
A mouse embryonic stem cell bank for inducible overexpression of human chromosome 21 genes
Upload
independentCategory
view
2download
0
European Journal of Immunogenetics (1992), 19, 141-158
D A M A G E IN B2m GENES A N D D N A
LOSS O F EXPRESSION OF CLASS I MHC PRODUCTS O N T H E MEMBRANE OF L R . 4 , A CELL
L5178Y
METHYLATION OF H - 2 GENES A R E INVOLVED I N
LINE D E R I V A T I V E O F THE T-CELL LYMPHOMA
G . N A V A , * R . O C A D I Z , ~ V . ORTEGA+ & G . A L F A R O *
* Departamento de Inrnunologia, Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, Mexico and *Division de Investigacion Basica, Instituto Nacional de
Cancerologia, Mexico
(Received 4 February 1992; accepted 24 February 1992)
S U M M A R Y
We have isolated an H-2 deficient cell line (LR.4) from the T-cell lymphoma L5178Y which grew without restrictions in the peritoneal cavity of different inbred strains of mice. The use of polyclonal anti-H-2 antiserum and complement indicated that LR.4 cells did not express class I determinants on the cell membrane. Southern blots of genomic DNA of LR.4 cells showed that B2m genes were severely damaged and that class I H-2 genes were extensively methylated. Consequently, LR.4 cells failed to transcribe mRNAs for both B2m and class I H-2 genes. On the other hand, specific immunity to LR.4 was demonstrated in C57BU6J mice since, in subsequent challenges with either LR.4 or EL4.4, LR.4 did not grow, whereas EL4.4 grew and killed the mice. In C57BL/6J mice, rejection of LR.4 was accompanied by the production of cytotoxic antibodies. The immune response induced in C57BL/6J mice was determined by non-H-2 antigenic determinants in LR.4 cells.
I N T R O D U C T I O N
Tumour progression and production of metastasis is the consequence of a complex and still poorly understood sequence of events in which both the malignant cell and the immunological status of the host play a fundamental role. Neoplastic cells are quite adaptable to different environments and so may escape immune surveillance by one or more mechanisms, such as antigenic modulation (Esmon & Little, 1976; Old et al., 1968), masking of antigenic determinants (Esmon & Little, 1976; Tokuyama & Migita, 1978; Celis et al . , 1979a, 1979b), or by qualitative or quantitative changes in the expression of membrane proteins (Schmidt & Festenstein, 1980;
Correspondence: Guillermo Alfaro, Departamento de Inmunologia, Instituto de Investigaciones Biome- dicas, Universidad Nacional Autonoma de Mexico, 04510 Mexico D.F., Mexico. Tel. 550 52 15; Telex: 177429.
141

142 G. Nava et al. Festenstein & Schmidt, 1981; Isakov et al., 1983; Katzav et al., 1983; Hui et al., 1986; Ostrand-Rosenberg et al . , 1989; Porgador et a l . , 1989).
The products of the MHC of the mouse (H-2) play an essential role in the development and regulation of an immune response. These molecules are encoded on chromosome 17 by a rather large number of polymorphic genes which can be classified into three main categories according to their genetic relationship and function (Hood et al., 1983; Klein etal . , 1983). Class I molecules, which include the classical transplantation antigens (K, D, and L) and which are expressed by most cells, restrict functions mediated by CTLs (Zinkernagel & Doherty, 1973). Class I1 molecules are the products of the immune response genes (IA and IE), have selective cellular distribution, and participate in the recognition of antigens by T helper lymphocytes (Schwartz, 1985). Class I11 genes encode some complement components (Hood et al., 1983; Klein et a l . , 1983).
Although a great deal of information about Class I molecules has been published in recent years, several questions relevant to their physiological role remain open, one being their participation in the systemic spread of malignant cells. In general, malignant cells tend to invade adjacent structures and produce metastasis. Variations in the expression of Class I molecules may alter the immunogenicity of the neoplastic cell, thereby altering its malignant potential. Cell variants which lack H-2 antigens on the membrane may escape an immune response mediated by CTLs and thus produce tumours. In fact, such situations have been described in distinct experimental systems in which tumours belonging to unrelated differentiation pathways were used (Gmiir et al . , 1980; Ostrand-Rosenberg et al., 1980; Isakov et al . , 1983; Eisenbach et al . , 1984; Katzav et al . , 1984; Potter et al . , 1984, 1985; Katzav et al . , 1985; Ljunggren & Karre, 1985; Wallich et al., 1985; Karre et al., 1986).
Most transplantable murine tumours grow in the strain of origin but fail to do so in other strains of mice. However in some cases cell variants lacking H-2 on the membrane can be successfully transplanted across allogeneic barriers (Eisenbach et al . , 1984; Katzav et al . , 1984). Lack of expression of these molecules may be considered as conferring a selective advantage to these cells when growing in vivo.
In this work, we used a mutant (LR.4) of an H-2 defective cell variant (LR) of the T-cell lymphoma L5178Y; LR.4 grew well in the peritoneal cavity of mice of distinct haplotypes. Since lack of expression of H-2 determinants on the cell surface may be a means by which a tumour cell escapes immune recognition, studies were carried out to gain insight on the immunological properties and genetic alterations which were involved in the unusual growth of this tumour cell variant across allogeneic barriers. We found that the cell phenotype was determined by mutations in B2rn genes and by changes in the pattern of DNA methylation in H-2 genes. Of the several inbred strains of mice investigated, only C57BLJ6J were capable of mounting an efficient immune response which led to rejection of the tumour after several weeks of growth.
M A T E R I A L S A N D M E T H O D S
Mice Eight to twelve week-old mice of the following inbred strains were used in this study: BALB/c and DBM2 (H-2d); BALB.b and C57BU6J (H-2b) and BALB.k (H-2k). An outbred strain, CD1, was also used. All mice were bred in the vivarium of the Instituto de Investifaciones Biomedicas.
Tumour cell lines A spontaneous cell variant, LR, that could grow across allogeneic barriers was isolated in our

Lack of expression of class I determinants in LR.4 143 laboratory from the methyl cholanthrene-induced T-cell lymphoma L5178Y (H-2d) (Fisher, 1958). LR.4, a spontaneous mutant resistant to 5-bromo-2'-deoxyuridine (Sigma Chemical Co., St Louis, MO, USA) was selected by exposing LR to increasing concentrations of this compound. Both LW.l and EL4.4 were obtained by mutation of L5178Y and EL4 (H-2b) (Gorer, 1950) respectively, with ethyl methanesulphonate (Thompson, 1979) and by selection for cells resistant to 8-azaguanine (both from Sigma). All these mutants were RPMI-HAT (Littlefield, 1966) sensitive and retained the growth characteristics, both in vivo and in vitro, of the respective parental cell line.
EL4.4, LW.1, and LR.4 were maintained either in vitro or in the peritoneal cavity of BALB.b, BALBlc, and BALB.k mice, respectively. The culture medium used for propagating these cell lines in culture conditions (37°C; humidified 5% C02-air) was RPMI 1640 supplemented with 10% heat-inactivated fetal calf serum, 1 mM L-glutamine, and 1% of the following: non-essential amino acids, sodium pyruvate, and a combination of antibiotics (all these products were from Gibco Laboratories, Grand Island, NY, USA).
Cell fusion experiments
For the fusion (Fazkas et al., 1980), a cell mixture consisting of 1 x lo6 cells of both LR.4 and EL4.4, each taken from log phase cultures, were used. The fusion was carried out in RPMI 1640 containing 50% polyethylene glycol (PEG 1000, Sigma). Cell hybrids were selected by seeding the fusion mixture in 96-well plates (Costar, Cambridge, MA, USA) containing RPMI-HAT medium and murine peritoneal macrophages as feeder layers. Wells showing growth of colonies one week after fusion were expanded and either cloned by limited dilution or transferred to non-selective medium for freezing and storage in liquid nitrogen until use. As a routine procedure, cell hybrids were cultured under selective pressure.
Conventional antisera and murine monoclonal antibodies
Anti-H-2 antisera were prepared by repeated intraperitoneal (i.p.) injections of 1 X 10' spleen cells from donor mice of the appropriate recipient strain: for anti-H-2b, BALB.b into BALBlc, and for anti-H-2d, BALBlc into BALB.b. The specificity of each antiserum was tested against target and control cells in a cytotoxicity assay in which fresh rabbit serum was used as a complement source.
