
University of Groningen Alpha-glucan acting enzymes in ...
169
University of Groningen Alpha-glucan acting enzymes in Aspergillus niger Kaaij, Rachel Maria van der IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2007 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Kaaij, R. M. V. D. (2007). Alpha-glucan acting enzymes in Aspergillus niger: diversity in enzymatic activities and functions. s.n. Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license. More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne- amendment. Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 13-07-2022
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of University of Groningen Alpha-glucan acting enzymes in ...

University of Groningen
Alpha-glucan acting enzymes in Aspergillus nigerKaaij, Rachel Maria van der
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2007
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Kaaij, R. M. V. D. (2007). Alpha-glucan acting enzymes in Aspergillus niger: diversity in enzymatic activitiesand functions. s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license.More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne-amendment.
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 13-07-2022

Alpha-glucan acting enzymes in Aspergillus niger Diversity in enzymatic activities and functions
Rachel van der Kaaij

R. M. van der Kaaij Alpha-glucan acting enzymes in Aspergillus niger: Diversity in enzymatic activities and functions ISBN 978-90-367-3175-1 Cover design : Ernst Arbouw Printed by: PrintPartners Ipskamp, Enschede The author gratefully acknowledges financial support from the Groningen Biomolecular Sciences and Biotechnology Institute and TNO Food and Biotechnology Innovations for the printing of this thesis.

RIJKSUNIVERSITEIT GRONINGEN
Alpha-glucan acting enzymes in Aspergillus niger Diversity in enzymatic activities and functions
Proefschrift
ter verkrijging van het doctoraat in de Wiskunde en Natuurwetenschappen aan de Rijksuniversiteit Groningen
op gezag van de Rector Magnificus, dr. F. Zwarts, in het openbaar te verdedigen op
vrijdag 12 oktober 2007 om 13:15 uur
door
Rachel Maria van der Kaaij geboren op 21 juli 1976
te Nijmegen

Promotores: Prof. dr. L. Dijkhuizen Prof. dr. M.J.E.C. van der Maarel Beoordelingscommissie: Prof. dr. B.W. Dijkstra Prof. dr. A.J. Driessen Prof. dr. J. Hille

Contents
Chapter 1 Introduction 7 Chapter 2 Aspergillus niger genome wide analysis reveals a large number of novel α-glucan acting enzymes with unexpected expression profiles 25 Chapter 3 Characterization of two novel, putatively cell wall associated and GPI-anchored, α-glucanotransferase enzymes of Aspergillus niger 61 Chapter 4 Biochemical characterization of two GPI-anchored α-glucanotransferase enzymes from Aspergillus niger reveals a novel reaction specificity in subfamily GH13_1 of fungal α-amylases 85 Chapter 5 Phylogenetic and biochemical characterization of a novel cluster of intracellular fungal α-amylase enzymes 99 Chapter 6 Summary and concluding remarks 121 Reference list 133 Nederlandse Samenvatting 143 List of publications 153 Dankwoord 155 Appendix 157

Abbreviations Agd Alpha-glucosidase Ags Alpha-glucan synthase Agt Alpha-glucanotransferase Amy Alpha-amylase CFW Calcofluor White CGTase Cyclodextrin glucanotransferase DMSO Dimethylsulfoxide DP Degree of polymerization EDTA Ethylenediamine tetraacetate ER Endoplasmic reticulum GH Glycoside Hydrolase GPI Glycosylphosphatidylinositol GT Glycosyltransferase HMM Hidden Markov Model kDa kilo Dalton PIR Protein with internal repeat SBD Starch binding domain SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis TLC Thin layer chromatography UDP Uridine 5’-diphosphate

Chapter 1 Introduction

Alpha-glucan acting enzymes in Aspergillus niger
8
The kingdom of fungi Fungi form a large and tremendously variable group of eukaryotic micro organisms. Historically, they have long been classified in the kingdom of plants, until it was realized that the life style of plants and fungi is completely different. Plants are phototrophs, utilizing energy from light, while fungi are chemotrophs, obtaining energy from the degradation of organic materials. Likewise, autotrophic plants synthesize their organic components from atmospheric carbon dioxide, while fungi are heterotrophs and need organic material as their carbon source. Consequently, fungi play an important role in the recycling of carbon and nitrogen, and they often have a saprophytic or symbiotic life style. Fungi can be opportunistic or obligate pathogens, causing many diseases in plants and animals. In agriculture, fungi are one of the main causes of crop destruction, either by decreasing yields or by the production of toxins in crops rendering them poisonous (Carlile and Watkinson, 1994). On the other hand, many mutualistic relationships exist between fungi and other organisms. Fungi forming arbuscular mycorrhiza are obligate symbionts of plants and belong to the order Glomales. They obtain their carbon and energy sources directly from the host plant, via the colonisation of plant roots. Almost all plant species are colonized by mycorrhizal fungi and benefit from the increased potential of taking up minerals and water, which has a large impact on crop yield (Smith and Read, 1997). Another example of an intimate symbiotic relationship of fungi and plants exists in the lichens, with lichen-forming fungi acquiring their carbon source from extracellularly located green algae or cyanobacteria (Honegger, 2001). Some insects, such as tropical ant species, make use of the ability of fungi to degrade plant material by growing them in “gardens” in their nests (Silva et al., 2003). In human culture, several yeast and fungal species have a prominent role in the production of food. Agaricus bisporus (common mushroom) and other edible mushroom species are grown on large scale for human consumption. The yeast Saccharomyces cerevisiae (Baker’s yeast) is used in bread baking and brewing, and Aspergillus oryzae has been used for the production of Japanese soy sauce and miso (fermented soy beans) since a long time (Hesseltine, 1983). Examples of more recent inventions of applications for fungi are the production of antibiotics by e.g. Penicillium species, and the use of Fusarium venenatum as a producer of food with a high protein content (Quorn) used as an alternative for meat. Most fungal species described to date can be classified into two phyla: the Basidiomycetes and the Ascomycetes. Basidiomycetes comprise for example the mushroom fungi and the rusts, which are primarily obligate plant pathogens. Ustilago maydis, the causal agent of corn smut disease in maize, and Cryptococcus neoformans,

Chapter 1 Introduction
9
an opportunistic human pathogen, are examples of Basidiomycetes of which the genomic sequences are available. The Ascomycetes can be divided over three subphyla: Taphrinomycotina, Saccharomycotina and Pezizomycotinia. The Taphrinomycotina form a diverse group of yeast-like and filamentous fungi, of which Schizosaccharomyces pombe (also called fission yeast) is an often used model species. The subphylum Saccharomycotina consists of the ‘true yeasts’, including S. cerevisiae and the human pathogen Candida albicans. The largest subphylum is formed by the Pezizomycotina comprising mainly filamentous fungi producing fruiting bodies. Among the nine classes of the Pezizomycotina are the Sordariomycetes, including Neurospora crassa and Magnaporthe grisea, and the Eurotiomycetes which include Histoplasma capsulatum and the aspergilli.
The aspergilli The filamentous fungi of the genus Aspergillus belong to the Ascomycetes. Up to now, 274 different species have been described (NCBI website, June 2007). The genus was first catalogued in 1729 by the Italian biologist and priest Pietro Antonio Micheli (1679-1753), who named it after the aspergillum, a brush used for the sprinkling of holy water (Micheli, 1729). The asexual spores or conidiospores of aspergilli form long rows diverting from the conidiophore which, observed under a microscope, resemble the hairs of a brush (Fig. 1). Aspergilli occur worldwide, mostly as saprophytic, soil borne fungi. Some species, such as Aspergillus fumigatus and Aspergillus flavus are opportunistic pathogens for humans and other higher animals. Most human systemic infections are found among immunocompromised patients. Other species, such as Aspergillus niger, are well known for their usage in food production and modern biotechnology.
Aspergillus niger A. niger is a common soil fungus, sometimes observed as a black mould spoiling fruit and other foods. Although this species is generally non-pathogenic, inhalation of large amounts of spores can lead to the lung disease aspergillosis (Schuster et al., 2002). Oral intake of A. niger has been assessed as harmless by the World Health Organization, which opened the opportunity to use this versatile producer for industrial production of acids, pharmaceuticals and enzymes. Various strains of A. niger are applied in the large scale industrial preparation of citric acid (also known as food additive E330) and gluconic acid (E574), which serve as ingredients for the production of various foods and

Alpha-glucan acting enzymes in Aspergillus niger
10
drinks (Lasure and Magnuson, 2004;Mourya and Jauhri, 2000). A. niger is also a very efficient producer of enzymes.
Fig. 1 Aspergillus species seen under a microscope. (Courtesy CBS Fungal Biodiversity Centre). The most widely used A. niger enzyme is glucoamylase, which is produced by selected industrial strains overproducing this enzyme, e.g. strain A. niger DS03043 (van Dijck et al., 2003). Glucoamylase is nowadays produced and sold by several companies under brand names like Stargen and Plusweet (from Genencor) and Fungamyl (Novozymes). These commercial preparations often combine glucoamylase with α-amylase for the release of glucose from starch granules, which can subsequently be converted into fructose by glucose isomerase yielding high fructose syrups (van der Maarel et al., 2002). Also, cocktails of glucoamylase in combination with α-amylases or glucose oxidases are applied to improve dough structure in bread baking and to improve the process of
brewing. Aspergilli, including A. niger, are also known for the production of various types of pectinases, which are used for the clarification of wine and fruit juices and production of fruit purees (Schauer and Boriss, 2004). Cellulases produced by aspergilli are in use in washing detergents to reduce pilling, and in the brewing industry. An example of a commercially produced cellulase from A. niger is Finizym (Novo Nordisk). A niger is not only deployed in the production of its own proteins, it is also a host for the heterologous expression of proteins from various eukaryotes, including those originating from mammals, with mainly applications in the pharmaceutical industry (Yoder and Lehmbeck, 2004).
A. niger is one of the main host organisms for the expression and production of enzymes by the Dutch biotechnology company DSM. To improve this process and

Chapter 1 Introduction
11
search for novel enzyme activities, the genome was sequenced by Biomax under the authority of DSM in 2002 (Pel et al., 2007). Since that time, the genome sequences of several other Aspergillus species have also become available. This has allowed us to compare the different species with regard to gene organization as well as sequence features of specific genes of interest. In this thesis, the focus will be on new A. niger enzymes with similarity to enzymes involved in the degradation of one of its main carbon sources: starch.
Starch and glycogen Starch is one of the most abundantly available carbon sources for a saprophytic fungus like A. niger. It is the main storage polysaccharide in plants, where it can be stored in the roots, tubers, seeds and fruits (Peters, 2006). Starch also constitutes a major component of the human diet. Rice, wheat, maize and potato are examples of plants with high starch content often used in human consumption. Products derived from starch are used for various applications in the food industry, for example as a thickener in sauces and soups or to change the texture of dairy products, but also in the production of paper and biodegradable plastics. Starch is composed of two types of molecules: amylose and amylopectin. Amylose is an unbranched chain of α-(1,4) linked glucose residues, which generally has a degree of polymerization (DP) between 250 and 5000. Amylopectin is also mainly composed of α-(1,4)-linked glucose residues, but approximately 3-5% of these glucose moieties is additionally linked with an α-(1,6) bond, creating a branched molecule of DP 10,000-100,000 (Fig. 2). In general, starch is composed of 15-30% amylose and 70-85% amylopectin, but the exact composition depends on the source (Robyt, 1998). Starch in plants is produced from ADP-glucose formed as a result of photosynthesis (Preiss, 2004). The enzyme starch synthase couples the ADP-glucose to an existing starch molecule via an α-(1,4) glycosidic bond, thereby releasing ADP. Alternatively, a new starch molecule may be formed via a glycosylated protein acting as a primer for the elongation reaction by a starch synthase. Subsequently, α-(1,6) branches are formed in the growing glucan chains by starch branching enzymes. Starch synthesis takes place in the chloroplast or, in non-photosynthetic storage tissues, in the amyloplast.

Alpha-glucan acting enzymes in Aspergillus niger
12
Fig. 2 Part of amylopectin or glycogen, showing four α-(1,4) linked glucose residues and one α-(1,6) linked glucose residue forming a branching point. Glycogen is the equivalent polysaccharide storage compound in bacteria, fungi and animals. Glycogen is a highly branched molecule, with between 7 and 12% α-(1,6) linked glucose molecules. It exists in dissolved form in cells and is therefore faster accessible for degradation than starch in plants, which is normally stored in a granular form. In analogy of the synthesis route of starch, glycogen is produced from UDP-glucose via three enzymes: (1) glycogenin, a self-glycosylating enzyme acting as a starter molecule, (2) glycogen synthase which elongates the chain with α-(1,4) linked glucose residues, and (3) a branching enzyme which incorporates the α-(1,6) glycosidic bonds. The degradation of glycogen is performed by another set of specialized enzymes. Glycogen phosphorylase releases glucose-1-phosphate but cannot act near branching points. Here, a debranching enzyme first transfers a maltosyl unit from the branching point to the end of a linear α-(1,4) chain and subsequently releases glucose by hydrolysing the remaining α-(1,6) linkage (Francois and Parrou, 2001).
Enzymes acting on starch Degradation of starch by heterotrophic organisms is performed by the concerted action of different enzymes, including α-amylases, α-glucosidases and glucoamylases. Αlpha-amylases hydrolyse the internal α-(1,4)-glycosidic bonds in starch, glycogen and maltooligosaccharides (α-(1,4) linked glucose oligomers) producing shorter maltooligosaccharides and maltose (4-O-α-D glucopyranosyl-D-glucose) (EC 3.2.1.1). A special type of amylases, the β-amylases, specifically release maltose from the reducing end of a substrate and are only found in bacteria and in plants. Glucoamylases

Chapter 1 Introduction
13
release β-glucose from the non-reducing end of maltooligosaccharides (EC 3.2.1.3), while α-glucosidase releases α-glucose from the non-reducing end (EC 3.2.1.20).
The enzymes performing these hydrolysis reactions on starch and related substrates are classified in different families according to the classification of glycoside hydrolase (GH) enzymes, which is based on sequence similarity of the proteins (see http://www.cazy.org) (Henrissat, 1991;Henrissat and Bairoch, 1993). Glucoamylases are grouped in family GH15, and most α-glucosidases group in family GH31, together with a few enzymes with different substrate specificities such as α-xylosidase and α-(1,3) glucosidase. The β-amylases belong to family GH14, while the majority of the α-amylases belong to family GH13. Some bacterial and archaeal enzymes with α-amylase activity have no apparent sequence similarity to GH13 enzymes and are placed in their own family GH57 (Henrissat and Bairoch, 1996).
Family GH13 enzymes The GH13 family is the largest sequence based glycoside hydrolase family. It contains more than 30 different enzymatic specificities, and proteins originating from all kingdoms of life. Most enzymes in this family either hydrolyse or form α-(1,4) or α-(1,6) glycosidic bonds. Additionally, some enzymes act on sucrose (β-D-fructofuranosyl α-D-glucopyranoside, with a β-(1,2) glycosidic bond) or trehalose (α-D-glucopyranosyl-α-D-glucopyranoside, with an α-(1,1) glycosidic bond). To improve the predictability of the enzymatic activity of family GH13 members, the family was recently divided into 35 subfamilies, which were assigned based on the phylogenetic analysis of a large number of protein sequences (Stam et al., 2006). Most of these subfamilies contain only one enzymatic activity, while several enzyme specificities, notably α-amylases, are divided over a number of subfamilies. For example, extracellular fungal α-amylases are grouped in family GH13_1, while α-amylases from animals belong to subfamily GH13_24, and cyclodextrin glucanotransferases cluster in subfamily GH13_2 (Table 1). In general, family GH13 enzymes can be recognised by the presence of four highly conserved amino acid regions, first identified by Nakajima et al. (Nakajima et al., 1986) which contain most of the amino acids present in the active site (MacGregor et al., 2001). Within these regions, 7 amino acids important for catalytic activity and binding and positioning of the substrate are generally conserved (Table 1). These include the two catalytic residues Asp227 and Glu251 (numbering according to acid amylase from A. niger, unless indicated otherwise). The third completely conserved amino acid is Asp318, which plays an important role in substrate binding and transition state stabilization. Mutation of any of these three residues in family GH13 enzymes resulted in a dramatic decrease of the enzymatic activity or rendered the enzymes completely
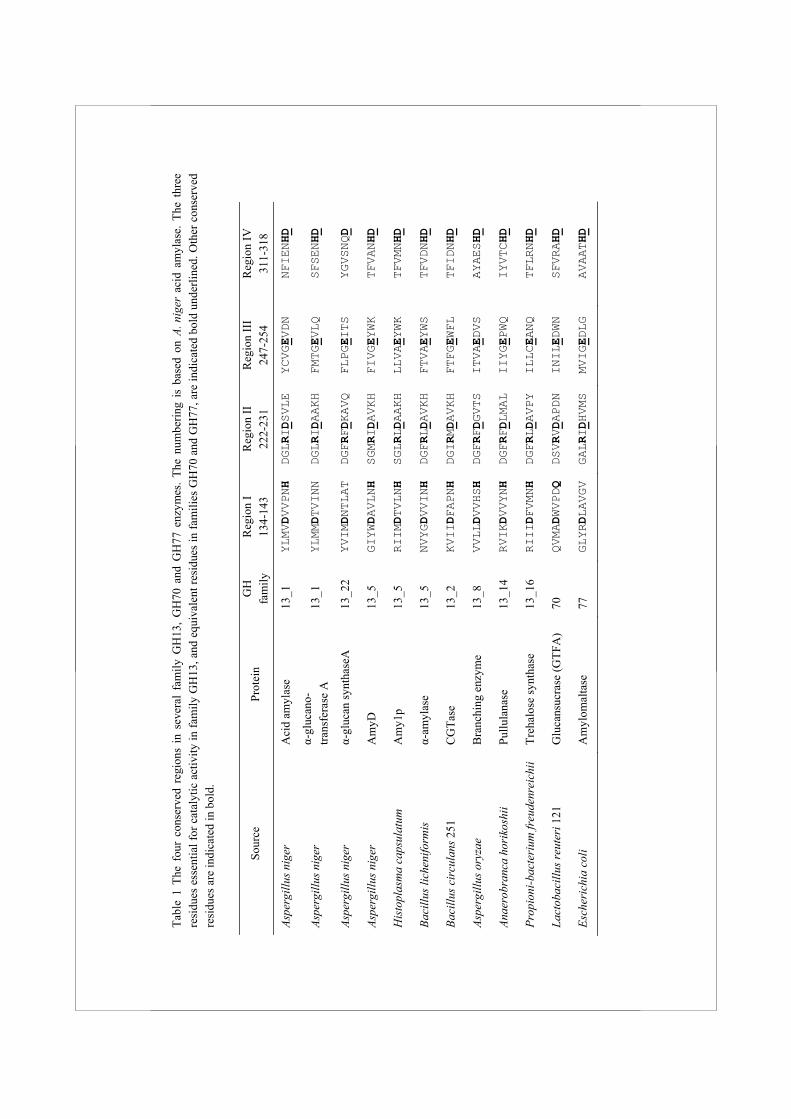
Tabl
e 1
The
four
con
serv
ed r
egio
ns i
n se
vera
l fa
mily
GH
13, G
H70
and
GH
77 e
nzym
es. T
he n
umbe
ring
is b
ased
on
A. n
iger
aci
d am
ylas
e. T
he t
hree
re
sidu
es e
ssen
tial f
or c
atal
ytic
act
ivity
in fa
mily
GH
13, a
nd e
quiv
alen
t res
idue
s in
fam
ilies
GH
70 a
nd G
H77
, are
indi
cate
d bo
ld u
nder
lined
. Oth
er c
onse
rved
re
sidu
es a
re in
dica
ted
in b
old.
Sour
ce
Prot
ein
GH
fa
mily
R
egio
n I
134-
143
Reg
ion
II
222-
231
Reg
ion
III
247-
254
Reg
ion
IV
311-
318
Aspe
rgill
us n
iger
A
cid
amyl
ase
13_1
YLMVDVVPNH
DGLRIDSVLE
YCVGEVDN
NFIENHD
Aspe
rgill
us n
iger
α-
gluc
ano-
tra
nsfe
rase
A
13_1
YLMMDTVINN
DGLRIDAAKH
FMTGEVLQ
SFSENHD
Aspe
rgill
us n
iger
α-
gluc
an sy
ntha
seA
13
_22
YVIMDNTLAT
DGFRFDKAVQ
FLPGEITS
YGVSNQD
Aspe
rgill
us n
iger
A
myD
13
_5
GIYWDAVLNH
SGMRIDAVKH
FIVGEYWK
TFVANHD
His
topl
asm
a ca
psul
atum
A
my1
p 13
_5
RIIMDTVLNH
SGLRLDAAKH
LLVAEYWK
TFVMNHD
Baci
llus l
iche
nifo
rmis
α-
amyl
ase
13_5
NVYGDVVINH
DGFRLDAVKH
FTVAEYWS
TFVDNHD
Baci
llus c
ircu
lans
251
C
GTa
se
13_2
KVIIDFAPNH
DGIRMDAVKH
FTFGEWFL
TFIDNHD
Aspe
rgill
us o
ryza
e B
ranc
hing
enz
yme
13_8
VVLLDVVHSH
DGFRFDGVTS
ITVAEDVS
AYAESHD
Anae
robr
anca
hor
ikos
hii
Pullu
lana
se
13_1
4 RVIKDVVYNH
DGFRFDLMAL
IIYGEPWQ
IYVTCHD
Prop
ioni
-bac
teri
um fr
eude
nrei
chii
Treh
alos
e sy
ntha
se
13_1
6 RIIIDFVMNH
DGFRLDAVPY
ILLCEANQ
TFLRNHD
Lact
obac
illus
reut
eri 1
21
Glu
cans
ucra
se (G
TFA
) 70
QVMADWVPDQ
DSVRVDAPDN
INILEDWN
SFVRAHD
Esch
eric
hia
coli
Am
ylom
alta
se
77
GLYRDLAVGV
GALRIDHVMS
MVIGEDLG
AVAATHD

Chapter 1 Introduction
15
inactive (Klein et al., 1992;Sogaard et al., 1993). The other conserved residues, as displayed in Table 1, often play a more indirect role in catalysis, for example in binding and positioning of the substrate (Leemhuis et al., 2002;Van der Veen et al., 2000). The reaction mechanism of family GH13 enzymes is described as a double replacement mechanism (McCarter and Withers, 1994;Uitdehaag et al., 1999). The substrate is bound in the active site with its non-reducing end in the minus subsites. In enzymes with glucanotransferase activity, this implies that the part of the donor substrate that is transferred is bound in the minus subsites, while the acceptor substrate is bound in the plus subsites (Davies et al., 1997). The actual reaction proceeds in two steps: first, the proton donor (Glu251) protonates the oxygen atom in the scissile bond, thereby cleaving the substrate. The remaining part of the substrate forms a covalent intermediate via a β-glycosidic linkage with the nucleophile (Asp227), and the leaving group departs from the plus subsite. The existence of a covalent intermediate has been demonstrated for different enzymes, including an α-amylase (Tao et al., 1989) and a CGTase (Mosi et al., 1997;Uitdehaag et al., 1999). In the second step, an acceptor molecule is deprotonated by Glu251, which is now acting as a general base, and attacks the β-glycosidic linkage, thereby forming a new α-glycosidic linkage. The acceptor molecule may be water, in the case of hydrolysis, or another saccharide molecule in case of a transglycosylation reaction (Fig. 3). The 3D structure of the enzymes in family GH13 is characterized by the presence of several domains (Jespersen et al., 1991). The main part of the proteins, domain A, is a (β/α)8 barrel or TIM-barrel, a symmetrical fold composed of 8 β-strands surrounded by 8 α-helices. The active site is located in the loops at the C-termini of several of the β-strands. In addition to domain A, all family GH13 enzymes contain a loop protruding between the third β-strand and the third α-helix of the (β/α)8 barrel called domain B (Janecek et al., 1997). Domain B is thought to be involved in the formation of the substrate binding cleft, and its composition and 3D structure can be highly variable between the different GH13 subfamilies. Studies on highly homologous barley α-amylases with different B-domains have shown the importance of this domain in substrate specificity and stability at low pH (Rodenburg et al., 1994;Juge et al., 1995). A variety of other domains may be present in family GH13 enzymes, depending on their enzymatic reaction and substrate specificities. For example, α-amylases (EC 3.2.1.1) often contain an additional C-terminal domain C which has an antiparallel β-sandwich fold (Jespersen et al., 1991). Cyclodextrin glucanotransferases contain two extra domains following the C-domain: domain D has an unknown function, while domain E is a starch binding domain (SBD) (Penninga et al., 1996). Such a SBD, which binds strongly to granular starch and cyclodextrins, belongs in Carbohydrate Binding Module family 20 (CBM20) (Knegtel et al., 1995). Some α-amylases and most glucoamylases

Alpha-glucan acting enzymes in Aspergillus niger
16
from family GH15 also contain a domain from family CBM20 (see e.g. Iefuji et al. (1996) and Svensson et al. (1986)).
Fig. 3 Reaction mechanism of family GH13 enzymes. This example shows the hydrolysis of an α-(1,4) glycosidic linkage. The reaction starts with the binding of the substrate and the formation of a covalent intermediate with the catalytic nucleophile. Subsequently, the acceptor molecule (in this case water) is activated by the acid/base catalyst and the product is formed. In case of a transglycosylation reaction, a sugar is used as acceptor substrate and a new glycosidic linkage is formed. The figure has been adapted from Uitdehaag et al. (1999). Family GH13 enzymes share their (β/α)8 barrel structure and their reaction mechanism with the families GH70 and GH77. Together, these three families form the α-amylase superfamily, or clan GH-H. The members of family GH70 are glucansucrases, large bacterial enzymes which use sucrose to form long α-glucan polymers with different

Chapter 1 Introduction
17
types of glycosidic linkages (EC 2.4.1.5 and 2.4.1.140) (Monchois et al., 1999). The (β/α)8 barrel of GH70 enzymes is thought to be circularly permuted compared to GH13 enzymes, and several additional domains with unknown function are present (MacGregor et al., 1996;van Hijum et al., 2006).Family GH77 contains amylomaltase enzymes from bacteria and archaea, including many thermostable examples with industrial applications (Kaper et al., 2004). These intracellular enzymes have disproportionating activity and transfer α-(1,4) linked glucans to an acceptor substrate, which may be starch, maltooligosaccharides or glucose (EC 2.1.4.25). The enzymes in the families GH13, GH70 and GH77 share their catalytic machinery, implicating that the amino acids involved in catalysis are largely conserved. However, there are also several differences within the four conserved regions of these families. For example, His143 is almost completely conserved in family GH13, and several studies have shown that mutation of this residue can have a strong effect on catalytic activity or reaction specificity of the enzyme (Chang et al., 2003;Nakamura et al., 1993). The equivalent of this His is, however, replaced by a conserved Glu in glucansucrase enzymes, while the equivalent residue is not conserved at all in amylomaltases of family GH77 (Table 1).
Function and regulation of family GH13 enzymes in fungi Filamentous fungi often live on decaying plant material. Consequently, they need to produce and secrete a full range of enzymes to degrade plant polysaccharides such as pectin, cellulose and starch. Indeed many fungi possess powerful enzyme mixtures to hydrolyse a variety of plant polysaccharides and form small oligosaccharides, which can be taken up via specialised transporters to serve as carbon and energy sources. To degrade starch, fungi produce extracellular α-amylases, α-glucosidases and glucoamylases. The extracellular fungal α-amylases identified thus far show considerable mutual similarity and have been classified in subfamily GH13_1. In A. niger, the enzymes from this subfamily include acid amylase (Boel et al., 1990) and the almost identical proteins AmyA/AmyB identified A. niger var awamori (Korman et al., 1990). These two identical enzymes are, however, not present in the genome sequence of A. niger strain ATCC 1015, which harbours only one orthologous protein with 72% identity to AmyA/B. In addition, a protein identical to A. oryzae TAKA-amylase has been identified in an industrial A. niger strain (Vujicic-Zagar and Dijkstra, 2006). Other previously identified starch-acting enzymes in A. niger include glucoamylase GlaA (Boel et al., 1984) and α-glucosidase AglA (Nakamura et al., 1997), which has been renamed AgdA to prevent confusion with α-galactosidases (den

Alpha-glucan acting enzymes in Aspergillus niger
18
Herder et al., 1992). Thus far, no enzymes from family GH70 or GH77 have been identified in Eukaryotes including fungi.
Expression of the starch degrading enzymes in Aspergillus species is regulated by AmyR, a positive transcriptional regulator with a GAL4-type Zn(II)2Cys6 cluster. The six conserved cysteine residues in this type of regulator need two Zn atoms (or occasionally Cd atoms) to form a so called binuclear cluster, which allows binding to a specific stretch of DNA (Pan and Coleman, 1990). The specific DNA sequence for binding of AmyR is identified as 5’-CGGN8(C/A)GG-3’ (Petersen et al., 1999;Ito et al., 2004). Expression of starch degrading enzymes is induced via AmyR when breakdown products of starch are present, notably maltose or isomaltose. In A. nidulans, isomaltose was found to be a stronger inducer of the AmyR system than maltose. It was suggested that maltose is converted into isomaltose by an α-glucosidase with α-(1,6) transglycosylation activity (Kato et al., 2002a;Kato et al., 2002b). The expression of starch degrading enzymes is repressed in the presence of high concentrations of an easily accessible, monosaccharide carbon source such as glucose. This process of carbon catabolite repression is mediated by the transcriptional repressor CreA (Dowzer and Kelly, 1991). Expression of the amyR gene is regulated both by CreA and by AmyR itself (Tani et al., 2001). Apart from starch catabolism, family GH13 enzymes in fungi are also involved in the formation and breakdown of the storage compound glycogen. The presence of intracellular glycogen has been demonstrated in several fungal species (Mattey and Allan, 1990;Bahia et al., 1997;Francois and Parrou, 2001), and the intracellular enzymes needed for its production and degradation are encoded in the available genome sequences. Two of these enzymes, glycogen branching and debranching enzyme, are members of family GH13. In S. cerevisiae, glycogen storage is regulated via the phosphorylation state of the main synthesising and degrading enzymes, glycogen synthase and glycogen phosphorylase, and is dependent on the availability of the carbon source (Francois and Parrou, 2001).
The fungal cell wall Recently, several studies have shown that different family GH13 enzymes of fungal origin may be involved in the formation of α-glucans in the fungal cell wall, rather than in starch degradation. The fungal cell wall is usually made up of chitin, β-glucan, α-glucan and covalently attached cell wall proteins. All these components form together an extensively crosslinked complex (Bowman and Free, 2006). The most abundant cell wall polysaccharide in yeasts and fungi is β-glucan, which mainly exists as β-(1,3) glucan with occasional β-(1,6) linked branches (Lesage and

Chapter 1 Introduction
19
Bussey, 2006). Additionally, a β-(1,6) linked glucan which is highly branched with β-(1,3) linkages is present in S. cerevisiae and S. pombe (Klis et al., 2001), but was not identified in several other ascomycete fungi. Among the aspergilli, the cell wall glucans of A. fumigatus are the best studied, due to the pathogenic nature of this species. Apart from β-(1,3) glucan, A. fumigatus contains a linear glucan with β-(1,3) and β-(1,4) linkages (Fontaine et al., 2000). β-Glucans are formed by plasma-membrane localized β-glucan synthases, which use UDP-glucose to form the glucan intracellularly, and subsequently transport it out of the cell (Cabib and Kang, 1987). Additionally, several enzymes localized in the cell wall area are known to either hydrolyze or transglycosylate β-glucans. In S. cerevisiae, these enzymes are mainly encoded by the GAS gene family (Ragni et al., 2007), and homologous proteins have been described in S. pombe (De Groot et al., 2007), C. albicans (Hartland et al., 1991) and A. fumigatus (Mouyna et al., 2000). As β-glucan forms a vital part of the cell wall for many pathogenic species, a group of anti-fungal drugs known as echinocandins is on the market, which act through the inhibition of β-glucan synthases (Beauvais and Latgé, 2001).
Chitin, a linear chain of β-(1,4)-N-acetyl-glucosamine, is present in most yeast species as well as filamentous fungi. In S. cerevisiae it is mainly located in the scars formed after budding and constitutes only 1-2% of the cell wall dry weight (Lesage and Bussey, 2006). In filamentous fungi, chitin forms a larger component of the cell wall (10-20%) because it is responsible for maintaining the cell wall structure (Bowman and Free, 2006). It is produced by chitin synthases, integral membrane proteins that require UDP-N-acetyl-glucosamine as donor molecule.
Cell wall mannoproteins (proteins with covalently linked mannose residues) represent a considerable part of up to 40% (w/w) of the fungal cell wall (Brul et al., 1997). Two types of cell wall localized proteins are described in yeasts and fungi: the proteins with internal repeats (PIR proteins) and those which are linked to the plasma membrane or cell wall via a glycosylphosphatidylinositol (GPI) anchor. This last category of proteins is found among all eukaryotes. The GPI-anchored proteins contain an N-terminal signal sequence for secretion, which targets them to the endoplasmic reticulum (ER) where they are glycosylated. Additionally, a specific C-terminal sequence motif, described by De Groot et al. (2003), is recognized and cleaved, and a ready-made GPI-anchor is attached to the new C-terminus (the ω-site). The complex GPI structure consists of mannose and glucosamine residues linked to an inositol - PO4 - lipid component referred to as phosphatidylinositol (PI), and is linked to the C-terminus of the protein via a phosphoethanolamine group (Fig 4). Upon secretion, the protein is bound to the plasma membrane via the fatty acid chains. Some GPI-anchored proteins are subsequently released from their GPI-anchors by the action of phospholipase C (PLC) which cleaves between the inositol - PO4 and the lipid anchoring. The proteins
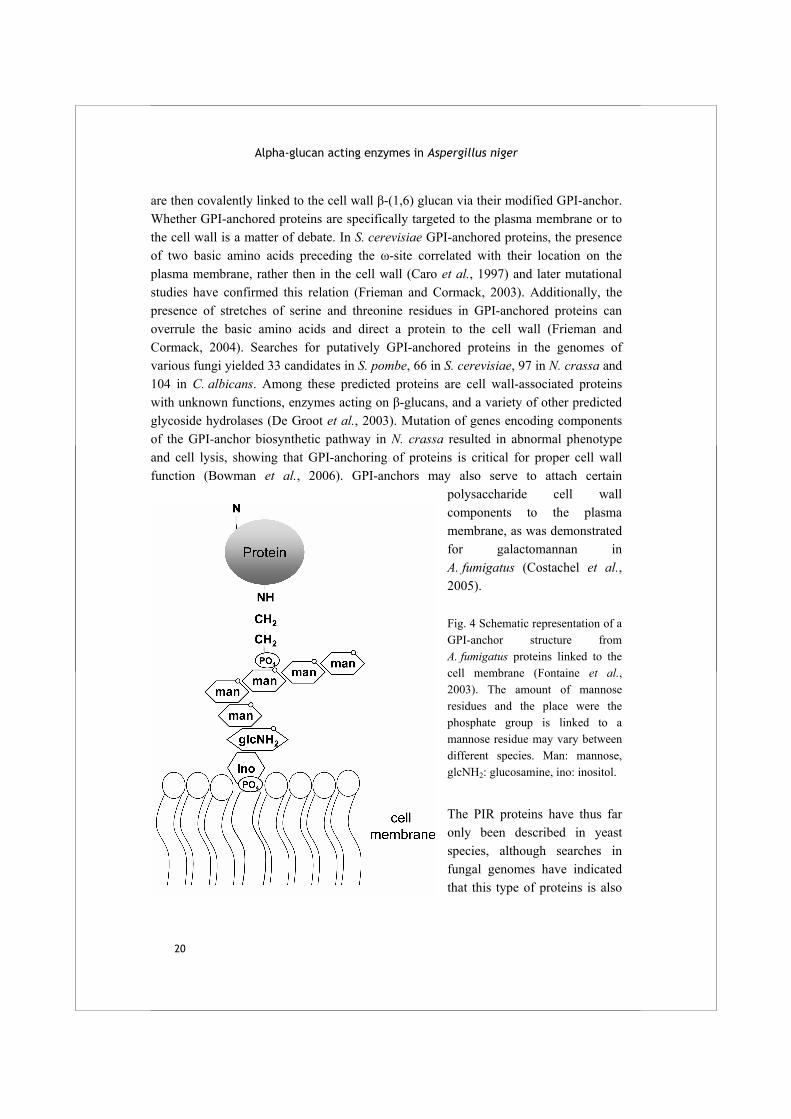
Alpha-glucan acting enzymes in Aspergillus niger
20
are then covalently linked to the cell wall β-(1,6) glucan via their modified GPI-anchor. Whether GPI-anchored proteins are specifically targeted to the plasma membrane or to the cell wall is a matter of debate. In S. cerevisiae GPI-anchored proteins, the presence of two basic amino acids preceding the ω-site correlated with their location on the plasma membrane, rather then in the cell wall (Caro et al., 1997) and later mutational studies have confirmed this relation (Frieman and Cormack, 2003). Additionally, the presence of stretches of serine and threonine residues in GPI-anchored proteins can overrule the basic amino acids and direct a protein to the cell wall (Frieman and Cormack, 2004). Searches for putatively GPI-anchored proteins in the genomes of various fungi yielded 33 candidates in S. pombe, 66 in S. cerevisiae, 97 in N. crassa and 104 in C. albicans. Among these predicted proteins are cell wall-associated proteins with unknown functions, enzymes acting on β-glucans, and a variety of other predicted glycoside hydrolases (De Groot et al., 2003). Mutation of genes encoding components of the GPI-anchor biosynthetic pathway in N. crassa resulted in abnormal phenotype and cell lysis, showing that GPI-anchoring of proteins is critical for proper cell wall function (Bowman et al., 2006). GPI-anchors may also serve to attach certain
polysaccharide cell wall components to the plasma membrane, as was demonstrated for galactomannan in A. fumigatus (Costachel et al., 2005). Fig. 4 Schematic representation of a GPI-anchor structure from A. fumigatus proteins linked to the cell membrane (Fontaine et al., 2003). The amount of mannose residues and the place were the phosphate group is linked to a mannose residue may vary between different species. Man: mannose, glcNH2: glucosamine, ino: inositol.
The PIR proteins have thus far only been described in yeast species, although searches in fungal genomes have indicated that this type of proteins is also

Chapter 1 Introduction
21
present in ascomycete fungi (De Groot et al., 2005). PIR proteins in S. cerevisiae contain repeats of a sequence of 19 amino acids and are linked to the cell wall β-(1,3) glucan via a mild-alkali sensitive linkage (Toh-e A et al., 1993;Kapteyn et al., 1999). Cell wall α-glucan is present in most Ascomycetes and Basidiomycetes, including S. pombe, but is absent from yeast species belonging to the Saccharomycetes. Generally, α-glucan in fungal cell walls is of the α-(1,3) type with a small percentage of α-(1,4) glycosidic bonds, also called pseudonigeran (Grün et al., 2005;Horisberger et al., 1972). Additionally, an α-glucan with alternating α-(1,3)/(1,4) glycosidic bonds (nigeran) has been identified in A. niger and several other Ascomycetes (Barker and Carrington, 1953;Woranovicz-Barreira et al., 1999;Johnston, 1965). A few cases of glucan types with mainly α-(1,4) bonds have been reported in S. pombe as well as A. niger (Garcia et al., 2006;Kirimura et al., 1999), but none of these were investigated in detail. It is generally believed that the α-glucan compound is produced by α-glucan synthases, although this synthesizing process was never directly demonstrated. Alpha-glucan synthases are predicted transmembrane enzymes with two catalytic domains, as shown in Fig. 5. The C-terminal, intracellular part has similarity to glycogen- and starch-synthases, members of family glycosyltransferase (GT) 5 which use UDP- or ADP glucose to form an α-glucan chain. The N-terminal, extracellular domain of α-glucan synthases has resemblance to family GH13 enzymes and is thought to be involved in the coupling of extruded glucan chains (Hochstenbach et al., 1998;Grün et al., 2005). In addition to α-glucan synthases, two other types of enzymes, both family GH13 homologs, were recently shown to play a role in fungal α-glucan formation. The first was Aah3p, a family GH13_1 protein identified in the fission yeast S. pombe (Morita et al., 2006). The knockout of the corresponding gene caused an aberrant cell shape and hypersensitivity towards cell wall degrading enzymes, indicating that cell wall biogenesis was somehow affected. Subsequent expression of mutated Aah3p in which the conserved aspartate and glutamate residues in the active site were replaced by alanine failed to rescue the phenotype of the knockout. This was an indication for the importance of the enzymatic activity of Aah3p, rather than its structural properties. Expression of a tagged version of Aah3p showed that the protein was localized in the cell membrane fraction, and that it was attached via a GPI-anchor, as was already predicted by De Groot et al. (2003). A second α-amylase homologue with a proposed role in cell wall formation is Amy1p from H. capsulatum. This predicted protein was shown to have high similarity to an α-amylase from Bacillus licheniformis, and had no recognisable signal sequence. A functional knockout of Amy1p lost completely the ability to form cell wall α-(1,3) glucan, which is a very relevant finding while α-(1,3) glucan is critical for virulence in this pathogenic fungus (Rappleye et al., 2004).

Alpha-glucan acting enzymes in Aspergillus niger
22
Fig. 5 Schematic representation of a fungal α-glucan synthase protein, based on the model of S. pombe Ags1p as proposed by Hochstenbach et al. (1998).
Scope of this thesis The starting point for the studies described in this thesis was the genome sequence of A. niger (Pel et al., 2007). The sequence was used to identify novel enzymes resembling family GH13, 15 and 31 enzymes putatively acting on starch. A selection of the predicted proteins was then studied with regard to regulation of expression, phylogeny, physiological function and biochemical characteristics. Chapter 1 describes the phylogenetic analysis of the family GH13, 15 and 31 enzymes in A. niger and related aspergilli. This is combined with the transcriptional analysis of these enzymes in A. niger grown on different carbon sources. The comparison of gene expression in a wild type strain and in an AmyR deletion strain allowed for the identification of genes encoding enzymes involved in starch degradation. Expression of some of the genes of interest appeared not to be regulated by AmyR or the presence of maltose, indicating that the encoded proteins may be involved in other physiological processes than starch degradation.

Chapter 1 Introduction
23
In chapter 2 two novel GH13 enzymes from A. niger are expressed, purified and characterised. The enzymes, named AgtA and AgtB, act as α-glucanotransferases on maltooligosaccharide substrates and starch. They are the first described family GH13 members which are predicted to be anchored to the cell wall or cell membrane with a GPI anchor. An A. niger knockout strain of agtA has an increased sensitivity for calcofluor white, a cell wall disturbing compound.
The two novel α-glucanotransferases are studied in more biochemical detail in chapter 3. By using a synthetically blocked substrate, we have determined the specific activity and Km for both enzymes. Additionally, the efficiency of both enzymes to use various acceptor substrates is compared.
In chapter 4, the phylogenetic analysis of a novel group of intracellular fungal α-amylases is presented. These enzymes are similar to H. capsulatum Amy1p and belong to the subfamily GH13_5. One enzyme from this novel group, AmyD from A. niger, is expressed in E. coli and biochemically characterised.
Finally, a summary of the data described in this thesis combined with a discussion of the results is included. A summary in Dutch, meant for those with little knowledge of microbiology and biochemistry, is also provided.


25
Chapter 2 Aspergillus niger genome wide analysis reveals a large number of novel α-glucan acting enzymes with unexpected expression profiles X-L. Yuan*, R.M. van der Kaaij*, C.A.M.J.J. van den Hondel, P.J. Punt, M.J.E.C van der Maarel, L. Dijkhuizen and A.F.J. Ram
*These authors contributed equally to this study
Submitted
Abstract The filamentous ascomycete Aspergillus niger is well known for its ability to produce a large variety of enzymes for the degradation of plant polysaccharide material. A major carbon and energy source for this soil fungus is starch, which can be degraded by the concerted action of α-amylase, glucoamylase and α-glucosidase enzymes, members of the glycoside hydrolase (GH) families 13, 15 and 31, respectively. In this study we have combined analysis of the genome sequence of A. niger CBS 513.88 with microarray experiments to identify novel enzymes from these GH families and to predict their physiological functions. Using HMM searches, we have identified a total of 17 previously unknown family GH13, 15 and 31 enzymes in the A. niger genome, all of which have orthologues in other aspergilli. Only two of the newly identified enzymes, a putative α-glucosidase (AgdB) and an α-amylase (AmyC), were predicted to play a role in starch degradation. The expression of the majority of the genes identified was not induced by maltose as carbon source, and not dependent on the presence of AmyR, the transcriptional regulator for starch degrading enzymes. The possible physiological functions of these other predicted family GH13, GH15 and GH31 enzymes, including predicted intracellular enzymes and cell wall associated proteins, in alternative α-glucan modifying processes are discussed.

Alpha-glucan acting enzymes in Aspergillus niger
26
Introduction Aspergillus niger is a saprophytic fungus well known for its production and secretion of a variety of hydrolytic enzymes contributing to its ability to degrade plant polysaccharides such as (hemi-)cellulose, pectin, starch and inulin (De Vries and Visser, 2001;Tsukagoshi et al., 2001;Yuan et al., 2006). Starch is the most abundant storage carbohydrate in the plant kingdom and is present in tubers, seeds and roots of a variety of crop plants including cereals, potatoes and manioc (Peters, 2006). Starch is composed of two different molecules: (i) amylose, an unbranched, single chain of α-(1,4)-linked glucose residues and (ii) amylopectin, consisting of a α-(1,4)-linked glucose chain with α-(1,6)-branches on every 12-25 glucose residues along the α-(1,4)-linked backbone (Robyt, 1998). The degradation of starch is performed by a variety of enzymes, which are divided over three Glycoside Hydrolase (GH) families based on their sequence similarity (http://www.cazy.org) (Coutinho and Henrissat, 1999). The first step in starch degradation is the endo-hydrolysis of the long polysaccharide chains into shorter maltooligosaccharides and α-limit dextrins by α-amylases (EC 3.2.1.1). Αlpha-amylases belong to family GH13, a large family containing various hydrolysing and transglycosylating enzymes, mostly acting on α-(1,4) or α-(1,6)-glycosidic bonds. Members of family GH13 have a (β/α)8 barrel structure and can be recognized by four highly conserved amino acid regions containing three catalytic residues (MacGregor et al., 2001;Nakajima et al., 1986). After endo-hydrolysis, subsequent steps in starch degradation involve exo-acting enzymes releasing glucose. This reaction is performed by glucoamylase type enzymes of family GH15 (EC 3.2.1.3), a relatively confined family with regard to enzyme specificity, as all its studied members hydrolyse either α-(1,4) or α-(1,6)-bonds to release β-glucose from the non-reducing end of maltooligosaccharides. Most GH15 enzymes described thus far have a starch binding domain (SBD) attached (Sauer et al., 2000), a discrete C-terminal region of the protein that can bind to starch and facilitates hydrolysis (Southall et al., 1999). Additionally, α-(1,4)-glucosidases of family GH31 may release α-glucose from the non-reducing end of starch (EC 3.2.1.20). This family also harbours other enzyme specificities such as α-xylosidase activity. Several A. niger enzymes involved in starch degradation, and their corresponding genes, have been characterized and isolated. A. niger glucoamylase GlaA (family GH15) is an important enzyme for the modification of starch in the food industry (Boel et al., 1984;van Dijck et al., 2003). Additionally, one GH31 α-glucosidase (AglA, renamed AgdA) (Nakamura et al., 1997) has been characterized previously, as well as three family GH13 α-amylases: acid amylase AamA, and the almost identical AmyA and

Chapter 2 Sequence and expression analysis
27
AmyB (Boel et al., 1990;Korman et al., 1990). Also, a protein identical to A. oryzae TAKA-amylase has been isolated for crystallization from an industrial A. niger strain (Vujicic-Zagar and Dijkstra, 2006). The transcriptional regulation of the genes encoding starch degrading enzymes has been studied in several aspergilli (Nakamura et al., 2006). In general, their expression is high on starch and induced by the presence of (iso)maltose (Tsukagoshi et al., 2001;Kato et al., 2002a). The presence of the inducer activates the Zn(II)2Cys6 transcription factor AmyR which binds to CGGN8(C/A)GG sequences in the promoter regions of AmyR target genes thereby activating their transcription (Petersen et al., 1999;Gomi et al., 2000;Tani et al., 2001;Ito et al., 2004).
Recent studies have indicated that some GH13 enzymes in fungi may be involved in the synthesis or remodeling of α-glucan in the fungal cell wall, rather than in starch degradation. The cell wall of aspergilli contains four major classes of polysaccharides: chitin, α-glucan, β-(1,3)-glucan and galactomannan (Fontaine et al., 2000;Beauvais and Latgé, 2001). The α-glucan fraction identified in A. niger consists of two types of molecules: a linear polymer with alternating α-(1,3)/(1,4)-glycosidic bonds called nigeran (Barker and Carrington, 1953) and pseudonigeran, a linear α-(1,3)-glucan molecule with some (3-10%) α-(1,4)-linkages (Johnston, 1965;Horisberger et al., 1972). Synthesis of α-glucan is thought to be carried out by α-glucan synthase enzymes encoded by ags genes. The first putative α-(1,3)-D-glucan synthase encoding gene (ags1) was identified in Schizosaccharomyces pombe (Hochstenbach et al., 1998). The ags1 gene encodes a large, three-domain protein. In addition to the multi-pass transmembrane domain in the C-terminal part of the protein, two predicted catalytic domains are present. The middle domain shows strong similarity to glycogen and starch synthases in family Glycosyltransferase (GT) 5 and is predicted to be involved in the synthesis of α-glucan. The N-terminal part of the protein is similar to α-amylases and belongs to family GH13. This part of the protein is predicted to be localized extracellularly, and might be involved in connecting two α-(1,3)-glucan chains (Grün et al., 2005). Apart from the α-glucan synthases, two types of family GH13 enzymes were recently identified in fungi to play a role in fungal cell wall biosynthesis. Marion et al. (2006) provided evidence for the involvement of a putative α-amylase (Amy1p) in the formation of α-(1,3)-glucan in the cell wall of Histoplasma capsulatum. An AMY1 knockout was unable to produce α-(1,3) glucan and showed reduced virulence. In dimorphic fungi like H. capsulatum, cell wall α-glucan is a known virulence factor (Rappleye et al., 2004;Rappleye and Goldman, 2006). The second α-amylase-like enzyme Aah3p was first studied in S. pombe (Morita et al., 2006). Disruption of aah3 encoding a GPI-anchored protein resulted in a hypersensitivity towards cell wall degrading enzymes and an aberrant cell shape, indicating that normal cell wall

Alpha-glucan acting enzymes in Aspergillus niger
28
biosynthesis was affected. Disruption of a homologous gene (agtA) in A. niger also had an effect on cell wall stability (van der Kaaij et al., 2007) (chapter 3). In this study, we surveyed the A. niger genome sequence (Pel et al., 2007) to identify for the first time all GH13, GH15 and GH31 family members present in this important industrial source for amylolytic enzymes. This resulted in identification of a surprisingly large number of previously unknown enzymes. By studying their phylogeny, the presence of specific protein features and synteny with other Aspergillus species, members of each GH family could be separated in several groups. Additionally, we studied the transcriptional regulation of the genes encoding these proteins in a wild type A. niger strain, and in a derived amyR deletion strain, during their growth on xylose and maltose. Only few of the identified proteins were induced by maltose. Expression of many of the identified groups of enzymes, including the homologues of both S. pombe Aah3p and H. capsulatum Amy1p, was not induced by maltose, and not dependent on the presence of AmyR. The possible involvement of these enzymes in cell wall α-glucan synthesis and modification is discussed.
Material and methods
Database mining of A. niger genome and analysis of predicted proteins
The full genome sequence of A. niger strain CBS 513.88, has been deposited at the EMBL database with accession numbers AM270980-AM270998 (Pel et al., 2007) and was used for database mining. The nucleotide accession numbers of A. niger genes as listed in Table 1 refer to this database. Hidden Markov Model (HMM) profiles were built with the HMMER package (Durbin and Eddy, 1998) (http://hmmer.wustl.edu/) based on the amino acid sequences of known members of GH13, GH15 and GH31. Proteins belonging to these families, originating from the different kingdoms of life, were retrieved from the CAZy website at http://www.cazy.org (Coutinho and Henrissat, 1999), and the protein sequences were extracted from the GenBank/GenPept database at http://www.ncbi.nlm.nih.gov/entrez/ and Swiss-Prot database at http://www.expasy.org/sprot/. The A. niger genome was searched with the HMM profiles using the WISE2 package (Birney et al., 2004) (http://www.ebi.ac.uk/Wise2/). The presence of signal peptidase cleavage sites, glycosylphosphatidylinositol (GPI) attachment sites and starch binding domains (SBD) in the obtained sequences were predicted by web-based tools at URL: http://www.cbs.dtu.dk/services/SignalP/ (Bendtsen et al., 2004), URL: http://mendel.imp.ac.at/sat/gpi/gpi_server (Eisenhaber et al., 2004), and URL: http://www.ncbi.nlm.nih.gov/BLAST/ (Marchler-Bauer and Bryant, 2004), respectively.

Chapter 2 Sequence and expression analysis
29
Multiple sequence alignments of GH13, 15 and 31 family members were performed using DNAMAN version 4.0 (Lynnon BioSoft, Canada). The alignments were based on the full length of the predicted proteins, except in case of predicted α-glucan synthases for which only the N-terminal part, encoding the family GH13 domain, was used for the alignment. The phylogenetic relationship was calculated by using Optimal Alignment (Thompson et al., 1994) with gap opening and gap extension penalties of 10 and 0.05, respectively. A bootstrapped test of phylogeny was performed by the neighbor-joining method using 1000 replicates. Wherever possible, one protein with described activity was included for each of the groups identified based on phylogenetic analysis.
Strains and transformations
The wild type A. niger strain used in this study is N402, a cpsA1 derivative of A. niger van Tieghem (CBS 120.49, ATCC 9029) (Bos et al., 1988). Strain AB4.1 is a pyrG negative derivative of N402 (van Hartingsveldt et al., 1987) and was used to construct the amyR disruption strain. A. niger strains were grown in Aspergillus minimal medium (MM) (Bennett and Lasure, 1991), or Aspergillus complete medium (CM) consisting of MM with the addition of 0.5% (w/v) yeast extract and 0.1% (w/v) casamino acids. Growth medium was supplemented with 10 mM uridine (Serva, Germany) when required. Transformation of A. niger AB4.1 was performed as described earlier (Punt and van den Hondel, 1992) using lysing enzymes (L1412, Sigma, U.S.A.) for protoplastation. The bacterial strain used for transformation and amplification of recombinant DNA was Escherichia coli XL1-Blue (Stratagene, U.S.A.). Transformation of XL1-Blue was performed according to the heat shock protocol (Inoue et al., 1990).
Disruption of the maltose utilization activator amyR in A. niger
Plasmid pJG01 containing the A. niger amyR gene as a 4.3 kb NsiI fragment in pGEM11 was kindly provided by P. vanKuyk (Wageningen University, the Netherlands) and used to disrupt the amyR gene. The construction of the amyR deletion cassette was performed as follows. The BamHI-EcoRI fragment and NsiI-SalI fragment flanking the amyR ORF at the 5’ and 3’ region respectively were isolated from pJG01. The isolated NsiI-SalI fragment was cloned into pUC19 to obtain plasmid pAmyRF3. Subsequently, a BamHI-SalI fragment carrying the A. oryzae pyrG gene, obtained from plasmid pAO4-13 (de Ruiter-Jacobs et al., 1989) was inserted into pAmyRF3 which resulted in plasmid pAmyRF3-pyrG. The BamHI-EcoRI fragment isolated from pJG10 was ligated into pAmyRF3-pyrG resulting in the amyR deletion plasmid (p∆amyR). Prior to transformation to AB4.1, p∆amyR was linearized with EcoRI. Uridine prototrophic transformants were selected by their ability to grow on MM without uridine. After two rounds of purification, transformants were tested for their ability to grow on starch. Approximately 10% of the pyrG+ transformants showed defective

Alpha-glucan acting enzymes in Aspergillus niger
30
growth on MM agar plates containing starch as sole carbon source. Six independent putative amyR deletion strains (YvdM1.1-1.6) with identical phenotypes were obtained. Southern blot analysis confirmed proper deletion and a single integration of the amyR disruption cassette at the amyR locus. Strain YvdM1.1 was used for further analysis and we will refer to this strain as the ΔamyR strain in the remaining of this paper.
Culture conditions, RNA preparation, microarray analysis and data analysis
RNA extracted from the A. niger ΔamyR strain and its parental strain (N402) grown on different carbon sources were used for microarray experiments using custom made ‘dsmM_ANIGERa_coll’ Affymetrix GeneChip® Microarrays kindly provided by DSM Food Specialties (Delft, The Netherlands). All experiments for each growth condition (culturing the mycelia, RNA extractions and microarray hybridizations) were performed twice as independent biological experiments. A. niger spores (2 x 106 spores ml-1) were inoculated in 250 ml MM supplemented with 2% (w/v) xylose (Sigma) and 0.1% (w/v) casamino acids and grown for 18 h at 30 °C on a rotary shaker at 300 rpm. The mycelium was harvested by suction over a nylon membrane and washed with MM without carbon source. Aliquots of 1.6 g wet weight of mycelium were transferred to 300 ml Erlenmeyer flasks containing 70 ml MM supplemented with 1% (w/v) carbon source (maltose (Sigma) or xylose) and incubated at 30 °C for a further 2 or 8 h. The pH of all cultures grown for 2 h was equal to the pH at the time of transfer (pH 6.2). Cultures grown for 8 h after transfer were buffered at pH 4 by addition of 100 mM of citric acid/sodium citrate to prevent strong acidification. The mycelium was harvested over Miracloth filter, frozen in liquid nitrogen and stored at -80 °C. Total RNA was isolated from mycelia using TRIzol reagent (Invitrogen) and RNA quality was verified by analyzing aliquots with glyoxal/DMSO gel electrophoresis and Agilent Bioanalyzer “Lab on chip” system (Agilent Technologies, U.S.A.). Processing, labeling and hybridization of cRNA to A. niger Affymetrix GeneChips were performed according to the corresponding Affymetrix protocols for “Eukaryotic Target Preparation” and “Eukaryotic Target hybridization”. For probe array washing and staining, the protocol “Antibody Amplification for Eukaryotic Targets” was followed. Hybridized probe array slides were scanned with Agilent technologies G2500A Gene Array Scanner at a 3 µm resolution and a wavelength of 570 nm. Affymetrix Microarray Suite software MAS5.0 was used to calculate the signal and p-values and to set the algorithm’s absolute call flag, which indicates the reliability of the data points according to P (present), M (marginal) and A (absent). The data on each chip were globally scaled to an arbitrary target gene intensity of 500.

