
CTT1 overexpression increases life span of calorie-restricted Saccharomyces cerevisiae deficient in...
9
RESEARCH ARTICLE CTT1 overexpression increases life span of calorie-restricted Saccharomyces cerevisiae deficient in Sod1 Germana Rona • Ricardo Herdeiro • Cristiane Juliano Mathias • Fernando Araripe Torres • Marcos Dias Pereira • Elis Eleutherio Received: 14 September 2014 / Accepted: 5 January 2015 Ó Springer Science+Business Media Dordrecht 2015 Abstract Studies using different organisms revealed that reducing calorie intake, without malnutrition, known as calorie restriction (CR), increases life span, but its mechanism is still unkown. Using the yeast Saccharomyces cerevisiae as eukaryotic model, we observed that Cu, Zn-superoxide dismutase (Sod1p) is required to increase longevity, as well as to confer protection against lipid and protein oxidation under CR. Old cells of sod1 strain also presented a premature induction of apoptosis. However, when CTT1 (which codes for cytosolic catalase) was overexpressed, sod1 and WT strains showed similar survival rates. Fur- thermore, CTT1 overexpression decreased lipid per- oxidation and delayed the induction of apoptotic process. Superoxide is rapidly converted to hydrogen peroxide by superoxide dismutase, but it also under- goes spontaneous dismutation albeit at a slower rate. However, the quantity of peroxide produced from superoxide in this way is two-fold higher. Peroxide degradation, catalyzed by catalase, is of vital importance, because in the presence of a reducer transition metal peroxide is reduced to the highly reactive hydroxyl radical, which reacts indiscrimi- nately with most cellular constituents. These findings might explain why overexpression of catalase was able to overcome the deficiency of Sod1p, increasing life span in response to CR. Keywords Calorie restriction Ctt1 Life span Saccharomyces cerevisiae Sod1p Introduction According to oxidative stress theory of aging, first proposed by Harman in 1954, aging is correlated with the accumulation of cellular damages triggered by reactive oxygen species (ROS) produced by normal cell metabolism. As the organism ages, antioxidants decrease, increasing oxidative damage (Harman 2006). A large amount of data about the oxidative cumulative values in healthy individuals related to age has been shown (reviewed by del Valle 2011). In general, the oxidative stress theory of aging was confirmed, but some studies presented conflicting and contradictory results concerning the level of antioxi- dants in older volunteers (Mecocci et al. 2000; Ozbay and Dulger 2002). Due to the complexity of higher eukaryotic systems, it is appropriate to choose a relatively simple, but still relevant, model system for a G. Rona (&) R. Herdeiro C. J. Mathias M. D. Pereira E. Eleutherio Department of Biochemistry, Institute of Chemistry, Federal University of Rio de Janeiro, Avenida Athos da Silveira Ramos 149, Bloco A, Lab 547, Rio de Janeiro, RJ 21941-909, Brazil e-mail: [email protected] F. A. Torres Department of Cellular Biology, Institute of Biology, University of Brası ´lia, Brası ´lia, DF, Brazil 123 Biogerontology DOI 10.1007/s10522-015-9550-7
Transcript of CTT1 overexpression increases life span of calorie-restricted Saccharomyces cerevisiae deficient in...

RESEARCH ARTICLE
CTT1 overexpression increases life span of calorie-restrictedSaccharomyces cerevisiae deficient in Sod1
Germana Rona • Ricardo Herdeiro • Cristiane Juliano Mathias •
Fernando Araripe Torres • Marcos Dias Pereira •
Elis Eleutherio
Received: 14 September 2014 / Accepted: 5 January 2015
� Springer Science+Business Media Dordrecht 2015
Abstract Studies using different organisms revealed
that reducing calorie intake, without malnutrition,
known as calorie restriction (CR), increases life span,
but its mechanism is still unkown. Using the yeast
Saccharomyces cerevisiae as eukaryotic model, we
observed that Cu, Zn-superoxide dismutase (Sod1p) is
required to increase longevity, as well as to confer
protection against lipid and protein oxidation under
CR. Old cells of sod1 strain also presented a premature
induction of apoptosis. However, when CTT1 (which
codes for cytosolic catalase) was overexpressed, sod1
and WT strains showed similar survival rates. Fur-
thermore, CTT1 overexpression decreased lipid per-
oxidation and delayed the induction of apoptotic
process. Superoxide is rapidly converted to hydrogen
peroxide by superoxide dismutase, but it also under-
goes spontaneous dismutation albeit at a slower rate.