Murine hybridomas producing monoclonal antibodies (MoAb) 34-5-8s to H-2Dd (Ozato et al., 1982), 28-14-8s to antigens ahred by H-2Ld, H-2Db andH-2Dq (Ozato etal., 1980), and SF1-1.1.1 to H-2Kd (Dr A. M. Stall, Stanford University, CA, USA) were obtained from the ATCC (Rockville, MA, USA). Indirect immunofluorescence was performed on unfixed cell suspensions using culture supernatants containing MoAb as described in details elsewhere (Janossy & Amlot, 1987; Alfaro et al., 1992). Fluorescinated goat anti-mouse Ig was purchased from Southern Biotechnology Associates Inc. (Birmingham, AL, USA).
Radiolabelling of cells and cytotoxicity assay
Each cell line was labelled by incubating 1 X lo6 cells in 0.5ml RPMI 1640 with 200pCi of 51Cr (Na2Cr04, 350 to 600 mCi mg-' Cr; (Amersham, Aylesbury, Buckinghamshire, UK)) for 1 h under culture conditions. Thereafter the cells were washed three times by centrifugation with cold RPMI 1640. The cells were adjusted to the desired concentration with RPMI 1640.
The presence of cytotoxic antibodies in sera was determined by the "Cr release assay (Celis et al., 1979a). In brief, "Cr labelled cells were distributed among test tubes (4 x lo4 cells in 25 p1 per tube) in an ice-water bath. To each tube, 25 pl of a dilution of the appropriate antisera was added
C

144 G. Nava et al. and incubated for 15 min. Thereafter, 25 pl of rabbit serum, as complement source, was added and the volume of each tube was brought to lOOpl with RPMI 1640 and all tubes were incubated for 1 h under culture conditions. After the incubation, 1.9ml of cold PBS was added to each tube and all tubes were centrifuged. After centrifugation, 1.0ml of supernatant was removed from each tube and transferred to fresh tubes to be counted. Controls to assess the toxicity of antibody, complement, or medium were included.
Yo specific release
experimental release - spontaneous release total release - spontaneous release x 100 - -
DNA isolation and Southern blots High molecular weight DNA was isolated from cell lines growing in the peritoneal cavity of appropriate inbred strains of mice and from liver of DBN2 mice as described by Sambrook et al. (1989). DNA samples (10 pg) were digested with selected restriction enzymes under conditions recommended by the manufacturer (Amersham, Aylesbury, UK) and electrophoresed in 0.8% agarose gels. The DNA was denatured in situ and transferred to nitrocellulose filters (Schleicher & Schuell Inc., Keene, NH, USA) according to the procedure of Southern (1975). Pre-hybridi- zation and hybridization with nick-translated DNA from plasmid subclones of genomic or cDNA clones were performed as described elsewhere (Alfaro et al., 1992). Washing conditions were: once in 2 x SSC, 0.1% NaDodS04 at room temperature, and three times with 0.1 X SSC at 50°C.
RNA blot analysis
Total RNA was prepared from the cell lines under investigation and from liver of DBN2 mice using the guanidine thiocyanate procedure (Chirwin et al., 1979), pelleting the RNA through a CsCl cushion. Ten micrograms of each RNA were electrophoresed through a 1.5% agarose gel containing 2 . 2 M formaldehyde (Sambrook et al., 1989). The RNA was transferred to nitrocellulose filters according to a method described by Thomas (1980) and hybridized to nick-translated DNA probes. Hybridization and wash conditions were as described in the preceding section.
DNA probes
The following DNA probes were used in this investigation: (a) a 0.66kb HpaII insert of a cDNA from the H-2Ld gene (Evans et al . , 1982); (b) the cDNA probe pH-2d-5b which contains a 164bp Pstl fragment of the 3' non-coding region of the H-2Kd gene (Xin et al . , 1982); (c) plasmids containing a 6 kb BamHI fragment from IAak or an 11.6 kb HindIII-EcoRI fragment encompass- ing the complete IAPk gene were a gift of Dr Margulies; (d) molecular probes containing a 3.4kb SalI fragment from IEad (Steinmetz et al., 1981; McNicholas et al., 1982) or an 1.8 kb EcoRI insert from IEPd (Steinmetz et al . , 1981); (e) a 0.6kb EcoRI-KpnI fragment of genomic DNA specific for the second exon of the B2m gene (Parnes & Seidman, 1982); and (f) the pBR322 derivative containing a 0.7 kb EcoRI fragment from histone H4 gene (Seyler-Tuyns & Birnstiel, 1981).
5-Azacytidine treatment
Five hundred thousand LR.4 cells were cultured in RPMI 1640 medium with 10 ~ L M AzaC (Sigma Chemical Co.) for 3d and subsequently cultured in the same medium without AzaC for another
Lack of expression of class I determinants in LR.4 T
100
90
80
70
30
20
10
Days after transplantation of the tumour
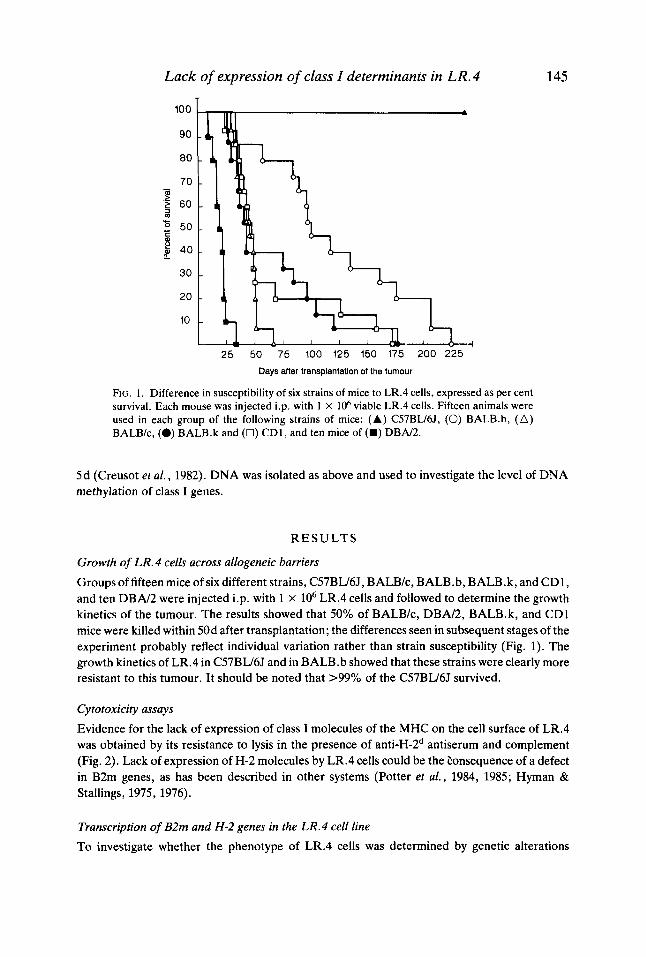
FIG. 1 . Difference in susceptibility of six strains of mice to LR.4 cells, expressed as per cent survival. Each mouse was injected i.p. with 1 X loh viable LR.4 cells. Fifteen animals were used in each group of the following strains of mice: ( A ) C57BU6J, (0) BALB.b, (A) BALB/c, (0) BALB.k and (0) CD1, and ten mice of (U) DBN2.
145
5d (Creusot et al., 1982). DNA was isolated as above and used to investigate the level of DNA methylation of class I genes.
R E S U L T S
Growth of LR.4 cells across allogeneic barriers
Groups of fifteen mice of six different strains, C57BLJ6J, BALB/c, BALB.b, BALB.k, and CD1, and ten D B N 2 were injected i.p. with 1 x lo6 LR.4 cells and followed to determine the growth kinetics of the tumour. The results showed that 50% of BALB/c, DBN2, BALB.k, and CD1 mice were killed within 50d after transplantation; the differences seen in subsequent stages of the experiment probably reflect individual variation rather than strain susceptibility (Fig. 1). The growth kinetics of LR.4 in C57BU6J and in BALB.b showed that these strains were clearly more resistant to this tumour. It should be noted that >99% of the C57BU6J survived.