Chapter 2 Sequence and expression analysis
31
The pre-scaled data from each hybridization experiment were then assigned for per chip normalization using Genespring 7.0 software (Silicon Genetics, U.S.A.). For genome wide analysis, we focused on maltose induced genes and therefore a pre-filtering of data was performed to select for genes whose detection calls are present in both maltose duplicate samples in the wild type strain (N402). The selected dataset was further used for 1-way ANOVA analysis under the test type of “parametric test, don’t assume variances equal”. Fold changes in expression between two different conditions were then computed for genes with p < 0.08 based on 1- way ANOVA analysis.
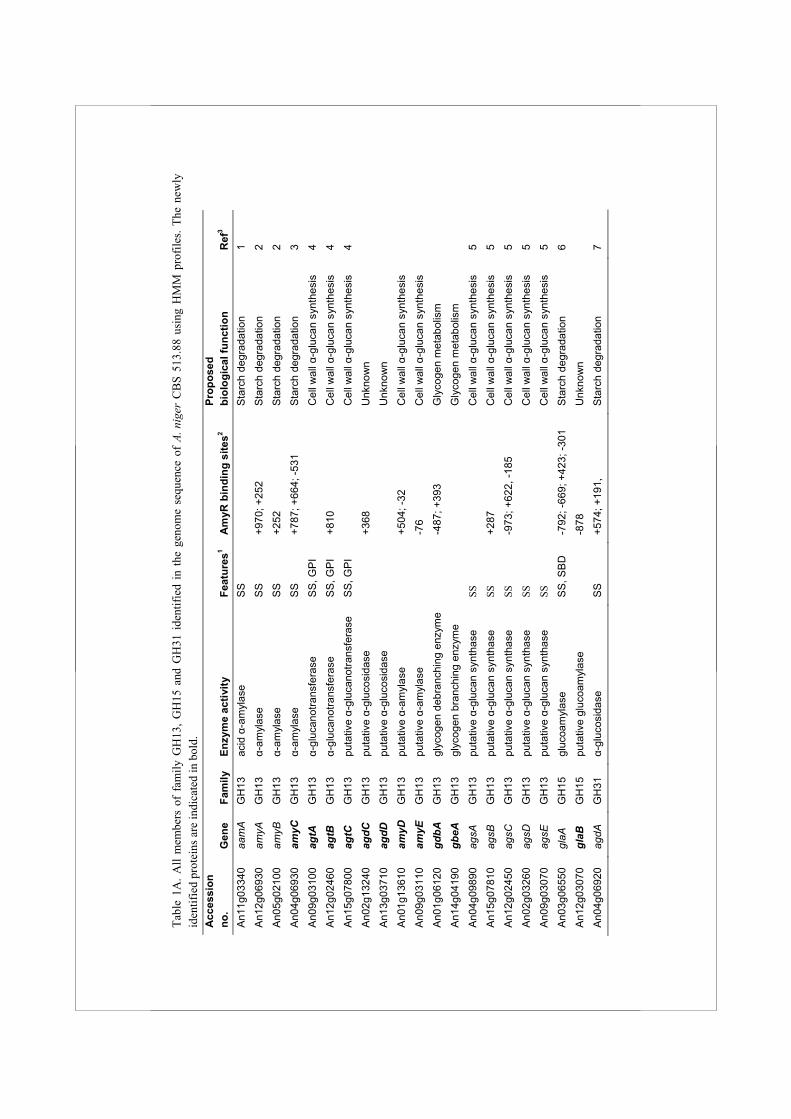
Tabl
e 1A
. All
mem
bers
of
fam
ily G
H13
, GH
15 a
nd G
H31
iden
tifie
d in
the
geno
me
sequ
ence
of
A. n
iger
CB
S 51
3.88
usi
ng H
MM
pro
files
. The
new
ly
iden
tifie
d pr
otei
ns a
re in
dica
ted
in b
old.
A
cces
sion
no
. G
ene
Fam
ily
Enzy
me
activ
ity
Feat
ures
1 A
myR
bin
ding
site
s2 Pr
opos
ed
biol
ogic
al fu
nctio
n R
ef3
An1
1g03
340
aam
A
GH
13
acid
α-a
myl
ase
SS
Sta
rch
degr
adat
ion
1
An1
2g06
930
amyA
G
H13
α-
amyl
ase
SS
+9
70; +
252
Sta
rch
degr
adat
ion
2
An0
5g02
100
amyB
G
H13
α-
amyl
ase
SS
+2
52
Sta
rch
degr
adat
ion
2
An0
4g06
930
amyC
G
H13
α-
amyl
ase
SS
+7
87; +
664;
-531
S
tarc
h de
grad
atio
n 3
An0
9g03
100
agtA
G
H13
α-
gluc
anot
rans
fera
se
SS
, GP
I
Cel
l wal
l α-g
luca
n sy
nthe
sis
4
An1
2g02
460
agtB
G
H13
α-
gluc
anot
rans
fera
se
SS
, GP
I +8
10
Cel
l wal
l α-g
luca
n sy
nthe
sis
4
An1
5g07
800
agtC
G
H13
pu
tativ
e α-
gluc
anot
rans
fera
se
SS
, GP
I
Cel
l wal
l α-g
luca
n sy
nthe
sis
4
An0
2g13
240
agdC
G
H13
pu
tativ
e α-
gluc
osid
ase
+3
68
Unk
now
n
An1
3g03
710
agdD
G
H13
pu
tativ
e α-
gluc
osid
ase
Unk
now
n
An0
1g13
610
amyD
G
H13
pu
tativ
e α-
amyl
ase
+5
04; -
32
Cel
l wal
l α-g
luca
n sy
nthe
sis
An0
9g03
110
amyE
G
H13
pu
tativ
e α-
amyl
ase
-7
6 C
ell w
all α
-glu
can
synt
hesi
s
An0
1g06
120
gdbA
G
H13
gl
ycog
en d
ebra
nchi
ng e
nzym
e -4
87; +
393
Gly
coge
n m
etab
olis
m
An1
4g04
190
gbeA
G
H13
gl
ycog
en b
ranc
hing
enz
yme
G
lyco
gen
met
abol
ism
An0
4g09
890
agsA
G
H13
pu
tativ
e α-
gluc
an s
ynth
ase
SS
C
ell w
all α
-glu
can
synt
hesi
s 5
An1
5g07
810
agsB
G
H13
pu
tativ
e α-
gluc
an s
ynth
ase
SS
+287
C
ell w
all α
-glu
can
synt
hesi
s 5
An1
2g02
450
agsC
G
H13
pu
tativ
e α-
gluc
an s
ynth
ase
SS
-973
; +62
2, -1
85
Cel
l wal
l α-g
luca
n sy
nthe
sis
5 A
n02g
0326
0 ag
sD
GH
13
puta
tive α-
gluc
an s
ynth
ase
SS
C
ell w
all α
-glu
can
synt
hesi
s 5
An0
9g03
070
agsE
G
H13
pu
tativ
e α-
gluc
an s
ynth
ase
SS
C
ell w
all α
-glu
can
synt
hesi
s 5
An0
3g06
550
glaA
G
H15
gl
ucoa
myl
ase
SS
, SB
D
-792
; -66
9; +
423;
-301
S
tarc
h de
grad
atio
n 6
An1
2g03
070
glaB
G
H15
pu
tativ
e gl
ucoa
myl
ase
-8
78
Unk
now
n
An0
4g06
920
agdA
G
H31
α-
gluc
osid
ase
SS
+5
74; +
191,
S
tarc
h de
grad
atio
n 7

Tabl
e 1A
(con
tinue
d)
Acc
essi
on
no.
Gen
e Fa
mily
En
zym
e ac
tivity
Fe
atur
es1
Am
yR b
indi
ng s
ites2
Prop
osed
bi
olog
ical
func
tion
Ref
3
An0
1g10
930
agdB
G
H31
pu
tativ
e α-
gluc
osid
ase
SS
+9
04; -
334
Sta
rch
degr
adat
ion
An0
9g05
880
agdE
G
H31
pu
tativ
e α-
gluc
osid
ase
II S
S
P
rote
in g
lyco
syla
tion
An1
8g05
620
agdF
G
H31
un
know
n
U
nkno
wn
An0
7g00
350
agdG
G
H31
un
know
n S
S
+402
U
nkno
wn
An0
9g03
300
axlA
G
H31
pu
tativ
e α-
xylo
sida
se
SS
+1
26
Xyl
oglu
can
degr
adat
ion
An0
1g04
880
axlB
G
H31
pu
tativ
e α-
xylo
sida
se
+4
30; +
138,
X
ylog
luca
n de
grad
atio
n

Tabl
e 1B
. Fun
ctio
nally
des
crib
ed f
amily
GH
13 a
nd G
H31
mem
bers
fro
m o
ther
org
anis
ms,
used
for
the
mul
tiple
seq
uenc
e al
ignm
ents
(Fi
g. 1
). Fo
r ea
ch
A. n
iger
pro
tein
iden
tifie
d, a
func
tiona
lly o
r bio
chem
ical
ly c
hara
cter
ized
pro
tein
with
the
high
est s
imila
rity
was
use
d in
the
phyl
ogen
etic
ana
lysi
s.
Acc
essi
on n
o.
Nam
e Fa
mily
En
zym
e ac
tivity
Fe
atur
es1
Bio
logi
cal f
unct
ion
Org
anis
m
Ref
3
BA
A78
714
A
ndG
be1
GH
13
glyc
ogen
bra
nchi
ng e
nzym
e G
lyco
gen
met
abol
ism
A
. nid
ulan
s 8
BA
A34
996
ScG
db1
GH
13
glyc
ogen
deb
ranc
hing
enz
yme
Gly
coge
n m
etab
olis
m
S. c
erev
isia
e 9
P19
571
BsA
myA
G
H13
α-
amyl
ase
SS
S
tarc
h de
grad
atio
n B
acill
us s
p.
10
CA
A54
266
BsA
glA
G
H13
α-
gluc
osid
ase
S
tarc
h de
grad
atio
n B
acill
us s
p.
11
CA
A21
237
S
pAah
1 G
H13
un
know
n SS
, GPI
α-
Glu
can
bios
ynth
esis
S
. pom
be
12
CA
A91
249
SpA
ah2
GH
13
unkn
own
SS, G
PI
α-G
luca
n bi
osyn
thes
is
S. p
ombe
12
C
AB
4000
6 S
pAah
3 G
H13
un
know
n SS
, GPI
α-
Glu
can
bios
ynth
esis
S
. pom
be
12
CA
A16
864
SpA
ah4
GH
13
unkn
own
SS, G
PI
α-G
luca
n bi
osyn
thes
is
S. p
ombe
12
A
BK
6285
4 H
cAm
y1
GH
13
unkn
own
α-G
luca
n bi
osyn
thes
is
H. c
apsu
latu
m
13
AB
F508
83
AN
7345
.2
GH
31
α/β-
gluc
osid
ase
SS
S
tarc
h/ce
llulo
se d
egra
datio
n A
. nid
ulan
s 14
A
BF5
0846
A
N75
05.2
G
H31
α-
xylo
sida
se
X
ylan
deg
rada
tion
A. n
idul
ans
14
BA
B39
856
And
Agd
B
GH
31
α-gl
ucos
idas
e S
S
Sta
rch
degr
adat
ion
A. n
idul
ans
15
AA
U87
580
Tr
Agu
II G
H31
α-
gluc
osid
ase
II S
S
Pro
tein
gly
cosy
latio
n T.
rees
ei
16
A45
249
CA
MA
L2
GH
31
mal
tase
Mal
tose
deg
rada
tion
C. a
lbic
ans
17
1 SS
= p
redi
cted
N-te
rmin
al S
igna
l Seq
uenc
e; G
PI =
pre
dict
ed G
lyco
sylp
hosp
hatid
ylin
osito
l anc
hor s
igna
l; SB
D =
pre
dict
ed st
arch
bin
ding
dom
ain.
2 Th
e pr
esen
ce o
f con
sens
us A
myR
bin
ding
site
s (C
GG
N8(
A/C
)GG
) was
ana
lyse
d in
the
prom
oter
regi
on u
p to
1 k
b up
stre
am o
f the
star
t cod
on.
The
num
bers
indi
cate
the
dist
ance
to th
e tra
nsla
tion
star
t cod
on. T
he c
onse
nsus
sequ
ence
mig
ht b
e pr
esen
t in
the
codi
ng (+
) or i
n th
e no
n-co
ding
(-) s
trand
. 3 R
efer
ence
s: 1
(Boe
l et a
l., 1
990)
; 2 (K
orm
an e
t al.,
199
0); 3
Van
der
Kaa
ij &
Yua
n. u
npub
l.; 4
(van
der
Kaa
ij et
al.,
200
7); 5
(Dam
veld
et a
l., 2
005b
);
6 (B
oel e
t al.,
198
4); 7
(Nak
amur
a et
al.,
199
7); 8
(Sas
angk
a et
al.,
200
2); 9
(Tes
te e
t al.,
200
0); 1
0 (T
suka
mot
o et
al.,
198
8); 1
1 (N
akao
et a
l., 1
994)
; 12
(Mor
ita e
t al
., 20
06);
13 (
Mar
ion
et a
l., 2
006)
; 14
(B
auer
et
al.,
2006
); 15
(K
ato
et a
l., 2
002b
); 16
(G
eyse
ns e
t al
., 20
05);
17 (
Geb
er e
t al
., 19
92).

Chapter 2 Sequence and expression analysis
35
Results
Identification of glycoside hydrolase family 13, 15 and 31 genes in the A. niger CBS 513.88 genome sequence
α-Amylases, glucoamylases and α-glucosidases, members of families GH13, 15 and 31, respectively, are the three main types of enzymes involved in breakdown of starch by aspergilli (Tsukagoshi et al., 2001). To identify all genes encoding enzymes that might play a role in starch utilization, or other α-glucan modifying processes in A. niger, the genome of A. niger CBS 513.88 was searched with HMM profiles based on known enzymes from families GH13, 15 and 31. This resulted in the retrieval of a total of 27 protein sequences (listed in Table 1A) including 17 previously unknown proteins. Two approaches were combined to predict putative functions in cellular processes for this surprisingly large number of newly identified proteins. First, phylogenetic trees were constructed using the GH13, GH15 and GH31 family members identified in the A. niger genome, as well as functionally characterized proteins from other organisms with similarity to the identified A. niger proteins (Fig. 1). Second, using DNA microarrays, the expression of all the A. niger genes encoding GH13, GH15 and GH31 enzymes was examined in both the A. niger wild type strain N402 and the ΔamyR strain derived, after growth on different carbon sources (Fig. 2 and Suppl. Table 1). Both the N402 and the ΔamyR strains were pregrown in xylose for 18 h, and mycelia were transferred to either xylose or maltose media and grown further for 2 h or 8 h. Expression levels were determined based on geometric mean data of biological duplicate samples. We will discuss each enzyme family in detail and combine the findings in A. niger with the predicted proteins present in the genomes of A. fumigatus (Nierman et al., 2005), A. nidulans (Galagan et al., 2005) and A. oryzae (Machida et al., 2005).
Identification and transcriptional regulation of GH13 family members
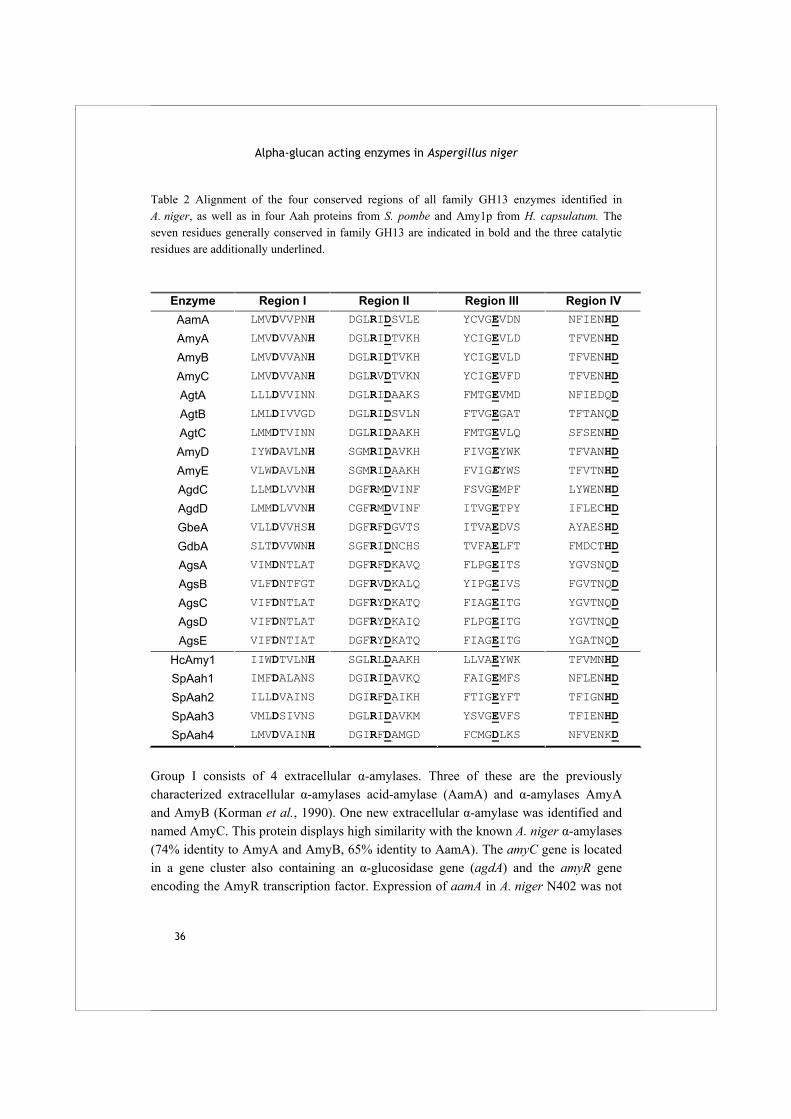
The HMMer search for family GH13 enzymes in the A. niger genome resulted in the identification of 18 protein sequences of which 10 had not been identified previously (Table 1A). Table 2 displays the four conserved regions typical for family GH13 proteins as identified in these enzymes. A phylogenetic tree was produced combining the A. niger family GH13 enzymes with several functionally characterized GH13 family proteins from other organisms (Fig. 1A). The combination of this phylogenetic analysis with a functional annotation of the proteins revealed 6 recognizable subgroups.
Alpha-glucan acting enzymes in Aspergillus niger
36
Table 2 Alignment of the four conserved regions of all family GH13 enzymes identified in A. niger, as well as in four Aah proteins from S. pombe and Amy1p from H. capsulatum. The seven residues generally conserved in family GH13 are indicated in bold and the three catalytic residues are additionally underlined.
Enzyme Region I Region II Region III Region IV AamA LMVDVVPNH DGLRIDSVLE YCVGEVDN NFIENHD
AmyA LMVDVVANH DGLRIDTVKH YCIGEVLD TFVENHD
AmyB LMVDVVANH DGLRIDTVKH YCIGEVLD TFVENHD
AmyC LMVDVVANH DGLRVDTVKN YCIGEVFD TFVENHD
AgtA LLLDVVINN DGLRIDAAKS FMTGEVMD NFIEDQD
AgtB LMLDIVVGD DGLRIDSVLN FTVGEGAT TFTANQD
AgtC LMMDTVINN DGLRIDAAKH FMTGEVLQ SFSENHD
AmyD IYWDAVLNH SGMRIDAVKH FIVGEYWK TFVANHD
AmyE VLWDAVLNH SGMRIDAAKH FVIGEYWS TFVTNHD
AgdC LLMDLVVNH DGFRMDVINF FSVGEMPF LYWENHD
AgdD LMMDLVVNH CGFRMDVINF ITVGETPY IFLECHD
GbeA VLLDVVHSH DGFRFDGVTS ITVAEDVS AYAESHD
GdbA SLTDVVWNH SGFRIDNCHS TVFAELFT FMDCTHD
AgsA VIMDNTLAT DGFRFDKAVQ FLPGEITS YGVSNQD
AgsB VLFDNTFGT DGFRVDKALQ YIPGEIVS FGVTNQD
AgsC VIFDNTLAT DGFRYDKATQ FIAGEITG YGVTNQD
AgsD VIFDNTLAT DGFRYDKAIQ FLPGEITG YGVTNQD
AgsE VIFDNTIAT DGFRYDKATQ FIAGEITG YGATNQD
HcAmy1 IIWDTVLNH SGLRLDAAKH LLVAEYWK TFVMNHD
SpAah1 IMFDALANS DGIRIDAVKQ FAIGEMFS NFLENHD
SpAah2 ILLDVAINS DGIRFDAIKH FTIGEYFT TFIGNHD
SpAah3 VMLDSIVNS DGLRIDAVKM YSVGEVFS TFIENHD
SpAah4 LMVDVAINH DGIRFDAMGD FCMGDLKS NFVENKD
Group I consists of 4 extracellular α-amylases. Three of these are the previously characterized extracellular α-amylases acid-amylase (AamA) and α-amylases AmyA and AmyB (Korman et al., 1990). One new extracellular α-amylase was identified and named AmyC. This protein displays high similarity with the known A. niger α-amylases (74% identity to AmyA and AmyB, 65% identity to AamA). The amyC gene is located in a gene cluster also containing an α-glucosidase gene (agdA) and the amyR gene encoding the AmyR transcription factor. Expression of aamA in A. niger N402 was not

Chapter 2 Sequence and expression analysis
37
detectable in xylose media, but was strongly induced in the presence of maltose (Fig. 2). The expression of aamA was reduced to a non-detectable level in the ΔamyR strain. Expression of amyA and amyB was not detected in A. niger N402 in any of the conditions tested (Fig. 2) (see Discussion). The expression of the newly identified amyC gene was relatively low compared to the aamA gene. At 2 h after transfer from the preculture, the expression level of amyC was independent of the carbon source, while after 8 h the expression level was reduced 3 fold on xylose compared to maltose. Additionally, the expression on maltose was reduced 2-3 fold in the ΔamyR strain (Suppl. Table 1). The presence of three putative AmyR binding elements in the promoter region of amyC further suggests that its expression is controlled by AmyR (Table 1A). Group II contains three putative GPI-anchored enzymes, recently identified as α-glucanotransferases, and named AgtA, AgtB and AgtC, respectively (van der Kaaij et al., 2007) (chapter 3). This subgroup of proteins is characterized by the presence of two hydrophobic signal sequences. The N-terminal signal sequence is predicted to serve for translocation to the endoplasmic reticulum (ER) while the C-terminal sequence is predicted to be replaced by a preassembled glycosylphosphatidylinositol (GPI) anchor in the ER (Orlean, 1997). The three enzymes cluster together with the α-amylases in the phylogenetic tree (Fig. 1A), but can be distinguished from the α-amylases by their catalytic domains which are clearly different from the consensus sequence for the α-amylase family. In all three proteins, one or two highly conserved histidines in conserved regions I and IV are replaced by other hydrophilic residues (Table 2). Family GH13 members without these histidine residues, which are part of the active site, are very rare. Mutation of these residues generally results in reduced activity or altered reaction specificity of the enzymes (Chang et al., 2003;Leemhuis et al., 2004). Interestingly, both the conserved His-residues in Regions I and IV are also missing in all (putative) α-glucan synthases (Table 2). Other Aspergillus genomes harbour two or three Agt homologues (Suppl. Fig. 1A, Suppl. Table 2), all sharing the aberrant conserved regions and predicted GPI-anchoring. In the A. niger genome, both agtB and agtC are located next to genes encoding putative α-glucan synthases, and this arrangement of genes is conserved in other aspergilli. The agtA gene is constitutively expressed in both the wild type strain N402 and the ΔamyR strain under all growth conditions examined (Fig. 2). Expression of agtB was only detected 8 h after transfer, regardless of the carbon source and independent of AmyR, while expression of agtC was not detected.

Alpha-glucan acting enzymes in Aspergillus niger
38
Fig. 1A Bootstrapped phylogenetic tree of A. niger GH13 enzymes and several closest homologues from other species. Newly identified proteins in the genome of A. niger are shown in bold. A description of each protein is listed in Tables 1A/B. Bootstrap values are indicated on the node of each branch. The tree was created with DNAMAN 4.0 using gap and extension penalties of 10 and 0.5 respectively. The scale bar corresponds to a genetic distance of 0.05 substitution per position.
Group III consists of two putative intracellular α-glucosidases, named agdC and agdD. The protein sequences contained all residues commonly conserved in the α-amylase superfamily (Table 2). The predicted intracellular proteins lack clear similarity to any previously characterized fungal protein, although similar enzymes are predicted in A. oryzae, A. nidulans and A. fumigatus (Suppl. Fig. 1A). Their most related functionally characterized homologue is an α-glucosidase from Bacillus sp. SAM1606

Chapter 2 Sequence and expression analysis
39
(Nakao et al., 1994). Expression of agdC was low and not induced on maltose, and expression of agdD was not observed (Fig. 2). Group IV contains two putative intracellular α-amylases, named AmyD and AmyE, which share 55% identity. The predicted proteins are similar to a recently identified protein Amy1p from H. capsulatum, which was shown to be involved in cell wall α-glucan synthesis. The latter protein has not been characterized biochemically. Functionally characterized enzymes with similarity to this cluster therefore included only bacterial enzymes, of which maltohexaose-forming α-amylase of alkalophilic Bacillus sp. #707 (Tsukamoto et al., 1988) had the highest similarity. No significant expression of amyD or amyE was detected in our experiments (Fig. 2). Predicted enzymes highly similar to A. niger AmyD and AmyE were also present in other Aspergillus species (Suppl. Fig. 1A, Suppl. Table 2). Like the A. niger amyE gene, the orthologues in A. nidulans (AN3309.3) and in A. oryzae (AO003001497) are clustered with genes encoding α-glucanotransferases and α-glucan synthases (Suppl. Table 2). The two A. niger proteins in Group V could be reliably annotated as enzymes involved in glycogen metabolism: glycogen branching enzyme (GbeA) and glycogen debranching enzyme (GdbA). A homologue for each of these enzymes is present in the other Aspergillus genome sequences. Transcriptional analysis in A. niger showed that both genes were constitutively expressed, and this was unaffected in the ΔamyR strain (Fig. 2). Group VI contains the five predicted α-glucan synthase genes (Damveld et al., 2005). The derived proteins are highly similar (66- 83%) to each other and all contain the two catalytic domains (GT5 and GH13) characteristic for these proteins. Both agsB and agsC are clustered in the genome with genes encoding the above mentioned α-glucanotransferases agtC and agtB, respectively. In both cases, the direction of transcription of the pair of genes is such that they can be transcribed from their intergenic region. Expression analysis in our microarray data collection, which includes data from various time points on several carbon sources (glucose, maltose, xylose, inulin, sucrose) indicate that agsC and agtB are co-expressed at later growth stage, independent of the carbon source (data not shown). Expression of both agsB and its neighboring gene agtC was not detected in any of the growth conditions. The expression levels of agsA and agsD were very low, or not detectable. AgsE was highly expressed in all experiments independent of AmyR (Fig. 2). As expected, several proteins with high similarity to the A. niger α-glucan synthases are predicted from the other Aspergillus genomes, although the number of homologues present is highest in A. niger (Suppl. Fig. 1A, Suppl. Table 2).
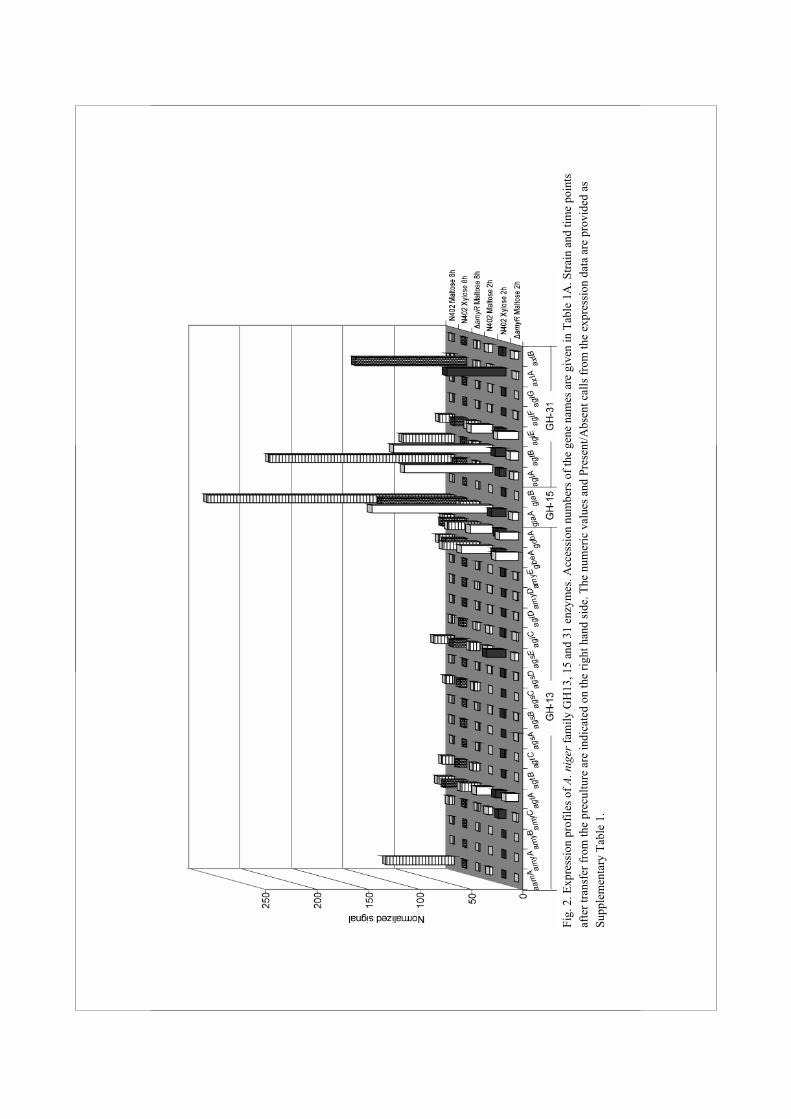
Fi
g. 2
. Exp
ress
ion
prof
iles o
f A. n
iger
fam
ily G
H13
, 15
and
31 e
nzym
es. A
cces
sion
num
bers
of t
he g
ene
nam
es a
re g
iven
in T
able
1A
. Stra
in a
nd ti
me
poin
ts
afte
r tra
nsfe
r fro
m th
e pr
ecul
ture
are
indi
cate
d on
the
right
han
d si
de. T
he n
umer
ic v
alue
s and
Pre
sent
/Abs
ent c
alls
from
the
expr
essi
on d
ata
are
prov
ided
as
Supp
lem
enta
ry T
able
1.

Chapter 2 Sequence and expression analysis
41
Identification and transcriptional regulation of GH15 family members
A HMMer search for GH15 family members in the A. niger genome returned two proteins: the previously described glucoamylase GlaA (Boel et al., 1984) and an unknown predicted protein named GlaB. The GlaB protein lacks both an N-terminal signal sequence and an SBD, and displays a low similarity of 26% with GlaA (Suppl. Fig. 1B). These two types of family GH15 enzymes are also recognized in other aspergilli studied. All predicted proteins similar to GlaB lack both a signal for secretion and an SBD, features typically present in the fungal GH15 enzymes described to date. A. niger glaA was expressed on xylose and strongly induced on maltose, as described previously (Fowler et al., 1990). The induction of glaA was AmyR dependent. Expression of glaB was not detected in any of the conditions tested (Fig. 2).
Identification and transcriptional regulation of GH31 family members
HMMer searches in the genome of A. niger revealed the presence of seven GH31 family members, of which only one was previously identified as an α-glucosidase (aglA) (Nakamura et al., 1997). We propose to name all (putative) α-glucosidases in A. niger Agd enzymes, similar to the nomenclature in A. nidulans, and to rename AglA as AgdA, to prevent confusion with α-galactosidases (den Herder et al., 1992). The phylogenetic tree (Fig. 1B) shows the presence of (at least) 4 subgroups within family GH31. In the group of the α-glucosidases, AgdA clusters with AgdB, a predicted extracellular protein. AgdB has some similarity to two A. nidulans enzymes: AN7345.2 (62% identity) with both α- and β-glucosidase activity (Bauer et al., 2006) and α-glucosidase B (AndAgdB/ AN8953.3, 53% identity), with strong transglycosylation activity (Kato et al., 2002b). However, none of the true orthologues of AgdB has been characterized (Suppl. Fig. 1C). Transcript analysis revealed that agdB was regulated similar to agdA, as the strong induction of both genes in the presence of maltose was dependent on the presence of amyR. However, where most AmyR regulated genes (aamA, glaA and agdA) reached their highest level of induction after 8 h growth on maltose, the expression level of agdB was decreased after 8 h compared to 2 h (Fig. 2). From the remaining five GH31 family members, AgdE shows similarity to Trichoderma reesei glucosidase II (TrAguII) (Geysens et al., 2005). This type of α-glucosidases is located in the ER where it is involved in the trimming of α-(1,3)-linked glucose residues from the N-glycan core structure Glc3Man9GlcNAc2, which may be attached to proteins designated to be secreted (Geysens et al., 2005). AgdE contains a predicted signal sequence which might serve to direct the protein to the ER. The agdE gene was constitutively expressed in all conditions tested (Fig. 2), consistent with its predicted function as an α-glucosidase II. Two additional GH31 family members, AgdF

Alpha-glucan acting enzymes in Aspergillus niger
42
and AgdG, lack similarity to any functionally described proteins. Expression of both genes was not detected in any of the tested growth conditions.
Fig. 1B Bootstrapped phylogenetic tree of A. niger GH31 enzymes and several closest homologues from other species. Newly identified proteins in the genome of A. niger are shown in bold. A description of each protein is listed in Tables 1A/B. Bootstrap values are indicated on the node of each branch. The tree was created with DNAMAN 4.0 using gap and extension penalties of 10 and 0.5 respectively. The scale bar corresponds to a genetic distance of 0.05 substitution per position. The two final family GH31 members, named AxlA and AxlB, cluster with AN7505.2 from A. nidulans, which was recently characterized as an α-xylosidase (Bauer et al., 2006). Gene axlA was highly expressed in the presence of xylose, while no expression was detected in maltose grown cultures (Fig. 2). The gene encoding the putatively intracellular AxlB was expressed at a very low level independent of the carbon source (Fig. 2). A BLASTP search in the genomes of A. fumigatus, A. nidulans and A. oryzae for family GH31 enzymes resulted in a similar collection of enzymes as identified in A. niger (Suppl. Fig. 1C). Several clusters of orthologous proteins are distinguishable, but the assignment of an enzymatic activity to these clusters is not yet possible due to a lack of well studied homologues for these enzymes.

Chapter 2 Sequence and expression analysis
43
Genome wide analysis of AmyR dependent maltose induced genes using microarrays
The expression analysis of genes encoding family GH13, GH15 and GH31 enzymes in the A. niger genome revealed that the expression of only a limited number of genes was induced by maltose in an AmyR dependent way. In fact, only four genes (aamA, glaA, agdA, and agdB) showed the predicted expression pattern for genes encoding enzymes involved in the breakdown of starch (Fig. 2). To identify additional genes with a possible role in starch metabolism, a genome wide expression analysis was performed. Comparison of the transcriptome of the wild type strain (N402), grown for 2 h on either xylose or maltose, identified 644 differentially expressed genes (>2-fold induction on maltose). A set of 13 genes was expressed > 2 fold higher in N402 compared to the ΔamyR strain. Combining the two gene sets resulted in a collection of 7 genes that were > 2-fold induced on maltose in an AmyR dependent manner (Fig. 3, Table 3). Six of these were assigned to the category of carbohydrate transport and metabolism according to FunCat (Ruepp et al., 2004), including the three genes encoding known extracellular starch degrading enzymes (aamA, glaA and agdA) and a putative α-glucosidase (agdB) (Table 3). The transcription factor gene amyR was also induced on maltose compared to xylose (2.6 fold), indicating that some transcriptional regulation of the amyR gene itself occurs as well. As expected, the amyR transcript was absent from the ΔamyR strain (Table 3). Gene An15g03940, encoding a protein with high similarity (68%) to a Candida intermedia glucose/xylose symporter (Leandro et al., 2006), was 2.3-fold induced by maltose; this induction was not observed in the ΔamyR strain. The seventh induced gene (An11g02550) encodes a protein highly similar (72%) to Kluyveromyces lactis phosphoenolpyruvate carboxykinase (Kitamoto et al., 1998) which functions in gluconeogenesis by catalyzing the conversion of oxaloacetate into phosphoenolpyruvate. A similar comparative analysis was performed for samples taken after 8 h of growth. A total of 29 genes were significantly induced by maltose in comparison to xylose, and 167 genes were 2 fold higher expressed in N402 compared to the ΔamyR strain. By combining the two sets, we identified 19 genes which were induced by maltose and whose induction was AmyR dependent (Fig. 3, Table 3). Ten of these genes encode proteins involved in carbohydrate transport and metabolism, from which 6 were also identified as differentially expressed after 2 h (aamA, glaA, agdA, agdB, amyR, An15g03940). Additionally identified genes included amyC, encoding an extracellular α-amylase, three genes encoding putative sugar transporters, and several other genes belonging to various functional categories (Table 3).

Alpha-glucan acting enzymes in Aspergillus niger
44
Fig. 3. Results of microarray analysis for maltose induced and AmyR dependent genes. Venn-diagram showing the number of genes induced on maltose compared to xylose in A. niger N402, and the number of genes induced in N402 compared to the ΔamyR deletion strain. The number of genes both induced by maltose and dependent on AmyR is indicated in bold. Both sets were analysed 2 h and 8 h after transfer from a preculture.
Tabl
e 3A
Ove
rvie
w o
f 7 m
alto
se in
duce
d an
d A
myR
dep
ende
nt g
enes
in A
. nig
er g
row
n fo
r 2 h
afte
r tra
nsfe
r fro
m p
recu
lture
. Fo
ld in
duct
ion
OR
F no
. N
402
mal
tose
N40
2 xy
lose
Δam
yR
mal
tose
mal
tose
vs
xyl
ose
N40
2 vs
Δam
yR
P- valu
e Pu
tativ
e fu
nctio
n
Car
bohy
drat
e tr
ansp
ort a
nd m
etab
olis
m
An1
1g03
3401
3.15
± 0
.122
P
0.08
± 0
.04
A
0.13
± 0
.09
A
38.6
24
.2
0.08
0 ac
id α
-am
ylas
e A
amA
An0
4g06
920
85.9
6 ±
22.5
4 P
6.
24 ±
0.9
8 P
5.
03 ±
0.0
3 P
13
.8
17.1
0.
064
extra
cellu
lar α
-glu
cosi
dase
Agd
A
An0
1g10
930
96.5
0 ±
31.5
0 P
10
.47
± 2.
59
P
8.35
± 2
.13
P
9.2
11.5
0.
062
extra
cellu
lar α
-glu
cosi
dase
Agd
B
An0
3g06
550
117.
80 ±
43.
16
P
13.7
8 ±
4.14
P
5.
85 ±
0.4
7 P
8.
5 20
.1
0.07
2 gl
ucoa
myl
ase
Gla
A
An0
4g06
910
14.0
2 ±
1.24
P
5.
47 ±
0.8
3 P
0.
05 ±
0.0
1 A
2.
6 30
9.8
0.01
4 tra
nscr
iptio
nal r
egul
ator
Am
yR
An1
5g03
940
152.
30 ±
19.
00
P
65.7
4 ±
3.04
P
70
.59
± 2.
88
P
2.3
2.2
0.07
4 pu
tativ
e m
onos
acch
arid
e tra
nspo
rter
Ener
gy
An1
1g02
550
58.8
7 ±
3.86
P
10
.13
± 1.
41
P
25.2
7 ±
5.35
P
5.
8 2.
3 0.
033
puta
tive
phos
phoe
nolp
yruv
ate
carb
oxyk
inas
e
1 Gen
e na
me
in b
old
indi
cate
s tha
t the
gen
e is
>2-
fold
indu
ced
on m
alto
se in
an
Am
yR d
epen
dant
way
afte
r bot
h 2
and
8 ho
urs.
2 Th
e ex
pres
sion
lev
el w
as b
ased
on
the
geom
etric
mea
n va
lue
of t
he d
uplic
ate
sam
ples
and
the
dev
iatio
n va
lues
bet
wee
n th
e du
plic
ate
sam
ples
are
indi
cate
d. P
, M o
r A, r
epre
sent
ing
dete
ctio
n ca
lls fo
r pre
sent
, mar
gina
l or a
bsen
t res
pect
ivel
y. T
he P
-val
ue w
as b
ased
on
1-w
ay
AN
OV
A a
naly
sis.
Gen
es w
ere
divi
ded
over
diff
eren
t fun
ctio
nal g
roup
s acc
ordi
ng to
Fun
Cat
(Rue
pp e
t al.,
200
4).
Tabl
e 3B
Ove
rvie
w o
f 19
mal
tose
indu
ced
and
Am
yR d
epen
dent
gen
es in
A. n
iger
gro
wn
for 8
h a
fter t
rans
fer f
rom
pre
cultu
re.
Fold
indu
ctio
n O
RF
no.
N40
2 m
alto
se
N
402
xylo
se
Δa
myR
m
alto
se
m
alto
se
vs x
ylos
e N
402
vs
Δam
yR
P-va
lue
Puta
tive
func
tion
Car
bohy
drat
e tr
ansp
ort a
nd m
etab
olis
m
A
n11g
0334
01 65
.26
± 7.
48
P
0.58
± 0
.57
A
0.08
± 0
.01
A
113.
0 80
8.4
0.00
4 ac
id α
-am
ylas
e A
amA
A
n01g
1093
0 49
.46
± 3.
42
P
2.37
± 0
.08
P
6.84
± 1
.99
P
20.9
7.
2 0.
006
extra
cellu
lar α
-glu
cosi
dase
Agd
B
An0
4g06
920
179.
10 ±
13.
90
P
9.89
± 3
.63
P
3.36
± 0
.46
P
17.6
51
.7
0.01
2 ex
trace
llula
r α-g
luco
sida
se A
gdA
A
n02g
0354
0 62
.79
± 8.
36
P
8.81
± 0
.61
P
0.96
± 0
.82
P,A
6.
7 61
.9
0.01
2 pu
tativ
e he
xose
tran
spor
t pro
tein
Mst
C
An1
5g03
940
144.
40 ±
6.8
0 P
35
.93
± 2.
72
P
3.04
± 0
.03
P,M
4.
0 47
.6
0.00
3 pu
tativ
e m
onos
acch
arid
e tra
nspo
rter
An0
3g06
550
239.
30 ±
2.0
5 P
83
.03
± 4.
35
P
19.0
3 ±
1.75
P
2.
9 12
.6
0.01
2 gl
ucoa
myl
ase
Gla
A
An0
4g06
930
4.36
± 0
.50
P
1.50
± 0
.16
P
1.40
± 0
.03
P
2.9
3.1
0.06
2 ex
trace
llula
r α-a
myl
ase
Am
yC
An0
4g06
910
18.0
9 ±
1.25
P
6.
52 ±
0.1
2 P
0.
12 ±
0.0
4 A
2.
8 15
3.6
0.02
7 tra
nscr
iptio
nal r
egul
ator
Am
yR
An0
9g04
810
18.4
7 ±
0.44
P
6.
46 ±
0.3
5 P
2.
52 ±
0.0
2 P
2.
9 7.
4 0.
003
puta
tive
hexo
se tr
ansp
orte
r A
n12g
0745
0 13
7.60
± 1
3.00
P
51
.57
± 11
.82
P
52.5
0 ±
6.31
P
2.
7 2.
6 0.
064
suga
r/H+
sym
porte
r Mst
A
Ener
gy
An0
3g06
270
4.72
± 0
.29
P
1.59
± 0
.14
A
0.20
± 0
.25
A
3.3
25.6
0.
035
puta
tive
isoa
myl
alc
ohol
oxi
dase
A
n16g
0601
0 5.
18 ±
1.5
5 P
1.
80 ±
0.3
9 P
0.
22 ±
0.0
2 A
2.
9 23
.8
0.03
2 pu
tativ
e ph
osph
ogly
cera
te m
utas
e A
min
o ac
id tr
ansp
ort a
nd m
etab
olis
m
A
n03g
0028
0 8.
35 ±
0.9
0 P
3.
57 ±
0.7
4 P
,A
0.12
± 0
.06
A
2.3
68.0
0.
050
sim
ilarit
y to
tyro
sina
se p
rote
in
Cel
l res
cue,
def
ence
and
viru
lenc
e
An1
4g05
730
3.99
± 0
.59
P
0.23
± 0
.03
A
1.37
± 0
.73
P,A
18
.8
3.2
0.03
7 si
mila
rity
to in
tegr
al m
embr
ane
prot
ein
A
n03g
0029
0 5.
96 ±
0.8
9 P
1.
68 ±
0.4
7 P
0.
64 ±
0.2
1 P
3.
5 9.
3 0.
068
sim
ilarit
y to
inte
gral
mem
bran
e pr
otei
n
An0
6g00
490
8.14
± 0
.89
P
3.81
± 0
.30
P,A
2.
23 ±
0.4
4 A
2.
1 3.
7 0.
066
sim
ilarit
y to
inte
gral
mem
bran
e pr
otei
n Pr
otei
n fa
te
An1
8g04
260
5.44
± 0
.40
P
2.57
± 0
.07
P
1.77
± 0
.28
M,A
2.
1 3.
1 0.
056
sim
ilarit
y to
UD
P-g
alac
tose
tran
spor
ter
uncl
assi
fied
An1
6g01
290
15.0
4 ±
0.28
P
5.
22 ±
1.6
9 P
,A
1.97
± 0
.26
A
2.6
6.9
0.02
7 un
know
n
An0
9g06
130
12.6
2 ±
2.51
P
5.
41 ±
0.9
3 P
2.
69 ±
0.3
0 P
,A
2.3
4.7
0.05
9 un
know
n

Chapter 2 Sequence and expression analysis
47
Discussion In the present study, we have mined the full genome sequence of A. niger strain CBS 513.88 (Pel et al., 2007) for the presence of glycoside hydrolases belonging to families GH13, GH15 and GH31. Members of these protein families in aspergilli, including A. niger, have been studied extensively, mainly because of their industrial relevance. Nevertheless, our study revealed the presence of several groups of enzymes that had not been identified previously. These novel enzymes are conserved among several Aspergillus species as well as other Ascomycetes, indicating that they may play an important role in fungal metabolism. The GH13 family in A. niger contains six separate groups of amylase-type enzymes (Fig. 1), of which three groups had not been described thus far. The best described group comprises the extracellular α-amylases, and is now extended with AmyC in addition to the three known α-amylases. Overexpression of the amyC gene in A. niger resulted in increased levels of endo-α-amylase activity in the medium, indicating that this gene indeed encodes an extracellular α-amylase (Van der Kaaij and Yuan, unpublished results). The relatively low expression of amyC compared to other starch degrading enzymes may explain why this protein has not been identified previously. The localization of the amyC gene in the genome is noteworthy, as it is part of a small cluster of amylolytic genes (with agdA) and their transcriptional regulator gene amyR. A similar organization is observed in the genomes of other aspergilli such as A. nidulans and A. oryzae RIB40 (Gomi et al., 2000). In A. fumigatus the same cluster is extended with a glucoamylase (similar to GlaA). Gene clusters of transcriptionally co-regulated genes in filamentous fungi are often involved in the same process, e.g. secondary metabolite production (Woloshuk et al., 1995) or catabolism of amino acids (Hull et al., 1989). A possible function of AmyC might be to act as a scouting enzyme for the presence of starch, resulting in the subsequent activation of AmyR by starch derived molecules (maltose or isomaltose). The lack of detectable expression of amyA and amyB in A. niger N402 is puzzling. Both genes were expressed in A. niger strain CBS 513.88 in a fed-batch fermentation using glucose as a carbon source (Pel et al., 2007), but batch cultivation of A. niger N402 on glucose did not result in detectable expression in microarray studies (E. Martens, H. Kools, P. Schaap, pers. comm.). These results indicate a difference between both strains with regard to transcriptional regulation of amyA/B. Alternatively, the amyA/B gene sequences may differ between the two strains, so that expression in N402 is not observed on the microarrays. On the other hand, our results are in line with the observation that the A. niger MGG029ΔaamA strain (N402 background) with gene

Alpha-glucan acting enzymes in Aspergillus niger
48
disruptions for both aamA and glaA grows very poorly on starch (Weenink et al., 2006), confirming that AmyA and AmyB do not contribute significantly to the amylolytic potential of N402. The A. niger acid amylase (encoded by aamA) in the CBS 513.88 strain does not contain a predicted SBD, while its homologues in A. nidulans, A. fumigatus and other aspergilli contain a full length SBD (Suppl. Table 2). Comparison of the aamA gene and its 3’ flanking region with homologous genes revealed that it harbours a deletion of 230 nucleotides right after the part encoding the (β/α)8 barrel, resulting in a stop codon. The genome sequence continues by encoding part of a predicted SBD, but this is not translated as part of the acid amylase (Suppl. Fig. 2). This organization of aamA and its homologues resulting in expression of a protein with or without SBD, was confirmed by PCR analysis. Interestingly, the AamA protein in A. niger N402 contains a functional SBD, as was confirmed by its purification from culture fluid of a N402 ΔglaA strain, and subsequent demonstration of the SBD with specific antibodies (personal communication D. Archer, University of Nottingham, UK). In this study we have identified three new groups within the GH13 family that had not been described before in aspergilli: α-amylase-like proteins predicted to be GPI-anchored enzymes (group II) (subfamily GH13_1, as defined by Stam et al., 2006), intracellular α-glucosidases (group III) (unknown subfamily) and intracellular α-amylases (group IV) (subfamily GH13_5). Detailed biochemical studies of two GPI-anchored proteins revealed that they are α-glucanotransferases, using donor substrates with α-(1,4)-glycosidic bonds and at least five anhydroglucose units. The reaction products reached a degree of polymerization of at least 30 glucose residues, all linked via α-(1,4)-glycosidic bonds (van der Kaaij et al., 2007). The expression analysis of the agt genes showed that agtA and agtB are expressed during submerged vegetative growth irrespective of the carbon source (Fig. 2). Their expression was not affected in the ΔamyR strain, indicating that these genes are not expressed in relation to starch metabolism. The Agt enzymes are homologues of a group of four GPI-anchored amylase-like proteins recently identified in S. pombe (Aah1-4p), which have similarly aberrant conserved regions (Table 2). A deletion of one of these homologues, aah3, resulted in aberrant morphology of the cell and an increased sensitivity towards cell wall degrading enzymes (Morita et al., 2006). A similar phenotype was observed for an A. niger knockout of agtA (van der Kaaij et al., 2007). Such a phenotype can be explained if α-glucan synthesis in the cell wall is affected. The clustering of agt genes with genes encoding α-glucan synthases in all four Aspergillus species studied, and the expression pattern of the A. niger agt genes both support the hypothesis that Agt enzymes play a role in α-glucan synthesis.

Chapter 2 Sequence and expression analysis
49
A. niger contains two predicted intracellular α-amylases, AmyD and AmyE, that belong to the recently identified subfamily GH13_5 (Stam et al., 2006). A homologous gene in H. capsulatum, AMY1, was shown to be essential for the formation of α-(1,3)-glucan in the cell wall (Marion et al., 2006). The possibility of a role for A. niger amyE in the cell wall is strengthened by its location in the genome: amyE is located next to the agtA gene, and agsE is located in close proximity of these genes. A similar cluster of genes, containing a homologue of amyE as well as ags and agt genes, is present in both A. nidulans and A. oryzae (Suppl. Table 2). The clustering of some of the amyD/E homologues with α-glucan synthases and GPI-anchored glucanotransferases is another indication that they might be involved in the same process of α-glucan formation or maintenance. Both the GPI-anchored α-glucanotransferases and the family GH13_5 α-amylase like proteins are only present in fungi with cell wall α-glucan and not in the true yeasts, which lack this type of cell wall glucans. The expression levels of amyD and amyE were below detection level, and thus could not confirm nor refute the possible involvement of these proteins in cell wall α-glucan synthesis in A. niger. The biochemical characterization of AmyD is described in chapter 5. At this moment, no function can be assigned to the predicted intracellular α-glucosidases AgdC and AgdD and their homologues identified in other fungi. Analysis of the expression levels of agdC and agdD did not yield any indication of their physiological role. The enzymes show similarity to a Bacillus α-glucosidase as well as to a maltase from C. albicans, and might therefore be involved in the intracellular degradation of maltose. Alternatively, these enzymes could play a role in trehalose metabolism, as they show some similarity to bacterial trehalose-6-phosphate hydrolases. Trehalose is known to be a common reserve carbohydrate in fungi including A. niger (Wolschek and Kubicek, 1997;Arguelles, 2000). The GH31 protein family in A. niger consists of seven members, many of which have unknown functions. Previously identified agdA (Nakamura et al., 1997) showed the expected regulation by AmyR, similar as observed for the α-glucosidase AgdB in A. nidulans (Nakamura et al., 2006). A. niger contains a second α-glucosidase (agdB) which showed a similar expression pattern as the agdA gene (Fig. 2). Orthologues of A. niger agdB are found in other aspergilli but none of them has been characterized biochemically (Suppl. Fig. 1C). A. niger seems to lack orthologues of two recently characterized proteins from A. nidulans: AN7345.3, with both α- and β-glucosidase activity (Bauer et al., 2006) and A. nidulans AgdB (AN8953.) which has transglycosylation activity (Kato et al., 2002b) (Suppl. Fig. 1C).

Alpha-glucan acting enzymes in Aspergillus niger
50
One of the GH31 family members in A. niger, designated AxlA, could be classified as a putative xylosidase, based on two observations. First, the gene was strongly induced on xylose, indicating that this gene might be part of the xylanolytic spectrum of A. niger. Second, the AxlA protein clusters with a subgroup of GH31 family members which also include the A. nidulans AN7505.2 gene which has been characterized as an α-xylosidase (Bauer et al., 2006). A surprising finding from the genome mining of A. niger was the high number of GH enzymes with a predicted intracellular location. This group included not only the starch-acting enzymes described here, but also intracellular glycoside hydrolases with other predicted activities such as β-glucosidase, chitinase, β-mannosidase, β-xylosidase, rhamnosidase and invertase activity (Pel et al., 2007;Goosen et al., 2007). Intracellular enzymes with these predicted activities are also present in various other Aspergillus genomes. Most of these proteins were predicted to be exo-acting, i.e. releasing terminal mono- or disaccharide residues from the non-reducing end of the substrate. In A. niger, the genes encoding these proteins are expressed at a relatively low level compared to the extracellular starch degrading enzymes, sometimes even below the detection limit. In a recent publication, expression of intracellular chitinases in A. nidulans was found to be induced during the autolysis phase of the culture (Yamazaki et al., 2007). A role for these and other predicted intracellular proteins during the autolysis phase would explain their low expression levels observed in A. niger during vegetative growth, as well as their lack of secretion signals. The genome wide expression analysis identified only six genes that were >2-fold induced by maltose in an AmyR dependent way after both 2 and 8 h of induction. Among them are genes encoding two well known starch degrading enzymes (aamA, glaA), two (putative) α-glucosidases (agdA, agdB), a putative sugar transporter protein (An15g03940) and the AmyR transcription factor itself. The specificity of the sugar transporter encoded by the An15g03940 gene has not been examined, but from its expression profile it is likely that it functions as glucose or maltose transporter. Three additional sugar transporters were higher expressed on maltose than on xylose after 8 h (Table 3B). One of them, MstA has been functionally characterized as a high affinity glucose transporter (Vankuyk et al., 2004) which is in line with its expression profile. The two additional sugar transporter encoding genes (mstC and An09g04810) showed an interesting expression pattern. Their expression was induced on maltose (6.7 and 4.0-fold increase on maltose versus xylose), and their expression on xylose was also relatively high. In the ΔamyR strain both their expression levels were strongly reduced, also relative to the xylose conditions, indicating that the deletion of AmyR might have had an indirect effect. A possible explanation is that disruption of the

Chapter 2 Sequence and expression analysis
51
AmyR transcription factor resulted in low levels of extracellular enzymes converting maltose to glucose, and consequently stress due to low glucose levels. The low availability of glucose might be a signal to downregulate mstC and An09g04810 if these genes encode low affinity sugar transporters. Interestingly, the amyR gene itself was also induced by the presence of maltose and belongs to the group of co-regulated genes, suggesting that its regulation takes place, at least partly, at the transcriptional level. Furthermore, conserved AmyR binding sites are present in the promoters of all co-regulated genes, with the exception of aamA (Table 1A). Possibly a currently unknown AmyR binding site different from the consensus binding site is present in the aamA promoter. The results presented in this paper suggest that A. niger can metabolize maltose by inducing the expression of a limited amount of enzymes and sugar transporters. Other members of the GH13, GH15 and GH31 protein families might function in starch metabolism in different conditions (solid state growth, or influenced by pH, nitrogen metabolism, etc.), or they may be involved in other processes such as the synthesis of cell wall α-glucan, or the glycosylation of proteins. To determine the exact function of these enzymes will require further biochemical characterization in combination with the detailed analysis of gene deletion mutants.
Acknowledgements
We thank Harrie Kools (Wageningen University, The Netherlands) for performing the HMM searches and Patricia vanKuyk (Wageningen University, The Netherlands) for the amyR plasmid.
Supplementary data
Supplementary Table 1 (next page) The numeric values and Present/Absent calls for expression data, and calculated fold-changes between different conditions for all genes encoding family GH13, 15 and 31 enzymes identified in A. niger.

