
Curcumin protects against acute liver damage in the rat by inhibiting NF-κB, proinflammatory...
8
Curcumin protects against acute liver damage in the rat by inhibiting NF-κB, proinflammatory cytokines production and oxidative stress Karina Reyes-Gordillo a , José Segovia b , Mineko Shibayama c , Paula Vergara b , Mario G. Moreno a , Pablo Muriel a, ⁎ a Sección Externa de Farmacología, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. México b Departamento de Fisiología, Biofísica y Neurociencias, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. México c Departamento de Patología Experimental, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. México Received 8 December 2006; received in revised form 7 February 2007; accepted 8 February 2007 Available online 22 February 2007 Abstract Curcumin, an anti-inflammatory and antioxidant compound, was evaluated for its ability to suppress acute carbon tetrachloride-induced liver damage. Acute hepatotoxicity was induced by oral administration of CCl 4 (4 g/kg, p.o.). Curcumin treatment (200 mg/kg, p.o.) was given before and 2 h after CCl 4 administration. Indicators of necrosis (alanine aminotransferase) and cholestasis (γ-glutamyl transpeptidase and bilirubins) resulted in significant increases after CCl 4 intoxication, but these effects were prevented by curcumin treatment. As an indicator of oxidative stress, GSH was oxidized and the GSH/GSSG ratio decreased significantly by CCl 4 , but was preserved within normal values by curcumin. In addition to its antioxidants properties, curcumin is capable of preventing NF-κB activation and therefore to prevent the secretion of proinflammatory cytokines. Therefore, in this study we determined the concentrations of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) mRNA, and NF-κB activation. CCl 4 -administered rats depicted significant increases in TNF-α, IL-1β, and IL-6 production, while curcumin remarkably suppressed these mediators of inflammation in liver damage. These results were confirmed by measuring TNF-α, and IL-1β protein production using Western Blot analysis. Accordingly, these proteins were increased by CCl 4 and this effect was abolished by curcumin. Administration of CCl 4 induced the translocation of NF-κB to the nucleus; CCl 4 induced NF-κB DNA binding activity was blocked by curcumin treatment. These findings suggest that curcumin prevents acute liver damage by at least two mechanisms: acting as an antioxidant and by inhibiting NF-κB activation and thus production of proinflammatory cytokines. © 2007 Elsevier B.V. All rights reserved. Keywords: Curcumin; Cytokines; Liver injury; Carbon tetrachloride; Necrosis; NF-κB. 1. Introduction Liver diseases constitute a major problem of world wide proportions. CCl 4 is a well known hepatotoxin that is widely used to induce acute-toxic liver injury in a large range of laboratory animals [1]. Acute hepatotoxicity induces inflamma- tion, necrosis, and oxidative stress of hepatocytes [2]. In the last few years, it has been suggested that in most cases hepatocellular injury is not due to the damaging agent itself but to the inflammatory cells that have been attacked by the stressed hepatocytes [3]. Inflammatory response is mediated by the action of cytokines, mainly tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) and these cytokines can induce an acute phase response or modulate the effects of interleukin-6 (IL-6) [4]. The importance of inflammatory cytokines as hepatotrophic factors is demonstrated by evidence that circulating levels of TNF-α and IL-1β are increased in rats that develop liver damage [5]. It has been shown that inflammatory molecular events are regulated following activa- tion of nuclear factor (NF)-κB by CCl 4 release of TNF-α, and IL-1β, [6]. NF-κB is a cardinal regulator of inflammatory response, controlling the expression of genes encoding cytokines [7]. NF-κB is also known to activate the transcription of TNF-α-inducible genes, including IL-1β [8]. Thus, a vicious cycle is established in the hepatocytes: TNF-α, and IL-1β Biochimica et Biophysica Acta 1770 (2007) 989 – 996 www.elsevier.com/locate/bbagen ⁎ Corresponding author. Tel.: +52 55 5061 3303; fax: +52 55 5061 3394. E-mail address: [email protected] (P. Muriel). 0304-4165/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.bbagen.2007.02.004
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Curcumin protects against acute liver damage in the rat by inhibiting NF-κB, proinflammatory...

a 1770 (2007) 989–996www.elsevier.com/locate/bbagen
Biochimica et Biophysica Act
Curcumin protects against acute liver damage in the rat by inhibiting NF-κB,proinflammatory cytokines production and oxidative stress
Karina Reyes-Gordillo a, José Segovia b, Mineko Shibayama c, Paula Vergara b,Mario G. Moreno a, Pablo Muriel a,⁎
a Sección Externa de Farmacología, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. Méxicob Departamento de Fisiología, Biofísica y Neurociencias, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. México
c Departamento de Patología Experimental, Cinvestav-IPN., Apdo. Postal 14-740. México 07000, D.F. México
Received 8 December 2006; received in revised form 7 February 2007; accepted 8 February 2007Available online 22 February 2007
Abstract
Curcumin, an anti-inflammatory and antioxidant compound, was evaluated for its ability to suppress acute carbon tetrachloride-induced liverdamage. Acute hepatotoxicity was induced by oral administration of CCl4 (4 g/kg, p.o.). Curcumin treatment (200 mg/kg, p.o.) was given beforeand 2 h after CCl4 administration. Indicators of necrosis (alanine aminotransferase) and cholestasis (γ-glutamyl transpeptidase and bilirubins)resulted in significant increases after CCl4 intoxication, but these effects were prevented by curcumin treatment. As an indicator of oxidativestress, GSH was oxidized and the GSH/GSSG ratio decreased significantly by CCl4, but was preserved within normal values by curcumin. Inaddition to its antioxidants properties, curcumin is capable of preventing NF-κB activation and therefore to prevent the secretion ofproinflammatory cytokines. Therefore, in this study we determined the concentrations of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β),and interleukin-6 (IL-6) mRNA, and NF-κB activation. CCl4-administered rats depicted significant increases in TNF-α, IL-1β, and IL-6production, while curcumin remarkably suppressed these mediators of inflammation in liver damage. These results were confirmed by measuringTNF-α, and IL-1β protein production using Western Blot analysis. Accordingly, these proteins were increased by CCl4 and this effect wasabolished by curcumin. Administration of CCl4 induced the translocation of NF-κB to the nucleus; CCl4 induced NF-κB DNA binding activitywas blocked by curcumin treatment. These findings suggest that curcumin prevents acute liver damage by at least two mechanisms: acting as anantioxidant and by inhibiting NF-κB activation and thus production of proinflammatory cytokines.© 2007 Elsevier B.V. All rights reserved.