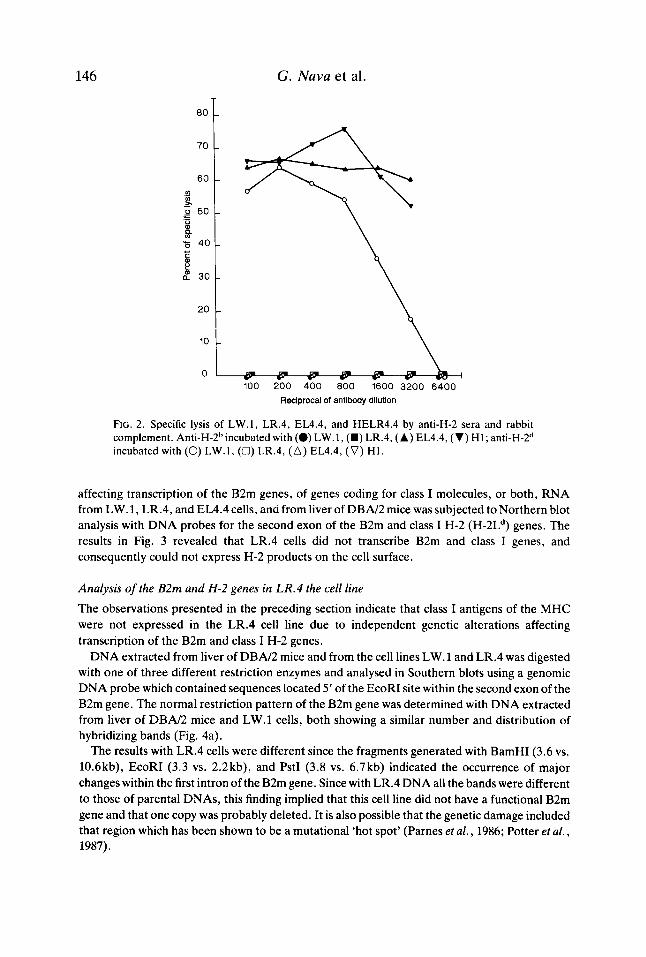
Cytotoxicity assays Evidence for the lack of expression of class I molecules of the MHC on the cell surface of LR.4 was obtained by its resistance to lysis in the presence of anti-H-2d antiserum and complement (Fig. 2). Lack of expression of H-2 molecules by LR.4 cells could be the eonsequence of a defect in B2m genes, as has been described in other systems (Potter et al., 1984, 1985; Hyman & Stallings, 1975, 1976).
Transcription of B2m and H-2 genes in the LR.4 cell line
To investigate whether the phenotype of LR.4 cells was determined by genetic alterations
146 G. Nava et al.
0 1 c a e a m Q a 100 200 400 800 1600 3200 6400
Reciprocal of antibody dilution
FIG. 2. Specific lysis of LW.1, LR.4, EL4.4, and HELR4.4 by anti-H-2 sera and rabbit complement. A11ti-H-2~incubated with (0) LW.1, (m) LR.4, ( A ) EL4.4, (V) H1; a11ti-H-2~ incubated with (0) LW.1, (0) LR.4, (A) EL4.4, (V) H1.
affecting transcription of the B2m genes, of genes coding for class I molecules, or both, RNA from LW.l, LR.4, and EL4.4 cells, and from liver of DBN2 mice was subjected to Northern blot analysis with DNA probes for the second exon of the B2m and class I H-2 (H-2Ld) genes. The results in Fig. 3 revealed that LR.4 cells did not transcribe B2m and class I genes, and consequently could not express H-2 products on the cell surface.
Analysis of the B2m and H-2 genes in LR.4 the cell line The observations presented in the preceding section indicate that class I antigens of the MHC were not expressed in the LR.4 cell line due to independent genetic alterations affecting transcription of the B2m and class I H-2 genes.
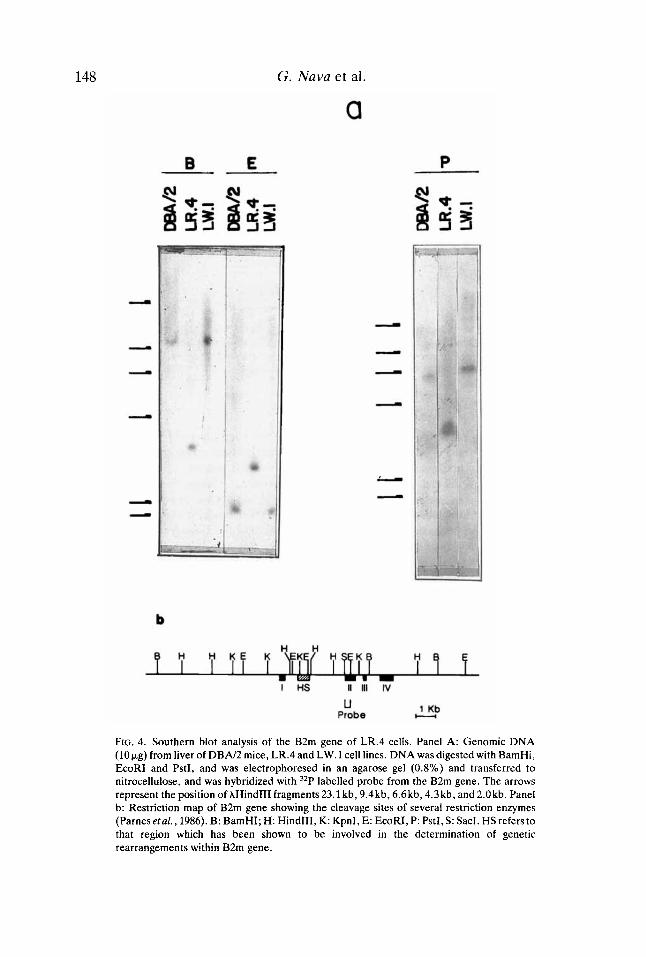
DNA extracted from liver of DBN2 mice and from the cell lines LW.l and LR.4 was digested with one of three different restriction enzymes and analysed in Southern blots using a genomic DNA probe which contained sequences located 5’ of the EcoRI site within the second exon of the B2m gene. The normal restriction pattern of the B2m gene was determined with DNA extracted from liver of DBN2 mice and LW.l cells, both showing a similar number and distribution of hybridizing bands (Fig. 4a).
The results with LR.4 cells were different since the fragments generated with BamHI (3.6 vs. 10.6kb), EcoRI (3.3 vs. 2.2kb), and PstI (3.8 vs. 6.7kb) indicated the occurrence of major changes within the first intron of the B2m gene. Since with LR.4 DNA all the bands were different to those of parental DNAs, this finding implied that this cell line did not have a functional B2m gene and that one copy was probably deleted. It is also possible that the genetic damage included that region which has been shown to be a mutational ‘hot spot’ (Parnes et al., 1986; Potter et al., 1987).

Lack of expression of class I determinants in LR.4 147
FIG. 3. Northern blot analysis of RNA of LR.4 cells. Total RNA (1011.8) extracted from the cell lines LW.l, LR.4, DBA/2, EL4.4, and LR.4 treated with AzaC, was electrophoresed in a 1.2% agarose-formaldehyde gel and transferred to nitrocellulose. The blots were hybridized with 32P labelled probes listed on the left side of the figure: the H-2L” cDNA and genomic clones for P2m and H4.
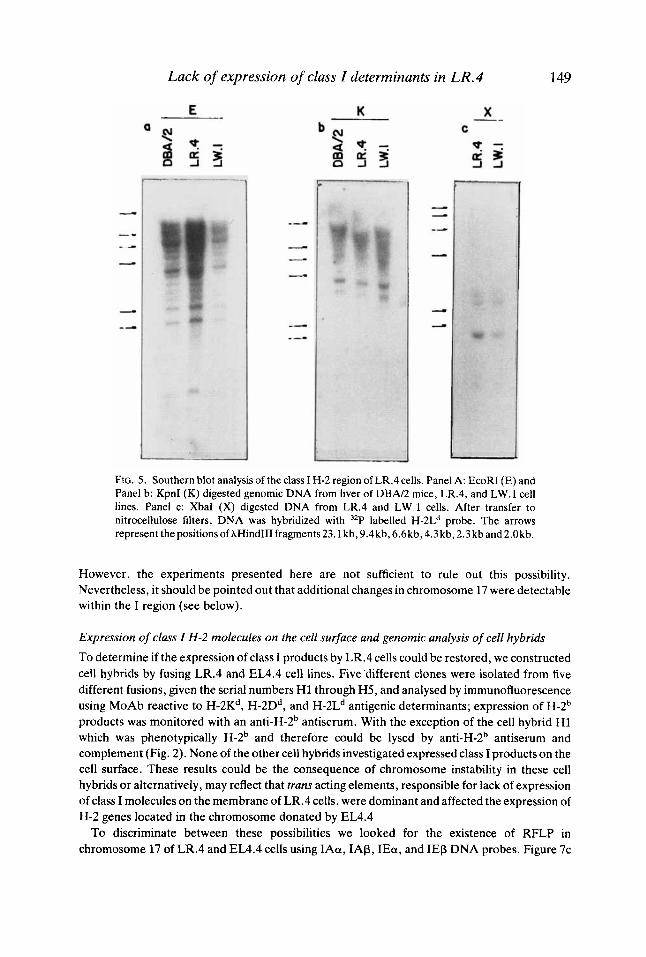
Hybridization of DNA extracted from the cell line LR.4 and LR.l and from liver of DBN2 mice with the class I H-2Ld probe, showed that there had not been major genetic changes in that region of LR.4 encompassing H-2 genes coding for class I antigens (Fig. 5). Two possible explanations for the lack of transcription of class I genes are: changes in the pattern of DNA methylation, the existence of mutations affecting more than one gene, or a combination of both.