Fo
ld in
duct
ion
Fold
indu
ctio
n
N
402
(mal
2h)
N40
2 (x
yl2h
)
Δam
yR
(mal
2h)
mal
tose
vs
xy
lose
N40
2 vs
Δ
amyR
N
402
(mal
8h)
N
402
(xyl
8h)
Δ
amyR
(m
al8h
)
mal
tose
vs
xy
lose
N40
2 vs
Δ
amyR
aam
A
3.15
± 0
.12
P
0.08
± 0
.04
A
0.13
± 0
.09
A
38.6
24
.2
65.2
6 ±
7.48
P
0.
58 ±
0.5
7 A
0.
08 ±
0.0
1 A
11
2.9
807.
7
amyA
0.
10 ±
0.0
3 A
0.
10 ±
0.0
3 A
0.
07 ±
0.0
2 A
1.
0 1.
4 0.
12 ±
0.0
2 A
0.
11 ±
0.0
1 A
0.
09 ±
0.0
5 A
1.
0 1.
2
amyB
0.
08 ±
0.0
0 A
0.
08 ±
0.0
1 A
0.
06 ±
0.0
0 A
1.
0 1.
3 0.
26 ±
0.1
7 A
0.
10 ±
0.0
1 A
0.
08 ±
0.0
3 A
2.
5 3.
1
amyC
4.
58 ±
1.6
1 P
6.
96 ±
0.7
3 P
2.
12 ±
0.0
5 P
0.
7 2.
2 4.
36 ±
0.5
0 P
1.
50 ±
0.1
6 P
1.
40 ±
0.0
3 P
2.
9 3.
1
agtA
16
.24
± 4.
45
P
9.73
± 1
.70
P
12.4
4 ±
0.15
P
1.
7 1.
3 14
.75
± 1.
16
P
21.3
8 ±
1.06
P
17
.71
± 0.
93
P
0.7
0.8
agtB
0.
26 ±
0.1
3 A
0.
21 ±
0.0
1 A
0.
16 ±
0.0
8 A
1.
3 1.
7 10
.76
± 2.
23
P
7.56
± 0
.38
P
6.15
± 0
.45
P
1.4
1.8
agtC
0.
10 ±
0.0
1 A
1.
04 ±
0.0
5 A
0.
08 ±
0.0
0 A
0.
1 1.
3 0.
37 ±
0.2
6 A
0.
43 ±
0.2
7 A
0.
43 ±
0.0
0 A
0.
9 0.
9
agsA
0.
47 ±
0.1
1 A
0.
20 ±
0.0
1 A
0.
64 ±
0.0
2 A
2.
4 0.
7 0.
90 ±
0.1
6 A
2.
42 ±
0.6
3 P
,A
0.50
± 0
.16
A
0.4
1.8
agsB
0.
76 ±
0.1
3 A
1.
51 ±
0.1
2 P
,A
0.79
± 0
.13
A
0.5
1.0
1.18
± 0
.10
A
0.86
± 0
.14
A
0.88
± 0
.11
A
1.4
1.3
agsC
0.
13 ±
0.0
2 A
0.
17 ±
0.0
2 A
0.
12 ±
0.0
3 A
0.
8 1.
1 7.
70 ±
0.0
6 P
6.
76 ±
0.4
3 P
5.
20 ±
0.4
2 P
1.
1 1.
5
agsD
1.
21 ±
0.0
7 A
0.
82 ±
0.2
5 M
,A
0.67
± 0
.14
A
1.5
1.8
1.17
± 0
.07
P,A
0.
84 ±
0.1
3 M
,A
0.52
± 0
.17
A
1.4
2.2
agsE
5.
70 ±
3.4
0 P
18
.55
± 0.
24
P
3.71
± 0
.40
P
0.3
1.5
18.6
1 ±
3.68
P
13
.81
± 0.
93
P
8.99
± 2
.97
P
1.3
2.1
amyD
1.
01 ±
0.1
1 A
0.
24 ±
0.1
7 A
1.
08 ±
0.0
9 A
4.
1 0.
9 0.
33 ±
0.0
1 A
0.
75 ±
0.2
2 A
0.
70 ±
0.3
1 A
0.
4 0.
5
amyE
0.
42 ±
0.5
1 P
,A
1.14
± 0
.08
A
1.17
± 0
.15
P
0.4
0.4
0.37
± 0
.03
A
0.80
± 0
.03
A
0.62
± 0
.20
A
0.5
0.6
gbeA
30
.82
± 12
.98
P
10.4
8 ±
1.61
P
22
.34
± 4.
05
P
2.9
1.4
13.6
9 ±
0.35
P
20
.42
± 2.
81
P
35.1
9 ±
2.50
P
0.
7 0.
4
gdbA
22
.08
± 6.
02
P
7.86
± 1
.10
P
19.0
6 ±
0.33
P
2.
8 1.
2 9.
49 ±
1.8
3 P
23
.58
± 8.
80
P
30.2
7 ±
4.89
P
0.
4 0.
3
agdA
85
.96
± 22
.54
P
6.24
± 0
.98
P
5.03
± 0
.03
P
13.8
17
.1
179.
10 ±
13.
90
P
9.89
± 3
.63
P
3.36
± 0
.46
P
18.1
53
.3
agdB
96
.50
± 31
.50
P
10.4
7 ±
2.59
P
8.
35 ±
2.1
3 P
9.
2 11
.6
49.4
6 ±
3.42
P
2.
37 ±
0.0
8 P
6.
84 ±
1.9
9 P
20
.9
7.2
agdC
2.
93 ±
0.6
1 P
1.
34 ±
0.0
2 M
,A
2.16
± 0
.18
P
2.2
1.4
1.40
± 0
.17
A
3.96
± 0
.35
P
2.71
± 0
.72
P,M
0.
4 0.
5
agdD
0.
74 ±
0.0
1 A
0.
48 ±
0.0
4 A
0.
65 ±
0.0
9 A
1.
5 1.
1 0.
59 ±
0.1
2 A
0.
76 ±
0.5
0 A
0.
64 ±
0.2
5 A
0.
8 0.
9
agdE
21
.31
± 1.
96
P
7.61
± 0
.83
P
20.4
1 ±
2.83
P
2.
8 1.
0 14
.10
± 1.
27
P
11.6
2 ±
1.83
P
11
.28
± 0.
07
P
1.2
1.3
agdF
1.
32 ±
0.2
2 P
0.
13 ±
0.0
2 A
1.
46 ±
0.1
9 P
,M
10.4
0.
9 0.
28 ±
0.0
3 A
0.
11 ±
0.0
1 A
0.
65 ±
0.4
0 P
,A
2.5
0.4
agdG
0.
73 ±
0.0
0 A
0.
48 ±
0.0
3 A
0.
32 ±
0.0
5 A
1.
5 2.
3 0.
34 ±
0.3
6 A
0.
57 ±
0.0
4 A
0.
28 ±
0.0
3 A
0.
6 1.
2
glaA
11
7.80
± 4
3.16
P
13
.78
± 4.
14
P
5.85
± 0
.47
P
8.5
20.1
23
9.30
± 2
.05
P
83.0
3 ±
4.35
P
19
.03
± 1.
75
P
2.9
12.6
glaB
0.
13 ±
0.0
5 A
0.
30 ±
0.0
7 A
0.
07 ±
0.0
0 A
0.
4 1.
9 0.
15 ±
0.0
6 A
0.
29 ±
0.0
4 A
0.
06 ±
0.0
9 A
0.
5 2.
4
axlA
1.
95 ±
0.0
5 A
57
.37
± 10
.72
P
2.27
± 0
.15
A
0.0
0.9
3.13
± 0
.88
M,A
10
8.40
± 3
.85
P
2.56
± 0
.23
A
0.0
1.2
axlB
4.
38 ±
2.1
1 P
2.
69 ±
0.2
5 P
4.
23 ±
0.7
6 P
1.
6 1.
0 0.
86 ±
0.2
7 A
1.
89 ±
0.1
3 P
,A
2.24
± 0
.15
P
0.5
0.4
Chapter 2 Supplementary data
53
Supplementary Table 2. Inventory of all family GH13, 15 and 31 enzymes in A. fumigatus, A. nidulans and A. oryzae. The proteins were identified via BLASTP using all family GH13, 15 and 31 enzymes from A. niger as query sequences. Aspergillus species
ORF no. Accession no.
Closest A. niger homologue§
Features
GH 13 family
A. fumigatus Group I Afu4g10130 EAL89775 AmyA; 5.6E-214 SS Afu2g00710 EAL87170 AamA; 1E-196 SS, SBD Group II Afu3g00900* EAL86373 AgtA; 2.4E-204 SS Afu2g03230 EAL87420 AgtA; 3.5E-182 SS, GPI Afu2g13460 EAL93641 AgtA; 9.1E-127 SS, GPI Group III Afu3g07380 EAL92833 AgdC; 1.5E-275 Afu7g06380 EAL86834 AgdC; 1.4E-146 Afu8g07070 EAL85467 AgdC; 1.1E-139 Afu2g11610 EAL93455 AgdC; 3.8E-128 Afu2g11620 EAL93456 AgdC; 1.9E-117 Group IV Afu1g15150 EAL90846 AmyD; 2.2E-203 Group V Afu1g02140 EAL87926 GdbA; 0.0 Afu5g10540 EAL91552 GbeA; 0.0 Group VI Afu2g11270 EAL93422 AgsD; 0.0 SS Afu3g00910 EAL86374 AgsE; 0.0 SS Afu1g15440 EAL90874 AgsA; 0.0 A. nidulans Group I AN2018.3 EAA64850 AmyA; 0.0 SS AN3388.3 EAA63356 AmyA; E-104 SS AN3402.3 EAA63370 AamA; 0.0 SS, SBD Group II AN3308.3* EAA63276 AgtA; 0.0 SS, GPI AN4507.3++ EAA60850 AgtA; E-179 SS, GPI AN6324.3 EAA58708 AgtA; E-138 SS, GPI Group III AN4843.2 ABF50870 AgdC; 0.0 AN10420.3 EAA59077 AgdC; 0.0 Group IV AN3309.3 EAA63277 AmyE; 0.0 Group V AN10060.3 EAA65758 GdbA; 0.0 AN2314.3 EAA64425 GbeA; 0.0 Group VI AN5885.3 EAA58394 AgsD; 0.0 SS AN3307.3 EAA63275 AgsE; 0.0 SS A. oryzae Group I AO090120000196 P0C1B3 AmyA; 0.0 SS AO090023000944 P0C1B3 AmyA; 0.0 SS Group II AO090003001498 * BAE58540 AgtA; 0.0 SS, GPI AO090120000263 BAE62865 AgtA; 5.00E-72 SS, GPI&
Group III AO090020000176 BAE63400 AgdC; 0.0 AO090026000034 BAE59534 AgdC; 0.0 AO090103000378 BAE65811 AgdC; 0.0
Alpha-glucan acting enzymes in Aspergillus niger
54
AO090038000234 BAE64051 AgdC; e-156 AO090103000129 BAE65599 AgdC; e-149 Group IV AO090003001497 BAE58539 AmyD; 0.0 AO090005001193 BAE56147 AmyD; 0.0 Group V AO090005000884 BAE55891 GdbA; 0.0 AO090010000483 Q96VA4 GbeA; 0.0 Group VI AO090003001500 BAE58541 AgsE 0.0 SS AO090010000106 BAE66017 AgsA; 0.0 SS AO090026000523 BAE59959 AgsD;0.0 SS GH15 family
A. fumigatus Afu2g00690 EAL87168 GlaA; 1.1E-219 SS, SBD Afu3g00610 EAL86343 GlaA; 8.9E-160 SS, SBD Afu4g10140 EAL89774 GlaA;1E-63 SS, SBD Afu8g01390 EAL85029 GlaB; 2.5E-245 Afu6g08080 EAL88661 GlaB; 2.7E-208 A. nidulans AN7402.3 EAA61773 GlaA; 0.0 SS, SBD AN11143.3 EAA64118 GlaA; 0.0 SS, SBD AN5463.2 EAA62623 GlaA; 2E-32 SS, SBD A. oryzae AO090010000746 P36914 GlaA; 0.0 SS, SBD AO090003000321 BAA25205 GlaA; e-164 SS, SBD AO090138000105 BAE64481 GlaB; 0.0
GH31 family
A. fumigatus Afu4g10150 EAL89773 agdA; 0.0 SS Afu1g16250 EAL90953 agdB; 0.0 SS Afu5g03500 EAL85956 AgIIA; 0.0 SS Afu1g03140 EAL88026 AxlB; 0.0 Afu2g05400 EAL87632 AxlB; 4.2E-47 Afu1g06560 EAL88367 AgIIB; 0.0 Afu2g00490 EAL87147 AxlA; 1.9E-48 A. nidulans AN2017.3 . EAA64849 agdA; 0.0 SS AN7345.3 EAA61716 agdB; 0.0 SS AN0941.3 EAA65970 agdB; 0.0 SS AN8953.3 EAA63748 agdB; 0.0 SS AN11054.3 EAA58873 AgIIA; 0.0 SS AN7120.3 EAA61325 AxlB; 0.0 AN0280.3 EAA66153 AxlB; 0.0 AN7505.3 EAA62085 AxlB; 2E-52 AN10935.3 EAA61743 AxlB; 2E-15 AN3504.3 EAA59065 AgIIB; 0.0 A. oryzae AO090003001209 BAE58289 agdA; 0.0 SS AO090005001084 BAE56058 agdB; 0.0 SS AO090038000471 BAE64257 agdB; 0.0 SS AO090102000559 BAE61578 AgIIA; 0.0 SS AO090701000558 BAE62256 AgIIC; 0.0 SS AO090701000639 BAE62323 AxlA; 0.0 SS

Chapter 2 Supplementary data
55
A. oryzae AO090005000767 BAE55790 AxlB; 0.0 AO090005000768 BAE55791 AxlB; E-153 AO090001000649 BAE57147 AxlB; 3E-50 AO090026000111 BAE59606 AxlB; 2E-30 AO090023000288 BAE58852 AgIIB; 0.0
§These proteins were obtained by using full length sequences of A. niger GH13, 15 and GH31 family proteins in a BLASTP search to identify homologous sequences in the genomes of A. fumigatus, A. nidulans and A. oryzae. *The genes encoding these proteins are clustered with genes encoding α(1,3)-glucan synthases in the genome. ++An earlier start codon was identified resulting in a predicted signal sequence for excretion which was lacking in the original annotation. & GPI-anchoring was not predicted by the online tool of Eisenhaber et al. (2004), but according to the consensus sequence described by De Groot et al. (2003), a GPI-anchoring signal may be present. Suppl. Figure 1 (next pages) Bootstrapped phylogenetic trees of family GH13 (A), GH15 (B), and GH31 (C) enzymes from A. niger, A. fumigatus, A. nidulans and A. oryzae. Bootstrap values are indicated on the node of each branch. The tree was created with DNAMAN 4.0 using gap and extension penalties of 10 and 0.5 respectively. The scale bar corresponds to a genetic distance of 0.05 substitution per position. A description of each protein is listed in Supplementary Table 2.
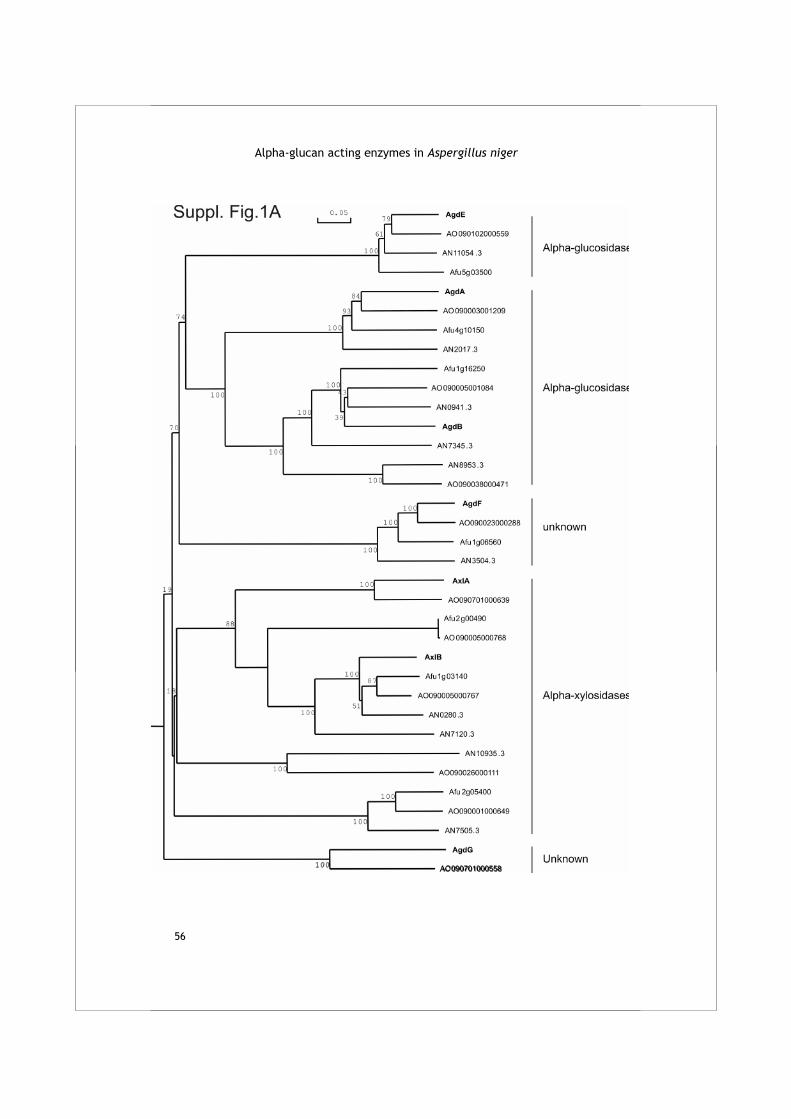
Alpha-glucan acting enzymes in Aspergillus niger
56
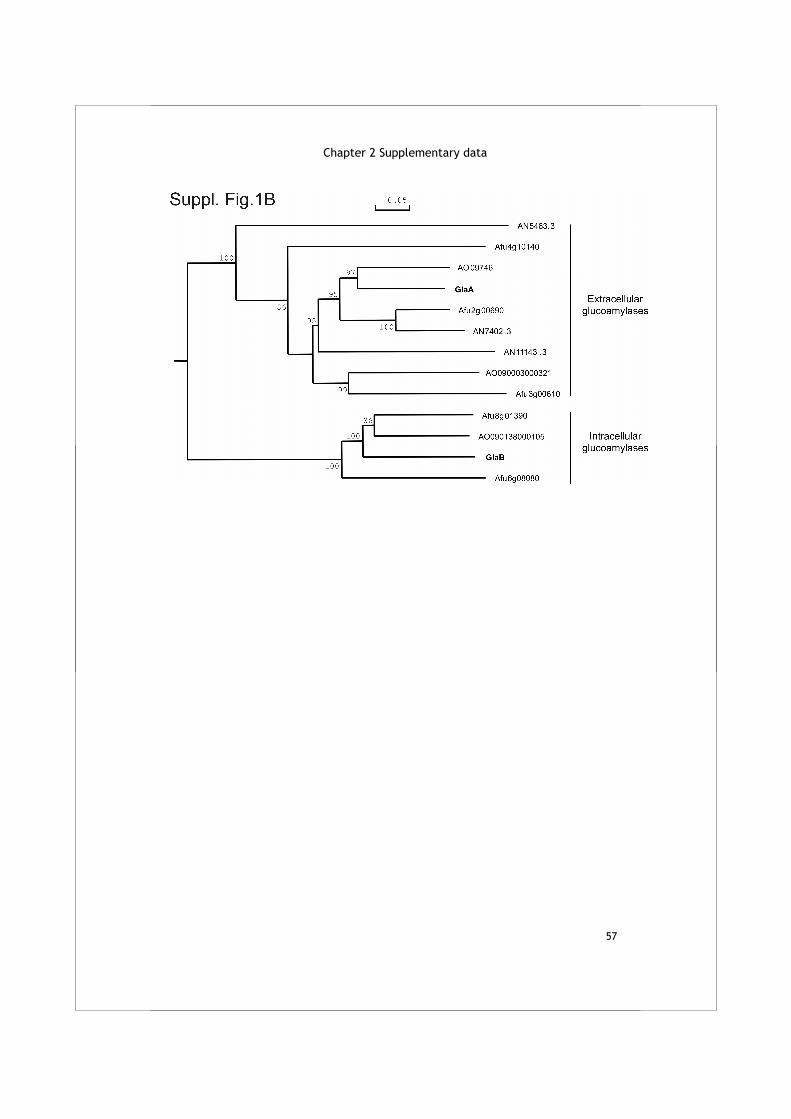
Chapter 2 Supplementary data
57
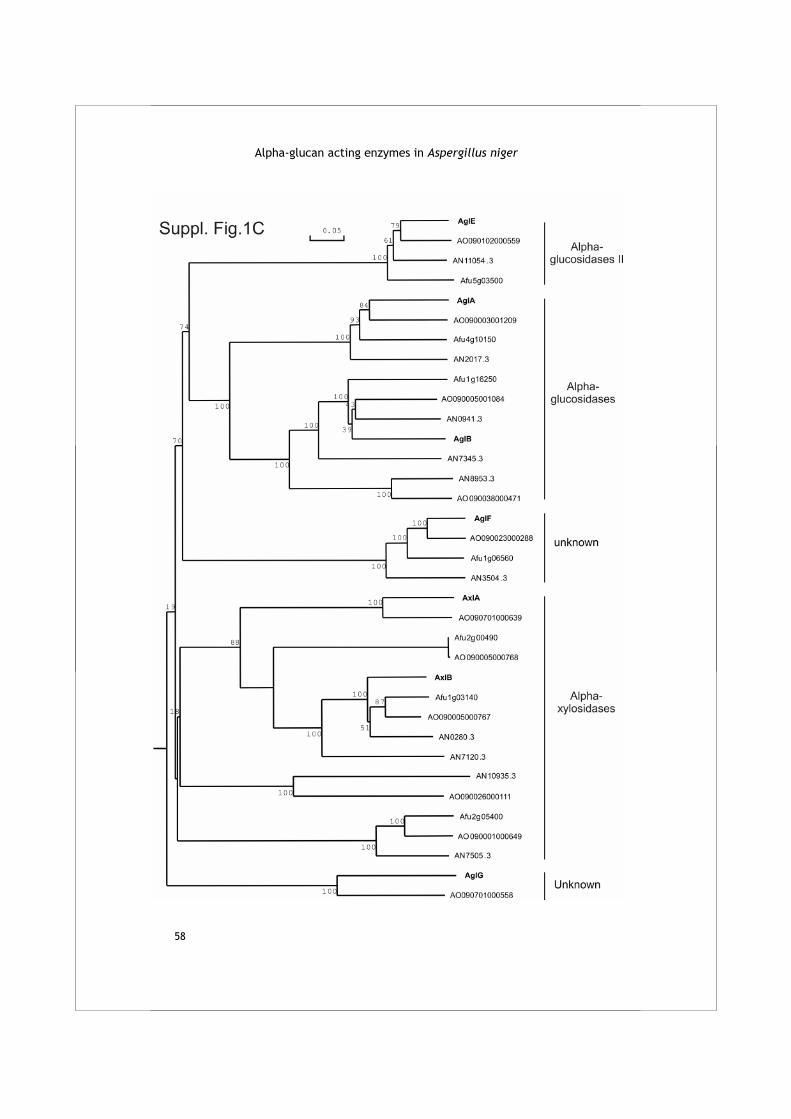
Alpha-glucan acting enzymes in Aspergillus niger
58

Chapter 2 Supplementary data
59
Suppl. figure 2. Alignments showing different AamA homologues from A. foetidus (CBS115.52), A. vadensis (CBS113.365), A. niger N402 (CBS120.49, ATCC1015), A. niger DSM (CBS513.88) and A. niger (NRRL3122). (A) Alignment of the 3’ end of aamA and homologues. The stop codon is indicated in bold, non-coding DNA sequences are underlined. (B) Alignment of the C-terminal part of AamA and homologous proteins. The C-terminal end is indicated with *. Suppl. Fig. 2A.
A.foetidus ACGTCCGGCACGAAGCTCATCGAAGCGTACACCTGCACGTCCGTGACGGTGGACTCGAAC A.vadensis ACGTCCGGCACGGAGCTCATCGAAGCGTACACCTGCACGTCCGTGACGGTGGACTCGAAC A.niger N402 ACATCCGGCACGAAGCTGATCGAAGCGTATACATGCACGTCCGTGACCGTGGACTCGAGC A.niger DSM ACATCCGGCACGAAGCTGATCGAAGCGTACACATGCACATCCGTGACCGTGGACTCGAGC A.niger NRRL ACATCCGGCACGAAGCTGATCGAAGCGTACACATGCACATCCGTGACCGTGGACTCGAGC A.foetidus GGGGATATCCCTGTGCCGATGGCTTCGGGATTACCTAGAGTTCTCCTCCCTGCTTCGGTG A.vadensis GGGGATATCCCCGTGCCGATGGCGTCGGGATTACCCAGGGTTCTTCTGCCTGCTTCGGTG A.niger N402 GGCGATATCCCCGTGCCGATGGCGTCGGGATTACCGAGAGTTCTTCTCCCCGCGTCCGTC A.niger DSM GGCGATATTCCCGTGCCGATGGCGTCGGGATTACCGAGAGTTCTTCTGCCCGCGTCCGTC A.niger NRRL GGCGATATTCCCGTGCCGATGGCGTCGGGATTACCGAGAGTTCTTCTGCCCGCGTCCGTC A.foetidus GTTGATAGTTCTTCGCTTTGTGGGGGGAGTGGTA......ACACAACCACGACCACAACT A.vadensis GTTGATAGTTCCTCGCTTTGTGGGGGGAGTGGTAG............CACAACTACAACC A.niger N402 GTCGATAGCTCTTCGCTCTGTGGCGGGAGCGGAAGTAATTCCTCAACTACAACCACAACA A.niger DSM GTCGATAGCTCTTCGCTCTGTGGCGGGAGCGGAAG......................... A.niger NRRL GTCGATAGCTCTTCGCTCTGTGGCGGGAGCGGAAG......................... A.foetidus GCTGCTACCTCCACATCCAAAGCCACCACCTCCTCTTCTTCTTCTTCTGCTGCTGCTACT A.vadensis ACAACTACCTCATCTTCCACGGCTACCTCCACCTCCACATCTAAAACCACAACAT..... A.niger N402 ACAGCTACCTCATCTTCCACTGCGACATCCAAATCCGCATCAACCTCGTCTA........ A.niger DSM ............................................................ A.niger NRRL ............................................................ A.foetidus ACTTCTTCATCATGCACCGCAACAAGCACCACCCTCCCCATCACCTTCGAAGAACTCGTC A.vadensis .CCTCTTCATCATGCACCGCAACAAGCACCACCCTCCCCATCACCTTCGAAGAACTCGTA A.niger N402 .CGTCGACGGCATGCACAGCTACCTCTACCTCCCTCGCGGTCACGTTCGAAGAGCTCGTC A.niger DSM ............................................................ A.niger NRRL ............................................................ A.foetidus ACCACTACCTACGGGGAAGAAGTCTACCTCAGCGGATCTATCTCCCAGCTCGGAGAGTGG A.vadensis ACCACCACCTACGGCGAAGAAGTCTACCTGAGCGGATCTATCTCCCAGCTCGGGGAGTGG A.niger N402 ACGACTACCTACGGGGAGGAAATCTACCTGAGCGGATCGATCTCCCAGCTTGGGGACTGG A.niger DSM ............................................................ A.niger NRRL ............................................................ A.foetidus GATACGAGTGACGCGGTGAAGTTGTCCGCGGATGATTATACCTCGAGTAACCCCGAGTGG A.vadensis GATACAAGTGACGCGGTGAAGTTGTCCGCGAATGATTATACCTCGAGTAACCCCGAATGG A.niger N402 GATACGAGTGATGCGGTGAAGATGTCCGCGGATGATTATACGTCGAGTAATCCGGAGTGG A.niger DSM ..................................ATTATACGTCGAGTAATCCGGAGTGG A.niger NRRL ..................................ATTATACGTCGAGTAATCCGGAGTGG

Alpha-glucan acting enzymes in Aspergillus niger
60
A.foetidus TCTGTTACTGTGTCGTTGCCGGTGGGGACGACCTTCGAGTATAAGTTTATTAAGGTCGAT A.vadensis TATGTTACTGTGTCGTTGCCGGTGGGGACGACCTTCGAGTATAAGTTTATTAAGGTCGAG A.niger N402 TCGGTTACTGTGACGTTGCCGGTGGGGACAACCTTTGAGTATAAGTTTATTAAGGTGGAG A.niger DSM TCGGTTACTGTGACGTTGCCGGTGGGGACCACTTTCGAGTATAAGTTTATTAAGGTGGAG A.niger NRRL TCGGTTACTGTGACGTTGCCGGTGGGGACCACTTTCGAGTATAAGTTTATTAAGGTGGAG A.foetidus GAGGGTGGAAGTGTGACTTGGGAAAGTGATCCGAATAGGGAGTATACTGTGCCTGAATGT A.vadensis GAGGATGGGAGTGTTACTTGGGAAAGTGATCCGAATAGGGAGTATACTGTGCCTGAATGT A.niger N402 TCGGATGGGACTGTTACTTGGGAGAGTGATCCGAATCGGGAGTATACGGTGCCCGAGTGT A.niger DSM TCGGATGGGACTGTTACTTGGGAAAGTGATTTGAATCGGGAGTATACGGTGCCGGAGTGT A.niger NRRL TCGGATGGGACTGTTACTTGGGAAAGTGATTCGAATCGGGAGTATACGGTGCCGGAGTGT A.foetidus GGGAGTGGGAGTGGGGAGACGGTGTTTGATACGTGGAGGTAG A.vadensis GGGAGTGGG......GAGACGGTGTTTGATACGTGGAGGTAG A.niger N402 GGGAGTGGG......GAGACGGTGGTTGATACGTGGAGGTAG A.niger DSM GGGAGTGGG......GAGACGGTGGTTGATACGTGGAGGTAG A.niger NRRL GGGAGTGGG......GAGACGGTGGTTGATACGTGGAGGTAG
Suppl. Fig. 2B. A.foetidus TSGTKLIEAYTCTSVTVDSNGDIPVPMASGLPRVLLPASVVDSSSLCGGSG.N..TTTTT A.vadensis TSGTELIEAYTCTSVTVDSNGDIPVPMASGLPRVLLPASVVDSSSLCGGSG...STTTTT A.niger N402 TSGTKLIEAYTCTSVTVDSSGDIPVPMASGLPRVLLPASVVDSSSLCGGSGSNSSTTTTT A.niger DSM TSGTKLIEAYTCTSVTVDSSGDIPVPMASGLPRVLLPASVVDSSSLCGGSGRLYVE*.. A.niger NRRL TSGTKLIEAYTCTSVTVDSSGDIPVPMASGLPRVLLPASVVDSSSLCGGSGRLYVE*.. A.foetidus TAATSTSKATTSSSSSSAAATTSSSCTATSTTLPITFEELVTTTYGEEVYLSGSISQLGE A.vadensis T..TSSSTAT..STSTSKTTTSSSSCTATSTTLPITFEELVTTTYGEEVYLSGSISQLGE A.niger N402 T.ATSSSTAT..SKSAS.TTSTSTACTATSTSLAVTFEELVTTTYGEEIYLSGSISQLGD A.foetidus WDTSDAVKLSADDYTSSNPEWSVTVSLPVGTTFEYKFIKVDEGGSVTWESDPNREYTVPE A.vadensis WDTSDAVKLSANDYTSSNPEWYVTVSLPVGTTFEYKFIKVEEDGSVTWESDPNREYTVPE A.niger N402 WDTSDAVKMSADDYTSSNPEWSVTVTLPVGTTFEYKFIKVESDGTVTWESDPNREYTVPE A.foetidus CGSGSGETVFDTWR* A.vadensis CGSG..ETVFDTWR* A.niger N402 CGSG..ETVVDTWR*

61
Chapter 3
Characterization of two novel, putatively cell wall associated and GPI-anchored, α-glucanotransferase enzymes of Aspergillus niger R.M. van der Kaaij, X-L. Yuan, A. Franken, A.F.J. Ram, P.J. Punt, M.J.E.C. van der Maarel and L. Dijkhuizen
Published in Eukaryotic Cell 2007 Jul;6(7):1178-88.
Abstract In the genome sequence of Aspergillus niger CBS 513.88 three genes were identified with high similarity to fungal α-amylases. The derived protein sequences were in two ways aberrant from all described fungal α-amylases: they were predicted to be glycosylphosphatidylinositol-anchored, and some highly conserved amino acids of the α-amylase family were absent. We expressed two of these enzymes in a suitable A. niger strain and characterized the purified proteins. Both enzymes showed transglycosylation activity on donor substrates with α-(1,4)-glycosidic bonds and at least five anhydroglucose units. The enzymes, designated AgtA and AgtB, produced new α-(1,4)-glycosidic bonds and therefore belong to the group of the 4-α-glucanotransferases (EC 2.4.1.25). Their reaction products reached a degree of polymerization of at least 30. Maltose and larger maltooligosaccharides were the most efficient acceptor substrates, although AgtA also used small nigerooligosaccharides containing α-(1,3)-glycosidic bonds as acceptor substrate. An agtA knockout of A. niger showed an increased susceptibility towards the cell wall disrupting compound Calcofluor White, indicating a cell wall integrity defect in this strain. Homologues of AgtA and AgtB are present in other fungal species with α-glucans in their cell walls, but not in yeast species lacking cell wall α-glucan. The possible role for these enzymes in the synthesis and/or maintenance of the fungal cell wall is discussed.

Alpha-glucan acting enzymes in Aspergillus niger
62
Introduction Aspergillus niger is a filamentous ascomycete fungus with a worldwide distribution. As a saprophyte, the fungus produces and secretes a large variety of extracellular enzymes especially proteases and polysaccharide hydrolases to convert plant cell walls and storage compounds into growth substrates (see e.g. Duarte and Costa-Ferreira (1994), Martens-Uzunova et al. (2006)). This quality is exploited for the production of enzymes for the food and feed industry on a large scale. Recently, the full genome sequence of A. niger CBS 513.88 was determined and annotated (Pel et al., 2007). A high level of synteny was observed between A. niger and other sequenced aspergilli, although more extracellular hydrolytic enzymes were annotated in A. niger. A detailed full genome search showed the presence of a considerable number of previously unknown, predicted enzymes belonging to the α-amylase superfamily. The α-amylase superfamily (Kuriki and Imanaka, 1999) comprises a large variety of enzymes that are active towards polysaccharides with α-glycosidic linkages such as starch and glycogen (MacGregor et al., 2001). Most members of this family are either involved in production of storage compounds such as glycogen or starch, or degradation of these compounds as extracellular carbon and energy sources. The tertiary structure of these enzymes is characterized by a (β/α)8 barrel containing four highly conserved amino acid regions that form the catalytic site (MacGregor et al., 2001) (see also the CAZy website at http://www.cazy.org). Based on sequence similarity, members of the α-amylase superfamily are divided over glycoside hydrolase (GH) families 13, 70 and 77. Here, we focus on family GH13, which mostly contains enzymes that perform a hydrolytic reaction, i.e. they cleave an α-glycosidic linkage using water as an acceptor molecule. The best known hydrolytic enzyme of family GH13 is α-amylase (EC 3.2.1.1), which hydrolyses the internal α-(1,4)-glycosidic bonds in starch, glycogen and maltooligosaccharides (α-(1,4) linked glucose oligomers) producing shorter maltooligosaccharides and maltose (O-α-D-Glc-(1,4)-α-D-Glc). Other GH13 family members perform a transglycosylation (or glucanotransferase) reaction in which they cleave an α-(1,4) glycosidic bond in a donor substrate and subsequently do not use water but instead another oligosaccharide as acceptor substrate to form a new α-glycosidic linkage. A. niger produces a number of extracellular enzymes classified as members of GH13, which are involved in the degradation of starch. These include acid amylase, which is well known for its stability at low pH (Boel et al., 1990) and two almost identical α-amylase enzymes (AmyA/B) (Korman et al., 1990). We have identified several additional members of family GH13 in the A. niger genome sequence, three of which clustered together in the phylogenetic tree of the GH13 family members and showed a

Chapter 3 Two novel alpha-glucanotransferases
63
typical glycosylphosphatidylinositol (GPI) anchoring signal at the protein C-terminus. A GPI-anchor serves as a targeting signal to the cell membrane and/or the cell wall. The cell wall of aspergilli has been shown to contain four major classes of polysaccharides: chitin, α-glucan, β-(1,3)-glucan and galactomannan (Bardalaye and Nordin, 1977;Johnston, 1965;Blumenthal and Roseman, 1957;Stagg and Feather, 1973). In addition, it contains covalently attached cell wall proteins (Brul et al., 1997). The α-glucan fraction is composed of an α-(1,3)-glucan with 3-10% α-(1,4)-glycosidic linkages (Horisberger et al., 1972;Johnston, 1965), and nigeran, a glucan with alternating α-(1,3) and α-(1,4) glycosidic bonds (Bobbitt et al., 1977). Alpha-(1,3)-Glucan synthases have been identified and functionally studied in a number of different fungi (Hochstenbach et al., 1998;Damveld et al., 2005b;Beauvais et al., 2005;Reese and Doering, 2003). These enzymes are large proteins (~2400 amino acids long) consisting of three conserved domains which are predicted to be involved in the synthesis, transport and crosslinking of the α-(1,3)-glucan (Hochstenbach et al., 1998). Detailed structural studies in Schizosaccharomyces pombe have revealed that the α-glucan is a linear glucose polymer of 260 residues in length consisting of two α-(1,3)-glucan chains that are interconnected via α-(1,4) linked glucose residues (Grün et al., 2005). A mutation in the N-terminal part of the α-(1,3)-glucan synthases (the proposed crosslinking domain) abolished the linkage between the two α-(1,3)-glucan chains indicating that this part of the protein acts as a glucanotransferase, connecting the glucan chains. Recently, two types of putative family GH13 enzymes have been shown to play a role in fungal cell wall α-(1,3)-glucan formation. Marion et al. (2006) showed the involvement of a putative α-amylase (Amy1p) in the formation of α-(1,3)-glucan in the cell wall of Histoplasma capsulatum. In this pathogenic fungus, α-(1,3)-glucan is known to play an important role in virulence (Rappleye et al., 2004). A functional knockout of AMY1 resulted in a lack of α-(1,3)-glucan formation and decreased virulence. The second α-amylase homolog, Aah3p, was studied in S. pombe (Morita et al., 2006). A knockout strain of this GPI-anchored protein was hypersensitive towards cell wall degrading enzymes, and showed aberrant cell shape. The enzymatic activities of Amy1p and Aah3p have not been studied. In this paper, we report the first biochemical characterization of two family GH13 enzymes putatively involved in α-(1,3)-glucan formation. We expressed and purified two GPI-anchored enzymes from A. niger, both homologues of Aah3p from S. pombe. The biochemical characterization showed that the two A. niger enzymes are family GH13 α-glucanotransferases, making them the first of their kind to be described for

Alpha-glucan acting enzymes in Aspergillus niger
64
fungi. A gene knockout of one of the enzymes in A. niger resulted in increased sensitivity towards Calcofluor White (CFW), a cell wall disrupting compound.
Materials and methods
Bioinformatics tools
The full genome sequence of A. niger strain CBS 513.88 was provided by DSM (Pel et al., 2007). A Hidden Markov model (HMM) profile was built using the HMMR package (Durbin and Eddy, 1998) based on the amino acid sequences of described α-amylases, which were retrieved from the CAZy website (http://www.CAZy.org) (Coutinho and Henrissat, 1999). The obtained profile was used to screen the A. niger CBS 513.88 genomic database using the WISE 2 package (Birney et al., 2004). The presence of a signal peptidase cleavage site and a GPI-attachment site were predicted by web-based search tools (http://www.cbs.dtu.dk/services/SignalP/ (Bendtsen et al., 2004) and http://mendel.imp.univie.ac.at/sat/gpi/gpi_server.html (Eisenhaber et al., 2004) respectively). The GPI-attachment prediction was confirmed by a manual comparison of the protein sequences with the consensus sequence for yeast GPI-proteins as described by De Groot et al. (2003). Amino acid sequence alignments and phylogenetic analysis were performed using MEGA3.1 (Kumar et al., 1993) and adjusted manually if necessary. Sequences from other fungal genomes were retrieved via the option genomic Blast at NCBI (http://www.ncbi.nlm.nih.gov/sutils/genom_table.cgi?organism=fungi).
Aspergillus niger strains, growth conditions and transformation
A. niger strain N402 (cspA1 derivative of ATCC9029 (Bos et al., 1988)) mRNA was used for the construction of a cDNA library. Genomic DNA of strains A. niger NRRL3122 and N402 was isolated and used as a template in PCR reactions. A. niger strain MGG029-ΔaamA (Weenink et al., 2006) was used as a host for protein overexpression. This strain, derived from strain MGG029 (prtT glaA::fleor pyrG), is deficient in the expression of several extracellular proteases, and it has no glucoamylase gene (glaA) and acid amylase gene (aamA) resulting in very poor growth on starch (Weenink et al., 2006). Strain MA70.15 (pyrG-, ku70::amdS) (Meyer et al., 2007) was used for the disruption of the agtA gene. Aspergillus strains were grown in Aspergillus Minimal Medium (MM) or Complete Medium (CM) which is MM with addition of 0.1% casaminoacids and 0.5% yeast extract (Oxoid, Basingstoke, UK) (Bennett and Lasure, 1991). Cultures for protein production were grown in CMS (CM supplemented with 2% (w/v) sucrose and 1% (w/v) glucose). Spores were obtained by growing A. niger on CM with 2% (w/v) agar

Chapter 3 Two novel alpha-glucanotransferases
65
for 4 days and scraping off the spores with 0.9% (w/v) NaCl. Liquid cultures were inoculated with 106 spores l-1 medium, and subsequently grown at 30 °C while shaking at 280 r.p.m. Transformation of A. niger was performed as described previously (Punt and van den Hondel, 1992) using lysing enzymes (Sigma, Zwijndrecht, The Netherlands). Selection of positive clones was performed on the basis of their ability to grow successively on MM containing 15 mM CsCl and 10 mM aceetamide or acrylamide as sole nitrogen source, brought about by the expression of the amdS gene (Kelly and Hynes, 1985).
Cloning procedures
All basic molecular techniques were performed according to standard procedures (Sambrook et al., 1989). E. coli TOP10 (Invitrogen, Carlsbad, U.S.A.) or DH5α (Stratagene, La Jolla, U.S.A.) were used for transformation and amplification of recombinant DNA. The primers used were obtained from Eurogentec (Seraing, Belgium) or Biolegio (Nijmegen, The Netherlands). All steps during the construction of the overexpression vectors were checked by restriction analysis, and the final constructs were checked by sequencing (GATC Biotech AG, Konstanz, Germany). Genomic DNA was isolated from A. niger N402 and NRRL3122 as described by Kolar et al. (1988). All PCR reactions were performed with 2.5 units of Pwo DNA polymerase (Roche, Indianapolis, U.S.A.), 1x buffer and 1 mM of each dNTP in a total volume of 25 μl. A cDNA library was produced from A. niger N402 grown on MM with addition of starch as sole carbon source. Primers used are indicated in Table 1. The overexpression vector for the transformation of A. niger was provided by Dr. J. Benen (Wageningen University, Wageningen, The Netherlands), and was produced as follows: Gene pgaII (encoding polygalacturonase II from A. niger) was cloned into pPROM-S (Benen et al., 1999) using NsiI and KpnI restriction sites. A NotI site was generated immediately downstream of the stop codon of the pgaII gene by site directed mutagenesis. The gene encoding acetamidase (amdS (Kolar et al., 1988)) was amplified by PCR with specific primers from plasmid p3SR2 (Wernars et al., 1985) and cloned in front of the pki-promoter region (Parenicova et al., 1998) using XbaI restriction sites, resulting in vector Ppki-pgaII-amdS. The construct for the overexpression of the chimaeric protein AgtA-SBD (a fusion between AgtA and the Starch Binding Domain (SBD) of A. niger glucoamylase GlaA) was made as follows: Genomic DNA of A. niger N402 was used as a template in a PCR reaction with specific primers to generate the DNA fragment encoding the SBD, including the linker region. The primers were designed to amplify nucleotide 3643 to 4149 (numbering according to glaA coding sequence EMBL ID AY250996). An ApaI restriction site was built into the forward primer to allow the subsequent cloning of the agtA gene fragment in frame with the

Alpha-glucan acting enzymes in Aspergillus niger
66
SBD. The SBD-encoding fragment was cloned into vector Ppki-pgaII-amdS using NsiI and NotI restriction sites, thereby replacing the pgaII gene. The cDNA fragment encoding agtA was generated by PCR with specific primers on the cDNA library. The primers were designed to amplify the gene up to nucleotide 1766 (in gene sequence), which does not include the C-terminal GPI-anchoring part. The AgtA cDNA was cloned in frame N-terminally of the SBD using NsiI and ApaI, resulting in the expression vector Ppki-agtA-SBD-amdS. Sequencing of the construct revealed one point mutation compared to the original gene sequence in the genomic database. Nucleotide 1420 (in the coding sequence of agtA) was changed from A to G, resulting in Ala474 instead of Thr474 in the derived amino acid sequence. This mutation was consistent in several independent clones and was therefore considered to be representing a strain difference between N402 and CBS 513.88. The same mutation was found in the equivalent protein sequence published by the DOE Joint Genome Institute (http://genome.jgi-psf.org/Aspni1/Aspni1.home.html).
Table 1 Primers used for the production of plasmids for overexpression and deletion of genes in A. niger. The restriction sites used for cloning are underlined.
Primer name Primer sequence Enzyme
Primers used for construction of pΔagtA
AgtAP1for ATAAGAATGCGGCCGCTGTCCTGTGTGTTCCAGCCT NotI
AgtAP2rev GCTCTAGAAATGATCAAGGGTTGCGTACA XbaI
AgtAP3for GCTCTAGATATGCTGATAGCTACAGATGG XbaI
AgtAP4rev CGGGATCCGGAGTGGATAGCTGGTAAGGC BamHI
Primers used for construction of Ppki-agtA-SBD-amdS
SBD-fw GGCCCTATGCATGGCCCTGGGCCCACCTGTGCGGCCACATCTGC NsiI, ApaI
SBD-rev CCCGCTGCGGCCGCCTACCGCCAGGTGTCAGTCAC NotI
AgtA-SBD-fw GGCCCTATGCATATGGTCTCAATGTCGGCCCTGC NsiI
AgtA-SBD-rev CCGGGAGGGCCCTCCGCACAGCCCACTGCC ApaI
Primers used for construction of Ppki-agtA-amdS and Ppki-agtB-amdS
AgtA-fw GGCCCTATGCATGTCTCAATGTCGGCCCTGC NsiI
AgtA-rev CCCGCTGCGGCCGCTTACCACATCCCCACAATCA NotI
AgtB-for GGCCCTATGCATTTTCGAAAATCCGCTTCCCTC NsiI
AgtB-rev CCCGCTGCGGCCGCTTATATCCGGAATGCCAAAAAT NotI

Chapter 3 Two novel alpha-glucanotransferases
67
The constructs for overexpression of agtA and agtB were produced as follows: the complete gene sequences of genes agtA and agtB were amplified with specific primers from genomic DNA isolated from A. niger NRRL3122 (van Dijck et al., 2003). The primers contained restriction sites for NsiI and NotI, which were used to clone the gene fragments into vector Ppki-pgaII-amdS, thereby replacing the pgaII gene. This resulted in the vectors Ppki-agtA-amdS and Ppki-agtB-amdS.
Protein production, purification and detection procedures Production of AgtA-SBD
Several stable transformants were checked for their level of extracellular production of the chimaeric protein AgtA-SBD by Western blotting with polyclonal antiserum raised against purified SBD kindly provided by Dr. D. Archer and Dr. D. McKensie (University of Nottingham & IFR Norwich, UK) (Le Gal-Coeffet et al., 1995). SDS-PAGE and Western blots were performed according to standard protocols. Immobilon-P (Millipore, Billerica, U.S.A.) was used as blotting membrane. The untransformed strain was included as negative control while purified SBD (Van der Maarel et al., unpublished data) served as positive control for immunodetection. The A. niger transformant producing highest levels of AgtA-SBD was grown in a 5 l batch fermentor (New Brunswick Scientific, Edison, U.S.A.) inoculated with 100 ml culture pregrown on Potato Dextrose Broth (Difco). The medium used for the batch fermentation consisted of the following components: 3% (w/v) glucose, 117 mM NaNO3, 3.25 mM MgSO4.7H2O, 14.7 mM KH2PO4, 0.69 mM CaCl2.H2O, 0.5% Yeast Extract and spore elements. Fermentor conditions were pH 4.5, a temperature of 30 °C and aeration at 1.2 l min-1. The feed contained 20% (w/v) glucose, 74 mM KH2PO4, 350 mM NaNO3, 1% yeast extract and 1% tryptone and was added at a rate of 5 ml h-1. Three days after starting the feed the growth medium was collected by filtration over miracloth (Calbiochem, EMD Biosciences, La Jolla, CA, U.S.A.). The pH of the medium was subsequently adjusted to pH 6 with 1 M NaOH and AgtA-SBD was extracted from the medium via binding of the SBD to starch granules based on the procedure described by Paldi et al. (2003).1 l medium was added to 13 g waxy maize starch that was prewashed with elution buffer (10 mM NaAc pH 6), and incubated for 2 h at 4 °C while shaking gently. The starch with bound proteins was collected by centrifugation (15 min at 5000 x g) and washed once with ice-cold elution buffer followed by another centrifugation step. AgtA-SBD was eluted from the starch granules by the addition of 25 ml of elution buffer per batch of 13 g starch and subsequent incubation at 40 °C for 1 h while shaking gently. After removing the starch granules by centrifugation the protein was collected from the supernatant. A second round of binding was performed to remove residual proteins and media components. The protein was concentrated and the buffer changed to 20 mM Tris-HCl pH 8 using a Centriprep YM50 column (Millipore, Bedford,

Alpha-glucan acting enzymes in Aspergillus niger
68
U.S.A.). The sample was applied to an anion exchange column (ResourceQ 1 ml, Amersham Biosciences, New Jersey, U.S.A.) equilibrated with 20 mM Tris-HCl pH 8. Proteins were eluted with a NaCl gradient (0-1 M NaCl) at a flow rate of 1 ml min-1. AgtA-SBD was eluted as a single activity peak at 280 mM NaCl. Purification of AgtA-SBD was confirmed by Western blot analysis. Deglycosylation was performed with 850 U of Endoglycosidase H (Endo H) (New England Biolabs, Ipswich, U.S.A.) on 1.5 μg of purified protein for 20 h in a total volume of 15 µl according to the manufacturer’s instructions.
Production of AgtA and AgtB
After transformation of each of the plasmids Ppki-agtA-amdS and Ppki-agtB-amdS into A. niger MGG029-ΔaamA, 9 transformants were selected which showed the best growth on selective medium. The best transformant for overexpression of each protein was selected by growth in liquid CMS and visual inspection of protein production on SDS-PAGE. The selected transformants were grown in CMS for 3 days at 30 °C and 200 r.p.m. Mycelium was removed from the culture medium by filtration over miracloth. The medium was concentrated over a Centriprep YM-50 membrane filter and the concentrated protein was taken up in 20 mM Tris-HCl buffer pH 8. The proteins were purified via anion exchange chromatography as described above. Both AgtA and AgtB were eluted as a single activity peak at a concentration of 150 mM NaCl. At each stage of the protein purification, the protein amount was measured using the Bradford method with reagent from Bio-Rad (Hercules, U.S.A.), and purity was checked using SDS-PAGE analysis (Laemmli, 1970) and staining with Biosafe Coomassie (Bio-Rad).
Enzymatic assays
All oligosaccharides used were obtained from Sigma, except nigerotriose (O-α-D-Glc-(1,3)-α-D-Glc-(1,3)-D-Glc) which was purchased from Dextra laboratories (Reading, U.K.) and nigerose (O-α-D-Glc-(1,3)-α-D-Glc) which was a kind gift from Nihon Shokuhin Kako Co. Ltd. (Shizuoka, Japan). Lactobacillus reuteri polysaccharide was a gift from Dr. S. Kralj (University of Groningen, The Netherlands) and α-(1,3)-glucan isolated from A. nidulans which was a kind gift from Dr. B. J. Zonneveld (Leiden University, Leiden, The Netherlands). As soluble starch, Paselli SA2 with an average degree of polymerization (DP) of 50 (AVEBE, Foxhol, The Netherlands), was used. All reactions were performed at 37 °C. To determine the optimum pH for activity of AgtA and AgtB, 0.5 μg purified enzyme was incubated with 20 mM maltopentaose (a maltooligosaccharide of DP 5) in a 20 μl reaction volume for 30 min at 11 different pH values. The reaction was buffered either by 30 mM NaAc buffer at pH 4.2 to 7.0, or K2HPO4/KH2PO4 buffer at pH 6.3 to 8.0. Subsequently, 2 μl of the reaction mixture was spotted on a Thin Layer Chromatography (TLC) plate (Silica gel 60 F254, Merck,

Chapter 3 Two novel alpha-glucanotransferases
69
Darmstadt, Germany) and, after drying, the plate was run for 6 h in 75 ml running buffer (butanol/ethanol/mQ 5/5/3 (v/v/v)) in a container of 22 x 6 x 22 cm. After running the plate was dried and sprayed with 50% sulphuric acid in methanol and left to develop for 10 min at 110 °C. Hydrolyzing activity on potato starch was determined by the incubation of 1 μg purified enzyme with 600 μl 0.02%, 0.2% or 2% (w/v) dissolved potato starch in 50 mM NaAc buffer containing 1 mM CaCl2. Reactions were buffered at pH 5.5 (AgtA) or pH 4.8 (AgtB) and performed in duplicate. Samples of 50 μl were taken after several time intervals up to 4 h and used for the determination of reducing ends and glucose. The formation of reducing ends was measured with the bicinchoninic acid method (Meeuwsen et al., 2000), and glucose formation was measured with the Glucose GOD-PAP assay (Roche, Mannheim, Germany). Appropriate calibration curves and negative controls were included for all assays and reactions. Standard assay conditions for all further enzymatic reactions were as follows: 0.4 µg of purified enzyme was incubated in 20 μl 25 mM NaAc buffer pH 5.5 containing 1 mM CaCl2 and 0.01% sodiumazide in the presence of 20 mM oligosaccharide substrate and/or 4% (w/v) soluble starch or other polysaccharide, except nigeran and α-(1,3)-glucan. Nigeran and α-(1,3)-glucan were dissolved in 1 M NaOH, after which the pH of a 1% solution was adjusted to pH 5.5 with HAc. The final concentration used in reaction mixtures was 0.5% (w/v). Reaction products were detected either by TLC (as described above) or HPLC (Dionex) analysis. For HPLC analysis, 5 μl of the reaction mixture was diluted in 1.5 ml 90% dimethyl sulfoxide. Separation of oligosaccharides was achieved as described earlier by Kralj et al. (2004b).
Disruption of the agtA gene
The plasmid used to disrupt the agtA gene was constructed as follows. The DNA fragments flanking the agtA gene were amplified by PCR using N402 genomic DNA as template: 1.5 kb of 5’ flanking DNA and 1.0 kb of 3’ flanking DNA were amplified by PCR using primers AgtAP1f and AgtAP2r, AgtAP3f and AgtAP4r (Table 1), respectively. Each primer was adapted with a restriction site for further cloning. The amplified PCR fragments were digested with NotI and XbaI or XbaI and BamHI, respectively, and cloned in a three way ligation into NotI and BamHI digested pBlue-ScriptII SK to obtain plasmid pAgtAF53. Subsequently, pAgtAF53 was digested with XbaI, and ligated with the 2.7 kb XbaI fragment containing the A. oryzae pyrG gene, obtained from plasmid pAO4-13 (de Ruiter-Jacobs et al., 1989) which resulted in the agtA disruption plasmid, pΔagtA. Before transformation to MA70.15, pΔagtA was linearized with NotI. Uridine prototrophic transformants were selected by incubating protoplasts on agar plates containing MM without uridine. Transformants were purified and genomic DNA was isolated and analysed by Southern Blot analysis to identify

Alpha-glucan acting enzymes in Aspergillus niger
70
possible ΔagtA strains. Deletion of the agtA locus by homologous recombination was expected to result in appearance of a 4.8 kb fragment after digestion of the genomic DNA with KpnI and the loss of a 2.0 kb fragment which was expected in the wild-type. For the hybridization, the NotI-XbaI fragment (containing the 5’ flanking region of the agtA gene) was used. Independently obtained transformants were purified and strains with the expected hybridization pattern (MA71.1, MA71.3, MA71.4, MA71.7) were used for phenotypic analysis.
Phenotypic characterization of the ΔagtA strain and AgtA/AgtB overexpression strains.
Sensitivity towards CFW was assayed as described (Ram and Klis, 2006). Conidiospores from the control strains (MA70.15 transformed with pAO4-13 containing A. oryzae pyrG, and MGG029-ΔaamA), the ΔagtA strains (MA71.1, MA71.3, MA71.4, MA71.7), and the agtA and agtB overexpression strains were spotted on CFW-plates. All strains were also checked for their ability to grow on starch as sole carbon source by the inoculation of 3 μl of spore solution containing 106, 105, 104 or 103 spores on plates containing MM with either 1% (w/v) potato starch or 1% (w/v) glucose as carbon source. Colony growth was followed daily.
Table 2 Alignment of the generally conserved regions of the α-amylase family as present in A. niger acid amylase and α-amylase AmyA compared to homologous regions in AgtA, AgtB and AgtC from A. niger and the homologous protein Aah3p from S. pombe. Catalytic residues are underlined, generally conserved residues are indicated in bold.
Enzyme Region I Region II Region III Region IV
Acid amylase LMVDVVPNH DGLRIDSVLE YCVGEVDN NFIENHD
AmyA LMVDVVANH DGLRIDTVKH YCIGEVLD TFVENHD
AgtA LMMDTVINN DGLRIDAAKH FMTGEVLQ SFSENHD
AgtB LLLDVVINN DGLRIDAAKS FMTGEVMD NFIEDQD
AgtC LMLDIVVGD DGLRIDSVLN FTVGEGAT TFTANQD
Aah3p (S. pombe) VMLDSIVNS DGLRIDAVKM YSVGEVFS TFIENHD
Amino acid numbering 1 135 – 143 222 - 231 247 – 254 312 - 318
1 Numbering according to A. niger acid amylase

Chapter 3 Two novel alpha-glucanotransferases
71
Gene accession numbers
The full genome sequence of A. niger strain CBS513.88, has been deposited at the EMBL database with accession numbers AM270980-AM270998 (Pel et al., 2007). The locus tags of the genes studied here are: An09g03100 (AgtA), An12g02460 (AgtB) and An15g07800 (AgtC).
Results
Sequence analysis
The full genome sequence of A. niger CBS 513.88 was searched for genes encoding proteins belonging to family GH13 using a HMM profile based on known α-amylases. Apart from 3 genes encoding previously described extracellular α-amylases, 8 genes encoding for yet undescribed family GH13 proteins were identified, as well as five genes predicted to encode membrane bound α-(1,3)-glucan synthases (Pel et al., 2007). Within the group of 8 α-amylase type proteins, three were characterized by the presence of an N-terminal signal sequence for secretion and a hydrophobic C-terminal sequence predicted to act as an attachment site for a GPI-anchor (Bendtsen et al., 2004;De Groot et al., 2003). These proteins, called AgtA, AgtB and AgtC, form the topic of this study. The predicted sites for the attachment of a GPI-anchor (the ω-sites) were largely in accordance with the consensus sequence for fungal GPI-proteins as described by De Groot et al. (2003) (Fig. 1). An exception to this consensus sequence was residue Thr543 at the ω+2 position in AgtC: an alternative ω-site could be residue Ser538 instead of Ser541 predicted by the online GPI-prediction tool (Eisenhaber et al., 2004). Homologous enzymes with predicted GPI-anchoring sites were also identified in the available genome sequences of other aspergilli as well as in Neurospora crassa, Magnaporthe grisea and S. pombe. In many cases, the genes encoding the Agt homologues are located next to genes encoding predicted α-(1,3) glucan synthases. No homologous family GH13 proteins containing GPI-anchoring sites were found in the genome sequences of Saccharomycotina like Candida albicans, Kluyveromyces lactis and Saccharomyces cerevisiae. The amino acid sequences of AgtA, AgtB and AgtC show 54-56% similarity to A. niger acid amylase (Boel et al., 1990). A phylogenetic analysis of the Agt proteins, compared to the A. niger α-amylases, the homologous proteins in A. oryzae and the Aah proteins in S. pombe, is shown in Fig 2. The Agt proteins contain most of the amino acids generally conserved in family GH13 (Table 2). However, in all three protein sequences the highly conserved His143, which is part of conserved region I, is absent (numbering used is according to acid α-amylase of A. niger, unless indicated otherwise). In AgtB and AgtC, a second conserved histidine in conserved region IV is replaced by glutamate (Table 2).

