
Delayed Recruitment of Immunocompetent Cells in Denervated Rat Periodontal Ligament Following...
8
http://jdr.sagepub.com/ Journal of Dental Research http://jdr.sagepub.com/content/78/6/1214 The online version of this article can be found at: DOI: 10.1177/00220345990780060401 1999 78: 1214 J DENT RES V. Vandevska-Radunovic, S. Kvinnsland and R. Jonsson Following Experimental Tooth Movement Delayed Recruitment of Immunocompetent Cells in Denervated Rat Periodontal Ligament Published by: http://www.sagepublications.com On behalf of: International and American Associations for Dental Research can be found at: Journal of Dental Research Additional services and information for http://jdr.sagepub.com/cgi/alerts Email Alerts: http://jdr.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: http://jdr.sagepub.com/content/78/6/1214.refs.html Citations: by guest on July 14, 2011 For personal use only. No other uses without permission. jdr.sagepub.com Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Delayed Recruitment of Immunocompetent Cells in Denervated Rat Periodontal Ligament Following...

http://jdr.sagepub.com/Journal of Dental Research
http://jdr.sagepub.com/content/78/6/1214The online version of this article can be found at:
DOI: 10.1177/00220345990780060401
1999 78: 1214J DENT RESV. Vandevska-Radunovic, S. Kvinnsland and R. Jonsson
Following Experimental Tooth MovementDelayed Recruitment of Immunocompetent Cells in Denervated Rat Periodontal Ligament
Published by:
http://www.sagepublications.com
On behalf of:
International and American Associations for Dental Research
can be found at:Journal of Dental ResearchAdditional services and information for
http://jdr.sagepub.com/cgi/alertsEmail Alerts:
http://jdr.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://jdr.sagepub.com/content/78/6/1214.refs.htmlCitations:
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

J Dent Res 78(6): 1214-1220, June, 1999
Delayed Recruitment of ImmunocompetentCells in Denervated Rat Periodontal LigamentFollowing Experimental Tooth MovementV. Vandevska-Radunovicl*, S. Kvinnsland1, and R. Jonsson2
lDepartment of Anatomy and Cell Biology, University of Bergen, Arstadveien 19, 5009 Bergen, Norway; and 2Broegelmann ResearchLaboratory, University of Bergen, Bergen, Norway; *corresponding author
Abstract. It has previously been shown that the number ofmononuclear phagocytic cells in the periodontal ligament(PDL) of orthodontically moved rat molars is significantlyincreased (p < 0.05) at 3, 7, and 14 days compared with thecontrols. Since these changes coincide with increased den-sity of peptidergic nerve fibers, it was of particular interestto investigate a possible relation between the immunocom-petent cells and sensory nerve fibers in the PDL of experi-mentally moved and denervated rat molars. Twenty-twoyoung animals had the first right mandibular molar movedmesially, 7, 14, and 21 days after ipsilateral inferior alveolarnerve axotomy. The left side served as unoperated control.An immunohistochemical procedure was carried out onalternate, serial, cryostat sections with antibodies againstCD11b (macrophages, dendritic cells) and class II major his-tocompatibility complex (MHC) molecules (RT1B). At 7 and14 days, the number of CDllb+- and RT1B-expressing cellsin the denervated PDL showed no significant differencecompared with the contralateral side. However, at 21 days,when periodontal tissue re-innervation is established, thenumber of the investigated immunocompetent cells in thePDL of the denervated and experimentally moved mandibu-lar molars demonstrated a significant difference comparedwith the contralateral and control molars (p < 0.05). It can beconcluded that axotomy of the inferior alveolar nerve delaysthe recruitment of macrophage-like and class II MHC mole-cule-expressing cells in the PDL of orthodontically movedrat molars. The results further indicate that sensory nervefibers interact with immunocompetent cells and participatein their mobilization to locally inflamed tissues.
Key words: dendritic cells, denervation, macrophages,periodontal ligament, orthodontic tooth movement.
Received November 19, 1997; Last Revision August 20, 1998;Accepted September 18,1998
Introduction
The immune and neuroendocrine systems represent anintegrated circuit that results from the sharing of ligandsand their receptors (Blalock, 1994). There is accumulatingevidence that immunocompetent cells can produce hor-mones and neuropeptides (Przewlocki et al., 1992; Maggianoet al., 1994), whereas neurons and endocrine cells are able toproduce various cytokines (Cunningham and De Souza,1993; Jafarian-Tehrani et al., 1995). In turn, these moleculesmediate the immune-neuroendocrine interplay in autocrineand paracrine fashion (Blalock, 1994).