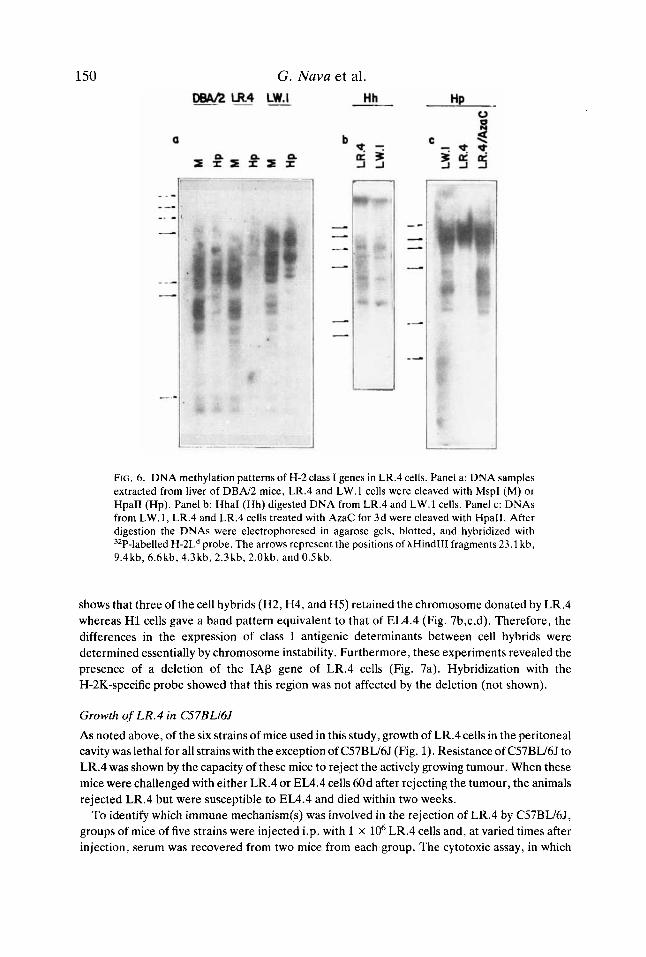
To determine whether the DNA of LR.4 was unusually methylated, the isoschizomers MspI and HpaII which recognize the sequence CCGG containing the predominant methylation site (CpG) in higher eukaryotes were used. HpaII will cut this sequence only if the internal C is unmodified in both strands, whereas MspI will cut regardless of its states of methylation (Bird, 1986; Lindsay & Bird, 1987). Genomic DNA from LR.4 and LW.l cells, and from liver of DBN2 mice, was digested with either MspI or HpaII and probed with H-2Ld (Fig. 6a). The electrophoretic patterns obtained after digesting the three DNAs with MspI were very similar. However, with HpaII we found that the region of the genome of LR.4 containing class I genes was extensively methylated compared with that from LW.l cells and from liver of DBA/2 mice.
To further investigate the level of DNA methylation and its role in the expression of class I molecules, DNA samples from LR.4 and LW.l cells were digested with an additional methylation sensitive enzyme, HhaI. Both LW.1 and LR.4 cells gave a discrete but similar distribution of DNA bands (Fig. 6b), thus suggesting that even if the sequence (CGCG) recognized by this enzyme was extensively methylated in both cell lines, it may not have played a relevant role in the expression of class I genes.
To assess whether DNA methylation was the only factor involved in the lack of transcription of class I genes, LR.4 cells were incubated in the presence of AzaC for 3d. The DNA was extracted 5d later and digested with HpaII. Treatment with AzaC induced extensive demethylation of LR.4 DNA (Fig. 6c). However, LR.4 cells remained incapable of transcribing these genes (Fig.
In relation to the role of mutation in determining the lack of transcription in LR.4 cells, spontaneous and independent mutations affecting H-2K, D and L genes are unlikely to occur.
3).
148 G. Nava et al.
a
FIG. 4. Southern blot analysis of the B2m gene of LR.4 cells. Panel A: Genomic DNA (10pg) from liver of D B N 2 mice, LR.4 and LW.1 cell lines. DNA was digested with BamHi, EcoRI and PstI, and was electrophoresed in an agarose gel (0.8%) and transferred to nitrocellulose, and was hybridized with 32P labelled probe from the B2m gene. The arrows represent the position of AHindIII fragments 23.1 kb, 9.4kb, 6.6kb, 4.3 kb, and 2.0kb. Panel b: Restriction map of B2m gene showing the cleavage sites of several restriction enzymes (Parnes etal., 1986). B: BamHI; H: HindIII, K: KpnI, E: EcoRI, P: PstI, S: SacI. HS refers to that region which has been shown to be involved in the determination of genetic rearrangements within B2m gene.
Lack of expression of class I determinants in LR.4 149
FIG. 5 . Southern blot analysis of the class I H-2 region of LR.4 cells. Panel A: EcoRI (E) and Panel b: KpnI (K) digested genomic DNA from liver of DBN2 mice, LR.4, and LW.l cell lines. Panel c: XbaI (X) digested DNA from LR.4 and L W . l cells. After transfer to nitrocellulose filters, DNA was hybridized with 32P labelled H-2Ld probe. The arrows represent thepositionsofAHindIIIfragments23.1 kb,9.4kb,6.6kb,4.3kb,2.3kband2.Okb.
However, the experiments presented here are not sufficient to rule out this possibility. Nevertheless, it should be pointed out that additional changes in chromosome 17 were detectable within the I region (see below).
Expression of class I H-2 molecules on the cell surface and genomic analysis of cell hybrids
To determine if the expression of class I products by LR.4 cells could be restored, we constructed cell hybrids by fusing LR.4 and EL4.4 cell lines. Five different clones were isolated from five different fusions, given the serial numbers H1 through H5, and analysed by immunofluorescence using MoAb reactive to H-2Kd, H-2Dd, and H-2Ld antigenic determinants; expression of H-2b products was monitored with an anti-H-2b antiserum. With the exception of the cell hybrid H1 which was phenotypically H-2b and therefore could be lysed by anti-H-2b antiserum and complement (Fig. 2). None of the other cell hybrids investigated expressed class I products on the cell surface. These results could be the consequence of chromosome instability in these cell hybrids or alternatively, may reflect that trans acting elements, responsible for lack of expression of class I molecules on the membrane of LR.4 cells, were dominant and affected the expression of H-2 genes located in the chromosome donated by EL4.4
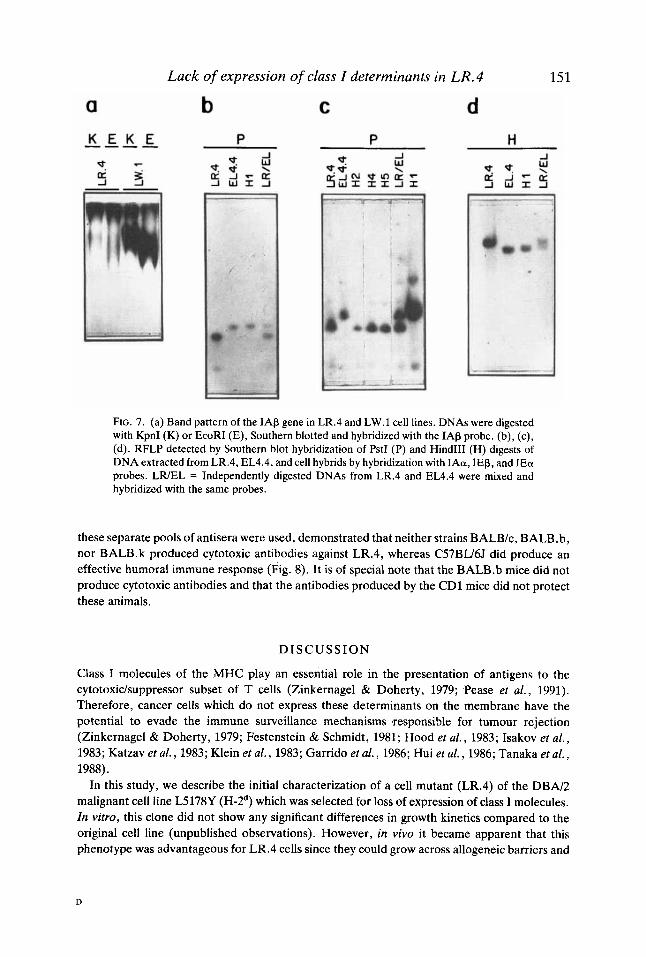
To discriminate between these possibilities we looked for the existence of RFLP in chromosome 17 of LR.4 and EL4.4 cells using IAa, IAP, IEa, and IEP DNA probes. Figure 7c