Alpha-glucan acting enzymes in Aspergillus niger
72
Fig. 1 GPI-anchor specific amino acid features in fungal proteins as described previously compared to the C-terminal sequence of AgtA, AgtB and AgtC. A) Consensus sequence for GPI-attachment according to De Groot et al. (2003), with X representing any amino acid. The ω-site is indicated in bold. B) C-terminal ends of AgtA, AgtB and AgtC indicating the potential GPI-modification site. The ω-site as predicted by the online tool for the prediction of a GPI-modification site (Eisenhaber et al., 2004) is indicated in bold, an alternative ω-site is indicated in bold underlined. The hydrophobic tail is underlined.
Purification and enzyme activity of AgtA fused to a starch binding domain
The agtA gene, lacking its C-terminal anchoring domain, was fused to the starch binding domain (SBD) of the A. niger glucoamylase gene serving as a protein tag, and the fusion construct was transformed into A. niger MGG029-ΔaamA resulting in 25 stable transformants. The transformant with the highest expression level of the AgtA-SBD fusion protein in the culture medium, as determined by Western blot with polyclonal antiserum raised against purified SBD (Le Gal-Coeffet et al., 1995), was selected for larger scale protein production. AgtA-SBD was isolated from the culture medium by binding it to waxy maize starch granules, and after further purification by anion exchange chromatography a single protein band with an apparent molecular mass of about 130 kDa was obtained (Fig. 3). This band was recognized by anti-SBD antibodies, a strong indication that this band contained the AgtA-SBD fusion protein (calculated molecular mass 73.6 kDa). Treatment with Endo H to remove N-glycosylation resulted in a small decrease of the apparent molecular mass indicating that the protein was N-glycosylated (Fig. 3). The high molecular mass of the fusion protein after Endo H treatment could be caused by heavy O-glycosylation of the linker region of the SBD (Williamson et al., 1992).

Chapter 3 Two novel alpha-glucanotransferases
73
Fig. 2 Bootstrapped phylogenetic tree of the A. niger extracellular α-amylases (acid amylase AamA, α-amylase A and B, and putative α-amylase An04g06930) and α-glucanotransferases (AgtA/B/C), the homologous putative proteins identified in the genome of A. oryzae, and the S. pombe Aah proteins (SpAah1p to SpAah4p). The alignment and phylogenetic analysis were performed with MEGA version 3.1 using default settings. A bootstrapped tree was constructed with the neighbor-joining method using 500 replicates.
M MAgtA-SBD AgtA AgtB
N EH N EH N EH209
12480
49
3529
M MAgtA-SBD AgtA AgtBAgtA-SBD AgtA AgtB
N EH N EH N EH209
12480
49
3529
Fig. 3 SDS-PAGE analysis of purified AgtA-SBD, AgtA and AgtB in native form (N) and after treatment with Endo H (EH). The marker is indicated by M, the size of the marker proteins (in kDa) is indicated on the side.

Alpha-glucan acting enzymes in Aspergillus niger
74
Purified AgtA-SBD was incubated with potato starch to determine its hydrolysing activity. Hydrolysis of starch may result in the formation of glucose, maltose or longer maltooligosaccharides, the formation of which can be quantified by the measurement of the reducing ends formed during the reaction. For AgtA-SBD, a low hydrolyzing activity of 0.46 ± 0.02 μmole reducing ends mg-1 min -1 was detected when incubated with 0.2% starch, and no formation of glucose was observed. TLC analysis of the reaction mixture showed that short oligosaccharides were formed in low amounts from soluble starch (Fig. 4, lane 1). Incubation with both maltose and soluble starch resulted in the formation of more oligosaccharides than made from starch alone (Fig. 4, lane 2). No products were formed from maltose alone (result not shown).
Fig. 4 TLC analysis of reaction products of AgtA-SBD from different substrates. Enzyme (0.4 µg) was incubated with 20 mM oligosaccharide and/or 4% soluble starch for 1 h at 37 °C. The arrow indicates the level where the following samples were loaded: Marker (lane M) containing a mix of maltooligosaccharides ranging from glucose (G1) to maltoheptaose (G7), unmodified soluble starch (lane S), reaction products of AgtA-SBD incubated with: soluble starch (lane 1), maltose and soluble starch (lane 2), maltopentaose (lane 3), maltoheptaose (lane 4).

Chapter 3 Two novel alpha-glucanotransferases
75
When AgtA-SBD was incubated with maltopentaose or maltoheptaose (maltooligosaccharide of DP 7), a variety of oligosaccharides ranging from maltose to oligosaccharides with a degree of polymerization (DP) of at least 13 to 18, respectively, were formed (Fig. 4, lanes 3-4). This result indicated that AgtA-SBD hydrolysed starch to some extent, but mainly acted as a glucanotransferase, transferring parts of a donor oligosaccharide, which might be starch, to an acceptor substrate, e.g. maltose, thereby producing a variety of oligosaccharides of different lengths.
Production and purification of AgtA and AgtB
To rule out any effect of the incorporated SBD on the enzymatic activity of AgtA, both AgtA and AgtB were overexpressed in A. niger in their native form for further biochemical analysis. All selected transformants overproduced a protein with an estimated molecular mass of 85 kDa not observed in the untransformed strain. In a similar procedure of cloning and transformation, we also attempted to produce the AgtC protein. Although insertion of the overexpression construct in selected transformants was confirmed by Southern blot analysis, none of the transformants overproduced the AgtC protein. The characterization of AgtC was therefore not included in this study. Proteins expressed in the culture medium of A. niger MGG029-ΔaamA-AgtA and of A. niger MGG029-ΔaamA-AgtB were concentrated and subsequently submitted to anion exchange chromatography, resulting in the purification of AgtA and AgtB. Both proteins had an apparent molecular mass of approximately 70 kDa, but after removal of N-glycosylation the apparent protein masses decreased to approximately 55 kDa, close to their theoretical masses (AgtA 58.8 kDa and AgtB 57.7 kDa, after removal of C-terminal end for GPI-anchoring) (Fig. 3).
Glucanotransferase activity of AgtA and AgtB on maltooligosaccharides
Both AgtA and AgtB were incubated with maltooligosaccharides ranging in size from maltose to maltohexaose (a DP 6 maltooligosaccharide) and the products were analysed by TLC. With maltopentaose or maltohexaose as substrates, both enzymes produced a range of oligosaccharides of DP 15 or larger, similar to what was observed previously for AgtA-SBD (Fig. 5, lanes 3, 4, 7 and 8). Incubation with maltopentaose at lower concentrations (2 or 10 mM) also resulted in the formation of products of DP 6 and larger (results not shown). Activity of both enzymes on the smaller maltooligosaccharides was limited (Fig. 5, lane 1, 2 and 5, 6). Neither of the enzymes produced glucose in detectable amounts. When incubated with dissolved potato starch both enzymes produced small amounts of reducing ends (hydrolysis on 2% starch 0.55 ± 0.19 μmole mg-1 min-1 for AgtA and 0.26 ± 0.16 μmole mg-1 min -1 for AgtB; the

Alpha-glucan acting enzymes in Aspergillus niger
76
hydrolysis rate was approx. 4 times lower when measured on 0.2% starch). TLC analysis of the products produced from maltoheptaose at different pH values indicated that AgtA was active between pH 4.5 and pH 6, and AgtB showed activity between pH 4 and pH 5.5 (data not shown). Glucose polymers with other types of glycosidic linkages, like dextran (a glucan polymer with α-(1,6)-glycosidic bonds), nigeran, A. nidulans α-(1,3) glucan, an L. reuteri polysaccharide containing α-(1,3)- and α-(1,6)-glycosidic bonds (Kralj et al., 2004a), and cellulose (β-(1,4)-glycosidic bonds) were tested alone or in combination with maltose as an acceptor substrate. None of these polysaccharides acted as substrates for AgtA or AgtB (data not shown).
Fig. 5 TLC analysis of the reaction products of AgtA (left side) and AgtB (right side) incubated with different substrates. Purified enzyme (0.4 μg) was incubated with 20 mM substrate for 1 h at 37 °C. The samples were applied at the spots where the polymeric material is visible. The marker lanes (indicated by M) contain a mix of maltooligosaccharides ranging from glucose (G1) to maltoheptaose (G7). The figure shows reaction products of AgtA/AgtB incubated with: maltotriose (lane 1/5), maltotetraose (lane 2/6), maltopentaose (lane 3/7), maltohexaose (lane 4/8).
Identification of the reaction products of AgtA and AgtB
To study the nature of the glycosidic linkages formed as well as the maximum length of the oligosaccharide products, reaction products of AgtA and AgtB were analysed qualitatively by HPLC. AgtA and AgtB produced a similar mixture of oligosaccharides with various lengths from maltoheptaose as substrate (Fig. 6B and C). The maximum detectable product length was approximately DP 28 after one hour of incubation for
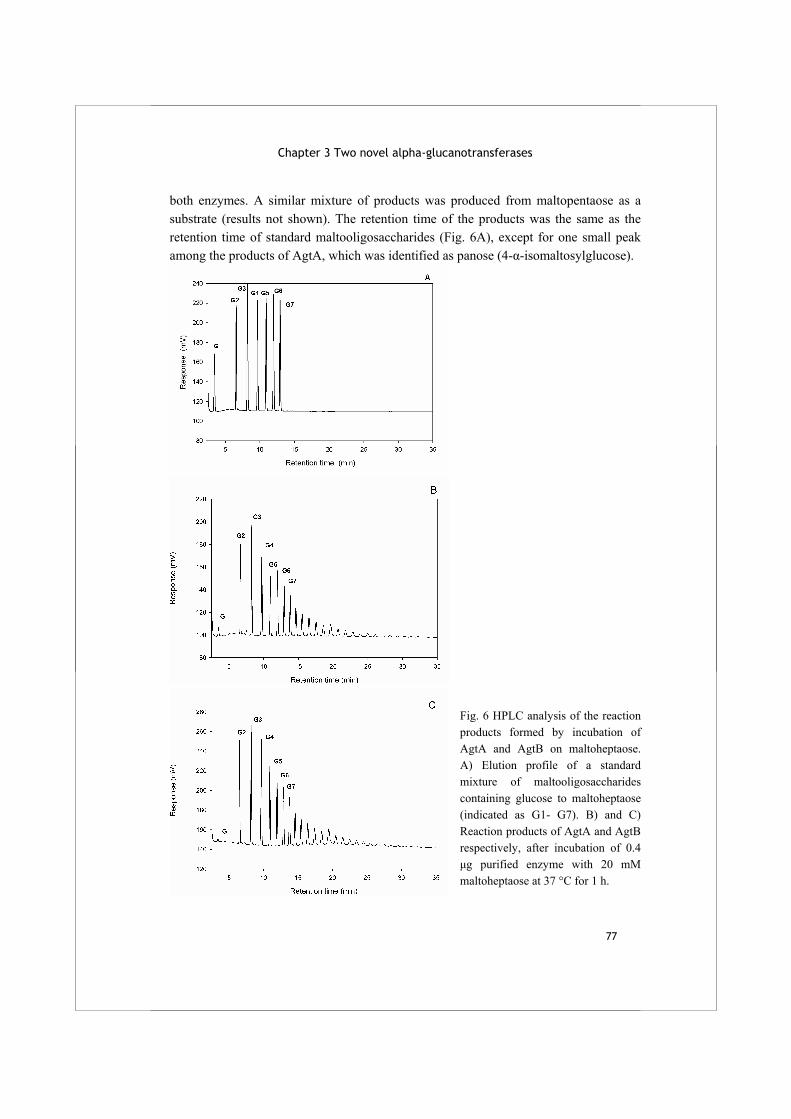
Chapter 3 Two novel alpha-glucanotransferases
77
both enzymes. A similar mixture of products was produced from maltopentaose as a substrate (results not shown). The retention time of the products was the same as the retention time of standard maltooligosaccharides (Fig. 6A), except for one small peak among the products of AgtA, which was identified as panose (4-α-isomaltosylglucose).
Fig. 6 HPLC analysis of the reaction products formed by incubation of AgtA and AgtB on maltoheptaose. A) Elution profile of a standard mixture of maltooligosaccharides containing glucose to maltoheptaose (indicated as G1- G7). B) and C) Reaction products of AgtA and AgtB respectively, after incubation of 0.4 μg purified enzyme with 20 mM maltoheptaose at 37 °C for 1 h.

Alpha-glucan acting enzymes in Aspergillus niger
78
AgtA and AgtB can use α-(1,3)-glucooligosaccharides as acceptor
The presence of the putative GPI-anchoring sequence in both AgtA and AgtB indicate that both enzymes are possibly present in the cell wall or cell membrane. The α-glucans present in the cell wall of A. niger are α-(1,3)-glucan (Horisberger et al., 1972) and nigeran (Bobbitt et al., 1977). In the previous paragraph we showed that neither nigeran nor α-(1,3)-glucan were used as (donor) substrate by AgtA and AgtB. To investigate whether small substrates with α-(1,3)-glycosidic bonds could be used as an acceptor substrate, both enzymes were incubated with soluble starch as donor substrate, combined with glucose, maltose, nigerose or nigerotriose as acceptor substrates. Analysis of the reaction products by TLC and HPLC revealed that glucose was not used as acceptor substrate by either of the enzymes; maltose was an efficient acceptor as shown previously for AgtA-SBD (results not shown). AgtA also formed a series of oligosaccharides using either nigerose or nigerotriose as acceptor substrates, although the amount of products formed was lower than when maltose acted as acceptor substrate (Fig. 7A). AgtB did not use nigerose or nigerotriose efficiently as acceptor substrate (Fig. 7B). No activity of AgtA or AgtB was observed on as sole substrates (result not shown). These results indicated that small α-(1,3)-linked oligosaccharides can be used as acceptor substrate by AgtA, and to a very limited extent by AgtB, but only in combination with an α-(1,4)-linked donor molecule.
ΔagtA and agtA/agtB overexpression strains are Calcofluor White hypersensitive
To examine the consequence of the loss of the agtA gene in A. niger and to analyse the physiological role of this enzyme, a deletion mutant of the agtA gene was constructed. Ten randomly chosen pΔagtA transformants were subjected to Southern blot analysis which revealed that in 8 out of the 10 transformants the agtA gene was properly deleted (data not shown). Phenotypic analysis of several ΔagtA strains revealed that their growth rate on solid media was slightly reduced, but no changes in the morphology of the hyphae or conidia were observed. We analysed the sensitivity of the ΔagtA strain to the cell wall disturbing compound Calcofluor White (CFW). Hypersensitivity towards CFW has been shown to be indicative for mutants with impaired cell wall strength (Ram and Klis, 2006;Damveld et al., 2005a). As shown in Fig. 8A, the ΔagtA strains showed an increased sensitivity towards CFW. The observed sensitivity is not as strong as for the deletion of other A. niger cell wall related proteins such as α-(1,3)-glucan synthaseA or the Cell wall proteinA (Damveld et al., 2005b;Damveld et al., 2005a). The overexpression of agtA and agtB also resulted in an increased sensitivity towards CFW (Fig. 8B). All deletion and overexpression strains were also tested for their ability
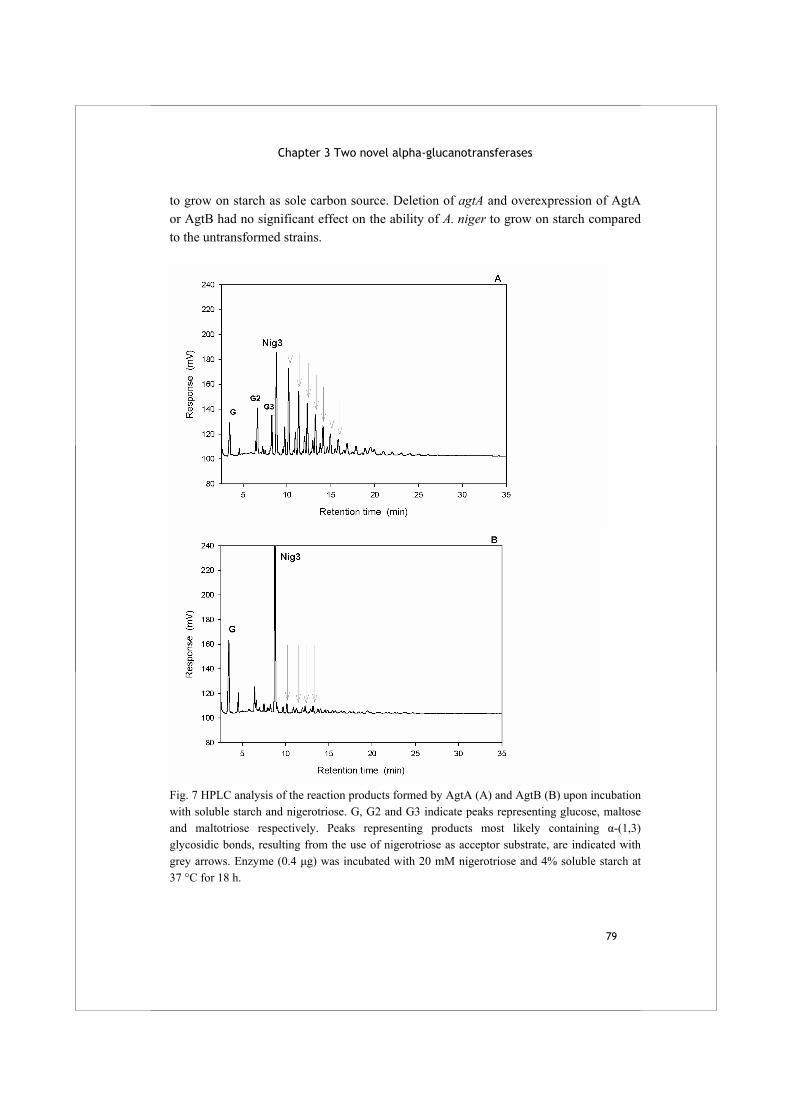
Chapter 3 Two novel alpha-glucanotransferases
79
to grow on starch as sole carbon source. Deletion of agtA and overexpression of AgtA or AgtB had no significant effect on the ability of A. niger to grow on starch compared to the untransformed strains.
Fig. 7 HPLC analysis of the reaction products formed by AgtA (A) and AgtB (B) upon incubation with soluble starch and nigerotriose. G, G2 and G3 indicate peaks representing glucose, maltose and maltotriose respectively. Peaks representing products most likely containing α-(1,3) glycosidic bonds, resulting from the use of nigerotriose as acceptor substrate, are indicated with grey arrows. Enzyme (0.4 μg) was incubated with 20 mM nigerotriose and 4% soluble starch at 37 °C for 18 h.

Alpha-glucan acting enzymes in Aspergillus niger
80
Fig. 8 The effect of agtA deletion (A) or agtA and agtB overexpression (B) on the susceptibility of the resulting strains towards Calcofluor White (CFW) induced cell wall stress. A) A. niger ΔagtA strains (MA71.1, MA71.3, MA71.4, MA71.7) and the control strain (MA70.15 transformed with pAO4-13), grown on complete medium containing 0, 100 or 400 μg ml-1 CFW for 96 hours. B) A. niger strains overexpressing AgtA or AgtB and the parental strain (MGG029-ΔaamA) grown on complete medium containing 0 or 200 μg ml-1 CFW for 72 h. The amount of spores applied per spot is indicated at the top of the figures.
Discussion All putative GPI-anchored family GH13 enzymes identified in the genome sequences of four aspergilli, as well as N. crassa, M. grisea and S. pombe, were originally annotated as α-amylases, because of their high similarity to known extracellular fungal α-amylases. However, most of the protein sequences missed the commonly conserved His143 in region I (Jespersen et al., 1991). Mutation of this residue, which is located in the active site (Uitdehaag et al., 1999), resulted in a strongly reduced activity in several α-amylases (Nakamura et al., 1993;Chang et al., 2003), or in altered reaction specificities in other family GH13 enzymes (see e.g. Leemhuis et al. (2004), Nakamura et al. (1993)). Despite the missing His143 residue, the A. niger Agt enzymes and their

Chapter 3 Two novel alpha-glucanotransferases
81
homologues are clearly members of family GH13, based on their high similarity with known proteins in this family. The second generally conserved His residue, His317 in conserved region IV, is replaced by Gln in AgtB and AgtC. Although His317 is overall highly conserved in the α-amylase family, this residue appears to be less important for the determination of the catalytic activity because some α-amylases are known which also do not posses a His in this position (Hoshiko et al., 1987;Kang et al., 2004). In conclusion, the Agt enzymes in A. niger and homologues in other fungi are highly similar to the well described fungal α-amylases, but aberrant conserved regions combined with the presence of a GPI-anchoring signal make them clearly distinguishable. We produced two of the A. niger Agt enzymes and determined their biochemical activities. As the enzymatic activity of the Agt proteins was not known at the start of this research, it was decided to start with the production of the enzyme encoded by gene An09g03100 (AgtA) fused with a clearly recognizable tag, the SBD. This allowed isolation and purification of the protein from media via binding to starch granules, as well as recognition of the purified protein using antibodies. Subsequently, both AgtA and AgtB (encoded by gene An12g02460) were produced in their native form for further biochemical characterization. A host A. niger strain with very low α-amylase activity was chosen to strongly reduce interference by native activities when searching for the biochemical activity of the investigated enzymes. AgtA-SBD as well as native AgtA and AgtB had a very low hydrolysing activity on starch, but clearly showed α-glucanotransferase activity on maltooligosaccharides alone, and on maltooligosaccharides plus starch. We therefore propose to name the A. niger GPI-anchored family GH13 enzymes Agt (α-glucanotransferase) A and B. As the predicted protein encoded by gene An15g07800 has the same sequence characteristics, we propose to name it AgtC. AgtA and AgtB were overproduced from their entire predicted coding sequences, which included the predicted GPI-anchoring signal. The presence of both AgtA and AgtB in the medium suggested that these proteins were not fully retained at the plasma membrane or the cell wall, but that at least part of the proteins was released into the medium. This might be an indication that the proteins are not GPI-anchored, although their C-terminal signal sequence as well as the investigation of the homologous Aah3p from S. pombe (Morita et al., 2006) suggest otherwise. Another explanation is that the enzymes had been released by endogenous phospholipase C activity, as was previously shown to occur with GPI-anchored proteins in A. niger and S. cerevisiae (Brul et al., 1997).

Alpha-glucan acting enzymes in Aspergillus niger
82
AgtA and AgtB produced a similar range of products consisting of maltooligosaccharides, indicating that both enzymes formed α-(1,4)-glycosidic bonds and can therefore be classified as 4-α-glucanotransferases (EC 2.4.1.25). Also a small amount of panose was produced by AgtA, indicating an ability to synthesize α-(1,6) linkages. Alternatively, panose may have been produced by a minor contamination of α-glucosidase, which is known to produce α-(1,6) linkages (Kato et al., 2002b). The enzymatic activity of AgtA and AgtB is unique among the α-glucanotransferases from bacteria as well as eukarya. The α-glucanotransferases that have been described until now usually release one glucose molecule for every transfer event (Takaha et al., 1993), but AgtA and AgtB did not produce glucose in significant amounts. Additionally, bacterial amylomaltases use very small donor and acceptor molecules (maltotriose and glucose, respectively) (Kaper et al., 2004), while AgtA and AgtB prefer longer donor molecules with a minimum length of 5 glucose residues, and maltose as smallest possible acceptor substrate. The use of the α-(1,3) linked oligosaccharides nigerose and nigerotriose as acceptor substrate by family GH13 α-glucanotransferases has not been reported before. We conclude that the A. niger α-glucanotransferases represent a new subgroup of family GH13 in view of their atypical donor and acceptor profiles and their C-terminal GPI-anchoring sequences. Based on their common putative cell wall associated location and amino acid sequences, it is expected that the closely related GPI-anchored family GH13 proteins in other fungi will show similar glucanotransferase activities, although their precise substrate and product profiles remain to be determined. Most extracellular members of family GH13 are involved in the degradation of starch to supply energy and carbon to the cells. There are strong indications that this is not the case for AgtA and AgtB. This study describes that neither a knockout of agtA nor overexpression of AgtA or AgtB had an effect on the ability of A. niger strains to grow on starch, even if the parental strain was severely hampered in this trait. In another study, we have also shown that expression of the agtA and agtB genes is not regulated by AmyR, the general regulator for starch processing enzymes in aspergilli (Yuan et al., in preparation) (Petersen et al., 1999), and similar results were found for the homologous genes in A. nidulans (Nakamura et al., 2006). Taken together, these data indicate that the Agt proteins most likely are not involved in starch catabolism. An alternative function could be the production or modification of α-glucans in the fungal cell wall, which was suggested for one of the GPI-anchored Agt homologues in the fission yeast S. pombe, Aah3p, in a functional study (Morita et al., 2006). Deletion of aah3 resulted in a morphological defect and hypersensitivity towards cell wall degrading enzymes. The knockout could not be rescued by transformation with the aah3 gene in which the catalytic residues had been mutated, showing the importance of the enzymatic activity rather than the structural properties of the protein. Our finding of a

Chapter 3 Two novel alpha-glucanotransferases
83
clear enzymatic activity for AgtA and AgtB confirms the importance of the catalytic residues for their physiological role. The proposed role for AgtA and its homologues in cell wall α-glucan production or maintenance is strengthened by the analysis of agtA knockout strains, which showed increased sensitivity towards CFW. The overexpression of AgtA and AgtB in A. niger caused a similar effect, which might be an indication that the unnaturally high amounts of these enzymes have a negative effect on cell wall strength. The function of the Agt proteins in the fungal cell wall might be analogous to the function of GPI-anchored β-glucanosyltransferases. These enzymes, identified in several yeasts and fungi including aspergilli, play a role in the cross linking of cell wall β-glucan (Popolo et al., 1993;Mouyna et al., 2000). In A. fumigatus, the β-glucan component of the cell wall is used as a target for antifungal drugs (Beauvais and Latgé, 2001). A knockout of one of its GPI-anchored β-glucanosyltransferases, Gel2p, resulted in an altered cell wall composition, increased sensitivity for CFW, and reduced virulence (Mouyna et al., 2005). We tested the ability of AgtA and AgtB to process the two α-glucan cell wall components, nigeran and α-(1,3) glucan, but no activity was detected. Because of their poor solubility at low pH, these substrates were offered at a relatively low concentration and partly in crystalline form, which might prevent the enzymes from acting on these cell wall components in vitro. It was shown, however, that AgtA performed a transglycosylation reaction involving an α-(1,4) linked donor substrate and an α-(1,3) linked acceptor substrate. A similar reaction was described to occur in S. pombe cell walls, where two linear polysaccharide chains of α-(1,3)-glucan with several α-(1,4)-linkages at the reducing end were interconnected by a transglycosylation reaction (Grün et al., 2005). Although this process was suggested to be performed by the transferase domain of α-(1,3) glucan synthase Ags1p, a similar cross linking reaction could be performed by AgtA. This would also explain the clustering of agt and ags (α-glucan synthase) genes conserved in many Ascomycetes. Clustering of genes involved in the same metabolic pathway is well described in fungi (Keller and Hohn, 1997). To conclude, we have studied two novel putatively GPI-anchored family GH13 enzymes of A. niger, with homologues in many other fungi. The enzymes, AgtA and AgtB, both showed a unique type of α-(1,4) glucanotransferase activity, and our functional characterization indicated that their involvement in the facilitation of growth on starch is unlikely. The characterization of a knockout of agtA suggested that this enzyme could be involved in cell wall α-glucan synthesis, which is in line with the results on a knockout of a homologous protein from S. pombe (Morita et al., 2006).

Alpha-glucan acting enzymes in Aspergillus niger
84
More study is needed to confirm the proposed physiological role for these α-glucanotransferases and to identify their exact in vivo reaction.
Acknowledgements
We are very grateful to Dr. Jacques Benen and Harrie Kools of Wageningen University for providing the pki-pgaII plasmid and the help with the HMMR searches. We also acknowledge the contribution of Peter Sanders (TNO Quality of Life, The Netherlands) for the HPLC analysis and Mark Arentshorst (Leiden University) for technical assistance with the knockout.

85
Chapter 4 Biochemical characterization of two GPI-anchored α-glucanotransferase enzymes from Aspergillus niger reveals a novel reaction specificity in subfamily GH13_1 of fungal α-amylases R.M. van der Kaaij, M.J.E.C. van der Maarel and L. Dijkhuizen
Submitted
Abstract Aspergillus niger AgtA and AgtB are the first family GH13 α-(1,4) glucanotransferase enzymes described in fungi. Their main activity is disproportionation of maltopentaose or longer maltooligosaccharides substrates, including starch. Biochemical and phylogenetic characterization of AgtA and AgtB revealed clear differences with prokaryotic disproportionating enzymes. AgtA and AgtB are GPI-anchored to the cell wall, where they most likely play a role in cell wall α-glucan maintenance. They represent a novel reaction specificity in subfamily GH13_1, also comprising fungal extracellular α-amylases involved in starch degradation, and can be distinguished on the basis of specific sequence features.

Alpha-glucan acting enzymes in Aspergillus niger
86
Introduction Family 13 (GH13) is the largest of all sequence based glycoside hydrolase (GH) families, comprising at least 26 different reaction specificities (http://www.cazy.org) (Coutinho and Henrissat, 1999;Stam et al., 2006). A well-known member of this family is α-amylase, an enzyme hydrolyzing starch. Members of family GH13 have a common 3D structure and catalytic machinery. The active site of these enzymes is composed of four conserved amino acid regions containing the three catalytic residues and four additional highly conserved residues involved in substrate binding and stabilization of the transition state of the reaction (MacGregor et al., 2001). Based on amino acid sequence similarity, family GH13 was recently divided into 35 subfamilies, which correlated relatively well with the biochemical activities reported for enzymes in each subfamily (Stam et al., 2006). Analysis of the α-amylase type enzymes (subfamily GH13_1) encoded in the genomes of several Aspergillus species revealed that they could be categorized in two distinct groups. Apart from the extracellular α-amylases necessary for the hydrolysis of starch, all species harboured several cell wall, or membrane bound, GPI-anchored enzymes with clear similarity to α-amylases, but with aberrant conserved regions (Yuan et al., in preparation). Two of these enzymes, AgtA and AgtB from A. niger, were identified as α-glucanotransferases acting on starch and maltooligosaccharides (EC 2.4.1.25) (van der Kaaij et al., 2007). The phenotype of an agtA knockout strain, and the expression patterns of agtA and agtB in wild type A. niger, suggested that these enzymes play a role in synthesis and/or maintenance of cell wall α-glucans (van der Kaaij et al., 2007). Fungal cell wall α-glucan is generally composed of two types of polymers: α-(1,3) glucans with a small percentage of α-(1,4) glycosidic bonds, and nigeran with alternating α-(1,3) and α-(1,4) glycosidic bonds. The proposed role for AgtA/B and homologous fungal enzymes is further supported by the phenotype of a knockout strain for the homologous GPI-anchored protein Aah3p in S. pombe, which showed an aberrant cell shape and hypersensitivity to cell wall degrading enzymes (Morita et al., 2006). Stam et al. (2006) expressed their concern with respect to the accuracy of sequence similarity-based annotations of Glycoside Hydrolase (GH) family enzymes in genome sequencing projects, especially in view of the “decreasing number of novel enzymatic characterization reports… reflecting the fact that the quest for increased impact factors renders journals reluctant to publish such characterizations”. This study provides a detailed characterization of the two novel subfamily GH13_1 AgtA/B enzymes from

Chapter 4 Biochemical characterization of AgtA and AgtB
87
A. niger. Biochemically, but not phylogenetically, they resemble prokaryotic disproportionating enzymes (e.g. amylomaltases, family GH77). We show that phylogenetic analysis, as performed by Stam et al., is not sufficient to distinguish between fungal α-amylases and Agt enzymes, but predicted GPI-anchoring and variations in the four conserved family GH13 regions clearly distinguish these two types of fungal enzymes.
Materials and methods
Enzyme production and purification
Production and purification of full length AgtA and AgtB were performed as described before (van der Kaaij et al., 2007). AgtA and AgtB were overexpressed in A. niger strain MGG029-ΔaamA (Weenink et al., 2006), which is a knockout for the genes encoding glucoamylase (glaA) and acid amylase (aamA) resulting in very low extracellular α-amylase activity. Both enzymes were purified from culture medium via anion exchange chromatography and stored in 50 mM NaAc pH 5.5 containing 1 mM CaCl2 at -20 °C.
General assay conditions
All oligosaccharides were obtained from Sigma, except nigerotriose which was purchased from Dextra laboratories and nigerose (O-α-D-Glc-(1,3)-α-D-Glc) which was a kind gift from Nihon Shokuhin Kako Co. Ltd. (Shizuoka, Japan). The α-(1,3)-glucan isolated from A. nidulans was a gift from Dr. B. J. Zonneveld (Leiden University, Leiden, The Netherlands). Nigeran and α-(1,3)-glucan were dissolved in 1 M NaOH and the pH of a 1% solution was adjusted to pH 5.5 with HAc. Quantitative measurements of glucanotransferase activity were performed with p-nitrophenyl-α-D-maltoheptaoside-(4,6)-O-benzylidine (PNP-G7) (Megazyme, County Wicklow, Ireland), a double blocked maltoheptaoside, as donor substrate (as adapted from Nakamura et al. (1994) and Van der Veen et al. (2000)). In the standard assay 100-500 ng AgtA or 400-1000 ng AgtB were incubated in a total reaction volume of 180 μl, containing 10 mM PNP-G7 as donor substrate, 40 mM acceptor substrate (maltose in standard reactions) and 1 mM CaCl2 in 50 mM NaAc buffer. AgtA reactions were buffered at pH 5.5 and AgtB reactions at pH 4.8, unless indicated otherwise. To measure hydrolysis, reactions were performed without acceptor substrate. Samples of 30 μl were taken after 0, 5, 10, 15 and 20 min and were immediately transferred to preheated reaction tubes and incubated for 15 min (AgtA) or 5 min (AgtB) at 78 °C to deactivate the enzymes. After cooling on ice, 120 μl 50 mM NaAc buffer pH 4.5 containing 1 U A. niger α-glucosidase (Megazyme) was added to each sample and samples were incubated for 30 min at 37 °C to liberate p-nitrophenol (PNP) from the

Alpha-glucan acting enzymes in Aspergillus niger
88
cleaved PNP-G7 product. Subsequently, 0.75 ml 1 M Na2CO3 was added to each sample and Abs402 was measured. In each experiment auto hydrolysis of PNP-G7 was always negligible. Reactions were performed at least in duplicate. Linear regression was applied on the data of each separate reaction and the slope of the trend line (with R2 >0.99) was used as a measure for the reaction rate. 1 U of enzyme activity was defined as the amount of enzyme liberating 1 μMol of PNP per minute under the described reaction conditions. A calibration curve was made which indicated that the molar extinction coefficient of PNP under these conditions was 16.4 mM-1 cm-1 at 402 nm. The experimental data were analysed using SigmaPlot version 9.0 (Systat Software).
Temperature optimum
Temperature optima were determined with 100 ng AgtA or 400 ng AgtB. Reactions with AgtA at 60 °C and AgtB at 48.5 °C and 53 °C were incubated 10 min (instead of 20 min), to prevent loss of activity due to heat inactivation.
Kinetic analysis
Kinetic analysis was performed at 45 °C for AgtA and 40 °C for AgtB, with 100 ng or 500 ng purified enzyme, respectively. For both enzymes, 11 different PNP-G7 concentrations were tested in the range of 0.5-18 mM (AgtA) or 0.5-22 mM (AgtB). The calculated molecular masses of the proteins were 58840 Da for AgtA and 57790 Da for AgtB, after removal of the C-terminal end for GPI-anchoring. The Km and Vmax values were determined via Lineweaver-Burk plots with R2 >99.
Acceptor substrates
AgtA (100 ng) or AgtB (500 ng) were incubated with different acceptor substrates (40 mM) at 37 °C and pH 5.5. Reaction rates were compared to the values observed with maltose, and corrected for autohydrolysis (release of PNP in the absence of acceptor substrate).
Subsite modelling and acarbose inhibition
Initial product formation was analysed using 800 ng AgtA or 1.5 μg AgtB in a total reaction volume of 66 μl containing 20 mM maltopentaose, maltohexaose or maltoheptaose. Samples of 12 μl were taken after 0, 5, 15, 30, 60 min of incubation and analysed by HPLC (Dionex) (van der Kaaij et al., 2007). Inhibition by acarbose (0, 1 or 10 mM) was checked using 100 ng AgtA or 300 ng AgtB with 20 mM maltoheptaose in a total volume of 15 µl. Samples were taken after 60 min and analysed by thin layer chromatography as described before (van der Kaaij et al., 2007).
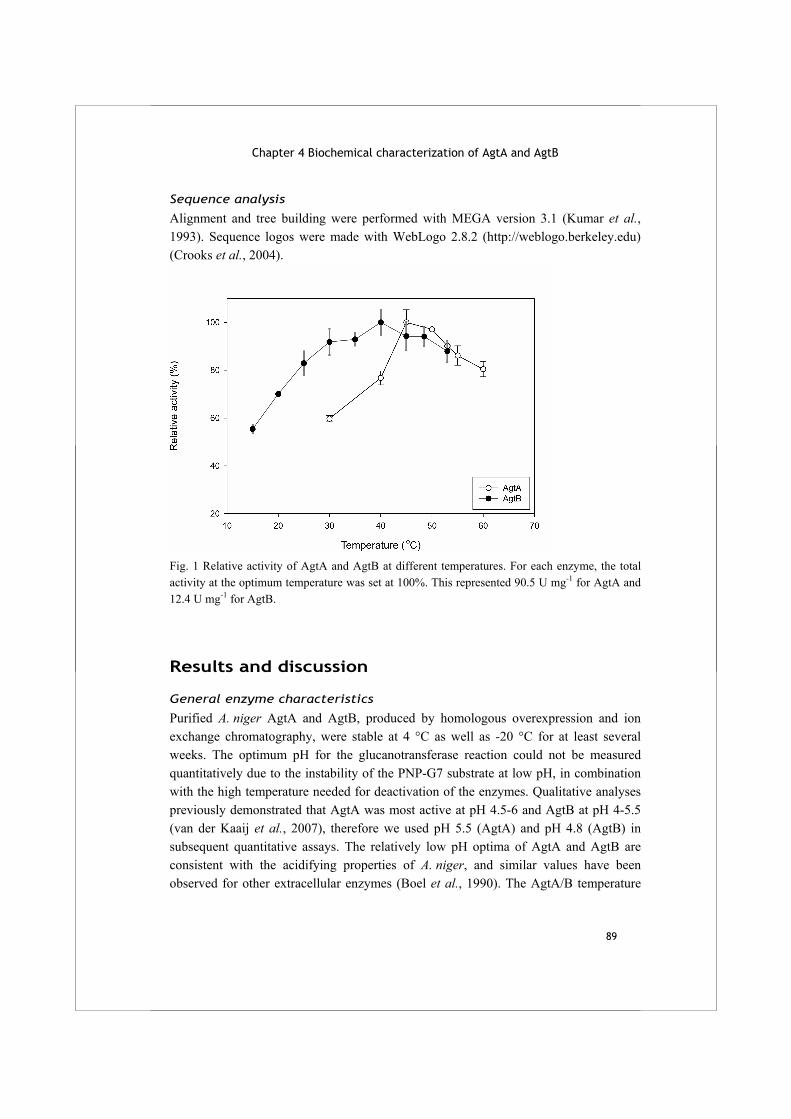
Chapter 4 Biochemical characterization of AgtA and AgtB
89
Sequence analysis
Alignment and tree building were performed with MEGA version 3.1 (Kumar et al., 1993). Sequence logos were made with WebLogo 2.8.2 (http://weblogo.berkeley.edu) (Crooks et al., 2004).
Fig. 1 Relative activity of AgtA and AgtB at different temperatures. For each enzyme, the total activity at the optimum temperature was set at 100%. This represented 90.5 U mg-1 for AgtA and 12.4 U mg-1 for AgtB.
Results and discussion
General enzyme characteristics
Purified A. niger AgtA and AgtB, produced by homologous overexpression and ion exchange chromatography, were stable at 4 °C as well as -20 °C for at least several weeks. The optimum pH for the glucanotransferase reaction could not be measured quantitatively due to the instability of the PNP-G7 substrate at low pH, in combination with the high temperature needed for deactivation of the enzymes. Qualitative analyses previously demonstrated that AgtA was most active at pH 4.5-6 and AgtB at pH 4-5.5 (van der Kaaij et al., 2007), therefore we used pH 5.5 (AgtA) and pH 4.8 (AgtB) in subsequent quantitative assays. The relatively low pH optima of AgtA and AgtB are consistent with the acidifying properties of A. niger, and similar values have been observed for other extracellular enzymes (Boel et al., 1990). The AgtA/B temperature

Alpha-glucan acting enzymes in Aspergillus niger
90
optima were determined using PNP-G7 as donor and maltose as acceptor substrate (Fig. 1). AgtA showed a relatively narrow temperature optimum between 45 – 50 °C, while AgtB showed a remarkably broad temperature optimum. Although the relative activity of both enzymes was similar between 47 °C and 53 °C, their stability was clearly different. AgtB lost activity rapidly at 50 °C and higher during the 20 min reaction period, while AgtA activity was stable at temperatures up to 55 °C (results not shown).
Fig. 2 Relative activity of AgtA and AgtB with PNP-G7 as donor substrate and different acceptor substrates. For each enzyme, the activity with maltose as acceptor substrate was set at 100%, representing 75.5 U mg-1 for AgtA and 10.2 U mg-1 for AgtB.
Kinetic analysis
Michaelis-Menten type kinetics was observed for the α-glucanotransferase reactions of AgtA and AgtB with PNP-G7 and maltose. The values obtained for various kinetic parameters differed strongly between the two enzymes (Table 1). Notably, the Vmax for AgtA was five times higher than for AgtB, and specific activities differed a factor 10. This relatively low specific activity of AgtB compared to AgtA was also observed in reactions with maltooligosaccharides analysed by HPLC (results not shown). Hydrolysis of PNP-G7 was low but in a similar range for both enzymes (between 2 and 3.5 U mg-1), and comparable to the starch hydrolysis rates (van der Kaaij et al., 2007). The Michaelis-Menten kinetics of AgtA and AgtB with regard to donor substrate is

Chapter 4 Biochemical characterization of AgtA and AgtB
91
generally also observed for prokaryotic disproportionating enzymes. The Km value of AgtA/B for PNP-G7 as donor substrate is, however, 10 times higher than for bacterial cyclodextrin glucanotransferases (CGTases) (Leemhuis et al., 2003;Leemhuis et al., 2002), and actually more in the range of the Km for PNP-G7 in the hydrolysis reaction of related fungal α-amylases, e.g. A. niger acid amylase (Boel et al., 1990). The Kcat values of AgtA and AgtB are rather low compared to those of CGTases (Kcat of disproportionation reaction around 1000 s-1) (Leemhuis et al., 2003;Leemhuis et al., 2002), but similar to those reported for amylomaltases, members of family GH77 (e.g. of Pyrobaculum aerophilum IM2, 115 s-1) (Kaper et al., 2005). Table 1 Kinetic parameters of AgtA and AgtB determined for the disproportionation reaction with PNP-G7 as donor substrate and maltose as acceptor substrate.
AgtA AgtB
Km (donor) 2.4 mM 4.9 mM
Vmax 95 U mg-1 18.4 U mg-1
Kcat 334 s-1 64 s-1
Kcat/Km 139 s-1 mM-1 13 s-1 mM-1
Acceptor substrates and subsite modelling
The relative efficiency of AgtA and AgtB towards different acceptor substrates, using PNP-G7 as donor substrate, was determined (Fig. 2). For both enzymes, the best acceptor substrate was maltotriose. Interestingly, nigerose and nigerotriose also were used as acceptor substrates, although not as efficiently as maltooligosaccharides. Maltopentaose and larger acceptor substrates were not tested because these may also act as donor substrates, thereby interfering with PNP-G7 as a quantifiable donor substrate. The polysaccharides nigeran and α-(1,3)-glucan were not used as acceptor substrate (data not shown). Activities with glucose and isomaltose were very low, as opposed to amylomaltase enzymes that efficiently use glucose as acceptor substrate (Kaper et al., 2004). In analogy to other family GH13 and GH77 enzymes, AgtA and AgtB most likely act by cleaving a donor substrate, covalently binding part of it to the amino acid acting as nucleophile, and the subsequent coupling of this donor substrate to the non-reducing end of an incoming acceptor molecule (see e.g. Uitdehaag et al. (1999)). We incubated AgtA and AgtB with different maltooligosaccharide substrates (G5-G7) and identified the initial products formed (Figs. 3 and 4).
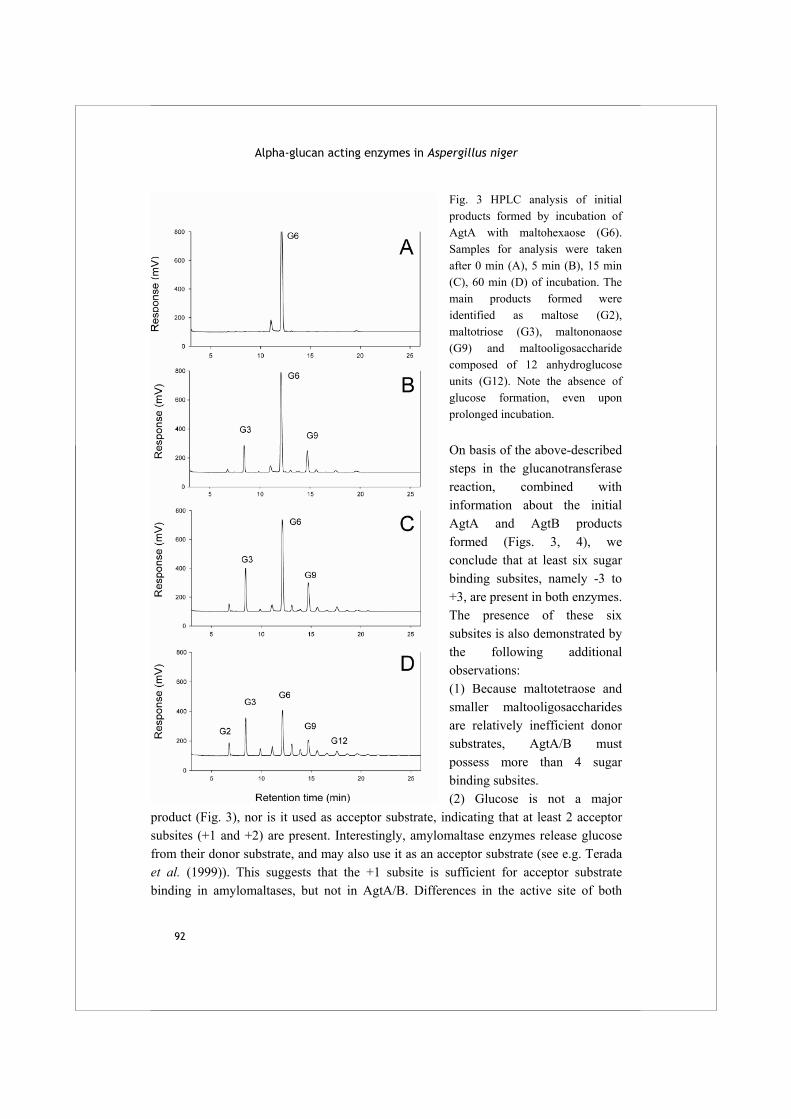
Alpha-glucan acting enzymes in Aspergillus niger
92
Fig. 3 HPLC analysis of initial products formed by incubation of AgtA with maltohexaose (G6). Samples for analysis were taken after 0 min (A), 5 min (B), 15 min (C), 60 min (D) of incubation. The main products formed were identified as maltose (G2), maltotriose (G3), maltononaose (G9) and maltooligosaccharide composed of 12 anhydroglucose units (G12). Note the absence of glucose formation, even upon prolonged incubation. On basis of the above-described steps in the glucanotransferase reaction, combined with information about the initial AgtA and AgtB products formed (Figs. 3, 4), we conclude that at least six sugar binding subsites, namely -3 to +3, are present in both enzymes. The presence of these six subsites is also demonstrated by the following additional observations: (1) Because maltotetraose and smaller maltooligosaccharides are relatively inefficient donor substrates, AgtA/B must possess more than 4 sugar binding subsites. (2) Glucose is not a major
product (Fig. 3), nor is it used as acceptor substrate, indicating that at least 2 acceptor subsites (+1 and +2) are present. Interestingly, amylomaltase enzymes release glucose from their donor substrate, and may also use it as an acceptor substrate (see e.g. Terada et al. (1999)). This suggests that the +1 subsite is sufficient for acceptor substrate binding in amylomaltases, but not in AgtA/B. Differences in the active site of both

Chapter 4 Biochemical characterization of AgtA and AgtB
93
enzyme types also become apparent from the comparison of their conserved regions, containing many of the amino acids composing the active site (Fig. 6). (3) The existence of a +3 acceptor binding site is indicated by the observation that maltotriose is a more efficient acceptor substrate than maltose (Fig. 2). (4) Initial formation of maltose and G8 from maltopentaose clearly indicates the presence of a -3 subsite. In general, the production of specific initial reaction products suggests that AgtA/B perform a non-progressive type of reaction, which is similar to amylomaltases (see e.g. Terada et al. (1999)).
Fig. 4 Initial products formed by AgtA and AgtB from different maltooligosaccharides. AgtA/B were incubated with maltopentaose, maltohexaose or maltoheptaose and initial reaction products, which were the same for both enzymes, were identified by HPLC analysis (see Fig. 3). Indicated is the predicted binding of donor substrates in the different subsites leading to the formation of the observed initial products. Subsites indicated as described by Davies et al. (1997). O, glucose unit; Ø, reducing end.
Phylogenetic analysis
AgtA and AgtB are the first studied representatives of a group of homologous enzymes identified in genome sequences of many filamentous fungi (Yuan et al., in preparation). Phylogenetically, this group is closely related to fungal α-amylases in subfamily GH13_1, but not to any of the prokaryotic disproportionating enzymes, despite the α-glucanotransferase activity of AgtA/B described here (Table 2; Fig. 5). The predicted Agt homologues can be distinguished from the α-amylases by their C-terminal GPI-anchoring signal and, in most cases, the absence of the equivalent of His147 (conserved region I) and sometimes also His324 (conserved region IV) (numbering according to AgtA) (Fig. 6), two highly conserved amino acids in family GH13 enzymes, located in the -1 subsite. In Agt homologues, His147 is mostly replaced by rather small, generally hydrophilic amino acids such as Asp, Asn or Ser. Interestingly, all family GH77 amylomaltase enzymes also lack the His147 residue (Fig. 6) (Kaper et al., 2007). One of the few known GH13 enzymes lacking both histidine residues is acarviosyltransferase from Actinoplanes sp. strain SE50, the only known enzyme that can efficiently use the α-amylase inhibitor acarbose as a substrate for α-glucanotransferase activity (Hemker et

Alpha-glucan acting enzymes in Aspergillus niger
94
al., 2001). AgtA and AgtB were only partly inhibited by 1 or 10 mM acarbose, which was not used as a substrate, suggesting that they are relatively insensitive for this inhibitor compared to other GH13 enzymes that generally have a Ki value for acarbose in the µmolar range (results not shown) (Kim et al., 1999). This acarbose insensitivity in AgtA/B may be based on the missing His residue(s), but this remains to be proven experimentally.
Fig. 5 Bootstrapped phylogenetic tree of A. niger AgtA/B/C and homologous proteins from A. fumigatus and S. pombe, predicted α-amylases from these fungi and several prokaryotic enzymes with disproportionation activity (listed in Table 2). The alignment and phylogenetic analysis were performed with MEGA version 3.1 using default settings. A bootstrapped tree was constructed with the neighbor-joining method using 500 replicates. The scale bar corresponds to a genetic distance of 0.2 substitution per position.

Chapter 4 Biochemical characterization of AgtA and AgtB
95
Fig. 6 Sequence logos of the four conserved sequence regions in (A) predicted GPI-anchored fungal α-glucanotransferase enzymes (based on 17 sequences), (B) predicted fungal extracellular α-amylases (17 sequences), and (C) family GH77 amylomaltase enzymes from bacteria and archaea (20 sequences). The first amino acid in each sequence logo is numbered according to AgtA (A), A. niger AamA (B) or E. coli MalQ (C).
Conclusions The A. niger AgtA and AgtB enzymes, for the first time described in biochemical detail in this study, are representatives of a large group of predicted fungal GH13_1 enzymes. Both Agt enzymes perform the same α-(1,4) glucanotransferase type of reaction, but also display clearly different features. The enzymes possess some unique properties compared to previously described enzymes with α-glucanotransferase activity, all from prokaryotic origin. The characteristic properties of the AgtA/B enzymes still remain to be compared to those of their homologues from other ascomycete fungi. The precise in vivo reaction and physiological function of these enzymes is most likely in cell wall synthesis and/or maintenance, but this remains to be proven experimentally. In future work, their biochemical characteristics as studied here need to be related to the dynamics of the structure of fungal cell wall, adding to our as yet poor understanding of the process of α-glucan formation and maintenance. Based on sequence similarity, AgtA/B and their homologues are classified into subfamily GH13_1, together with the fungal extracellular α-amylases (Stam et al., 2006). The activity of AgtA and AgtB has now been proven to be mainly of the glucanotransferase type, clearly different from that of the α-amylases, leading to a ‘polyspecific’ subfamily GH13_1. Other polyspecific subfamilies identified by Stam et al. generally displayed strongly related activities in which the variety was merely a matter of semantics or specificity of the applied assays. AgtA and AgtB as well as their homologues are not distinguishable from fungal α-amylases based on the phylogenetic analysis applied for the formation of subfamilies. They can, however, be distinguished
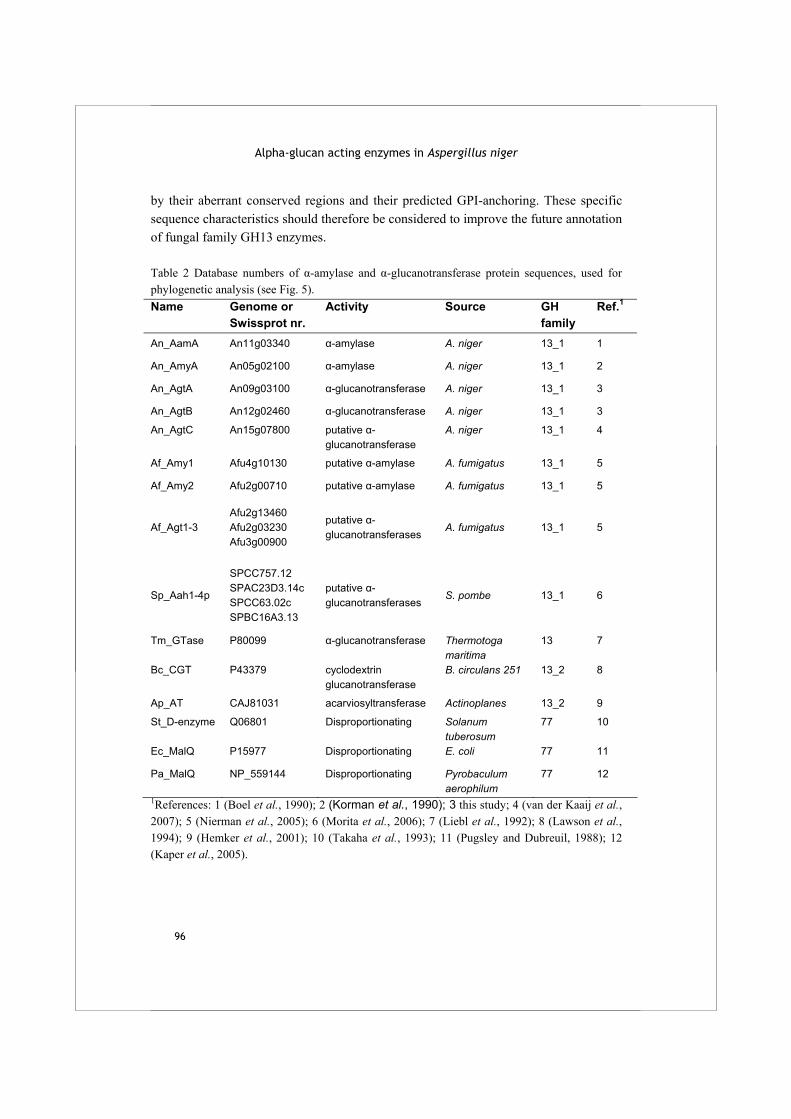
Alpha-glucan acting enzymes in Aspergillus niger
96
by their aberrant conserved regions and their predicted GPI-anchoring. These specific sequence characteristics should therefore be considered to improve the future annotation of fungal family GH13 enzymes. Table 2 Database numbers of α-amylase and α-glucanotransferase protein sequences, used for phylogenetic analysis (see Fig. 5). Name Genome or
Swissprot nr. Activity Source GH
family Ref.1
An_AamA An11g03340 α-amylase A. niger 13_1 1
An_AmyA An05g02100 α-amylase A. niger 13_1 2
An_AgtA An09g03100 α-glucanotransferase A. niger 13_1 3
An_AgtB An12g02460 α-glucanotransferase A. niger 13_1 3
An_AgtC An15g07800 putative α-glucanotransferase
A. niger 13_1 4
Af_Amy1 Afu4g10130 putative α-amylase A. fumigatus 13_1 5
Af_Amy2 Afu2g00710 putative α-amylase A. fumigatus 13_1 5
Af_Agt1-3 Afu2g13460 Afu2g03230 Afu3g00900
putative α-glucanotransferases
A. fumigatus 13_1 5
Sp_Aah1-4p
SPCC757.12 SPAC23D3.14c SPCC63.02c SPBC16A3.13
putative α-glucanotransferases
S. pombe 13_1 6
Tm_GTase P80099 α-glucanotransferase Thermotoga maritima
13 7
Bc_CGT P43379 cyclodextrin glucanotransferase
B. circulans 251 13_2 8
Ap_AT CAJ81031 acarviosyltransferase Actinoplanes 13_2 9
St_D-enzyme Q06801 Disproportionating Solanum tuberosum
77 10
Ec_MalQ P15977 Disproportionating E. coli 77 11
Pa_MalQ NP_559144 Disproportionating Pyrobaculum aerophilum
77 12
1References: 1 (Boel et al., 1990); 2 (Korman et al., 1990); 3 this study; 4 (van der Kaaij et al., 2007); 5 (Nierman et al., 2005); 6 (Morita et al., 2006); 7 (Liebl et al., 1992); 8 (Lawson et al., 1994); 9 (Hemker et al., 2001); 10 (Takaha et al., 1993); 11 (Pugsley and Dubreuil, 1988); 12 (Kaper et al., 2005).