Interactions, particularly between nerve and immuno-competent cells, can have a regulatory role in the process ofconnective and bone tissue remodeling during orthodontictooth movement (Davidovitch, 1991). Teeth subjected tomechanical forces demonstrate increased periodontal levelsof various cytokines (Davidovitch et al., 1988; Saito et al.,1991), increased recruitment of mononuclear phagocyticcells (Jager et al., 1993), as well as increased sprouting ofnerve fibers immunoreactive to neuropeptides such as calci-tonin gene-related peptide (CGRP) and substance P (SP)(Kvinnsland and Kvinnsland, 1990; Nicolay et al., 1990;Vandevska-Radunovic et al., 1997a). These changes co-local-ize and coincide with alterations in blood vessel density andenhanced blood flow (Vandevska-Radunovic et al., 1994), allbeing part of a comprehensive inflammatory dental tissuereaction.
Periodontal and pulpal inflammatory responses incidentto experimental tooth movement may well be controlled byneurogenic mechanisms. Neuropeptides are shown to initi-ate and control vascular events in various tissues that aretypical for inflammation (Mayer et al., 1988; Smith et al.,1993; Olgart, 1996). However, deprivation of sensory tissueinnervation either abolishes or suspends the development ofan inflammatory reaction (Gyorfi et al., 1994; Vandevska-Radunovic et al., 1998). In addition, characterization of spe-cific neuropeptide receptors on immunocompetent cells(Umeda and Arisawa, 1989; Abello et al., 1991) undoubtedly
1214 by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Immunocompetent Cells
connects the nervous and immune systems, suggesting thatneuropeptides have biologically relevant effects on theimmunological function.
In a previous experiment conducted on maxillary ratmolars, we have shown that experimental tooth movementinduces increased periodontal recruitment of cells belongingto the mononuclear phagocytic system, but has no signifi-cant effect on lymphocytes and granulocytes (Vandevska-Radunovic et al., 1997b). Macrophage-like cells on theexperimental side were significantly increased at 3 (74%), 7(76%), and 14 (59%) days, while cells expressing class II mol-ecules demonstrated significant increase at 3 (49%) and 7(34%) days compared with the controls. Both cell typesshowed a tendency to approach the control values at the lastexperimental period of 14 days (Vandevska-Radunovic et al.,1997b). These cellular changes coincide and co-localize withchanges in density and distribution of peptidergic nervefibers (Vandevska-Radunovic et al., 1997a). Therefore, theaim of this study was to investigate a possible interactionbetween the sensory nerve fibers and the mononuclearphagocytic cells in the periodontal ligament (PDL) of ratmolars in conjunction with experimental tooth movement.Since denervation of the maxillary rat molars requires exten-sive surgical procedure and tissue damage, we have chosento conduct the experiment in the lower jaw. For that pur-pose, the inferior alveolar nerve was axotomized, and itseffect on the recruitment of macrophages and cells express-ing class II major histocompatibility complex (MHC) mole-cules was quantitatively evaluated.
Materials and methods
AnimalsTwenty-two Mol:WIST male rats, approximately 200 g in weight,were used in this study. The experimental group was comprisedof 18 animals, while 4 animals served as an unoperated controlgroup. The rats were housed in polycarbonate cages in a conven-tional animal room and fed standard pellet diet with water adlibitum. All experiments were registered and approved by theNorwegian Experimental Animal Board (NEAB).
Experimental procedureThe operations were carried out while the animals were undergeneral anesthesia, for which purpose a subcutaneous injectionof fentanyl/fluanizon midazolam (Hypnorm Dormicum), 0.2mL/100 g body weight, was used. An orthodontic appliance,consisting of a coiled spring ligated to the first right mandibularmolar and connected to the mandibular incisors, was insertedinto the animals of the experimental group. The coil exerted aforce of approximately 0.5 N. Throughout the experimental peri-ods, the force attenuated, and by day 14 the coil was inactive.Immediately after the insertion of the appliance, unilateral axot-omy of the right inferior alveolar nerve was done as earlierdescribed (Vandevska-Radunovic et al., 1998). A horizontal skinincision was made on the lateral aspects of the mandible. Themasseter muscle was bluntly dissected, thus providing access to
the bone surface right above the mandibular foramen. The over-
J Dent Res 78(6) 1999
for each tooth, so that the mean value could be obtained. Allstructures that demonstrated evident brown staining, and only
in Denervated PDL 1215
lying bone was removed with a dental bur, and the exposedalveolar nerve was elevated and then sectioned with scissors,leaving the blood vessels intact. The incision was closed by threesutures (Supramid, 5/0, Resorba, Niirnberg, Germany), and theanimals were allowed to recover. The left contralateral sideserved as an intragroup control. After the experimental proce-dures, the animals received daily doses of analgesic for two tothree days (Temgesis/Buprenorphinum, Reckitt & Colman,Hull, England, 0.1 mL/100 g, twice a day) and were fed a softdiet. All animals were weighed before the start of the operationand prior to their death. Recovery was uneventful, and all ratsshowed an increase in their body weight, from 5% in the seven-day group to 32% in the 21-day group.