150 G. Nava et al.
m.4 L m Hh Hp -
FIG. 6. DNA methylation patterns of H-2 class I genes in LR.4 cells. Panel a: DNA samples extracted from liver of D B N 2 mice, LR.4 and LW.1 cells were cleaved with MspI (M) or HpaII (Hp). Panel b: HhaI (Hh) digested DNA from LR.4 and LW.1 cells. Panel c: DNAs from LW.l, LR.4 and LR.4 cells treated with AzaC for 3d were cleaved with HpaII. After digestion the DNAs were electrophoresed in agarose gels, blotted, and hybridized with 'ZP-labelled H-2L" probe. The arrows represent the positions of XHindIII fragments 23.1 kb, 9.4kb, 6.6kb,4.3kb,2.3kb,2.0kb, and0.5kb.
shows that three of the cell hybrids (H2, H4, and H5) retained the chromosome donated by LR.4 whereas H1 cells gave a band pattern equivalent to that of EL4.4 (Fig. 7b,c,d). Therefore, the differences in the expression of class I antigenic determinants between cell hybrids were determined essentially by chromosome instability. Furthermore, these experiments revealed the presence of a deletion of the IAP gene of LR.4 cells (Fig. 7a). Hybridization with the H-2K-specific probe showed that this region was not affected by the deletion (not shown).
Growth of LR.4 in C57BLl6J As noted above, of the six strains of mice used in this study, growth of LR.4 cells in the peritoneal cavity was lethal for all strains with the exception of C57BW6.l (Fig. 1). Resistance of C57BW6J to LR.4 was shown by the capacity of these mice to reject the actively growing tumour. When these mice were challenged with either LR.4 or EL4.4 cells 60d after rejecting the tumour, the animals rejected LR.4 but were susceptible to EL4.4 and died within two weeks.
To identify which immune mechanism(s) was involved in the rejection of LR.4 by C57BL/6J, groups of mice of five strains were injected i.p. with 1 x lo6 LR.4 cells and, at varied times after injection, serum was recovered from two mice from each group. The cytotoxic assay, in which
a Lack of expression of class I determinants in LR.4
b C d 15 1
FIG. 7. (a) Band pattern of the IAP gene in LR.4 and LW.l cell lines. DNAs were digested with KpnI (K) or EcoRI (E), Southern blotted and hybridized with the IAP probe. (b), (c), (d). RFLP detected by Southern blot hybridization of PstI (P) and Hind111 (H) digests of DNA extracted from LR.4, EL4.4, and cell hybrids by hybridization with IAa, IEP, and IEa probes. LWEL = Independently digested DNAs from LR.4 and EL4.4 were mixed and hybridized with the same probes.
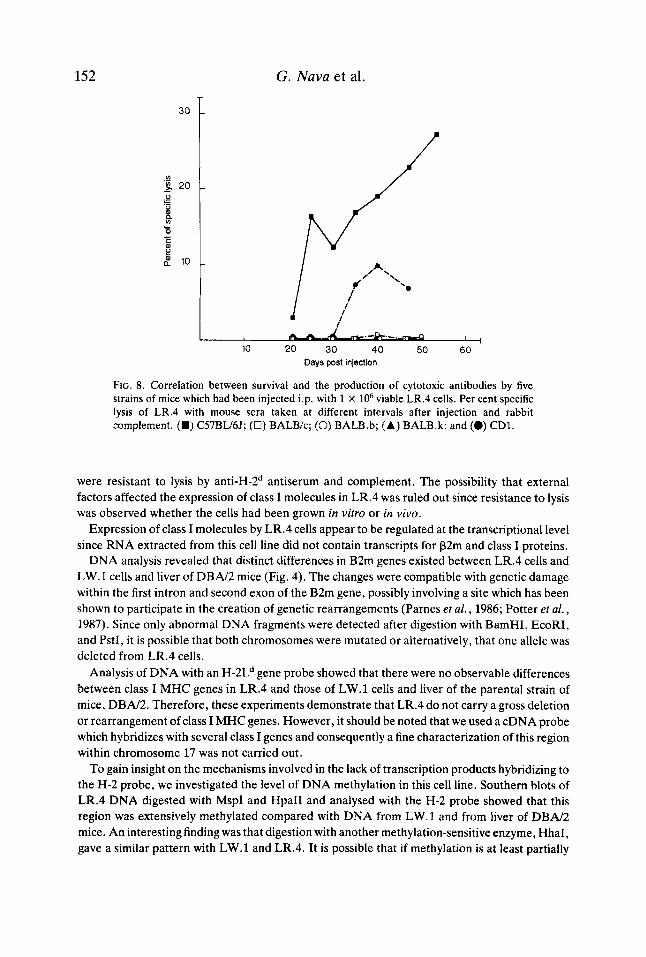
these separate pools of antisera were used, demonstrated that neither strains BALBk, BALB.b, nor BALB.k produced cytotoxic antibodies against LR.4, whereas C57BW6J did produce an effective humoral immune response (Fig. 8). It is of special note that the BALB.b mice did not produce cytotoxic antibodies and that the antibodies produced by the CD1 mice did not protect these animals.
D I S C U S S I O N
Class I molecules of the MHC play an essential role in the presentation of antigens to the cytotoxidsuppressor subset of T cells (Zinkernagel & Doherty, 1979; Pease et al . , 1991). Therefore, cancer cells which do not express these determinants on the membrane have the potential to evade the immune surveillance mechanisms *responsible for tumour rejection (Zinkernagel & Doherty, 1979; Festenstein & Schmidt, 1981; Hood et al., 1983; Isakov et al . , 1983; Katzav et al., 1983; Klein et al . , 1983; Garrido et al., 1986; Hui et al., 1986; Tanaka et al . , 1988).
In this study, we describe the initial characterization of a cell mutant (LR.4) of the DBN2 malignant cell line L5178Y (H-2d) which was selected for loss of expression of class I molecules. In vitro, this clone did not show any significant differences in growth kinetics compared to the original cell line (unpublished observations). However, in vivo it became apparent that this phenotype was advantageous for LR.4 cells since they could grow across allogeneic barriers and
D
152 G. Nuva et al.
30
1 1 I ,
10 20 30 40 50 60 Days post injection
FIG. 8. Correlation between survival and the production of cytotoxic antibodies by five strains of mice which had been injected i.p. with 1 X lo6 viable LR.4 cells. Per cent specific lysis of LR.4 with mouse sera taken at different intervals after injection and rabbit complement. (H) C57BU6J; (U) BALB/c; (0) BALB.b; (A) BALB.k; and (0) CDI.
were resistant to lysis by anti-H-2d antiserum and complement. The possibility that external factors affected the expression of class I molecules in LR.4 was ruled out since resistance to lysis was observed whether the cells had been grown in vitro or in vivo.
Expression of class I molecules by LR.4 cells appear to be regulated at the transcriptional level since RNA extracted from this cell line did not contain transcripts for P2m and class I proteins.
DNA analysis revealed that distinct differences in B2m genes existed between LR.4 cells and LW.1 cells and liver of DBN2 mice (Fig. 4). The changes were compatible with genetic damage within the first intron and second exon of the B2m gene, possibly involving a site which has been shown to participate in the creation of genetic rearrangements (Parnes el al., 1986; Potter et af., 1987). Since only abnormal DNA fragments were detected after digestion with BamHI, EcoRI, and PstI, it is possible that both chromosomes were mutated or alternatively, that one allele was deleted from LR.4 cells.
Analysis of DNA with an H-2Ld gene probe showed that there were no observable differences between class I MHC genes in LR.4 and those of LW.l cells and liver of the parental strain of mice, DBN2. Therefore, these experiments demonstrate that LR.4 do not carry a gross deletion or rearrangement of class I MHC genes. However, it should be noted that we used a cDNA probe which hybridizes with several class I genes and consequently a fine characterization of this region within chromosome 17 was not carried out.