Chapter 4 Biochemical characterization of AgtA and AgtB
97
Acknowledgement
We thank P. Sanders (TNO Quality of Life, The Netherlands) for Dionex analyses.


99
Chapter 5
Phylogenetic and biochemical characterization of a novel cluster of intracellular fungal α-amylase enzymes R.M. van der Kaaij, Š. Janeček, M.J.E.C. van der Maarel and L. Dijkhuizen
Accepted for publication in Microbiology
Abstract Currently known fungal α-amylases are well-characterized extracellular enzymes that are classified into glycoside hydrolase subfamily GH13_1. This study describes the identification, phylogenetic and biochemical analysis of novel intracellular fungal α-amylases. The phylogenetic analysis shows that they cluster in the recently identified subfamily GH13_5 and display very low similarity to fungal α-amylases of subfamily GH13_1. Homologues of these intracellular enzymes are present in the genome sequences of all filamentous fungi studied, including Ascomycetes and Basidiomycetes. One of the enzymes belonging to this new group, Amy1p from Histoplasma capsulatum, recently has been functionally linked to the formation of cell wall α-glucan (Marion et al., 2006). To study the biochemical characteristics of this novel cluster of α-amylases, we overexpressed and purified a homologue from Aspergillus niger, AmyD, and studied its activity product profile with starch and related substrates. AmyD has a relatively low hydrolyzing activity on starch (2.2 U mg-1) producing mainly maltotriose. A possible function of these enzymes in relation to cell wall α-glucan synthesis is discussed.

Alpha-glucan acting enzymes in Aspergillus niger
100
Introduction Alpha-amylases are widely occurring enzymes which hydrolyze the α-(1,4)-glycosidic bonds in starch and glycogen, producing short maltooligosaccharides and maltose. Based on sequence similarity, most α-amylases (E.C. 3.2.1.1) are classified in glycoside hydrolase (GH) family 13, although some α-amylases originating from extremophilic organisms belong to family GH57 (Henrissat, 1991;Henrissat and Bairoch, 1996) (see also the CAZy website at http://www.cazy.org). Based on a phylogenetic analysis of 1691 different members of family GH13, the family was recently divided into 35 subfamilies, all acting on α-glycosidic bonds (Stam et al., 2006). Several of these subfamilies display α-amylase specificity, but many other enzyme reaction specificities are also represented. The tertiary structure of these enzymes is characterized by a (β/α)8
barrel containing four highly conserved amino acid regions that form the active site (MacGregor et al., 2001), but the overall sequence similarity can be as low as 10% and only a catalytic triad of amino acids is conserved invariantly (Machovic and Janecek, 2003). The shared (β/α)8 barrel structure and catalytic mechanism within family GH13 enzymes is believed to represent a common evolutionary origin (Kuriki and Imanaka, 1999;Janecek, 1997). The presence of the four conserved regions and a common secondary and tertiary structure allows construction of alignments and phylogenetic studies within the family. The phylogeny of α-amylases is generally in agreement with their origin, e.g. all fungal α-amylases are more related to each other than to the α-amylases originating from plants or animals. Alpha-amylases from bacteria, however, are scattered over several clusters, which group either with animal, plant or fungal α-amylases, or form a separate branch (Janecek, 1994). Several α-amylases from yeasts and fungi have been studied previously (see e.g. Steyn et al. (1995), Matsuura et al. (1984), Moreira et al. (2004), Boel et al. (1990)). In all cases, these enzymes are secreted into the extracellular environment, where they are involved in degradation of starch and glycogen into small oligosaccharides, which can be imported into the cells to serve as energy and carbon source. The expression levels of extracellular starch degrading enzymes are generally increased during growth in the presence of maltose or isomaltose. This system is particularly well studied in aspergilli, where expression of these enzymes is regulated by AmyR (Petersen et al., 1999;Tani et al., 2001;Nakamura et al., 1997). Some recent studies have shown that different fungal GH13 enzymes may be involved in formation and/or modification of α-glucans in fungal cell walls, rather than in starch degradation. The fungal cell wall is usually made up of chitin, β-glucan, α-glucan, galactomannan and attached cell wall proteins (Klis et al., 2002;Beauvais and Latgé,

Chapter 5 A novel cluster of intracellular alpha-amylases
101
2001). Generally, α-glucan in fungal cell walls is of the α-(1,3) type with a small percentage of α-(1,4) glycosidic bonds (Grün et al., 2005). Additionally, an α-glucan with alternating α-(1,3)/(1,4) glycosidic bonds (nigeran) has been identified in Aspergillus niger and some other Ascomycete species (Barker and Carrington, 1953;Woranovicz-Barreira et al., 1999). It is generally believed that these fungal α-glucans are produced by α-glucan synthases, although this has never been demonstrated directly. These transmembrane enzymes contain two catalytic domains. The C-terminal, intracellular domain exhibits similarity to members of glycosyltransferase family 5 (Coutinho et al., 2003) and probably uses UDP-glucose to produce a glucan chain. The N-terminal, extracellular domain has resemblance to family GH13 enzymes and is thought to be involved in the coupling of extruded glucan chains (Grün et al., 2005;Hochstenbach et al., 1998). Two novel types of family GH13 homologues were recently shown to play a role in fungal α-glucan formation. The first of these is Aah3p, a glycosylphosphatidylinositol (GPI) anchored protein identified in the fission yeast Schizosaccharomyces pombe (Morita et al., 2006). A knockout of the corresponding gene caused an aberrant cell shape and hypersensitivity towards cell wall degrading enzymes, indicating a role for the Aah3p protein in cell wall integrity. The biochemical characterization of two homologous GPI-anchored proteins from A. niger (AgtA and AgtB) revealed that they have 4-α-glucanotransferase activity on maltooligosaccharides and starch (van der Kaaij et al., 2007). A second type of GH13 enzyme with a role in cell wall formation is Amy1p from Histoplasma capsulatum, a close relative of the aspergilli (James et al., 2006). In this pathogenic, dimorphic fungus, α-(1,3) glucan is critical for virulence (Rappleye et al., 2004). A functional knockout strain of Amy1p, a putative intracellular α-amylase, lost completely the ability to form cell wall α-(1,3) glucan and attenuated its virulence (Marion et al., 2006). No biochemical characterization of Amy1p has been reported. The recent publication of the genome sequences of four aspergilli (Machida et al., 2005;Pel et al., 2007;Nierman et al., 2005;Galagan et al., 2005) allowed for the identification of all α-amylase homologues in these species. In a first analysis (Pel et al., 2007), it became apparent that these Aspergillus species encode several novel putative GH13 enzymes with relatively low similarity to the known, extracellular fungal α-amylases. Some of these proteins are, however, highly homologous to Amy1p from H. capsulatum. In the present study, we have identified homologues of Amy1p in aspergilli and other fungi, and performed sequence analysis as well as phylogenetic analyses on this group of novel fungal α-amylases. This information is combined with the heterologous expression, purification and characterization of one of these enzymes,

Alpha-glucan acting enzymes in Aspergillus niger
102
AmyD from A. niger, to gain a first time insight into the biochemical properties of a representative of this group of novel fungal enzymes.
Materials and methods
Sequence retrieval and analysis
The full genome sequence of A. niger strain CBS 513.88 has been deposited at the EMBL database with accession numbers AM270980-AM270998 (Pel et al., 2007). Previously identified genes with locus tags An01g13610 (AmyD) and An09g03110 (AmyE) from this database were used as query sequences in BLAST searches (Altschul et al., 1997) in all non-redundant GenBank CDS translations, RefSeq Proteins, PDB and SwissProt. The possible presence of a signal peptidase cleavage site or the possibility of non-classical secretion was analysed by web-based search tools SignalP and SecretomeP (http://www.cbs.dtu.dk/services/) (Bendtsen et al., 2004;Bendtsen et al., 2005).
Sequence alignment and evolutionary tree
A set of sequences encoding α-amylases was retrieved from GenBank (Benson et al., 2006) and SwissProt (Wu et al., 2006) (Table 1). The set was supplemented with the twelve fungal sequences identified as described above, and three additional sequences: Amy1p from H. capsulatum (Marion et al., 2006) and AgtA (An09g03100) and AgtB (An12g02460) from A. niger. The alignment strategy was based on the approach described by Da Lage et al. (2004). In short: (i) the best conserved regions - the strands β1, β2, β3, β4, β5, β7 and β8 of the catalytic (β/α)8 barrel and the region V in domain B (Janecek, 2002) - were identified in each sequence; (ii) the segments preceding and succeeding the regions around strands β1 and β8, respectively, were cut off; (iii) the shortened sequences (amino acids 34 to 435 in AmyD) were aligned by the program CLUSTALW (Thompson et al., 1994); (iv) the identified conserved sequence regions were aligned manually, if necessary; and (v) the remaining parts of the alignment (between the regions) were manually tuned where applicable. The evolutionary tree was calculated with the neighbour-joining method (Saitou and Nei, 1987) implemented in the CLUSTALX package (Jeanmougin et al., 1998) using the final alignment including the gaps; the number of bootstrap trials used was 1,000. The tree was displayed with the program TreeView (Page, 1996).
Strains, plasmids and cloning procedure
All basic molecular techniques were performed according to standard procedures (Sambrook et al., 1989). E. coli TOP10 (Invitrogen) was used for transformation and

Chapter 5 A novel cluster of intracellular alpha-amylases
103
amplification of recombinant DNA. E. coli BL21 STAR (DE3) (Invitrogen) was used for protein production. A synthetic gene with the coding sequence of An01g13610 was made by Geneart and cloned into pDONR221 using two ClaI restriction sites. The construct pDONR221- An01g13610 was subsequently used to make the final construct pDEST17- An01g13610 according to the manufacturer’s instructions. All intermediate steps in the production of the construct were checked by restriction analysis, and the final construct was checked by sequencing (GATC Biotech). As a positive control for cloning and purification, and a negative control for enzyme activity assays, we also cloned the gene encoding β-glucuronidase (gus) into pDEST-17 according to the manufacturer’s instructions.
Protein production and purification
E. coli BL21 STAR (DE3) transformed with pDEST17- An01g13610 or pDEST17-gus was grown in LB medium (Ausubel et al., 1987) containing 100 μg ampicillin ml-1 at 16 °C until an OD600 of 0.4 was reached. Expression was induced by the addition of 1 mM IPTG and cultures were grown until OD600 was 0.8-1. Cells were harvested by centrifugation (10 min, 5,000 g, 4 °C) and washed with 50 mM Tris-HCl buffer pH 8. Cell pellets were resuspended in binding buffer (50 mM Tris-HCl buffer pH 8 containing 500 mM NaCl, 10 mM imidazole, 5 mM β-mercaptoethanol). Cell free extract was produced by sonication of the resuspended cells (8 x 15 sec with 40 sec intervals, on ice) and subsequent centrifugation (20 min, 4 °C, 10,000 g). The cell lysate was applied to washed Ni-NTA column material (Qiagen) and incubated for 2 h at 4 °C. After washing the column material with binding buffer, His-tagged proteins were eluted with 50 mM Tris-HCl buffer pH 8 containing 250 mM NaCl, 1 mM β-mercaptoethanol and 100 mM imidazole. At each stage of the protein purification, the protein amount was measured using the Bradford method with reagent from Bio-Rad and purity was checked using SDS-PAGE analysis (Laemmli, 1970). The enzyme was concentrated over a YM10 filter (Millipore) and stored at -20 °C in Na-barbital buffer pH 6.5 containing 15% glycerol. Na-barbital buffer contained 28.5 mM NaAc, 28.5 mM Na-barbital and 116 mM NaCl and was brought to the desired pH by addition of HCl. After Ni-NTA purification, AmyD was used in biochemical assays for a maximum of 4 days.
Analysis of enzyme activity
Substrates were obtained from Sigma-Aldrich, except for the following: nigerotriose was purchased from Dextra laboratories, nigerose was a kind gift from Nihon Shokuhin Kako (Shizuoka, Japan), an α-(1,3)-glucan isolated from A. nidulans was a kind gift from Dr. B. J. Zonneveld (Leiden University, Leiden, The Netherlands) and a Lactobacillus reuteri exopolysaccharide (Kralj et al., 2004a) was supplied by Dr. S. Kralj (University of Groningen, The Netherlands). Starch, amylopectin and amylose

Alpha-glucan acting enzymes in Aspergillus niger
104
type III (all from Sigma-Aldrich) used in activity assays all originated from potato, glycogen originated from oyster. The standard reaction conditions used to measure hydrolysing activity were as follows: the enzyme was incubated with 0.2% (w/v) potato starch (or another substrate) in Na-barbital buffer pH 6.5 at 37 °C. Reactions were performed in a total volume of 350 μl. Samples of 50 μl were taken from the reaction every 3 min, diluted in 50 μl Na-barbital buffer pH 6.5 and subsequently used for determination of reducing ends using the bicinchoninic acid method (Meeuwsen et al., 2000). Six samples were taken from each reaction and all reactions were performed at least in duplicate. The amount of enzyme added depended on the batch, but generally 1 μg was added, representing between 2 10-3 and 2.5 10-3 U, with 1 U defined as the amount of enzyme producing 1 μmol reducing ends min-1. In all assays, reactions with 5 µg of Ni-NTA purified Gus were included to check for background activity. Relative enzyme activities under different conditions were assayed with the same batch of enzyme. The pH optimum was determined by performing the standard reaction at pH values between 4 and 8.5 in Na-barbital buffer. Temperature stability of AmyD was determined using the standard test for starch hydrolysis, performed with enzyme diluted in 10 µl Na-barbital pH 6.5, incubated for 10 min at different temperatures. The Km value of AmyD for starch in the hydrolysis reaction was determined by measuring its activity with 8 different concentrations of starch, varying between 0.01% and 1% (w/v) under standard conditions in triplicate. The effect of NaCl was measured by addition of various concentrations of NaCl (between 50 mM and 0.5 M) to the standard reaction. The hydrolysis of substrates other than potato starch was determined as described above. Proper calibration curves were included in every measurement. Enzymatic reactions for qualitative analysis were performed as follows: standard reactions were performed in a total volume of 10 µl, containing 20 mM disaccharide or oligosaccharide substrate, or 0.2% polysaccharide substrate, or a combination of these, in Na-barbital pH 6.5. Additional reactions with 100 mM or 500 mM maltoheptaose were performed in the same way. AmyD enzyme (1 μg) representing approximately 2 10-3 U, or 5 µg Ni-NTA purified Gus, was added to the reaction mixture and was incubated for 30 min at 37 °C. A total of 2.4 μl of the reaction product was spotted on a TLC plate (Silica gel 60 F254, Merck) and, after drying, the plate was run for 6 h in a small amount of running buffer (butanol/ethanol/MQ 5/5/3 (v/v/v)). After running the plate was dried and sprayed with 50% sulphuric acid in methanol and developed for 10 min at 110 °C. Samples for HPLC (Dionex) analyses were prepared as follows: reactions were performed in 1 ml 1% amylopectin (w/v) or 1% amylose (w/v) in Na-barbital buffer pH 6.5 at 37 °C. AmyD enzyme (2 μg) representing approximately 4 10-3 U, or 10 µg Ni-NTA purified Gus, was added to the reaction and samples of 250 μl were taken after 0,

Chapter 5 A novel cluster of intracellular alpha-amylases
105
10 and 60 min of incubation. Samples were diluted in 1250 μl 90% dimethyl sulfoxide and subsequently used for HPLC analysis performed as described before (Kralj et al., 2004b). Activity staining of AmyD was performed by running 2 μg of Ni-NTA purified protein on SDS-PAGE containing 10% polyacrylamide and 0.12% (w/v) amylopectin. The protein samples were not boiled and neither were denaturing components added in the loading buffer as to preserve enzymatic activity. After separation, the gel was washed and incubated in Na-barbital buffer pH 6.5 at 37 °C for 18 h, and subsequently stained with diluted iodine solution. Iodine stains the amylopectin in the gel purple, except in places where it is degraded by α-amylase activity. Subsequently, the gel was washed and used for the staining of 6xHis-tagged proteins with the InVision staining method (Invitrogen) according to the manufacturer’s instructions. Afterwards, the same gel was stained with Biosafe Coomassie and Silver stain plus (both from Bio-Rad).
Results
Sequence retrieval and analysis
Previous analysis of family GH13 enzymes encoded in the genome sequence of A. niger CBS 513.88 resulted in annotation of two predicted intracellular enzymes, An01g13610 (AmyD) and An01g13610 (AmyE), as novel α-amylases, distantly related to extracellular fungal α-amylases (Pel et al., 2007). A BLASTp search with these two sequences was performed in the available databases (November 2006), yielding nine fungal homologues from A. oryzae, A. fumigatus, A. nidulans, Neurospora crassa and Magnaporthe grisea. A more distantly related homologous sequence from the genome of Cryptococcus neoformans (Loftus et al., 2005) was also included. This set of predicted proteins, combined with the A. niger AmyD and AmyE proteins and Amy1p from H. capsulatum (Marion et al., 2006), was used for a detailed sequence comparison. In A. niger, A. oryzae, A. nidulans and M. grisea, the genes encoding the Amy1p homologues are part of a small cluster of genes which are predicted to be involved in production of cell wall α-glucan. Apart from an Amy1p homolog, these clusters contain genes encoding an α-glucan synthase and a novel type of GPI-anchored family GH13 enzyme. A homologue of these GPI-anchored enzymes in S. pombe was recently identified as important for cell wall α-glucan synthesis (Morita et al., 2006). The retrieved protein sequences of 7 out of the 12 Amy1p homologues were missing one or more of the highly conserved regions specific for family GH13 (Janecek, 2002;Kuriki and Imanaka, 1999). After careful analysis of the original gene sequences, it appeared that the prediction of introns in these genes was not correct. The intron prediction was corrected in compliance with intron consensus elements (Kupfer et al., 2004), thereby

Alpha-glucan acting enzymes in Aspergillus niger
106
restoring full length protein sequences containing all conserved family GH13 residues and regions. In gene EEA30628.1 from N. crassa the position of two expected introns could not be predicted with certainty and thus no correction was made. Although the protein sequence was extended, it could nevertheless be used in alignments because no frameshift occurred within the sequence extensions. In A. oryzae BAE58539.1 the third conserved region could not be restored due to a frame shift, probably caused by a sequencing error. The sequence was nevertheless restored by detailed comparison with the homologous gene from another A. oryzae sequencing project (Uniprot accession number AB078784). None of the (corrected) protein sequences were predicted to have an N-terminal signal for secretion. Two proteins were slightly above the threshold level in the analysis for non-classical secretion (EAA33974.1 and An09g03110). The 13 (corrected) protein sequences together with two novel α-glucanotransferases from A. niger (An-AgtA and An-AgtB) were aligned with 25 α-amylases from representative taxa of the three kingdoms of life: bacteria, archaea and eukarya. The alignment that spanned the entire catalytic (β/α)8-barrel, including domain B (the β3→α3 insertion), served for calculating the phylogenetic tree (Fig. 1). The tree clearly shows the high mutual similarity of the 13 novel GH13 proteins described in this study, and their high similarity to a group of bacterial α-amylases of the liquefying type recently grouped in subfamily GH13_5, represented here by proteins from Bacillus stearothermophilus (Bacst in Fig. 1) and Streptococcus mutans (Stcmu) (Stam et al., 2006). For example, the similarity of AmyD with the bacterial proteins Bacst and Stcmu was 51-56%, while its similarity with the archaeal proteins included was 33%. Interestingly, a protein sequence from C. neoformans (Crcne), a basidiomycete, clustered between the GH13_5 proteins from ascomycete fungi and bacteria (Fig. 1). The 13 intracellular fungal proteins are clearly unrelated to all previously identified extracellular fungal α-amylases grouped presently in the subfamily GH13_1, represented by A. niger acid amylase (Aspni) and A. oryzae TAKA-amylase (Aspor). The recently identified α-glucanotransferases AgtA and AgtB cluster with the extracellular fungal α-amylases, rather than with the intracellular group. The bacterial α-amylases form several clusters in the tree, reflecting their sequence similarities to enzymes from either fungi, plants or animals, as described previously (Janecek, 1994;Janecek et al., 1999;Da Lage et al., 2004).
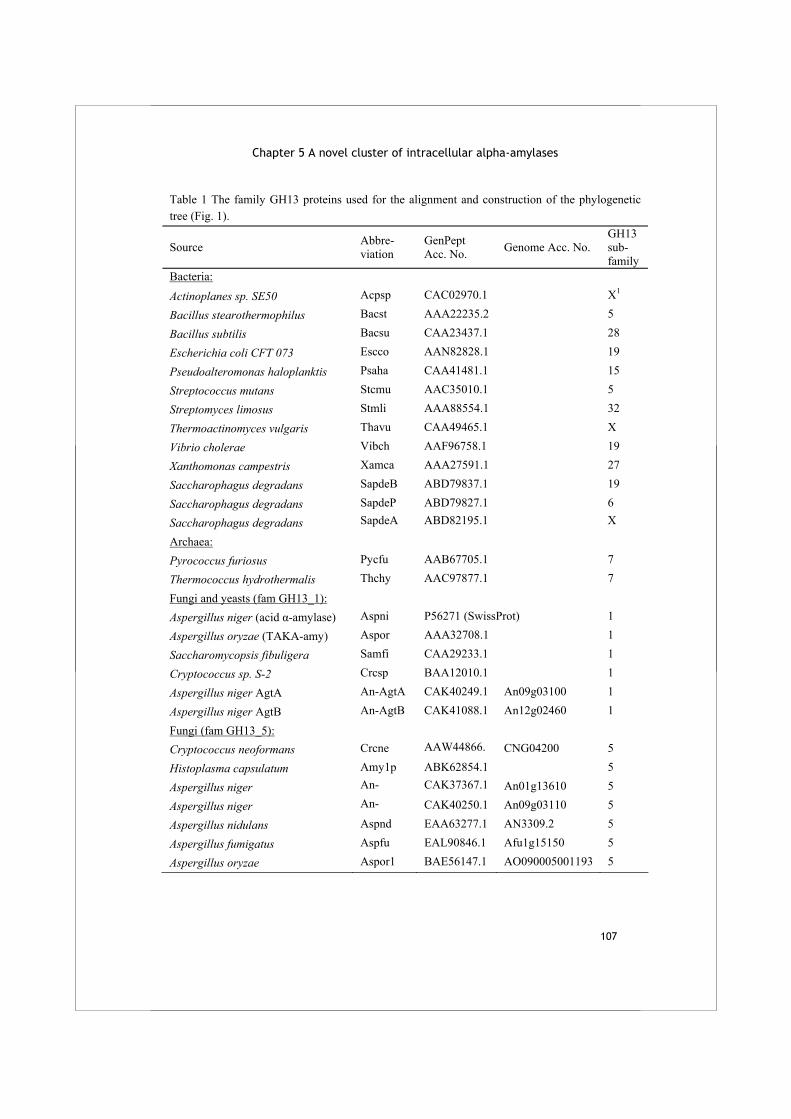
Chapter 5 A novel cluster of intracellular alpha-amylases
107
Table 1 The family GH13 proteins used for the alignment and construction of the phylogenetic tree (Fig. 1).
Source Abbre- viation
GenPept Acc. No. Genome Acc. No.
GH13 sub- family
Bacteria: Actinoplanes sp. SE50 Acpsp CAC02970.1 X1
Bacillus stearothermophilus Bacst AAA22235.2 5
Bacillus subtilis Bacsu CAA23437.1 28
Escherichia coli CFT 073 Escco AAN82828.1 19
Pseudoalteromonas haloplanktis Psaha CAA41481.1 15
Streptococcus mutans Stcmu AAC35010.1 5
Streptomyces limosus Stmli AAA88554.1 32
Thermoactinomyces vulgaris Thavu CAA49465.1 X
Vibrio cholerae Vibch AAF96758.1 19
Xanthomonas campestris Xamca AAA27591.1 27
Saccharophagus degradans SapdeB ABD79837.1 19
Saccharophagus degradans SapdeP ABD79827.1 6
Saccharophagus degradans SapdeA ABD82195.1 X
Archaea: Pyrococcus furiosus Pycfu AAB67705.1 7
Thermococcus hydrothermalis Thchy AAC97877.1 7
Fungi and yeasts (fam GH13_1): Aspergillus niger (acid α-amylase) Aspni P56271 (SwissProt) 1
Aspergillus oryzae (TAKA-amy) Aspor AAA32708.1 1
Saccharomycopsis fibuligera Samfi CAA29233.1 1
Cryptococcus sp. S-2 Crcsp BAA12010.1 1
Aspergillus niger AgtA An-AgtA CAK40249.1 An09g03100 1
Aspergillus niger AgtB An-AgtB CAK41088.1 An12g02460 1
Fungi (fam GH13_5):
Cryptococcus neoformans Crcne AAW44866. CNG04200 5
Histoplasma capsulatum Amy1p ABK62854.1 5
Aspergillus niger An- CAK37367.1 An01g13610 5
Aspergillus niger An- CAK40250.1 An09g03110 5
Aspergillus nidulans Aspnd EAA63277.1 AN3309.2 5
Aspergillus fumigatus Aspfu EAL90846.1 Afu1g15150 5
Aspergillus oryzae Aspor1 BAE56147.1 AO090005001193 5

Alpha-glucan acting enzymes in Aspergillus niger
108
Table 1 (continued) Aspergillus oryzae Aspor2 BAE58539.1 AO090003001497 5
Neurospora crassa Neucr1 EEA30628.1 NCU05873.1 5
Neurospora crassa Neucr2 EAA33974.1 NCU09486.1 5
Magnaporthe grisea Maggr1 EAA51692.1 MG03287.4 5 Magnaporthe grisea Maggr2 EAA48105.1 MG09642.4 5
Magnaporthe grisea Maggr3 EAA48034.1 MG09164.4 5 Plants: Hordeum vulgare (barley – high pI) HorvuH AAA98790.1 6
Malus domestica (apple) Maldo AAF63239.1 6
Phaseolus vulgaris (kidney bean) Phavu BAA33879.1 6
Animals: Drosophila melanogaster (fruit fly) Drome CAA28238.1 15
Homo sapiens (human, saliva) Homsa AAA52279.1 24
Litopenaeus vannamei (shrimp) Penva CAA54524.1 24 1 X, not assigned to a subfamily The thirteen putative intracellular fungal α-amylases share several sequence features with the bacterial enzymes in family GH13_5. These features are, or may be, invariant among the intracellular fungal enzymes and the related bacterial enzymes, but in most cases have no (conserved) equivalent in the other α-amylases studied here (Fig. 2). These specific sequence features include: (i) histidine (His57, AmyD numbering) in the region flanking the strand β2 from the N-terminus; (ii) arginine (Arg60) and cysteine (Cys82) flanking strand β2 at the C-terminus, which, although occurring highly specifically in most fungal GH13_5 enzymes, are absent from the C. neoformans sequence representing the basidiomycete fungi; (iii) an almost invariant leucine residue preceding the conserved NH in conserved region I, at the end of the β3-strand region (136_DAVLNH); (iv) two aromatic residues succeeding the catalytic glutamate proton donor in conserved region III (around strand β5) (297_EYWR); (v) a cysteine and a leucine in the region covering the β8-strand (392_GQPCIFWGDL), which are invariant in all fungal proteins but only partly conserved among the related bacterial sequences. In addition to the catalytic (β/α)8-barrel, domain B is also highly specific for the intracellular fungal enzymes and the bacterial enzymes in family GH13_5. With approximately 105 amino acids, this domain is longer than typically found in fungal α-amylases in family GH13_1, with a B-domain of around 65 amino acids.

Chapter 5 A novel cluster of intracellular alpha-amylases
109
Fig. 1 Phylogenetic tree of family GH13 enzymes from a variety of organisms. The abbreviations used are explained in Table 1. The sources of the α-amylase enzymes are indicated. The tree is based on the alignment made in ClustalW of the part of the sequences encoding the (β/α)8 barrel. The two separate groups of fungal enzymes, intracellular (encircled, GH13_5) and extracellular (GH13_1), are clearly distinguishable. The branch lengths are proportional to the sequence divergence. Numbers along branches are bootstrap values (1,000 replicates). The scale bar (bottom left corner) indicates 0.1 amino acid replacements per site.

Alpha-glucan acting enzymes in Aspergillus niger
110
Fig. 2 Parts of the alignment of family GH13 α-amylase proteins used for construction of a phylogenetic tree (Fig. 1) showing the amino acids specifically conserved among subfamily GH13_5. Abbreviations are explained in Table 1. Names of fungal proteins are underlined.
Biochemical properties of A. niger AmyD
AmyD from A. niger, encoded by An01g13610, was overproduced, purified over Ni-NTA, and characterized to gain insight in the biochemical properties of the group of intracellular fungal α-amylases. PCR with specific primers did not result in retrieval of amyD or amyC (An09g03110) cDNA from cDNA libraries constructed from A. niger strain N402 grown on starch or inulin (van der Kaaij et al., 2007). Therefore, a synthetic gene encoding the predicted coding sequence of An01g13610 was made, and codon usage was optimised for expression in E. coli. Production of AmyD in E. coli was only observed when the host was grown at 16-18 °C, and the yield was relatively low
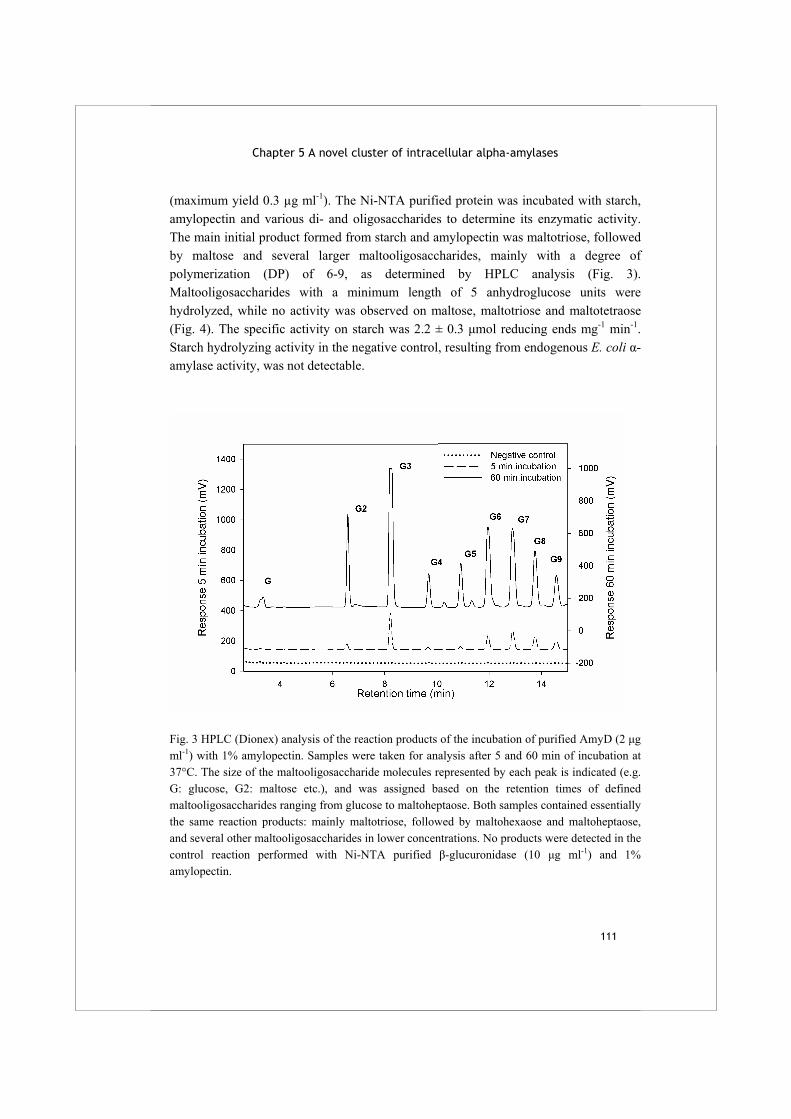
Chapter 5 A novel cluster of intracellular alpha-amylases
111
(maximum yield 0.3 µg ml-1). The Ni-NTA purified protein was incubated with starch, amylopectin and various di- and oligosaccharides to determine its enzymatic activity. The main initial product formed from starch and amylopectin was maltotriose, followed by maltose and several larger maltooligosaccharides, mainly with a degree of polymerization (DP) of 6-9, as determined by HPLC analysis (Fig. 3). Maltooligosaccharides with a minimum length of 5 anhydroglucose units were hydrolyzed, while no activity was observed on maltose, maltotriose and maltotetraose (Fig. 4). The specific activity on starch was 2.2 ± 0.3 μmol reducing ends mg-1 min-1. Starch hydrolyzing activity in the negative control, resulting from endogenous E. coli α-amylase activity, was not detectable.
Fig. 3 HPLC (Dionex) analysis of the reaction products of the incubation of purified AmyD (2 μg ml-1) with 1% amylopectin. Samples were taken for analysis after 5 and 60 min of incubation at 37°C. The size of the maltooligosaccharide molecules represented by each peak is indicated (e.g. G: glucose, G2: maltose etc.), and was assigned based on the retention times of defined maltooligosaccharides ranging from glucose to maltoheptaose. Both samples contained essentially the same reaction products: mainly maltotriose, followed by maltohexaose and maltoheptaose, and several other maltooligosaccharides in lower concentrations. No products were detected in the control reaction performed with Ni-NTA purified β-glucuronidase (10 μg ml-1) and 1% amylopectin.
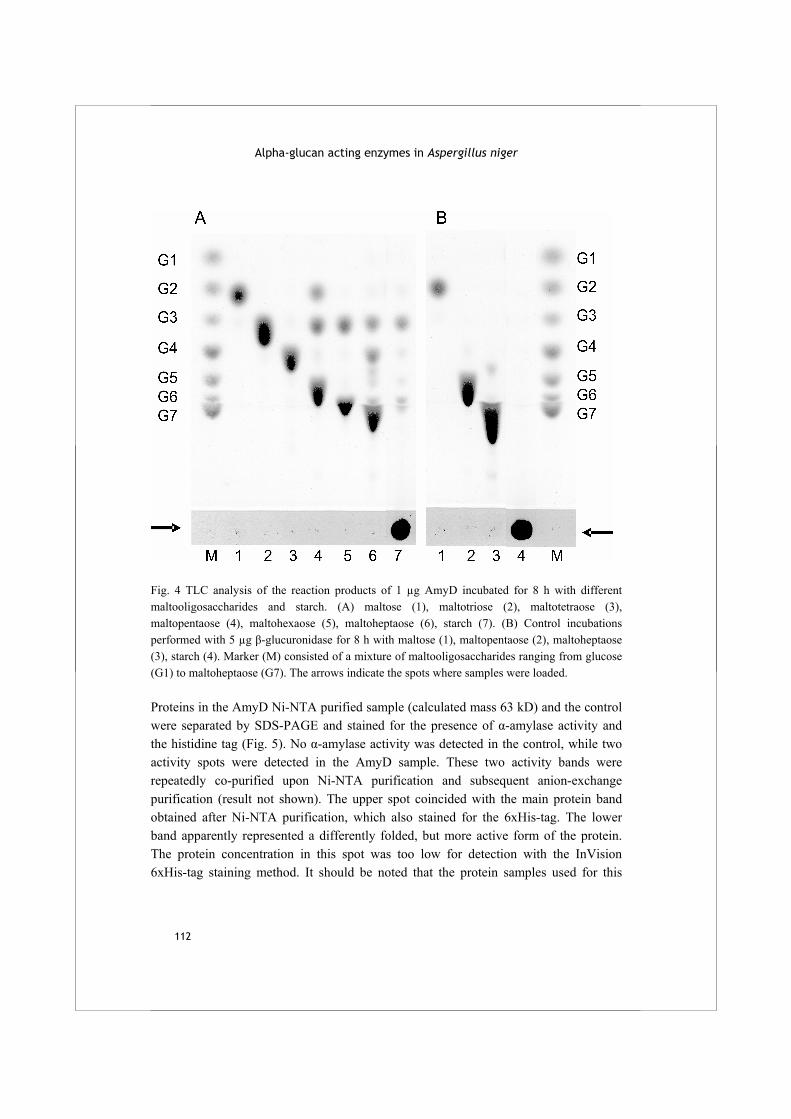
Alpha-glucan acting enzymes in Aspergillus niger
112
Fig. 4 TLC analysis of the reaction products of 1 µg AmyD incubated for 8 h with different maltooligosaccharides and starch. (A) maltose (1), maltotriose (2), maltotetraose (3), maltopentaose (4), maltohexaose (5), maltoheptaose (6), starch (7). (B) Control incubations performed with 5 µg β-glucuronidase for 8 h with maltose (1), maltopentaose (2), maltoheptaose (3), starch (4). Marker (M) consisted of a mixture of maltooligosaccharides ranging from glucose (G1) to maltoheptaose (G7). The arrows indicate the spots where samples were loaded. Proteins in the AmyD Ni-NTA purified sample (calculated mass 63 kD) and the control were separated by SDS-PAGE and stained for the presence of α-amylase activity and the histidine tag (Fig. 5). No α-amylase activity was detected in the control, while two activity spots were detected in the AmyD sample. These two activity bands were repeatedly co-purified upon Ni-NTA purification and subsequent anion-exchange purification (result not shown). The upper spot coincided with the main protein band obtained after Ni-NTA purification, which also stained for the 6xHis-tag. The lower band apparently represented a differently folded, but more active form of the protein. The protein concentration in this spot was too low for detection with the InVision 6xHis-tag staining method. It should be noted that the protein samples used for this
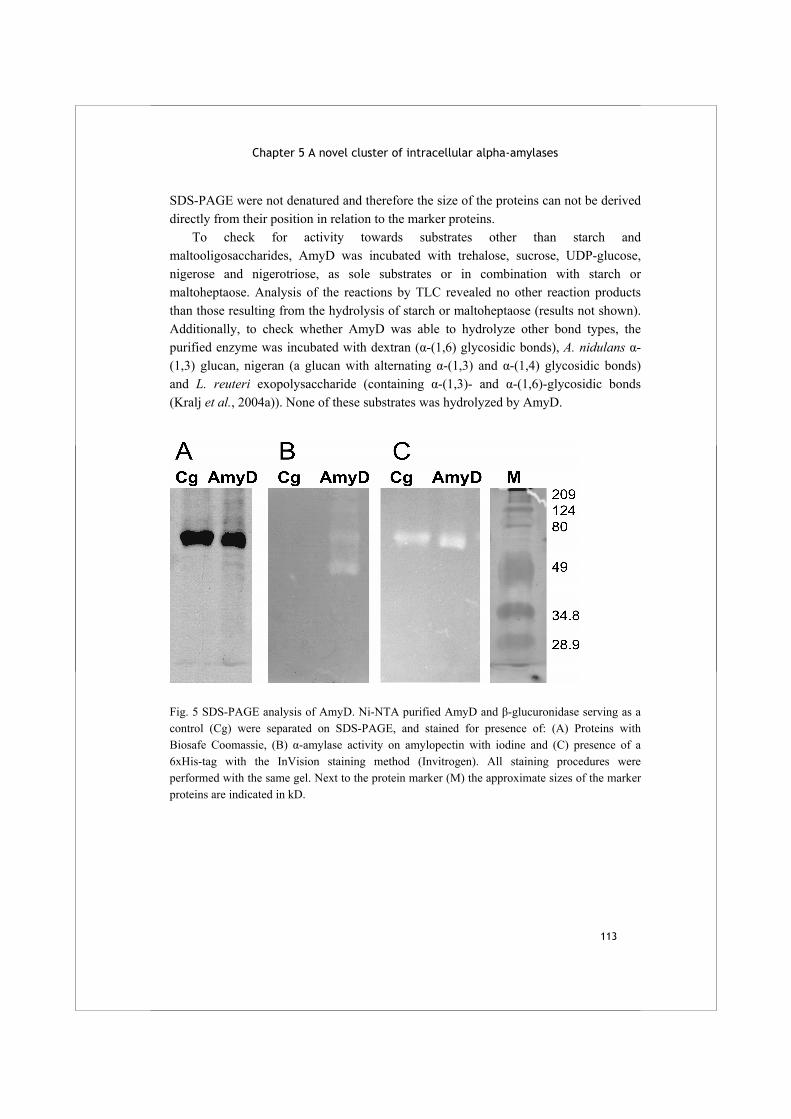
Chapter 5 A novel cluster of intracellular alpha-amylases
113
SDS-PAGE were not denatured and therefore the size of the proteins can not be derived directly from their position in relation to the marker proteins. To check for activity towards substrates other than starch and maltooligosaccharides, AmyD was incubated with trehalose, sucrose, UDP-glucose, nigerose and nigerotriose, as sole substrates or in combination with starch or maltoheptaose. Analysis of the reactions by TLC revealed no other reaction products than those resulting from the hydrolysis of starch or maltoheptaose (results not shown). Additionally, to check whether AmyD was able to hydrolyze other bond types, the purified enzyme was incubated with dextran (α-(1,6) glycosidic bonds), A. nidulans α-(1,3) glucan, nigeran (a glucan with alternating α-(1,3) and α-(1,4) glycosidic bonds) and L. reuteri exopolysaccharide (containing α-(1,3)- and α-(1,6)-glycosidic bonds (Kralj et al., 2004a)). None of these substrates was hydrolyzed by AmyD.
Fig. 5 SDS-PAGE analysis of AmyD. Ni-NTA purified AmyD and β-glucuronidase serving as a control (Cg) were separated on SDS-PAGE, and stained for presence of: (A) Proteins with Biosafe Coomassie, (B) α-amylase activity on amylopectin with iodine and (C) presence of a 6xHis-tag with the InVision staining method (Invitrogen). All staining procedures were performed with the same gel. Next to the protein marker (M) the approximate sizes of the marker proteins are indicated in kD.
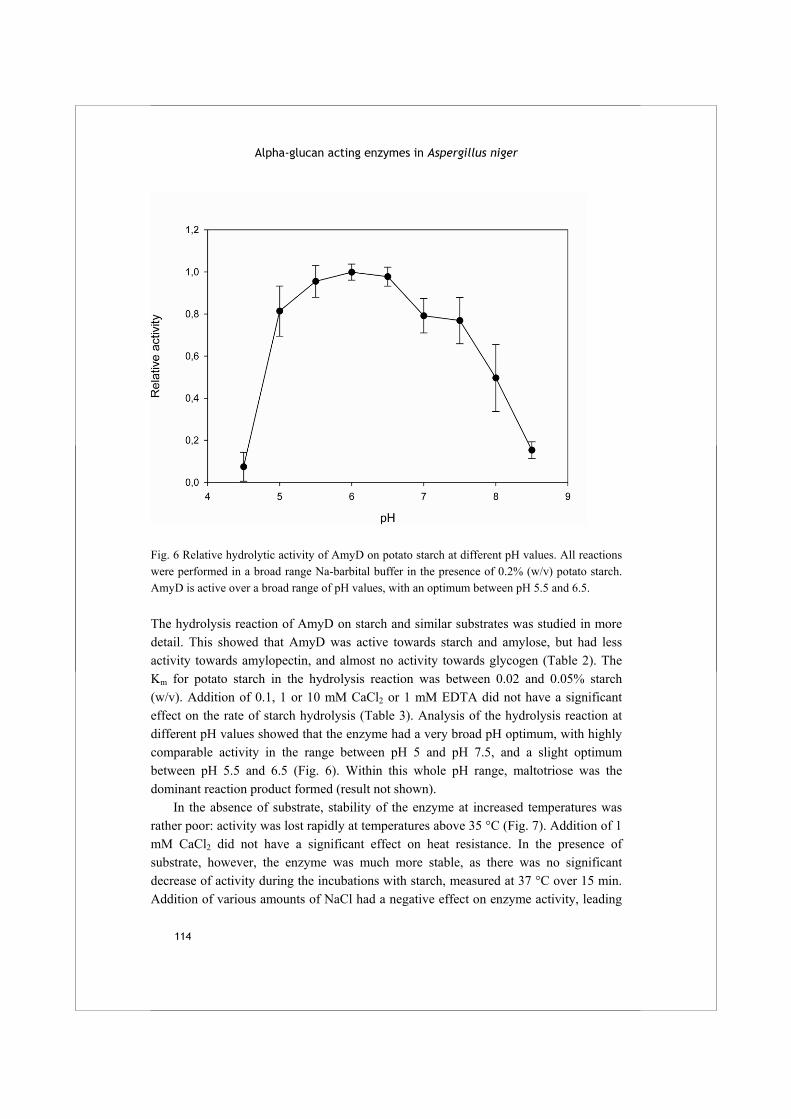
Alpha-glucan acting enzymes in Aspergillus niger
114
Fig. 6 Relative hydrolytic activity of AmyD on potato starch at different pH values. All reactions were performed in a broad range Na-barbital buffer in the presence of 0.2% (w/v) potato starch. AmyD is active over a broad range of pH values, with an optimum between pH 5.5 and 6.5. The hydrolysis reaction of AmyD on starch and similar substrates was studied in more detail. This showed that AmyD was active towards starch and amylose, but had less activity towards amylopectin, and almost no activity towards glycogen (Table 2). The Km for potato starch in the hydrolysis reaction was between 0.02 and 0.05% starch (w/v). Addition of 0.1, 1 or 10 mM CaCl2 or 1 mM EDTA did not have a significant effect on the rate of starch hydrolysis (Table 3). Analysis of the hydrolysis reaction at different pH values showed that the enzyme had a very broad pH optimum, with highly comparable activity in the range between pH 5 and pH 7.5, and a slight optimum between pH 5.5 and 6.5 (Fig. 6). Within this whole pH range, maltotriose was the dominant reaction product formed (result not shown). In the absence of substrate, stability of the enzyme at increased temperatures was rather poor: activity was lost rapidly at temperatures above 35 °C (Fig. 7). Addition of 1 mM CaCl2 did not have a significant effect on heat resistance. In the presence of substrate, however, the enzyme was much more stable, as there was no significant decrease of activity during the incubations with starch, measured at 37 °C over 15 min. Addition of various amounts of NaCl had a negative effect on enzyme activity, leading

Chapter 5 A novel cluster of intracellular alpha-amylases
115
to approximately 75% of activity in the presence of 500 mM NaCl, compared to the standard reaction conditions (Table 4). Table 2: Relative hydrolytic activity of AmyD on different substrates. All reactions were performed with the same batch of purified enzyme and under standard conditions (0.2% (w/v) substrate concentration, pH 6.5). The 100% activity level was 2.2 U mg-1.
Substrate Relative activity Potato starch 100 ± 2.6
Amylose 101 ± 4.2 Amylopectin 66 ± 2.6
Glycogen 3.6 ± 5.2 Table 3 Relative hydrolytic activity of AmyD in the presence of Ca2+ or EDTA. The 100% activity level was 2.2 U mg-1.
[Ca2+] (mM) Relative activity 0 100 ± 5.8
0.1 105 ± 7.1 1 98.0 ± 16.8
10 96.3 ± 18.8 [EDTA] (1 mM) 100.2 ± 13.9
Table 4 Relative hydrolytic activity of AmyD in the presence of different concentrations of NaCl. The 100% activity level was 2.4 U mg-1
[NaCl] (mM) Relative activity 0 100 ± 2.7
50 89.0 ± 7.9 100 83.7 ± 4.3 500 76.2 ± 0.2 1000 74.3 ± 2.9
Discussion The recent release of the full genomic sequence of several fungal species has boosted research into various physiological processes in these organisms. Even in areas which where relatively well studied, such as the degradation of starch, the information on novel sequences has raised many new questions. In the aspergilli, as first observed by Pel et al. (2007), a novel group of α-amylases was observed with very low similarity to all other fungal α-amylases described thus far. Proteins belonging to this new fungal α-

Alpha-glucan acting enzymes in Aspergillus niger
116
amylase group characteristically have high mutual sequence similarity and a predicted intracellular location. According to the recent division of family GH13 enzymes into subfamilies, the extracellular fungal α-amylases are classified into group GH13_1. We conclude that the novel intracellular fungal α-amylases described here are members of subfamily GH13_5, displaying - within the frame of the entire α-amylase family GH13 - the highest sequence similarity to the bacterial enzymes earlier assigned to this subfamily (Stam et al., 2006). Their close evolutionary relationship was confirmed by the existence of several amino acids specifically conserved within this subfamily (Fig. 2). However, these did not include amino acids which are known to be predictive for certain enzyme reaction or product specificities within family GH13, such as hydrolysis of α-(1,6) glycosidic bonds, or α-glucanotransferase activity (as reviewed by MacGregor et al. (2001)). The specific functions of these conserved amino acid residues thus remain to be determined. The 13 protein sequences subjected to sequence analysis in this paper can be considered examples of an undoubtedly much larger group of fungal α-amylases. Homologous proteins were identified in the genome sequences of the Ascomycetes Botrytis cinerea, Chaetomium globosum and Sclerotinia sclerotiorum (http://www.broad.mit.edu/annotation/fungi/fgi/index.html). In many fungal species, the organisation of the genes encoding the GH13_5 enzymes is highly conserved, strengthening the hypothesis that these enzymes have the same, or strongly related physiological functions. The phylogenetic analysis of the fungal sequences has shown that these α-amylases are positioned on a common branch with a group of bacterial α-amylases previously described as related to plant α-amylases (Da Lage et al., 2004). The similarity between the two groups implicates an evolutionary relationship, as observed previously for a cluster of bacterial enzymes with similarity to animal α-amylases, suggested to be a result of horizontal gene transfer (Da Lage et al., 2004). The bacterial α-amylases belonging to subfamily GH13_5 include enzymes from Bacillus and Cytophaga species (Yuuki et al., 1985;Jeang et al., 2002;Kanai et al., 2004), some of which are thermostable (Kim et al., 2000) and used in industry (Guzman-Maldonado and Paredes-Lopez, 1995). Several of these bacterial enzymes are known to produce oligosaccharides with specific lengths from starch, including for example a maltohexaose forming α-amylase from Bacillus (Kanai et al., 2004). To analyse whether similar properties are also encountered in the related fungal enzymes, we produced and characterised a randomly chosen homologue from this cluster, AmyD from A. niger. The biochemical analysis showed that, similar to the related bacterial enzymes, AmyD produces mainly maltotriose from starch, amylose and amylopectin. Similar to other α-amylases, AmyD displayed a relatively low activity on highly branched substrates like glycogen. Increased stability or activity in the presence of Ca2+,
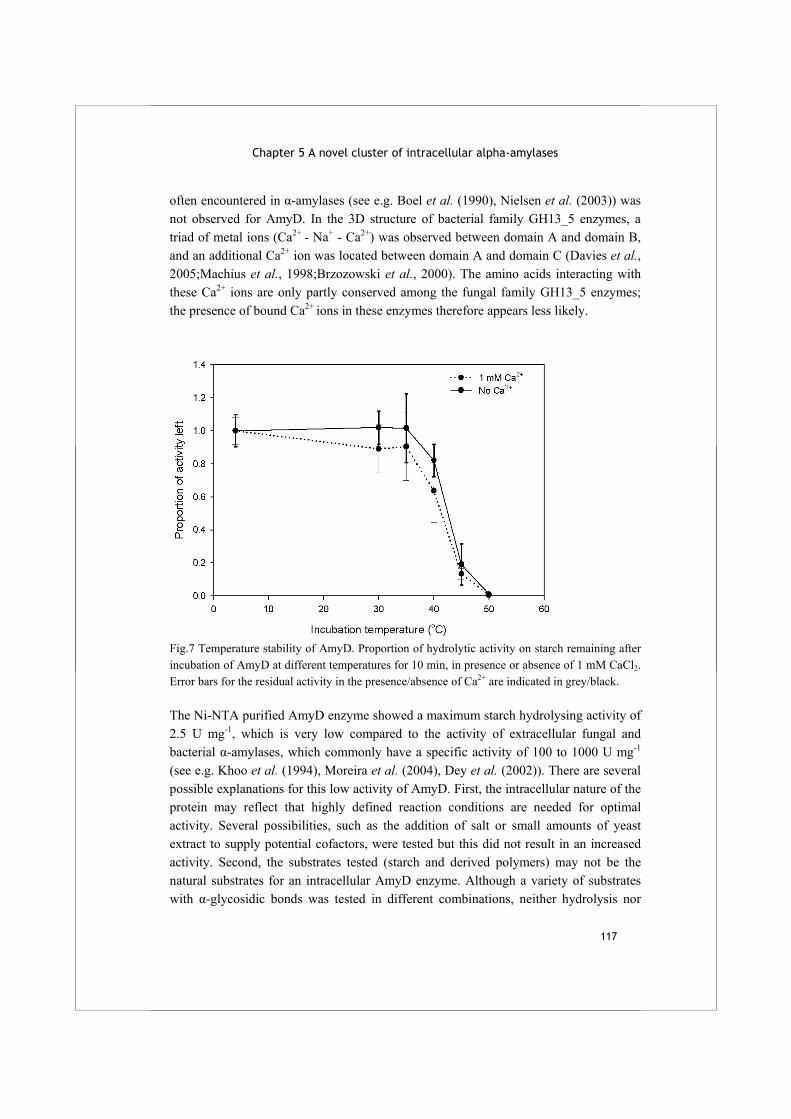
Chapter 5 A novel cluster of intracellular alpha-amylases
117
often encountered in α-amylases (see e.g. Boel et al. (1990), Nielsen et al. (2003)) was not observed for AmyD. In the 3D structure of bacterial family GH13_5 enzymes, a triad of metal ions (Ca2+ - Na+ - Ca2+) was observed between domain A and domain B, and an additional Ca2+ ion was located between domain A and domain C (Davies et al., 2005;Machius et al., 1998;Brzozowski et al., 2000). The amino acids interacting with these Ca2+ ions are only partly conserved among the fungal family GH13_5 enzymes; the presence of bound Ca2+ ions in these enzymes therefore appears less likely.
Fig.7 Temperature stability of AmyD. Proportion of hydrolytic activity on starch remaining after incubation of AmyD at different temperatures for 10 min, in presence or absence of 1 mM CaCl2. Error bars for the residual activity in the presence/absence of Ca2+ are indicated in grey/black. The Ni-NTA purified AmyD enzyme showed a maximum starch hydrolysing activity of 2.5 U mg-1, which is very low compared to the activity of extracellular fungal and bacterial α-amylases, which commonly have a specific activity of 100 to 1000 U mg-1 (see e.g. Khoo et al. (1994), Moreira et al. (2004), Dey et al. (2002)). There are several possible explanations for this low activity of AmyD. First, the intracellular nature of the protein may reflect that highly defined reaction conditions are needed for optimal activity. Several possibilities, such as the addition of salt or small amounts of yeast extract to supply potential cofactors, were tested but this did not result in an increased activity. Second, the substrates tested (starch and derived polymers) may not be the natural substrates for an intracellular AmyD enzyme. Although a variety of substrates with α-glycosidic bonds was tested in different combinations, neither hydrolysis nor

Alpha-glucan acting enzymes in Aspergillus niger
118
transglycosylation reactions were observed to occur on any of these substrates. Therefore it appears likely that the observed hydrolysis of maltooligosaccharides is the natural reaction of AmyD. A third explanation is that a high AmyD activity is not needed for its physiological function in the cell. The data available on specific activities all relate to fungal α-amylases acting in the extracellular environment. The latter enzymes have been selected for their ability to rapidly hydrolyse substrates, in order to minimize the chances for competing organisms to use the same carbon source. Therefore, a comparison of the activity levels of intra- and extracellular α-amylases enzymes is not a proper one. A possible physiological function for the fungal intracellular α-amylases was provided by the study of the AmyD homologue Amy1p of H. capsulatum. Although the enzymatic activity of this protein has not been studied, a clear link was made with the production of cell wall α-glucan, as a functional knockout was completely unable of producing α-(1,3) glucan (Marion et al., 2006). This hypothesis on the function of fungal GH13_5 α-amylases is strengthened by the genomic organization of the encoding genes, many of which are arranged in a small cluster of genes also encoding an α-glucan synthase and a GPI-anchored α-glucanotransferase. Such an arrangement of genes is observed for e.g. A. niger amyE but not for amyD, and for A. oryzae AO090003001497 but not for AO090005001193. Additionally, regulation of these α-amylases by AmyR was not observed in A. niger nor in A. nidulans, contrary to what would have been expected for enzymes involved in starch degradation (see Chapter 2) (Nakamura et al., 2006). Recently, the first detailed structural analysis of fungal cell wall α-glucan was performed in S. pombe (Grün et al., 2005). On the basis of these data, the authors proposed a model which suggested that a small molecule consisting of α-(1,4) linked glucose residues acts as a primer for the formation of longer, α-(1,3) linked polymers by α-glucan synthases. As proposed previously by Marion et al. (2006), the function of Amy1p, AmyD and related proteins could be to produce such a primer molecule. In this study, we have shown that AmyD indeed produces small maltooligosaccharides, with a preference for maltotriose in vitro. The combination of previously published data on Amy1p, the genomic organization of fungal family GH13_5 genes, and the biochemical characterization of A. niger AmyD strongly suggest the involvement of these enzymes in cell wall α-glucan formation. Further studies on the formation of cell wall α-glucans in fungi, a process which is still relatively obscure, are needed to confirm this involvement and to determine the role of AmyD and other enzymes involved.

