
Amphid defective mutant of Caenorhabditis elegans
8
Genetica 94: 195-202, 1994. 195 @ 1994 KluwerAcademic Publishers. Printedin the Netherlands. Amphid defective mutant of Caenorhabditis elegans Laura De Riso, Filomena Ristoratore, Marisa Sebastiano & Paolo Bazzic~alupo* Istituto InternazionaIe di Genetica e Bio~sica. CNR. Via G. Marconi 10, 80125, Napoli, Italy *Author for correspondence Received and accepted 17 May 1994 Abstract Studies are reported on a chemoreception mutant which arose in a mutator strain. The mutant sensory neurons do not stain with fluoresceine isothiocyanate (Dyf phenotype), hence the name, dyf-1, given to the gene it identifies. The gene maps on LGI, 0.4 map units from dpy-5 on the unc-ll side. The response of mutant worms to various repellents has been studied and shown to be partially altered. Other chemoreception based behaviors are less affected. The cilia of the sensory neurons of the amphid are shorter than normal and the primary defect may be in the capacity of the sheath cells to secrete the matrix material that fills the space between cilia in the amphid channel. Progress toward the molecular cloning of the gene is also reported. Relevant results from other laboratories are briefly reviewed. Chemoreception and the nervous system The nervous system of Caenorhabditis elegans has been completely described at the cellular and sub- cellular level (White et al., 1986). It is made of a limited number of neurons whose development and wiring is described in greater detail than in any other organism. The genetic and molecular analysis of sensitivity to light touch has led in Caenorhabditis elegans to the identification of several genes (mec genes) involved in determining the structures underlying this behavior and has already given important information on the role of genetic information in specifying organization and function of a simple sensory system (Chalfie & White, 1988; Chalfie & Au, 1989). Chemoreception is another sensory behavior which can be analysed genet- ically in C. elegans and can offer the opportunity of correlating the results of the genetic analysis with a wealth of information at the anatomical, developmen- tal and physiological level. Many aspects of C. ele- gans behavior are influenced by chemicals present in the environment. Ward (1973) has studied chemotaxis, identifying a number of water soluble attractants, and with the use of mutants has been able to establish that orientation along a gradient of attractant is achieved through klinotaxis. Mutants failing to be attracted, designated Che for chemotaxis defective, have been isolated and the ultra structural analysis of their senso- ry organs has confirmed that the two amphids are the main chemosensory organs in C. elegans (Dusenbery, Sheridan & Russell, 1975; Lewis & Hodgkin, 1977). There are also noxious chemicals and unfavourable environmental conditions that act as repellent on C. elegans and from which worms move away. Culotti and Russell (1978) have studied the avoidance reac- tion of worms to high osmotic strength in the medium and have isolated mutants unable to sense it, called Osm, for osmotic avoidance defective. Chemical signals in the environment are also important in determining the pathway of larval devel- opment. When food is present and when worms are not too crowded, development proceeds normally from larval stage L2 to L3 and L4. When food is scarce and/or worms are crowded, L2 worms moult instead to dauers. These have a more resistant cuticle, do not feed and have specific behavior that confers to them a role of resistance and dispersal. When food is available, dauers start to feed, moult to L4, and normal devel- opment resumes. The ability to sense food and crowd- ing is used to determine entry into and exit from the dauer developmental pathway. Mutants unable to enter this pathway (dauer defective) and mutants entering it in the presence of abundant food (dauer constitutive) have been isolated and placed in epistatic pathway(s)
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Amphid defective mutant of Caenorhabditis elegans

Genetica 94: 195-202, 1994. 195 @ 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Amphid defective mutant of Caenorhabditis elegans
L a u r a D e Riso , F i l o m e n a Ri s to ra to re , M a r i s a S e b a s t i a n o & P a o l o Bazzic~alupo* Istituto InternazionaIe di Genetica e Bio~sica. CNR. Via G. Marconi 10, 80125, Napoli, Italy *Author for correspondence
Received and accepted 17 May 1994
Abstract
Studies are reported on a chemoreception mutant which arose in a mutator strain. The mutant sensory neurons do not stain with fluoresceine isothiocyanate (Dyf phenotype), hence the name, dyf-1, given to the gene it identifies. The gene maps on LGI, 0.4 map units from dpy-5 on the unc-l l side. The response of mutant worms to various repellents has been studied and shown to be partially altered. Other chemoreception based behaviors are less affected. The cilia of the sensory neurons of the amphid are shorter than normal and the primary defect may be in the capacity of the sheath cells to secrete the matrix material that fills the space between cilia in the amphid channel. Progress toward the molecular cloning of the gene is also reported. Relevant results from other laboratories are briefly reviewed.