To gain insight on the mechanisms involved in the lack of transcription products hybridizing to the H-2 probe, we investigated the level of DNA methylation in this cell line. Southern blots of LR.4 DNA digested with MspI and HpaII and analysed with the H-2 probe showed that this region was extensively methylated compared with DNA from LW. 1 and from liver of DBN2 mice. An interesting finding was that digestion with another methylation-sensitive enzyme, HhaI, gave a similar pattern with LW.l and LR.4. It is possible that if methylation is at least partially

Lack of expression of class I determinants in LR.4 153 involved in lack of expression of H-2 genes in the LR.4 cell line, the sequence recognized by MspI and HpaII, but not that one recognized by HhaI, may be located predominantly in regions which are crucial for expression of these genes.
Methylation has been shown to be necessary for transcription of H-2K genes in F9 cells and of QlO genes in the liver (Singer & Maguire, 1990; Tanaka etal., 1983,1986). However, the findings obtained with different cell lines suggest that highly methylated DNA prevents expression of class I H-2 genes (Young & Tilghman, 1984; Carrington et al., 1985; Bonal et al., 1986; Meruelo et al., 1986; Miyada & Wallace, 1986). Also, our results indicate that if DNA methylation plays a relevant role in gene expression, this is not the only factor involved, since treatment of LR.4 cells with AzaC resulted in substantial demethylation H-2 genes in LR.4 cells but was not sufficient to induce expression of these genes, a finding which is in agreement with those reported in other experimental systems (Young & Tilghman, 1984; Keeney & Hansen, 1989; Keeney et al., 1989; Maschek et a l . , 1989). Two different cis-acting factors which may affect transcription of class I genes are: changes in the structure of chromatine and the presence of mutations in the regulatory or structural regions of these genes. In fact, such mechanisms appear to be involved in the complete or selective lack of transcription of class I genes in S49 tumour sublines (Keeney & Hansen, 1989; Keeney et al . , 1989) and in metastatic variants cloned from the T10 sarcoma (Aboud et al., 1991).
Several lines of evidence indicate that nuclear factors are implicated in the positive and negative regulation of transcription of class I genes in normal and malignant cells. Mascheck et al. (1989) and Henseling et al. (1990) have described H-2K-deficient cell lines which express reduced amounts of the nuclear factor H2TFUKBFl and that harbour DNase I hypersensitive sites at the regulatory region recognized by this protein (Israel et al., 1987). It is also noteworthy that RadLV and MoLV code for nuclear factors which enhance the expression of class I genes (Brown et al., 1988). On the other hand, in cells transfected or infected with Ad12, the product encoded by the E1A region suppresses transcription of class I genes (Paabo et al., 1989).
It is now well established that P2m plays an essential role in the conformation, intracellular transport, and surface expression of class I molecules (Myers et al., 1989; Williams et al., 1989). Coordinate regulation of class I and p2m genes has been suggested since the nuclear factor H2TFlIKBFl binds with the same affinity to regulatory regions of these genes (Croce et al., 1981; Israel et al., 1987). Furthermore, most cell lines with mutated B2m genes do not express class I molecules on the cell membrane (Hyman & Stallings, 1975; Parnes & Seidman, 1982; Parnes et al., 1986; Potter et al., 1987). However, it should be mentioned that surface expression of class I determinants in the absence of P2m has been described (Potter et al., 1984, 1985).
In the case of LR.4 cells it is unlikely that the genetic alterations affecting P2m and class I genes could have been determined by common regulatory mechanisms. We would like to suggest that lack of expression of these genes is coincidental and the result of independent events.
Our attempts to rescue the expression of class I genes from LR.4 cells in cell hybrids were not successful because they retained only one of the two chromosomes 17. The dominant cell appears to be LR.4 and segregation probably occurred in the early stages after the fusion since the pattern of DNA bands detected in Southern blots of cloned (H2-H3) and uncloned (H4H5) cells was that of the cell line under investigation. Chromosome segregation is a common finding in cell hybrids but the mechanisms responsible for this phenomenon remain unclear. Further attempts to isolate stable clones after fusing LR.4 and EL4.4 cells were not pursued.
Another interesting property of this cell variant was that it was capable of growing across allogeneic barriers, i.e. in strains of inbred mice of different haplotype (H-2’, H-2d, and H-2k), as well as in CDI, an outbred strain. With the exception of C57BL/6J, all the strains investigated were killed by the tumour, albeit with different kinetics (Fig. 1).

154 G. Nuvu et al. In the C57BU6J mice, the neoplastic cells grew but >99% of the mice ultimately rejected the
tumour. Variations in the size of the inoculum gave equivalent results (not shown), a finding which is in disagreement with other reports (Eisenbach el al . , 1984; Katzav et al . , 1984). The finding that mice which had rejected the tumour were resistant to subsequent challenges with the same cell line, implied that the immune system was involved in the rejection.
Therefore, to study the immune response, we injected other groups of the same five strains of mice with LR.4 and assayed for the production of circulating antibodies to this cell variant. Only C57BU6J mice, in which LR.4 was either growing or had been rejected, developed an immune response mediated by antibodies (Fig. 7). This was not the case for the other strains of mice.
The importance of the genetic background of inbred strains of mice in the development of resistance to some lymphoid tumours has been demonstrated by different groups (Fuji & Murakami, 1983; Zalman & Meruelo, 1986). In particular, Fuji & Murakami (1983) found in F1 mice, that a combination of both H-2 and non-H-2 genes is important in raising an immune response against a T-cell tumour. Our results on the resistance of C57BU6J mice to LR.4 are in agreement with this finding. After being inoculated with LR.4, C57BU6J mounted an effective immune response, whereas BALB.b, which is also H-2b, did not. However, the BALB.b mice had a longer period of survival than did the remaining three strains in the study.
The combined participation of the cellular membrane of the neoplastic cell and the immune system of the animal is of fundamental importance in tumour progression and in the production of metastatic nodules. The results presented in this article suggest that the capacity to grow across allogeneic barriers, which is characteristic of LR.4, is probably determined by the lack of expression of class I H-2 determinants. Failure to express class I molecules on the cell surface would allow a tumour cell to evade H-Zrestricted immune mechanisms.
In relation to the nature of the antigenic determinant(s) responsible for the induction of an immune response against LR.4 in C57BW6J mice it should be mentioned that specific cytotoxic T lymphocytes directed against non-H2 antigen(s) of the parental cell line L5178Y have been induced in DBN2 mice (Alexander et al . , 1966). However, we think that the antigen(s) recognized in LR.4 by C57BU6J mice may be different since repeated immunizations of DBN2 and BALB/c mice with mitomycin treated cells did not protect these animals from subsequent challenges with live cells (results not shown).
We conclude that the specific protection against LR.4 in C57BU6J mice was mediated by an immune response elicited by a non-H-2 antigen(s).
A C K N O W L E D G M E N T S
This work was in part supported by grant PCCBBNA-020583 from the Consejo Nacional de Ciencia y Tecnologia, Mexico. We are grateful to Dr C. Lomeli for technical assistance, to Carla Archer for reviewing the manuscript, and to S . Campos, S . Barbosa, and J. AvilCs for art work.
R E F E R E N C E S
ABOUD, M., AMITAI, H. , HULEIHEL, M., HAR-VARDI, I . , GOPAS, J. & SEGAL, S. (1991) Differential transcriptional control of the H-2K and H-2D loci of the major histocompatibility complex in fibrosarcoma cells. Immunological Investigations, 20, 475.
ALEXANDER, P. , CONNELL, D.I. & MIKULSKA, Z.B. (1966) Treatment of a murine leukemia with spleen cells or sera from allogeneic mice immunized against the tumor. Cancer Research, 26, 1508.

Lack of expression of class I determinants in LR.4 155 ALFARO, G., LOMELI, C., OCADIZ, R., ORTEGA, V., BARRERA, R., RAMIREZ, M. & NAVA, G . (1992)
Immunologic and genetic characterization of S180, a cell line of murine origin capable of growing in different inbred strains of mice. Veterinary Immunology and Immunopathology, 30,385.
BIRD, A.P. (1986) CpG-rich islands and the function of DNA methylation. Nature, 321,209. BONAL, F.J., PAREJA, E., MARTIN, J., ROMERO, C. & GARRIDO, F. (1986) Repression of class I H-2K, H-2D
antigens on GR9 methylcholanthrene-induced tumor cell clones is related to the level of DNA methylation. Journal of Immunogenetics, 13, 179.
BROWN, G.D., CHOI, Y., PAMPENO, C. & MERUELO, D. (1988) Regulation of H-2 class I gene expression in virally transformed and infected cells. Critical Reviews in Immunology, 8, 175.