Seven (n = 6), 14 (n = 6), and 21 days (n = 6) after the opera-tion, the animals were re-anaesthetized with an overdose ofHypnorm Dormicum and transcardially perfused withheparinized phosphate buffer, followed by 10% EDTA, contain-ing 7.5% polyvinylpyrrolidone (PVP; Aldrich Chemicals,Steinheim, Germany). The jaws were excised and demineralizedat 4°C for 4 wks in 10% EDTA containing PVP (Jonsson et al.,1986). The demineralized sections were then frozen in methyl-butane, pre-chilled with liquid nitrogen and kept at -80°C untilsectioned sagitally at 10 to 12 ptm in a cryostat.
ImmunohistochemistryThe immunohistochemical procedure has been previouslydescribed (Vandevska-Radunovic et al., 1997). In short, the sec-tions were fixed in acetone for 5 min, washed in Tris-bufferedsaline (TBS), and treated with 0.3% H202 for 10 min. Thereafter,20% normal horse serum and 4% bovine serum albumin (BSA;Sigma, St. Louis, MO, USA) in TBS were used to incubate thesections for 20 min. Alternate serial sections were then incubatedfor 60 min in a humidity chamber with the following mono-clonal antibodies (Table 1): OX6 (class II MHC molecules; kindlydonated by Dr. Rikard Holmdahl, University of Lund, Sweden)and OX42 (CD11b; Seralab, Crawley Down, Sussex, UK). Afterbeing washed in TBS, the sections were incubated in biotinylatedhorse anti-mouse Ig, rinsed in TBS, and treated with avidin-biotin-peroxidase complex (DAKO, Glostrup, Denmark) for 30min. Antigen-antibody complexes were visualized for 15 min bythe use of H202 and 3-amino-9-ethyl carbazole (AldrichChemicals, Steinheim, Germany) containing buffer. Finally, thesections were counterstained with Mayer's hematoxylin andembedded in an aqueous mounting medium (Immu-mount,Shandon, Pittsburgh, PA, USA). We tested the specificity of theimmune reaction routinely by omitting the primary antibodyand substituting it with phosphate-buffered saline (PBS). Ratspleen tissue sections served as a positive control for theimmunoreactive cells.
Evaluation and statistical proceduresFor each monoclonal antibody, four sections from the centralpart of an experimental, contralateral, and control first mandibu-lar molar were used for evaluation. The labeled cells were
counted in a light microscope (2.5 x 10) throughout the PDL,summarized, and divided by the number of evaluated sections
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from
1216 Vandevska-Radunovic et al.
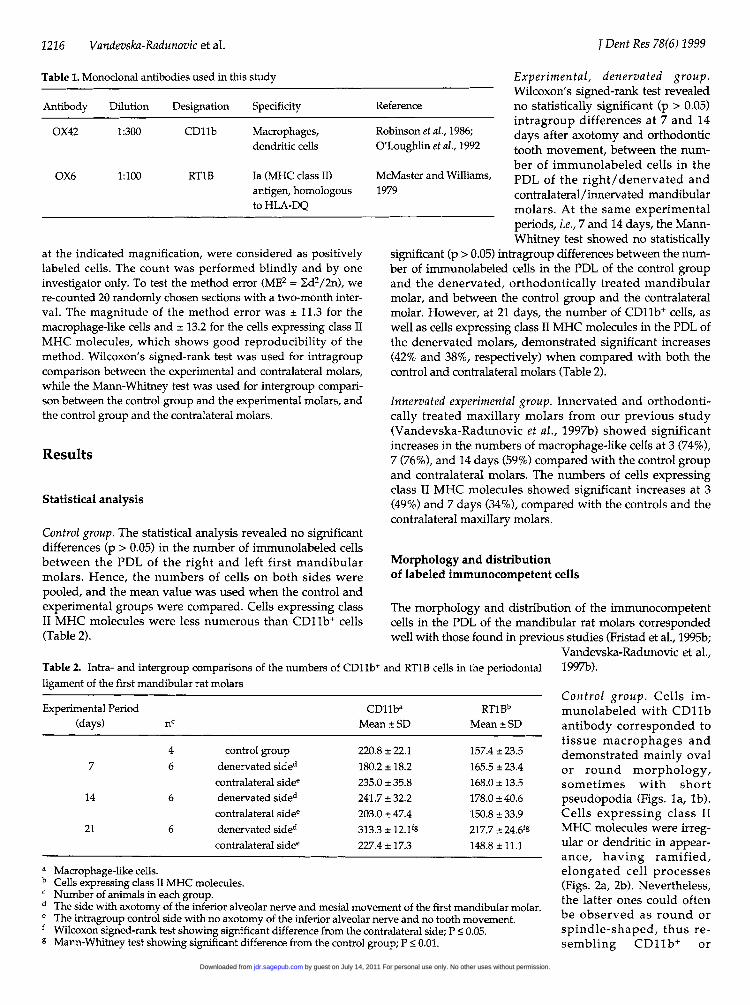
Table 1. Monoclonal antibodies used in this study
Antibody Dilution Designation Specificity
OX42 1:300 CD11b Macrophages,dendritic cells
OX6 1:100 RT1B Ia (MHC class II)antigen, homologousto HLA-DQ
at the indicated magnification, were considered as positivelylabeled cells. The count was performed blindly and by oneinvestigator only. To test the method error (ME2 = Ed2/2n), were-counted 20 randomly chosen sections with a two-month inter-val. The magnitude of the method error was ± 11.3 for themacrophage-like cells and ± 13.2 for the cells expressing class IIMHC molecules, which shows good reproducibility of themethod. Wilcoxon's signed-rank test was used for intragroupcomparison between the experimental and contralateral molars,while the Mann-Whitney test was used for intergroup compari-son between the control group and the experimental molars, andthe control group and the contralateral molars.