Chemoreception and the nervous system
The nervous system of Caenorhabditis elegans has been completely described at the cellular and sub- cellular level (White et al., 1986). It is made of a limited number of neurons whose development and wiring is described in greater detail than in any other organism. The genetic and molecular analysis of sensitivity to light touch has led in Caenorhabditis elegans to the identification of several genes (mec genes) involved in determining the structures underlying this behavior and has already given important information on the role of genetic information in specifying organization and function of a simple sensory system (Chalfie & White, 1988; Chalfie & Au, 1989). Chemoreception is another sensory behavior which can be analysed genet- ically in C. elegans and can offer the opportunity of correlating the results of the genetic analysis with a wealth of information at the anatomical, developmen- tal and physiological level. Many aspects of C. ele- gans behavior are influenced by chemicals present in the environment. Ward (1973) has studied chemotaxis, identifying a number of water soluble attractants, and with the use of mutants has been able to establish that orientation along a gradient of attractant is achieved through klinotaxis. Mutants failing to be attracted, designated Che for chemotaxis defective, have been
isolated and the ultra structural analysis of their senso- ry organs has confirmed that the two amphids are the main chemosensory organs in C. elegans (Dusenbery, Sheridan & Russell, 1975; Lewis & Hodgkin, 1977). There are also noxious chemicals and unfavourable environmental conditions that act as repellent on C. elegans and from which worms move away. Culotti and Russell (1978) have studied the avoidance reac- tion of worms to high osmotic strength in the medium and have isolated mutants unable to sense it, called Osm, for osmotic avoidance defective.
Chemical signals in the environment are also important in determining the pathway of larval devel- opment. When food is present and when worms are not too crowded, development proceeds normally from larval stage L2 to L3 and L4. When food is scarce and/or worms are crowded, L2 worms moult instead to dauers. These have a more resistant cuticle, do not feed and have specific behavior that confers to them a role of resistance and dispersal. When food is available, dauers start to feed, moult to L4, and normal devel- opment resumes. The ability to sense food and crowd- ing is used to determine entry into and exit from the dauer developmental pathway. Mutants unable to enter this pathway (dauer defective) and mutants entering it in the presence of abundant food (dauer constitutive) have been isolated and placed in epistatic pathway(s)

196
(Riddle, 1988; Vowels & Thomas, 1992). A subset of these mutants has been shown to bear defects in the sen- sory cilia of the amphids, suggesting that the amphid neurons sense the relevant signals (for a review see Riddle, 1988).
The variety of effects that chemicals in the envi- ronment have on C. elegans indicates that chemore- ception is more complex than the response to light touch. This greater complexity is already apparent at the level of sensory neurons: in C. elegans there are six light touch receptor cells belonging to three dif- ferent classes, whereas chemical sensitivity uses 30 or more different chemoreception neurons, belonging to 15 different classes (White et al., 1986; Chalfie & White, 1988).
The cell-lineage and anatomy of chemoreception organs have been described (Ward et al., 1975; Sulston et aI., 1993; Perkins et al., 1986). Most of the connec- tions of the sensory neurons of these organs have also been described (White et al., 1986). Laser ablation experiments have been used to identify the function of single sensory cells of the amphids in the detec- tion of the stimuli for dauer formation, chemotaxis, avoidance, attraction to volatile odorants (Bargmann, Thomas & Horvitz, 1990; Bargmann & Horvitz, 1991a, b; Bargmann, Hartweig & Horvitz, 1993; Kaplan & Horvitz, 1993). Some general features of the organization of the amphid that emerge from these studies are:
a) Each sensory neuron of the amphids usually bears receptors for more than one stimulus; one neu- ron, ASH, has been shown to sense high osmotic strength but also to respond to mechanical stimuli (Kaplan & Horvitz, 1993);
b) Receptors for a given stimulus are usually present on more than one but not on all sensory neurons. They are not equally distributed on these neurons and thus the roles of the various neurons in the response to a given stimulus are quantitatively and probably qualitatively different from each other (Bargmann, Thomas & Horvitz, 1990; Bargmann & Horvitz, 1991a, b).
Dyf mutants
In 1986 Perkins et al. showed that eight pairs of chemosensory neurons of live wild type animals take up Fluoresceine isothiocyanate (FITC) when it is present in the medium. The dye enters the neurons through their exposed endings; within minutes pro- cesses and cell bodies become fluorescent, stay that way for several hours and can be recognized easily with a fluorescence microscope. The cells that stain are six pairs of amphid neurons in the head and the two pairs of phasmid neurons in the tail. Since then, other lipophylic dyes have been used with similar results. Mutants have been found in which the staining of amphid neurons is lacking or altered (Fig. 1). This class of mutants has been named Dyf, for dye filling alterations.
The Dyf phenotype, unlike behavioral phenotypes, can be easily detected on single live worms. This makes genetic analysis far easier and several groups, interest- ed in chemoreception, have focused their attention on genes which can mutate to a Dyf phenotype. Perkins et aI. (1986), as well as researchers in other laboratories, have addressed the question of the overlap between the Dyf phenotype and defects in chemoreception. Mutants in most of the genes identified on the basis of behavioral alteration also have a Dyf phenotype (Perkins et al., 1986). In addition, new Dyf mutants have been isolated following various protocols which usually included an enriching step such as collecting worms unable to avi~id high osmotic strength or dauer defective revertants of dauer constitutive mutants (R. Herman & D. Riddle, personal communication). In our laboratory we screened directly for lack of staining, and chose only mutants in which none of the amphid cells, that normally do, would stain. All our Dyf mutants show some chemoreception defect, especially in the avoidance response to repellents, but are normal for sensitivity to light touch (Bazzicalupo et al., 1994). Dyf alleles, all showing some chemoreception defect, have so far been identified in more than 20 genes with about half of them falling into genes previously iden- tified on the basis of altered behavior: osm, che, and daf .