However, the quantity of peroxide produced from
superoxide in this way is two-fold higher. Peroxide
degradation, catalyzed by catalase, is of vital
importance, because in the presence of a reducer
transition metal peroxide is reduced to the highly
reactive hydroxyl radical, which reacts indiscrimi-
nately with most cellular constituents. These findings
might explain why overexpression of catalase was
able to overcome the deficiency of Sod1p, increasing
life span in response to CR.
Keywords Calorie restriction � Ctt1 � Life span �Saccharomyces cerevisiae � Sod1p
Introduction
According to oxidative stress theory of aging, first
proposed by Harman in 1954, aging is correlated with
the accumulation of cellular damages triggered by
reactive oxygen species (ROS) produced by normal
cell metabolism. As the organism ages, antioxidants
decrease, increasing oxidative damage (Harman
2006). A large amount of data about the oxidative
cumulative values in healthy individuals related to age
has been shown (reviewed by del Valle 2011). In
general, the oxidative stress theory of aging was
confirmed, but some studies presented conflicting and
contradictory results concerning the level of antioxi-
dants in older volunteers (Mecocci et al. 2000; Ozbay
and Dulger 2002). Due to the complexity of higher
eukaryotic systems, it is appropriate to choose a
relatively simple, but still relevant, model system for a
G. Rona (&) � R. Herdeiro � C. J. Mathias �M. D. Pereira � E. Eleutherio
Department of Biochemistry, Institute of Chemistry,
Federal University of Rio de Janeiro, Avenida Athos da
Silveira Ramos 149, Bloco A, Lab 547, Rio de Janeiro,
RJ 21941-909, Brazil
e-mail: [email protected]
F. A. Torres
Department of Cellular Biology, Institute of Biology,
University of Brasılia, Brasılia, DF, Brazil
123
Biogerontology
DOI 10.1007/s10522-015-9550-7

deeper understanding of the role of oxidative stress in
longevity. Yeast can go through the aging process by
many ways, the most important are the chronological
(CLS) and replicative aging (RLS). Yeast CLS is a
measure of the survival of a non-dividing population
of cells (Parrella and Longo 2008). The RLS deter-
mines the number of divisions an individual mother
cell undergoes before death, the daughter cell, obtains
the undamaged material (Carmona-Gutierrez and
Buttner 2014).
In the past, studies using the yeast Saccharomyces
cerevisiae enabled major breakthroughs in the under-
standing of basic cellular and molecular processes
(Botstein and Fink 2011). Today, the use of yeast is
undergoing a ‘rebirth’ in both fundamental and
applied research. Studies of budding yeast have made
immense contributions to our understanding of the
aging process (Kaeberlein 2010). The reasons for this
success are experimental tractability, especially in
applying classical and molecular genetic methods to
associate genes with proteins and functions within the
cell.
Another reason for its applicability in aging-
related research is that some pathways involved in
degenerative diseases are conserved from yeast to
human. One-third of genes involved in human
diseases have S. cerevisiae orthologues. A common
strategy used to understand the molecular events of
diseases is the expression of human disease proteins
in yeast. S. cerevisiae also offers an attractive cellular
environment to investigate aging disease-related
proteins that have no apparent homologous counter-
part in yeast, holding great promises for the eluci-
dation of the molecular processes of these diseases
and the development of new therapies (Khurana and
Lindquist 2010; Mirisola et al. 2013). As the S.
cerevisiae is the most investigated eukaryotic micro-
organism, functional genomics techniques and bio-
informatics analysis of the transcriptome, proteome,
metabolome and interactome were originally devel-
oped using these yeast. This has resulted in dat-
abases, such as http://www.yeastgenome.org, that
represent valuable sources for research on aging.
Yeast cells are typically grown in media containing
high levels of glucose (2 %) and amino acids.
However, independent studies have shown that RLS
and CLS can be increased by reducing either glucose
or amino acid concentration (or both) (Mannarino
et al. 2008; Reverter-Branchat et al. 2007). In our
lab, we have used this microorganism to understand
the mechanism by which calorie restriction (CR)
increases life span (Mannarino et al. 2008; Huberts
et al. 2014). This intervention is able to extend the
life span of organisms ranging from yeast to mam-
mals and is thought to produce the higher increase in
human life expectancy (Baur 2010). However, the
potential significance of CR on human lifespan was
put in check in light of the recently published
research on CR in primates, which suggests that
reduction in calories intake does not affect life span
(Mattison et al. 2012). In 2009, another study con-
cluded that CR did extend life in rhesus monkeys
(Colman et al. 2009). Genetics probably explains part
of the variation between the monkey studies,
increasing the importance of the use of a less com-
plex model, such as yeast. Although the mechanism
by which CR increases life span is not completely
clear, it well known that oxidative damages are
reduced in calorie-restricted organisms (Agarwal
et al. 2005; Poljsak 2011). CR seems to induce
adaptive cell responses, which promote increased
resistance to adverse conditions, thus delaying the
onset of age-associated damages and extending life
span (Ribaric et al. 2012).