Keywords: Curcumin; Cytokines; Liver injury; Carbon tetrachloride; Necrosis; NF-κB.
1. Introduction
Liver diseases constitute a major problem of world wideproportions. CCl4 is a well known hepatotoxin that is widelyused to induce acute-toxic liver injury in a large range oflaboratory animals [1]. Acute hepatotoxicity induces inflamma-tion, necrosis, and oxidative stress of hepatocytes [2]. In the lastfew years, it has been suggested that in most caseshepatocellular injury is not due to the damaging agent itselfbut to the inflammatory cells that have been attacked by thestressed hepatocytes [3]. Inflammatory response is mediated by
⁎ Corresponding author. Tel.: +52 55 5061 3303; fax: +52 55 5061 3394.E-mail address: [email protected] (P. Muriel).
0304-4165/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.bbagen.2007.02.004
the action of cytokines, mainly tumor necrosis factor-α (TNF-α)and interleukin-1β (IL-1β) and these cytokines can induce anacute phase response or modulate the effects of interleukin-6(IL-6) [4]. The importance of inflammatory cytokines ashepatotrophic factors is demonstrated by evidence thatcirculating levels of TNF-α and IL-1β are increased in ratsthat develop liver damage [5]. It has been shown thatinflammatory molecular events are regulated following activa-tion of nuclear factor (NF)-κB by CCl4 release of TNF-α, andIL-1β, [6]. NF-κB is a cardinal regulator of inflammatoryresponse, controlling the expression of genes encodingcytokines [7]. NF-κB is also known to activate the transcriptionof TNF-α-inducible genes, including IL-1β [8]. Thus, a viciouscycle is established in the hepatocytes: TNF-α, and IL-1β

990 K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
promotes NF-κB activation, and NF-κB leads to enhancedproduction of additional TNF-α, and IL-1β. This cycleeventually alters the structure of the hepatocytes, and impairstheir function [9], consequently, prolonged activation of NF-κBleads to perpetuated inflammatory responses and therefore thistranscription factor is a target for a range of anti-inflammatorydrugs used to treat different diseases [8].
Curcumin (1,7-bis(4-hidroxy-3-methoxyphenyl)-1,6-hepa-diene-3,5-dione), obtained from Curcuma longa L. (Zengiber-aceae family) rhizomes, has been widely used in indigenousmedicine for centuries and has shown a variety of physiologicaland pharmacological activities [10]. Interestingly, curcumin wasshown to inhibit the production of various cytokines, includingTNF-α and IL-1β [11]. In addition, studies involving systemicadministration of curcumin have shown its beneficial effects ona variety of inflammatory conditions by modulating NF-κBactivity [12]. The other salient feature of curcumin is thatexhibits strong antioxidant activity, comparable to vitamin Cand E, and it was shown to be a potent scavenger of a variety ofreactive oxygen species [13].
These observations prompted us to investigate whethercurcumin might play a protective role against necrosis andinflammation induced by CCl4 intoxication by blocking NF-κBactivation, the release of inflammatory cytokines and thecytotoxic free radicals. We conclude that curcumin preventsNF-κB activation, cytokine production, oxidative stress andliver damage induced by CCl4 in the rat.
2. Materials and methods
2.1. Chemicals
Curcumin, carboxymethylcellulose, thiobarbarbituric acid, γ-glutamyl-p-nitroanilide, L-γ-glutamyl-p-nitroaniline, p-nitrophenyl phosphate and bovineserum albumin were purchased from the Sigma Chemical Company, (St. Louis,MO, USA). Carbon tetrachloride, sodium acetate, sodium hydroxide, glacialacetic acid, hydrochloric acid, sulfuric acid, ethanol, methanol, potassium hydro-xide and formaldehyde were obtained from J.T. Backer (Xalostoc, Mexico).
2.2. Study design
A total of 24 male Wistar rats (weighing 250–300 g) were used in this study.They were maintained on standard rat chow diet with free access to drinkingwater. The animals were housed four or five per polycarbonate cage and werekept under controlled conditions of 22±2 °C, 50–60% relative humidity and 12-h light–dark cycles. All animals received human care and the study complieswith the institution's guidelines and the Mexican official regulation (NOM-062-ZOO-1999) regarding technical specifications for production, care and use oflaboratory animals.
Rats were divided into 4 groups of 6 rats each. Group I consisted of controlanimals that received vehicle only (0.1 mL of mineral oil, p.o.) which served ascontrol. In group II, acute liver damage was produced by o.p. administration ofCCl4 (4 g/kg) dissolved in mineral oil. In group III the rats received CCl4 asmentioned above and also received oral curcumin treatment (200 mg/kg) 2 hbefore and 2 h after CCl4 administration. Finally rats in group IV received thevehicle only as in group I and curcumin before and after vehicle administration.
2.3. Biochemical estimations
Animals were sacrificed 24 h after CCl4 administration under light etheranesthesia; blood was collected by cardiac puncture and the liver was rapidlyremoved. Serumwas obtained for determination of liver damage by measuring the
activities of γ-glutamyl transpeptidase (γ-GTP) [14], alanine aminotransferase(ALT) [15], and bilirubins content (kit Bioxon-Mexico).
2.4. Assessment of lipid peroxidation
The extent of lipid peroxidation was esteemed in liver homogenates bymeasurement of malondialdehyde (MDA) formation using the thiobarbituricacid method [16]. Protein was determined according to Bradford [17] usingbovine serum albumin as standard.