c) The amphid neurons involved in sensing water sol- uble chemicals are distinct from those that detect volatile odorants (Bargmann, Hartweig & Horvitz, 1993). Thus, also in C. elegans, smell and taste are distinct.
dyf-1 (mn335) We have studied in more detail one Dyf mutant, mn335, which was isolated by Todd Starich in R. Herman's laboratory at the University of Minnesota. It arose as a spontaneous mutant in a mutator strain. The mutation,
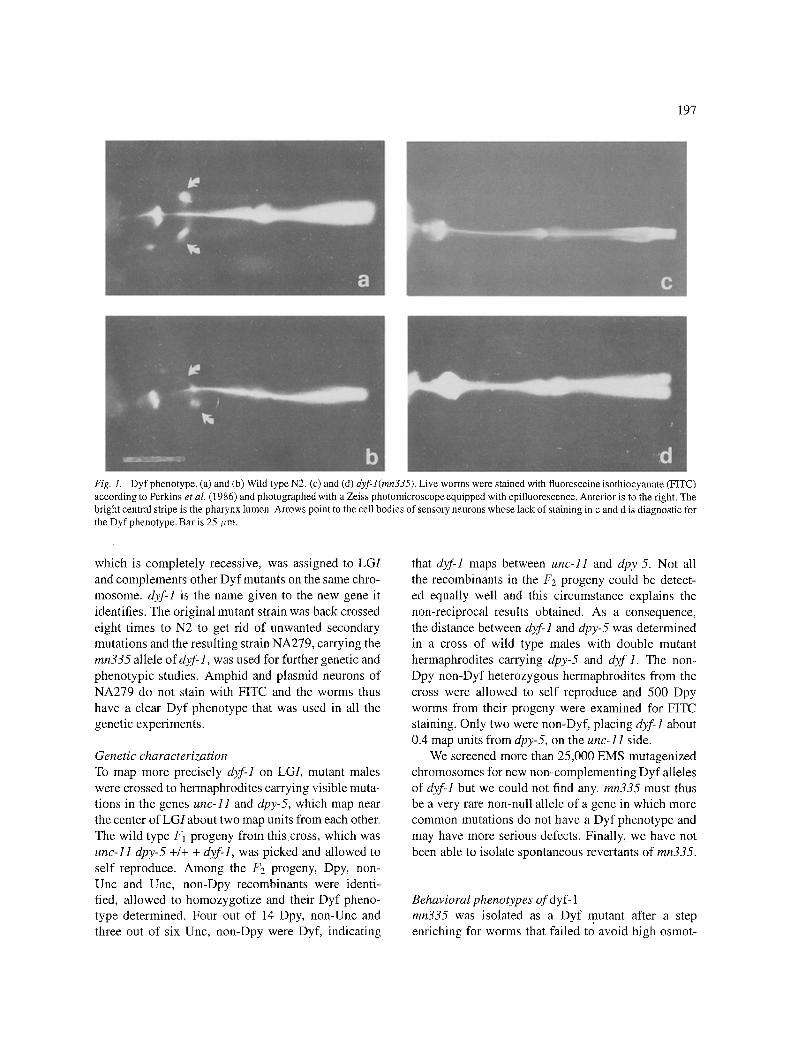
197
Fig. 1. Dyf phenotype. (a) and (b) Wild type N2. (c) and (d) d2f-l(mn335). Live worms were stained with fluoresceine isothiocyanate (FITC) according to Perkins et al. (1986) and photographed with a Zeiss photomicroscope equipped with epifluorescence. Anterior is to the right. The bright central stripe is the pharynx lumen. Arrows point to the cell bodies of sensory neurons whose lack of staining in c and d is diagnostic for the Dyf phenotype. Bar is 25 ~m.
which is completely recessive, was assigned to LGI and complements other Dyf mutants on the same chro- mosome, dyf-1 is the name given to the new gene it identifies. The original mutant strain was back crossed eight times to N2 to get rid of unwanted secondary mutations and the resulting strain NA279, carrying the mn335 allele of dyf-1, was used for further genetic and phenotypic studies. Amphid and plasmid neurons of NA279 do not stain with FITC and the worms thus have a clear Dyf phenotype that was used in all the genetic experiments.
Genetic characterization To map more precisely dyf-1 on LGI, mutant males were crossed to hermaphrodites carrying visible muta- tions in the genes unc-ll and dpy-5, which map near the center of L G I about two map units from each other. The wild type F1 progeny from this cross, which was unc-ll dpy-5 +/+ + dyf-I, was picked and allowed to self reproduce. Among the F2 progeny, Dpy, non- Uric and Uric, non-Dpy recombinants were identi- fied, al lowed to homozygotize and their Dyf pheno- type determined. Four out of 14 Dpy, non-Uric and three out of six Unc, non-Dpy were Dyf, indicat ing
that dyf-1 maps between unc-ll and dpy-5. Not all the recombinants in the F2 progeny could be detect- ed equally well and this circumstance explains the non-reciprocal results obtained. As a consequence, the distance between dyf-1 and dpy-5 was determined in a cross of wild type males with double mutant hermaphrodites carrying dpy-5 and dyf-1. The non- Dpy non-Dyf heterozygous hermaphrodites from the cross were allowed to self reproduce and 500 Dpy worms from their progeny were examined for FITC staining. Only two were non-Dyf, placing dyf-1 about 0.4 map units from dpy-5, on the unc-ll side.