Chapter 5 A novel cluster of intracellular alpha-amylases
119
Acknowledgements
We are very grateful to Harrie Kools of Wageningen University for help with the HMMR searches and to Peter Sanders of TNO Quality of Life for the HPLC analysis. We thank DSM (The Netherlands) for providing the genome sequence of Aspergillus niger. SJ thanks the Slovak Grant Agency for the VEGA grant No. 2/5067/27.


121
Chapter 6 Summary and concluding remarks

Alpha-glucan acting enzymes in Aspergillus niger
122
Fungi are eukaryotic organisms, characteristically highly variable in size, shape and natural habitat. Within the phylum of ascomycete fungi, the aspergilli form a well studied genus with more than 250 separate species. Some of these, such as Aspergillus fumigatus and Aspergillus flavus, are known to be human pathogens. The filamentous fungus Aspergillus niger is a common soil fungus with many industrial applications. It produces a large variety of enzymes with applications in the food and paper industry. This mould, named after the black appearance of its sporulating colonies, is the organism studied in this thesis. Fitting to their chemotrophic and heterotrophic lifestyle, fungi are able to degrade various polysaccharides; they import the small carbohydrates released to serve as energy and carbon source. Fungi produce various types of extracellular enzymes specialized in hydrolysis of specific carbohydrates. The expression of these enzymes is often tightly regulated, so that only enzymes needed to degrade the carbohydrate at hand are produced and excreted by the fungus. An example of such a system of carbohydrate degradation by fungi is the hydrolysis of starch, a polymer abundantly present in plant storage organs. Starch consists of amylose, a linear chain of α-(1,4) linked glucose molecules, and amylopectin, a branched glucan with α-(1,4) and approximately 3-5 % α-(1,6) glycosidic bonds. Fungi produce a mixture of different extracellular enzymes for starch degradation. Alpha-amylases hydrolyze some of the internal α-(1,4) glycosidic bonds, and the resulting maltooligosaccharides are subsequently hydrolyzed into β- and α-glucose by glucoamylases and α-glucosidases, respectively. In aspergilli, the expression of starch-degrading enzymes is induced in the presence of maltose or isomaltose, molecules formed by the partial hydrolysis of starch. This regulation is mediated by the AmyR transcription factor, acting as a positive regulator (Petersen et al., 1999). Alpha-amylases are ubiquitous enzymes, present in all kingdoms of life. Most α-amylases belong to the family 13 of glycoside hydrolases (GH13), according to the classification of carbohydrate acting enzymes based on sequence similarity (see CAZy website at http://www.cazy.org) (Coutinho and Henrissat, 1999). Also family GH57 contains enzymes with α-amylase activity, but these proteins only occur in prokaryotes. Other types of starch acting enzymes, both from prokaryotes and eukaryotes, are classified in family GH15 (glucoamylases) or in family GH31 (α-glucosidases). Apart from enzymes hydrolyzing α-(1,4) glycosidic bonds, family GH13 also comprises enzymes hydrolyzing α-(1,6) glycosidic bonds, and enzymes with α-glucanotransferase activities. Most of the family GH13 enzymes known are involved in degradation or synthesis of starch, glycogen and similar α-glucan polymers. Some enzymes may be

Chapter 6 Summary and concluding remarks
123
involved in metabolism of trehalose, a eukaryotic storage compound, and others act on sucrose. The overall sequence similarity of GH13 enzymes with different reaction specificities may be very low, but they share four conserved amino acid regions which together form the active site of the proteins. Three amino acids within these regions are invariable, and are known to be essential for catalysis. Four other amino acids are highly conserved within the family, and play a role in binding and positioning of the substrate (Nakajima et al., 1986). At the start of this PhD study, the aim of the project was defined as follows: to identify the enzymes from A. niger involved in starch degradation, and to assess their individual roles in this process by studying their biochemical properties and the transcriptional regulation of their encoding genes. These studies were greatly facilitated by the availability of the recently completed genome sequence of A. niger CBS 513.88. Chapter 2 describes the identification of genes encoding enzymes putatively involved in starch degradation in the genome of A. niger, and the effect of maltose on the expression of these genes. Additionally, an amyR deletion strain was constructed to study the effect of this transcriptional regulator on the expression of the genes identified. The genome search for family GH13, GH15 and GH31 enzymes yielded a total of 17 novel predicted proteins. Thus, although A. niger already represents a rich source for commercial starch acting enzymes, analysis of its genome sequence revealed that it encodes a surprisingly large number of unknown, putatively starch acting enzymes. These enzymes are potentially interesting both from a scientific and from an industrial point of view. Only a small number of these newly identified enzymes were subsequently predicted to act in vivo on starch in the extracellular environment. These predictions were based on the presence or absence of a putative signal sequence for secretion in combination with expression data. Apart from the previously described acid amylase (Boel et al., 1990) and AmyA and AmyB (Korman et al., 1990), we identified one extracellular α-amylase designated AmyC. The amyC gene is located in close proximity to the amyR gene, but its expression was relatively low and not as tightly regulated by AmyR as that of the other α-amylases. This protein possibly acts as a scouting enzyme, producing maltose to initiate the expression of other starch degrading enzymes regulated by AmyR. Among the α-glucosidases of family GH31, only a few members were clearly regulated by AmyR and expressed in the presence of maltose, suggesting a role in starch degradation. Many other family GH13, GH15 and GH31 enzymes identified in the A. niger genome were predicted not to be involved in starch degradation, for a variety of reasons. For example, many of the predicted enzymes from all three families lacked a signal sequence for secretion. These enzymes are therefore thought to function in the

Alpha-glucan acting enzymes in Aspergillus niger
124
intracellular environment, although it is unclear what their functions in the cell might be. Alternatively, the enzymes might be released into the environment upon autolysis of the mycelium. This hypothesis is supported by the observed upregulation of chitinases during the autolysis phase of A. niger (Yamazaki et al., 2007), and the finding of a predicted intracellular laccase in the culture filtrate of the white rot fungus Termitomyces clypeatus (Bose et al., 2007). Expression of the intracellular GH13, 15, 31 enzymes in A. niger was very poor or not detectable in our experiments, all of which were performed with rather young, submerged cultures. An exception was formed by enzymes involved in glycogen metabolism. The production and degradation of this intracellular storage molecule involves, amongst others, two family GH13 enzymes (glycogen branching and debranching enzyme), and both these enzymes were expressed in our A. niger cultures. Based on a phylogenetic analysis, we identified three novel subgroups of family GH13 enzymes in the genome sequence of A. niger (chapter 2), comprising respectively two predicted intracellular α-glucosidases, two predicted intracellular α-amylases, and three extracellular enzymes that were predicted to be anchored on the cell wall with a glycosylphosphatidylinositol (GPI) anchor. The same groups with homologous enzymes were also predicted from the genome sequences of other aspergilli, and partly in other Ascomycetes and Basidiomycetes as well. The characteristics of the predicted intracellular α-amylases and the GPI-anchored enzymes were studied in more detail, as described in chapters 3, 4 and 5 of this thesis. Chapters 3 and 4 describe the detailed characterization of two novel family GH13 α-glucanotransferase enzymes from A. niger, predicted to be localized in the cell wall. The enzymes, named α-glucanotransferase (Agt) A and B, showed transglycosylation activity on substrates with α-(1,4)-glycosidic bonds. The minimum size of the donor substrate was maltopentaose, and the minimal size for the acceptor substrate was maltose. The enzymes produced new α-(1,4)-glycosidic bonds (EC 2.4.1.25) and their reaction products reached a degree of polymerization (DP) of at least 30. This is the first time that enzymes with such a disproportionating type of reaction are described in fungi. Based on similar sequence properties, a third enzyme predicted from the genome sequence was named AgtC. Although the basic reaction of the two studied Agt enzymes is very similar, a more detailed characterization (chapter 4) revealed significant differences. When tested with a synthetic donor substrate, the specific transglycosylation activity of AgtA was ten times higher than the activity of AgtB. AgtB showed a broad temperature optimum and relatively poor heat stability, compared to AgtA. Both enzymes where able to use nigerose and nigerotriose as acceptor substrates (with α-(1,3) glycosidic bonds), although only to a limited extent compared to

Chapter 6 Summary and concluding remarks
125
maltooligosaccharide substrates. The enzymatic activity of AgtA and AgtB is similar to that of bacterial amylomaltases, which act on smaller maltooligosaccharides substrates, using maltose as a donor and glucose as acceptor molecule (Kaper et al., 2004). They usually release one glucose molecule for every transglycosylation event, a clear difference with AgtA and B, which do not produce significant amounts of glucose. Most amylomaltases are members of family GH77, although some belong in family GH13. Despite the similar reaction performed by AgtA/B and bacterial disproportionating enzymes such as amylomaltases, there is no significant overall sequence similarity between these groups of enzymes. There is, however, a specific sequence trait shared between fungal Agt enzymes and family GH77 amylomaltases: one of the 7 highly conserved amino acids in GH13, the equivalent of His143 in A. niger acid amylase, is not present (MacGregor et al., 2001;Kaper et al., 2007). It is possible that the absence of His143 plays a decisive role in the determination of disproportionating activity in the Agt enzymes. Interestingly, the same His residue is missing in the GH13 domain of α-glucan synthase (Ags) enzymes, which are also predicted to act as α-glucanotransferases (Grün et al., 2005). On the other hand, His143 is present in other GH13 enzymes with disproportionating activity, notably Thermotoga maritima amylomaltase (Liebl et al., 1992) and cyclodextrin glucanotransferase (CGTase) enzymes, and is known to be important for catalytic activity in these enzymes (Mattsson et al., 1995;Leemhuis et al., 2004). Alternatively, the absence of the equivalent of His143 might have something to do with the in vivo substrate of Agt and Ags enzymes, which is probably a mixed type of α-glucan with both α-(1,4) and α-(1,3) glycosidic linkages (see below). To investigate the possible role of the equivalent of His143 in Agt and Ags enzymes, the in vivo reaction and physiological role of these enzymes need to be elucidated, combined with biochemical studies of wild type and mutant enzymes, with modified amino residues at this position. Homologues of AgtA/B were identified in the genome sequences of many Ascomycetes and Basidiomycetes, e.g. in all available Aspergillus genomes, but also in Botrytis cinerea, Neurospora crassa, the fission yeast Schizosaccharomyces pombe and the Basidiomycete Cryptococcus neoformans. Homologues of Agt enzymes may be recognized by their high sequence similarity to fungal α-amylases, the lack of His143 and the presence of a GPI-anchoring signal. The identified homologues of the Agt enzymes in other fungi have all been annotated as α-amylases, but the biochemical characterization of AgtA and AgtB presented in this thesis shows that reannotation of these predicted enzymes as α-glucanotransferases is required. Expression of the agt genes in A. niger was not induced by maltose and not dependent on the presence of AmyR (chapter 2), suggesting that the proteins are not involved in

Alpha-glucan acting enzymes in Aspergillus niger
126
the catabolism of starch. Rather, agtA was constitutively expressed, and agtB was co-regulated with its neighbouring gene agsC, one of five genes encoding an α-glucan synthase in A. niger. The latter are membrane bound enzymes responsible for the production of cell wall α-glucan (Hochstenbach et al., 1998). The cell wall of aspergilli contains α-(1,3) glucan and nigeran, with alternating α-(1,3) and α-(1,4) glycosidic bonds, as well as other types of polysaccharides. Many of the agt genes in different fungi are located next to, or in the neighbourhood of, ags genes. This conserved clustering and co-expression of agt and ags genes are strong indications that the two types of enzymes serve in the same physiological process, namely the synthesis of cell wall α-glucan. This hypothesis is in line with the absence of agt homologues in the Saccharomycotina which do not produce cell wall α-glucan. A deletion of an agt homologue (aah3) in S. pombe resulted in a strain with an aberrant cell shape which was hypersensitive towards cell wall degrading enzymes, suggesting that the deletion had an effect on cell wall integrity (Morita et al., 2006). In chapter 3, we have shown that both the knockout of agtA and the overexpression of either AgtA or AgtB in A. niger had a clear negative effect on its susceptibility towards CalcoFluorWhite (CFW), a cell wall disrupting compound (Ram and Klis, 2006). This is a strong indication that cell wall strength is impaired in these strains, caused by the knockout or overexpression of agt genes. However, such an effect of CFW might also be indirect and caused by the disturbance of other cellular functions. The deletion of S. pombe aah3, for example, also caused a defect in vacuolar protein sorting, probably representing an indirect effect of the impaired integrity of the cell wall (Iwaki et al., 2007). More evidence for the function of Agt enzymes might be obtained from the comparative analysis of the cell wall α-glucan components of A. niger wild type, and the ΔagtA and AgtA/B overexpression strains. At present we cannot exclude that AgtB or AgtC partly replace the function of AgtA in the ΔagtA strain. Therefore, also agtA/B/C double and triple knockouts should be constructed in further work, possibly showing even more pronounced effects on cell wall physiology. In chapter 5, a cluster of predicted intracellular fungal α-amylases was studied in more detail. Phylogenetic analysis of this group of 12 protein sequences from 7 different fungal species demonstrated that they belong to subfamily GH13_5, a subfamily previously thought to contain only bacterial α-amylases (Stam et al., 2006). These proteins show only very little similarity to the fungal α-amylases described previously, which all belong to subfamily GH13_1. We expressed one of these enzymes, AmyD from A. niger, in E. coli and studied its biochemical characteristics. The enzyme showed a low hydrolyzing activity on starch and similar substrates, a broad pH optimum and rather poor temperature stability. The activity or stability of the enzyme was not dependent on Ca2+ ions, although analysis of the 3D structures of related bacterial

Chapter 6 Summary and concluding remarks
127
enzymes revealed the presence of least three Ca2+ ions. A homologue of AmyD, Amy1p from Histoplasma capsulatum, was identified in a search for genes involved in biosynthesis of cell wall α-(1,3) glucan (Marion et al., 2006). A functional knockout of AMY1 in H. capsulatum was unable to produce α-(1,3) glucan and showed reduced virulence. The exact function of Amy1p in α-glucan formation in H. capsulatum has not been studied, but the suggested involvement of family GH13_5 enzymes with α-glucan formation is strengthened by the organization of the genes encoding these enzymes, which are often clustered with agt and ags genes (Fig. 1).
Fig. 1 Schematic representation of the chromosomal organization of some of the A. niger genes discussed in this thesis. An overview of all genes discussed is given in chapter 2, Table 1. Putative AmyR binding sites are indicated with *. Abbreviations used: agdA; α-glucosidase A, amy; amylase, ags; α-glucan synthase, agt; α-glucanotransferase. Two genes encoding hypothetical proteins are indicated with hyp. Gene sizes and distances are not to scale.
The results described in this thesis suggest that enzymes homologous to A. niger AgtA/B and AmyD/E play a role in fungal cell wall α-glucan formation or maintenance. Cell wall α-glucan is thought to be produced by α-glucan synthases (Ags enzymes), but the biochemistry of this process is still largely unknown. Ags enzymes have two predicted catalytic domains; an extracellular domain that resembles family GH13 enzymes, and an intracellular one resembling enzymes of Glycosyltransferase family 5 (GT5) (Hochstenbach et al., 1998). Family GT5 also comprises glycogen synthases, which require a short oligosaccharide as a primer molecule to produce glycogen from UDP- or ADP-glucose (Lomako et al., 2004). A similar oligosaccharide primer might be provided to the GT5 domain of α-glucan synthases by AmyD, which was shown to release mainly maltotriose when incubated with maltooligosaccharides. The amyD and

Alpha-glucan acting enzymes in Aspergillus niger
128
amyE genes, however, were not expressed in A. niger under the conditions tested, although expression of several ags genes was observed (chapter 2). This implies that activity of AmyD/E was not necessary for α-glucan formation under the conditions studied, or that their expression level was too low for detection. A third possibility is that amyD/E were expressed but not detected, as the microarrays used were based on the genome of A. niger CBS 513.88, while experiments were done with strain N402 which might have some differences at DNA level. It is well described that different Ags enzymes have specific physiological functions within a fungal colony, which is also reflected by their expression patterns (Beauvais et al., 2005;Damveld et al., 2005b;Garcia et al., 2006). AmyD/E expression might be regulated similarly, if their activity is only needed in combination with one of the five Ags enzymes specifically. An additional conundrum with respect to the reaction and function of fungal GH13_5 enzymes is their in vivo substrate; AmyD has very poor hydrolyzing activity on glycogen, the only known intracellular glucan polymer. Alternatively, these enzymes might be acting on small intracellular maltooligosaccharides, or even on the cell wall α-glucan itself during the synthesizing process performed by the intracellular GT5 domain of Ags enzymes. This remains to be studied. According to the hypothesis postulated by Hochstenbach et al. (1998), the glucan polymer formed by the intracellular GT5 domain of Ags enzymes is transported over the membrane to the extracellular environment. Studies on the α-glucan composition in S. pombe suggested that the function of the GH13 domain of one of its glucan synthases, Ags1, was to interconnect two α-(1,3) glucan chains via a transglycosylation reaction involving a short stretch of α-(1,4) glycosidic bonds (Grün et al., 2005). The function of AgtA and AgtB might be to perform a similar transglycosylation reaction to couple different α-glucan chains. Assuming that AgtA and AgtB (mainly) act on α-(1,4) linked oligosaccharides in vivo, as they do in vitro, it is unclear which part of the cell wall α-glucan is their natural substrate. The cell walls of S. pombe spores contain an α-(1,4) linked glucan which can be stained with iodine (Garcia et al., 2006). A recent study showed that overexpression of Ags1 resulted in overproduction of α-(1,4) linked glucan in S. pombe (Vos et al., 2007). One study also reported the formation of an amylose-like glucan in A. niger grown under physical stress in shaking cultures, but this polysaccharide was only loosely attached to the mycelium and did not appear to be a structural component of the cell wall (Kirimura et al., 1999). Additionally, mycelium and spores of A. niger strain MGG029-ΔaamA, a mutant strain with white mycelium, did not stain with iodine, confirming previous reports that the A. niger cell wall does not contain long stretches of α-(1,4) linked glucan which might serve as the natural substrate for Agt enzymes (van der Kaaij, unpublished results). Alternatively, the Agt enzymes might act on the α-(1,4) bonds in nigeran, but this type of glucan is only

Chapter 6 Summary and concluding remarks
129
present in a limited number of fungal species, while Agt homologues occur widespread. In vitro activity of AgtA/B on nigeran could not be demonstrated, but a more straightforward way to exclude it as a substrate would be to offer well dissolvable nigerooligosaccharides to these enzymes and check for formation of elongated products. Possibly, Agt enzymes act on short α-(1,4) linked stretches in α-(1,3) glucan, as was also suggested for the Ags GH13 domain. Indeed, early studies report small percentages of α-(1,4) linkages in A. niger α-(1,3) glucan, which could represent α-(1,4) linked stretches similar to those identified in S. pombe α-glucan (Horisberger et al., 1972;Johnston, 1965). However, to perform a crosslinking reaction between α-(1,3) glucan chains with short α-(1,4) stretches would involve an α-(1,3) linked acceptor molecule, which is a poor acceptor substrate for both AgtA and AgtB (chapter 4). With the present knowledge on Aspergillus α-glucan, the exact in vivo reaction of Agt enzymes is uncertain, but this topic might be addressed in future studies by determining the composition of the cell walls of different Agt and Ags mutants. In contrast to most filamentous Ascomycetes, yeasts like Saccharomyces cerevisiae and Candida albicans do not produce cell wall α-glucan. Other components such as chitin and β-glucan are shared among both yeast-like and filamentous Ascomycetes, and the enzymes involved in e.g. β-glucan synthesis and remodelling are highly homologous between the species. Simultaneous expression of all enzymes involved in α-glucan synthesis in a yeast host might result in α-glucan production, thereby offering a system to study α-glucan synthesis in detail. Such an approach may provide definite proof for the involvement of the different types of enzymes. Additionally, it would offer an experimental model to study the in vivo effects of mutagenesis of specific amino acids in Agt, Ags and family GH13_5 enzymes. Considering the arguments mentioned above, we hereby propose a highly speculative model for the function of AgtA/B and AmyD/E in the A. niger cell wall (Fig. 2). It is already clear that this model does not provide the full picture, experimental evidence lacking for most points raised. Such a model, however, combines the present knowledge on the topic, and offers a starting-point for future experiments: In the cell, AmyD/E provides an α-(1,4) linked oligosaccharide primer molecule to the intracellular GT5 domain of Ags. Subsequently, UDP/ADP-glucose is used for elongation of the glucan chain. The growing chain, which might be of the α-(1,3) glucan or nigeran type, is transported over the membrane via the membrane spanning domain of Ags, and subsequently coupled to another α-glucan chain by the GH13 domain of Ags. Agt enzymes assist in the coupling of newly formed α-glucan chains to

Alpha-glucan acting enzymes in Aspergillus niger
130
existing chains, and allow for growth and branching of the mycelium by keeping the α-glucan component flexible. The stretch of α-(1,4) linkages, possibly originating from the oligosaccharide primer molecule supplied by AmyD/E, might be the main part of the α-glucan available for a coupling reaction. Coregulated agt and ags genes encode for proteins producing or acting on the same type of α-glucan.
Fig. 2 Hypothetical model of α-glucan synthesis in A. niger as described in this chapter. AmyD/E is located in the cell, where it may provide a primer molecule to an α-glucan synthase (Ags). Subsequently, the primer is elongated by the GT5 domain of the Ags enzyme using UDP- or ADP-glucose, and the α-glucan produced is transported out of the cell via the Ags transmembrane regions. In the cell wall, the α-glucan chains may be linked to each other or other cell wall components by the action of the GH13 domain of Ags enzymes or by GPI-anchored α-glucanotransferases.
The hypothetical model on α-glucan biosynthesis in fungal cell walls has some resemblance to the system for β-glucan production and maintenance. Fungal β-(1,3) glucan is produced by β-glucan synthases, members of family GT48 and related to plant cellulose synthases, which have been identified in several yeasts and filamentous fungi (Lesage and Bussey, 2006). Like α-glucan synthases, the enzymes comprise several domains, including two cytoplasmic domains and two separate domains with membrane spanning regions for localization. Although β-glucan formation has been studied more

Chapter 6 Summary and concluding remarks
131
extensively than α-glucan formation, the biochemistry of the synthesis process is still not known exactly. S. cerevisiae harbors two similar β-glucan synthases, FKS1 and FKS2, which use UDP-glucose to form a β-(1,3) glucan that is subsequently transported to the outside of the cell. A double knockout for both FKS genes is not viable (Mazur et al., 1995). Additional enzymes involved in β-glucan synthesis are GPI-anchored β-(1,3) glucanosyltransferases, called GAS proteins in S. cerevisiae, PHR in C. albicans and Gel proteins in A. fumigatus. Deletion strains for these enzymes affected virulence of C. albicans and A. fumigatus, the causative agent of aspergillosis (De Bernardis et al., 1998;Mouyna et al., 2005). The genes encoding the different enzymes involved in β-glucan synthesis are not clustered, as is the case for the ags, agt, and amyD/E genes.
The fungal cell wall is an important structure for the targeting of antifungal drugs; it forms an essential part of the cell, it is easily accessible and it is composed of fungus specific components not produced by mammalian hosts. A relatively novel type of antifungal drugs, the echinocandins, act as inhibitors of the enzymes producing cell wall β-(1,3) glucan and are used against a wide variety of human fungal pathogens (Morris and Villmann, 2006). In contrast to cell wall β-glucan, α-glucan has not been a major target for the development of antifungal drugs, mainly because it does not occur in C. albicans, the most frequently encountered fungal pathogen of humans. As for the aspergilli, it was suggested that α-glucan is not important for pathogenicity of A. fumigatus (Beauvais et al., 2005;Maubon et al., 2006). However, α-glucans are known to be important or even essential factors for pathogenicity in a number of other fungi, mainly dimorphic species such as H. capsulatum (Rappleye et al., 2004), Blastomyces dermatitidis (Hogan and Klein, 1994) and Paracoccidioides brasiliensis (Borges-Walmsley et al., 2002), but also in the pathogenic Basidiomycete C. neoformans (Reese et al., 2007). Although the molecular mechanism for the involvement of α-glucan in virulence is often unknown, it is clear that the α-glucan component of fungal cell walls deserves to be studied as a potential target for antifungal drugs.


133
Reference List
1. Altschul,S.F., Madden,T.L., Schaffer,A.A., Zhang,J., Zhang,Z., Miller,W., and Lipman,D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402.
2. Arguelles,J.C. (2000) Physiological roles of trehalose in bacteria and yeasts: a comparative analysis. Arch Microbiol 174: 217-224.
3. Ausubel,F.M., Brent,R.E., Kingston,D.D., Moore,J.G., Seidman,J.G., Smith,J.A., and Struhl,K. (1987) Current Protocols in molecular biology New York: John Wiley & Sons, Inc.
4. Bahia,M.C., Vieira,R.P., Mulloy,B., Hartmann,R., and Bergter,E.B. (1997) The structures of polysaccharides and glycolipids of Aspergillus fumigatus grown in the presence of human serum. Mycopathologia 137: 17-25.
5. Bardalaye,P.C., and Nordin,J.H. (1977) Chemical structure of the galactomannan from the cell wall of Aspergillus niger. J Biol Chem 252: 2584-2591.
6. Barker,S.A., and Carrington,T.R. (1953) Studies of Aspergillus niger. Part II. Transglycosylation by Aspergillus niger. J Chem Soc 1953: 3588-3593.
7. Bauer,S., Vasu,P., Persson,S., Mort,A.J., and Somerville,C.R. (2006) Development and application of a suite of polysaccharide-degrading enzymes for analyzing plant cell walls. Proc Natl Acad Sci U S A 103: 11417-11422.
8. Beauvais,A., and Latgé,J.P. (2001) Membrane and cell wall targets in Aspergillus fumigatus. Drug Resist Updat 4: 38-49.
9. Beauvais,A., Maubon,D., Park,S., Morelle,W., Tanguy,M., Huerre,M. et al. (2005) Two alpha(1-3) glucan synthases with different functions in Aspergillus fumigatus. Appl Environ Microbiol 71: 1531-1538.
10. Bendtsen,J.D., Kiemer,L., Fausboll,A., and Brunak,S. (2005) Non-classical protein secretion in bacteria. BMC Microbiol 5: 58.
11. Bendtsen,J.D., Nielsen,H., Von Heijne,G., and Brunak,S. (2004) Improved prediction of signal peptides: SignalP 3.0. J Mol Biol 340: 783-795.
12. Benen,J.A., Kester,H.C., and Visser,J. (1999) Kinetic characterization of Aspergillus niger N400 endopolygalacturonases I, II and C. Eur J Biochem 259: 577-585.
13. Bennett,J.W., and Lasure,L.L. (1991) More gene manipulations in fungi. San Diego: Academic Press, pp. 441-447.
14. Benson,D.A., Karsch-Mizrachi,I., Lipman,D.J., Ostell,J., and Wheeler,D.L. (2006) GenBank. Nucleic Acids Res 34: D16-D20.
15. Birney,E., Clamp,M., and Durbin,R. (2004) GeneWise and Genomewise. Genome Res 14: 988-995. 16. Blumenthal,H.J., and Roseman,S. (1957) Quantitative estimation of chitin in fungi. J Bacteriol 74: 222-
224. 17. Bobbitt,T.F., Nordin,J.H., Roux,M., Revol,J.F., and Marchessault,R.H. (1977) Distribution and
conformation of crystalline nigeran in hyphal walls of Aspergillus niger and Aspergillus awamori. J Bacteriol 132: 691-703.
18. Boel,E., Brady,L., Brzozowski,A.M., Derewenda,Z., Dodson,G.G., Jensen,V.J. et al. (1990) Calcium binding in alpha-amylases: an X-ray diffraction study at 2.1-A resolution of two enzymes from Aspergillus. Biochemistry 29: 6244-6249.
19. Boel,E., Hjort,I., Svensson,B., Norris,F., Norris,K.E., and Fiil,N.P. (1984) Glucoamylases G1 and G2 from Aspergillus niger are synthesized from two different but closely related mRNAs. EMBO J 3: 1097-1102.
20. Borges-Walmsley,M.I., Chen,D., Shu,X., and Walmsley,A.R. (2002) The pathobiology of Paracoccidioides brasiliensis. Trends Microbiol 10: 80-87.
21. Bos,C.J., Debets,A.J., Swart,K., Huybers,A., Kobus,G., and Slakhorst,S.M. (1988) Genetic analysis and the construction of master strains for assignment of genes to six linkage groups in Aspergillus niger. Curr Genet 14: 437-443.
22. Bose,S., Mazumder,S., and Mukherjee,M. (2007) Laccase production by the white-rot fungus Termitomyces clypeatus. J Basic Microbiol 47: 127-131.
23. Bowman,S.M., and Free,S.J. (2006) The structure and synthesis of the fungal cell wall. Bioessays 28: 799-808.

Alpha-glucan acting enzymes in Aspergillus niger
134
24. Bowman,S.M., Piwowar,A., Al,D.M., Vierula,J., and Free,S.J. (2006) Mutational analysis of the Glycosylphosphatidylinositol (GPI) anchor pathway demonstrates that GPI-anchored proteins are required for cell wall biogenesis and normal hyphal growth in Neurospora crassa. Eukaryot Cell 5: 587-600.
25. Brul,S., King,A., van der Vaart,J.M., Chapman,J., Klis,F., and Verrips,C.T. (1997) The incorporation of mannoproteins in the cell wall of S. cerevisiae and filamentous Ascomycetes. Antonie Van Leeuwenhoek 72: 229-237.
26. Brzozowski,A.M., Lawson,D.M., Turkenburg,J.P., Bisgaard-Frantzen,H., Svendsen,A., Borchert,T.V. et al. (2000) Structural analysis of a chimeric bacterial alpha-amylase. High-resolution analysis of native and ligand complexes. Biochemistry 39: 9099-9107.
27. Cabib,E., and Kang,M.S. (1987) Fungal 1,3-beta-glucan synthase. Methods Enzymol 138: 637-642. 28. Carlile,M.J., and Watkinson,S.C. (1994) The Fungi London: Academic Press Limited. 29. Caro,L.H., Tettelin,H., Vossen,J.H., Ram,A.F., Van Den,E.H., and Klis,F.M. (1997) In silicio
identification of glycosyl-phosphatidylinositol-anchored plasma-membrane and cell wall proteins of Saccharomyces cerevisiae. Yeast 13: 1477-1489.
30. Chang,C.T., Lo,H.F., Chi,M.C., Yao,C.Y., Hsu,W.H., and Lin,L.L. (2003) Identification of essential histidine residues in a recombinant alpha-amylase of thermophilic and alkaliphilic Bacillus sp. strain TS-23. Extremophiles 7: 505-509.
31. Costachel,C., Coddeville,B., Latge,J.P., and Fontaine,T. (2005) Glycosylphosphatidylinositol-anchored fungal polysaccharide in Aspergillus fumigatus. J Biol Chem 280: 39835-39842.
32. Coutinho,P.M., Deleury,E., Davies,G.J., and Henrissat,B. (2003) An evolving hierarchical family classification for glycosyltransferases. J Mol Biol 328: 307-317.
33. Coutinho,P.M., and Henrissat,B. (1999) Carbohydrate-active enzymes: an integrated database approach. In Recent Advances in Carbohydrate Bioengineering. Gilbert,H.J., Davies,G.J., Henrissat,B., and Svensson,B. (eds). Cambridge: The Royal Society of Chemistry, pp. 3-12.
34. Crooks,G.E., Hon,G., Chandonia,J.M., and Brenner,S.E. (2004) WebLogo: a sequence logo generator. Genome Res 14: 1188-90.
35. Da Lage,J.L., Feller,G., and Janecek,S. (2004) Horizontal gene transfer from Eukarya to bacteria and domain shuffling: the alpha-amylase model. Cell Mol Life Sci 61: 97-109.
36. Damveld,R.A., Arentshorst,M., Vankuyk,P.A., Klis,F.M., van den Hondel,C.A., and Ram,A.F. (2005a) Characterisation of CwpA, a putative glycosylphosphatidylinositol-anchored cell wall mannoprotein in the filamentous fungus Aspergillus niger. Fungal Genet Biol 42: 873-885.
37. Damveld,R.A., Vankuyk,P.A., Arentshorst,M., Klis,F.M., van den Hondel,C.A., and Ram,A.F. (2005b) Expression of agsA, one of five 1,3-alpha-d-glucan synthase-encoding genes in Aspergillus niger, is induced in response to cell wall stress. Fungal Genet Biol 42: 165-177.
38. Davies,G.J., Brzozowski,A.M., Dauter,Z., Rasmussen,M.D., Borchert,T.V., and Wilson,K.S. (2005) Structure of a Bacillus halmapalus family 13 alpha-amylase, BHA, in complex with an acarbose-derived nonasaccharide at 2.1 A resolution. Acta Crystallogr D Biol Crystallogr 61: 190-193.
39. Davies,G.J., Wilson,K.S., and Henrissat,B. (1997) Nomenclature for sugar-binding subsites in glycosyl hydrolases. Biochem J 321: 557-559.
40. De Bernardis,F., Muhlschlegel,F.A., Cassone,A., and Fonzi,W.A. (1998) The pH of the host niche controls gene expression in and virulence of Candida albicans. Infect Immun 66: 3317-3325.
41. De Groot,P.W., Hellingwerf,K.J., and Klis,F.M. (2003) Genome-wide identification of fungal GPI proteins. Yeast 20: 781-796.
42. De Groot,P.W., Ram,A.F., and Klis,F.M. (2005) Features and functions of covalently linked proteins in fungal cell walls. Fungal Genet Biol 42: 657-675.
43. De Groot,P.W., Yin,Q.Y., Weig,M., Sosinska,G.J., Klis,F.M., and de Koster,C.G. (2007) Mass spectrometric identification of covalently bound cell wall proteins from the fission yeast Schizosaccharomyces pombe. Yeast 24: 267-78.
44. de Ruiter-Jacobs,Y.M., Broekhuijsen,M., Unkles,S.E., Campbell,E.I., Kinghorn,J.R., Contreras,R. et al. (1989) A gene transfer system based on the homologous pyrG gene and efficient expression of bacterial genes in Aspergillus oryzae. Curr Genet 16: 159-163.
45. De Vries,R.P., and Visser,J. (2001) Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol Mol Biol Rev 65: 497-522.
46. den Herder,I.F., Rosell,A.M., van Zuilen,C.M., Punt,P.J., and van den Hondel,C.A. (1992) Cloning and expression of a member of the Aspergillus niger gene family encoding alpha-galactosidase. Mol Gen Genet 233: 404-410.

Reference list
135
47. Dey,G., Palit,S., Banerjee,R., and Maiti,B.R. (2002) Purification and characterization of maltooligosaccharide-forming amylase from Bacillus circulans GRS 313. J Ind Microbiol Biotechnol 28: 193-200.
48. Dowzer,C.E., and Kelly,J.M. (1991) Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol Cell Biol 11: 5701-5709.
49. Duarte,J.C., and Costa-Ferreira,M. (1994) Aspergilli and lignocellulosics: enzymology and biotechnological applications. FEMS Microbiol Rev 13: 377-386.
50. Durbin,R., and Eddy,S. (1998) Biological Sequence analysis: Probabilistic Models of Proteins and Nucleic acids. A tutorial introduction to hidden Markov models and other probabilistic modelling approaches in computational sequence analysis. Cambridge: Cambridge University Press.
51. Eisenhaber,B., Schneider,G., Wildpaner,M., and Eisenhaber,F. (2004) A sensitive predictor for potential GPI lipid modification sites in fungal protein sequences and its application to genome-wide studies for Aspergillus nidulans, Candida albicans, Neurospora crassa, Saccharomyces cerevisiae and Schizosaccharomyces pombe. J Mol Biol 337: 243-253.
52. Fontaine,T., Magnin,T., Melhert,A., Lamont,D., Latgé,J.P., and Ferguson,M.A. (2003) Structures of the glycosylphosphatidylinositol membrane anchors from Aspergillus fumigatus membrane proteins. Glycobiology 13: 169-177.
53. Fontaine,T., Simenel,C., Dubreucq,G., Adam,O., Delepierre,M., Lemoine,J. et al. (2000) Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall. J Biol Chem 275: 41528.
54. Fowler,T., Berka,R.M., and Ward,M. (1990) Regulation of the glaA gene of Aspergillus niger. Curr Genet 18: 537-545.
55. Francois,J., and Parrou,J.L. (2001) Reserve carbohydrates metabolism in the yeast Saccharomyces cerevisiae. FEMS Microbiol Rev 25: 125-145.
56. Frieman,M.B., and Cormack,B.P. (2003) The omega-site sequence of glycosylphosphatidylinositol-anchored proteins in Saccharomyces cerevisiae can determine distribution between the membrane and the cell wall. Mol Microbiol 50: 883-896.
57. Frieman,M.B., and Cormack,B.P. (2004) Multiple sequence signals determine the distribution of glycosylphosphatidylinositol proteins between the plasma membrane and cell wall in Saccharomyces cerevisiae. Microbiology 150: 3105-3114.
58. Galagan,J.E., Calvo,S.E., Cuomo,C., Ma,L.J., Wortman,J.R., Batzoglou,S. et al. (2005) Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 438: 1105-1115.
59. Garcia,I., Tajadura,V., Martin,V., Toda,T., and Sanchez,Y. (2006) Synthesis of alpha-glucans in fission yeast spores is carried out by three alpha-glucan synthase paralogues, Mok12p, Mok13p and Mok14p. Mol Microbiol 59: 836-853.
60. Geber,A., Williamson,P.R., Rex,J.H., Sweeney,E.C., and Bennett,J.E. (1992) Cloning and characterization of a Candida albicans maltase gene involved in sucrose utilization. J Bacteriol 174: 6992-6996.
61. Geysens,S., Pakula,T., Uusitalo,J., Dewerte,I., Penttila,M., and Contreras,R. (2005) Cloning and characterization of the glucosidase II alpha subunit gene of Trichoderma reesei: a frameshift mutation results in the aberrant glycosylation profile of the hypercellulolytic strain Rut-C30. Appl Environ Microbiol 71: 2910-2924.
62. Gomi,K., Akeno,T., Minetoki,T., Ozeki,K., Kumagai,C., Okazaki,N., and Iimura,Y. (2000) Molecular cloning and characterization of a transcriptional activator gene, amyR, involved in the amylolytic gene expression in Aspergillus oryzae. Biosci Biotechnol Biochem 64: 816-827.
63. Goosen,C., Yuan,X.L., van Munster,J.M., Ram,A.F., van der Maarel,M.J., and Dijkhuizen,L. (2007) Molecular and biochemical characterization of a novel intracellular invertase from Aspergillus niger with transfructosylating activity. Eukaryot Cell 6: 674-681.
64. Grün,C.H., Hochstenbach,F., Humbel,B.M., Verkleij,A.J., Sietsma,J.H., Klis,F.M. et al. (2005) The structure of cell wall alpha-glucan from fission yeast. Glycobiology 15: 245-257.
65. Guzman-Maldonado,H., and Paredes-Lopez,O. (1995) Amylolytic enzymes and products derived from starch: a review. Crit Rev Food Sci Nutr 35: 373-403.
66. Hartland,R.P., Emerson,G.W., and Sullivan,P.A. (1991) A secreted beta-glucan-branching enzyme from Candida albicans. Proc Biol Sci 246: 155-160.
67. Hemker,M., Stratmann,A., Goeke,K., Schroder,W., Lenz,J., Piepersberg,W., and Pape,H. (2001) Identification, cloning, expression, and characterization of the extracellular acarbose-modifying glycosyltransferase, AcbD, from Actinoplanes sp. strain SE50. J Bacteriol 183: 4484-4492.

Alpha-glucan acting enzymes in Aspergillus niger
136
68. Henrissat,B. (1991) A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem J 280: 309-316.
69. Henrissat,B., and Bairoch,A. (1993) New families in the classification of glycosyl hydrolases based on amino acid sequence similarities. Biochemistry Journal 293: 781-788.
70. Henrissat,B., and Bairoch,A. (1996) Updating the sequence-based classification of glycosyl hydrolases. Biochem J 316: 695-696.
71. Hesseltine,C.W. (1983) Microbiology of oriental fermented foods. Annu Rev Microbiol 37: 575-601. 72. Hochstenbach,F., Klis,F.M., Van Den,E.H., van,D.E., Peters,P.J., and Klausner,R.D. (1998)
Identification of a putative alpha-glucan synthase essential for cell wall construction and morphogenesis in fission yeast. Proc Natl Acad Sci U S A 95: 9161-9166.
73. Hogan,L.H., and Klein,B.S. (1994) Altered expression of surface alpha-1,3-glucan in genetically related strains of Blastomyces dermatitidis that differ in virulence. Infect Immun 62: 3543-3546.
74. Honegger,R. (2001) The symbiotic phenotype of lichen-forming Ascomycetes. In Fungal Associations. Hock,B. (ed). Berlin: Springer-Verlag, pp. 165-188.
75. Horisberger,M., Lewis,B.A., and Smith,F. (1972) Structure of a (1, 3)-α-D-glucan (pseudonigeran) of Aspergillus niger NNRL 326 cell wall. Carbohydr Res 23: 183-188.
76. Hoshiko,S., Makabe,O., Nojiri,C., Katsumata,K., Satoh,E., and Nagaoka,K. (1987) Molecular cloning and characterization of the Streptomyces hygroscopicus alpha-amylase gene. J Bacteriol 169: 1029-1036.
77. Hull,E.P., Green,P.M., Arst,H.N., Jr., and Scazzocchio,C. (1989) Cloning and physical characterization of the L-proline catabolism gene cluster of Aspergillus nidulans. Mol Microbiol 3: 553-559.
78. Iefuji,H., Chino,M., Kato,M., and Iimura,Y. (1996) Raw-starch-digesting and thermostable alpha-amylase from the yeast Cryptococcus sp. S-2: purification, characterization, cloning and sequencing. Biochem J 318: 989-996.
79. Inoue,H., Nojima,H., and Okayama,H. (1990) High efficiency transformation of Escherichia coli with plasmids. Gene 96: 23-28.
80. Ito,T., Tani,S., Itoh,T., Tsukagoshi,N., Kato,M., and Kobayashi,T. (2004) Mode of AmyR binding to the CGGN8AGG sequence in the Aspergillus oryzae taaG2 promoter. Biosci Biotechnol Biochem 68: 1906-1911.
81. Iwaki,T., Morita,T., Tanaka,N., Giga-Hama,Y., and Takegawa,K. (2007) Loss of a GPI-anchored membrane protein Aah3p causes a defect in vacuolar protein sorting in Schizosaccharomyces pombe. Biosci Biotechnol Biochem 71: 623-626.
82. James,T.Y., Kauff,F., Schoch,C.L., Matheny,P.B., Hofstetter,V., Cox,C.J. et al. (2006) Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 443: 818-822.
83. Janecek,S. (1994) Sequence similarities and evolutionary relationships of microbial, plant and animal alpha-amylases. Eur J Biochem 224: 519-524.
84. Janecek,S. (1997) Alpha-Amylase family: molecular biology and evolution. Prog Biophys Mol Biol 67: 67-97.
85. Janecek,S. (2002) How many conserved sequence regions are there in the α-amylase family? Biologia 57 (Suppl. 11): 29-41.
86. Janecek,S., Leveque,E., Belarbi,A., and Haye,B. (1999) Close evolutionary relatedness of alpha-amylases from Archaea and plants. J Mol Evol 48: 421-426.
87. Janecek,S., Svensson,B., and Henrissat,B. (1997) Domain evolution in the alpha-amylase family. J Mol Evol 45: 322-331.
88. Jeang,C.L., Chen,L.S., Chen,M.Y., and Shiau,R.J. (2002) Cloning of a gene encoding raw-starch-digesting amylase from a Cytophaga sp. and its expression in Escherichia coli. Appl Environ Microbiol 68: 3651-3654.
89. Jeanmougin,F., Thompson,J.D., Gouy,M., Higgins,D.G., and Gibson,T.J. (1998) Multiple sequence alignment with Clustal X. Trends Biochem Sci 23: 403-405.
90. Jespersen,H.M., MacGregor,E.A., Sierks,M.R., and Svensson,B. (1991) Comparison of the domain-level organization of starch hydrolases and related enzymes. Biochem J 280: 51-55.
91. Johnston,I.R. (1965) The composition of the cell wall of Aspergillus niger. Biochem J 96: 651-658. 92. Juge,N., Rodenburg,K.W., Guo,X.J., Chaix,J.C., and Svensson,B. (1995) Isozyme hybrids within the
protruding third loop domain of the barley alpha-amylase (beta/alpha)8-barrel. Implication for BASI sensitivity and substrate affinity. FEBS Lett 363: 299-303.
93. Kanai,R., Haga,K., Akiba,T., Yamane,K., and Harata,K. (2004) Biochemical and crystallographic analyses of maltohexaose-producing amylase from alkalophilic Bacillus sp. 707. Biochemistry 43: 14047-14056.

Reference list
137
94. Kang,H.K., Lee,J.H., Kim,D., Day,D.F., Robyt,J.F., Park,K.H., and Moon,T.W. (2004) Cloning and expression of Lipomyces starkeyi alpha-amylase in Escherichia coli and determination of some of its properties. FEMS Microbiol Lett 233: 53-64.
95. Kaper,T., Leemhuis,H., Uitdehaag,J.C., Veen,B.A., Dijkstra,B.W., Maarel,M.J., and Dijkhuizen,L. (2007) Identification of acceptor substrate binding subsites +2 and +3 in the amylomaltase from Thermus thermophilus HB8. Biochemistry 46: 5261-5269.
96. Kaper,T., Talik,B., Ettema,T.J., Bos,H., van der Maarel,M.J., and Dijkhuizen,L. (2005) Amylomaltase of Pyrobaculum aerophilum IM2 produces thermoreversible starch gels. Appl Environ Microbiol 71: 5098-5106.
97. Kaper,T., van der Maarel,M.J., Euverink,G.J., and Dijkhuizen,L. (2004) Exploring and exploiting starch-modifying amylomaltases from thermophiles. Biochem Soc Trans 32: 279-282.
98. Kapteyn,J.C., Van Egmond,P., Sievi,E., Van Den Ende,H., Makarow,M., and Klis,F.M. (1999) The contribution of the O-glycosylated protein Pir2p/Hsp150 to the construction of the yeast cell wall in wild-type cells and beta 1,6-glucan-deficient mutants. Mol Microbiol 31: 1835-1844.
99. Kato,N., Murakoshi,Y., Kato,M., Kobayashi,T., and Tsukagoshi,N. (2002a) Isomaltose formed by alpha-glucosidases triggers amylase induction in Aspergillus nidulans. Curr Genet 42: 43-50.
100. Kato,N., Suyama,S., Shirokane,M., Kato,M., Kobayashi,T., and Tsukagoshi,N. (2002b) Novel alpha-glucosidase from Aspergillus nidulans with strong transglycosylation activity. Appl Environ Microbiol 68: 1250-1256.
101. Keller,N.P., and Hohn,T.M. (1997) Metabolic pathway gene clusters in filamentous fungi. Fungal Genet Biol 21: 17-29.
102. Kelly,J.M., and Hynes,M.J. (1985) Transformation of Aspergillus niger by the amdS gene of Aspergillus nidulans. EMBO J 4: 475-479.
103. Khoo,S.L., Amirul,A.A., Kamaruzaman,M., Nazalan,N., and Azizan,M.N. (1994) Purification and characterization of alpha-amylase from Aspergillus flavus. Folia Microbiol (Praha) 39: 392-398.
104. Kim,M.J., Lee,S.B., Lee,H.S., Lee,S.Y., Baek,J.S., Kim,D. et al. (1999) Comparative study of the inhibition of alpha-glucosidase, alpha-amylase, and cyclomaltodextrin glucanosyltransferase by acarbose, isoacarbose, and acarviosine-glucose. Arch Biochem Biophys 371: 277-283.
105. Kim,T.J., Park,C.S., Cho,H.Y., Cha,S.S., Kim,J.S., Lee,S.B. et al. (2000) Role of the glutamate 332 residue in the transglycosylation activity of Thermus maltogenic amylase. Biochemistry 39: 6773-6780.
106. Kirimura,K., Yusa,S., Rugsaseel,S., Nakagawa,H., Osumi,M., and Usami,S. (1999) Amylose-like polysaccharide accumulation and hyphal cell-surface structure in relation to citric acid production by Aspergillus niger in shake culture. Appl Microbiol Biotechnol 52: 421-428.
107. Kitamoto,H.K., Ohmomo,S., and Iimura,Y. (1998) Isolation and nucleotide sequence of the gene encoding phosphoenolpyruvate carboxykinase from Kluyveromyces lactis. Yeast 14: 963-967.
108. Klein,C., Hollender,J., Bender,H., and Schulz,G.E. (1992) Catalytic center of cyclodextrin glycosyltransferase derived from X-ray structure analysis combined with site-directed mutagenesis. Biochemistry 31: 8740-8746.
109. Klis,F.M., de Groot,P., and Hellingwerf,K. (2001) Molecular organization of the cell wall of Candida albicans. Med Mycol 39 Suppl 1: 1-8.
110. Klis,F.M., Mol,P., Hellingwerf,K., and Brul,S. (2002) Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol Rev 26: 239-256.
111. Knegtel,R.M., Strokopytov,B., Penninga,D., Faber,O.G., Rozeboom,H.J., Kalk,K.H. et al. (1995) Crystallographic studies of the interaction of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 with natural substrates and products. J Biol Chem 270: 29256-29264.
112. Kolar,M., Punt,P.J., van den Hondel,C.A., and Schwab,H. (1988) Transformation of Penicillium chrysogenum using dominant selection markers and expression of an Escherichia coli lacZ fusion gene. Gene 62: 127-134.
113. Korman,D.R., Bayliss,F.T., Barnett,C.C., Carmona,C.L., Kodama,K.H., Royer,T.J. et al. (1990) Cloning, characterization, and expression of two alpha-amylase genes from Aspergillus niger var. awamori. Curr Genet 17: 203-212.
114. Kralj,S., van Geel-Schutten,G.H., Dondorff,M.M., Kirsanovs,S., van der Maarel,M.J., and Dijkhuizen,L. (2004a) Glucan synthesis in the genus Lactobacillus: isolation and characterization of glucansucrase genes, enzymes and glucan products from six different strains. Microbiology 150: 3681-3690.
115. Kralj,S., van Geel-Schutten,G.H., van der Maarel,M.J., and Dijkhuizen,L. (2004b) Biochemical and molecular characterization of Lactobacillus reuteri 121 reuteransucrase. Microbiology 150: 2099-2112.

Alpha-glucan acting enzymes in Aspergillus niger
138
116. Kumar,S., Tamura,K., and Nei,M. (1993) MEGA: Molecular Evolutionary Analysis, version 1.01. The Pennsylvania State University.
117. Kupfer,D.M., Drabenstot,S.D., Buchanan,K.L., Lai,H., Zhu,H., Dyer,D.W. et al. (2004) Introns and splicing elements of five diverse fungi. Eukaryot Cell 3: 1088-1100.
118. Kuriki,T., and Imanaka,T. (1999) The concept of the alpha-amylase family: structural similarity and common catalytic mechanism. J Biosci Bioeng 87: 557-565.
119. Laemmli,U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
120. Lasure,L.L., and Magnuson,J.K. (2004) Organic acid production by filamentous fungi. In Advances in Fungal Biotechnology for Industry, Agriculture and Medicine. Tkacz,J.S., and Lange,L. (eds). New York: Kluwer Academic & Plenum Publishers, pp. 307-340.
121. Lawson,C.L., van Montfort,R., Strokopytov,B., Rozeboom,H.J., Kalk,K.H., de Vries,G.E. et al. (1994) Nucleotide sequence and X-ray structure of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 in a maltose-dependent crystal form. J Mol Biol 236: 590-600.
122. Le Gal-Coeffet,M.F., Jacks,A.J., Sorimachi,K., Williamson,M.P., Williamson,G., and Archer,D.B. (1995) Expression in Aspergillus niger of the starch-binding domain of glucoamylase. Comparison with the proteolytically produced starch-binding domain. Eur J Biochem 233: 561-567.
123. Leandro,M.J., Goncalves,P., and Spencer-Martins,I. (2006) Two glucose/xylose transporter genes from the yeast Candida intermedia: first molecular characterization of a yeast xylose-H+ symporter. Biochem J 395: 543-549.
124. Leemhuis,H., Dijkstra,B.W., and Dijkhuizen,L. (2003) Thermoanaerobacterium thermosulfurigenes cyclodextrin glycosyltransferase. Eur J Biochem 270: 155-162.
125. Leemhuis,H., Uitdehaag,J.C., Rozeboom,H.J., Dijkstra,B.W., and Dijkhuizen,L. (2002) The remote substrate binding subsite -6 in cyclodextrin-glycosyltransferase controls the transferase activity of the enzyme via an induced-fit mechanism. J Biol Chem 277: 1113-1119.
126. Leemhuis,H., Wehmeier,U.F., and Dijkhuizen,L. (2004) Single amino acid mutations interchange the reaction specificities of Cyclodextrin Glycosyltransferase and the acarbose-modifying enzyme Acarviosyl Transferase. Biochemistry 43: 13204-13213.
127. Lesage,G., and Bussey,H. (2006) Cell wall assembly in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 70: 317-343.
128. Liebl,W., Feil,R., Gabelsberger,J., Kellermann,J., and Schleifer,K.H. (1992) Purification and characterization of a novel thermostable 4-alpha-glucanotransferase of Thermotoga maritima cloned in Escherichia coli. Eur J Biochem 207: 81-88.
129. Loftus,B.J., Fung,E., Roncaglia,P., Rowley,D., Amedeo,P., Bruno,D. et al. (2005) The genome of the basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science 307: 1321-1324.
130. Lomako,J., Lomako,W.M., and Whelan,W.J. (2004) Glycogenin: the primer for mammalian and yeast glycogen synthesis. Biochim Biophys Acta 1673: 45-55.
131. MacGregor,E.A., Janecek,S., and Svensson,B. (2001) Relationship of sequence and structure to specificity in the alpha-amylase family of enzymes. Biochim Biophys Acta 1546: 1-20.
132. MacGregor,E.A., Jespersen,H.M., and Svensson,B. (1996) A circularly permuted alpha-amylase-type alpha/beta-barrel structure in glucan-synthesizing glucosyltransferases. FEBS Lett 378: 263-266.
133. Machida,M., Asai,K., Sano,M., Tanaka,T., Kumagai,T., Terai,G. et al. (2005) Genome sequencing and analysis of Aspergillus oryzae. Nature 438: 1157-1161.
134. Machius,M., Declerck,N., Huber,R., and Wiegand,G. (1998) Activation of Bacillus licheniformis alpha-amylase through a disorder-->order transition of the substrate-binding site mediated by a calcium-sodium-calcium metal triad. Structure 6: 281-292.
135. Machovic,M., and Janecek,S. (2003) The invariant residues in the alpha-amylase family: just the catalytic triad. Biologia 58: 1127-1132.
136. Marchler-Bauer,A., and Bryant,S.H. (2004) CD-Search: protein domain annotations on the fly. Nucleic Acids Res 32: W327-W331.
137. Marion,C.L., Rappleye,C.A., Engle,J.T., and Goldman,W.E. (2006) An alpha-(1,4)-amylase is essential for alpha-(1,3)-glucan production and virulence in Histoplasma capsulatum. Mol Microbiol 62: 970-983.
138. Martens-Uzunova,E.S., Zandleven,J.S., Benen,J.A., Awad,H., Kools,H.J., Beldman,G. et al. (2006) A new group of exo-acting family 28 glycoside hydrolases of Aspergillus niger that are involved in pectin degradation. Biochem J 400: 43-52.
139. Matsuura,Y., Kusunoki,M., Harada,W., and Kakudo,M. (1984) Structure and possible catalytic residues of Taka-amylase A. J Biochem (Tokyo) 95: 697-702.