2.5. Reduced and oxidized glutathione determinations in liver
GSH and GSSG were prepared daily in 0.1 M sodium phosphate 0.005 MEDTA buffer (pH 8.0) and kept on ice until used. OPT solution was prepareddaily in reagent-grade absolute methanol just prior to use.
Liver samples were homogenized on ice using a polytron homogenizer. Thesolution used for homogenization consisted of 3.75 mL of the phosphate-EDTAbuffer and 1 mL of 25% H3PO4, which was used as a protein precipitant. Thetotal homogenate was centrifuged at 4 °C at 100,000×g for 20 min to obtain thesupernant for the assay of GSH and GSSG.
Determination of GSH was performed accordingly to Hissin and Hilf [18].To 0.5 mL of the 100,000×g supernatant, 4.5 mL of the phosphate-EDTAbuffer pH 8.0 was added. The final assay mixture (2.0 mL) contained 100 μLof the diluted tissue supernatant, 1.8 mL of phosphate-EDTA buffer, and100 μL of the OPT solution, containing 100 μg of OPT. After thoroughmixing and incubation at room temperature for 15 min, the solution wastransferred to a quartz cuvette. Fluorescence at 420 nm was determined withthe activation at 350 nm.
In GSSG assay, O.5-mL portion of the original 100,000× supernatant wasincubated at room temperature with 200 μL of 0.04 M N-ethylmaleimida(NEM) for 30 min to interact with GSH present in the tissue. To this mixture,4.3 mL of 0.1 N NaOH was added. A 100-μL portion of this mixture wastaken for measurement of GSSG, using outlined above for GSH assay, exceptthat 0.1 N NaOH was employed as diluent rather than phosphate-EDTAbuffer.
2.6. Western blot assays
The TriPure reagent (Roche Diagnostics) was used to isolate total proteinfrom samples of liver tissue. Fresh tissue was homogenized in 1 mL of TriPurereagent, then, 0.2 mL of chloroform was added to homogenates, the lowerphase was treated with isopropanol to precipitate total protein. Samples werecentrifuged at 12,000 rpm for 10 min at 4 °C, then, 3 washes were performedwith 0.3 M guanidine hydrochloride in 95% ethanol. A final wash wasperformed with 100% ethanol, samples were centrifuged as previouslydescribed, and the pellet resuspended in 1% SDS. Volumes equivalent to50 μg of proteins were transferred on 12% polyacrylamide gel; separatedproteins were transferred onto nitrocellulose paper (Amersham). Next, blotswere blocked with 5% skim milk and 0.05% Tween-20 for 30 min at roomtemperature and independently incubated overnight at 4 °C with antibodiesselective against each cytokine isoform, TNF-α, and IL-1β, (AD1837P andAB1832 respectively from Chemicon Int. Inc.). The following day, themembranes were washed and then exposed to a secondary peroxidase-labeledantibody (Zymed) diluted 1:4000 in the blocking solution for 1 h at roomtemperature. Blots were washed and protein developed using the ECL detectionsystem (NEN Life Sciences products). Blots were stripped and incubated with amonoclonal antibody directed against β-actin, which was used as a control tonormalize cytokine protein expression levels. The procedure to stripmembranes was as follows. First, blots were washed four times withphosphate-saline buffer pH 7.4 (0.015 M, 0.9% NaCl), then immersed instripping buffer (2-mercaptoethanol 100 mM, sodium dodecyl sulfate 2% andTris–HCl 62.5 mM, pH 6.7) for 30 min at 60 °C with gentle shaking,membranes were then washed five times with 0.05% Tween-20 in phosphate-saline buffer.
Images were digitalized using the BioDoc-It System (UVP) and thenanalyzed densitometrically using the Lab Works 4.0 Image Acquisition andAnalysis software (UVP), as previously described [19].

Table 1Effect of acute administration of carbon tetrachloride (CCl4) on serum alanine aminotransferase (ALT), γ-gamma-glutamyl transpeptidase (γ-GTP) activities, and totalbilirubins in control group (vehicle), CCl4-treated rats (CCl4), CCl4-rats treated with curcumin (CCl4+Curc.), and rats administered with curcumin alone (curcumin)
Parameter Group
Vehicle CCl4 CCl4+Curc. Curcumin
ALT (μmol/l min) 20.796±3.820 156.105±0.759a 94.087±4.894ab 22.332±1.560γ-GTP (μmol/l min) 4.868±0.740 20.987±2.948a 15.351±1.055ab 7.953±0.961BILIRUBINS (μmol/l) 11.821±0.979 48.514±4.152a 23.385±3.437ab 8.660±1.210
Values are means±S.E. (n=6) “a” means significantly different from control at p<0.05; “b” means significantly different from CCl4-rats treated at p<0.05.
991K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
2.7. RT-PCR analysis
Cytokine transcript levels of TNF-α, IL-1β and IL-6 in liver tissues of ratswere measured using a reverse transcriptase (RT)-PCR technique. Total RNAwas isolated using the TriPure reagent (Roche Diagnostics), RNA concentra-tions and purity were determined by measuring the absorbance A260 nm–A280 nm ratio, cDNA obtained by reverse transcriptase (Invitrogen), usingpolydt as a primer, and Taq polymerase was used for the PCR reactions(Invitrogen). For IL-1β mRNA, we used these primers: 5′ ATGGCAAC-TGTTCCTGAACTCAACT 3′ and 5′ CAGGACAGGTATAGATTC-TTTCCTTT 3′. Reactions were performed as follows: an initial step at 94 °Cfor 1 min 30 s was followed by 35 cycles (1 min at 94 °C, 1 min at 64 °C and1 min at 72 °C), and finally 3-min extension step at 72 °C. For TNF-α mRNA,we used these primers: 5′-TTCTGTCTACTGAACTTGGGGGTGATC-GGTCC-3′ and 5-′ GTATGAGATAGCAAATCGGCTGACGGTGTGGG-3′.Reactions were performed as follows: an initial step at 94 °C for 1 min 30 s wasfollowed by 35 cycles (1 min at 94 °C, 1 min at 66 °C and 1 min at 72 °C), andfinally 3-min extension step at 72 °C. For IL-6 mRNA, we used these primers:5′-ATGAAGTTCCTCTCTGCAAGAGACT-3′ and 5′-CACTAGGTTTGCC-GAGTAGATCTC-3′. Reactions were performed as follows: an initial step at94 °C for 1 min 30 s was followed by 35 cycles (1 min at 94 °C, 1 min at 62 °Cand 1 min at 72 °C), and finally 3-min extension step at 72 °C. To determineβ-actin, we used sequences and reaction conditions previously described [20].After PCR, products were electrophoresed on agarose gels, stained withethidium bromide, and images were recorded using a BioDoc-It System (UVP).