Results
Statistical analysis
Control group. The statistical analysis revealed no significantdifferences (p > 0.05) in the number of immunolabeled cellsbetween the PDL of the right and left first mandibularmolars. Hence, the numbers of cells on both sides werepooled, and the mean value was used when the control andexperimental groups were compared. Cells expressing classII MHC molecules were less numerous than CDlb+ cells(Table 2).
Table 2. Intra- and intergroup comparisons of the numbers of CD11Iligament of the first mandibular rat molars
Experimental, denervated group.Wilcoxon's signed-rank test revealed
Reference no statistically significant (p > 0.05)intragroup differences at 7 and 14
Robinson et al., 1986; days after axotomy and orthodonticO'Loughlin et al., 1992 tooth movement, between the num-
ber of immunolabeled cells in theMcMaster and Williams, PDL of the right/denervated and1979 contralateral/innervated mandibular
molars. At the same experimentalperiods, i.e., 7 and 14 days, the Mann-Whitney test showed no statistically
significant (p > 0.05) intragroup differences between the num-ber of immunolabeled cells in the PDL of the control groupand the denervated, orthodontically treated mandibularmolar, and between the control group and the contralateralmolar. However, at 21 days, the number of CD11b+ cells, aswell as cells expressing class II MHC molecules in the PDL ofthe denervated molars, demonstrated significant increases(42% and 38%, respectively) when compared with both thecontrol and contralateral molars (Table 2).
Innervated experimental group. Innervated and orthodonti-cally treated maxillary molars from our previous study(Vandevska-Radunovic et al., 1997b) showed significantincreases in the numbers of macrophage-like cells at 3 (74%),7 (76%), and 14 days (59%) compared with the control groupand contralateral molars. The numbers of cells expressingclass II MHC molecules showed significant increases at 3(49%) and 7 days (34%), compared with the controls and thecontralateral maxillary molars.
Morphology and distributionof labeled immunocompetent cells
The morphology and distribution of the immunocompetentcells in the PDL of the mandibular rat molars correspondedwell with those found in previous studies (Fristad et al., 1995b;
Vandevska-Radunovic et al.,+ and RT1B cells in the periodontal 1997b).
Experimental Period CD11ba RTlBb(days) nc Mean ± SD Mean ± SD
4 control group 220.8 ± 22.1 157.4 ± 23.57 6 denervated sided 180.2 ± 18.2 165.5 ± 23.4
contralateral sidee 235.0 ± 35.8 168.0 ± 13.514 6 denervated sided 241.7 ± 32.2 178.0 ± 40.6
contralateral sidee 203.0 ± 47.4 150.8 ± 33.921 6 denervated sided 313.3 ± 12.19 217.7 ± 24.6fg
contralateral sidee 227.4 ± 17.3 148.8 ±11.1
a
b
de
f
g
Macrophage-like cells.Cells expressing class II MHC molecules.Number of animals in each group.The side with axotomy of the inferior alveolar nerve and mesial movement of the first mandibular molar.The intragroup control side with no axotomy of the inferior alveolar nerve and no tooth movement.Wilcoxon signed-rank test showing significant difference from the contralateral side; P < 0.05.Mann-Whitney test showing significant difference from the control group; P < 0.01.