Reference list
139
140. Mattey,M., and Allan,A. (1990) Glycogen accumulation in Aspergillus niger. Biochem Soc Trans 18: 1020-1021.
141. Mattsson,P., Battchikova,N., Sippola,K., and Korpela,T. (1995) The role of histidine residues in the catalytic act of cyclomaltodextrin glucanotransferase from Bacillus circulans var. alkalophilus. Biochim Biophys Acta 1247: 97-103.
142. Maubon,D., Park,S., Tanguy,M., Huerre,M., Schmitt,C., Prevost,M.C. et al. (2006) AGS3, an alpha(1-3)glucan synthase gene family member of Aspergillus fumigatus, modulates mycelium growth in the lung of experimentally infected mice. Fungal Genet Biol 43: 366-375.
143. Mazur,P., Morin,N., Baginsky,W., el Sherbeini,M., Clemas,J.A., Nielsen,J.B., and Foor,F. (1995) Differential expression and function of two homologous subunits of yeast 1,3-beta-D-glucan synthase. Mol Cell Biol 15: 5671-5681.
144. McCarter,J.D., and Withers,S.G. (1994) Mechanisms of enzymatic glycoside hydrolysis. Curr Opin Struct Biol 4: 885-892.
145. Meeuwsen,P.J., Vincken,J.P., Beldman,G., and Voragen,A.G. (2000) A universal assay for screening expression libraries for carbohydrases. J Biosci Bioeng 89: 107-109.
146. Meyer,V., Arentshorst,M., El Ghezal,A., Drews,A.C., Kooistra,R., van den Hondel,C.A., and Ram,A.F. (2007) Highly efficient gene targeting in the Aspergillus niger kusA mutant. J Biotechnol 128: 770-775.
147. Micheli,P.A. (1729) Nova plantarum genera juxta Turnefortii methodum disposita. Florence. 148. Monchois,V., Willemot,R.M., and Monsan,P. (1999) Glucansucrases: mechanism of action and
structure-function relationships. FEMS Microbiol Rev 23: 131-151. 149. Moreira,F.G., Lenartovicz,V., and Peralta,R.M. (2004) A thermostable maltose-tolerant alpha-amylase
from Aspergillus tamarii. J Basic Microbiol 44: 29-35. 150. Morita,T., Tanaka,N., Hosomi,A., Giga-Hama,Y., and Takegawa,K. (2006) An alpha-amylase
homologue, aah3, encodes a GPI-anchored membrane protein required for cell wall integrity and morphogenesis in Schizosaccharomyces pombe. Biosci Biotechnol Biochem 70: 1454-1463.
151. Morris,M.I., and Villmann,M. (2006) Echinocandins in the management of invasive fungal infections, Part 2. Am J Health Syst Pharm 63: 1813-1820.
152. Mosi,R., He,S., Uitdehaag,J., Dijkstra,B.W., and Withers,S.G. (1997) Trapping and characterization of the reaction intermediate in cyclodextrin glycosyltransferase by use of activated substrates and a mutant enzyme. Biochemistry 36: 9927-9934.
153. Mourya,S., and Jauhri,K.S. (2000) Production of citric acid from starch-hydrolysate by Aspergillus niger. Microbiol Res 155: 37-44.
154. Mouyna,I., Fontaine,T., Vai,M., Monod,M., Fonzi,W.A., Diaquin,M. et al. (2000) Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J Biol Chem 19;275: 14882-14889.
155. Mouyna,I., Morelle,W., Vai,M., Monod,M., Lechenne,B., Fontaine,T. et al. (2005) Deletion of GEL2 encoding for a beta(1-3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus. Mol Microbiol 56: 1675-1688.
156. Nakajima,T., Imanaka,T., and Aiba,S. (1986) Comparison of amino acid sequences of eleven different alpha-amylases. Appl Microbiol Biotechnol 23: 355-360.
157. Nakamura,A., Haga,K., and Yamane,K. (1993) Three histidine residues in the active center of cyclodextrin glucanotransferase from alkalophilic Bacillus sp. 1011: effects of the replacement on pH dependence and transition-state stabilization. Biochemistry 32: 6624-6631.
158. Nakamura,A., Haga,K., and Yamane,K. (1994) The transglycosylation reaction of cyclodextrin glucanotransferase is operated by a Ping-Pong mechanism. FEBS Lett 337: 66-70.
159. Nakamura,A., Nishimura,I., Yokoyama,A., Lee,D.G., Hidaka,M., Masaki,H. et al. (1997) Cloning and sequencing of an alpha-glucosidase gene from Aspergillus niger and its expression in A. nidulans. J Biotechnol 53: 75-84.
160. Nakamura,T., Maeda,Y., Tanoue,N., Makita,T., Kato,M., and Kobayashi,T. (2006) Expression profile of amylolytic genes in Aspergillus nidulans. Biosci Biotechnol Biochem 70: 2363-2370.
161. Nakao,M., Nakayama,T., Kakudo,A., Inohara,M., Harada,M., Omura,F., and Shibano,Y. (1994) Structure and expression of a gene coding for thermostable alpha-glucosidase with a broad substrate specificity from Bacillus sp. SAM1606. Eur J Biochem 220: 293-300.
162. Nielsen,A.D., Fuglsang,C.C., and Westh,P. (2003) Effect of calcium ions on the irreversible denaturation of a recombinant Bacillus halmapalus alpha-amylase: a calorimetric investigation. Biochem J 373: 337-343.

Alpha-glucan acting enzymes in Aspergillus niger
140
163. Nierman,W.C., Pain,A., Anderson,M.J., Wortman,J.R., Kim,H.S., Arroyo,J. et al. (2005) Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 438: 1151-1156.
164. Orlean,P. (1997) Biogenesis of yeast wall and surface components. In Cell Cycle and Cell Biology. Pringle,J., Broach,J.R., and Jones,E.W. (eds). New York: Cold Spring Harbor Laboratory Press, pp. 229-362.
165. Page,R.D.M. (1996) TREEVIEW: An application to display phylogenetic trees on personal computers. Computer Applications in the Biosciences 12: 357-358.
166. Paldi,T., Levy,I., and Shoseyov,O. (2003) Glucoamylase starch-binding domain of Aspergillus niger B1: molecular cloning and functional characterization. Biochem J 372: 905-910.
167. Pan,T., and Coleman,J.E. (1990) GAL4 transcription factor is not a "zinc finger" but forms a Zn(II)2Cys6 binuclear cluster. Proc Natl Acad Sci U S A 87: 2077-2081.
168. Parenicova,L., Benen,J.A., Kester,H.C., and Visser,J. (1998) pgaE encodes a fourth member of the endopolygalacturonase gene family from Aspergillus niger. Eur J Biochem 251: 72-80.
169. Pel,H.J., de Winde,J.H., Archer,D.B., Dyer,P.S., Hofmann,G., Schaap,P.J. et al. (2007) Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat Biotechnol 25: 221-231.
170. Penninga,D., van der Veen,B.A., Knegtel,R.M., van Hijum,S.A., Rozeboom,H.J., Kalk,K.H. et al. (1996) The raw starch binding domain of cyclodextrin glycosyltransferase from Bacillus circulans strain 251. J Biol Chem 271: 32777-32784.
171. Peters,D. (2006) Carbohydrates for fermentation. Biotechnol J 1: 806-814. 172. Petersen,K.L., Lehmbeck,J., and Christensen,T. (1999) A new transcriptional activator for amylase genes
in Aspergillus. Mol Gen Genet 262: 668-676. 173. Popolo,L., Vai,M., Gatti,E., Porello,S., Bonfante,P., Balestrini,R., and Alberghina,L. (1993)
Physiological analysis of mutants indicates involvement of the Saccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesis and cell separation. J Bacteriol 175: 1879-1885.
174. Preiss,J. (2004) Plant starch synthesis. In Starch in food. Eliasson,A.-C. (ed). Cambridge: Woodhead Publishing Limited, pp. 3-56.
175. Pugsley,A.P., and Dubreuil,C. (1988) Molecular characterization of malQ, the structural gene for the Escherichia coli enzyme amylomaltase. Mol Microbiol 2: 473-479.
176. Punt,P.J., and van den Hondel,C.A. (1992) Transformation of filamentous fungi based on hygromycin B and phleomycin resistance markers. Methods Enzymol 216: 447-457.
177. Ragni,E., Fontaine,T., Gissi,C., Latge,J.P., and Popolo,L. (2007) The Gas family of proteins of Saccharomyces cerevisiae: characterization and evolutionary analysis. Yeast 24: 297-308.
178. Ram,A.F., and Klis,F.M. (2006) Identification of fungal cell wall mutants using susceptibility assays based on Calcofluor white and Congo red. Nat Protoc 1: 2253-2256.
179. Rappleye,C.A., Engle,J.T., and Goldman,W.E. (2004) RNA interference in Histoplasma capsulatum demonstrates a role for alpha-(1,3)-glucan in virulence. Mol Microbiol 53: 153-165.
180. Rappleye,C.A., and Goldman,W.E. (2006) Defining virulence genes in the dimorphic fungi. Annu Rev Microbiol 60: 281-303.
181. Reese,A.J., and Doering,T.L. (2003) Cell wall alpha-1,3-glucan is required to anchor the Cryptococcus neoformans capsule. Mol Microbiol 50: 1401-1409.
182. Reese,A.J., Yoneda,A., Breger,J.A., Beauvais,A., Liu,H., Griffith,C.L. et al. (2007) Loss of cell wall alpha(1-3) glucan affects Cryptococcus neoformans from ultrastructure to virulence. Mol Microbiol 63: 1385-1398.
183. Robyt,J.F. (1998) Essentials of Carbohydrate Chemistry New York: Springer-Verlag New York Inc. 184. Rodenburg,K.W., Juge,N., Guo,X.J., Sogaard,M., Chaix,J.C., and Svensson,B. (1994) Domain B
protruding at the third beta strand of the alpha/beta barrel in barley alpha-amylase confers distinct isozyme-specific properties. Eur J Biochem 221: 277-284.
185. Ruepp,A., Zollner,A., Maier,D., Albermann,K., Hani,J., Mokrejs,M. et al. (2004) The FunCat, a functional annotation scheme for systematic classification of proteins from whole genomes. Nucleic Acids Res 32: 5539-5545.
186. Saitou,N., and Nei,M. (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425.
187. Sambrook,J., Fritsch,E.F., and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual Cold spring Harbor, NY: Cold Spring Harbor Laboratory Press.
188. Sasangka,P., Matsuno,A., Tanaka,A., Akasaka,Y., Suyama,S., Kano,S. et al. (2002) Structural features of the glycogen branching enzyme encoding genes from Aspergilli. Microbiol Res 157: 337-344.

Reference list
141
189. Sauer,J., Sigurskjold,B.W., Christensen,U., Frandsen,T.P., Mirgorodskaya,E., Harrison,M. et al. (2000) Glucoamylase: structure/function relationships, and protein engineering. Biochim Biophys Acta 1543: 275-293.
190. Schauer,F., and Boriss,R. (2004) Biocatalysis and biotransformation. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine. Tkacz,J.S., and Lange,L. (eds). New York: Kluwer Academic & Plenum Publishers, pp. 237-306.
191. Schuster,E., Dunn-Coleman,N., Frisvad,J.C., and van Dijck,P.W. (2002) On the safety of Aspergillus niger -a review. Appl Microbiol Biotechnol 59: 426-435.
192. Silva,A., Bacci,M., Jr., Gomes de Siqueira,C., Correa Bueno,O., Pagnocca,F.C., and Aparecida Hebling,M.J. (2003) Survival of Atta sexdens workers on different food sources. J Insect Physiol 49: 307-313.
193. Smith,S.E., and Read,D.J. (1997) Mycorrhizal Symbiosis. London: Academic Press Ltd. 194. Sogaard,M., Kadziola,A., Haser,R., and Svensson,B. (1993) Site-directed mutagenesis of histidine 93,
aspartic acid 180, glutamic acid 205, histidine 290, and aspartic acid 291 at the active site and tryptophan 279 at the raw starch binding site in barley alpha-amylase 1. J Biol Chem 268: 22480-22484.
195. Southall,S.M., Simpson,P.J., Gilbert,H.J., Williamson,G., and Williamson,M.P. (1999) The starch-binding domain from glucoamylase disrupts the structure of starch. FEBS Letters 447: 58-60.
196. Stagg,C.M., and Feather,M.S. (1973) The characterization of a chitin-associated D-glucan from the cell walls of Aspergillus niger. Biochim Biophys Acta 320: 64-72.
197. Stam,M.R., Danchin,E.G., Rancurel,C., Coutinho,P.M., and Henrissat,B. (2006) Dividing the large glycoside hydrolase family 13 into subfamilies: towards improved functional annotations of alpha-amylase-related proteins. Protein Eng Des Sel 19: 555-562.
198. Steyn,A.J., Marmur,J., and Pretorius,I.S. (1995) Cloning, sequence analysis and expression in yeasts of a cDNA containing a Lipomyces kononenkoae alpha-amylase-encoding gene. Gene 166: 65-71.
199. Svensson,B., Larsen,K., and Gunnarsson,A. (1986) Characterization of a glucoamylase G2 from Aspergillus niger. Eur J Biochem 154: 497-502.
200. Takaha,T., Yanase,M., Okada,S., and Smith,S.M. (1993) Disproportionating enzyme (4-alpha-glucanotransferase; EC 2.4.1.25) of potato. Purification, molecular cloning, and potential role in starch metabolism. J Biol Chem 268: 1391-1396.
201. Tani,S., Katsuyama,Y., Hayashi,T., Suzuki,H., Kato,M., Gomi,K. et al. (2001) Characterization of the amyR gene encoding a transcriptional activator for the amylase genes in Aspergillus nidulans. Curr Genet 39: 10-15.
202. Tao,B.Y., Reilly,P.J., and Robyt,J.F. (1989) Detection of a covalent intermediate in the mechanism of action of porcine pancreatic alpha-amylase by using 13C nuclear magnetic resonance. Biochim Biophys Acta 995: 214-220.
203. Terada,Y., Fujii,K., Takaha,T., and Okada,S. (1999) Thermus aquaticus ATCC 33923 amylomaltase gene cloning and expression and enzyme characterization: production of cycloamylose. Appl Environ Microbiol 65: 910-915.
204. Teste,M.A., Enjalbert,B., Parrou,J.L., and Francois,J.M. (2000) The Saccharomyces cerevisiae YPR184w gene encodes the glycogen debranching enzyme. FEMS Microbiol Lett 193: 105-110.
205. Thompson,J.D., Higgins,D.G., and Gibson,T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673-4680.
206. Toh-e A, Yasunaga,S., Nisogi,H., Tanaka,K., Oguchi,T., and Matsui,Y. (1993) Three yeast genes, PIR1, PIR2 and PIR3, containing internal tandem repeats, are related to each other, and PIR1 and PIR2 are required for tolerance to heat shock. Yeast 9: 481-494.
207. Tsukagoshi,N., Kobayashi,T., and Kato,M. (2001) Regulation of the amylolytic and (hemi-)cellulolytic genes in Aspergilli. J Gen Appl Microbiol 47: 1-19.
208. Tsukamoto,A., Kimura,K., Ishii,Y., Takano,T., and Yamane,K. (1988) Nucleotide sequence of the maltohexaose-producing amylase gene from an alkalophilic Bacillus sp. #707 and structural similarity to liquefying type alpha-amylases. Biochem Biophys Res Commun 151: 25-31.
209. Uitdehaag,J.C., Mosi,R., Kalk,K.H., van der Veen,B.A., Dijkhuizen,L., Withers,S.G., and Dijkstra,B.W. (1999) X-ray structures along the reaction pathway of cyclodextrin glycosyltransferase elucidate catalysis in the alpha-amylase family. Nat Struct Biol 6: 432-436.
210. van der Kaaij,R.M., Yuan,X.L., Franken,A., Ram,A.F.J., Punt,P.J., Maarel,M.J.E.C., and Dijkhuizen,L. (2007) Characterization of two novel, putatively cell wall associated and GPI-anchored, alpha-glucanotransferase enzymes of Aspergillus niger. Eukaryot Cell doi:10.1128.

Alpha-glucan acting enzymes in Aspergillus niger
142
211. van der Maarel,M.J., van,d., V, Uitdehaag,J.C., Leemhuis,H., and Dijkhuizen,L. (2002) Properties and applications of starch-converting enzymes of the alpha-amylase family. J Biotechnol 94: 137-155.
212. Van der Veen,B., Uitdehaag,J.C., Dijkstra,B.W., and Dijkhuizen,L. (2000) Engineering of cyclodextrin glycosyltransferase reaction and product specificity. Biochim Biophys Acta 1543: 336-360.
213. van Dijck,P.W., Selten,G.C., and Hempenius,R.A. (2003) On the safety of a new generation of DSM Aspergillus niger enzyme production strains. Regul Toxicol Pharmacol 38: 27-35.
214. van Hartingsveldt,W., Mattern,I.E., van Zeijl,C.M., Pouwels,P.H., and van den Hondel,C.A. (1987) Development of a homologous transformation system for Aspergillus niger based on the pyrG gene. Mol Gen Genet 206: 71-75.
215. van Hijum,S.A., Kralj,S., Ozimek,L.K., Dijkhuizen,L., and van Geel-Schutten,I.G. (2006) Structure-function relationships of glucansucrase and fructansucrase enzymes from lactic acid bacteria. Microbiol Mol Biol Rev 70: 157-176.
216. Vankuyk,P.A., Diderich,J.A., MacCabe,A.P., Hererro,O., Ruijter,G.J., and Visser,J. (2004) Aspergillus niger mstA encodes a high-affinity sugar/H+ symporter which is regulated in response to extracellular pH. Biochem J 379: 375-383.
217. Vos,A., Dekker,N., Distel,B., Leunissen,J.A., and Hochstenbach,F. (2007) Role of the SYN domain of Ags1p in cell wall alpha -glucan biosynthesis in fission yeast. J Biol Chem 282: 18969-79.
218. Vujicic-Zagar,A., and Dijkstra,B.W. (2006) Monoclinic crystal form of Aspergillus niger alpha-amylase in complex with maltose at 1.8 angstroms resolution. Acta Crystallograph Sect F Struct Biol Cryst Commun 62: 716-721.
219. Weenink,X.O., Punt,P.J., van den Hondel,C.A., and Ram,A.F. (2006) A new method for screening and isolation of hypersecretion mutants in Aspergillus niger. Appl Microbiol Biotechnol 69: 711-717.
220. Wernars,K., Goosen,T., Wennekes,L.M., Visser,J., Bos,C.J., van den Broek,H.W. et al. (1985) Gene amplification in Aspergillus nidulans by transformation with vectors containing the amdS gene. Curr Genet 9: 361-368.
221. Williamson,G., Belshaw,N.J., and Williamson,M.P. (1992) O-glycosylation in Aspergillus glucoamylase. Conformation and role in binding. Biochem J 282: 423-428.
222. Woloshuk,C.P., Yousibova,G.L., Rollins,J.A., Bhatnagar,D., and Payne,G.A. (1995) Molecular characterization of the afl-1 locus in Aspergillus flavus. Appl Environ Microbiol 61: 3019-3023.
223. Wolschek,M.F., and Kubicek,C.P. (1997) The filamentous fungus Aspergillus niger contains two "differentially regulated" trehalose-6-phosphate synthase-encoding genes, tpsA and tpsB. J Biol Chem 272: 2729-2735.
224. Woranovicz-Barreira,S.M., Gorin,P.A., Sassaki,P.L., Marcelli,M.P., and Iacomini,M. (1999) Galactomannoglucans of lichenized fungi of Cladonia spp.: significance as chemotypes. FEMS Microbiol Lett 181: 313-317.
225. Wu,C.H., Apweiler,R., Bairoch,A., Natale,D.A., Barker,W.C., Boeckmann,B. et al. (2006) The Universal Protein Resource (UniProt): an expanding universe of protein information. Nucleic Acids Res 34: D187-D191.
226. Yamazaki,H., Yamazaki,D., Takaya,N., Takagi,M., Ohta,A., and Horiuchi,H. (2007) A chitinase gene, chiB, involved in the autolytic process of Aspergillus nidulans. Curr Genet 51: 89-98.
227. Yoder,W.T., and Lehmbeck,J. (2004) Heterologous Expression and Protein Secretion. In Advances in Fungal Biotechnology for Industry, Agriculture and Medicine. Tkacz,J.S., and Lange,L. (eds). New York: Kluwer Academic & Plenum Publishers, pp. 201-219.
228. Yuan,X.L., Goosen,C., Kools,H., van der Maarel,M.J., van den Hondel,C.A., Dijkhuizen,L., and Ram,A.F. (2006) Database mining and transcriptional analysis of genes encoding inulin-modifying enzymes of Aspergillus niger. Microbiology 152: 3061-3073.
229. Yuuki,T., Nomura,T., Tezuka,H., Tsuboi,A., Yamagata,H., Tsukagoshi,N., and Udaka,S. (1985) Complete nucleotide sequence of a gene coding for heat- and pH-stable alpha-amylase of Bacillus licheniformis: comparison of the amino acid sequences of three bacterial liquefying alpha-amylases deduced from the DNA sequences. J Biochem (Tokyo) 98: 1147-1156.

143
Nederlandse Samenvatting
Dit is een vereenvoudigde samenvatting van de inhoud van dit proefschrift. De lezer die op zoek is naar wetenschappelijke verantwoording en inhoudelijke details wordt verwezen naar de Engelstalige samenvatting en hoofdstuk 1 t/m 5.

Alpha-glucan acting enzymes in Aspergillus niger
144
Schimmels zijn overal, al zie je ze meestal niet. Schimmels zitten in de grond, in de kaas, in de koelkast en de badkamer, en op je pizza ai fungi. Veel mensen weten niet dat schimmels, ook wel fungi genoemd, hun eigen rijk toegewezen hebben gekregen, naast bijvoorbeeld het dierenrijk en het plantenrijk. Tot het rijk der schimmels behoren de paddestoelen, de gisten, en de filamenteuze (draadvormige) schimmels. Sommige soorten schimmels kunnen ziektes veroorzaken bij planten of dieren, of ze kunnen voedsel bederven door de vorming van gifstoffen. Andere soorten, de zogenoemde mycorrhiza, zijn juist van groot belang voor het plantenleven: ze leven in of rond plantenwortels en helpen de plant om voldoende water en zouten op te nemen uit de bodem. Ook mensen kunnen belang hebben bij schimmels, bijvoorbeeld wanneer ze worden gebruikt bij het bereiden van voedsel. Het bekendst is waarschijnlijk het gebruik van gist bij het maken van bier en brood, maar ook voor de bereiding van brie en blauwe kaas, of meer exotische producten als Quorn en Japanse sojasaus en miso zijn schimmels nodig. In dit proefschrift is de algemeen voorkomende draadvormige bodemschimmel Aspergillus niger bestudeerd. Een soort die sommige mensen misschien kennen als een zwarte aanslag op vochtige plaatsen in huis. A. niger is voor mensen niet schadelijk en wordt veel gebruikt in de moderne biotechnologie; de schimmel kan heel efficiënt citroenzuur produceren, een ingrediënt van bijvoorbeeld frisdrank, en wordt daarvoor op grote schaal gekweekt. Schimmels groeien op allerlei soorten organische stoffen, vaak resten van dode planten of dieren. Voordat de grote organische moleculen uit de voedingsbodem door de schimmel kunnen worden opgenomen, moeten ze buiten de schimmelcel worden afgebroken tot kleinere moleculen. Daarvoor maakt de schimmel zogeheten enzymen, eiwitten die een chemische reactie versnellen zonder daarbij zelf verbruikt te worden (zg. biokatalysatoren). De stoffen die betrokken zijn bij de reactie van een enzym, in dit geval de grote moleculen die in stukjes worden geknipt, worden het substraat genoemd. Een voorbeeld van zo’n systeem is de afbraak van zetmeel door schimmelenzymen. Zetmeel wordt gemaakt als opslag van reservevoedsel in planten, en kan door bijvoorbeeld bodemschimmels als voedsel worden gebruikt als de plant dood gaat. Zetmeel bestaat uit lange moleculen van aan elkaar verbonden suikers (glucose, zie Figuur 1A). Amylases zijn enzymen die zetmeel kunnen afbreken. Deze enzymen worden door de schimmel gemaakt en naar buiten gebracht, waar ze zetmeelketens in kleinere moleculen knippen. Dit proces, waarbij zetmeel het substraat is van amylase, wordt hydrolyse genoemd.

Samenvatting
145
Fig. 1A Schematische weergave van een glucosemolecuul, een ringvormige structuur met 6 koolstofatomen. De nummers geven aan waar de zes koolstofatomen zich bevinden en wat hun nummering is. Ieder koolstofatoom heeft een OH-groep die potentieel een binding kan vormen met een ander glucose molecuul. B Schematische weergave van een deel van een zetmeelmolecuul (de OH-groepen zijn niet weergegeven). Zetmeel is opgebouwd uit glucosemoleculen die via een zuurstofmolecuul (O) aan hun eerste koolstofatoom verbonden zijn met het vierde koolstofatoom van het volgende glucosemolecuul (een α-(1,4) binding). Een
paar procent van de glucosemoleculen is bovendien via het eerste koolstofatoom aan het zesde koolstofatoom van een ander molecuul gebonden (α-(1,6) binding). In werkelijkheid bestaat een zetmeel molecuul uit 5000 tot 100,000 verbonden glucosemoleculen. C Schematische weergave van een binding zoals die veel voorkomt in de α-glucanen van schimmelcelwanden. De meeste glucosemoleculen zijn verbonden via hun eerste en derde koolstofatoom (een α-(1,3) binding). Deze α-glucanen bestaan in werkelijkheid uit tenminste enkele honderden aan elkaar gekoppelde glucosemoleculen. De verkorte zetmeelketens worden nog verder afgebroken door andere schimmelenzymen, glucosidases en glucoamylases, zodat uiteindelijk glucose wordt gevormd. Een glucosemolecuul is klein genoeg om als voedselbron te worden opgenomen in de cellen van A. niger en andere schimmels. Ook voor mensen en veel andere dieren is zetmeel een belangrijke bron van energie. Wij hebben dus ook

Alpha-glucan acting enzymes in Aspergillus niger
146
amylases, glucosidases en glucoamylases om zetmeel, uit bijvoorbeeld brood of aardappels, af te breken tot suikermoleculen die worden opgenomen in het bloed.
Enzymen werken over het algemeen maar op één specifiek substraat. Een schimmel moet daarom de enzymen produceren voor de voedselbron die op een bepaald moment beschikbaar is. Amylase wordt bijvoorbeeld vooral gemaakt als er zetmeel in de buurt is. Maar hoe “weet” een schimmelcel dat? Daarvoor is een uitgebreid regulatiesysteem nodig, en hoewel de details van dit systeem op dit moment nog niet bekend zijn, is het wel duidelijk dat een eiwit uit A. niger hierin een grote rol speelt. Zonder dit eiwit, dat AmyR wordt genoemd (van Amylase-Regulator), groeit de schimmel erg slecht op zetmeel omdat er geen amylases en andere enzymen meer worden aangemaakt die het voedsel kunnen afbreken. Eiwitten, dus ook enzymen, bestaan uit een keten van aminozuren. De volgorde waarin de aminozuren aan elkaar gekoppeld zijn is vastgelegd in een stukje van het DNA dat een gen wordt genoemd. Een gen codeert zo voor een eiwit. De keten van aminozuren vouwt zich op tot een driedimensionale structuur, waardoor het functionele eiwit wordt gevormd. Enzymen met dezelfde functie, zoals amylases uit verschillende schimmelsoorten, lijken vaak sterk op elkaar, niet alleen wat betreft de volgorde van de aminozuren maar ook in hun driedimensionale structuur. Vooral vier regio’s in de aminozuurketen zijn vrijwel identiek in alle amylases, of ze nu gemaakt worden door schimmels, bacteriën of mensen. Deze vier regio’s zijn belangrijk voor het maken van een goedwerkend enzym, omdat ze na de vouwing de plek vormen waar zetmeelketens binden aan het enzym en geknipt worden. Deze regio’s zijn ook herkenbaar in de genen die coderen voor de amylases. Daarom kunnen ze gebruikt worden om te zoeken naar genen die coderen voor nog onbekende amylases in het totale DNA van de schimmel, dat is opgeslagen in een database. Het totale DNA van een soort heet het genoom, en de laatste jaren is van steeds meer soorten, inclusief de mens en veel soorten schimmels, het genoom bekend geworden. Zoals beschreven in hoofdstuk 2 hebben we op deze manier in A. niger in totaal negen amylases gevonden, waarvan er drie al eerder beschreven waren. We hebben getest of deze enzymen vooral door de schimmel gemaakt werden als zetmeel de voedselbron is, en of ze gereguleerd worden door AmyR. Zo konden we een uitspraak doen over de mogelijke functie van al die enzymen. Eén van de nieuwe amylases, AmyC genaamd, lijkt inderdaad betrokken bij de afbraak van zetmeel buiten de cel. Voor de andere enzymen, die verdeeld zijn over twee groepen, lijkt zo’n functie onwaarschijnlijk. De eigenschappen van deze enzymen en hun mogelijke functie in de schimmel is in de latere hoofdstukken onderzocht.

Samenvatting
147
De eerste nieuwe groep, beschreven in hoofdstuk 3 en 4, bestaat uit drie enzymen die op elkaar lijken en in een aantal opzichten verschillend zijn van de bekende amylases. Een van de vier belangrijke regio’s is verschillend van andere amylases en bovendien kan op basis van de volgorde van aminozuren worden voorspeld dat de drie eiwitten worden vastgezet op de buitenkant van de cel. Dit gebeurt via een GPI-anker, een structuur van suikers en vetzuren, die het uiteinde van het eiwit verbindt met de celwand. Er zijn maar relatief weinig eiwitten die met een GPI-anker worden vastgezet: een schimmel maakt ongeveer honderd verschillende eiwitten die op die manier worden vastgezet, terwijl het duizenden andere eiwitten maakt die de cel uitgestuurd worden zonder verankering. In hoofdstuk 3 en 4 wordt beschreven hoe we twee van deze bijzondere eiwitten hebben geproduceerd in A. niger, en hoe we ze daarna hebben gezuiverd en gekarakteriseerd. Beide enzymen bleken zetmeel niet te hydrolyseren zoals amylases, maar knipten een stuk van een zetmeelmolecuul af en plakten dat deel weer aan een ander molecuul, dat zo verlengd werd (Van der Kaaij et al., 2007). Een enzym dat een deel van een substraat overbrengt naar een ander substraat heet een glucanotransferase en deze twee A. niger enzymen zijn daarom α(alpha)-glucanotransferase A en B genoemd, afgekort tot AgtA en AgtB. Enzymen die op amylases lijken en zo’n transglycosylasereactie uitvoeren zijn eerder beschreven in bijvoorbeeld bacteriën en planten, maar dit soort enzymen waren nog niet ontdekt in schimmels. Toch blijkt uit analyse van het DNA van andere filamenteuze schimmels dat ze ook dit soort enzymen kunnen maken. De aanwezigheid van dit type enzymen in veel schimmelsoorten is een aanwijzing dat ze een rol spelen bij een proces dat in al deze schimmels van belang is. Om te onderzoeken wat de functie van Agt-enzymen is hebben we een A. niger stam gemaakt die geen AgtA meer kan maken, een zogeheten knockout stam. Het effect daarvan werd zichtbaar door de knockout te laten groeien in aanwezigheid van een stof die de celwanden van schimmels destabiliseert. In aanwezigheid van deze stof groeide de knockout veel langzamer dan de originele A. niger stam met AgtA (zie figuur 8 van hoofdstuk 3). Blijkbaar is AgtA nodig om een sterke celwand te vormen. In een recente publicatie wordt een vergelijkbare proef gedaan met een andere schimmel, Schizosaccharomyces pombe (Morita et al., 2006). Deze schimmel heeft een aantal enzymen die lijken op AgtA, en de knockout van één ervan leidde ook tot een zwakkere celwand. Al met al hebben we een aantal aanwijzingen dat de Agt-enzymen niet worden gemaakt om zetmeel af te breken, maar dat ze een rol spelen in de celwand van A. niger en andere filamenteuze schimmels. Zo zien we dat AmyR geen effect heeft op het maken van de Agt-enzymen, terwijl alle enzymen waarvan we weten dat ze zetmeel afbreken wel door AmyR gereguleerd worden. Ook hebben we laten zien dat de knockout van AgtA geen effect had op de groei van de schimmel op zetmeel, terwijl het

Alpha-glucan acting enzymes in Aspergillus niger
148
wel een effect had op stevigheid van de celwand. Ook het GPI-anker is een aanwijzing, want het verankert deze enzymen in de buurt van hun vermoedelijke substraat, de celwand. Bovendien zijn er geen gewone amylases bekend die zo’n anker hebben. Om het definitieve bewijs te leveren voor de rol van Agt-enzymen zou je de samenstelling van de celwand van een knockout en een gewone A. niger moeten vergelijken; volgens onze hypothese zouden die verschillend moeten zijn. Dergelijke experimenten zijn in het onderzoek beschreven in dit proefschrift niet uitgevoerd. De tweede nieuwe groep van amylases die we in het genoom van A. niger geïdentificeerd hebben bestaat uit een tweetal verwante enzymen die AmyD en AmyE zijn genoemd. Vergeleken met alle schimmelamylases die eerder in de literatuur zijn beschreven, hebben deze twee enzymen een sterk afwijkende aminozuurvolgorde. Het is opvallend dat ze meer lijken op een groep amylases uit bacteriën dan op de bekende amylases uit schimmels (zie figuur 2). Ook in het genoom van andere schimmelsoorten blijken zulke ‘vreemde’ amylases voor te komen. Op basis van de aminozuurvolgorde voorspellen we dat al deze eiwitten in de cel blijven, zodat een rol bij de afbraak van grote zetmeelmoleculen buiten de cel onwaarschijnlijk is. In hoofdstuk 5 staat beschreven dat het enzym AmyD weliswaar amylase-activiteit heeft, maar dat die activiteit honderd tot duizend keer lager is dan die van bekende schimmelamylases. Het enzym maakt voornamelijk maltotriose vrij, dat zijn drie aan elkaar gekoppelde glucosemoleculen. Omdat AmyD en AmyE in de in hoofdstuk 2 beschreven experimenten niet door A. niger geproduceerd werden, zijn er weinig aanknopingspunten voor een hypothese over hun functie. Zo’n hypothese wordt wel geleverd in een publicatie over een enzym uit Histoplasma capsulatum, een schimmel nauw verwant aan de aspergilli. Dit eiwit, genaamd Amy1p, heeft dezelfde afwijkende aminozuurvolgorde als AmyD en AmyE. Een knockout stam die geen Amy1p meer kon produceren bleek een onderdeel van de celwand, de α-glucanen, niet meer te kunnen maken (Marion et al., 2006). Naast de Agt-enzymen lijken dus ook deze nieuwe amylase-achtige AmyD/E enzymen een rol te spelen in de celwand van schimmels. De celwand van schimmels vormt de buitenkant van het organisme en beschermt deze tegen uitdroging en chemische invloeden van buitenaf. De celwand kan ook stevigheid, structuur en kleur geven, zoals goed zichtbaar is bij paddestoelen. Een van de onderdelen van zo’n schimmel celwand zijn de α-glucanen, die lijken op zetmeel. Beide soorten moleculen bestaan uit ketens van gekoppelde glucosemoleculen, maar de binding tussen de glucosemoleculen is bij α-glucanen meestal van een andere soort. In zetmeel bestaan de bindingen voornamelijk tussen het eerste en het vierde koolstofatoom van de glucosemoleculen (een α-(1,4) binding), terwijl in celwand α-glucanen de binding meestal bestaat tussen het eerste en het derde koolstofatoom (een α-(1,3) binding) hoewel er ook α-(1,4) bindingen in kunnen voorkomen (zie Fig. 1B en

Samenvatting
149
Fig. 2 Schematische weergave van de verwantschap tussen amylases uit verschillende organismen. Hoe langer de lijn tussen twee eiwitten, hoe groter de verschillen in hun aminozuurvolgorde. Er wordt aangenomen dat enzymen die veel op elkaar lijken zich relatief recent in de evolutie uitgesplitst hebben. De ‘gewone’ schimmel amylases en de Agt-enzymen lijken relatief veel op elkaar, en vallen in dezelfde groep. AmyD en AmyE, samen met andere enzymen uit filamenteuze schimmels, hebben een andere aminozuurvolgorde en vormen daarom hun eigen groep. Ook is zichtbaar dat bijvoorbeeld amylases uit planten een aparte groep vormen. C). De samenstelling van α-glucanen verschilt per schimmelsoort en is lang niet altijd in detail bekend. De α-glucanen worden gemaakt door speciale enzymen die in het celmembraan zitten, zogeheten α-glucansynthases. Een deel van het α-glucansynthase bevindt zich aan de binnenkant, waar het geactiveerde glucosemoleculen (UDP-glucose) aan elkaar koppelt en zo een α-glucanketen vormt (Fig. 3). Deze glucanketens worden vervolgens naar de buitenkant van de cel gebracht, waar ze een plek krijgen tussen de andere onderdelen van de celwand. De α-glucansynthase enzymen hebben nog een ander onderdeel dat op amylases lijkt en zich aan de buitenkant van de cel bevindt. De functie van dit gedeelte is nog niet

Alpha-glucan acting enzymes in Aspergillus niger
150
helemaal duidelijk, maar mogelijk knipt het de α-glucanketens om ze op een andere manier weer met elkaar te verbinden. Dit bestaande, deels hypothetische model van glucanproductie in schimmels kan nu worden uitgebreid met de twee groepen enzymen die in dit proefschrift voor het eerst beschreven zijn (Fig. 3). De AmyD/E-enzymen die in de cel zitten produceren mogelijk een kort molecuul, zoals maltotriose, dat kan dienen als startpunt voor de aankoppeling van UDP-glucose door α-glucansynthases. Dit molecuul is vormt zo een ‘primer’ voor de vorming van α-glucan. De Agt-enzymen, die met hun GPI-anker op de buitenkant van de cel zitten, kunnen misschien de α-glucanketens knippen en ze op andere manieren weer aan elkaar plakken. Zo spelen ze mogelijk een rol bij het flexibel houden van de α-glucanen in de celwand, zodat groei en vertakking van de schimmeldraden mogelijk blijft. Dit proefschrift laat duidelijk zien dat wetenschappelijk onderzoek zich niet altijd laat plannen. We begonnen met het zoeken naar nieuwe amylases voor de afbraak van zetmeel, maar eindigden bij de enzymatische synthese en modificatie van α-glucanen in schimmelcelwanden. Hiermee hebben we een begin gemaakt met het onderzoeken van een wetenschappelijk interessant en heel nieuw gebied van de schimmelfysiologie. Behalve de inhoud zijn ook de mogelijke praktische toepassingen van de resultaten van dit onderzoek veranderd. Het afbreken van zetmeel of het veranderen van de structuur ervan met enzymen is belangrijk in de voedingsmiddelenindustrie, dat de aanleiding gaf tot het bestuderen van dit systeem in A. niger. Aan de andere kant kan kennis over de celwanden van schimmels van belang zijn voor de ontwikkeling van medicijnen tegen schimmelinfecties. Een nauwe verwant van A. niger, Aspergillus fumigatus, veroorzaakt de ernstige ziekte aspergillosis. Hierbij infecteert de schimmel de luchtwegen, en verspreidt zich soms ook in de hersenen en andere organen. Een van de belangrijkste beschikbare medicijnen tegen aspergillosis werkt in op de celwand: het blokkeert de vorming van β(beta)-glucanen, een van de andere celwand onderdelen. Zonder β-glucanen raakt de schimmel verzwakt en is hij veel gevoeliger voor het menselijke immuunsysteem. Tot nu toe is er weinig aandacht geweest voor de α-glucanen als mogelijk aangrijpingspunt voor medicijnen. De uitbreiding van kennis over de synthese van α-glucanen, waaraan het onderzoek beschreven in dit proefschrift een bijdrage heeft geleverd, kan hopelijk resulteren in de ontwikkeling van nieuwe medicijnen tegen ziektes veroorzaakt door schimmels.

Samenvatting
151
Fig. 3 Schematische weergave van het hypothetische model van de vorming van α-glucan in de celwand van filamenteuze schimmels. AmyD of AmyE maken een korte suiker, zoals maltotriose, als primer voor de aankoppeling van UDP glucose door het binnendeel van het α-glucansynthase (Ags). De gevormde α-glucanketen wordt dan via een pore in de celmembraan naar de buitenkant van de cel getransporteerd. Hier worden de α-glucanketens aan elkaar gekoppeld door het buitenste, amylase-achtige deel van Ags, of door een van de α-glucanotransferase (Agt)-enzymen. Referenties Marion, C.L., Rappleye, C.A., Engle, J.T., and Goldman, W.E. (2006) An alpha-(1,4)-amylase is essential for alpha-(1,3)-glucan production and virulence in Histoplasma capsulatum. Molecular Microbiology 62: 970-983. Morita, T., Tanaka, N., Hosomi, A., Giga-Hama, Y., and Takegawa, K. (2006) An alpha-amylase homologue, aah3, encodes a GPI-anchored membrane protein required for cell wall integrity and morphogenesis in Schizosaccharomyces pombe. Bioscience, Biotechnology & Biochemistry 70: 1454-1463. Van der Kaaij R.M., Yuan X.L., Franken A., Ram A.F.J., Punt P.J., Maarel M.J.E.C. and Dijkhuizen L. (2007) Characterization of two novel, putatively cell wall associated and GPI-anchored, alpha-glucanotransferase enzymes of Aspergillus niger. Eukaryotic Cell 2007 Jul;6(7):1178-88.


153
List of publications A. Schouten, L. Wagemakers, F.L. Stefanato, R.M. van der Kaaij and J.A. van Kan (2003). Resveratrol acts as a natural profungicide and induces self-intoxication by a specific laccase. Molecular Microbiology 43(4):883-94. Patent No. 06122600.7-2405 ‘Glucanotransferase’. Filed on 19/10/2006 by DSM. Inventors: R.M. van der Kaaij, M.J.E.C.van der Maarel and L. Dijkhuizen H.J. Pel, J.H. de Winde, D.B. Archer, P.S. Dyer, G. Hofmann, P.J. Schaap, G. Turner, R.P. de Vries, R. Albang, K. Albermann, M.R. Andersen, J.D. Bendtsen, J.A. Benen, M. van den Berg, S. Breestraat, M.X. Caddick, R. Contreras, M. Cornell, P.M. Coutinho, E.G. Danchin, A.J. Debets, P. Dekker, P.W. van Dijck, A. van Dijk, L. Dijkhuizen, A.J. Driessen, C. d'Enfert, S. Geysens, C. Goosen, G.S. Groot, P.W. de Groot, T. Guillemette, B. Henrissat, M. Herweijer, J.P. van den Hombergh, C.A. van den Hondel, R.T. van der Heijden, R.M. van der Kaaij, F.M. Klis, H.J. Kools, C.P. Kubicek, P.A. van Kuyk, J. Lauber, X. Lu, M.J. van der Maarel, R. Meulenberg, H. Menke, M.A. Mortimer, J. Nielsen, S.G. Oliver, M. Olsthoorn, K. Pal, N.N. van Peij, A.F. Ram, U. Rinas, J.A. Roubos, C.M. Sagt, M. Schmoll, J. Sun, D. Ussery, J. Varga, W. Vervecken, P.J. van de Vondervoort, H. Wedler, H.A. Wosten, A.P. Zeng, A.J. van Ooyen, J. Visser and H. Stam (2007). Genome sequence and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nature Biotechnology 25; 221-231. W.H. van der Putten, G.A. Kowalchuk, E.P. Brinkman, G.T.A. Doodeman, R.M. van der Kaaij, A.F.D. Kamp, F.B.J. Menting and E.M. Veenendaal (2007). Neutral to positive soil feedback of non-native savannah grass relates to absence of soil pathogens and selective mycorrhizal infection. Ecology 88(4):978–988. R.M. van der Kaaij, X-L. Yuan, A. Franken, A.F.J. Ram, P.J. Punt, M.J.E.C. van der Maarel, and L. Dijkhuizen (2007). Characterization of two novel, putatively cell wall associated and GPI-anchored, α-glucanotransferase enzymes of Aspergillus niger. Eukaryotic Cell 2007 Jul;6(7):1178-88. R.M. van der Kaaij, Š. Janeček, M.J.E.C. van der Maarel and L. Dijkhuizen (2007). Phylogenetic and biochemical characterization of a novel group of intracellular fungal α-amylase enzymes. Accepted for publication in Microbiology.

Alpha-glucan acting enzymes in Aspergillus niger
154
R.M. van der Kaaij, M.J.E.C. van der Maarel and L. Dijkhuizen (2007). Biochemical characterization of GPI-anchored α-glucanotransferase enzymes from Aspergillus niger reveals a novel reaction specificity in subfamily GH13_1. Submitted. X-L Yuan, R.M. van der Kaaij, C.A.M.J.J. van den Hondel, P.J. Punt, M.J.E.C. van der Maarel, L. Dijkhuizen and A.F.J. Ram. Aspergillus niger genome wide analysis reveals a large number of novel α-glucan acting enzymes with unexpected expression profiles. Submitted.

155
Dankwoord Een van mijn stellingen had kunnen zijn, dat wetenschap en kunst met elkaar gemeen hebben dat het eindresultaat er zo makkelijk uit kan zien, terwijl de weg ernaar toe heel moeilijk is geweest. Maar dan geldt ook voor beiden, dat de bijdrage van alle mensen die helpen de hobbels in die weg te overwinnen niet direct in het eindproduct terug te vinden is. Daarom is, althans voor proefschriften, het dankwoord uitgevonden. Laat ik bij het begin beginnen. Ik had, waarschijnlijk, geen idee waar ik aan begon, maar in het eerste gesprek met Lubbert, Marc en Gert-Jan voelde ik mij direct op mijn gemak, en dat bleek in de daarop volgende jaren niet onterecht. Gert-Jan, ik ben blij dat je me nog hebt geholpen met ‘opstarten’. Marc, dank voor je constante vertrouwen in het project en mijn mogelijkheden daarin. Zonder je pep-talk had ik het misschien wel opgegeven. Hoe beter we elkaar leerden kennen, hoe beter de samenwerking ging, dus laten we dat nog even voortzetten. Lubbert, je bent als een rots in de branding voor Micphys. Ondanks je betrokkenheid bij zoveel verschillende projecten wist je altijd de juiste kritische vragen te stellen, en heb je me enorm geholpen bij het schrijfproces. Arthur, jij hoort zeker ook thuis in dit rijtje ‘begeleiders’. Dank voor je betrokkenheid bij mijn deel van het project. Inhoudelijk was je inbreng onmisbaar, maar ook je rustige en geduldige manier van werken heb ik erg gewaardeerd. Ook, of eigenlijk vooral, in het lab worden veel hobbels in de weg van het onderzoek overwonnen. Coenie, baie dankie voor het delen van je kennis, en voor het voeren van menige felle discussie. All other Micphys colleagues, especially those working within the CCB, thanks for all your support, sharing your knowledge and for creating a very good working atmosphere. Speciale dank aan Theo voor de levendige discussies tijdens de lunch. Ook dank aan alle collega’s binnen het IOP-project voor de uitwisseling van kennis en materiaal. Especially, thanks to Yuan for sharing her data. Iris, thanks for being around, I hope you learned as much from it as I did. Štefan, it was nice working together, I am glad we have been successful despite the physical distance. Voor ik aan dit project begon was Groningen voor mijn een verre, onbekende stad. Gelukkig waren er mensen bij wie ik meteen welkom was: bij Chris & Sonja, bij Nienke, en bij de zangers van Bonton. Mijn verhuizing betekende ook de overstap naar een andere (nóg leukere) rol bij Nivon Jeugd & Jongeren. Nivon-vrienden, ik heb het al vaker gezegd: van jullie heb ik de afgelopen tien jaar misschien nog wel het meest

Alpha-glucan acting enzymes in Aspergillus niger
156
geleerd. En wintervrienden, met jullie heb ik de meest verfrissende winteruitstapjes gemaakt die je je kunt wensen. Ik hoop nog veel (niet te grote) avonturen met jullie te beleven. Het echte begin ligt natuurlijk bij mijn ouders. Peter en Sylvia, en met jullie Simone en Ruud, jullie hebben mij altijd gesteund in mijn keuzes, ook als ze niet erg voor de hand lagen. Veel dank daarvoor. Inger en Michiel, ik ben blij dat jullie er zijn! Ik hoop dat jullie eigen weg zonder al te grote hobbels zal zijn. Ernst, jij hoort bij mij. Als geen ander maak je me vrolijk als je vraagt of ik vandaag weer een belangrijke uitvinding heb gedaan, of een nobelprijs heb gewonnen. Ik hoop dat je voor altijd naast me staat, want ik heb je lief. Rachel Augustus 2007

157
Appendix Genome sequence and analysis of the versatile cell factory Aspergillus niger CBS 513.88 Nature biotechnology 25; 221-231

Genome sequencing and analysis of the versatile cellfactory Aspergillus niger CBS 513.88Herman J Pel1, Johannes H de Winde1,2, David B Archer3, Paul S Dyer3, Gerald Hofmann4, Peter J Schaap5,Geoffrey Turner6, Ronald P de Vries7, Richard Albang8, Kaj Albermann8, Mikael R Andersen4,Jannick D Bendtsen9, Jacques A E Benen5, Marco van den Berg10, Stefaan Breestraat1, Mark X Caddick11,Roland Contreras12, Michael Cornell13, Pedro M Coutinho14, Etienne G J Danchin14, Alfons J M Debets15,Peter Dekker1, Piet W M van Dijck1, Alard van Dijk1, Lubbert Dijkhuizen16,17, Arnold J M Driessen17,Christophe d’Enfert18, Steven Geysens12, Coenie Goosen16,17, Gert S P Groot1, Piet W J de Groot19,Thomas Guillemette20, Bernard Henrissat14, Marga Herweijer1, Johannes P T W van den Hombergh1,Cees A M J J van den Hondel21, Rene T J M van der Heijden22, Rachel M van der Kaaij16,17, Frans M Klis19,Harrie J Kools5, Christian P Kubicek23, Patricia A van Kuyk21, Jurgen Lauber24, Xin Lu25, Marc J E C vander Maarel16, Rogier Meulenberg1, Hildegard Menke1, Martin A Mortimer11, Jens Nielsen4, Stephen G Oliver13,Maurien Olsthoorn1, Karoly Pal15,26, Noel N M E van Peij1, Arthur F J Ram21, Ursula Rinas25,Johannes A Roubos1, Cees M J Sagt1, Monika Schmoll23, Jibin Sun25, David Ussery27, Janos Varga26,28,Wouter Vervecken12, Peter J J van de Vondervoort21, Holger Wedler24, Han A B Wosten7, An-Ping Zeng25,Albert J J van Ooyen1, Jaap Visser29 & Hein Stam1
The filamentous fungus Aspergillus niger is widely exploited by the fermentation industry for the production of enzymes and
organic acids, particularly citric acid. We sequenced the 33.9-megabase genome of A. niger CBS 513.88, the ancestor of
currently used enzyme production strains. A high level of synteny was observed with other aspergilli sequenced. Strong function
predictions were made for 6,506 of the 14,165 open reading frames identified. A detailed description of the components of the
protein secretion pathway was made and striking differences in the hydrolytic enzyme spectra of aspergilli were observed. A
reconstructed metabolic network comprising 1,069 unique reactions illustrates the versatile metabolism of A. niger. Noteworthyis the large number of major facilitator superfamily transporters and fungal zinc binuclear cluster transcription factors, and the
presence of putative gene clusters for fumonisin and ochratoxin A synthesis.
Received 17 July 2006; accepted 27 November 2006; published online 28 January 2007; doi:10.1038/nbt1282
1DSM Food Specialties, PO Box 1, 2600 MA Delft, The Netherlands. 2Kluyver Centre for Genomics of Industrial Fermentation, Department for Biotechnology, DelftUniversity of Technology, Julianalaan 67, 2628 BC Delft, The Netherlands. 3School of Biology, University of Nottingham, University Park, Nottingham, NG7 2RD, UK.4Center for Microbial Biotechnology, Technical University of Denmark, Building 223, Søltofts Plads, DK-2800 Kgs. Lyngby, Denmark. 5Section Fungal Genomics,Laboratory of Microbiology, Wageningen University, Dreijenlaan 2, 6703 HA Wageningen, The Netherlands. 6Department of Molecular Biology and Biotechnology,University of Sheffield, Sheffield S10 2TN, UK. 7Microbiology, Science Faculty, Utrecht University, Padualaan 8, 3584 CH Utrecht, The Netherlands. 8Biomax InformaticsAG, Lochhamer Str. 9, 82152 Martinsried, Germany. 9CLC bio, Gustav Wieds Vej 10, 8000 Aarhus C, Denmark. 10DSM Anti Infectives, PO Box 425, 2600 KA Delft, TheNetherlands. 11The University of Liverpool, School of Biological Sciences, Biosciences Bld., Crownstreet, Liverpool L69 7ZB, UK. 12Ghent University and VIB, Dept.Molecular Biomedical Research, Unit of Fundamental and Applied Molecular Biology, Technologiepark 927, 9052 Gent, Belgium. 13Centre for the Analysis of BiologicalComplexity, Faculty of Life Sciences, The University of Manchester, Michael Smith Building, Oxford Road, Manchester M13 9PT, UK. 14Architecture et Fonction desMacromolecules Biologiques, UMR6098, CNRS. Universites Aix-Marseille I & II, Case 932, 163 Avenue de Luminy, 13288 Marseille, France. 15Laboratory of Genetics,Wageningen University , Arboretumlaan 4, 6703 BDWageningen, The Netherlands. 16Centre for Carbohydrate Bioprocessing (CCB) TNO-RuG, PO Box 14, 9750 AA Haren,The Netherlands. 17Department of Microbiology, Groningen Biomolecular Sciences and Biotechnology Institute and Materials Science Center Plus, University ofGroningen, PO Box 14, 9750 AA Haren, The Netherlands. 18Unite Postulante Biologie et Pathogenicite Fongiques, INRA USC 2019, Institut Pasteur, 75724 Paris Cedex15, France. 19Swammerdam Institute for Life Sciences, University of Amsterdam, Nieuwe Achtergracht 166, 1018 WV Amsterdam, The Netherlands. 20Laboratoire deMicrobiologie, UMR 77, Pathologie Vegetale, Universite d’Angers, 49045 Angers Cedex, France. 21Institute of Biology Leiden, Leiden University, Molecular Microbiology,Wassenaarseweg 64 2333 AL Leiden, The Netherlands. 22CMBI, Radboud University Medical Centre, PO Box 9101, 6500 HB Nijmegen, The Netherlands. 23Institute ofChemical Engineering, Research Area Gene Technology and Applied Biochemistry, Technical University Vienna, 1060 Vienna, Austria. 24Qiagen Genomics Services,Qiagen GmbH., 40724 Hilden, Germany. 25Helmholtz Center for Infection Research (former GBF-German Research Centre for Biotechnology), Inhoffenstrasse 7, D-38124Braunschweig, Germany. 26Department of Microbiology, Faculty of Sciences, University of Szeged, PO Box 533, H-6701 Szeged, Hungary. 27Center for BiologicalSequence Analysis, BioCentrum-DTU, Building 301, The Technical University of Denmark, DK-2800 Kgs. Lyngby, Denmark. 28CBS Fungal Biodiversity Centre,Uppsalalaan 8, 3584 CT Utrecht, The Netherlands. 29Fungal Genetics and Technology Consultancy, PO Box 396, 6700 AJ Wageningen, The Netherlands. Correspondenceshould be addressed to H.S. ([email protected]).
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 221
ART I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

Aspergillus niger, a member of the black aspergilli, is widely used inbiotechnology for the production of food ingredients, pharmaceuticalsand industrial enzymes. In their natural habitat A. niger strainssecrete large amounts of a wide variety of enzymes needed to releasenutrients from biopolymers. This high secretory capacity is exploitedby industry in both solid state and submerged fermentations1,2.A. niger has a long tradition of safe use in the production ofenzymes and organic acids. Many of these products have obtainedGRAS (generally regarded as safe) status3. Aspergillus enzymes are usedin starch processing, baking, brewing and beverage industries, inanimal feed and in the paper and pulping industry. Furthermore,A. niger is used as host for the production of heterologous proteins4,5
and as cell factory for the production of citric acid and gluconic acid6.A. niger exhibits a remarkably versatile metabolism, enabling growthon a wide range of substrates under various environmental conditions.Its ability to degrade a range of xenobiotics through various oxidative,hydroxylation and demethylation reactions provides potential foruse in bioremediation7.In this paper we describe the genomic DNA sequence of A. niger
strain CBS 513.88, its annotation and an initial gene expression studyusing Affymetrix DNA microarrays. CBS 513.88 is an early ancestor ofcurrently used enzyme production strains. Several features are com-pared with the recently published genomes of the aspergilli A. oryzae8,A. nidulans9 and A. fumigatus10. The availability of the genome willfacilitate the development of new products, improved strains andmore efficient processes.
RESULTS
Genome sequence and analysis
The genome of A. niger CBS 513.88 was sequenced using anordered set of large insert Escherichia coli bacterial artificial chromo-somes (BACs) in a process called BAC walking. First, a large insert(up to 150 kb) BAC library covering the genome more than20 times was constructed and used to generate over 18,000 BACend sequences with an average length of 500 base pairs (bp).In parallel, several of the larger BACs were sequenced using a7.5� coverage shotgun approach. Subsequently, sequences of thesefully sequenced ‘seed’ BACs were compared to the BAC ends,allowing the selection of a new set of BACs showing overlap withone of the sequenced BACs. The newly selected BACs were used forshotgun sequencing. A total of 505 BACs, representing a minimaltiling set covering the genome, were selected and sequenced using theBAC walking approach. The assembled genome sequence consists of468 DNA contigs spanning a total of 33.9 million unique bp (Mbp)arranged in 19 supercontigs. Recently an 8.9� shotgun coverage draftsequence of the citric acid–producing A. niger ATCC 1015 was madeavailable by the DOE Joint Genome Institute (http://www.jgi.doe.gov/aspergillus). Based on Blastn analysis the B37.2-Mbp genomesequence is 97% identical to and largely colinear with the genomesequence published here.
Gene identification and annotation
Algorithms specifically trained on the basis of genes known in A. nigerand related filamentous fungi were used to predict 14,165 proteincoding genes (Table 1). Because automated gene-model predictionsfrequently fail to correctly predict all intron and exon boundaries,extensive manual curation was used to verify the predicted genemodels and to assign gene functions on the basis of similarity toknown proteins with an established function. In this way we improvedmainly the models of genes encoding proteins with similarity toknown proteins.
The specificity of the 269 tRNA genes and the codon usage arepresented in Supplementary Tables 1 and 2 online and confirm thepublished usage11. A. oryzae has a comparable set of tRNAs (245),whereas A. nidulans and A. fumigatus contain 188 and 179 tRNAs,respectively. Whereas the observed tRNA density is higher for A. nigerand A. oryzae, the codon distribution is similar for all four species. Thefunctional catalog (FunCat) classification system12 was applied tofunctionally describe the A. niger predicted proteome. Of the 14,165predicted proteins, approximately half (6,505) could be assigned tofunctional protein classes relating to metabolism, cellular transportand protein fate (Fig. 1). A detailed comparison was made betweenA. niger and eight other filamentous fungi (Fig. 1): 9,253 A. nigerproteins have an ortholog in at least one of the nine queried species,1,992 proteins have orthologs in all species, whereas 3,373 proteins areshared between the filamentous fungi. The distribution of orthologousproteins over the various FunCat classes is shown (Fig. 1). Comparedto the other filamentous fungi A. niger contains a remarkably largenumber of unique proteins involved in C-compound, carbohydrate,lipid, fatty-acid and isoprenoid metabolism and secondary metabo-lism, reflecting the versatility of A. niger as a cell factory. In contrast,the number of unique proteins involved in cellular transport, proteinsecretion and fate are rather constant between the various filamentousfungi. Apparently A. niger is able to use its secretion machinery ina very efficient way.
Phylogenetic relationship and synteny between aspergilli
Phylogenetic relationships between filamentous fungi have often beenbased on ribosomal DNA sequences or single-gene families13. Weselected twenty strictly orthologous sequences from A. niger and eightfilamentous fungi. For A. terreus and A. clavatus no annotatedproteome is available, hence translations of orthologous gene frag-ments were used instead. The multiple sequence alignment wasmanually curated and used to build a maximum likelihood tree(Supplementary Table 3 and Supplementary Fig. 1 online). In thistree A. niger is closely related to both A. terreus and A. oryzae andseparate from A. fumigatus and A. clavatus, which have smallergenomes (29–30 Mb), and A. nidulans, which branches earlier.Of the 8,695 A. niger genes that have an ortholog in A. nidulans,
A. oryzae and A. fumigatus, 6,755 (78%) show conservation ofneighboring orthologs (synteny) in at least one of the three otherspecies; 4,189 genes (48%) are syntenic in all four Aspergillus speciesand were plotted on the proposed physical map of A. niger. Largeparts of the A. niger genome show a high conservation of gene orderwith one of the other Aspergillus species, in line with previousobservations9. However, within those regions numerous intrachro-mosomal rearrangements and microinversions have occurred (Fig. 2).
Table 1 Genome characteristics of A. niger CBS 513.88
Size assembled genome (Mb) 33.9
GC content assembled genome (%) 50.4
Protein-coding genes (all) 14,165
Protein-coding genes (o100 aa) 927
GC content protein-coding genes (%) 53.4
Average gene length (bp) 1,572
Gene density (genes/kb) 0.42
Average number of introns per gene 2.57
Average intron size (bp) 97
Average exon size (bp) 370
Number of tRNA genes 269
222 VOLUME 25 NUMBER 2 FEBRUARY 2007 NATURE BIOTECHNOLOGY
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

