
Mechanistic analysis of the search behaviour of Caenorhabditis elegans
11
, 20131092, published 15 January 2014 11 2014 J. R. Soc. Interface Liliana C. M. Salvador, Frederic Bartumeus, Simon A. Levin and William S. Ryu Caenorhabditis elegans Mechanistic analysis of the search behaviour of Supplementary data l http://rsif.royalsocietypublishing.org/content/suppl/2014/01/14/rsif.2013.1092.DC1.htm "Data Supplement" References http://rsif.royalsocietypublishing.org/content/11/92/20131092.full.html#ref-list-1 This article cites 43 articles, 9 of which can be accessed free Email alerting service here right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top http://rsif.royalsocietypublishing.org/subscriptions go to: J. R. Soc. Interface To subscribe to on January 15, 2014 rsif.royalsocietypublishing.org Downloaded from on January 15, 2014 rsif.royalsocietypublishing.org Downloaded from
Transcript of Mechanistic analysis of the search behaviour of Caenorhabditis elegans

20131092 published 15 January 201411 2014 J R Soc Interface Liliana C M Salvador Frederic Bartumeus Simon A Levin and William S Ryu Caenorhabditis elegansMechanistic analysis of the search behaviour of
Supplementary data
l httprsifroyalsocietypublishingorgcontentsuppl20140114rsif20131092DC1htm
Data Supplement
Referenceshttprsifroyalsocietypublishingorgcontent119220131092fullhtmlref-list-1
This article cites 43 articles 9 of which can be accessed free
Email alerting service hereright-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top
httprsifroyalsocietypublishingorgsubscriptions go to J R Soc InterfaceTo subscribe to
on January 15 2014rsifroyalsocietypublishingorgDownloaded from on January 15 2014rsifroyalsocietypublishingorgDownloaded from
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
rsifroyalsocietypublishingorg
ResearchCite this article Salvador LCM Bartumeus F
Levin SA Ryu WS 2014 Mechanistic analysis of
the search behaviour of Caenorhabditis elegans
J R Soc Interface 11 20131092
httpdxdoiorg101098rsif20131092
Received 23 November 2013
Accepted 16 December 2013
Subject Areasbiocomplexity biophysics
computational biology
Keywordssearch behaviour reorientation patterns
environmental uncertainty stochasticity
superdiffusion Caenorhabditis elegans
Author for correspondenceLiliana C M Salvador
e-mail lilianasalvadorglasgowacuk
daggerThese authors contributed equally to this
study
Electronic supplementary material is available
at httpdxdoiorg101098rsif20131092 or
via httprsifroyalsocietypublishingorg
amp 2014 The Author(s) Published by the Royal Society All rights reserved
Mechanistic analysis of the searchbehaviour of Caenorhabditis elegans
Liliana C M Salvador123dagger Frederic Bartumeus24dagger Simon A Levin1
and William S Ryu5
1Department of Ecology and Evolutionary Biology Princeton University Guyot Hall Princeton NJ 08542 USA2ICREA-Movement Ecology Laboratory Centre for Advanced Studies of Blanes (CEAB-CSIC) Cala St Francesc 14Blanes 17300 Spain3Departamento de Biologia Vegetal Faculdade de Ciencias da Universidade de Lisboa Campo GrandeLisboa 1749-016 Portugal4CREAF Cerdanyola del Valles Barcelona 08193 Spain5Department of Physics and the Donnelly Centre University of Toronto 60 St George St TorontoCanada M5S1A7
A central question in movement research is how animals use information and
movement to promote encounter success Current random search theory
identifies reorientation patterns as key to the compromise between optimizing
encounters for both nearby and faraway targets but how the balance between
intrinsic motor programmes and previous environmental experience deter-
mines the occurrence of these reorientation behaviours remains unknown
We used high-resolution tracking and imaging data to describe the complete
motor behaviour of Caenorhabditis elegans when placed in a novel environment
(one in which food is absent) Movement in C elegans is structured around
different reorientation behaviours and we measured how these contributed
to changing search strategies as worms became familiar with their new
environment This behavioural transition shows that different reorientation
behaviours are governed by two processes (i) an environmentally informed
lsquoextrinsicrsquo strategy that is influenced by recent experience and that controls
for area-restricted search behaviour and (ii) a time-independent lsquointrinsicrsquo
strategy that reduces spatial oversampling and improves random encounter
success Our results show how movement strategies arise from a balance
between intrinsic and extrinsic mechanisms that search behaviour in C elegansis initially determined by expectations developed from previous environ-
mental experiences and which reorientation behaviours are modified as
information is acquired from new environments
1 IntroductionNearly all organisms from single cells to large animals forage by controlling their
motor patterns [1ndash3] Foraging is a complex and multifaceted behaviour
It includes pre-detection components such as search and taxis and complex
post-detection events such as pursuing (chasing down) or handling (opening
subduing swallowing) prey [245] Foraging animals change their motor patterns
in response to surrounding environmental cues or as a result of previous experi-
ences and learning processes [2] An animalrsquos response to environmental cues
depends substantially on the spatial and temporal structure of environmental
information [1ndash3] Highly localized cues may trigger area-restricted searching
[67] or acute avoidance reactions [8] whereas environmental gradients are
exploited by both performing taxis where animals move towards a desirable
environment or avoid an unfavourable one or by different types of kinesis (eg
klinokinesis orthokinesis) where movement is modulated by a non-directional
response of the local cue intensity [29] Foraging movement should not only be
thought of as a set of stereotyped behavioural reactions to specific environmental
stimuli [10] External cues may be diffuse (non-directional) of variable quality
(low signal-to-noise ratio) intermittent (complex presencendashabsence dynamics)
or simply absent Under such conditions organisms may undertake specific
rsifroyalsocietypublishingorgJRSocInterface
1120131092
2
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
search strategies to efficiently explore space [11] until some
useful environmental information can be followed and
exploited and such strategies can be thought of as a major com-
ponent of pre-detection foraging behaviour when information
is substantially limited
How should animals interact with the environment and use
information when cues are scarce On the one hand current
random search theory [1213] has shown that reorientation
behaviour can provide an adequate balance between inten-
sive (local) and extensive (non-local) search [1415] releasing
the tension between optimizing for local and distant targets
without previous information about the target spatial dis-
tribution On the other hand we know animals learn and
have expectations based on previous experiences about their
environment Unfortunately even though attempts have
been made [16ndash20] it is still extremely difficult to relate
field-recorded animal trajectories convincingly to recently
developed stochastic search theory [2122] Accurate control
of sensory inputs memory and internal states of the animal
in the wild is overwhelmingly difficult and that limits the
possibility of showing how animals use information to
modify their reorientation patterns or to what extent in the
absence of information intrinsic motor programmes optimize
the balance between intensive (local) and extensive (non-
local) search In movement ecology studies [23] the size
sampling quality and statistical analysis of animal movement
datasets are critical factors Ideally data should be collected
continuously at high frequency to capture fast dynamics
but also over a long timescale to generate statistically
significant samples The determination of behavioural states
[2324] needs to be quantified in a comprehensive and objective
way Environmental conditions also need to be handled
carefully to control the amount of information available to
the animal
An experiment that fits these requirements is the study of
Caenorhabditis elegans search behaviour in a well-controlled
homogeneous laboratory environment C elegans is a small
nematode (1 mm) with a compact well-described neuronal
system and can be studied at many scales genes neurons
and networks [25ndash28] It has a complex yet tractable search
strategy that is generated by a relatively simple locomotive
repertoire [29] On the surface of agar plates worms crawl
forward (crawl) and backwards (reversal) by propagating
undulatory waves along their body These worms can also
interrupt their crawling motions (ie reorientation beha-
viours) by bending their body deeply to form the shape of
the Greek letter omega (omega) by pausing ( pause) or by
executing a more complex composite behaviour ( pirouette)
which is a sequence of a reversal and an omega performed
closely in time [30]
We quantified the wormrsquos search behaviour with a high-
level of detail to characterize the underlying behavioural
mechanisms governing their search patterns We performed
a relocation experiment from a resourced environment to
one without resources in order to investigate how past experi-
ence modulates search with minimal intervention of external
cues and to determine whether innate stochastic search be-
haviour exists Before the experiment C elegans were well
fed and during the experiment they searched the surface of
an agar plate without food The tracking data were sampled
at high resolution and magnification capturing both the tra-
jectory and body posture of single worms freely behaving as
they move on the surface of agar plates [3132] We improved
previous image processing software [31] to be able to detect
worm behaviours with high accuracy and provide a complete
behavioural dataset of individual searching distinguishing
crawling and reorientation types that enable us to characterize
the worm search patterns
2 Material and methods21 Experimental set-up211 Tracking microscopyThe tracking microscope has been described previously [3132] and
is similar to other tracking systems [33ndash36] In summary the ima-
ging system was designed to translocate around a fixed-position
assay plate As the worm moves on the surface of the plate the
system follows the movements of the worm while capturing
images at 4 Hz and recording its centre-of-mass position
212 Worm preparationFifty-two individuals C elegans strain N2 were grown at 208C and
maintained under standard laboratory conditions [37] Fresh
nematode growth medium (NGM) assay plates (17 Bacto agar
025 Bacto-peptone 03 NaCl 1 mM CaCl2 1 mM MgSO4
25 mM potassium phosphate buffer 5 mg ml21 cholesterol) were
partially dried by leaving uncovered for 1 h A copper ring
(51 cm inner diameter) pressed into the agar surface prevented
worms from crawling to the side of the plate Young adults were
rinsed of Escherichia coli by transferring them from OP50 food
plates into NGM buffer (same inorganic ion concentration as
NGM assay plates) and letting them swim for 1 min Individual
worms were transferred from the NGM buffer to the centre
of the assay plate (9 cm Petri dish) The plates were covered and
tracking began after 1 min and lasted no longer than 60 min
22 Diffusive properties of searchTo quantify the worm diffusive properties of the 52 individuals we
computed the mean-squared displacement (MSD) kx2ethtTHORNl of all
worm trajectories (population level) and checked whether the diffu-
sion process was normal (ie MSD increasing linearly with time) or
anomalous (nonlinear relationship with time) The MSD of a set of Nindividual displacements xn from an origin location to a location at
time t is given by kx2ethtTHORNl frac14PN
nfrac141 x2n and kx2ethtTHORNl Ksts where Ks
is the generalized diffusion coefficient and s is the diffusion expo-
nent Diffusivity domains can be distinguished by analysis of the
anomalous diffusion exponent s subdiffusion for 0 s 1 super-
diffusion for s 1 normal Brownian diffusion for s frac14 1 and
ballistic motion for s frac14 2
23 Behavioural flaggingReorientation events of the population of 52 worms were detected
from the segmented binary images using standard imaging pro-
cessing computer vision techniques [37ndash40] and eigenworm
analysis [3141] Binary images were skeletonized to derive the
wormrsquos centreline Omegas were detected by calculating solidity
(less than 070) and errors in skeletonization which occur when
worms touch or cross during omegas If a reversal followed
within two frames (05 s) then the behaviour is flagged as a pirou-
ette The frequency distribution of the time between omegas
following reversals was bimodal therefore we used the maximum
time of the first peak (two frames) as a threshold for the pirouette
behaviour Velocity of the wormrsquos undulatory cycle was measured
using eigenworm analysis [3141] and used to flag reversals and
pauses The behavioural detection algorithm (see electronic sup-
plementary material figure S1) was applied to the image dataset