2.8. Nuclear protein isolation and shift assays for NF-κB
The isolation of intact and stable nuclei consisted in iso-osmotic lysisprocedure obtained as reported by Dyer et al. [21], With this lysis procedure, thenuclear envelope remains intact even during the further manipulations of
Fig. 1. Reduced (GSH) and oxidized (GSSG) glutathione, GSH/GSSG ratio and total gtreated rats (CCl4), CCl4 treated with curcumin (CCl4+Curc.), and rats administeexperiments performed in duplicate assays±S.E. (n=6). “a” Means significantly diffp<0.05.
washing, freezing and ultracentrifugation and provides nuclear extract for use inNF-κB assay.
NF-κB was estimated using a commercial kit (Non-Radioactive NF-κB p50/p65 Transcription Factor Assay, SGT510 Chemicon, Mexico City). This kitcombines the principle of the electrophoretic mobility shift assay (EMSA) withthe enzyme-linked immunosorbent assay (ELISA).
2.9. Histology
Samples were taken from all the animals and fixed with 10% formaldehydein phosphate-buffered saline for 24 h. Then, they were washed with tap water,dehydrated in alcohol and embedded in paraffin. Sections of 6–7 μm weremounted in glass slides covered with silane. Hematoxylin and eosin stains wereperformed in each slide.
2.10. Statistical analysis
Data are expressed as mean values±SE. Comparisons were carried out byanalysis of variance followed by Dunnett's or Tukey's test, as appropriate, usingSigma Stat for Windows (2.0 Version). Differences were considered statisticallysignificant when p<0.05.
3. Results
Alanine amino transferase (ALT), γ-glutamyl transpeptidase(γ-GTP) and total bilirubins are shown in Table 1. ALT is acytosolic enzyme of the hepatocyte and an increase of enzymeactivity reflects an increase in plasma membrane permeability,which, in turn, is associated with cell death [22]. In acutelyliver-damaged rats, serum levels of ALT were significantly
lutathione (GSH+GSSG) determined in livers from control rats (vehicle), CCl4-red with curcumin alone (curcumin). Each bar represents the mean value oferent from control, p<0.05. “b” Means significantly different from CCl4 group,

Fig. 2. Degree of lipid peroxidation expressed as the content of malondialdehyde(MDA) formation determined in livers from control rats (vehicle), CCl4-treatedrats (CCl4), CCl4 treated with curcumin (CCl4+Curc.), and rats administeredwith curcumin alone (curcumin). Each bar represents the mean value ofexperiments performed in duplicate assays±S.E. (n=6). “a”Means significantlydifferent from control, p<0.05.
992 K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
higher (7.5 times) than controls. Curcumin treatment signifi-cantly lowered serum activity of ALT. γ-GTP is an enzymeembedded in the hepatocyte plasma membrane, mainly in thecanalicular domain, and is considered to be one of the bestindicators of cholestasis [22]. γ-GTP serum activity wasstatistically significantly augmented 24 h after CCl4 intoxica-tion. In rats treated with curcumin+CCl4, serum γ-GTP levelswere significantly lower than in rats treated with CCl4 alone,indicating the occurrence of an active protective process. Totalbilirubins were increased after CCl4 intoxication indicatingacute cholestasis, but curcumin treatment prevented signifi-cantly this effect. In rats treated with vehicle (mineral oil), theserum biochemical parameters were not significantly differentwhen compared with rats treated with curcumin alone.
As an indicator of oxidative stress at the hydrophilic level,we measured glutathione in the liver. Reduced (GSH), oxidized(GSSG), GSH/GSSG ratio and total (GSH+GSSG) liverglutathione are show in Fig. 1. Reduced glutathione decreasedsignificantly in the CCl4-treated rats, while curcumin adminis-
Fig. 3. Hematoxylin and eosin staining of liver sections from: (A) control rats (vehicwith curcumin alone. (100×).
tration prevented the decrement in GSH induced by CCl4.Moreover, the group receiving both curcumin and CCl4 showedsignificantly higher levels of GSH than control. In the groupreceiving CCl4 and curcumin, GSSG increased significantly.The GSH/GSSG ratio decreased following CCl4 administration,while curcumin preserved it at normal levels. Total glutathione(GSH+GSSG) decreased significantly in CCl4-treated rats, butincreased significantly in the group that received both curcuminand CCl4, and in rats administered with curcumin alone.
Lipid peroxidation is known to play a critical role in CCl4induced liver injury [23] and malondialdehyde (MDA) is one ofits end products. Thus, measurement of MDA can be used toassess lipid peroxidation. Liver MDA content increasedsignificantly by acute CCl4 administration. Interestingly,curcumin was not capable of preventing the increase in lipidperoxidation (Fig. 2).
Liver injury was also evaluated by a histological approach(Fig. 3). Acute liver damage was accompanied by extendednecrotic areas and distortion of liver around portal triads (panelB), as compared with control (panel A). Treatment of CCl4-group with curcumin (panel C) resulted in the absence ofnecrotic areas and alterations observed with CCl4 alone,compare panel C with panel B (CCl4-group). This histopatho-logical analysis is in agreement with the observed result in theALT activity, a biochemical indicator of necrosis that wassignificantly prevented by curcumin (Table 1).