Control group. Cells im-munolabeled with CD11bantibody corresponded totissue macrophages anddemonstrated mainly ovalor round morphology,sometimes with shortpseudopodia (Figs. la, lb).Cells expressing class IIMHC molecules were irreg-ular or dendritic in appear-ance, having ramified,elongated cell processes(Figs. 2a, 2b). Nevertheless,the latter ones could oftenbe observed as round orspindle-shaped, thus re-sembling CDllb+ or
j Dent Res 78(6) 1999
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Immunoco7npetent Cells in Denervated PDL
macrophage-like cells (Fig. 2b). BothCD11b' cells and cells expressing classII MHC molecules were rather evenlydistributed throughout the PDL. Theywere consistently seen in bone-marrowspaces and close to blood vessels, whilein some sections they were foundbetween the dentin and cellular cemen-tum and close to Malassez epithelialcells (Figs. I a, 2a).
Experimental grotup. At 7 and 14 days, theimmunolabeled cells showed no appar-ent differences in morphology or distri-bution compared with the contralateraland control first mandibular molars,being rather evenly distributed through-out the entire PDL. Hyalinized tissuecould be seen in the PDL of three molarsin the seven-day group (n 6), and inthe PDL of one molar in the 14-daygroup (n = 6). Although both CD11b+cells and cells expressing class II MHCmolecules were seen close to the hyalin- Figure la. Inter-riized zone (Fig. 3) in both of the experi- first mandibularmental periods, none of the cell types (CDllb+) cells (ainvaded the hyalinized tissue. An over- marrow spaces ('invadedepithelial cells (M)all increase in the distribution of dentin. Bar 0.1 mimmunolabeled cells was seen at 21days. In the PDL of the only molar thatdemonstrated hyalinized tissue at thisexperimental period (n = 6), a number ofCD11b cells and fewer cells expressingclass II MHC molecules were seenaround and invading the hyalinizedarea (Fig. 4). In some of the sections, atboth 14 and 21 days, immunolabeledcells were observed close to and insideroot resorption lacunae (not shown inthe micrographs), as well as in the vicin-ity of Malassez epithelial cells.
Discussion
The results of the present studydemonstrate that two weeks after theapplication of orthodontic force and Figure lb. Cellsaxotomy of the inferior alveolar nerve, CDIIb antibody aiheads) or roundthe number and distribution of the times with shortinvestigated immunocompetent cells in arrow). AB = alve(the PDL of rat molars were not signifi- Bar = 0.1 mm.cantly changed. However, three weeksafter the initiation of experimentaltooth movement, macrophage-like and class II MHC mole-cule-expressing cells, both belonging to the mononuclearphagocytic system, were abundant and significantlyincreased in number throughout the entire PDL.
The choice of the experimental periods in this investiga-tion, namely, 7, 14, and 21 days, was based on results
radicular area of a control Figure 2a. Inter-radicular area of a controlmolar. Macrophage-like first mandibular molar demonstrating classrrows) are seen in bone II antigen-expressing cells (RT1B) (arrows).') and close to Malassez AB = alveolar bone; D = dentin. Bar = 0.1[)AB = alveolar bone; D = mm.nm.
immunopositive to b .* WYT -
Ie mainly oval (arrow- Figure 2b. Most of the immunolabeled(open arrow), some- cells (RTIB) in the periodontal ligamentpseudopodia (small (PDL) are irregular or dendritic in shapeAlar bone; D = dentin. (arrowheads), while few are round or
oval (small arrow). Bar = 0.1 mm.
obtained from our previous investigations (Vandevska-Radunovic et al., 1997b, 1998). Using the same experimentalmodel, but in innervated, maxillary rat molars (Vandevska-Radunovic et al., 1997b), we have found that the recruitmentof immunocompetent, phagocytic cells showed no signifi-cance at day 1. The first significant changes occurred at 3
j Deiit Res 78(6) 1999 1217
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

1218 Vandcevska-Radudnovic et al.
Figure 3. Experimental first mandibularmolar, seven days after orthodontic toothmovement. CDllb-immunopositive cells(arrows) are seen close to, but not invading,the hyalinized tissue (H). D = dentin; P =pulp. Bar = 0.1 mm.