of 52 individuals 30 min each (total of 374 400 frames) and in
rsifroyalsocietypublishingorgJR
3
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
total it detected 937 reversals 1064 omegas 1125 pirouettes and
387 pauses This method was validated by computing the percen-
tage of behaviours detected by the algorithm to the ones detected
by human observation We used a control dataset of behaviours
observed by eye for five experimental runs (a total of 36 000
frames) representing 10 of the entire dataset The algorithm
detected over 92 (on average) of the number of omegas and rever-
sals that were identified by eye (see the electronic supplementary
material table S1) The 8 misidentified behavioural responses
were due to noisy images in which the worm body could not be
extracted Omegas with low body compressibility (lsquowidersquo omega
bends) and pauses coinciding with a straight body posture were
misidentified by crawls and reversals respectively
SocInterface1120131092
24 Pre-filtering data before analysisIf the worm stopped moving before the completion of the run
then the data were excluded We could see a clear and separable
behavioural effect caused by mechanical stimulus on the reorien-
tation frequency time series for the first 760 frames (32 min)
which we explicitly removed in our final dataset In the end
the following analyses are based on tracks and images of 52 indi-
viduals Tracks were of 268 s each and with a total number
of 2889 reorientation behaviours 788 reversals 860 omegas
935 pirouettes and 306 pauses
25 Orientational memory lossTo study the role of reorientation behaviours on the loss of orien-
tational memory we quantified the effect of each reorientation
type on the overall direction of the trajectory For that we com-
puted (i) the turning angle distribution generated by each
reorientation type and (ii) the angular correlation of the trajec-
tory as a function of reorientation type for each one of the
52 individual worms of our study
Turning angle distribution generated by each reorientation typereorientations are located in between two crawls (periods of
forward motion) The turning angle u generated via a specific reor-
ientation was computed by calculating the difference between the
absolute angles of the vectors following the overall direction of
the crawl just before and just after the reorientation The vectors
are defined using the first and last time frames of each crawl
Angular correlation of the trajectory as a function of reorientation typethe angular correlation function
CaethtTHORN frac141
n
Xn
jfrac141
kcosfrac12aethtthorn tTHORN aethtTHORNlj
where a(t) is the local tangent angle at time t t is the time lag
(from 1 to 13 s) j is the worm id and n is the total number of
worms (52) was calculated from the centre-of-mass data (trajec-
tory) to study the wormrsquos directionality of motion over time We
combined the behavioural data with the trajectory data and
sampled segment types ST from the trajectory that are crawls sep-
arated by a specific type T of reorientation (eg crawlndashreversalndash
crawl) The angular correlation function was computed for both
the original segments ST and their correspondent null model
which is a bootstrapping (499 times) of random segments of the
same length as ST of the same trajectory (see the electronic sup-
plementary material figure S8) The correlation function Ca(t)
was computed for each ST using different lag sizes t and it was
averaged over all the segments over all the 52 individuals (black
line in figure 3) For the null model case standard error bars of
the mean of Ca(t) were computed for each lag size t (dashed line
in figure 3) The difference between the solid lines representing
sequences of crawls interrupted by reorientation types and the
dashed line representing the null model (random segments of
the original trajectory) show how strong the contribution of each
behaviour is towards changes in direction The further the lines
are from each other the larger the contribution of the reorientation
behaviour is evident
26 Probability distribution of omega time inter-eventsWe selected among three probabilistic models of the exponential
family (simple double and stretched exponentials) to identify the
best-fitting probability distribution of the time intervals between
omegas at both the individual and population levels of the 52 indi-
vidual worms In particular we used KolmogorovndashSmirnov (KS)
tests [42] and the likelihood functions of the probability density
functions (pdfs of the three models) over a bounded range and
for pre-binned data [17] We described the three pdfs their corre-
sponding transformation when considering pre-binned data and
their resulting likelihood function in the electronic supplementary
material text S1 For model selection we include only those indi-
viduals that had performed at least 30 omegas during the
tracking period for the individual level analysis
27 Relationship between reorientation and crawl typesTo test the statistical independence between reorientation and
crawl types in our worm population we performed a contingency
table analysis followed by a chi-squared approximation for
proportions [43] We determined both the relationship between
reorientations and crawls occurring immediately after and
between reorientations and crawls that preceded them For the
dependent relationship between reorientations and their previous
crawls we studied in more detail the connection between the
different types of behaviours In particular we performed a two-
tailed Z-score test statistic Z (a frac14 005) [43] to test the null
hypothesis H0 pethCijRjTHORN frac14 pethCiTHORN against the alternative hypothesis
HA pethCijRjTHORN= pethCiTHORN where i and j represent respectively the
different crawl and reorientation types presented in figure 5b
For the behaviours with dependent relationships we used the
departures from the expected frequency between reorientation
events and their previous crawls to determine whether they are
positively (above expected frequency) or negatively (below the
expected frequency) correlated
28 Average time to close a loop and characteristicinter-event times for omegas
The average time to close a loop was calculated as the average time
to close a circle t frac14 2prs where r frac14 1jkj and s is the speed These
averages are taken over all the trajectories that contain loops The
average speed of a loop is 03 mm s21 and the median of loopsrsquo
curvature k is 41 mmndash 1 so t frac14 54 s The characteristic inter-
event time for omegas q frac14 67 s was taken directly from the
stretched exponential likelihood fit at the population level
29 Models of search behaviourInspired by our data analyses we have developed a modelling
framework that allows us to compare the search efficiency in a
patchy landscape of simple correlated random walks with dif-
ferent degrees of sinuosity r (crawling behaviour) to strategies
that additionally incorporate both stationarynon-modulated
(omega) and non-stationarysignal-modulated (pirouette) reor-
ientation behaviours To do so we use four behavioural
combinations according to the models in the electronic sup-
plementary material table S2 model 1 crawling behaviour
(correlated random walk) model 2 crawling behaviour with
omegas model 3 crawling behaviour with pirouettes and
model 4 crawling behaviour with both omegas and pirouettes
The details of the modelling framework are provided in the elec-
tronic supplementary material Search efficiency was computed
as the number of encounters per travelled distance averaged
over 500 individuals
0 103
(a)
(c)
(b)
102
s1 = 137
s2 = 046
10
1
10ndash1
5 pause
pirou
omeg
rev
crawls
10
15
20
25
30indi
vidu
als
35
40
45
50
5 10 15time (min)
20 25
10 102
log(t)
log(
MSD
)
103
end start
behavioursreversalsomegas pauses
pirouettes
25 5 mm
Figure 1 Individual worm track and population ethogram (a) A 30 min tracking run showing the centre-of-mass movements of a single C elegans worm Insetshows raw worm image Behavioural events are labelled as crawls (black) reversals (blue) omegas (cyan) pirouettes (orange) and pauses (red) (b) Spreadingcapacity of the worm population measured as the MSD across time (52 worm trajectories starting from the same origin point) Two regimes were found super-diffusive with slope s1 frac14 137 1 (solid line) and subdiffusive with 0 s2 frac14 046 1 (dashed line) The worm population does not passively diffuse throughthe environment Their spreading is accelerated covering a range of spatio-temporal scales up to the limiting scale of the experimental system (51 cm) wherespreading then becomes subdiffusive (c) Population ethogram (n frac14 52) showing individual behavioural variability Behavioural events are colour coded as aboveexcept crawls that are in grey
rsifroyalsocietypublishingorgJRSocInterface
1120131092
4
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
3 ResultsThe tracking system captures both the trajectory and the body
postures of single worms freely moving on the surface of agar
plates (Material and methods) Figure 1a shows representative
high-resolution tracking data and a detailed image of the
worm captured during the experiment The scaling behaviour
of the MSD across time shows that the worms do not passively
diffuse while searching but instead perform more complex
movements such that the population spreading while searching
is superdiffusive with an anomalous diffusion exponent s1 frac14
137 (greater than 1) up to the limiting scale of the experimen-
tal system that can be associated with the crossover towards a
subdiffusive regime s2 frac14 046 (less than 1 figure 1b) The
pairing of large-scale tracks (worm centre-of-mass trajectory)
and small-scale behavioural data (body postures) allows us to
flag crawls and reorientation behavioural events and produce
comprehensive trajectories (figure 1a) and ethograms (figure
1c) that show the transition between a number of crawling
and reorientation behaviours used by worms to explore the
environment Our analysis indicates a number of different
types of crawling motions and reorientation behaviours Crawl-
ing motions are not always straight but often form arcing or
looping trajectories [32] We flagged crawling behaviour
based on curvature and angular concordance [44] into four cat-
egories lines open arcs closed arcs and loops (see the
electronic supplementary material text S2 and figures S2 and
S3) We identified four types of reorientation behaviour
reversals omegas pauses and pirouettes (figure 1c Material
and methods)
The overall movement patterns did not show any direction-
al bias (Rayleigh tests of uniformity [45] applied to the first and
0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

on January 15 2014rsifroyalsocietypublishingorgDownloaded from
rsifroyalsocietypublishingorg
ResearchCite this article Salvador LCM Bartumeus F
Levin SA Ryu WS 2014 Mechanistic analysis of
the search behaviour of Caenorhabditis elegans
J R Soc Interface 11 20131092
httpdxdoiorg101098rsif20131092
Received 23 November 2013
Accepted 16 December 2013
Subject Areasbiocomplexity biophysics
computational biology
Keywordssearch behaviour reorientation patterns
environmental uncertainty stochasticity
superdiffusion Caenorhabditis elegans
Author for correspondenceLiliana C M Salvador
e-mail lilianasalvadorglasgowacuk
daggerThese authors contributed equally to this
study
Electronic supplementary material is available
at httpdxdoiorg101098rsif20131092 or
via httprsifroyalsocietypublishingorg
amp 2014 The Author(s) Published by the Royal Society All rights reserved
Mechanistic analysis of the searchbehaviour of Caenorhabditis elegans
Liliana C M Salvador123dagger Frederic Bartumeus24dagger Simon A Levin1
and William S Ryu5
1Department of Ecology and Evolutionary Biology Princeton University Guyot Hall Princeton NJ 08542 USA2ICREA-Movement Ecology Laboratory Centre for Advanced Studies of Blanes (CEAB-CSIC) Cala St Francesc 14Blanes 17300 Spain3Departamento de Biologia Vegetal Faculdade de Ciencias da Universidade de Lisboa Campo GrandeLisboa 1749-016 Portugal4CREAF Cerdanyola del Valles Barcelona 08193 Spain5Department of Physics and the Donnelly Centre University of Toronto 60 St George St TorontoCanada M5S1A7
A central question in movement research is how animals use information and
movement to promote encounter success Current random search theory
identifies reorientation patterns as key to the compromise between optimizing
encounters for both nearby and faraway targets but how the balance between
intrinsic motor programmes and previous environmental experience deter-
mines the occurrence of these reorientation behaviours remains unknown
We used high-resolution tracking and imaging data to describe the complete
motor behaviour of Caenorhabditis elegans when placed in a novel environment
(one in which food is absent) Movement in C elegans is structured around
different reorientation behaviours and we measured how these contributed
to changing search strategies as worms became familiar with their new
environment This behavioural transition shows that different reorientation
behaviours are governed by two processes (i) an environmentally informed
lsquoextrinsicrsquo strategy that is influenced by recent experience and that controls