We screened more than 25,000 EMS mutagenized chromosomes for new non-complementing Dyf alleles of dyf-1 but we could not find any. mn335 must thus be a very rare non-null allele of a gene in which more common mutations do not have a Dyf phenotype and may have more serious defects. Finally, we have not been able to isolate spontaneous revertants of mn335.
Behavioral phenotypes of dyf- 1 mn335 was isolated as a Dyf mutant after a step enriching for worms that failed to avoid high osmot-
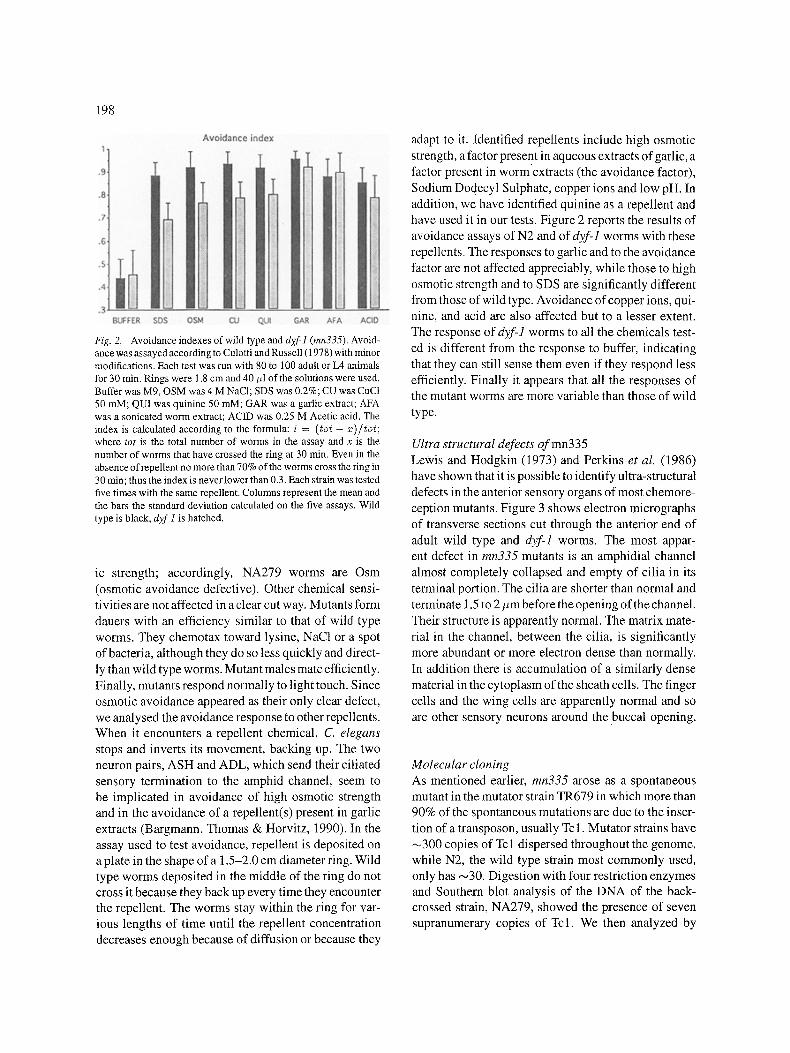
198
Fig,. 2. Avoidance indexes 0fwild type and d;~/"l (mn335). Avoid- ance was assayed according to Culotti and Russell (1978) with minor modifications. Each test was run with 80 to 100 adult or L4 animals for 30 rain. Rings were 1.8 cm and 40 >1 of the solutions were used. Buffer was M9, OSM was 4 M NaCI; SDS was 0.2%; CU was CuCI 50 raM; QUI was quinine 50 raM; GAR was a garlic extract; AFA was a sonicated worm extract; ACID was 0.25 M Acetic acid. The index is calculated according to the formula: i = (tot - x)/toL where tot is the total number of worms in the assay and x is the number of worms that have crossed the ring at 30 min. Even in the absence of repellent no more than 70% of the worms cross the ring in 30 min; thus the index is never lower than 0.3. Each strain was tested five times with the same repellent. Columns represent the mean and the bars the standard deviation calculated on the five assays. Wild type is black, dyfl is hatched.