In this work, we analyzed if CTT1 overexpression,
which codes for a cytoplasmic catalase, would be able
to surpass Sod1p deficiency to increase life span of S.
cerevisiae in response to CR. This yeast, like most
other eukaryotes, possesses two isoforms of superox-
ide dismutase, the Cu/Zn-Sod1p and the Mn-Sod2p.
Both remove superoxide radical by converting it to O2
and H2O2, which must be further disproportionated by
catalases and peroxidases (Herrero et al. 2008).
Peroxide is very dangerous because in the presence
of some metals produces hydroxyl radical, one of the
most reactive and toxic free radical. While Sod2p
stays in mitochondria, Sod1p is found in cytoplasm,
nucleus and in the mitochondrial intermembrane
space, comprising the greatest proportion of the total
superoxide dismutase (Miao and St. Clair 2009).
Furthermore, Sod1p presents a pronounced physio-
logical significance and therapeutic potential, being
involved in diseases frequently associated with aging,
such as familial amyotrophic lateral sclerosis (FALS),
Parkinson’s disease and several neurological disorders
(Fukai and Ushio-Fukai 2011).
Biogerontology
123

Materials and methods
Yeast strains and plasmid construction
CTT1 was amplified from genomic DNA of wild-type
strain BY4741 (MAT a his3 leu2 met15 ura3—
Euroscarf, Germany) by PCR, using Platinum Taq
DNA Polymerase High Fidelity (Invitrogen) and
primers CTT1-F (50-ggatccAAATGAACGTGTTCG
GTAAAAAAG) and CTT1-R (50-ggatccTTAATTGG-
CACTTGCAATGGAC), including a BamHI site
(lower case letters). The PCR product was purified
using the illustra GFX PCR DNA and Gel Band
Purification kit (GE Healthcare) and cloned into
pGEM-T vector (Promega). After digestion with
BamHI, a 1.7 Kb purified amplicon was inserted into
the YEp351-PGK (de Moraes et al. 1995), previously
digested with BglII, to obtain pPGK-CTT1. Plasmids
were propagated in Escherichia coli XL10 Gold grown
on LB medium with 100 lg/ml ampicillin. Standard
molecular biology techniques were used (Sambrook
et al. 1989). Following, pPGK-CTT1, harboring CTT1
under control of the PGK1 promoter and LEU2 as
selectable marker, was used to transform both BY4741
and its isogenic sod1 strain (sod1::KanMX4—Euro-
scarf, Germany), using the lithium acetate protocol
(Gietz et al. 1992).In the same way BY4741 and its
isogenic sod1 were also transformed with the YEp351-
PGK vector to use as a control. Transformants were
selected in synthetic medium (0.67 % yeast nitrogen
base without aminoacids, 0.01 % histidine, 0.01 %
uracil, 0.01 % methionine and 2 % agar) supple-
mented with 2 % glucose (SD 2 %).
Growth conditions, life span and oxidative damage
For the experiments, cells were grown in liquid media
SD 2 % or SD 0.5 % (containing 0.5 % glucose instead
of 2 %), until the middle of exponential growth phase
(OD570 = 0.6), with the ratio of flask volume/medium
of 5/1, at 28 �C and 160 rpm. To analyze life span and
oxidative damage accumulation as a function of time,
cells were switched from culture media to water (non-
proliferating conditions) and incubated at 37 �C/
160 rpm (Mannarino et al. 2008, 2011). Cell viability
was determined over time by standard dilution plate
counts on solid YPD medium (2 % glucose, 1 % yeast
extract 2 % peptone). Viability was expressed as the
percentage of colony forming units. The oxidant-
sensitive probe 2,7-dichlorofluorescin diacetate
(DCFDA) was used to measure the level of oxidative
stress (Mannarino et al. 2008). Protein carbonylation
and lipid peroxidation were determined as previously
described (Adamis et al. 2007). Slot blot using an anti-
DNP antibody revealed ROS-dependent protein car-
bonylation. Lipid oxidation was measured by TBARS
(thiobarbituric acid reactive species) method, which
detects malondialdehyde, a final product of lipid
peroxidation. Cells were centrifuged and washed with
cold distilled water. The pellets were resuspended in
0.5 ml of 10 % TCA (w/v) followed by addition of
1.5 g of glass beads. The samples were lysed by 6
cycles of 20 s agitation on a vortex followed by 20 s on
ice. Extracts were centrifuged and the supernatant
mixed with 0.1 ml of 0.1 M EDTA and 0.6 ml of 1 %
(w/v) thiobarbituric acid prepared in 0.05 M NaOH.