for area-restricted search behaviour and (ii) a time-independent lsquointrinsicrsquo
strategy that reduces spatial oversampling and improves random encounter
success Our results show how movement strategies arise from a balance
between intrinsic and extrinsic mechanisms that search behaviour in C elegansis initially determined by expectations developed from previous environ-
mental experiences and which reorientation behaviours are modified as
information is acquired from new environments
1 IntroductionNearly all organisms from single cells to large animals forage by controlling their
motor patterns [1ndash3] Foraging is a complex and multifaceted behaviour
It includes pre-detection components such as search and taxis and complex
post-detection events such as pursuing (chasing down) or handling (opening
subduing swallowing) prey [245] Foraging animals change their motor patterns
in response to surrounding environmental cues or as a result of previous experi-
ences and learning processes [2] An animalrsquos response to environmental cues
depends substantially on the spatial and temporal structure of environmental
information [1ndash3] Highly localized cues may trigger area-restricted searching
[67] or acute avoidance reactions [8] whereas environmental gradients are
exploited by both performing taxis where animals move towards a desirable
environment or avoid an unfavourable one or by different types of kinesis (eg
klinokinesis orthokinesis) where movement is modulated by a non-directional
response of the local cue intensity [29] Foraging movement should not only be
thought of as a set of stereotyped behavioural reactions to specific environmental
stimuli [10] External cues may be diffuse (non-directional) of variable quality
(low signal-to-noise ratio) intermittent (complex presencendashabsence dynamics)
or simply absent Under such conditions organisms may undertake specific
rsifroyalsocietypublishingorgJRSocInterface
1120131092
2
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
search strategies to efficiently explore space [11] until some
useful environmental information can be followed and
exploited and such strategies can be thought of as a major com-
ponent of pre-detection foraging behaviour when information
is substantially limited
How should animals interact with the environment and use
information when cues are scarce On the one hand current
random search theory [1213] has shown that reorientation
behaviour can provide an adequate balance between inten-
sive (local) and extensive (non-local) search [1415] releasing
the tension between optimizing for local and distant targets
without previous information about the target spatial dis-
tribution On the other hand we know animals learn and
have expectations based on previous experiences about their
environment Unfortunately even though attempts have
been made [16ndash20] it is still extremely difficult to relate
field-recorded animal trajectories convincingly to recently
developed stochastic search theory [2122] Accurate control
of sensory inputs memory and internal states of the animal
in the wild is overwhelmingly difficult and that limits the
possibility of showing how animals use information to
modify their reorientation patterns or to what extent in the
absence of information intrinsic motor programmes optimize
the balance between intensive (local) and extensive (non-
local) search In movement ecology studies [23] the size
sampling quality and statistical analysis of animal movement
datasets are critical factors Ideally data should be collected
continuously at high frequency to capture fast dynamics
but also over a long timescale to generate statistically
significant samples The determination of behavioural states
[2324] needs to be quantified in a comprehensive and objective
way Environmental conditions also need to be handled
carefully to control the amount of information available to
the animal
An experiment that fits these requirements is the study of
Caenorhabditis elegans search behaviour in a well-controlled
homogeneous laboratory environment C elegans is a small
nematode (1 mm) with a compact well-described neuronal
system and can be studied at many scales genes neurons
and networks [25ndash28] It has a complex yet tractable search
strategy that is generated by a relatively simple locomotive
repertoire [29] On the surface of agar plates worms crawl
forward (crawl) and backwards (reversal) by propagating
undulatory waves along their body These worms can also
interrupt their crawling motions (ie reorientation beha-
viours) by bending their body deeply to form the shape of
the Greek letter omega (omega) by pausing ( pause) or by
executing a more complex composite behaviour ( pirouette)
which is a sequence of a reversal and an omega performed
closely in time [30]
We quantified the wormrsquos search behaviour with a high-
level of detail to characterize the underlying behavioural
mechanisms governing their search patterns We performed
a relocation experiment from a resourced environment to
one without resources in order to investigate how past experi-
ence modulates search with minimal intervention of external
cues and to determine whether innate stochastic search be-
haviour exists Before the experiment C elegans were well
fed and during the experiment they searched the surface of
an agar plate without food The tracking data were sampled
at high resolution and magnification capturing both the tra-
jectory and body posture of single worms freely behaving as
they move on the surface of agar plates [3132] We improved
previous image processing software [31] to be able to detect
worm behaviours with high accuracy and provide a complete
behavioural dataset of individual searching distinguishing
crawling and reorientation types that enable us to characterize
the worm search patterns
2 Material and methods21 Experimental set-up211 Tracking microscopyThe tracking microscope has been described previously [3132] and
is similar to other tracking systems [33ndash36] In summary the ima-
ging system was designed to translocate around a fixed-position
assay plate As the worm moves on the surface of the plate the
system follows the movements of the worm while capturing
images at 4 Hz and recording its centre-of-mass position
212 Worm preparationFifty-two individuals C elegans strain N2 were grown at 208C and
maintained under standard laboratory conditions [37] Fresh
nematode growth medium (NGM) assay plates (17 Bacto agar
025 Bacto-peptone 03 NaCl 1 mM CaCl2 1 mM MgSO4
25 mM potassium phosphate buffer 5 mg ml21 cholesterol) were
partially dried by leaving uncovered for 1 h A copper ring
(51 cm inner diameter) pressed into the agar surface prevented
worms from crawling to the side of the plate Young adults were
rinsed of Escherichia coli by transferring them from OP50 food
plates into NGM buffer (same inorganic ion concentration as
NGM assay plates) and letting them swim for 1 min Individual
worms were transferred from the NGM buffer to the centre
of the assay plate (9 cm Petri dish) The plates were covered and
tracking began after 1 min and lasted no longer than 60 min
22 Diffusive properties of searchTo quantify the worm diffusive properties of the 52 individuals we
computed the mean-squared displacement (MSD) kx2ethtTHORNl of all
worm trajectories (population level) and checked whether the diffu-
sion process was normal (ie MSD increasing linearly with time) or
anomalous (nonlinear relationship with time) The MSD of a set of Nindividual displacements xn from an origin location to a location at
time t is given by kx2ethtTHORNl frac14PN
nfrac141 x2n and kx2ethtTHORNl Ksts where Ks
is the generalized diffusion coefficient and s is the diffusion expo-
nent Diffusivity domains can be distinguished by analysis of the
anomalous diffusion exponent s subdiffusion for 0 s 1 super-
diffusion for s 1 normal Brownian diffusion for s frac14 1 and
ballistic motion for s frac14 2
23 Behavioural flaggingReorientation events of the population of 52 worms were detected
from the segmented binary images using standard imaging pro-
cessing computer vision techniques [37ndash40] and eigenworm
analysis [3141] Binary images were skeletonized to derive the
wormrsquos centreline Omegas were detected by calculating solidity
(less than 070) and errors in skeletonization which occur when
worms touch or cross during omegas If a reversal followed
within two frames (05 s) then the behaviour is flagged as a pirou-
ette The frequency distribution of the time between omegas
following reversals was bimodal therefore we used the maximum
time of the first peak (two frames) as a threshold for the pirouette
behaviour Velocity of the wormrsquos undulatory cycle was measured
using eigenworm analysis [3141] and used to flag reversals and
pauses The behavioural detection algorithm (see electronic sup-
plementary material figure S1) was applied to the image dataset
of 52 individuals 30 min each (total of 374 400 frames) and in
rsifroyalsocietypublishingorgJR
3
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
total it detected 937 reversals 1064 omegas 1125 pirouettes and
387 pauses This method was validated by computing the percen-
tage of behaviours detected by the algorithm to the ones detected
by human observation We used a control dataset of behaviours
observed by eye for five experimental runs (a total of 36 000
frames) representing 10 of the entire dataset The algorithm
detected over 92 (on average) of the number of omegas and rever-
sals that were identified by eye (see the electronic supplementary
material table S1) The 8 misidentified behavioural responses
were due to noisy images in which the worm body could not be
extracted Omegas with low body compressibility (lsquowidersquo omega
bends) and pauses coinciding with a straight body posture were
misidentified by crawls and reversals respectively
SocInterface1120131092
24 Pre-filtering data before analysisIf the worm stopped moving before the completion of the run
then the data were excluded We could see a clear and separable
behavioural effect caused by mechanical stimulus on the reorien-
tation frequency time series for the first 760 frames (32 min)
which we explicitly removed in our final dataset In the end
the following analyses are based on tracks and images of 52 indi-
viduals Tracks were of 268 s each and with a total number
of 2889 reorientation behaviours 788 reversals 860 omegas
935 pirouettes and 306 pauses
25 Orientational memory lossTo study the role of reorientation behaviours on the loss of orien-
tational memory we quantified the effect of each reorientation
type on the overall direction of the trajectory For that we com-
puted (i) the turning angle distribution generated by each
reorientation type and (ii) the angular correlation of the trajec-
tory as a function of reorientation type for each one of the
52 individual worms of our study
Turning angle distribution generated by each reorientation typereorientations are located in between two crawls (periods of
forward motion) The turning angle u generated via a specific reor-
ientation was computed by calculating the difference between the
absolute angles of the vectors following the overall direction of
the crawl just before and just after the reorientation The vectors
are defined using the first and last time frames of each crawl
Angular correlation of the trajectory as a function of reorientation typethe angular correlation function
CaethtTHORN frac141
n
Xn
jfrac141
kcosfrac12aethtthorn tTHORN aethtTHORNlj
where a(t) is the local tangent angle at time t t is the time lag
(from 1 to 13 s) j is the worm id and n is the total number of
worms (52) was calculated from the centre-of-mass data (trajec-
tory) to study the wormrsquos directionality of motion over time We
combined the behavioural data with the trajectory data and
sampled segment types ST from the trajectory that are crawls sep-
arated by a specific type T of reorientation (eg crawlndashreversalndash
crawl) The angular correlation function was computed for both
the original segments ST and their correspondent null model
which is a bootstrapping (499 times) of random segments of the
same length as ST of the same trajectory (see the electronic sup-
plementary material figure S8) The correlation function Ca(t)
was computed for each ST using different lag sizes t and it was
averaged over all the segments over all the 52 individuals (black
line in figure 3) For the null model case standard error bars of
the mean of Ca(t) were computed for each lag size t (dashed line
in figure 3) The difference between the solid lines representing
sequences of crawls interrupted by reorientation types and the
dashed line representing the null model (random segments of
the original trajectory) show how strong the contribution of each
behaviour is towards changes in direction The further the lines
are from each other the larger the contribution of the reorientation
behaviour is evident
26 Probability distribution of omega time inter-eventsWe selected among three probabilistic models of the exponential
family (simple double and stretched exponentials) to identify the
best-fitting probability distribution of the time intervals between