CARRINGTON, M.N., SALTER, R.D., CRESSWELL, P. & TING, J.P.-Y. (1985) Evidence for methylation as a regulatory mechanism in HLA-DR gene expression. Immunogenetics, 22,219.
CELIS, E., HALE, A.H., RUSSELL, J.H. & EISEN, H.N. (1979a) Cyclicalchangesinsusceptibilityofamyeloma tumor (LPC-1) to immune destruction. I. Changes in reactivity with cytotoxic T lymphocytes and a11ti-H-2~ sera. Journal of Immunology, 122, 954.
CELIS, E., CHANG, T.W. & EISEN, H.N. (1979b) Cyclical changes in susceptibility of a myeloma tumor (LPC-1) to immune destruction. 111. Periodic production of a cell surface glycoprotein and changes in reactivity with cytotoxic T cells and a11ti-H-2~ sera. Journal of Immunology, 122, 2245.
CHIRWIN, J.M., PRZYBYLA, A.E., MACDONALD, R.J. & RUTTER, W.J. (1979) Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry, 18, 5294.
CREUSOT, F., Acs, G. & CHRISTMAN, J.K. (1982) Inhibition of DNA methyltransferases and induction of Friend erythroleukemia cell differentiation by 5-azacytidine and 5-aza-2’-deoxycytidine. Journal of Biological Chemistry, 257, 2041.
CROCE, C.M., LINNENBACH, A., HUEBNER, K., PARNES, J.R., MARGULIES, D.H., APPELLA, E. & SEIDMAN, J.G. (1981) Control of expression of histocompatibility antigens (H-2) and P2-microglobulin in F9 teratocarcinoma stem cells. Proceedings of the National Academy of Sciences, USA, 78, 5754.
EISENBACH, L., HOLLANDER, N., GREENFELD, L., YAKOR, H., SEGAL, S. & FELDMAN, M. (1984) The differential expression of H-2K versus H-2D antigens, distinguishing high-metastatic from low metastatic clones, is correlated with the immunogenic properties of the tumor cells. International Journal of Cancer, 34,567.
ESMON, N.L. & LIITLE, J.R. (1976) Different mechanisms for the modulation of TL antigens on murine lymphoid cells. Journal of Immunology, 117,919.
EVANS, G.A., MARGULIES, D.H., CAMERINI-OTERO, R.D., OZATO, K. & SEIDMAN, J.G. (1982) Structure and expression of a mouse major histocompatibility antigen gene, H-2Ld. Proceedings of the National Academy of Sciences, USA, 79, 1994.
FAZEKAS DE ST GROTH, S. & SCHEIDEGGER D. (1980) Production of monoclonal antibodies: Strategy and tactics. Journal of Immunological Methods, 35, 1.
FESTENSTEIN, H. & SCHMIDT, W. (1981) Variation in MHC antigenic profiles of tumor cells and its biological effects. Immunological Reviews, 60, 85.
FISHER, G.A. (1958) Studies of the culture of leukemic cells in vitro. Annals ofthe New York Academy of Sciences, 76, 673.
FUJI, H. & MURAKAMI, M. (1983) Differential tumor immunogenicity of D B N 2 mouse lymphoma L1210 and its sublines. 111. Control of host resistance to drug-resistance L1210 sublines by H-2 linked and non-H-2 linked genes. Journal of the National Cancer Institute, 70, 119.
GARRIDO, M.L., PEREZ, M., DELGADO, C., ROJANO, J. , ALGARRA, I., GARRIDO, A. & GARRIDO, F. (1986) Immunogenicity of H-2 positive and H-2 negative clones of a mouse tumor, GR9. Journal of Immunogenetics, 13, 159.
GMUR, R., SOLTER, D. & KNOWLES, B.B. (1980) Independent regulation of H-2K and H2D gene expression in murine teratocarcinoma somatic cell hybrids. Journal of Experimental Medicine, 151, 1349.
GORER, P.A. (1950) Studies in antibody response of mice to tumor inoculation. British Journal of Cancer, 4, 372.
HENSELING, U., SCHMIDT, W., SCHOLER, H.R., GRUSS, P. & HATZOPOULOS, A.K. (1990) A transcription factor interacting with the class I gene enhancer is inactive in tumorigenic cell lines which suppress major histocompatibility complex class I genes. Molecular and Cellular Biology, 10,4100.
HOOD, L., STEINMETZ, M. & MALISSEN, 8. (1983) Genes of the major histocompatibility complex of the mouse. Annual Review of Immunology, 1,529.
HUI, K.M., MINAMIDE, L., PRANDONI, N., FESTENSTEIN, H. & GROSVELD, F.G. (1986) Structural variation in the H-2 genes of AKR lymphomas. Journal oflmmunogenetics, 13, 117.

156 G. Nuvu et al. HYMAN, R. & STALLINGS, V. (1975) Characterization of a TL- variant of a homozygous TL+ mouse
lymphoma. Immunogenetics, 3, 75. HYMAN, R. & STALLINGS, V. (1976) Analysis of hybrids between H-2+, TL- lymphoma and an H-2+, TL+
lymphoma and its H-2-, TL- variant subline. Immunogenetics, 4, 171. ISAKOV, N., KATZAV, S., FELDMAN, M. & SEGAL, S. (1983) Loss of expression of transplantation antigens
encoded by the H-2K locus on Lewis Lung carcinoma cells and its relevance to the tumor's metastatic properties. Journal of the National Cancer Institute, 11, 139.
ISRAEL, A., KIMURA, A., KIERAN, M., YANO, O., KANELLOPOULOS, J. , LE BAIL, 0. & KOURILSKY, P. (1987) A common positive tram acting factor binds to enhancer sequences in the promoters of H-2 and p-2 microglobulin genes. Proceedings of the National Academy of Sciences, USA, 84, 2653.
JANOSSY, G. & AMLOT, P. (1987) Immunofluorescence and Immunohistochemistry. In: Lymphocytes. A Practical Approach (ed. by G.G.B. Klaus), pp. 67-108. IRL Press, Oxford.
KARRE, K., LJUNGREEN, H.G., PIONTEK, G. & KIESSLING, R. (1986) Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature, 319,675.
KATZAV, S., DEBASTELIER, P., TARTAKOVSKY, B., FELDMAN, M. & SEGAL, S. (1983) Alterations in MHC phenotypes of cloned T10 sarcoma cells are associated with shifts from non-metastatic to metastatic cells. Journal of the National Cancer Institute, 11, 317.
KATZAV, S., SEGAL, S. & FELDMAN, M. (1984) Immuno-selection in vivo of H-2D phenotypic variants from a metastatic clone of sarcoma cells results in cell lines of altered metastatic competence. International Journal of Cancer, 33, 407.
KATZAV, S., SEGAL, S. & FELDMAN, M. (1985) Metastatic capacity of cloned T10 sarcoma cells that differ in H-2 expression: inverse relationship to their immunogeneic potency. Journal of the National Cancer Institute, 15, 307.
KEENEY, J.B. & HANSEN, T.H. (1989) Cis-acting elements determine the locus-specific shutoff of class I major histocompatibility genes in murine S49 lymphoma sublines. Proceedings of the National Academy of Sciences, USA, 86,6288.
KEENEY, J.B., HEDAYAT, M., MYERS, N.M., CONNOLLY, J.M. & HANSEN, T.H. (1989) Locus-specific regulation of Kd, Dd, and Ld class I genes in the BALBlcS49 lymphoma sublines. Journal of Immunology, 143, 2364.
KLEIN, J. , FIGUEROA, F. & NAGY, Z.A. (1983) Genetics of the major histocompatibility complex: the final act. Annual Review of Immunology, 1, 119.
LINDSAY, S. & BIRD, A.P. (1987) Use of restriction enzymes to detect potential gene sequences in mammalian DNA. Nature, 321,336.
LITTLEFIELD, J.W. (1966) The use of drug-resistant markers to study the hybridization of mouse fibroblasts. Experimental Cell Research, 41, 190.
LJUNGGREN, H.G. & KARRE, K. (1985) Host resistance directed selectively against H-2-deficient lymphoma variants. Analysis of the mechanism. Journal of Experimental Medicine, 162, 1745.
MASCHECK, U., PULM, W. & HAMMERLING, G.J. (1989) Altered regulation of MHC class I genes in different tumor cell lines is reflected by distinct sets of DNase I hypersensitive sites. EMBO Journal, 8, 2297.
MASCHEK, U., PULM, W., SEGAL, S. & HAMMERLING, G.J. (1989) Major histocompatibility complex class I genes in murine fibrosarcoma IC9 are down regulated at the level of the chromatin structure. Molecular and Cellular Biology, 9, 3136.