ic strength; accordingly, NA279 worms are Osm (osmotic avoidance defective). Other chemical sensi-
tivities are not affected in a clear cut way. Mutants form dauers with an efficiency similar to that of wild type worms. They chemotax toward lysine, NaC1 or a spot of bacteria, although they do so less quickly and direct-
ly than wild type worms. Mutant males mate efficiently. Finally, mutants respond normally to light touch. Since osmotic avoidance appeared as their only clear defect,
we analysed the avoidance response to other repellents. When it encounters a repellent chemical, C. elegans stops and inverts its movement, backing up. The two neuron pairs, ASH and ADL, which send their ciliated
sensory termination to the amphid channel, seem to be implicated in avoidance of high osmotic strength and in the avoidance of a repellent(s) present in garlic
extracts (Bargmann, Thomas & Horvitz, 1990). In the assay used to test avoidance, repellent is deposited on a plate in the shape of a 1.5-2.0 cm diameter ring. Wild type worms deposited in the middle of the ring do not cross it because they back up every time they encounter the repellent. The worms stay within the ring for var- ious lengths of time until the repellent concentration decreases enough because of diffusion or because they
adapt to it. Identified repellents include high osmotic
strength, a factor present in aqueous extracts of garlic, a factor present in worm extracts (the avoidance factor),
Sodium Dodecyl Sulphate, copper ions and low pH. In addition, we have identified quinine as a repellent and have used it in our tests. Figure 2 reports the results of avoidance assays of N2 and of dyf-1 worms with these repellents. The responses to garlic and to the avoidance
factor are not affected appreciably, while those to high osmotic strength and to SDS are significantly different from those of wild type. Avoidance of copper ions, qui- nine, and acid are also affected but to a lesser extent. The response of dyf-1 worms to all the chemicals test- ed is different from the response to buffer, indicating
that they can still sense them even if they respond less efficiently. Finally it appears that all the responses of the mutant worms are more variable than those of wild type.
Ultra structural defects ofmn335 Lewis and Hodgkin (1973) and Perkins et al. (1986) have shown that it is possible to identify ultra-structural defects in the anterior sensory organs of most chemore-
ception mutants. Figure 3 shows electron micrographs of transverse sections cut through the anterior end of