omegas at both the individual and population levels of the 52 indi-
vidual worms In particular we used KolmogorovndashSmirnov (KS)
tests [42] and the likelihood functions of the probability density
functions (pdfs of the three models) over a bounded range and
for pre-binned data [17] We described the three pdfs their corre-
sponding transformation when considering pre-binned data and
their resulting likelihood function in the electronic supplementary
material text S1 For model selection we include only those indi-
viduals that had performed at least 30 omegas during the
tracking period for the individual level analysis
27 Relationship between reorientation and crawl typesTo test the statistical independence between reorientation and
crawl types in our worm population we performed a contingency
table analysis followed by a chi-squared approximation for
proportions [43] We determined both the relationship between
reorientations and crawls occurring immediately after and
between reorientations and crawls that preceded them For the
dependent relationship between reorientations and their previous
crawls we studied in more detail the connection between the
different types of behaviours In particular we performed a two-
tailed Z-score test statistic Z (a frac14 005) [43] to test the null
hypothesis H0 pethCijRjTHORN frac14 pethCiTHORN against the alternative hypothesis
HA pethCijRjTHORN= pethCiTHORN where i and j represent respectively the
different crawl and reorientation types presented in figure 5b
For the behaviours with dependent relationships we used the
departures from the expected frequency between reorientation
events and their previous crawls to determine whether they are
positively (above expected frequency) or negatively (below the
expected frequency) correlated
28 Average time to close a loop and characteristicinter-event times for omegas
The average time to close a loop was calculated as the average time
to close a circle t frac14 2prs where r frac14 1jkj and s is the speed These
averages are taken over all the trajectories that contain loops The
average speed of a loop is 03 mm s21 and the median of loopsrsquo
curvature k is 41 mmndash 1 so t frac14 54 s The characteristic inter-
event time for omegas q frac14 67 s was taken directly from the
stretched exponential likelihood fit at the population level
29 Models of search behaviourInspired by our data analyses we have developed a modelling
framework that allows us to compare the search efficiency in a
patchy landscape of simple correlated random walks with dif-
ferent degrees of sinuosity r (crawling behaviour) to strategies
that additionally incorporate both stationarynon-modulated
(omega) and non-stationarysignal-modulated (pirouette) reor-
ientation behaviours To do so we use four behavioural
combinations according to the models in the electronic sup-
plementary material table S2 model 1 crawling behaviour
(correlated random walk) model 2 crawling behaviour with
omegas model 3 crawling behaviour with pirouettes and
model 4 crawling behaviour with both omegas and pirouettes
The details of the modelling framework are provided in the elec-
tronic supplementary material Search efficiency was computed
as the number of encounters per travelled distance averaged
over 500 individuals
0 103
(a)
(c)
(b)
102
s1 = 137
s2 = 046
10
1
10ndash1
5 pause
pirou
omeg
rev
crawls
10
15
20
25
30indi
vidu
als
35
40
45
50
5 10 15time (min)
20 25
10 102
log(t)
log(
MSD
)
103
end start
behavioursreversalsomegas pauses
pirouettes
25 5 mm
Figure 1 Individual worm track and population ethogram (a) A 30 min tracking run showing the centre-of-mass movements of a single C elegans worm Insetshows raw worm image Behavioural events are labelled as crawls (black) reversals (blue) omegas (cyan) pirouettes (orange) and pauses (red) (b) Spreadingcapacity of the worm population measured as the MSD across time (52 worm trajectories starting from the same origin point) Two regimes were found super-diffusive with slope s1 frac14 137 1 (solid line) and subdiffusive with 0 s2 frac14 046 1 (dashed line) The worm population does not passively diffuse throughthe environment Their spreading is accelerated covering a range of spatio-temporal scales up to the limiting scale of the experimental system (51 cm) wherespreading then becomes subdiffusive (c) Population ethogram (n frac14 52) showing individual behavioural variability Behavioural events are colour coded as aboveexcept crawls that are in grey
rsifroyalsocietypublishingorgJRSocInterface
1120131092
4
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
3 ResultsThe tracking system captures both the trajectory and the body
postures of single worms freely moving on the surface of agar
plates (Material and methods) Figure 1a shows representative
high-resolution tracking data and a detailed image of the
worm captured during the experiment The scaling behaviour
of the MSD across time shows that the worms do not passively
diffuse while searching but instead perform more complex
movements such that the population spreading while searching
is superdiffusive with an anomalous diffusion exponent s1 frac14
137 (greater than 1) up to the limiting scale of the experimen-
tal system that can be associated with the crossover towards a
subdiffusive regime s2 frac14 046 (less than 1 figure 1b) The
pairing of large-scale tracks (worm centre-of-mass trajectory)
and small-scale behavioural data (body postures) allows us to
flag crawls and reorientation behavioural events and produce
comprehensive trajectories (figure 1a) and ethograms (figure
1c) that show the transition between a number of crawling
and reorientation behaviours used by worms to explore the
environment Our analysis indicates a number of different
types of crawling motions and reorientation behaviours Crawl-
ing motions are not always straight but often form arcing or
looping trajectories [32] We flagged crawling behaviour
based on curvature and angular concordance [44] into four cat-
egories lines open arcs closed arcs and loops (see the
electronic supplementary material text S2 and figures S2 and
S3) We identified four types of reorientation behaviour
reversals omegas pauses and pirouettes (figure 1c Material
and methods)
The overall movement patterns did not show any direction-
al bias (Rayleigh tests of uniformity [45] applied to the first and
0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

rsifroyalsocietypublishingorgJRSocInterface
1120131092
2
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
search strategies to efficiently explore space [11] until some
useful environmental information can be followed and
exploited and such strategies can be thought of as a major com-
ponent of pre-detection foraging behaviour when information
is substantially limited
How should animals interact with the environment and use
information when cues are scarce On the one hand current
random search theory [1213] has shown that reorientation
behaviour can provide an adequate balance between inten-
sive (local) and extensive (non-local) search [1415] releasing
the tension between optimizing for local and distant targets
without previous information about the target spatial dis-
tribution On the other hand we know animals learn and
have expectations based on previous experiences about their
environment Unfortunately even though attempts have
been made [16ndash20] it is still extremely difficult to relate
field-recorded animal trajectories convincingly to recently
developed stochastic search theory [2122] Accurate control
of sensory inputs memory and internal states of the animal
in the wild is overwhelmingly difficult and that limits the
possibility of showing how animals use information to
modify their reorientation patterns or to what extent in the
absence of information intrinsic motor programmes optimize
the balance between intensive (local) and extensive (non-
local) search In movement ecology studies [23] the size
sampling quality and statistical analysis of animal movement
datasets are critical factors Ideally data should be collected
continuously at high frequency to capture fast dynamics
but also over a long timescale to generate statistically
significant samples The determination of behavioural states
[2324] needs to be quantified in a comprehensive and objective
way Environmental conditions also need to be handled
carefully to control the amount of information available to
the animal
An experiment that fits these requirements is the study of
Caenorhabditis elegans search behaviour in a well-controlled
homogeneous laboratory environment C elegans is a small
nematode (1 mm) with a compact well-described neuronal
system and can be studied at many scales genes neurons
and networks [25ndash28] It has a complex yet tractable search
strategy that is generated by a relatively simple locomotive
repertoire [29] On the surface of agar plates worms crawl
forward (crawl) and backwards (reversal) by propagating
undulatory waves along their body These worms can also
interrupt their crawling motions (ie reorientation beha-
viours) by bending their body deeply to form the shape of
the Greek letter omega (omega) by pausing ( pause) or by
executing a more complex composite behaviour ( pirouette)
which is a sequence of a reversal and an omega performed
closely in time [30]
We quantified the wormrsquos search behaviour with a high-
level of detail to characterize the underlying behavioural
mechanisms governing their search patterns We performed
a relocation experiment from a resourced environment to
one without resources in order to investigate how past experi-
ence modulates search with minimal intervention of external
cues and to determine whether innate stochastic search be-
haviour exists Before the experiment C elegans were well
fed and during the experiment they searched the surface of
an agar plate without food The tracking data were sampled
at high resolution and magnification capturing both the tra-
jectory and body posture of single worms freely behaving as
they move on the surface of agar plates [3132] We improved
previous image processing software [31] to be able to detect
worm behaviours with high accuracy and provide a complete
behavioural dataset of individual searching distinguishing
crawling and reorientation types that enable us to characterize
the worm search patterns
2 Material and methods21 Experimental set-up211 Tracking microscopyThe tracking microscope has been described previously [3132] and
is similar to other tracking systems [33ndash36] In summary the ima-
ging system was designed to translocate around a fixed-position
assay plate As the worm moves on the surface of the plate the
system follows the movements of the worm while capturing
images at 4 Hz and recording its centre-of-mass position
212 Worm preparationFifty-two individuals C elegans strain N2 were grown at 208C and
maintained under standard laboratory conditions [37] Fresh
nematode growth medium (NGM) assay plates (17 Bacto agar
025 Bacto-peptone 03 NaCl 1 mM CaCl2 1 mM MgSO4
25 mM potassium phosphate buffer 5 mg ml21 cholesterol) were
partially dried by leaving uncovered for 1 h A copper ring
(51 cm inner diameter) pressed into the agar surface prevented
worms from crawling to the side of the plate Young adults were
rinsed of Escherichia coli by transferring them from OP50 food
plates into NGM buffer (same inorganic ion concentration as
NGM assay plates) and letting them swim for 1 min Individual
worms were transferred from the NGM buffer to the centre
of the assay plate (9 cm Petri dish) The plates were covered and
tracking began after 1 min and lasted no longer than 60 min
22 Diffusive properties of searchTo quantify the worm diffusive properties of the 52 individuals we
computed the mean-squared displacement (MSD) kx2ethtTHORNl of all
worm trajectories (population level) and checked whether the diffu-
sion process was normal (ie MSD increasing linearly with time) or
anomalous (nonlinear relationship with time) The MSD of a set of Nindividual displacements xn from an origin location to a location at
time t is given by kx2ethtTHORNl frac14PN
nfrac141 x2n and kx2ethtTHORNl Ksts where Ks
is the generalized diffusion coefficient and s is the diffusion expo-
nent Diffusivity domains can be distinguished by analysis of the
anomalous diffusion exponent s subdiffusion for 0 s 1 super-
diffusion for s 1 normal Brownian diffusion for s frac14 1 and
ballistic motion for s frac14 2
23 Behavioural flaggingReorientation events of the population of 52 worms were detected
from the segmented binary images using standard imaging pro-
cessing computer vision techniques [37ndash40] and eigenworm
analysis [3141] Binary images were skeletonized to derive the
wormrsquos centreline Omegas were detected by calculating solidity
(less than 070) and errors in skeletonization which occur when
worms touch or cross during omegas If a reversal followed
within two frames (05 s) then the behaviour is flagged as a pirou-
ette The frequency distribution of the time between omegas
following reversals was bimodal therefore we used the maximum
time of the first peak (two frames) as a threshold for the pirouette
behaviour Velocity of the wormrsquos undulatory cycle was measured