MCNICHOLAS, J., STEINMETZ, M., HUNKAPILLER, T., JONES, P. & HOOD, L. (1982) DNA sequence of the gene encoding the E, Ia polypeptide of the BALB/c mouse. Science, 218, 1229.
MERUELO, D., KORNREICH, K., ROSSOMANDO, A., PAMPENO, C., BORAL, A., SILVER, J.L., BUXBAUM, J . , WEISS, E.H., DEULIN, L.J., MELLOR, A.L., FLAVELL, R.A. & PELLICER, A. (1986) Lack of class I H-2 antigens in cells transformed by radiation leukemia virus is associated with methylation and rearrangement of H-2 DNA. Proceedings of the National Academy ofsciences, USA, 83,4504.
MIYADA, C.G. & WALLACE, B. (1986) Liver specific expression of Qa-associated class I gene is associated with DNA hypomethylation. Molecular and Cellular Biology, 6, 315.
MYERS, N.B., LIE, W.-L., NE'IT, M., RUBOCKI, R.J. & HANSEN, T.H. (1989) The conformation of Ld induced by P2-microglobulin is fixed during de novo synthesis and irreversible by exchange or dissociation. Journal of Immunology, 142,2751.
OLD, L.J., STOCKERT, E., BOYSE, E.A. & KIM, J.H. (1968) Antigenic modulation. Loss of TL antigen from cells exposed to TL antibody. Study of the phenomenon in vitro. Journal of Experimental Medicine, 127, 523.

Lack of expression of class I determinants in LR.4 157 OSTRAND-ROSENBERG, S., RIDER, T.A. & TWAROWSKI, A. (1980) Susceptibility of allogeneic mice to
teratocarcinoma 402AX. Immunogenetics, 10,607. OSTRAND-ROSENBERG, S . , CLEMENTS, V.K., TAHAKUR, A., & COLE, G.A. (1989) Transfection of major
histocompatibility complex class I and class I1 genes causes tumor rejection. Journal of Immunogenetics, 16,343.
OZATO, K., HANSEN, T.H. & SACHS, D.H. (1980) Monoclonal antibodies to mouse MHC antigens. 11. Antibodies to the H-2Ld antigen, the product of a third polymorphic locus of the mouse major histocompatibility complex. Journal of Immunology, 125,2473.
OZATO, K., MAYER, N.M. & SACHS, D.H. (1982) Monoclonal antibodies to mouse antigens. IV. A series of hybridoma clones producing anti-H-Zd antibodies and an examination of expression of H-2d antigens on these cells. Transplantation, 34, 113.
PAABo, S., SEVERINSSON, L., ANDERSSON, M., MARTENS, I., NILSSON, T. & PETERSON, P.A. (1989) Adenovirus proteins and MHC expression. Advances in Cancer Research, 52, 151.
PARNES, J.R. & SEIDMAN, J.G. (1982) Structure of wild-type and mutant p2-microglobulin genes. Cell, 29, 661.
PARNES, J.R., SIZER, K.C., SEIDMAN, J.G., STALLINGS, V. & HYMAN, R. (1986) A mutational hot-spot within an intron of the mouse p2-microglobulin gene. EMBO Journal, 5 , 103.
PEASE, L.R., HORTON, R.M., PULLEN, J.K. & CAI, Z. (1991) Structure and diversity of class I antigen presenting molecules in the mouse. Critical Reviews in Immunology, 11, 1.
PORGADOR, A,, FELDMAN, M. & EISENBACH, L. (1989) Transfection of B16 melanoma cells results in reduced tumorigenic and metastatic competence. Journal of Immunogenetics, 16, 291.
POTTER, T.A., BOYER, C., SCHMITTVERHULST, A.-M., GOLSTEIN, P. & RAJAN, T.V. (1984) Expression on (H-2Dd) on the cell surface in the absence of detectable P2-microglobulin. Journal of Experimental Medicine, 160, 317.
POTTER, T.A., ZEFF, R.A., SCHMITT VERHULST, A.-M. & RAJAN, T.V. (1985) Molecular analysis of an EL4 cell line that expresses H-2Dh but not H-2Kh or P2-microglobulin. Proceedings of the National Academy of Sciences, USA, 82,2950.
POTTER, T.A., FRANKEL, W., ZEFF, R.A. & RAJAN, T.V. (1987) Spontaneous deletion at the p2-m locus: Evidence for the site-specific genetic rearrangement. Journal of Immunology, 138, 1270.
SAMBROOK, J., FFRITSCH, E.F. & MANIATIS, T. (1989) Analysis and Cloning of Eukaryotic Genomic DNA. In: Molecular Cloning. A Laboratory Manual (ed. by ), p. 9.14. Cold Spring Harbor Laboratory Press, New York.
SCHMIDT, W. & FESTENSTEIN, H. (1980) Serological and immunochemical studies of H-2 allospecificities on K36.16 syngeneic tumor of AKR. Journal of Immunogenetics, 7,7.
SCHWARTZ, R.H. (1985) T-lymphocyte recognition of antigen in association with gene products of the major histocompatibility complex. Annual Review of Immunology, 3,237.
SEYLER-TUYNS, A. & BIRNSTIEL, M.L. (1981) Structure and expression in L-cells of a cloned H4 histone gene of the mouse. Journal of Molecular Biology, 151,607.
SINGER, D.S. & MAGUIRE, J.E. (1990) Regulation of the expression of class I MHC genes. Critical Reviews in Immunology, 10,235.
SOUTHERN, E.M. (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. Journal of Molecular Biology, 98, 503.
STEINMETZ, M., MYNARD, K., HORVATH, S. , MCNICHOLAS, J., SRELINGER, J . , WAKE, C., LONG, E., MACH, B. & HOOD, L. (1981) A molecular map of the immune response region from the major histocompatibi- lity complex of the mouse. Nature, 300,35.
TANAKA, K., APPELLA, E. &JAY, G. (1983) Developmental activation of the H-2Kgene iscorrelated with an increase in DNA methylation. Cell, 35, 457.
TANAKA, K., BARRA, Y., ISSELBACHER, K.J., KOURY, G. & JAY, G. (1986) Developmental and tissue-specific regulation of the Q10 class I gene by DNA methylation. Proceedings of the National Academy of Sciences, USA, 83,75.
TANAKA, K., YOSHIOKA, T., BIEBERICH, C. &JAY, G. (1988) Role of the major histocompatibilitycomplex class I antigens in tumor growth and metastasis. Annual Review of Immunology, 6 , 359.
THOMAS, P.S. (1980) Hybridization of denatured RNA and small DNA fragments transferred to nitrocellulose. Proceedings of the National Academy of Sciences, USA, 77,5201.
THOMPSON, L.H. (1979) Mutant isolation. In: Cell Culture. Methods in Enzymology Vol. 58 (ed. by W.B. Jacoby and I.H. Pastan), p. 308. Academic Press, New York.

158 G. Nuvu et al. TOKUYAMA, H. & MIGITA, S. (1978) Cell-surface major glycoprotein of BALB/c mouse plasmocytoma 58-8
cells. Journal of the National Cancer Institute, 61, 203. WALLICH, R. , BALBUC, N . , HAMMERLING, G.J., KATZAV, S . , SEGAL, S. & FELDMAN, M. (1985) Abrogation
of metastatic properties of tumor cells by de novo expression of H-2K antigens following H-2 gene transfection. Nature, 315, 301.
WILLIAMS, D.B., BARBER, B.H., FLAVELL, R.A. & ALLEN, H. (1989) Role of p2m in the intracellular transport and surface expression of murine class I histocompatibility molecules. Journalof Immunology, 142,2796.
XIN, J.-H., KVIST, S . & DOBBERSTEIN, B. (1982) Identification of an H-2Kd gene using a specific DNA probe. EMBO Journal, 1,467.
YOUNG, P.R. & TILGHMAN, S.J. (1984) Induction of a-fetoprotein synthesis in differentiating F9 teratocarcinoma cell is accompanied by a genome-wide loss of DNA methylation. Molecular and Cellular Biology, 4, 898.
ZALMAN, M.A. & MERUELO, D. (1986) Analysis of H-2-linked immune responses involved in resistance to AKR tumor growth. Immunogenetics, 24,51.
ZINKERNAGEL, R.M. & DOHERTY, P.C. (1979) MHC-restricted cytotoxic T cells: Studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction-specific, function, and responsiveness. Advances in Immunology, 27,51.
Copyright © 2022 FDOKUMEN




