Extensive studies have revealed key genes mediating liverinjury. Genes regulating hepatocellular necrosis and inflamma-tion influence the extent of hepatic damage [24]. We firstexamined the effect of curcumin on acutely liver-damaged ratsproinflammatory gene expression by measuring TNF-α, IL-1βand IL-6 mRNA accumulation by RT-PCR (Fig. 4). The resultsshow concomitant increases in TNF-α, IL-1β, and IL-6mRNA in livers derived from CCl4-treated rats. Importantly,
le); (B) CCl4-treated rats; (C) CCl4 treated with curcumin; (D) rats administered
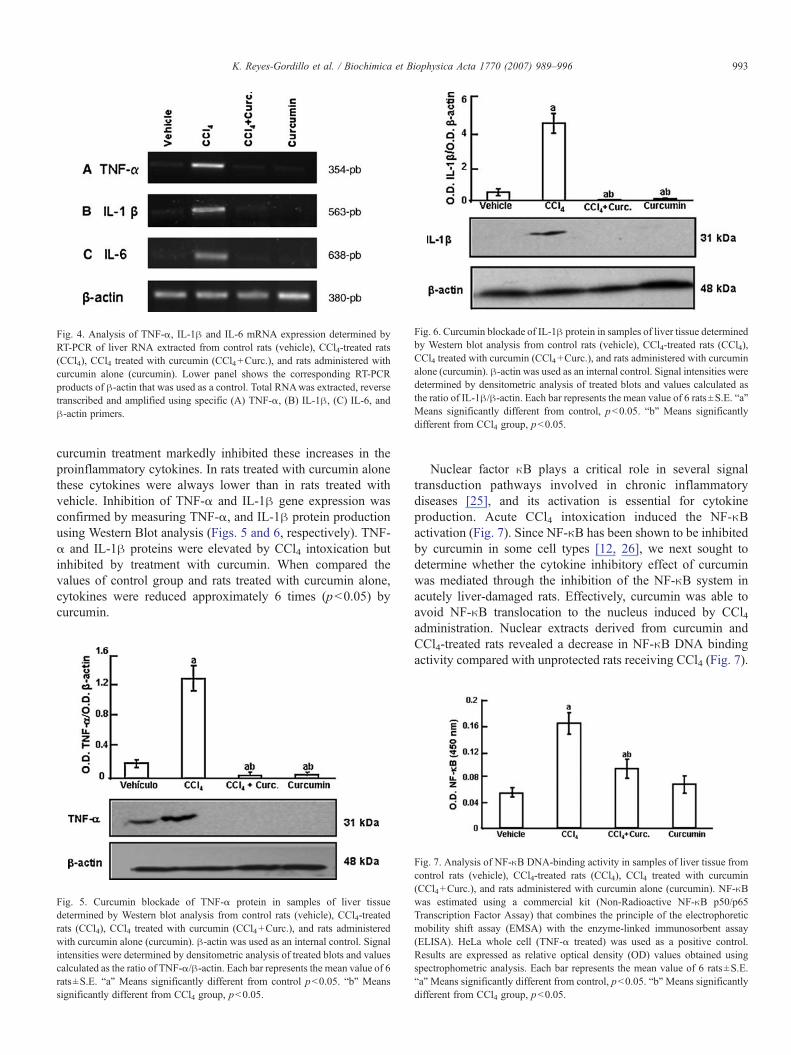
Fig. 4. Analysis of TNF-α, IL-1β and IL-6 mRNA expression determined byRT-PCR of liver RNA extracted from control rats (vehicle), CCl4-treated rats(CCl4), CCl4 treated with curcumin (CCl4+Curc.), and rats administered withcurcumin alone (curcumin). Lower panel shows the corresponding RT-PCRproducts of β-actin that was used as a control. Total RNAwas extracted, reversetranscribed and amplified using specific (A) TNF-α, (B) IL-1β, (C) IL-6, andβ-actin primers.
Fig. 6. Curcumin blockade of IL-1β protein in samples of liver tissue determinedby Western blot analysis from control rats (vehicle), CCl4-treated rats (CCl4),CCl4 treated with curcumin (CCl4+Curc.), and rats administered with curcuminalone (curcumin). β-actin was used as an internal control. Signal intensities weredetermined by densitometric analysis of treated blots and values calculated asthe ratio of IL-1β/β-actin. Each bar represents the mean value of 6 rats±S.E. “a”Means significantly different from control, p<0.05. “b” Means significantlydifferent from CCl4 group, p<0.05.
993K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
curcumin treatment markedly inhibited these increases in theproinflammatory cytokines. In rats treated with curcumin alonethese cytokines were always lower than in rats treated withvehicle. Inhibition of TNF-α and IL-1β gene expression wasconfirmed by measuring TNF-α, and IL-1β protein productionusing Western Blot analysis (Figs. 5 and 6, respectively). TNF-α and IL-1β proteins were elevated by CCl4 intoxication butinhibited by treatment with curcumin. When compared thevalues of control group and rats treated with curcumin alone,cytokines were reduced approximately 6 times (p<0.05) bycurcumin.
Fig. 5. Curcumin blockade of TNF-α protein in samples of liver tissuedetermined by Western blot analysis from control rats (vehicle), CCl4-treatedrats (CCl4), CCl4 treated with curcumin (CCl4+Curc.), and rats administeredwith curcumin alone (curcumin). β-actin was used as an internal control. Signalintensities were determined by densitometric analysis of treated blots and valuescalculated as the ratio of TNF-α/β-actin. Each bar represents the mean value of 6rats±S.E. “a” Means significantly different from control p<0.05. “b” Meanssignificantly different from CCl4 group, p<0.05.
Nuclear factor κB plays a critical role in several signaltransduction pathways involved in chronic inflammatorydiseases [25], and its activation is essential for cytokineproduction. Acute CCl4 intoxication induced the NF-κBactivation (Fig. 7). Since NF-κB has been shown to be inhibitedby curcumin in some cell types [12, 26], we next sought todetermine whether the cytokine inhibitory effect of curcuminwas mediated through the inhibition of the NF-κB system inacutely liver-damaged rats. Effectively, curcumin was able toavoid NF-κB translocation to the nucleus induced by CCl4administration. Nuclear extracts derived from curcumin andCCl4-treated rats revealed a decrease in NF-κB DNA bindingactivity compared with unprotected rats receiving CCl4 (Fig. 7).