Figure 4. A number(CD11b+) cells (M) surr(hyalinized area (H) ofafter experimental todentin. Bar = 0.1 mm.
days after orthodontic tooth movement, and the changeswere maintained at 7 days. At 14 days, the macrophage-likecells and cells expressing class II MHC molecules showed atendency to approach the control values, although the formerones maintained their significant increase on the experimen-tal side. In the present study, the number of the respectivecells showed no significant increase at either 7 or 14 daysafter the initiation of tooth movement, but at 21 days, thechanges were significant. The prolongation of the experimen-tal periods to 21 days in the present investigation was justi-fied by the finding that the rat PDL demonstrates almostcomplete sensory re-innervation at this stage (Vandevska-Radunovic et al., 1998). Thus, we could study the mobiliza-tion of immunocompetent phagocytic cells in orthodonticallystressed PDL not only during complete denervation (7 dayspost-axotomy), but also during initial (14 days post-axo-tomy) and established periodontal re-innervation (21 dayspost-axotomy). Compared with the innervated and ortho-dontically treated maxillary molars, the increase inmacrophage-like and class II molecule-expressing cells in thedenervated PDL is delayed, and concurs with the re-innerva-tion processes of periodontal tissues after axotomy of theinferior alveolar nerve (Berger et al., 1983; Wakisaka et al.,1987; Vandevska-Radunovic et al., 1998). This supports thefindings from previous studies that have shown more favor-able mobilization of immune cells in innervated than dener-vated pulps after injury (Fristad et al., 1995a), as well asaccelerated loss of pulp tissue in denervated than innervatedteeth following occlusal exposure (Byers and Taylor, 1993).
Rat dental tissues demonstrate almost complete deple-
tion of neurogenic peptides two daysafter axotomy of the inferior alveolarnerve (Berger et al., 1983), while thefirst re-innervation processes takeplace two weeks afterward (Wakisakaet al., 1987; Vandevska-Radunovic etal., 1998). The unchanged number ofimmunocompetent cells in the dener-vated and experimentally treated PDLwithin this period is not an unex-pected finding, and is in agreementwith previous reports (Fristad et al.,1995b). If one remembers that the lifes-pan of tissue macrophages is abouttwo months (Fawcett, 1994), it is veryprobable that the population of tissuemacrophages would not suffer severealterations in number two weeks afterdenervation. However, the unusualobservation was the absence ofmacrophages surrounding and invad-ing hyalinized tissues, which is in con-trast to previous findings.
Of macrophage-like Mononucleated, macrophage-like cellsound and invade the are abundant in the periphery of'he PDL, three weeks hyalinized zones and initiate theoth movement. D phagocytosis of necrotic, hyalinized
tissue in the PDL of innervated andexperimentally moved rat molars(Brudvik and Rygh, 1993; Vandevska-
Radunovic et al., 1997b). Therefore, it is possible that, in den-ervated PDL, the lack of local stimulus, presumablyprovided by the nerve fibers, might have prevented residentmacrophages from transforming into activated ones, acquir-ing enhanced phagocytic and antigen-processing activity(Peck, 1987). Once nerve fibers had re-innervated the PDL,both macrophage-like cells and cells expressing class IIMHC molecules encircled and invaded hyalinized tissue 21days after sensory fiber denervation.
Recruitment of immunocompetent cells during experi-mental tooth movement is part of a comprehensive inflam-matory response and coincides and co-localizes with bothvascular (Rygh et al., 1986; Vandevska-Radunovic et al.,1994) and neural (Kvinnsland and Kvinnsland, 1990;Vandevska-Radunovic et al., 1997a) dental tissue changes.Previous work has shown that, in denervated and experi-mentally moved molars, the periodontal blood flow increaseis postponed until a sensory tissue re-innervation is estab-lished (Vandevska-Radunovic et al., 1998). Both the re-inner-vation processes and the blood flow increase in the PDLcoincided and preceded the recruitment of mononuclearphagocytic cells shown in this study. Knowing that bloodcirculating monocytes constitute a large cell reserve that canrapidly be mobilized at sites of injury and infection(Fawcett, 1994), we found it very conceivable that not onlyregeneration of sensory nerve fibers but also enhancedblood flow is necessary for increased deployment ofimmunocompetent cells to the inflamed tissues.
One of the most important aspects of an inflammatoryprocess involves cell adhesion events. Prior to extravasation,
j Deiit Res 78(6) 1999
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

Immunocompetent Cells in Denervated PDL
leukocytes must adhere to endothelial sites (Lasky, 1992), aprocess that can be initiated by continuous mechanical stim-ulation of postcapillary venules, thus simulating the effect ofan orthodontic force (Iida et al., 1996). Several neuropeptidesare shown to up-regulate the expression of endothelial celladhesion molecules and elicit granulocyte infiltration (Smithet al., 1993). They are also shown to induce monocyte and T-lymphocyte chemotaxis (Ruff et al., 1985; Foster et al., 1992)and stimulate formation of new blood vessels through pro-liferation of endothelial cells (Hagerstrand et al., 1990).These neuropeptide properties, together with their potentvasodilatory effect (Brain et al., 1985), signify that sensorynerve fibers have both direct and indirect impact on themigratory ability of immunocompetent cells.