using eigenworm analysis [3141] and used to flag reversals and
pauses The behavioural detection algorithm (see electronic sup-
plementary material figure S1) was applied to the image dataset
of 52 individuals 30 min each (total of 374 400 frames) and in
rsifroyalsocietypublishingorgJR
3
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
total it detected 937 reversals 1064 omegas 1125 pirouettes and
387 pauses This method was validated by computing the percen-
tage of behaviours detected by the algorithm to the ones detected
by human observation We used a control dataset of behaviours
observed by eye for five experimental runs (a total of 36 000
frames) representing 10 of the entire dataset The algorithm
detected over 92 (on average) of the number of omegas and rever-
sals that were identified by eye (see the electronic supplementary
material table S1) The 8 misidentified behavioural responses
were due to noisy images in which the worm body could not be
extracted Omegas with low body compressibility (lsquowidersquo omega
bends) and pauses coinciding with a straight body posture were
misidentified by crawls and reversals respectively
SocInterface1120131092
24 Pre-filtering data before analysisIf the worm stopped moving before the completion of the run
then the data were excluded We could see a clear and separable
behavioural effect caused by mechanical stimulus on the reorien-
tation frequency time series for the first 760 frames (32 min)
which we explicitly removed in our final dataset In the end
the following analyses are based on tracks and images of 52 indi-
viduals Tracks were of 268 s each and with a total number
of 2889 reorientation behaviours 788 reversals 860 omegas
935 pirouettes and 306 pauses
25 Orientational memory lossTo study the role of reorientation behaviours on the loss of orien-
tational memory we quantified the effect of each reorientation
type on the overall direction of the trajectory For that we com-
puted (i) the turning angle distribution generated by each
reorientation type and (ii) the angular correlation of the trajec-
tory as a function of reorientation type for each one of the
52 individual worms of our study
Turning angle distribution generated by each reorientation typereorientations are located in between two crawls (periods of
forward motion) The turning angle u generated via a specific reor-
ientation was computed by calculating the difference between the
absolute angles of the vectors following the overall direction of
the crawl just before and just after the reorientation The vectors
are defined using the first and last time frames of each crawl
Angular correlation of the trajectory as a function of reorientation typethe angular correlation function
CaethtTHORN frac141
n
Xn
jfrac141
kcosfrac12aethtthorn tTHORN aethtTHORNlj
where a(t) is the local tangent angle at time t t is the time lag
(from 1 to 13 s) j is the worm id and n is the total number of
worms (52) was calculated from the centre-of-mass data (trajec-
tory) to study the wormrsquos directionality of motion over time We
combined the behavioural data with the trajectory data and
sampled segment types ST from the trajectory that are crawls sep-
arated by a specific type T of reorientation (eg crawlndashreversalndash
crawl) The angular correlation function was computed for both
the original segments ST and their correspondent null model
which is a bootstrapping (499 times) of random segments of the
same length as ST of the same trajectory (see the electronic sup-
plementary material figure S8) The correlation function Ca(t)
was computed for each ST using different lag sizes t and it was
averaged over all the segments over all the 52 individuals (black
line in figure 3) For the null model case standard error bars of
the mean of Ca(t) were computed for each lag size t (dashed line
in figure 3) The difference between the solid lines representing
sequences of crawls interrupted by reorientation types and the
dashed line representing the null model (random segments of
the original trajectory) show how strong the contribution of each
behaviour is towards changes in direction The further the lines
are from each other the larger the contribution of the reorientation
behaviour is evident
26 Probability distribution of omega time inter-eventsWe selected among three probabilistic models of the exponential
family (simple double and stretched exponentials) to identify the
best-fitting probability distribution of the time intervals between
omegas at both the individual and population levels of the 52 indi-
vidual worms In particular we used KolmogorovndashSmirnov (KS)
tests [42] and the likelihood functions of the probability density
functions (pdfs of the three models) over a bounded range and
for pre-binned data [17] We described the three pdfs their corre-
sponding transformation when considering pre-binned data and
their resulting likelihood function in the electronic supplementary
material text S1 For model selection we include only those indi-
viduals that had performed at least 30 omegas during the
tracking period for the individual level analysis
27 Relationship between reorientation and crawl typesTo test the statistical independence between reorientation and
crawl types in our worm population we performed a contingency
table analysis followed by a chi-squared approximation for
proportions [43] We determined both the relationship between
reorientations and crawls occurring immediately after and
between reorientations and crawls that preceded them For the
dependent relationship between reorientations and their previous
crawls we studied in more detail the connection between the
different types of behaviours In particular we performed a two-
tailed Z-score test statistic Z (a frac14 005) [43] to test the null
hypothesis H0 pethCijRjTHORN frac14 pethCiTHORN against the alternative hypothesis
HA pethCijRjTHORN= pethCiTHORN where i and j represent respectively the
different crawl and reorientation types presented in figure 5b
For the behaviours with dependent relationships we used the
departures from the expected frequency between reorientation
events and their previous crawls to determine whether they are
positively (above expected frequency) or negatively (below the
expected frequency) correlated
28 Average time to close a loop and characteristicinter-event times for omegas
The average time to close a loop was calculated as the average time
to close a circle t frac14 2prs where r frac14 1jkj and s is the speed These
averages are taken over all the trajectories that contain loops The
average speed of a loop is 03 mm s21 and the median of loopsrsquo
curvature k is 41 mmndash 1 so t frac14 54 s The characteristic inter-
event time for omegas q frac14 67 s was taken directly from the
stretched exponential likelihood fit at the population level
29 Models of search behaviourInspired by our data analyses we have developed a modelling
framework that allows us to compare the search efficiency in a
patchy landscape of simple correlated random walks with dif-
ferent degrees of sinuosity r (crawling behaviour) to strategies
that additionally incorporate both stationarynon-modulated
(omega) and non-stationarysignal-modulated (pirouette) reor-
ientation behaviours To do so we use four behavioural
combinations according to the models in the electronic sup-
plementary material table S2 model 1 crawling behaviour
(correlated random walk) model 2 crawling behaviour with
omegas model 3 crawling behaviour with pirouettes and
model 4 crawling behaviour with both omegas and pirouettes
The details of the modelling framework are provided in the elec-
tronic supplementary material Search efficiency was computed
as the number of encounters per travelled distance averaged
over 500 individuals
0 103
(a)
(c)
(b)
102
s1 = 137
s2 = 046
10
1
10ndash1
5 pause
pirou
omeg
rev
crawls
10
15
20
25
30indi
vidu
als
35
40
45
50
5 10 15time (min)
20 25
10 102
log(t)
log(
MSD
)
103
end start
behavioursreversalsomegas pauses
pirouettes
25 5 mm
Figure 1 Individual worm track and population ethogram (a) A 30 min tracking run showing the centre-of-mass movements of a single C elegans worm Insetshows raw worm image Behavioural events are labelled as crawls (black) reversals (blue) omegas (cyan) pirouettes (orange) and pauses (red) (b) Spreadingcapacity of the worm population measured as the MSD across time (52 worm trajectories starting from the same origin point) Two regimes were found super-diffusive with slope s1 frac14 137 1 (solid line) and subdiffusive with 0 s2 frac14 046 1 (dashed line) The worm population does not passively diffuse throughthe environment Their spreading is accelerated covering a range of spatio-temporal scales up to the limiting scale of the experimental system (51 cm) wherespreading then becomes subdiffusive (c) Population ethogram (n frac14 52) showing individual behavioural variability Behavioural events are colour coded as aboveexcept crawls that are in grey
rsifroyalsocietypublishingorgJRSocInterface
1120131092
4
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
3 ResultsThe tracking system captures both the trajectory and the body
postures of single worms freely moving on the surface of agar
plates (Material and methods) Figure 1a shows representative
high-resolution tracking data and a detailed image of the
worm captured during the experiment The scaling behaviour
of the MSD across time shows that the worms do not passively
diffuse while searching but instead perform more complex
movements such that the population spreading while searching
is superdiffusive with an anomalous diffusion exponent s1 frac14
137 (greater than 1) up to the limiting scale of the experimen-
tal system that can be associated with the crossover towards a
subdiffusive regime s2 frac14 046 (less than 1 figure 1b) The
pairing of large-scale tracks (worm centre-of-mass trajectory)
and small-scale behavioural data (body postures) allows us to
flag crawls and reorientation behavioural events and produce
comprehensive trajectories (figure 1a) and ethograms (figure
1c) that show the transition between a number of crawling
and reorientation behaviours used by worms to explore the
environment Our analysis indicates a number of different
types of crawling motions and reorientation behaviours Crawl-
ing motions are not always straight but often form arcing or
looping trajectories [32] We flagged crawling behaviour
based on curvature and angular concordance [44] into four cat-
egories lines open arcs closed arcs and loops (see the
electronic supplementary material text S2 and figures S2 and
S3) We identified four types of reorientation behaviour
reversals omegas pauses and pirouettes (figure 1c Material
and methods)
The overall movement patterns did not show any direction-
al bias (Rayleigh tests of uniformity [45] applied to the first and
0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

rsifroyalsocietypublishingorgJR
3
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
total it detected 937 reversals 1064 omegas 1125 pirouettes and
387 pauses This method was validated by computing the percen-
tage of behaviours detected by the algorithm to the ones detected
by human observation We used a control dataset of behaviours
observed by eye for five experimental runs (a total of 36 000
frames) representing 10 of the entire dataset The algorithm
detected over 92 (on average) of the number of omegas and rever-
sals that were identified by eye (see the electronic supplementary
material table S1) The 8 misidentified behavioural responses
were due to noisy images in which the worm body could not be
extracted Omegas with low body compressibility (lsquowidersquo omega
bends) and pauses coinciding with a straight body posture were
misidentified by crawls and reversals respectively
SocInterface1120131092
24 Pre-filtering data before analysisIf the worm stopped moving before the completion of the run
then the data were excluded We could see a clear and separable
behavioural effect caused by mechanical stimulus on the reorien-
tation frequency time series for the first 760 frames (32 min)
which we explicitly removed in our final dataset In the end
the following analyses are based on tracks and images of 52 indi-
viduals Tracks were of 268 s each and with a total number
of 2889 reorientation behaviours 788 reversals 860 omegas
935 pirouettes and 306 pauses
25 Orientational memory lossTo study the role of reorientation behaviours on the loss of orien-
tational memory we quantified the effect of each reorientation
type on the overall direction of the trajectory For that we com-
puted (i) the turning angle distribution generated by each
reorientation type and (ii) the angular correlation of the trajec-
tory as a function of reorientation type for each one of the
52 individual worms of our study
Turning angle distribution generated by each reorientation typereorientations are located in between two crawls (periods of
forward motion) The turning angle u generated via a specific reor-
ientation was computed by calculating the difference between the
absolute angles of the vectors following the overall direction of
the crawl just before and just after the reorientation The vectors