adult wild type and dyf-1 worms. The most appar- ent defect in mn335 mutants is an amphidial channel almost completely collapsed and empty of cilia in its
terminal portion. The cilia are shorter than normal and terminate 1.5 to 2 #m before the opening of the channel.
Their structure is apparently normal. The matrix mate- rial in the channel, between the cilia, is significantly more abundant or more electron dense than normally.
In addition there is accumulation of a similarly dense material in the cytoplasm of the sheath cells. The finger cells and the wing cells are apparently normal and so are other sensory neurons around the buccal opening.
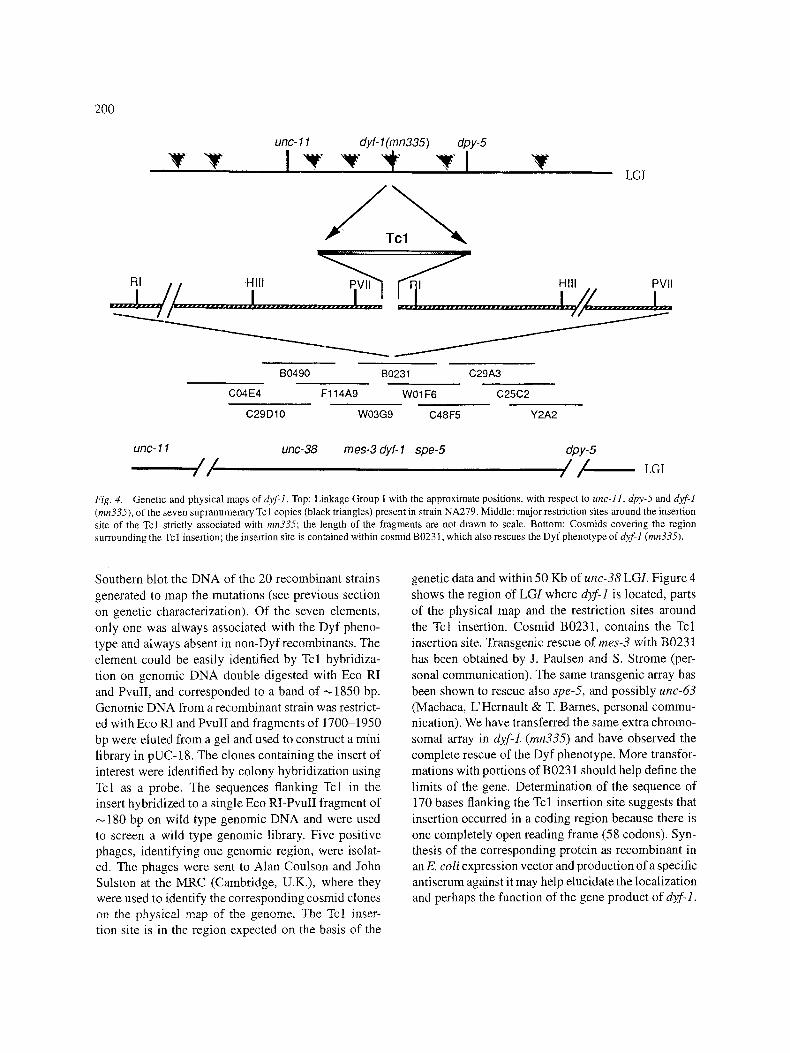
Molecular cloning As mentioned earlier, mn335 arose as a spontaneous mutant in the mutator strain TR679 in which more than 90% of the spontaneous mutations are due to the inser- tion of a transposon, usually Tcl. Mutator strains have ~300 copies of Tcl dispersed throughout the genome, while N2, the wild type strain most commonly used, only has ,-,30. Digestion with four restriction enzymes and Southern blot analysis of the DNA of the back- crossed strain, NA279, showed the presence of seven supranumerary copies of Tcl. We then analyzed by
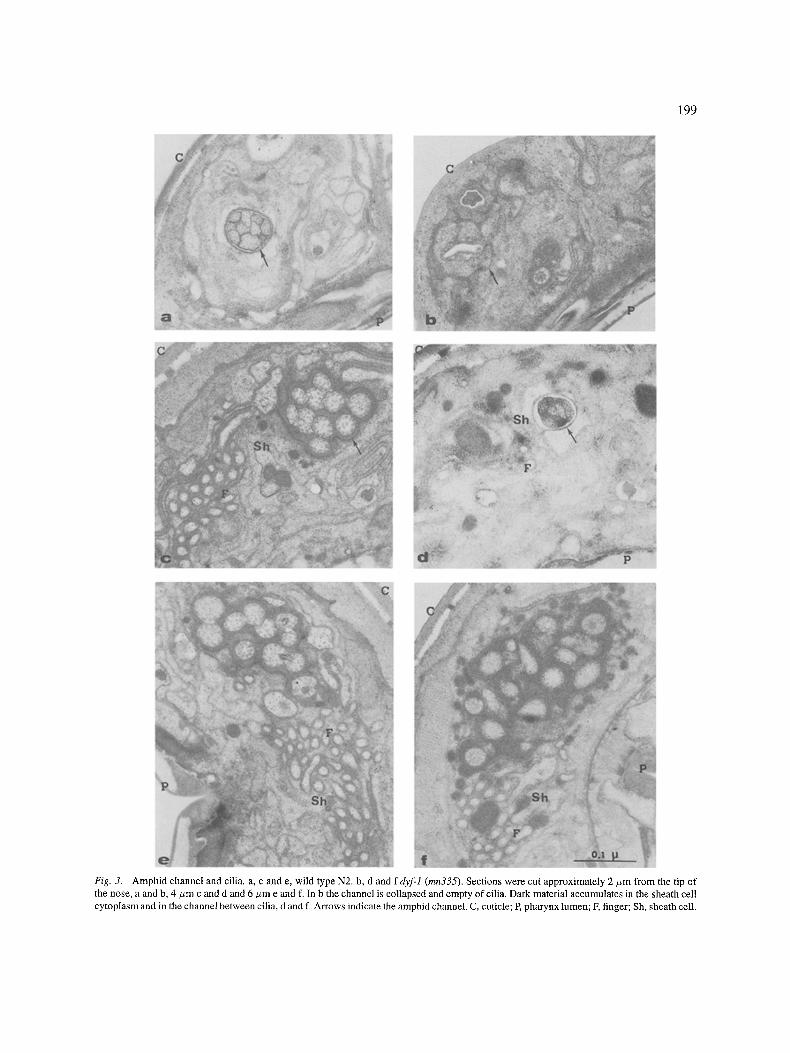
199
Fig. 3. Amphid channel and cilia, a, c and e, wild type N2. b, d and f dyfl (mn335). Sections were cut approximately 2/zm from the tip of the nose, a and b, 4 #m c and d and 6/ ,m e and f. In b the channel is collapsed and empty of cilia. Dark material accumulates in the sheath cell cytoplasm and in the channel between cilia, d and f. Arrows indicate the amphid channel C, cuticle; E pharynx lumen; F, finger; Sh, sheath cell.
200
unc-11 dyf-1(mn335) dpy-5
I
RI , , H i l l
I / I I
Y LGI
Hill PVll
J .............................. ] _ / / ............ L
B0490
C04E4
B0231 C29A3
F114A9 W01 F6 C25C2
C29 D 10 W03G9 C48F5 Y2A2
unc- 11 unc-38 mes-3 dyf- 1 spe-5 dpy-5 / / , , / / LGI
Fig. 4. Genetic and physical maps of dyt:l. Top: Linkage Group I with the approximate positions, with respect to unc-I1, dpy-5 and d2f-1 (mn335), of the seven supranumerary Tcl copies (black triangles) present in strain NA279. Middle: major restriction sites around the insertion site of the Tcl strictly associated with mn335; the length of the fragments are not drawn to scale. Bottom: Cosmids covering the region surrounding the Tc 1 insertion; the insertion site is contained within cosmid B0231, which also rescues the Dyf phenotype of dzf-1 (mn335).