Fig. 7. Analysis of NF-κB DNA-binding activity in samples of liver tissue fromcontrol rats (vehicle), CCl4-treated rats (CCl4), CCl4 treated with curcumin(CCl4+Curc.), and rats administered with curcumin alone (curcumin). NF-κBwas estimated using a commercial kit (Non-Radioactive NF-κB p50/p65Transcription Factor Assay) that combines the principle of the electrophoreticmobility shift assay (EMSA) with the enzyme-linked immunosorbent assay(ELISA). HeLa whole cell (TNF-α treated) was used as a positive control.Results are expressed as relative optical density (OD) values obtained usingspectrophometric analysis. Each bar represents the mean value of 6 rats±S.E.“a”Means significantly different from control, p<0.05. “b”Means significantlydifferent from CCl4 group, p<0.05.

994 K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
4. Discussion
In the present study we found that acute CCl4 intoxicationinduced liver damage, measured by increased serum markers ofhepatic injury and histology. The damage was associated with adecreased hepatic GSH/GSSG ratio, indicating oxidative stress.In addition, NF-κB was activated and proinflammatorycytokines regulated by this factor, TNF-α, IL-1β and IL-6were very high. Curcumin treatment prevented all the altera-tions produced by CCl4. We believe that the antioxidantproperties of curcumin [13] and its ability to inactivate NF-κB[12], and thus proinflammatory cytokines production [11], arethe most likely mechanisms of action of curcumin.
Carbon tetrachloride (CCl4) once was used widely as asolvent, cleaner, and degreaser both for industrial and home use.Today, the scientific database on effects of haloalkanes is so vastthat it is no longer used but as a model of experimental liverinjury [6]. Today CCl4 proves highly useful as an experimentalmodel for the study of certain hepatotoxic effects [1,23]. Itconsistently produces liver injury in many species, includingnon-human primates [27].
CCl4-metabolism begins with the formation of the trichlor-omethyl free radical, CCl3
· through the action of the mixedfunction cytochrome P450 oxygenase system of the endoplas-mic reticulum. This process involves reductive cleavage of acarbon-chlorine bond. The major cytochrome isozyme toexecute biotransformation of CCl4 is CYP2E1, but CYP2B1and CYP2B2 are also able to attack the CCl4 molecule [1].There is no evidence that curcumin inhibits CCl4 activation.
The CCl3U, in the presence of oxygen, is transformed into the
trichloromethyl peroxy radical, CCl300U. This radical is more
reactive and thus shorter lived than the CCl3· radical [28]. The
lifetime of the CCl300Uradical is in the millisecond range and
disappears from the tissue by reacting with suitable substrates tocomplete its electron pair. CCl300
Uis more likely than CCl3
Uto
abstract hydrogen from polyunsaturated fatty acids (PUFA) [29]leading to lipid peroxidation [23]. The abstraction of hydrogenfrom PUFA starts sequential reactions that finish in thecomplete disintegration of the fatty acid molecules with theconsequent formation of aldehydes, other carbonyls, andalkanes in a process called lipid peroxidation. It is importantto point out that lipid peroxidation may produce injury bycompromising the integrity of membranes and by covalentbinding of reactive intermediates to important biologicalmolecules like GSH; finally the process leads to necrosis andliver damage in general. Therefore, free radicals scavengers andantioxidants may interfere with the toxicity of CCl4.
Curcumin is a representative polyphenolic compound foundin the dietary spice turmeric. Among a wide range of biologicaland biochemical activities, curcumin therapeutic effects arebeing investigated on well-defined models of disease, such asinflammatory renal, pancreatic, and pulmonary diseases [30–32]. Recently, the protective effect of curcumin in rat liverinjury induced by CCl4 was demonstrated by Park et al. [33]where curcumin administration prevented ALT, aspartateaminotransferase, and alkaline phosphatase increases. Inagreement with Park et al. [33] results, we also observed
hepatoprotection by curcumin; however in that work the actionmechanism of curcumin was not investigated as we did in thepresent work.
Carbon tetrachloride decreased GSH and GSH/GSSG ratioindicating oxidative stress at the hydrophilic level. Curcumin, awell known antioxidant [13], not only prevented thesedecrements but in fact elevated them, and total (GSH+GSSG)glutathione over controls. These results suggest that curcumin isincreasing de novo synthesis of glutathione. Supporting ourobservations, Sharma et al. [28] and Singh et al. [34] haveshown that curcumin increases glutathione S-transferaseactivity. This enzyme is involved in the synthesis of GSH;therefore providing an explanation for our results.
The present work shows that lipid peroxidation (measured asMDA) increased significantly by acute CCl4 administration.However, curcumin was not able to prevent MDA production.The most likely explanation is that oral consumption ofcurcumin in rats resulted in approximately 75% being excretedin the feces and only traces appeared in the urine, suggestingpoor absorption of curcumin. It has been shown that curcumin isbio-transformed to dihydrocurcumin, tetrahidrocurcumin, andhexahidrocurcumin; subsequently, these products are convertedto glucuronide conjugates [13], which are more polar and havebetter absorption than curcumin [35]. Therefore, it is likely thatthe pharmacological actions of curcumin are caused by itshydrosoluble derivatives. Hydrophilic compounds with anti-oxidant properties are more likely to act on GSH than on lipids,where lipid peroxidation occurs, explaining the incapacity ofcurcumin to prevent lipid peroxidation.
On the other hand, the liver resembles a central organ ofcytokines activity; the production of cytokines in liver oftendepends upon the initial induction of early-response cytokinesreleased from tissue-resident macrophages (Kupffer cells) [2,4].Hepatotoxins such as CCl4 rapidly induce proinflammatorycytokines by Kupffer cells [36]. In this study curcumin has beenshown to inhibit the expression of a sequence of inflammatorycytokines such as TNF-α, IL-1β and IL-6 in CCl4-treated rats.In agreement with the results obtained herein, Surh [37], andJobin et al. [38], have demonstrated various anti-inflammatoryproperties of curcumin.