Based on the above, it can be concluded that axotomy ofthe inferior alveolar nerve delays the recruitment ofmacrophage-like and class II MHC molecule-expressingcells in the PDL of experimentally moved rat molars. Thisfurther indicates that sensory nerve fibers participate in themobilization of immunocompetent cells and modulate theirresponse to local tissue challenges, which includes the PDLduring experimental tooth movement.
Acknowledgments
The authors thank I. Sandvik Gavlen and M. Eidsheim forskillful technical assistance. Financial support for this studywas received from the Department of Anatomy and CellBiology, and the Broegelmann Foundation, Medical Faculty,University of Bergen.
References
Abello J, Kaiserlian D, Cuber JC, Revillard JP, Chayvialle JA(1991). Characterization of calcitonin gene-related peptidereceptors and adenylate cyclase response in the murinemacrophage cell line P338D1. Neuropeptides 19:43-49.
Berger RL, Byers MR, Calkins DF (1983). Dental nerve regenera-tion in rats. I. Electrophysiological studies of molar sensorydeficit and recovery. Pain 15:345-375.
Blalock JE (1994). The syntax of immune-neuroendocrine com-munication. Immunol Today 15:504-511.
Brain SD, Williams TJ, Tippins JR, Morris HR, Maclntyre 1 (1985).Calcitonin gene-related peptide is a potent vasodilator.Nature 313:54-56.
Brudvik P, Rygh P (1993). The initial phase of orthodontic root
resorption incident to local compression of the periodontalligament. Eur J Orthod 15:249-263.
Byers MR, Taylor PE (1993). Effect of sensory denervation on theresponse of rat molar pulp to exposure injury. J Dent Res72:613-618.
Cunningham ET Jr, De Souza EB (1993). Interleukin-1 receptorsin the brain and endocrine tissues. Immunol Today 14:171-176.
Davidovitch Z (1991). Tooth movement. Crit Rev Oral Biol Med2:411-450.
Davidovitch Z, Nicolay OF, Ngan PW, Shanfeld JL (1988).Neurotransmitters, cytokines and the control of alveolar
bone remodeling in orthodontics. Dent Clin North Am 32:411-434.
Fawcett DW (1994). A textbook of histology. 12th ed. New York:Chapman & Hall, p. 151.
Foster CA, Mandak B, Kromer E, Rot A (1992). Calcitonin gene-related peptide is chemotactic for human T lymphocytes.Ann NY Acad Sci 657:397-404.
Fristad I, Heyeraas KJ, Kvinnsland IH, Jonsson R (1995a).Recruitment of immunocompetent cells after dentinalinjuries in innervated and denervated young rat molars: animmunohistochemical study. J Histochem Cytochem 43:871-879.
Fristad I, Heyeraas KJ, Jonsson R, Kvinnsland IH (1995b). Effectof inferior alveolar nerve axotomy on immune cells andnerve fibres in young rat molars. Arch Oral Biol 40:1053-1062.
Gyorfi A, Fazekas A, Suba Z, Ender F, Rosival L (1994).Neurogenic component in ligature-induced periodontitis inthe rat. J Clin Periodontol 21:601-605.
Haegerstrand A, Dalsgaard CJ, Jonzon B, Larsson 0, Nilsson J(1990). Calcitonin gene-related peptide stimulates prolifera-tion of human endothelial cells. Proc Natl Acad Sci USA87:3299-3303.
lida J, Warita H, Nakagawa N, Soma K (1996). White blood cellmovement changes in post-capillary venule during intermit-tent or continuous compressions. In: Biological mechanismsof tooth movement and craniofacial adaptation. DavidovitchZ, Norton LA, editors. Boston: Harvard Society for theAdvancement of Orthodontics, pp. 189-194.
Jafarian-Tehrani M, Amrani A, Homo-Delarche F, Marquette C,Dardenne M, Haour F (1995). Localization and characteriza-tion of interleukin-1 receptors in the islet of Langerhansfrom control and non-obese diabetic mice. Endocrinology136:609-613.
Jager A, Radlanski RJ, Gotz W (1993). Demonstration of cells ofthe mononuclear phagocytic lineage in the periodontiumfollowing experimental tooth movement in the rat. Animmunohistochemical study using monoclonal antibodiesEDI and ED2 on paraffin-embedded tissues. Histochemistry100:161-166.
Jonsson R, Tarkowski A, Klareskog L (1986). A demineralizationprocedure for immunopathological use. EDTA treatmentpreserves lymphoid cell surface antigens. J Immunol Meth88:109-114.