Southern blot the DNA of the 20 recombinant strains generated to map the mutations (see previous section on genetic characterization). Of the seven elements, only one was always associated with the Dyf pheno- type and atways absent in non-Dyfrecombinants. The element could be easily identified by Tcl hybridiza- tion on genomic DNA double digested with Eco RI and PvuII, and corresponded to a band of ,-~1850 bp. Genomic DNA from a recombinant strain was restrict- ed with Eco RI and PvuII and fragments of 1700-1950 bp were eluted from a gel and used to construct a mini library in pUC-18. The clones containing the insert of interest were identified by colony hybridization using Tcl as a probe. The sequences flanking Tcl in the insert hybridized to a single Eco RI-PvuII fragment of
180 bp on wild type genomic DNA and were used to screen a wild type genomic library. Five positive phages, identifying one genomic region, were isolat- ed. The phages were sent to Alan Coulson and John Sulston at the MRC (Cambridge, U.K.), where they were used to identify the corresponding cosmid clones on the physical map of the genome. The Tcl inser- tion site is in the region expected on the basis of the
genetic data and within 50 Kb of unc-38 LGI. Figure 4 shows the region of LGI where d}f-1 is located, parts of the physical map and the restriction sites around the Tcl insertion. Cosmid B0231, contains the Tcl insertion site. Transgenic rescue of rues-3 with B0231 has been obtained by J. Paulsen and S. Strome (per- sonal communication). The same transgenic array has been shown to rescue also spe-5, and possibly uric-63 (Machaca, L'Hernault & T. Barnes, personal commu- nication). We have transferred the same extra chromo- somal array in dyf-1. (mn335) and have observed the complete rescue of the Dyf phenotype. More transfor- mations with portions of B0231 should help define the limits of the gene. Determination of the sequence of 170 bases flanking the Tcl insertion site suggests that insertion occurred in a coding region because there is one completely open reading frame (58 codons). Syn- thesis of the corresponding protein as recombinant in an E. coli expression vector and production of a specific antiserum against it may help elucidate the localization and perhaps the function of the gene product of dyf-].

201
Discussion and conclusions
The behavioral alterations of Dyf mutants are pleiotropic with sensitivity to many different chemi- cals usually decreased at the same time. A few Dyf mutants also show mechano-sensory defects but more often they have a normal response to light touch, indi- cating that they usually identify functions that are spe- cific to the chemosensory organs and are not general mutants of the nervous system. These characteristics are shared by mn335. This description of Dyf mutants applies in fact also to the majority of chemoreception mutants isolated so far on the basis of altered behav- ior, chemotaxis (Che), osmotic avoidance (Osm) and those dauer formation (Daf) mutants which alter the reception of the stimuli that trigger entry and exit from the dauer pathway. All these mutants, although isolat- ed because of a single altered behavior, are pleiotropic and are defective in other chemical sensitivities as well. Most of these mutants also alter FITC uptake, resulting in not stained or poorly stained Dyf animals.
Ultra-structural defects of the sensory cilia are com- mon to all the Dyf, Che and Osm mutants that have been analysed at the electron microscope level. Often all sensory cilia are altered; in some mutants, and mn335 appears to be one of them, only amphid and phasmid cilia are defective. Cilia are often shorter and variously disorganized and, as in mn335, the amphidi- al channel is partly collapsed and the matrix material secreted by the sheath cell is more abundant. These defects fully account for the Dyf phenotype and indi- cate that the mutants identify genes involved either in the growth, assembly and functioning of sensory cilia, or in the formation of the amphid channel, or in the secreted matrix material that fills it.
Shakir et al. (1993) have shown that the'gene osm- 3, which is involved in osmotic avoidance, chemotaxis, dauer formation and also has a Dyf phenotype~ encodes a kinesin-like protein, which may suggest a role of the osm-3 gene product in axonal transport. It is likely that also the products of the other Dyf genes function in the chemosensory neurons, or in the support ceils of the amphid. As for mn335, the accumulation of matrix material throughout the cytoplasm of sheath cells may indicate that the primary defect is in the capacity of these cells to secrete the matrix properly. In turn, this defect would result in stunted growth of cilia which are apparently normal in structure. The lack of oth- er Dyf alleles in dyf-1 suggests that the gene product may also be required in other cells besides the amphids and phasmids. Laser ablation of the sheath cell of the
amphid results in a Dyf phenotype and in a variety of chemoreception defects that resemble the pheno- types of most Dyf, Che and Osm mutants (Bargmann, Thomas & Horvitz, 1990). The primary defect of che- 12 mutants also seems to be in the sheath cell (Perkins et al., 1986).
The behavioral assays indicate that much shorter cilia, like those of mn335, still function, albeit less efficiently. This observatior~ is intriguing. It is possible that the changes in the shape of the amphid channel, consequent to the defects in the growth of cilia, result, in turn, in chemoreception defects because of changes in the accessibility of the chemicals to the specific receptors. Probably the machinery of sensory neurons for binding the chemical ligand and transmitting a sig- nal to the post synaptic neurons is intact in mn335 and possibly in other Dyf mutants. The sensory defects are only quantitative and result in clearly altered responses only because a relatively complex behavior is tested. The observation that osmotic avoidance is the most commonly lost response in Dyf mutants supports the notion that the general architecture and proper func- tioning of the amphidial channel are necessary to trans- form the high osmotic strength of the medium into a secondary signal capable of activating the sensory neu- ron (ASH). The sensitivity to other repellents such as garlic may be less dependent on the shape of the chan- nel and rely more directly on specific receptors. Thus it would not usually be lost in Dyf mutants (Bazzicalupo et al., 1994). The increased variability of the avoid- ance responses of dyf-I worms also agrees with this iaterpretatien.