Jobin et al. [38] investigated the effect of curcumin onintestinal gene expression and determined the molecularmechanism of curcumin action on the Iκ/NF-κB signalingpathway. Their results demonstrate that curcumin blocks geneexpression by inhibiting the signal leading to IκB kinase (IKK)activation without directly interfering with NF-κB-inducingkinase or IKK kinase complexes. Blockade of IKK activationcauses inhibition of IκBα phosphorylation/degradation and NF-κB activation [38].
NF-κB plays a critical role in the transcriptional control ofexpression of proinflammatory genes in various cells. Cyto-kines mediated induction of NF-κB requires activation ofvarious kinases, which ultimately lead to the phosphorylationand degradation of IκB, the NF-κB cytoplasmic inhibitor [39].Several lines of evidence suggest that the induction of NF-κBdependent gene expression in Kupffer cells contributes to CCl4-induced liver injury. In support of this hypothesis, it would be

995K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
highly desirable to develop a pharmacological strategy tosuppress the activation of NF-κB. Although curcumin is knownto inhibit the activation of NF-κB and suppress inflammation[38,40], this is the first time that it has been shown thatcurcumin prevents NF-κB activation on acute liver damage. Inthe present work we showed that CCl4 leads to increased NF-κBbinding activity when compared with the control group.Interestingly, curcumin prevented the activation of NF-κB inCCl4-treated rats; consequently, this study suggests that thehepatoprotective effects of curcumin are mediated through theinhibition of the NF-κB system in acutely liver-damaged rats.However, other actions of curcumin such as its antioxidantproperties cannot be discarded.
In summary, this study demonstrates that curcumin washighly effective in preventing CCl4-acute liver damage, whichblocked oxidative stress, the expression of proinflammatorycytokines, and the activation of NF-κB, suggesting that agentsthat prevent the activation of transcription factor NF-κB maysuppress expression of a series of proinflammatory moleculesand thereby prevent liver injury. In addition, the food additivecurcumin has the advantage of being a non-toxic and non-mutagen natural product [10].
Acknowledgements
The authors express their gratitude to Mr. Benjamín SalinasHernández, and Mr. Ramón Hernández, for their excellenttechnical assistance. The authors also acknowledge I.F. EnriqueChavez for his help with nuclear protein isolation. M. Sc.Karina Reyes-Gordillo was fellow of Conacyt. This work wassupported in part by Conacyt Grant 42721.
References
[1] P. Muriel, Some experimental models of liver damage, in: S. Sahu (Ed.),Hepatotoxicity: from genomics to in vitro and in vivo models. WestSussex, UK: John Wiley and Sons LTD (in press).
[2] P. Muriel, Cytokines in liver diseases, in: S. Sahu (Ed.), Hepatotoxicity:from genomics to in vitro and in vivo models. West Sussex, UK: JohnWiley and Sons LTD (in press).
[3] G. Ramadori, T. Armbrust, Cytokines in the liver, Eur. J. Gastroenterol.Hepatol. 13 (2001) 777–784.
[4] K.J. Simpson, N.W. Lukacs, L. Colleti, R.M. Strieter, S.L. Kunkel,Cytokines and the liver, J. Hepatol. 27 (1997) 1120–1132.
[5] S.Q. Yang, H.Z. Lin, M. Yin, J.H. Albrecht, A.M. Diehl, Effect of chronicethanol consumption on cytokine regulation of liver regeneration, Am. J.Physiol. 275 (1998) G696–G704.
[6] L.W. Weber, M. Boll, A. Stampfl, Hepatotoxicity and mechanism of actionof haloalkanes: carbon tetrachloride as a toxicological model, Crit. Rev.,Toxicol. 33 (2003) 105–136.
[7] M. Pinzanie, F. Marra, V. Carloni, Signal transduction in hepatic stellatecells, Liver 18 (1998) 2–13.
[8] F. Oakley, J. Mann, S. Nailard, D.E. Smart, N. Mungalsingh, C.Constandinou, et al., Nuclear factor-(B1 (p50) limits the inflammatoryand fibrogenic responses to chronic injury, Am. J. Pathol. 166 (2005)695–708.
[9] M.G. Neuman, Cytokine-central factor in alcoholic liver disease, AlcoholRes. Health 27 (2003) 307–316.
[10] H.P. Ammon, M.A. Walh, Pharmacology of Curcuma longa, Planta Med.57 (1991) 1–7.
[11] Y. Abe, S. Hashimoto, T. Horie, Curcumin inhibition of inflammatory
cytokine production by human peripheral blood monocytes and alveolarmacrophages, Pharmacol. Res. 39 (1999) 41–47.
[12] S. Singh, B.B. Aggarwal, Activation of transcription factor NF-κB issuppressed by curcumin (dyferulolylmethane), J. Biol. Chem. 270 (1995)24995–25000.
[13] R.K. Maheswari, A.K. Singh, J. Gaddipati, R.C. Srimal, Multiplebiological activities of curcumin: a short review, Life Sci. 78 (2006)2081–2087.
[14] M. Glossman, D.M. Neville, Glutamyl transferase in kidney brush bordermembranes, FEBS Lett. 19 (1972) 340–344.
[15] S. Reitman, S. Frankel, A colorimetric method for determination of serumoxaloacetic and glutamic pyruvic transaminases, Am. J. Clin. Pathol. 28(1957) 56–63.
[16] H. Okawa, N. Ohmishi, K. Yagi, Assay for lipid peroxides in animaltissues by the thiobarbituric acid reaction, Anal. Biochem. 95 (1979)351–358.
[17] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein dyebinding, Anal. Biochem. 72 (1976) 248–254.
[18] P.J. Hissin, R. Holf, A fluorometric method for determination of oxidizedand reduced glutathione in tissues, Anal. Biochem. 74 (1976) 214–226.