are defined using the first and last time frames of each crawl
Angular correlation of the trajectory as a function of reorientation typethe angular correlation function
CaethtTHORN frac141
n
Xn
jfrac141
kcosfrac12aethtthorn tTHORN aethtTHORNlj
where a(t) is the local tangent angle at time t t is the time lag
(from 1 to 13 s) j is the worm id and n is the total number of
worms (52) was calculated from the centre-of-mass data (trajec-
tory) to study the wormrsquos directionality of motion over time We
combined the behavioural data with the trajectory data and
sampled segment types ST from the trajectory that are crawls sep-
arated by a specific type T of reorientation (eg crawlndashreversalndash
crawl) The angular correlation function was computed for both
the original segments ST and their correspondent null model
which is a bootstrapping (499 times) of random segments of the
same length as ST of the same trajectory (see the electronic sup-
plementary material figure S8) The correlation function Ca(t)
was computed for each ST using different lag sizes t and it was
averaged over all the segments over all the 52 individuals (black
line in figure 3) For the null model case standard error bars of
the mean of Ca(t) were computed for each lag size t (dashed line
in figure 3) The difference between the solid lines representing
sequences of crawls interrupted by reorientation types and the
dashed line representing the null model (random segments of
the original trajectory) show how strong the contribution of each
behaviour is towards changes in direction The further the lines
are from each other the larger the contribution of the reorientation
behaviour is evident
26 Probability distribution of omega time inter-eventsWe selected among three probabilistic models of the exponential
family (simple double and stretched exponentials) to identify the
best-fitting probability distribution of the time intervals between
omegas at both the individual and population levels of the 52 indi-
vidual worms In particular we used KolmogorovndashSmirnov (KS)
tests [42] and the likelihood functions of the probability density
functions (pdfs of the three models) over a bounded range and
for pre-binned data [17] We described the three pdfs their corre-
sponding transformation when considering pre-binned data and
their resulting likelihood function in the electronic supplementary
material text S1 For model selection we include only those indi-
viduals that had performed at least 30 omegas during the
tracking period for the individual level analysis
27 Relationship between reorientation and crawl typesTo test the statistical independence between reorientation and
crawl types in our worm population we performed a contingency
table analysis followed by a chi-squared approximation for
proportions [43] We determined both the relationship between
reorientations and crawls occurring immediately after and
between reorientations and crawls that preceded them For the
dependent relationship between reorientations and their previous
crawls we studied in more detail the connection between the
different types of behaviours In particular we performed a two-
tailed Z-score test statistic Z (a frac14 005) [43] to test the null
hypothesis H0 pethCijRjTHORN frac14 pethCiTHORN against the alternative hypothesis
HA pethCijRjTHORN= pethCiTHORN where i and j represent respectively the
different crawl and reorientation types presented in figure 5b
For the behaviours with dependent relationships we used the
departures from the expected frequency between reorientation
events and their previous crawls to determine whether they are
positively (above expected frequency) or negatively (below the
expected frequency) correlated
28 Average time to close a loop and characteristicinter-event times for omegas
The average time to close a loop was calculated as the average time
to close a circle t frac14 2prs where r frac14 1jkj and s is the speed These
averages are taken over all the trajectories that contain loops The
average speed of a loop is 03 mm s21 and the median of loopsrsquo
curvature k is 41 mmndash 1 so t frac14 54 s The characteristic inter-
event time for omegas q frac14 67 s was taken directly from the
stretched exponential likelihood fit at the population level
29 Models of search behaviourInspired by our data analyses we have developed a modelling
framework that allows us to compare the search efficiency in a
patchy landscape of simple correlated random walks with dif-
ferent degrees of sinuosity r (crawling behaviour) to strategies
that additionally incorporate both stationarynon-modulated
(omega) and non-stationarysignal-modulated (pirouette) reor-
ientation behaviours To do so we use four behavioural
combinations according to the models in the electronic sup-
plementary material table S2 model 1 crawling behaviour
(correlated random walk) model 2 crawling behaviour with
omegas model 3 crawling behaviour with pirouettes and
model 4 crawling behaviour with both omegas and pirouettes
The details of the modelling framework are provided in the elec-
tronic supplementary material Search efficiency was computed
as the number of encounters per travelled distance averaged
over 500 individuals
0 103
(a)
(c)
(b)
102
s1 = 137
s2 = 046
10
1
10ndash1
5 pause
pirou
omeg
rev
crawls
10
15
20
25
30indi
vidu
als
35
40
45
50
5 10 15time (min)
20 25
10 102
log(t)
log(
MSD
)
103
end start
behavioursreversalsomegas pauses
pirouettes
25 5 mm
Figure 1 Individual worm track and population ethogram (a) A 30 min tracking run showing the centre-of-mass movements of a single C elegans worm Insetshows raw worm image Behavioural events are labelled as crawls (black) reversals (blue) omegas (cyan) pirouettes (orange) and pauses (red) (b) Spreadingcapacity of the worm population measured as the MSD across time (52 worm trajectories starting from the same origin point) Two regimes were found super-diffusive with slope s1 frac14 137 1 (solid line) and subdiffusive with 0 s2 frac14 046 1 (dashed line) The worm population does not passively diffuse throughthe environment Their spreading is accelerated covering a range of spatio-temporal scales up to the limiting scale of the experimental system (51 cm) wherespreading then becomes subdiffusive (c) Population ethogram (n frac14 52) showing individual behavioural variability Behavioural events are colour coded as aboveexcept crawls that are in grey
rsifroyalsocietypublishingorgJRSocInterface
1120131092
4
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
3 ResultsThe tracking system captures both the trajectory and the body
postures of single worms freely moving on the surface of agar
plates (Material and methods) Figure 1a shows representative
high-resolution tracking data and a detailed image of the
worm captured during the experiment The scaling behaviour
of the MSD across time shows that the worms do not passively
diffuse while searching but instead perform more complex
movements such that the population spreading while searching
is superdiffusive with an anomalous diffusion exponent s1 frac14
137 (greater than 1) up to the limiting scale of the experimen-
tal system that can be associated with the crossover towards a
subdiffusive regime s2 frac14 046 (less than 1 figure 1b) The
pairing of large-scale tracks (worm centre-of-mass trajectory)
and small-scale behavioural data (body postures) allows us to
flag crawls and reorientation behavioural events and produce
comprehensive trajectories (figure 1a) and ethograms (figure
1c) that show the transition between a number of crawling
and reorientation behaviours used by worms to explore the
environment Our analysis indicates a number of different
types of crawling motions and reorientation behaviours Crawl-
ing motions are not always straight but often form arcing or
looping trajectories [32] We flagged crawling behaviour
based on curvature and angular concordance [44] into four cat-
egories lines open arcs closed arcs and loops (see the
electronic supplementary material text S2 and figures S2 and
S3) We identified four types of reorientation behaviour
reversals omegas pauses and pirouettes (figure 1c Material
and methods)
The overall movement patterns did not show any direction-
al bias (Rayleigh tests of uniformity [45] applied to the first and
0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

0 103
(a)
(c)
(b)
102
s1 = 137
s2 = 046
10
1
10ndash1
5 pause
pirou
omeg
rev
crawls
10
15
20
25
30indi
vidu
als
35
40
45
50
5 10 15time (min)
20 25
10 102
log(t)
log(
MSD
)
103
end start
behavioursreversalsomegas pauses
pirouettes
25 5 mm
Figure 1 Individual worm track and population ethogram (a) A 30 min tracking run showing the centre-of-mass movements of a single C elegans worm Insetshows raw worm image Behavioural events are labelled as crawls (black) reversals (blue) omegas (cyan) pirouettes (orange) and pauses (red) (b) Spreadingcapacity of the worm population measured as the MSD across time (52 worm trajectories starting from the same origin point) Two regimes were found super-diffusive with slope s1 frac14 137 1 (solid line) and subdiffusive with 0 s2 frac14 046 1 (dashed line) The worm population does not passively diffuse throughthe environment Their spreading is accelerated covering a range of spatio-temporal scales up to the limiting scale of the experimental system (51 cm) wherespreading then becomes subdiffusive (c) Population ethogram (n frac14 52) showing individual behavioural variability Behavioural events are colour coded as aboveexcept crawls that are in grey
rsifroyalsocietypublishingorgJRSocInterface
1120131092
4
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
3 ResultsThe tracking system captures both the trajectory and the body
postures of single worms freely moving on the surface of agar
plates (Material and methods) Figure 1a shows representative
high-resolution tracking data and a detailed image of the
worm captured during the experiment The scaling behaviour
of the MSD across time shows that the worms do not passively
diffuse while searching but instead perform more complex
movements such that the population spreading while searching
is superdiffusive with an anomalous diffusion exponent s1 frac14
137 (greater than 1) up to the limiting scale of the experimen-
tal system that can be associated with the crossover towards a
subdiffusive regime s2 frac14 046 (less than 1 figure 1b) The
pairing of large-scale tracks (worm centre-of-mass trajectory)
and small-scale behavioural data (body postures) allows us to
flag crawls and reorientation behavioural events and produce
comprehensive trajectories (figure 1a) and ethograms (figure
1c) that show the transition between a number of crawling
and reorientation behaviours used by worms to explore the
environment Our analysis indicates a number of different
types of crawling motions and reorientation behaviours Crawl-
ing motions are not always straight but often form arcing or
looping trajectories [32] We flagged crawling behaviour
based on curvature and angular concordance [44] into four cat-
egories lines open arcs closed arcs and loops (see the
electronic supplementary material text S2 and figures S2 and
S3) We identified four types of reorientation behaviour
reversals omegas pauses and pirouettes (figure 1c Material
and methods)
The overall movement patterns did not show any direction-
al bias (Rayleigh tests of uniformity [45] applied to the first and
0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

0
005
010
015
020
025
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
reversals (n = 788) omegas (n = 860)
0 50 100 150
005
010
015
020
025
turning angle q (deg) turning angle q (deg)
rela
tive
freq
uenc
y of
reo
rien
tatio
ns p
er 1
8deg in
terv
al
pirouettes (n = 935)
0 50 100 150
pauses (n = 306)
(b)(a)
(c) (d )
Figure 2 The frequency distribution of turning angles generated by each reorientation behaviour The frequency distribution of turning angles (188 interval bars)generated by (a) reversals (b) omegas (c) pirouettes and (d ) pauses was computed from the analysis of 52 worms during the approximately 27 min assay periodReversal and omega distributions are close to uniformity The pirouette frequency distribution is centred around large turning angles (90 ndash 1808) values whereaspause frequency distribution is centred around small turning angles (0 ndash 908)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
5
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
last positions of each trajectory p-value frac14 056 n frac14 52) con-
firming the absence of landscape gradients or cue-biased
large-scale movement ie search and not taxis was the main
driver of the overall movement pattern We quantified how
each type of reorientation contributed to the loss of orientation-
al memory by both calculating turning angle distributions and
performing angular correlation analyses of trajectories (figures
2 and 3) Pirouettes generated a distribution centred at 1808turns and accounted for the strongest effect on the loss of orien-
tational memory at the trajectory scale Contrary to what has
been reported for insects [46] pauses generated almost no turn-
ing at all Reversals and omegas generated almost uniformly