A. nidulans centromere-flanking genes mapped without exception toeither the 5¢ or 3¢ end of a large A. niger supercontig and for eachlinkage group two centromere flanking supercontig ends were found.However, no clear centromeric sequences were identified. Inferredtelomeric regions show little to no synteny (Fig. 2).
Linking the A. niger physical map to the genetic map
Parasexual recombination and electrophoretic karyotyping were pre-viously used to establish the presence of eight linkage groups in thewidely studied A. niger strain CBS 120.49 (refs. 14,15). We used thesemethods to assign 72 characterized or newly selected genes to specificchromosomes (Supplementary Table 4 online). This positioningunambiguously links all supercontigs of CBS 513.88 to single chromo-somes of CBS 120.49. Electrophoretic karyotype data indicate thatchromosomes VI and VII may not be completely covered by sequencedata (Table 2). For chromosome VI this most likely relates to thepresence of rDNA repeats whereas for chromosome VII this may bedue to a large deletion in strain CBS 513.88. Four different methodswere used to establish the orientation of supercontigs on the chromo-somes resulting in a proposed alignment of the 19 supercontigs ofstrain CBS 513.88 with the eight linkage groups of strain CBS 120.49(Table 2 and Supplementary Data online).
Life cycle and reproduction
The majority of aspergilli, including A. niger are only known toreproduce by asexual means, forming conidiospores16. The identifiedset of genes involved in signal transduction and conidiophore develop-ment is essentially the same as that of A. nidulans (SupplementaryTable 5 online). Asexuality is thought to be derived from an ancestralsexual state17. We therefore screened the genome of A. niger for thepresence of genes involved in incompatibility (Supplementary Table 6online) and sexual reproduction (mating processes, signal transduc-tion and ascomata development). A full complement of apparently
functional, early sexual development genes, including a key mat-1alpha domain mating-type gene was identified (Supplementary Table5 and Supplementary Data). Two genes were only partially retrievedand one (pro1) contained a premature stop codon. mat-1 andpheromone-precursor and receptor genes were not expressed duringfed-batch fermentation, but this was not surprising given that matingand pheromone signaling require specific environmental conditionsfor induction (Supplementary Table 7 online). It has recently beensuggested that the ‘asexual’ species, A. fumigatus and A. oryzae mighthave sexual potential9,18. A similar situation could apply to A. niger,which would be of great value for strain improvement.
Cell wall development
The fungal cell wall determines biotechnologically relevant featuressuch as morphology during fermentation and cell integrity. The cellwall of A. niger consists of chitin, 1,3-b-glucan, 1,6-b-glucan, 1,3-a-glucan, galactosaminogalactan and galactomannan, similar toA. fumigatus19. Genes required for the biosynthesis of these cell wallcomponents were identified in the genome (Supplementary Table 8online). As in other aspergilli, a high degree of redundancy is observedfor chitin synthases and chitinases compared to Saccharomycescerevisiae (Supplementary Table 9 online). A. niger contains fiveputative 1,3-a-glucan synthase genes, the highest number in thefour published Aspergillus genomes. Specific members of each genefamily were expressed during vegetative growth in a fed-batch cultureon glucose and ammonium (Supplementary Table 7).The cell wall integrity (CWI) signaling pathway is responsible for
cell wall remodeling and reinforcement in response to a changingenvironment20. Several components of this pathway are present in thegenome (Supplementary Table 8) and indeed expressed during fed-batch culture, ensuring the growth of A. niger under harsh industrial-process conditions (Supplementary Table 7 and SupplementaryFig. 2 online).
Transposable elements50
Subcellular organization2,509
Metabolism3,002
Homeostasis242
Energy326
Cell cycle 506
Transcription944
Protein synthesis273Protein fate
901
Transport819
01 Metabolism: 3,002
01.05 01.20 01.06 Other 08.07 08.16 08.19 Other 06.04 06.07 Other
1316 574 504 608 152 238 378 207 189 417 295
SceMgiNcrSnoAniAfuAorAng
08 Transport: 975 06 Protein fate: 901
Cellular transport975
Communication326
Rescue/death946
a
b
Figure 1 Functional classification of A. niger
ORFs. (a) FunCat classes12 are, 01, Metabolism;
02, Energy; 03, Cell cycle and DNA processing;
04, Transcription; 05, Protein synthesis; 06,
Protein fate (folding, modification, destination),
67, Transport facilitation; 08, Cellular transport
and transport mechanisms; 10, Cellular
communication/signal transduction mechanism;11, Cell rescue, defense and virulence;
13, Regulation of/interaction with cellular
environment; 40, Subcellular localization; and
29, Transposable elements. The numbers
following class names represent the number
of genes in each category. (b) Comparison of
functional classes in different fungi. Sce,
Saccharomyces cerevisiae; Mgi, Magnaporthe
grisea; Ncr, Neurospora crassa; Sno,
Stagonospora nodulum; Ani, Aspergillus nidulans;
Afu, Aspergillus fumigatus; Aor, Aspergillus
oryzae; Ang, Aspergillus niger. Metabolism (01):
C-compound and carbohydrate metabolism
(01.05), lipid, fatty-acid and isoprenoid
metabolism (01.06) and secondary metabolism
(01.20); Cellular transport (08): vesicular
transport (08.07), extracellular transport,
exocytosis and secretion (08.16) and cellular
import (08.19); Protein fate (06): proteinsecretion and fate (06.04) and cellular transport
(06.07). The lengths of the bars are proportional
to the number of genes in each category.
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 223
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

Hydrophobins are important determinants of cell morphologythrough their amphipathic monolayer structure21 (SupplementaryTable 8). Two out of seven of the predicted hydrophobin genesare expressed during submerged fermentation (SupplementaryTable 7). By modifying the expression of hydrophobin genes, thecell wall composition and hyphal morphology of A. niger maybe altered, for example, to improve protein production22. Severalgenes involved in autolysis are present16,23. Reduced autolysisthrough controlled expression of these genes is relevant for industrialprocesses to minimize proteolysis of secreted proteins and to simplifydownstream processing24.
Central metabolism and organic acid production
A novel genome-scale model of A. niger central metabolism wasdeveloped on the basis of the A. niger genome. This metabolicmodel was constructed by supplementing literature on A. nigermetabolism with pathway databases and literature on other asper-gilli25,26. The reconstructed metabolic network consists of 1,069
unique reactions, of which 733 are based directly on literature and832 are supported by genomic data, with an overlap of 570 reactions.Reactions not supported by literature or genomic data were includedto ensure connectivity of the different pathways. For central metabo-lism only 24 enzymes out of 785 (excluding transport) are not linkedto an annotated open reading frame (ORF, 3%), indicating anexcellent coverage of metabolic genes. An overview of the subset of785 unique reactions is presented in Supplementary Table 10 online.Examining the entire metabolic model, only 65 enzymes were notfound in the annotation (Supplementary Table 11 online). A moredetailed discussion on A. niger central metabolism and transportcharacteristics is presented in Supplementary Data, SupplementaryFigures 3 and 4 online.The extremely flexible metabolism and high nutritional versatility
of A. niger is confirmed by the presence of various solute-transporterclasses (Supplementary Table 12 online). Members of the majorfacilitator superfamily appear to be exceptionally abundant inA. niger with 461 genes, which is in the same range as A. oryzae(507), but greater than in A. fumigatus (275) and A. nidulans (358)(Supplementary Table 13 online). These proteins are predicted to beinvolved in the transport of a wide range of substrates and several mayfunction as nutrient sensors. Aspergillus species have also undergone asignificant expansion in the number of fungal zinc binuclear clusterproteins (PF00172)27 (Supplementary Table 14 online). This distri-bution is mirrored by a second PFAM domain (PF04082), which wasoriginally identified as occurring in a proportion of proteins withPF00172 domain. This is most dramatic in A. niger and is not reflectedin other transcription factor domains (Supplementary Table 15online). The characterized examples of these proteins are involvedin regulating diverse aspects of primary and secondary metabolismincluding polysaccharide degradation.Using the reconstructed metabolic network, we investigated why
A. niger is an efficient organic acid producer28. Several enzymesinvolved in the formation of the citrate precursor oxaloacetate werefound, for example, two pyruvate carboxylases (one cytoplasmic, onemitochondrial), four malate dehydrogenases (three cytoplasmic andone mitochondrial). In addition a mitochondrial oxaloacetate trans-porter was found (Fig. 3). A. niger contains one cytosolic and threeputative mitochondrial citrate synthases. Phylogenetic analysis of theA. niger citrate synthases in various aspergilli reveals two clusterscontaining one and three members, respectively (SupplementaryFig. 5). The second cluster may have originated from a duplicationof an ancestral citrate synthase that was lost in all organisms except insome fungi and plants. At least one additional gene duplication eventoccurred (An01g09940 and An08g10920). Similar redundancy wasfound for aconitase, for which two putative cytoplasmic and twomitochondrial forms were found. The various gene duplications maybe important in view of the efficient production of citrate by A. niger.Moreover, the necessary transport steps could be facilitated by twomitochondrial and several cytoplasmic membrane tricarboxylate
1 2 3 4 5 6 7 8SC9|1L SC13|2L SC12|3L SC7|4L SC10|5EL
SC17|5L
SC1|2R
SC14|1R
SC15|3R
SC2|4R
SC3|6R
SC8|8R
SC6|8ER
SC19|4ER
Chromosomeof origin:1 =2 =3 =4 =5 =6 =7 =8 =U =
detail*
SC16|5R*
SC11|7R
SC4|6L SC5|7L SC18|8LFigure 2 Aspergillus comparative genome maps. A. niger supercontigs
(SC1-SC19) are ordered from left to right by their proposed chromosomal
assignment (1 to 8) and are projected vertically, left arm (L) on top.
Syntenic regions between A. niger and A. nidulans, A. fumigatus and
A. oryzae are plotted in three separate columns on the A. niger
chromosomes. The colored areas refer to the chromosome assignment in the
compared aspergilli (linkage group of origin). Gray blocks are syntenic with
contigs for which this chromosome assignment is unknown (U). The zoom-inon SC16 displays the position of orthologs on the chromosome (black bars
at the right) and regions of reverse orientation (black bars at the left).
224 VOLUME 25 NUMBER 2 FEBRUARY 2007 NATURE BIOTECHNOLOGY
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy
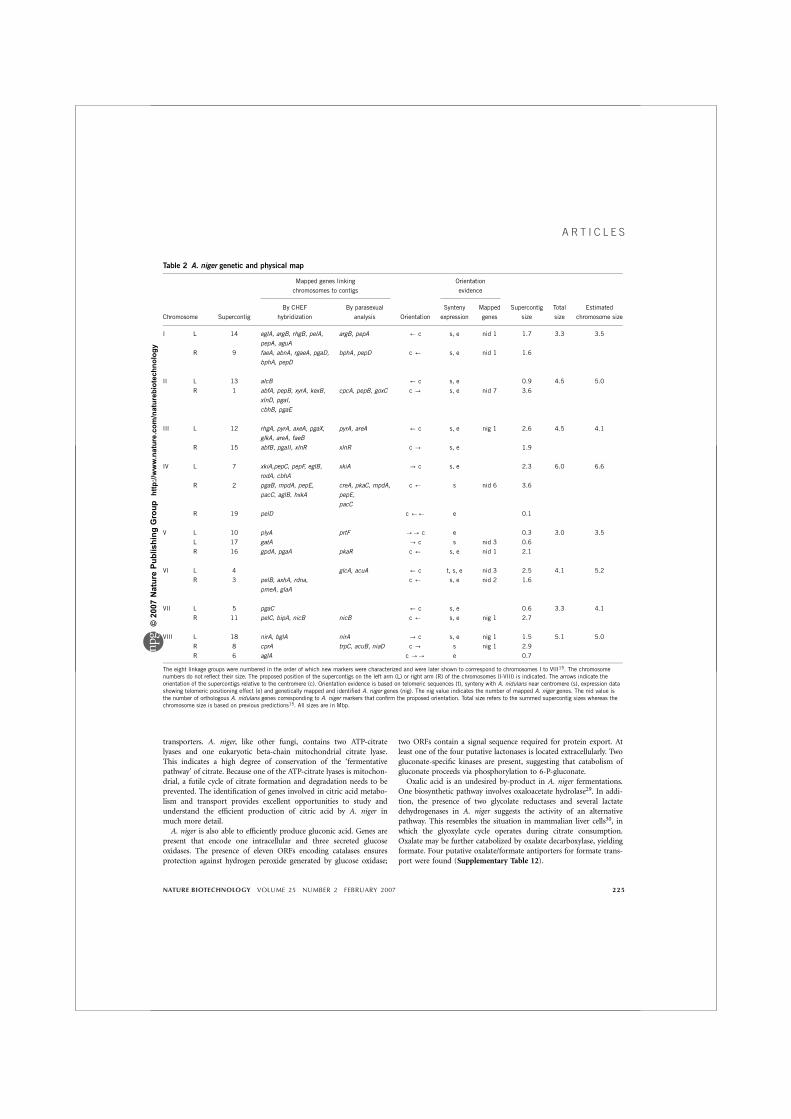
transporters. A. niger, like other fungi, contains two ATP-citratelyases and one eukaryotic beta-chain mitochondrial citrate lyase.This indicates a high degree of conservation of the ‘fermentativepathway’ of citrate. Because one of the ATP-citrate lyases is mitochon-drial, a futile cycle of citrate formation and degradation needs to beprevented. The identification of genes involved in citric acid metabo-lism and transport provides excellent opportunities to study andunderstand the efficient production of citric acid by A. niger inmuch more detail.A. niger is also able to efficiently produce gluconic acid. Genes are
present that encode one intracellular and three secreted glucoseoxidases. The presence of eleven ORFs encoding catalases ensuresprotection against hydrogen peroxide generated by glucose oxidase;
two ORFs contain a signal sequence required for protein export. Atleast one of the four putative lactonases is located extracellularly. Twogluconate-specific kinases are present, suggesting that catabolism ofgluconate proceeds via phosphorylation to 6-P-gluconate.Oxalic acid is an undesired by-product in A. niger fermentations.
One biosynthetic pathway involves oxaloacetate hydrolase29. In addi-tion, the presence of two glycolate reductases and several lactatedehydrogenases in A. niger suggests the activity of an alternativepathway. This resembles the situation in mammalian liver cells30, inwhich the glyoxylate cycle operates during citrate consumption.Oxalate may be further catabolized by oxalate decarboxylase, yieldingformate. Four putative oxalate/formate antiporters for formate trans-port were found (Supplementary Table 12).
Table 2 A. niger genetic and physical map
Mapped genes linking
chromosomes to contigs
Orientation
evidence
Chromosome Supercontig
By CHEF
hybridization
By parasexual
analysis Orientation
Synteny
expression
Mapped
genes
Supercontig
size
Total
size
Estimated
chromosome size
I L 14 eglA, argB, rhgB, pelA,
pepA, aguA
argB, pepA ’ c s, e nid 1 1.7 3.3 3.5
R 9 faeA, abnA, rgaeA, pgaD,
bphA, pepD
bphA, pepD c ’ s, e nid 1 1.6
II L 13 alcB ’ c s, e 0.9 4.5 5.0
R 1 abfA, pepB, xyrA, kexB,
xlnD, pgaI,
cbhB, pgaE
cpcA, pepB, goxC c - s, e nid 7 3.6
III L 12 rhgA, pyrA, axeA, pgaX,
glkA, areA, faeB
pyrA, areA ’ c s, e nig 1 2.6 4.5 4.1
R 15 abfB, pgaII, xlnR xlnR c - s, e 1.9
IV L 7 xkiA,pepC, pepF, eglB,
rodA, cbhA
xkiA - c s, e 2.3 6.0 6.6
R 2 pgaB, mpdA, pepE,
pacC, aglB, hxkA
creA, pkaC, mpdA,
pepE,
pacC
c ’ s nid 6 3.6
R 19 pelD c ’’ e 0.1
V L 10 plyA prtF -- c e 0.3 3.0 3.5
L 17 gatA - c s nid 3 0.6
R 16 gpdA, pgaA pkaR c ’ s, e nid 1 2.1
VI L 4 glcA, acuA ’ c t, s, e nid 3 2.5 4.1 5.2
R 3 pelB, axhA, rdna,
pmeA, glaA
c ’ s, e nid 2 1.6
VII L 5 pgaC ’ c s, e 0.6 3.3 4.1
R 11 pelC, bipA, nicB nicB c ’ s, e nig 1 2.7
VIII L 18 nirA, bglA nirA - c s, e nig 1 1.5 5.1 5.0
R 8 cprA trpC, acuB, niaD c - s nig 1 2.9
R 6 aglA c -- e 0.7
The eight linkage groups were numbered in the order of which new markers were characterized and were later shown to corrrespond to chromosomes I to VIII15. The chromosomenumbers do not reflect their size. The proposed position of the supercontigs on the left arm (L) or right arm (R) of the chromosomes (I-VIII) is indicated. The arrows indicate theorientation of the supercontigs relative to the centromere (c). Orientation evidence is based on telomeric sequences (t), synteny with A. nidulans near centromere (s), expression datashowing telomeric positioning effect (e) and genetically mapped and identified A. niger genes (nig). The nig value indicates the number of mapped A. niger genes. The nid value isthe number of orthologous A. nidulans genes corresponding to A. niger markers that confirm the proposed orientation. Total size refers to the summed supercontig sizes whereas thechromosome size is based on previous predictions15. All sizes are in Mbp.
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 225
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

Protein secretion in A. nigerFor reasons unknown, A. niger is a far more effective natural secretor ofproteins than the well-studied yeast S. cerevisiae31. In eukaryotesprotein secretion involves transport via endoplasmatic reticulum(ER), Golgi apparatus and vesicles to the cell membrane (Fig. 4 andSupplementary Table 16 online). Translocation from the cytoplasm tothe ER occurs in A. niger through the established signal recognitionparticle (SRP)-dependent and SRP-independent pathways. As in mam-mals, no ortholog of the essential yeast signal recognition and dockingprotein Srp21p was found. ER lumenal HSP70-type protein-foldingchaperones, including BipA an LhsA are present. In S. cerevisiae, Kar2p(equivalent of BipA in A. niger) functions together with the nucleotide-exchange factor Sil1p32. Surprisingly, no ortholog of yeast Sil1p can be
found in A. niger and other aspergilli, whereas a putative ortholog inNeurospora crassa could be detected. A. niger encodes three solublelumenal and one putative membrane-bound protein disulphideisomerase (PDI). Only PdiA (ortholog of yeast Pdi1p) and themembrane-associated EpsA (Eps1p) appear to be close orthologs ofknown yeast proteins. Finally, in contrast to several yeast species, plantsand mammals, the ER lumenal protein EroA of the sequenced aspergillicontains a predicted C-terminal ER-retention signal.The A. niger unfolded protein response (UPR) signaling pathway is
strikingly different from that in yeast31,33. In mammalian cells theprotein p58 interacts with and inhibits the ER-localized eIF2alphakinase PERK. This kinase is involved in translational regulation,attenuating the UPR during ER stress34. Orthologs of p58 andPERK are not found in yeast. The presence of a putative p58 orthologin A. niger is remarkable as no ortholog of PERK was found.Most components of the glycosylation machinery are readily
identified by sequence similarity. Although present in A. fumigatusand A. nidulans, no ortholog of yeast Alg14p was identified in A. niger.Interestingly, the dolichol-P-mannose synthase gene is more mamma-lian- than yeast-like as it does not contain a C-terminal hydrophobictransmembrane region. A. niger possesses a glycosylation-dependentquality-control system that is clearly distinct from that of S. cerevisiae.A. niger contains an ortholog of the protein-folding sensor UDP-Glc:glycoprotein glucosyltransferase absent in yeast. Calnexin is moresimilar to its counterpart in Schizosaccharomyces pombe.ER-associated protein degradation (ERAD) directs misfolded or
unassembled proteins to the proteasome35. A. niger lacks clearorthologs of the yeast ERAD proteins Cue1p, Rad23p, Ubx2p andYos9p and shows little homology to other ERAD components such asDer1p, Hrd1p, Doa10p and Hrd3p. Orthologs of all subunits of theyeast 26S proteasome are found, but orthologs of the regulatoryproteins Rpn13p and Rpn14p36 appear to have low similarity inA. niger or to be absent (Supplementary Data). The A. niger secretionsystem is equipped with a well-developed machinery for glycoprotein
Figure 3 Citrate biosynthesis and degradation.
The putative localization of enzymes and
transporters is indicated. Their symbols
are underlined when expression of their
corresponding A. niger genes (under noncitric
acid–producing conditions) was observed; the
numbers are in bold in this legend. The numbers
in parentheses next to the enzyme symbolsindicate the number of genes found for a specific
function whereas in the case of the pyruvate
dehydrogenase complex they indicate the total
number of genes involved in the complex.
cPYC, cytoplasmic pyruvate carboxylase
(An04g02090); cMDH, cytoplasmic malate
dehydrogenase (An11g07190, An12g00160,
An15g00070); cACO, cytoplasmic aconitase
(An 02g11040, An09g03870); cIDH,
NADP-specific isocitrate dehydrogenase
(An02g12430; note that cytoplasmic or
mitochondrial localization occurs by differential
translational start point utilization); cACL,
cytoplasmic ATP-citrate lyase (An11g00530);
PYT, putative pyruvate carrier, not identified because of lack of characterization in other organisms; OAT, oxaloacetate transporter (An14g06860); CMC,
citrate/malate carrier (An11g11230, An18g00070); PDH, pyruvate dehydrogenase complex E1-alpha subunit (An07g09530, An11g04550), E1-beta
subunit (An01g00100), E2 dihydrolipoamide acetyltransferase (An07g02180), E3 lipoamide dehydrogenase (An07g06840), E3 binding protein
(An02g11910); mPYC, mitochondrial pyruvate carboxylase (An15g02820); mMDH, mitochondrial malate dehydrogenase (An16g00120); mCL,
mitochondrial citrate lyase (An01g08610); AK, acetate kinase (An02g06420) and ACS, acetyl-CoA synthase (An05g00390); CS, citrate synthase(An01g09940, An08g10920, An09g06680); mACL, mitochondrial ATP-citrate lyase (An11g00510); mACO, mitochondrial aconitase (An08g10530,
An16g05760); mIDH, mitochondrial NADP- and NAD-isocitrate dehydrogenases (An02g12430, An08g05580, An18g06760).
Cytosol
PYT (0)
Citrate
Citrate
Isocitrate
IsocitrateFatty acids
Acetyl-CoA
2-ketoglutarate
2-ketoglutarate
Mitochondrion
Pyruvate
Oxaloacetate Oxaloacetate
Acetate
Acetyl-CoAACS (1)
AK (1)
Malate Malate
Pyruvate
cPYC (1)
CMC (2)cMDH (3) mMDH (1)
PDH (6)
mACL (1)
mACO (2)TC
A cycle
mIDH (3)
cIDH (1)
cACO (2)
cACL (1)CS (3)
CMC (2)
mCL (1)
OAT (1)mPYC (1)
anig
Protein misfolding47 47 47 4746 46
P
V EVesicle formation
and docking35 33 34 34 34 34
21
ER to Golgi andintra-Golgi transport
ER
N
Entry into ER18 18 17 15 16 20
17 17 17 17 17 15
39 38 39
Protein complexesinvolved in protein transport
38 38 40
19 18 19
Golgi and post-Golgiprotein sorting
18 18 19
119 97 101Glycosylation
G
101 90 66
Processes in ER
20 20 20 20 21
anid afum aory ncra sce
Figure 4 The secretion pathway of A. niger and other fungi. The number of
identified genes from A. niger (anig) and their putative orthologs in severalother fungi (anid, A. nidulans; afum, A. fumigatus; aory, A. oryzae; ncra,
N. crassa and sce, S. cerevisiae) are shown for various steps of the secretory
pathway. E, endosome; ER, endoplasmic reticulum; G, Golgi; P, proteasome;
V, vacuole; N, nucleus. See Supplementary Table 16 for details.
226 VOLUME 25 NUMBER 2 FEBRUARY 2007 NATURE BIOTECHNOLOGY
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

Table 3 Comparison of carbohydrate active enzymes in aspergilli
Enzyme CAZy family A. niger A. nidulans A. fumigatus A. oryzae
Glycosyl hydrolases
b-glucosidase GH1 3 3 5 3
b-galactosidase GH2 0 3 2 1
b-mannosidase GH2 3 3 2 3
b-glucuronidase GH2 3 3 1 2
Exo-b-d-glucosaminidase GH2 0 0 2 1
b-glucosidase GH3 12 15 12 16
b-xylosidase GH3 2 3 2 5
b-N-acetylhexosaminidase GH3 3 2 3 2
b-1,4-endoglucanase GH5 4 4 6 4
b-1,4-endomannanase GH5 1 6 2 2
Exo-1,3-b-glucanase GH5 4 3 2 3
Endo-1,6-b-glucosidase GH5 0 1 1 2
Endoglycoceramidase GH5 1 1 2 2
Cellobiohydrolase (non-reducing end) GH6 2 2 1 1
Cellobiohydrolase (reducing end) GH7 2 2 2 2
b-1,4-endoglucanase GH7 0 1 2 1
b-1,4-endoxylanase GH10 1 3 4 4
b-1,4-endoxylanase GH11 4 2 3 4
Xyloglucan-active b-1,4-endoglucanase GH12 3 1 4 4
a-amylase GH13 4 3 2 3
a-1,6-glucosidase GH13 2 2 5 5
a-1,3-glucan synthase GH13 5 2 3 3
GPI-anchored a-amylase-like GH13 3 3 3 2
Intracellular a-amylase-like GH13 2 1 1 2
Glycogen debranching enzyme GH13 2 2 1 2
Glucoamylase GH15 2 2 5 3
Endomannanase GH26 1 3 0 1
a-galactosidase GH27 5 3 5 3
Endopolygalacturonase GH28 7 3 4 4
Endorhamnogalacturonase GH28 6 1 2 6
Exopolygalacturonase GH28 4 3 3 2
Exorhamnogalacturonase GH28 3 1 2 4
Xylogalacturonan hydrolase GH28 1 1 1 2
a-l-fucosidase GH29 1 0 0 0
a-glucosidase GH31 7 9 6 10
a-xylosidase GH31 0 1 1 1
Endoinulinase GH32 1 0 1 0
Exoinulinase GH32 1 1 1 0
Invertase GH32 3 1 2 4
b-galactosidase GH35 5 4 5 6
a-galactosidase GH36 2 4 4 3
a-l-arabinofuranosidase GH43 2 3 3 3
Endoarabinanase GH43 4 4 6 6
b-xylosidase GH43 2 5 2 5
b-1,4-xylosidase/a-arabinofuranosidase GH43 1 1 4 3
Galactan 1,3-b-galactosidase GH43 1 2 1 3
b-1,4-endoglucanase GH45 0 1 1 0
a-l-arabinofuranosidase GH51 3 2 2 3
Endo-1,4-b-galactanase GH53 2 1 1 1
a-l-arabinofuranosidase GH54 1 1 1 1
b-1,4-endoglucanase GH61 7 9 7 8
Arabinoxylan arabinofuranohydrolase GH62 1 2 2 2
a-glucuronidase GH67 1 1 1 1
Endoglucanase GH74 1 2 2 0
a-l-rhamnosidase GH78 8 8 6 8
d-4,5 unsaturated glucuronyl hydrolase GH88 1 2 2 3
Exoarabinanase GH93 0 2 3 3
a-l-fucosidase GH95 2 2 2 3
Continued on next page.
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 227
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

quality-control, combined with an effective ERAD pathway, whichdiffers from that in yeast. In conclusion, the A. niger secretory systemshares some components with mammals and some with S. cerevisiae.
Biopolymer-degrading hydrolases
Aspergilli contain a wide spectrum of enzymes for polysaccharide,protein and lipid degradation. The two industrial aspergilli (A. nigerand A. oryzae) contain the highest percentage of extracellular enzymes.Cellulases, hemicellulases, pectinases, amylases, inulinases, lipases andproteases are used in a range of industrial applications. Here we focuson polysaccharide-degrading enzymes and proteases.Glycosyl hydrolase, lyase and esterase families involved in poly-
saccharide degradation in the aspergilli sequenced were identifiedusing the carbohydrate-active enzymes (CAZy) classification (http://www.cazy.org/, Table 3). A detailed overview of the A. niger enzymes ispresented in Supplementary Table 17 online. Specific differencesbetween the aspergilli were noticed. In contrast to the other threeaspergilli, A. niger contains only one GH10 and four GH11 endox-ylanases, a xylanase family liberating larger oligosaccharides. Forinulin degradation A. niger and A. fumigatus both contain invertasesand endo- and exoinulinases. However, A. nidulans does not haveendoinulinases, whereas A. oryzae appears to contain only invertases.This suggests that the latter two species degrade inulin extracellularly.For the degradation of pectin a large set of enzymes is required37.
A. niger has the largest set (21) of (GH28) enzymes, including sevencandidate endopolygalacturonases (Supplementary Table 17). Incontrast to the other aspergilli, A. niger has only a single pectatelyase (PL1). The pectinolytic machinery is completed by a singleputative delta-4,5-unsaturated glucuronyl hydrolase (GH88), twoputative delta-4,5-unsaturated rhamnogalacturonyl hydrolases(GH105), three putative pectin methylesterases (CE8) and one candi-date rhamnogalacturonan acetyl esterase (CE12) (Table 3). The singlepectate lyase is consistent with the acidifying properties of A. niger,given that pectate lyases show little activity at acidic pH. Moreover,A. niger does not contain exoarabinanases, indicating that A. niger
relies on the combined action of endoarabinanase and a-L-arabino-furanosidase for the hydrolysis of the arabinan side-chains of pectin.The glycoside hydrolase family GH13 involved in starch degrada-
tion is larger in aspergilli compared to other fungi. The GH13 familycontains three separate groups of amylase-type enzymes in A. nigerbased on phylogenetic clustering (Supplementary Fig. 6). A. nigercontains four putative extracellular a-amylases (SupplementaryTable 17). A second group consists of enzymes with relatively lowhomology to known alpha-amylases, which do not contain signalsequences. A third group consists of glycosyl phosphatidylinositol–anchored enzymes, which may play a role in the maintenance of thecell walls containing a-glucan38. The A. niger genome contains 42enzymes classified as CAZy family members without a signal sequence.The majority of these are likely to be involved in the degradation ofdisaccharides or glycosides imported by the fungus.A. niger has a full set of enzymes to degrade polypeptides (Supple-
mentary Table 18 online). Secreted proteases are applied in deter-gents, food applications and as biocatalysts in the production of finechemicals. The A. niger genome encodes 198 proteins involved inproteolytic degradation including a variety of secreted aspartylendoproteases (9), serine carboxypeptidases (10) and di- and tri-peptidylaminopeptidases (9). Compared to A. nidulans and A. oryzae,the number of putative secreted aminopeptidases (3) is low. Theabundance of aspartyl endoproteases and carboxypeptidases (mostlyactive at low pH) and the low number of aminopeptidases (mostlyactive at neutral or high pH) matches the acidifying properties ofA. niger as previously noted for A. oryzae8. Unlike the aminopeptidasesin the other sequenced aspergilli, seven aminopeptidases of A. niger arepredicted to be intracellular, indicating that at least part of the amino-terminal degradation of external proteins is taken care of by secreteddi- and tri-peptidylpeptidases.
Secondary metabolism and safety
Among the secondary metabolites produced by filamentous fungi,mycotoxins are most relevant from a safety point of view39. A. niger
Table 3 continued
Enzyme CAZy family A. niger A. nidulans A. fumigatus A. oryzae
Rhamnogalacturonyl hydrolase GH105 2 3 2 2
Polysaccharide lyases
Pectate lyase PL1 1 3 2 4
Pectin lyase PL1 5 5 4 8
Pectate lyase PL3 0 5 3 3
Rhamnogalacturonan lyase PL4 2 4 3 4
Pectate lyase PL9 0 1 1 1
Rhamnogalacturonan lyase PL11 0 1 0 0
Carbohydrate esterases
Acetyl xylan esterase CE1 1 1 2 1
Feruloyl esterase CE1 1 1 0 2
Unknown esterase CE1 1 1 2 2
Pectin methylesterase CE8 3 3 4 5
Rhamnogalacturonan acetylesterase CE12 2 2 2 4
Total 171 186 186 217
Secreted 131 (77%) 134 (72%) 124 (66%) 159 (73%)
Intracellular 40 (23%) 52 (28%) 62 (34%) 58 (27%)
ORFs are grouped by CAZy family and putative function based on the closest characterized homolog. GPI, glycosyl phosphatidylinositol.
228 VOLUME 25 NUMBER 2 FEBRUARY 2007 NATURE BIOTECHNOLOGY
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

contains several secondary metabolite clusters (SupplementaryTable 19 online). The genome contains 17 nonribosomal peptidesynthase (NRPS) and 34 polyketide synthase (PKS)-encoding genes,most of which are located in clusters. A. niger contains seven hybridPKS-NRPS synthases whereas other sequenced filamentous fungicontain a single hybrid PKS-NRPS. The sometimes unusual domainpatterns for these NRPS, PKS and hybrid genes are given inSupplementary Table 20 online. Of particular interest is thepresence of a gene cluster that shares a number of genes with theGibberella moniliformis gene cluster, which encodes the mycotoxinfumonisin (Supplementary Fig. 7)40. This cluster is absent from thegenomes of other sequenced aspergilli such as A. fumigatus, A. oryzaeand A. nidulans8–10.A putative ochratoxin cluster was identified on the basis of a PKS
fragment of A. ochraceus involved in ochratoxin biosynthesis41. SomeA. niger strains have been reported to synthesize this toxin but little isknown about the biosynthetic pathway42. In A. ochraceus, genesencoding chloroperoxidase, reductase, esterase, dehydratase andNRPS may be involved. Of these only an NRPS-like gene with theunusual domain structure ACPA is found in A. niger. Neither gene isexpressed at a detectable level (Supplementary Table 7).Siderophores are secondary metabolites involved in iron assimila-
tion and storage43 and protection against oxidative stress. One clearexample of a siderophore cluster was identified in the genome(Supplementary Table 19). The vast majority of the remainingnonribosomal peptide and polyketide synthases do not share ortho-logs with the other fungal genomes sequenced to date. In addition, noorthologs of genes for the biosynthesis of penicillin, prenylatedalkaloids (fumitremorgins), clavines, gliotoxin, aflatoxin or knownfungal terpenes (aristolochene and trichodiene) were found.
METHODSGenome sequencing and assembly. A library with B20.7� coverage of the
A. niger CBS 513.88 genome was constructed in pBeloBAC 11 and characterized
by end-sequencing and restriction digestion. Insert sizes of BAC clones ranged
from o50 to 4150 kb per clone. A total of 11,136 BAC clones were generated
with an average insert size of 68 kb. We generated 18,014 BAC end-sequences
with 504 bases average read length (phred20). The BAC end-sequencing success
rate was 80.9%. Using the BAC walking approach, we selected 595 BAC clones
for analysis, and 505 of those, representing the minimal tiling path, for shotgun
sequencing. The minimal tiling set of sequenced BACs was confirmed by the
high density map of BAC end-sequences (1 end per 2 kb). Repeated sequencing
of the estimated 100 rDNA copies was avoided by not sequencing BACs that
have end sequences matching rDNA. The 505 BACs selected have an average
insert size of 76.8 kb and have been sequenced with a 7.5� coverage. The
assembled BACs contained on average 2.14 contigs per BAC, corresponding
with an average of 1.14 sequence gaps per BAC. Shotgun sequences were
assembled with Phrap and edited after import into Gap4. BAC assemblies and
raw data were visualized and edited using the STADEN package. The genome
was assembled and logically joined using BAC clones physically bridging known
gaps to form supercontigs with a total unique sequence size of 33.9 Mb. The
estimated 5% of the genome not sequenced includes telomeric regions,
additional rDNA repeats and small gaps. Full details of the genome sequencing
and assembly can be found in Supplementary Methods online. CBS 513.88 can
be obtained from Centraal Bureau Schimmelcultures (http://www.cbs.knaw.nl/).
Annotation. Analysis and annotation of the genomic sequences of A. niger was
performed with a combined automatic and manual approach. ORFs were
predicted by a version of FGENESH44 trained on known A. niger and related
organism sequences as well as other gene prediction algorithms (Supplemen-
tary Methods). ORFs were named after the organism (An), supercontig
number (two digits) followed by g (gene) and a five-digit number matching
the order of the ORFs on the contig. tRNA genes were identified as described in
Supplementary Methods. For all ORFs identified, exhaustive automatic
bioinformatic analysis with respect to function and structure of the respective
protein was carried out using the PEDANT-ProTM software45. For each ORF
the automatically predicted functional features were manually verified and the
following features were manually annotated: gene model (based on Blast
comparisons with known proteins), title, functional categories according to
the MIPS functional catalog12 and EC numbers.
The manual annotation of the title involved (i) selection of a similar protein
through Blast comparison and (ii) a description of the degree of similarity with
this protein. If possible a similar protein with published experimental data was
selected. The description of the degree of similarity was assigned as follows:
questionable ORF, no Blast hit and questionable gene structure; no similarity,
no Blast hit; weak similarity, Blast e-value 4e-05; similarity, Blast e-value
oe-05 and 4e-20; strong similarity, Blast e-value oe-20; known protein,
identical to known A. niger gene.
Manual FunCat and EC number assignments were based on the described
function of the selected similar protein and took into account the degree of
similarity as illustrated by the following example. An A. niger protein matching
a well-known transcriptional activator was annotated in the manual FunCat as
follows: weak similarity, 04.05 mRNA transcription; similarity, 04.05.01 mRNA
synthesis; strong similarity, 04.05.01.04 transcriptional control; known protein,
04.05.01.04.01 transcriptional activator.
Phylogenetic analysis. Orthologous genes were grouped using all-against-all,
pairwise Blast similarity searches at the level of predicted proteins keeping
reciprocally best-matching genes. Twenty fungal panorthologous genes encod-
ing housekeeping functions and showing reciprocal best matches between all
selected organisms were selected (Supplementary Table 3) and the predicted
protein sequences were aligned with ClustalW46. Bidirectional Blast similarity
searching using the Blastx and tBlastn algorithms was applied to find additional
orthologs in the genome of A. clavatus. After deletion of ambiguously aligned
regions, the protein sequences were concatenated (7,767 amino acid sites).
Maximum likelihood phylogenetic analysis was performed with Tree Puzzle47
using the VTmodel48 and a gamma model of rate heterogeneity with alpha ¼0.65. Reliable bootstrap values were obtained for all nodes of the tree except for
the terminal nodes linking A. niger, A. oryzae and A. terreus, indicating their
close relationship.
For phylogenetic analysis of protein families, the protein sequences were
aligned first with ClustalW or ClustalX. Phylogenetic analyses were carried out
in MEGA 2.1 or 3.1 using Maximum Likelihood, Neighbor Joining and
Minimum Evolution49. Stability of clades was evaluated by 500 to 1,000
bootstrap rearrangements. Genome sequences used were obtained from the
National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/
Genomes/) except for A. clavatus (http://msc.tigr.org/aspergillus/aspergillus_
clavatus_nrrl_1/index.shtml) and A. terreus (http://www.broad.mit.edu/
annotation/genome/aspergillus_terreus/).
Ortholog detection and synteny analysis. To detect conservation of gene order,
we obtained pairwise lists of orthologs using bidirectional Blast searching of the
A. niger predicted coding sequence (CDS) and predicted CDS of each of
A. fumigatus, A. nidulans and A. oryzae. Sequences having a length r100
amino acids were omitted to get high confidence e-values. Bidirectional best hits
having an e-value o1e-10 in both directions were considered as orthologous
gene pairs. Conservation of synteny was determined by comparing adjacent
orthologous gene pairs allowing for inversions in gene order between species. If
there was a match, the two orthologous gene pairs were considered to be part of
a syntenous region. Such regions were extended by directional scanning along
the A. niger genome. Gene order and inversions were also recorded.
A. niger genetic and physical map. The genetic location of cloned genes was
established using pulsed field electrophoresis followed by Southern blot
analysis15. Parasexual analysis14 was used to determine the genetic location of
cpcA by linkage of the phleomycin resistance to hisD4. The methods used to
establish the physical map including supercontig orientation on chromosomes
are described in the Supplementary Data.
Fermentation and transcriptional profiling. A. niger was grown on defined
medium with glucose as a carbon source and ammonia as nitrogen source
using 20-liter submerged stirred (Rushton turbines) fermenters at controlled
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 229
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

pH of 4.5 at 35 1C. Glucose feeding started after 24 h. Biomass samples for
mRNA analysis were taken after 72 and 120 h. Sample treatment and
Affymetrix array (DSM proprietary GeneChips) analysis is described in
Supplementary Methods.
Reconstruction and analysis of the metabolic network. A comprehensive
literature search was carried out to identify reactions and/or enzymes that are
present in A. niger, and this compilation was supplemented by information
present in the enzyme databases BRENDA (http://www.brenda.uni-koeln.de/)
and SwissProt (http://www.expasy.org/). The KEGG database (http://www.
genome.ad.jp/kegg/metabolism.html) and the reconstructed metabolic network
of S. cerevisiae26 were used to fill gaps in incomplete pathways. Finally, the
information obtained through the annotation of all ORFs (see above) was
compared to this metabolic framework and as many ORFs as possible were
assigned to reactions. PSORT II (http://psort.ims.u-tokyo.ac.jp/form2.html)
and TargetP (http://www.cbs.dtu.dk/services/TargetP/) were used for analysis of
theoretical subcellular localization. Prediction of signal peptide cleavage sites
was carried out with SignalP 2.0 (http://www.cbs.dtu.dk/services/SignalP/).
Protein sequences were also analyzed with InterProScan (http://www.ebi.
ac.uk/InterProScan/).
Carbohydrate-active enzyme classification. The search for carbohydrate-active
enzymes was performed according to the routine update strategy of the CAZy
database (http://afmb.cnrs-mrs.fr/CAZY/). Sequences of the proteins in CAZy
were cut into their constitutive modules (catalytic modules, carbohydrate-
binding module (CBMs) and other noncatalytic modules) and the resulting
fragments were assembled in a sequence library for Blast searches50. Each
A. niger protein model was compared by Blast analysis against the library of
around 50,000 individual modules. Models with an e-value o0.1 were
manually analyzed to predict their function based on multiple sequence
alignment using ClustalW and a search for conserved signatures/motifs
characteristic of each family including the presence of the catalytic machinery.
SignalP (http://www.cbs.dtu.dk/services/SignalP/) was used to detect the pre-
sence of possible signal sequences.
Genome data and availability. The annotated genome has been deposited at
the EMBL database with the accession numbers AM270980–AM270998 (DNA
supercontigs) and AM269948–AM270415 (DNA contigs).
Requests for materials. A scalable vector graphic that plots all syntenous
regions on the A. niger chromosomes, including relative orientation of the
conserved synteny and blocks representing each gene, is available on request
from H.J.K. ([email protected]).
Note: Supplementary information is available on the Nature Biotechnology website.
ACKNOWLEDGMENTSE.G.J.D., P.M.C., B.H. and F.M.K. wish to acknowledge the financial supportfrom the European Commission (STREP FungWall grant, contract:LSHB-CT-2004-511952) and the French Ministry of Research (program ACI-BCMS, Enzywall). The work of X.L, U.R., J.S. and A.-P.Z. was partlyfunded by the Sonderforschungsbereich 578 (SFB578) of the DeutscheForschungsgemeinschaft, Germany. We acknowledge the Department of Energy,Joint Genome Institute, TIGR and the Broad Institute for allowing somecomparative genome analysis. Part of this work was supported by SENTER(BTS project BTS00010, TSGE 3012). Array hybridizations were performedat the MicroArray Department (MAD) in Amsterdam. Ulrike Jacobi isacknowledged for assistance in transcriptome analysis. We thank Bea denDekker for excellent organizational support.
AUTHOR CONTRIBUTIONSH.J.P. coordinated the A. niger sequencing and annotation work. J.H.de W.contributed to the microarray analysis, and metabolism and developmentalaspects. R.A. and K.A were coordinators of the Biomax annotation team.M.v.d.B. and A.v.D. were members of the DSM sequencing team. J.L. and H.W.at Qiagen developed the sequencing strategy and coordinated the sequencing.N.N.M.E.v.P. was responsible for the microarray work. R.M. and S.B. wereresponsible for the fermentation. H.M. was involved in transcriptome analysis.G.S.P.G. contributed to the manual annotation. J.A.R. was responsible for theDSM bioinformatics work. M.O. contributed to protein analysis andcharacterization. P.J.S., R.T.J.M.v.d.H. and H.J.K. worked out the phylogeny andsynteny between the aspergilli. P.J.I.v.d.V. and A.J.M.De. compared the physical
and genetic map. P.S.D. and H.A.B.W. investigated life cycle and reproductionwhereas K.P., J.Va. and A.J.M.De. looked for incompatibility genes. Cell wallfunctions were analyzed by A.F.J.R., F.M.K., P.W.J.de G. and H.A.B.W.Transcription factors were analyzed by M.X.C., A.M.M., S.G.O. and M.C.J.D.B. and D.U. analyzed the A. niger secretome. Analysis of the metabolism wascarried out by G.H. (coordination), M.R.A., J.N., C.d’E., C.P.K., M.S. and J.Vi.,whereas A.J.M.Dr. and P.A.v.K. analysed the genome for transport functions.D.B.A. coordinated the secretion team, which further included T.G.,C.A.M.J.J.v.d.H., X.L., N.M.E.v.P., A.F.J.R., U.R., C.M.J.S., J.S. and A.-P.Z. Theglycosylation aspects were covered by R.C., S.G. and W.V. The general analysisof the extracellular enzyme spectrum was performed by R.P.deV. (coordination),P.M.C., E.G.J.D., M.H. and B.H., whereas J.A.E.B. (pectinases), P.D. (proteases),R.M.v.d.K., M.J.E.C.v.d.M., C.G. and L.D. (amylases) covered specific areas.G.T., M.C. and P.W.M.v.D. analysed secondary metabolism and safety aspects.J.P.T.W.v.d.H. and A.J.J.v.O initiated the DSM sequencing project. H.J.P.,J.H.d.W., J.Vi. and H.S. (project leader) wrote the final text of the manuscript.
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Published online at http://www.nature.com/naturebiotechnology/
Reprints and permissions information is available online at http://npg.nature.com/
reprintsandpermissions
1. Pandey, A., Selvakumar, P., Soccol, C.R. & Nigam, P. Solid state fermentation for theproduction of industrial enzymes. Curr. Sci. 77, 149–162 (1999).
2. Berka, R.M., Dunn-Coleman, N. & Ward, M. Industrial enzymes from Aspergillusspecies. in Aspergillus Biology and Industrial Applications (eds. Bennett, J.W. &Klich, M.A.) 155–202, (Butterworth–Heinemann, Stoneham, MA, 1992).
3. Schuster, E., Dunn-Coleman, N., Frisvad, J.C. & van Dijck, P.W. On the safety ofAspergillus niger–a review. Appl. Microbiol. Biotechnol. 59, 426–435 (2002).
4. Punt, P.J. et al. Filamentous fungi as cell factories for heterologous protein production.Trends Biotechnol. 20, 200–206 (2002).
5. Archer, D.B. & Turner, G. Genomics of protein secretion and hyphal growth inAspergillus. in The Mycota XIII (ed. Brown, A.J.P.) 75–96, (Springer, Berlin Heidelberg,2006).
6. Ruijter, G.J.G., Kubicek, C.P. & Visser, J. Production of organic acids by fungi. in TheMycota X Industrial Applications (ed. Osiewacz, H.D.) 213–230, (Springer, BerlinHeidelberg, 2002).
7. Raj, H.G., Saxena, M. & Allameh, A. Metabolism of foreign compounds by fungi. inHandbook of Applied Mycology vol. 4: Fungal Biotechnology (eds. Arora, D.K., Elander,R.P. & Mukerji, K.G.) 881–904, (Marcel Dekker Inc., New York, 1992).
8. Machida, M. et al. Genome sequencing and analysis of Aspergillus oryzae. Nature 438,1157–1161 (2005).
9. Galagan, J.E. et al. Sequencing of Aspergillus nidulans and comparative analysis withA. fumigatus and A. oryzae. Nature 438, 1105–1115 (2005).
10.Nierman, W.C. et al. Genomic sequence of the pathogenic and allergenic filamentousfungus Aspergillus fumigatus. Nature 438, 1151–1156 (2005).
11.Nakamura, Y., Gojobori, T.S. & Ikemura, T. Codon usage tabulated from the interna-tional DNA sequence databases: status for the year 2000. Nucleic Acids Res. 28, 292(2000).
12.Ruepp, A. et al. The FunCat, a functional annotation scheme for systematic classifica-tion of proteins from whole genomes. Nucleic Acids Res. 32, 5539–5545 (2004).
13. Samson, R.A. Current taxonomic schemes of the genus Aspergillus and its teleo-morphs. in Aspergillus Biology and Industrial Applications (eds. Bennett, J.W. & Klich,M.A.) 355–390, (Butterworth-Heinemann, Stoneham, MA, 1992).
14.Debets, F., Swart, K., Hoekstra, R.F. & Bos, C.J. Genetic maps of eight linkage groups ofAspergillus niger based on mitotic mapping. Curr. Genet. 23, 47–53 (1993).
15. Verdoes, J.C. et al. The complete karyotype of Aspergillus niger: the use of introducedelectrophoretic mobility variation of chromosomes for gene assignment studies. Mol.Gen. Genet. 244, 75–80 (1994).
16. Adams, T.H., Wieser, J.K. & Yu, J.H. Asexual sporulation in Aspergillus nidulans.Microbiol. Mol. Biol. Rev. 62, 35–54 (1998).
17.Geiser, D.M., Timberlake, W.E. & Arnold, M.L. Loss of meiosis in Aspergillus.Mol. Biol.Evol. 13, 809–817 (1996).
18. Paoletti, M. et al. Evidence for sexuality in the opportunistic fungal pathogenAspergillus fumigatus. Curr. Biol. 15, 1242–1248 (2005).
19.Bernard, M. & Latge, J.P. Aspergillus fumigatus cell wall: composition and biosynth-esis. Med. Mycol. 1 Suppl 9–17 (2001).
20. Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol.Biol. Rev. 69, 262–291 (2005).
21.Wosten, H.A.B. Hydrophobins: multipurpose proteins. Annu. Rev. Microbiol. 55,625–646 (2001).
22. Askolin, S., Penttila, M., Wosten, H.A.B. & Nakari-Setala, T. The Trichoderma reeseihydrophobin genes hfb1 and hfb2 have diverse functions in fungal development. FEMSMicrobiol. Lett. 253, 281–288 (2005).
23. Emri, T., Molnar, Z., Pusztahelyi, T., Varecza, Z. & Pocsi, I. The FluG-BrlA pathwaycontributes to the initiation of autolysis in submerged Aspergillus nidulans cultures.Mycol. Res. 109, 757–763 (2005).
230 VOLUME 25 NUMBER 2 FEBRUARY 2007 NATURE BIOTECHNOLOGY
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy

24.White, S., McIntyre, M., Berry, D.R. & McNeil, B. The autolysis of industrial filamen-tous fungi. Crit. Rev. Biotechnol. 22, 1–14 (2002).
25.David, H., Akesson, M. & Nielsen, J. Reconstruction of the central metabolism ofAspergillus niger. Eur. J. Biochem. 270, 4243–4253 (2003).
26. Forster, J., Famili, I., Fu, P., Palsson, B.O. & Nielsen, J. Genome-scale reconstructionof the Saccharomyces cerevisiae metabolic network. Genome Res. 13, 244–253(2003).
27. Todd, R.B. & Andrianopoulos, A. Evolution of a fungal regulatory gene family: the Zn(II)Cys6 binuclear cluster DNA binding motif. Fungal Genet. Biol. 21, 388–405(1997).
28.Karaffa, L. & Kubicek, C.P. Aspergillus niger citric acid accumulation: do weunderstand this well working black box. Appl. Microbiol. Biotechnol. 61, 189–196(2003).
29.Pedersen, H., Hjort, C. & Nielsen, J. Cloning and characterization of oah, the geneencoding oxaloacetate hydrolase in Aspergillus niger. Mol. Gen. Genet. 263, 281–286(2000).
30.Poore, R.E., Hurst, C.H., Assimos, D.G. & Holmes, R.P. Pathways of hepaticoxalate synthesis and their regulation. Am. J. Physiol. 272, C289–C294(1997).
31.Schroder, M. & Kaufman, R.J. ER stress and the unfolded protein response. Mutat.Res. 569, 29–63 (2005).
32.Steel, G.J., Fullerton, D.M., Tyson, J.R. & Stirling, C.J. Coordinated activation of Hsp70chaperones. Science 303, 98–101 (2004).
33. Travers, K.J. et al. Functional and genomic analyses reveal an essential coordinationbetween the unfolded protein response and ER-associated degradation. Cell 101,249–258 (2000).
34. van Huizen, R., Martindale, J.L., Gorospe, M. & Holbrook, N.J. P58IPK, a novelendoplasmatic reticulum stress-inducible protein and potential negative regulator ofEIF2alpha signalling. J. Biol. Chem. 278, 15558–15564 (2003).
35.Romisch, K. Endoplasmatic reticulum associated degradation. Annu. Rev. Cell Dev.Biol. 21, 435–456 (2005).
36. Verma, R. et al. Proteasomal proteomics: identification of nucleotide-sensitive protea-some-interacting proteins by mass spectrometric analysis of affinity-purified protea-somes. Mol. Biol. Cell 11, 3425–3439 (2000).
37. de Vries, R.P., Benen, J.A.E., de Graaff, L.H. & Visser, J. Plant cell wall degradingenzymes produced by Aspergillus. in The Mycota X Industrial Applications (ed.Osiewacz, H.D.) 263–279, (Springer, Berlin Heidelberg, 2002).
38. Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 150,2029–2035 (2004).
39.Keller, N.P., Turner, G. & Bennett, J.W. Fungal secondary metabolism – from biochem-istry to genomics. Nat. Rev. Microbiol. 3, 937–947 (2005).
40. Proctor, R.H., Brown, D.W., Plattner, R.D. & Desjardins, A.E. Co-expression of 15contiguous genes delineates a fumonisin biosynthetic gene cluster in Gibberellamoniliformis. Fungal Genet. Biol. 38, 237–249 (2003).
41.O’Callaghan. Caddick, .X. & Dobson, D.W. A polyketide synthase gene required forochratoxin A biosynthesis in Aspergillus ochraceus. Microbiology 149, 3485–3491(2003).
42.Moss, M.O. Recent studies of mycotoxins. J. Applied Microbiol. Symp. Suppl. 84,62S–76S (1998).
43.Reiber, K. et al. The expression of selected non-ribosomal peptide synthetases inAspergillus fumigatus is controlled by the availability of free iron. FEMSMicrobiol. Lett.248, 83–91 (2005).
44. Salamov, A.A. & Solovyev, V.V. Ab initio gene finding in Drosophila genomic DNA.Genome Res. 10, 516–522 (2000).
45. Frishman, D. et al. Functional and structural genomics using PEDANT. Bioinformatics17, 44–57 (2001).
46. Thompson, J.D., Plewniak, F., Thierry, J. & Poch, O. DbClustal: rapid and reliable globalmultiple alignments of protein sequences detected by database searches. NucleicAcids Res. 28, 2919–2926 (2000).
47. Schmidt, H.A., Strimmer, K., Vingron, M. & von Haeseler, A. TREE-PUZZLE: maximumlikelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics18, 502–504 (2002).
48.Muller, T. & Vingron, M. Modeling amino acid replacement. J. Comput. Biol. 7,761–776 (2000).
49.Kumar, S., Tamura, K. & Nei, M. MEGA3: Integrated software for Molecular EvolutionaryGenetics Analysis and Sequence Alignment. Brief. Bioinform. 5, 150–163 (2004).
50. Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. Basic local alignmentsearch tool. J. Mol. Biol. 215, 403–410 (1990).
NATURE BIOTECHNOLOGY VOLUME 25 NUMBER 2 FEBRUARY 2007 231
A R T I C L E S
©20
07 N
atur
e Pu
blis
hing
Gro
up h
ttp://
ww
w.n
atur
e.co
m/n
atur
ebio
tech
nolo
gy




