[19] F. Pérez-Severiano, B. Escalante, P. Vergara, C. Ríos, J. Segovia, Age-dependent changes in nitric oxide synthase activity and protein expressionin striata of mice transgenic for the Huntington's disease mutation, BrainRes. 951 (2002) 36–42.
[20] D.L. Choi-Lundberg, M.C. Bohn, Ontogeny and distribution of glial cellline-derived neurotrophic factor (GDNF) mRNA in rat, Dev. Brain Res. 85(1995) 80–88.
[21] R.B. Dyer, N.K. Herzog, Isolation of intact nuclei for nuclear extractpreparation from a fragile B Lymphocyte cell line, BioTechniques 19(1995) 192–195.
[22] H.R. Rosen, H.R. Keeffe, Evaluation of abnormal liver enzymes, use ofliver test, and the serology of viral hepatitis, in: B.R. Bacon, A.M. DiBisceglie (Eds.), Liver Disease Diagnosis and Management, Churchill,New York, 2000, pp. 24–35.
[23] P. Muriel, Peroxidation of lipids and liver damage, in: S.I. Baskin, H.Salem (Eds.), Antioxidants, Oxidants, and Free radicals, Taylor andFrancis, Washington DC, 1997, pp. 237–257.
[24] R. Bataller, D.A. Brenner, Liver fibrosis, J. Clin. Invest. 115 (2005)209–218.
[25] S. Bengmark, Curcumin: an atoxic antioxidant and natural NF-κB,ciclooxygenase-2, lipooxygenase, and inducible nitric oxide synthaseinhibitor: a shield against acute and chronic diseases, J. Parenter. EnteralNutr. 30 (2005) 1–7.
[26] A. Kumar, S. Dhawan, N.J. Hardegen, B.B. Aggarwal, Curcumin(dyferulolylmethane) inhibition of tumor necrosis factor (TNF)-mediatedadhesion of monocytes to endothelial cells by suppression of cell surfaceexpression of adhesion molecules and of nuclear factor-κB activation,Biochem. Pharmacol. 55 (1998) 775–783.
[27] T. Yoshida, E. Adachi, H. Nigi, S. Fujii, M. Yanagi, Changes of sinusoidalbasement membrane collagens in early hepatic fibrosis induced with CCl4in cynomolgus monkeys, Pathology 31 (1999) 29–35.
[28] B.A. Mico, L.R. Pohl, Reductive oxygenation of carbon tetrachloride:trichloromethyl peroxy radical as a possible intermediate in the conversionof carbon tetrachloride to electrophilic chlorine, Arch. Biochem. Biophys.225 (1983) 596–609.
[29] L.G. Forni, J.E. Packer, T.F. Slater, R.L. Willson, Reaction of thetrichloromethyl and halothane-derived peroxy radicals with unsaturatedfatty acids: a pulse radiolysis study, Chem.-Biol. Interact. 45 (1983)171–177.
[30] J. Gaedeke, A.N. Noble, A.W. Border, Curcumin blocks multiple sites ofthe TGF-β signaling cascade in renal cells, Kidney Int. 66 (2004)112–120.
[31] R.A. Sharma, C.R. Ireson, R.D. Verschoyle, K.A. Hill, M.L. Williams, C.Leuratti, et al., Effects of dietary curcumin on glutathione S-transferase andmalondialdehyde-DNA adducts in rat liver and colon mucosa: relationshipwith drug levels, Clin. Cancer Res. 7 (2001) 1452–1458.
[32] D. Punithavathi, N. Venkatesan, M. Babu, Curcumin inhibition of

996 K. Reyes-Gordillo et al. / Biochimica et Biophysica Acta 1770 (2007) 989–996
bleomycin-induced pulmonary fibrosis in rats, Br. J. Pharmacol. 131(2000) 169–172.
[33] E.J. Park, C.H. Jeon, G. Ko, J. Kim, H.D. Sohn, Protective effect ofcurcumin in rat liver injury induced by carbon tetrachloride, J. Pharm.Pharmacol. 52 (2000) 437–440.
[34] S.V. Singh, X. Hu, S.K. Srivastava, M. Singh, H. Xia, J.L. Orchard, et al.,Mechanism of inhibition of benzo[a]pyrene-induced for stomach cancer inmice by dietary curcumin, Carcinogenesis 19 (1998) 1357–1360.
[35] M.H. Pan, T.M. Huang, J.K. Lin, Biotransformation of curcumin throughreduction and glucuronidation in mice, Drug Metab. Dispos. 27 (1999)486–494.
[36] A. Geier, S.K. Kim, T. Gerloff, C.G. Dietrich, F. Lammert, S.J. Karpen,et al., Hepatobiliary organic anion transporters are differentially regulatedin acute toxic liver injury induced by carbon tetrachloride, J. Hepatol. 37(2002) 198–205.
[37] Y.J. Surh, Anti-tumor promoting potential of selected spice ingredientswith antioxidative and anti-inflammatory activities: a short review, FoodChem. Toxicol. 40 (2002) 1091–1097.
[38] C. Jobin, C. Braham, M.P. Russo, B. Juma, A.S. Narula, D.A. Brenner,et al., Curcumin blocks cytokine-mediated NF-κB activation andproinflammatory gene expression by inhibiting inhibitory factor I-κBkinase activity, J. Immunol. 163 (1999) 3474–3483.
[39] P.M. Russo, B.L. Bennett, M.A. Manning, A.D. Brenner, Differentialrequirement for NF-κB inducing kinase in the induction of NF-κB by IL-1β, TNF-α, and Fas, Am. J. Physiol.: Cell Physiol. 283 (2002)C347–C357.
[40] S. Shishodia, M.H. Amin, R. Lai, B.B. Aggarwal, Curcumin (diferuloyl-methane) inhibit constitutive NF-κB activation, induces G1/S arrest.Suppresses proliferation, and induces apoptosis in mantle cell lymphoma,Biochem. Pharmacol. 70 (2005) 700–713.




