distributed turning angles showing a notable effect on orienta-
tional memory loss (figure 3) but not as strong as with
pirouetting Our results show that different types of reorienta-
tions generate different turning angle distributions and break
the directional persistence of the animal to different degrees
suggesting that distinct reorientation strategies may have
different roles within the search process
We further investigated the temporal dynamics of the
different types of reorientations and crawls (figure 4 electronic
supplementary material text S3) We found that the wormrsquos
searching behaviour is a combination of time-dependent and
time-independent components The frequency of certain
types of reorientations (pirouettes and reversals) and crawls
(lines and arcs) decreased through time (Spearmanrsquos cor-
relation range rs [ [21 2058] p-value 005 electronic
supplementary material tables S2 and S3) whereas the fre-
quency of omegas pausing and looping is time-independent
(rs[[-030 020] p-value frac14 031 051 and 099 respectively
electronic supplementary material tables S2 and S3)
For the observation window of our experiments (about
30 min) our results indicate that omegas which lead to uni-
formly distributed turning angle distributions (figure 2)
control for basal time-independent exploratory behaviour
In comparison pirouette and reversals are related to a behav-
ioural or physiological memory that decays through time
Assuming that time-dependent and time-independent reor-
ientations represent two separate behavioural modules we
characterized the inter-event time distribution for omegas to
explore the efficiency of the underlying stationary stochastic
process in promoting encounter success [16] The shape of
the distribution determines whether large but rare inter-
event times are less (simple exponential) or more probable
(double exponential stretched exponential) The latter feature
implies a wide range of crawl lengths and the presence of inten-
siveextensive search patterns We found that the omega
inter-event time distribution is best characterized by a stretched
exponential at the population level (figure 5a Material and
methods electronic supplementary material tables S4
and S5 KS test p-value frac14 059 G-test p-value frac14 092) whereas
the double exponential distribution is the best fit at the individ-
ual level (see the electronic supplementary material figure S4
tables S6 and S7 text S4) Despite the presence of characteristic
times for omegas both the combination of fast and slow omega
turning rates (double exponential [47]) or the presence of a
heavy-tailed time inter-event distribution (stretched exponen-
tial) can promote intensiveextensive search patterns and
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)
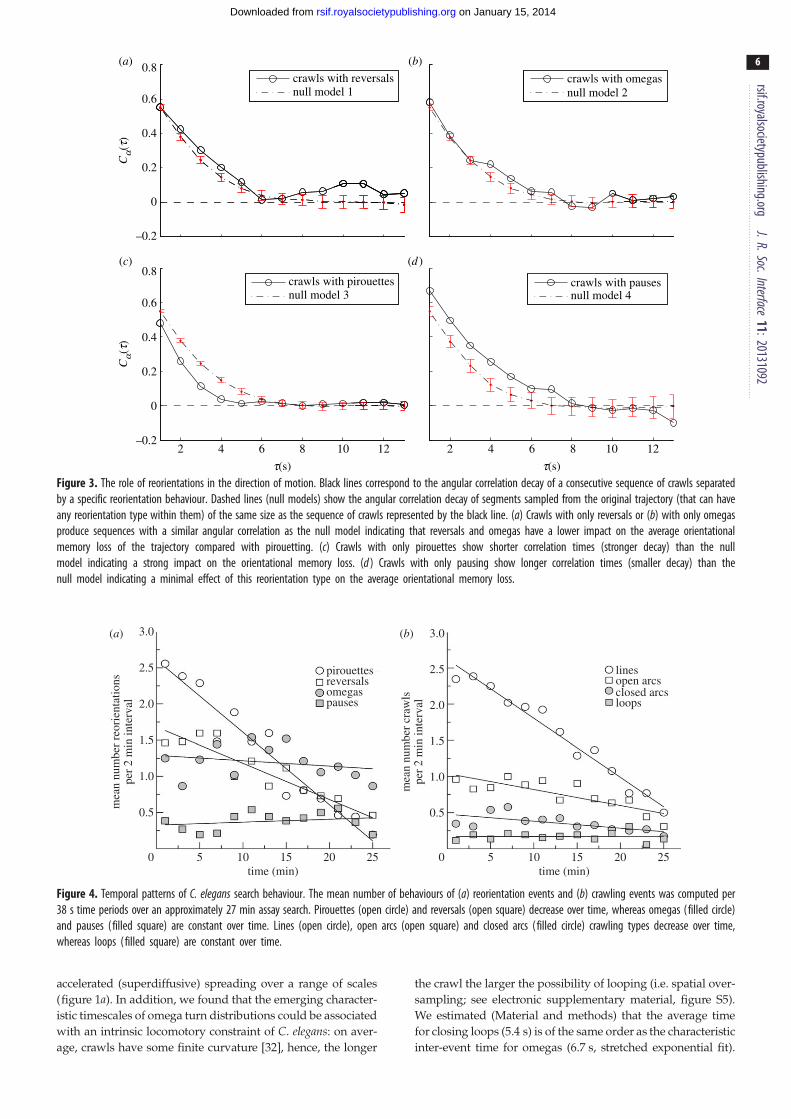
ndash02
0
02
04
06
08(a)
(c)
(b)
(d )
Ca(
t)C
a(t)
crawls with reversalsnull model 1
crawls with omegasnull model 2
2 4 6 8 10 12ndash02
0
02
04
06
08
t(s) t(s)
crawls with pirouettesnull model 3
2 4 6 8 10 12
crawls with pausesnull model 4
Figure 3 The role of reorientations in the direction of motion Black lines correspond to the angular correlation decay of a consecutive sequence of crawls separatedby a specific reorientation behaviour Dashed lines (null models) show the angular correlation decay of segments sampled from the original trajectory (that can haveany reorientation type within them) of the same size as the sequence of crawls represented by the black line (a) Crawls with only reversals or (b) with only omegasproduce sequences with a similar angular correlation as the null model indicating that reversals and omegas have a lower impact on the average orientationalmemory loss of the trajectory compared with pirouetting (c) Crawls with only pirouettes show shorter correlation times (stronger decay) than the nullmodel indicating a strong impact on the orientational memory loss (d ) Crawls with only pausing show longer correlation times (smaller decay) than thenull model indicating a minimal effect of this reorientation type on the average orientational memory loss
mea
n nu
mbe
r re
orie
ntat
ions
pe
r 2
min
inte
rval
mea
n nu
mbe
r cr
awls
pe
r 2
min
inte
rval
30(a) (b)
pirouettesreversalsomegaspauses loops
closed arcsopen arcslines
20
15
10
05
0 5 10 15time (min)
20 25 0 5 10 15time (min)
20 25
25
30
20
15
10
05
25
Figure 4 Temporal patterns of C elegans search behaviour The mean number of behaviours of (a) reorientation events and (b) crawling events was computed per38 s time periods over an approximately 27 min assay search Pirouettes (open circle) and reversals (open square) decrease over time whereas omegas (filled circle)and pauses (filled square) are constant over time Lines (open circle) open arcs (open square) and closed arcs ( filled circle) crawling types decrease over timewhereas loops (filled square) are constant over time
rsifroyalsocietypublishingorgJRSocInterface
1120131092
6
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
accelerated (superdiffusive) spreading over a range of scales
(figure 1a) In addition we found that the emerging character-
istic timescales of omega turn distributions could be associated
with an intrinsic locomotory constraint of C elegans on aver-
age crawls have some finite curvature [32] hence the longer
the crawl the larger the possibility of looping (ie spatial over-
sampling see electronic supplementary material figure S5)
We estimated (Material and methods) that the average time
for closing loops (54 s) is of the same order as the characteristic
inter-event time for omegas (67 s stretched exponential fit)
0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

0
loop rev
arc
line pir
W
1 2 3 4log(t)
ndash3
ndash2
ndash1
0(a)
(b)
P(T
gtt)
datastretcheddoublesimple
Figure 5 Caenorhabditis elegans omega-based search template and interdepen-dencies between reorientation-crawl pairings (a) Probability distribution ofomega inter-events at the population level (52 worms) The fit between theempirical data and the stretched exponential distribution (solid line) is betterthan those obtained with the double exponential (dashed line) and thesimple exponential (dotted line) distributions (b) Loop ndash omega and arc ndashomega pairings are positively correlated (solid arrows) whereas loop ndash pirouettearc ndash reversal and linendash omega pairings are negatively correlated (dashedarrows) The non-existence of an arrow between a reorientation ndash crawl pairingindicates independency between them
rsifroyalsocietypublishingorgJRSocInterface
1120131092
7
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
Hence an important function of omegas in the exploratory
behavioural template may be to avoid spatial oversampling
by preventing excessive looping
To understand better the inherent structure of C eleganssearch patterns we also investigated whether crawling and reor-
ientation behaviours were independent of each other (Material
and methods) We found that reorientations were all indepen-
dent of the following crawl type ( p-value frac14 006 n frac14 1717) but
dependent on the previous crawl type ( p-value frac14 328 1027
n frac14 1714 see the electronic supplementary material figure S6
tables S8 and S9) Omegas were positively correlated with
previous loops ( p-value frac14 0037 n frac14 41) and arcs ( p-value frac14
0004 n frac14 214) and negatively correlated with lines
( p-value frac14 12 1024 n frac14 204) Pirouettes and reversals rarely
finished loops or arcs respectively (figures 5b electronic sup-
plementary material table S10) The hypothesis that omegas
are commonly used to break curved crawls was not only sup-
ported by the latter results but we also found that there was a
significant negative correlation between the number of omegas
in a trajectory and the average length of loops (Spearmanrsquos
rank correlation r frac14 2053 p-value 001)
Our simulations (Material and methods and the electronic
supplementary material) showed that stationary reorienta-
tion templates (omegas) interrupting sinuous movements
at times drawn from stretched exponentials can indeed
improve the search efficiency (figure 6) If signal-modulated
then area-restricted search behaviour is added to the station-
ary reorientation template (pirouettes) then the search
efficiency further improves As expected the search efficiency
improves much more based on reactive (non-stationary) turn-
ing behaviour linked to environmental cues or past memory
than based on non-reactivestationary stochastic reorienta-
tion templates Nonetheless the best search strategy comes
out when combining both types of reorientation behaviours
4 DiscussionCaenorhabditis elegans moves with a limited locomotory reper-
toire but we have shown that in homogeneous information-
limited environments it produces complex and flexible search
behaviours by combining locomotion primitives These basic
movement behaviours are not independently controlled There
are at least first-order correlations between behavioural states
where reorientation events are dependent on the previous crawl-
ing events The movement of many organisms has been
described by a run-and-tumble model [9164648] however
the wormrsquos search movement cannot be represented by such a
strategy [32] for a number of reasons (i) crawl types differ in
their straightness (ii) there are correlations between reorientation
and crawling events (iii) some of the behavioural transitions
are time-dependent and (iv) the stationary components of
behaviour generate complex movement patterns
Omegas are responsible for a stationary multi-scale search
component that holds over 30 min and covers a wide range of
spatial scales These features have been recognized as efficient
random search behaviour that adequately trades-off for
nearby and distant targets in heterogeneous landscapes
[121316] where scale-free reorientation patterns comprise the
limiting optimal case [1516] Multi-scale search templates are
also observed in other organisms [4649ndash51] but there has
been little understanding on what sets these scales Despite
the huge variability of omega turn inter-event times here we
found that C elegans exhibits a characteristic timescale for
omegas connected to intrinsic locomotory constraints such as
not being able to move straight for a long time Worms tend to
end looping trajectories with omegas reducing the risk of self-
crossing paths Examples of other motor behaviours responding
to similar constraints are spiral motions of microorganisms [49]
which are thought to allow the organism to average out locomo-
tory biases and to swim in straighter paths and characteristic
turning frequencies in flagellated bacteria which are constrained
by sampling limits for diffusive sensing [52]
In addition to a stationary and multi-scale search
movement template C elegans also exhibits a non-stationary
080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

080 085 090r
095 100
sear
ch e
ffic
ienc
y (times
10ndash2
)
0
10
10
20
20
30
30
40
40
50(a)
(c)
(b)
50
165
160
155
150
145
140
135
crawling
reorientationclocks
onoff
omegasmulti-scale
search
onoff
model 1model 2model 3model 4
pirouettesarea-restricted
search
Figure 6 Caenorhabditis elegans search modelling framework (a) Conceptual diagram showing the presence of signal-modulated (non-stationary) and non-modulated (stationary) reorientation behaviour and the four potential behavioural combinations implemented Round shapes C elegans behaviours Grey polygonsbehavioural switches (b) Visualization of the path generated by model 4 (crawling behaviour plus omegas frac14 On and pirouettes frac14 On) with sinuosity r frac14 09 ina heterogeneous target landscape Black dotted line trajectory Grey circles targets Blue circles omegas Red circles pirouettes Red-dotted line subsets of thetrajectory influenced by area-restricted-search behaviour (c) Average search efficiency (number of targets found per distance travelled) for different values of direc-tional correlation (r) and the different random walk models Model 1 crawling behaviour (correlated random walk with sinuosity r) Model 2 crawling withomegas Model 3 crawling with pirouettes Model 4 crawling with both omegas and pirouettes
rsifroyalsocietypublishingorgJRSocInterface
1120131092
8
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
adjustable reorientation pattern (pirouettes and reversal turns)
that changes in time in relation to expectations of food location
It is well known that the worm intensifies the search to a
restricted area (high pirouette rates) when food or rewarding
environmental cues are present [2753] Here we show that
well-fed C elegans influenced by past environmental
memory or a present measure of internal state [53] also per-
forms area-restricted search (based on both pirouettes and
reversals) where food encounter expectations are high and gradu-
ally decreases pirouette and reversal rates when it fails to find
food expanding its search range
Previous experiments have revealed that as the memory
of a past resource-rich environment is gradually lost and
cues for new resources are missing animals switch their strat-
egies from intensive to extensive search modes [4654] The
relevance of our results is to show that at least for C elegans
rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

rsifroyalsocietypublishingorgJRSocInterface
1120131092
9
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
the story is not that simple Our results reveal that these
worms constantly maintain in the background a statio-
nary and complex movement template that combines both
intensive and extensive searches and controls for excessive
oversampling An important question remains as to why
such a random search template is needed for the survival
of animals that can learn and use environmental information
for their own profit Learning and memory generate expec-
tations on the environment which undoubtedly animals
use for search and survival However the environment is
noisy and the animalrsquos expectations (their model of the
world) are not always accurate For example in our exper-
iment C elegans performs an intense area-restricted search
in an empty area for about 20 min based on an erroneous
association of past environmental information The worms
expect to find food nearby based on a pre-condition that
does not hold anymore Yet animals have a way to hedge
their bets on their world model by generating an efficient
background search template This background search tem-
plate is an important behavioural module often disregarded
(see [1051]) but may be present in animals to deal with
environmental uncertainty We suggest that such a template
should accommodate motor constraints and incorporate
generalized views on target locations for example the expec-
tation of targets being nearby and faraway from onersquos
position More generally our results suggest that motor
behaviour that is not reinforced by environmental stimuli
(eg taxis) can be constructed on the basis of empirically
grounded expectation reflecting both information learned by
recent individual experiences (flexible and adjustable com-
ponents) or fixed in motor programmes across evolutionary
times (intrinsic less flexible templates)
Based on the above arguments one could make the follow-
ing predictions (i) incorporating complex reorientation patterns
(eg stretched exponential omega turns) should increase
the search efficiency when compared with pure correlated
random walking (pure crawling behaviour) (ii) the impact of
such (omega) reorientation templates on search efficiency
should be larger as we generate more straight-lined crawls
(iii) signal- or memory-modulated reorientations (pirouettes)
should have a much stronger positive effect on search efficiency
than stationary reorientation templates (omegas) (iv) the incor-
poration of both stationary and non-stationary (responding to
memory or to environmental cues) reorientation components
into a search strategy should lead to the best search efficiency
outcome All of these predictions are fulfilled in our simulations
run in patchy landscapes where reorientation mechanisms
become important [1415] Our simulation results depend on
specific parametrizations and should be taken as a qualitative
demonstration of these concepts
Our results show the great potential of studying the motor
mechanisms of C elegans in controlled laboratory environ-
ments to unravel both internal and external drivers of animal
movement behaviour Many of the current (but also classic)
questions about search optimal foraging theory and more
generally movement ecology [23] can be addressed by means
of model organisms Future research on C elegans can uncover
neuronal [26] and genetic [25] mechanisms underlying the
intrinsic stochastic behaviour of organisms as well as quantify
higher correlations between behavioural templates and the
wormrsquos adaptiveness for survival Our results extend beyond
C elegans (see the electronic supplementary material figure S7)
and suggest that in a search process animals perform reorienta-
tion patterns that not only respond to external cues andor
gradients but also are driven by their past memory and stochas-
tic search templates which highlight fundamental principles of
organismsrsquo probabilistic models of the world and how to explore
efficiently in the absence of environmental information
Acknowledgements LCMS was part of the PhD programme in Compu-tational Biology at Gulbenkian Institute of Science Portugal and isgrateful for the financial support from Portuguese Foundation forScience and Technology (FCT) Portugal SFRHBD329602006FB acknowledges the Ramon y Cajal Programme Ministry ofScience and Innovation Spain ref RyC-2009-04133 FB andLCMS work was also supported by Plan Nacional I thorn D thorn i Min-istry of Science and Innovation Spain ref BFU2010-22337 WSRand FB acknowledge the HFSP grant ref RGY00842011 SusanaBernal Katie Hampson and Daniel T Haydon provided valuablefeedback
Data accessibility The C elegans image data are stored in the followingrepository httpwwwryulabca5000fbsharingP1Ncl80x
References
1 Swingland R Greenwood PJ 1984 The ecology ofanimal movement Oxford UK Clarendon Press
2 Bell WJ 1991 Searching behaviour the behaviouralecology of finding resources London UK Chapmanand Hall
3 Stephens DW Brown JS Ydenberg RC 2007Foraging behavior and ecology Chicago ILChicago University Press
4 Schoener TW 1971 Theory of feeding strategiesAnnu Rev Ecol Syst 2 369 ndash 404 (doi101146annureves02110171002101)
5 Green RF 1980 Bayesian birds a simple example ofOatenrsquos stochastic model of optimal foraging TheorPopul Biol 18 244 ndash 256 (doi1010160040-5809(80)90051-9)
6 Heinrich B 1979 Resource heterogeneity andpatterns of movement in foraging bumblebeesOecologia 40 235 ndash 245 (doi101007BF00345321)
7 Weimerskirch H Pinaud D Pawlowski F Bost C-A2007 Does prey capture induce area-restrictedsearch A fine-scale study using GPS in a marinepredator the wandering albatross Am Nat 170734 ndash 743 (doi101086522059)
8 Kioslashrboe T 2008 A mechanistic approach to planktonecology Princeton NJ Princeton University Press
9 Berg HC 1993 Random walks in biology PrincetonNJ Princeton University Press
10 Heisenberg M 2009 Is free will an illusion Nature459 164 ndash 165 (doi101038459164a)
11 Bartumeus F da Luz MGE Viswanathan GM CatalanJ 2005 Animal search strategies a quantitativerandom-walk analysis Ecology 86 3078 ndash 3087(doi10189004-1806)
12 Raposo EP Bartumeus F da Luz MGE Ribeiro-NetoPJ Souza TA Viswanathan GM 2011 Howlandscape heterogeneity frames optimal diffusivity
in searching processes PLoS Comput Biol 7e1002233 (doi101371journalpcbi1002233)
13 Bartumeus F Raposo EP Viswanathan GM da Luz MGE2013 Stochastic optimal foraging theory In Dispersalindividual movement and spatial ecology a mathematicalperspective (eds MA Lewis PK Maini SV Petrovskii)pp 3-32 Berlin Germany Springer ndash Verlag
14 Bartumeus F 2007 Levy processes in animalmovement an evolutionary hypothesis Fractals 15151 ndash 162 (doi101142S0218348X07003460)
15 Bartumeus F Levin SA 2008 Fractal reorientationclocks linking animal behavior to statistical patternsof search Proc Natl Acad Sci USA 105 19 072 ndash19 077 (doi101073pnas0801926105)
16 Viswanathan GM Buldyrev SV Havlin S da Luz MGRaposo EP Stanley HE 1999 Optimizing the successof random searches Nature 401 911 ndash 914 (doi10103844831)
rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)

rsifroyalsocietypublishingorgJRSocInterface
1120131092
10
on January 15 2014rsifroyalsocietypublishingorgDownloaded from
17 Edwards AM et al 2007 Revisiting Levy flight searchpatterns of wandering albatrosses bumblebees anddeer Nature 449 1044 ndash 1048 (doi101038nature06199)
18 Sims DW et al 2008 Scaling laws of marinepredator search behaviour Nature 4511098 ndash 1102 (doi101038nature06518)
19 Humphries NE et al 2010 Environmental contextexplains Levy and Brownian movement patternsof marine predators Nature 465 1066 ndash 1069(doi101038nature09116)
20 Humphries NE Weimerskirch H Queiroz NSouthall EJ Sims DW 2012 Foraging success ofbiological Levy flights recorded in situ Proc NatlAcad Sci USA 109 7169 ndash 7174 (doi101073pnas1121201109)
21 Viswanathan GM Luz MGE Raposo EP Stanley EH2011 The physics of foraging an introduction torandom searches and biological encountersCambridge UK Cambridge University Press
22 Benichou O Loverdo C Moreau M Voituriez R 2011Intermittent search strategies Rev Mod Phys 8381 ndash 129 (doi101103RevModPhys8381)
23 Nathan R Getz WM Revilla E Holyoak M KadmonR Saltz D Smouse PE 2008 A movement ecologyparadigm for unifying organismal movementresearch Proc Natl Acad Sci USA 105 19 052 ndash19 059 (doi101073pnas0800375105)
24 Morales JM Ellner SP 2002 Scaling up animalmovements in heterogeneous landscapes theimportance of behavior Ecology 83 2240 ndash 2247(doi1018900012-9658(2002)083[2240SUAMIH]20CO2)
25 Bargmann CI 1993 Genetic and cellular analysisof behavior in C elegans Annu Rev Neurosci 1647 ndash 71 (doi101146annurevne16030193000403)
26 De Bono M Maricq AV 2005 Neuronal substrates ofcomplex behaviors in C elegans Annu RevNeurosci 28 451 ndash 501 (doi101146annurevneuro27070203144259)
27 Gray JM Hill JJ Bargmann CI 2005 A circuit fornavigation in Caenorhabditis elegans Proc NatlAcad Sci USA 102 3184 ndash 3191 (doi101073pnas0409009101)
28 Mori I 1999 Genetics of chemotaxis andthermotaxis in the nematode Caenorhabditiselegans Annu Rev Genet 33 399 ndash 422 (doi101146annurevgenet331399)
29 Croll NA 1975 Components and patterns in thebehaviour of the nematode C elegans J Zool 176159 ndash 176 (doi101111j1469-79981975tb03191x)
30 Pierce-Shimomura JT Morse TM Lockery SR 1999The fundamental role of pirouettes inCaenorhabditis elegans chemotaxis J Neurosci 199557 ndash 9569
31 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2008 Dimensionality and dynamics in the behaviorof C elegans PLoS Comput Biol 4 e1000028(doi101371journalpcbi1000028)
32 Stephens GJ Johnson-Kerner B Bialek W Ryu WS2010 From modes to movement in the behavior ofCaenorhabditis elegans PLoS ONE 5 e13914(doi101371journalpone0013914)
33 Williams PL Dusenbery DB 1990 A promisingindicator of neurobehavioral toxicity using thenematode Caenorhabditis elegans and computertracking Toxicol Ind Health 6 425 ndash 440 (doi101177074823379000600306)
34 Cronin CJ Mendel JE Mukhtar S Kim Y-M StirblRC Bruck J Sternberg PW 2005 An automatedsystem for measuring parameters of nematodesinusoidal movement BMC Genet 6 5 (doi1011861471-2156-6-5)
35 Wang SJ Wang Z-W 2013 Track-a-worm an open-source system for quantitative assessment ofC elegans locomotory and bending behavior PLoSONE 8 e69653 (doi101371journalpone0069653)
36 Peliti M Chuang JS Shaham S 2013 Directionallocomotion of C elegans in the absence of externalstimuli PLoS ONE 8 e78535 (doi101371journalpone0078535)
37 Geng W Cosman P Baek J-H Berry CC Schafer WR2003 Quantitative classification and naturalclustering of Caenorhabditis elegans behavioralphenotypes Genetics 165 1117 ndash 1126
38 Cronin CJ Feng Z Schafer WR 2006 Automatedimaging of C elegans behavior MethodsMol Biol 351 241 ndash 251 (doi1013851-59745-151-7241)
39 Hoshi K Shingai R 2006 Computer-drivenautomatic identification of locomotion states inCaenorhabditis elegans J Neurosci Methods 157355 ndash 363 (doi101016jjneumeth200605002)
40 Huang K-M Cosman P Schafer WR 2006 Machinevision based detection of omega bends and
reversals in C elegans J Neurosci Methods 158323 ndash 336 (doi101016jjneumeth200606007)
41 Likitlersuang J Stephens G Palanski K Ryu WS2012 C elegans tracking and behavioralmeasurement J Vis Exp 69 e4094 (doi1037914094)
42 Clauset A Shalizi CR Newman MEJ 2009 Power-law distributions in empirical data SIAM Rev 51661 ndash 703 (doi101137070710111)
43 Zar JH 2010 Biostatistical analysis 5th edn NewJersey NJ Prentice-Hall Inc
44 Fortin M-J Dale MRT 2005 Spatial analysis a guide forecologists Cambridge UK Cambridge University Press
45 Batschelet E 1981 Circular statistics in biologyNew York NY Academic Press
46 Bazazi S Bartumeus F Hale JJ Couzin ID 2012Intermittent motion in desert locusts behaviouralcomplexity in simple environments PLoS Comput Biol8 e1002498 (doi101371journalpcbi1002498)
47 Srivastava N Clark DA Samuel ADT 2009 Temporalanalysis of stochastic turning behavior of swimmingC elegans J Neurophysiol 102 1172 ndash 1179(doi101152jn909522008)
48 Turchin P 1998 Quantitative analysis of movementmeasuring and modeling population redistribution inanimals and plants Sunderland MA SinauerAssociates
49 Jennings HS 1901 On the significance of the spiralswimming of organisms Am Nat 35 369 ndash 378(doi101086277922)
50 Korobkova E Emonet T Vilar JMG Shimizu TSCluzel P 2004 From molecular noise to behaviouralvariability in a single bacterium Nature 428574 ndash 578 (doi101038nature02404)
51 Brembs B 2011 Towards a scientific concept of freewill as a biological trait spontaneous actions anddecision-making in invertebrates Proc R Soc B278 930 ndash 939 (doi101098rspb20102325)
52 Berg HC Purcell EM 1977 Physics ofchemoreception Biophys J 29 193 ndash 219 (doi101016S0006-3495(77)85544-6)
53 Hills T Brockie PJ Maricq AV 2004 Dopamine andglutamate control area-restricted search behavior inCaenorhabditis elegans J Neurosci 24 1217 ndash 1225(doi101523JNEUROSCI1569-032004)
54 Kraemer PJ Golding JM 1997 Adaptive forgettingin animals Psychon Bull Rev 4 480 ndash 491 (doi103758BF03214337)




