Kvinnsland I, Kvinnsland S (1990). Changes in CGRP-immunore-active nerve fibres during experimental tooth movement in
rats. Eur J Orthod 12:320-329.Lasky LA (1992). Selectins: interpreters of cell-specific carbohy-
drate information during inflammation. Science 258:964-969.Maggiano N, Piantelli M, Ricci R, Larocca LM, Capelli A,
Ranelletti FO (1994). Detection of growth hormone-produ-cing cells in human thymus by immunohistochemistry andnon radioactive in situ hybridization. J Histochem Cytochem42:1349-1354.
Mayer EA, Raybould H, Koelbel C (1988). Neuropeptides,inflammation, and motility. Dig Dis Sci 33(Suppl):71S-77S.
McMaster WR, Williams AF (1979). Identification of Ia glycopro-teins in rat thymus and purification from rat spleen. Eur JImmunol 9:426-433.
Nicolay OF, Davidovitch Z, Shanfeld JL, Alley K (1990).Substance P immunoreactivity in periodontal tissues during
j Dent Res 78(6) 1999 1219
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from

1220 Vandevska-Radunovic et al.
orthodontic tooth movement. Bone Miner 11:19-29.Olgart L (1996). Neural control of pulpal blood flow. Crit Rev Oral
Biol Med 7:159-171.O'Laughlin S, Braverman M, Smith-Jefferies M, Buckley P (1992).
Macrophages (histiocytes) in various reactive and inflamma-tory conditions express different antigenic phenotypes. HumPathol 23:1410-1418.
Peck R (1987). Neuropeptides modulating macrophage function.Ann NY Acad Sci 496:264-270.
Przewlocki R, Hassan AHS, Lason W, Epplen C, Herz A, Stein C(1992). Gene expression and localization of opioid peptidesin immune cells of inflamed tissue: functional role inantinociception. Neuroscience 48:491-500.
Robinson AP, White TM, Mason DW (1986). Macrophage hetero-geneity in the rat as delineated by two monoclonal antibod-ies MRC OX-41 and OX-42, the latter recognizingcomplement receptor type 3. Immunology 57:239-247.
Ruff MR, Wahl SM, Pert CB (1985). Substance P receptor-medi-ated chemotaxis of human monocytes. Peptides 6:107-111.
Rygh P, Bowling K, Hovlandsdal L, Williams S (1986). Activationof the vascular system: a main mediator of periodontal fiberremodeling in orthodontic tooth movement. Am J Orthod89:453-468.
Saito I, Ishii K, Hanada K, Sato 0, Maeda T (1991). Responses ofcalcitonin gene-related peptide-immunopositive nerve fibresin the periodontal ligament of rat molars to experimentaltooth movement. Arch Oral Biol 36:689-692.
Smith CH. Barker JN, Morris RW, MacDonald DM, Lee TH
(1993). Neuropeptides induce rapid expression of endothe-hal cell adhesion molecules and elicit granulocyte infiltrationin human skin. JImmunol 151:3274-3282.
Umeda Y, Arisawa M (1989). Characterization of the calcitoningene-related peptide receptor in mouse T lymphocytes.Neuropeptides 14:237-242.
Vandevska-Radunovic V, Kristiansen AB, Heyeraas KJ,Kvinnsland S (1994). Changes in blood circulation in teethand supporting tissues incident to experimental tooth move-ment. Eur I Orthod 16:361-369.
Vandevska-Radunovic V, Kvinnsland S, Kvinnsland IH (1997a).Effect of experimental tooth movement on nerve fibresimmunoreactive to calcitonin gene-related peptide, proteingene product 9.5, and blood vessel density and distributionin rats. Eur J Orthod 19:517-529.
Vandevska-Radunovic V, Kvinnsland IH, Kvinnsland S, JonssonR (1997b). Immunocompetent cells in rat periodontal liga-ment and their recruitment incident to experimental ortho-dontic tooth movement. Eur J Oral Sci 105:36-44.
Vandevska-Radunovic V, Kvinnsland IH, Kvinnsland S (1998).Effect of inferior alveolar nerve axotomy on the periodontaland pulpal blood flow subsequent to experimental toothmovement in rats. Acta Odontol Scand 56:57-64.
Wakisaka S, Ichikawa H, Nishikawa S, Uchiyama T, Matsuo S,Takano Y, et al. (1987). Immunohistochemical study onregeneration of substance P-like immunoreactivity in ratmolar pulp and periodontal ligament following resection ofinferior alveolar nerve. Arch Oral Biol 32:225-227.
j Dent Res 78(6) 1999
by guest on July 14, 2011 For personal use only. No other uses without permission.jdr.sagepub.comDownloaded from




