The molecular cloning and analysis of d)f-] and other Dyf genes will clarify this picture and contribute to some important aspects of neurobiology especially regarding the growth and assembly of axonemal cilia of sensory neurons.
Acknowledgements
We thank Robert H. Herman and Todd Starich for shar- ing many unpublished results and for providing mn335. We also thank the C. elegans community and partic- ularly Thomas Barnes for sharing results and a strain carrying the cosmid B0231 in a tandem array. We thank Michele Cermola for help with the electron microscopy and Teresa Vespa and Vito Rita for technical assis- tance. Some nematode strains used in this work were

202
provided by the Caenorhabditis Genetic Center, which is funded by the National Institutes of Health, National Center for Research Resources. This work was in part supported by the EEC program STD, contract CT92- 0096-I, and by CNR Project, Biotecnologie e Biostru- mentazione, to R B. Support was also received from Consorzio Interuniversitario per la Ricerca nei Paesi in via di Sviluppo, Universittt di Roma and by COMIPA, Roma to R B.
References
Bargmann, C. I., E. Hartweig & R. H. Horvitz, 1993. Odorant- selective genes and neurons mediate olfaction in C. elegans. Cell 74: 515-527.
Bargmann, C. I. & H. R. Horvitz, 1991a. Control of larval devel- opment by chemosensory neurons in Caenorhabditis elegans. Science 251: 1243-I246.
Bargmann, C. I. & H. R. Horvitz, 1991b. Chemosensory neurons with overlapping functions direct chemotaxis to multiple chem- icals in C. elegans. Neuron 7: 729-742.
Bargmann, C. I., J. H. Thomas & H. R. Hortivz, 1990. Chemosensory cell function in the behavior and develpment of Caenorhabditis elegans. Cold Spring Harbor Syrup. Quant. Biol. LV: 529-538.
Bazzicalupo, R, L. De Riso, E Ristoratore, E Maimone & M. Sebastiano, 1994. Chemoreception in nematodes, in Advances in Molecular Plant Nematology, edited by C. De Giorgi, E Lamberti and D. Bird. Plenum Press, New York, N.Y. In press.
Chalfie, M. & M. Au, 1989. Genetic control of differentiation of the Caenorhabditis elegans touch receptor neurons. Science 243: 1027-1033.
Chalfie, M. & J. G. White, 1988. The nervous system, pp. 337-391 in The Nematode Caenorhabditis elegans, edited by W. B. Wood, C. S. H. Press, Cold Spring Harbor, N.Y.
Culotti, J. &. D. Russell, 1978. Osmotic avoidance defective mutants of the nematode Caenorhabditiselegans. Genetics 90: 243-256.
Dusenbery, D. B., R. A. Sheridan & R. L. Russell, 1975. Chemo- taxis defective mutants of the nematode Caenorhabditis elegans. Genetics 80: 297-309.
Kaplan, J. M. & H. R. Horvitz, 1993. A dual mechanosensory and chemosensory neuron in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 90: 2227-2231.
Lewis, J. A. & J. Hodgkin, 1977. Specific neuroanatomical changes in chemosensory mutants of the nematode Caenorhabditis ele- gans. J. Comp. Neurol. 172: 489-510.
Perkins, L. A., E. M. Hedgecock, N. Thomson & J. G. Culotti, 1986. Mutant sensory cilia in the nematode Caenorhabditis elegans. Dev. Biol. 117: 456-487.
Riddle, D. L., 1988. The dauer larva, pp. 393-412 in The Nematode Caenorhabditis elegans, edited by W. B. Wood. C.S.H. Press, Cold Spring Harbor, N.Y.
Shakir, M. A., T. Fukushige, H. Yasuda, J. Miwa & S. S. Sid- diqui, 1993. C. elegans osm-3 gene mediating osmotic avoid- ance behaviour encodes a kinesin-like protein. Neuroreport 4: 891-894.
Sulston, J. E., E. Schieremberg, J. G. White & N. Thomson, 1983. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100:64-119.
Vowels, J. J. & J. H. Thomas, 1992. Genetic analysis ofchemosenso- ry control of dauer formation in Caenorhabditis elegans. Genet- ics. 130: 105-127.
Ward, S., 1973. Chemotaxis by the nematode Caenorhabditis ele- gans: Identification of attractants and analysis of the response by the use of mutants. Proc. Natl. Acad. Sci. USA 70: 817-821.
Ward, S., N. Thomson, J. G. White & S. Brenner, 1975. Electron microscopical reconstruction of the anterior sensory anatomy of the nematode Caenorhabditis elegans. J. Comp. Neurol. 160: 313-338.
White, J. G., E. Southgate, J. N. Thomson & S. Brenner, 1986. The structure of the nervous system of the nematode Caenorhabditis elegans. Phil. Trans. Roy. Soc. London B 314: 1-340.




















