired in glutamine regulation. J. BacterioL 155:1.
Hern~indez, G., S~inchez-Pescador, R., Palacios, R., and Mora, J. (1983). Nitrogen source regulates glutamate dehydrogenase NADP synthesis in Neurospora crassa. J. Bacteriol. 154:524.
Holder, A. A., Wooton, J. C., Baron, A. J., Chambers, G. K., and Fincham, J. R. S. (1975). The amino acid sequence of Neurospora crassa NADP-specific glutamate dehydrogenase. Biochem. J. 149:757.
Hummelt, G., and Mora, J. (1980a). NADH-dependent glutamate synthase and nitrogen metabolism in Neurospora crassa. Biochem. Biophys. Res. Commun. 92:127.
Hummelt, G., and Mora, J. (1980b). Regulation and function of glutamate synthase. Biochem. Biophys. Res. Commun. 96:168.
Jarai, G., and Marzluf, G. A. (1990). Analysis of conventional and in vitro generated mutants of nmr, the negatively acting nitrogen regulatory gene of Neurospora crassa. MoL Gen. Genet. 222:233.
Kinnaird, J. H., Keighren, M. A., Kinsey, J. A., Eaton, M., and Fincham, J. R. S. (1982). Cloning of the am glutamate dehydrogenase gene of Neurospora crassa through the use of synthetic DNA probe. Gene 20:387.
Lara, M., Blanco, L., Campomanes, M., Calva, E., Palacios, R., and Mora, J. (1982). Physiology of ammonium assimilation in Neurospora crassa. J. Bacteriol. 150:105.
Limon-Lason, J., Lara, M., Resendiz, B., and Mora, J. (1977). Regulation of glutamine synthetase in fed-batch cultures of Neurospora crassa. Biochem. Biophys. Res. Commun. 78:1234.
Lomnitz, A., Calder6n, J., Hern~indez, G., and Mora, J. (1987). Functional analysis of ammonium assimilation enzymes in Neurospora crassa. J. Gen. Microbiol. 133:2333.
Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. J. BioL Chem. 193:265.
Marzluf, G. A. (1981). Regulation of nitrogen metabolism and gene expression in fungi. Microbiol. Rev. 45:437.
Mora, J., Calder6n, J., and Hern~ndez, G. (1988). Search, assimilation, and turnover in some fungi, In Sgmchez-Esquivel (ed.), Nitrogen Control of Metabolic Processes, CRC Press, Boca Raton, Fla., pp. 58-81.
Mora, Y., Chfivez, O., and Mora, J. (1980). Regulation of Neurospora crassa glutamine synthetase by carbon and nitrogen source. J. Gen. MicrobioL 118:455.
Palacios, R. (1976). Neurospora crassa glutamine synthetase: Purification by affinity chromatog- raphy and characterization of subunit structure. J. Biol. Chem. 251:4787.
Palacios, R., Campomanes, M., and Quinto, C. (1977). Neurospora crassa glutamine synthetase

Regulation of GS, GDH, and GOGAT in Neurospora crassa nit-2 and ms-5 Mutant Strains 439
translation of specific messenger ribonucleic acid in a cell-flee system derived from reticulocytes. J. Biol. Chem. 252:3028.
Premakumar, R., Sorger, G. J., and Goodman, D. (1980). Physiological characterization of a Neurospora crassa mutant with impaired regulation of nitrate reductase. J. Bacteriol. 144:542.
Reinert, W. R., and Marzluf, G. A. (1975). Genetic and metabolic control of the purine catabolic enzymes of Neurospora crassa. 3/Iol. Gen. Genet. 139:39.
Ronzio, R. A., and Meister, A. (1968). Phosphorylation of methionine sulfoximine by glutamine synthetase. Proc. Natl. Acad. Sci. USA 59:164.
S~inchez, F., Campomanes, M., Quinto, C., Hansberg, W., Mora, J., and Palacios, R. (1978). Nitrogen sources regulate glutamine synthetase mRNA levels in Neurospora crassa. J. Bacteriol. 136:880.
Sgnchez, F., Calva, E., Campomanes, M., Blanco, L., Guzm~in, J., Saborio, J. L., and Palacios, R. (1980). Heterogeneity of glutamine synthetase polypeptides in Neurospora crassa. Z Biol. Chem. 255:2231.
Seale, T. W., Brett, M., Baron, A. J., and Fincham, J. R. S, (1977). Amino acid replacements resulting from suppression of missense of a chain terminator mutation in Neurospora. Genetics 86:261.
Sidding, M. A. M., Kinsey, J. A., Fincham, J. R. S., and Keighren, M. (1980). Frameshift mutations affecting the N-terminal sequence of Neurospora NADP-specific glutamate dehydrogenase. Z Mol. Biol. 137:125.
Sorger, G. J., Debanne, M. T., and Davies, J. (1974). Effect of nitrate on the synthesis and decay of nitrate reductase in Neurospora. Bh)chem. Z 140:395.
Stewart, U., and Vollmer, S. J. (1986). Molecular cloning of nit-2, a regulatory gene required for nitrogen metabolite repression in Neurospora crassa. Gene 46:291.
Vaca, G., and Mora, J. (1977). Nitrogen regulation of arginase in Neurospora crassa. J. Bactetiol. 131:719.
Van Pilsum, J. F., Martin, R. P., Kito, E., and Hess, J. (1956). Determination of creatine, creatinine, arginine, guanido-acetic acid, guanidine and methyl-guanidine in biological fluids. J. Biol. Chem. 222:225.
Vichido, I., Mora, Y., Quinto, C., Palacios, R., and Mora, J. (1978). Nitrogen regulation of glutamine synthetase in Neurospora crassa. J. Gen. Microbiol. 106:251.
Vogel, H. J. (1964). Distribution of lysine pathways among fungi: evolutionary implications. Am. Nat. 98:435.
Westergaard, M., and Mitchell, H. K. (1947). Neurospora V.A. synthetic medium favoring sexual production. Am. J. Bot. 34:573.




