The reaction mixture was incubated in a boiling water
bath for 15 min and, after cooling, the absorbance was
measured at 532 nm (Steels et al. 1994).
Enzyme activities and apoptotic markers
Extracts for enzymatic determinations were obtained by
the disruption of cells with glass beads in 0.1 M Tris–
HCl buffer, pH 6.0 (Pereira et al. 2003). Protein
concentration was determined according to (Stickland
1951), using bovine serum albumin as standard. The
catalase activity was determined based on H2O2
consumption (Aebi 1964). Apoptosis was analyzed by
caspase activity and Annexin V-FITC staining. Caspase
assays were based on the measurement of the hydrolysis
of the peptide substrate acetyl-Asp-Glu-Val-Asp
p-nitroanilide, resulting in the release of the p-nitroan-
iline (Caspase assay kit, CASP-3-C, Sigma-Aldrich,
USA). Annexin V conjugated with FITC fluorochrome
staining (CLONTECH Laboratories, Inc.) was per-
formed as previously described (Gomes et al. 2008). To
determine frequencies of phenotypes, at least 300 cells
of three independent experiments were evaluated.
Results and discussion
Irrespective of the model organism, C. elegans, Dro-
sophila or yeast, the reduction of oxidative damage is
one of the mechanisms that extend life span in response
to CR (Shimokawa et al. 2008). Oxygen consumption
and intracellular levels of ROS are higher in calorie-
Biogerontology
123

restricted than in non-restricted yeasts, since there is a
higher respiratory chain activity under CR (Agarwal
et al. 2005; Mesquita et al. 2010). Because of such mild
stress, some antioxidant systems are overexpressed
(Agarwal et al. 2005), making calorie-restricted yeasts
more efficient in the destruction of ROS.
In this work, to understand the biological mecha-
nism underlying the anti-aging action of CR, the
relationship between antioxidants systems and some
markers of oxidative stress in calorie-restricted yeasts
was investigated. For this, we used a mutant yeast
strain unable to express SOD1, containing or not the
high copy plasmid Yep351PGK-CTT1, which endows
cells with constitutive CTT1 overexpression. The
catalase overexpression was confirmed in the mutants
WT CTT1 and sod1 CTT1, under caloric restriction,
showing that both lineages presented similar catalase
activities (Table 1). These scavenging enzymes were
chosen because it was previously found that their
expressions are induced in CR (Agarwal et al. 2005).
Yeasts grew in media containing 2 or 0.5 % glucose
(to simulate CR) until the middle of exponential
growth phase and shifted to water for measuring how
long a cell can survive in a non-dividing state. This
strategy has been applied by us and other groups
(Fabrizio and Longo 2003; Harris et al. 2005;
Mannarino et al. 2008, 2011; Piper 2006). In the wild,
an environment that lacks nutrients is common for
microorganisms and even to certain mammals (hiber-
nation of bears and rodents). When yeast cells are
incubated in water, they are not starving, because they
metabolize the intracellular stored nutrients.
As expected, longevity of calorie-restricted cells was
significantly higher than non-restricted cells (Fig. 1). At
high levels of glucose (2 %), yeasts use glucose to obtain
energy exclusively via glycolysis, their cellular defense
systems are repressed and, for this reason, they are very
sensitive to a severe oxidative stress, as occurs during
incubation in water at 37 �C. Damages accumulate over
time in water because cell division and new synthesis are
not occurring. Furthermore, when cells are shifted to
37 �C, the accumulation of oxidative damages and
consequently the aging process are accelerated. Accord-
ing to previous results, differences in life span measured
at 28 �C are reproducible at 37 �C, i.e., the differences in
life span between a mutant and its parental strain are the
same at both temperatures but appear earlier at 37 �C
(Piper 2006; Mannarino et al. 2008, 2011). As can be seen
in Fig. 1a, both WT and sod1 mutant strains presented a
similar life span when incubated in water at 37 �C when
previously grown in 2 % glucose. This result had already
been observed when cells were grown and kept in rich
(YPD) media containing 2 % glucose at 28 �C for several
days: cells deficient in Sod1p showed a life span similar to
WT (Demir and Koc 2010). However, CR was not able to
increase cell longevity in the absence of Sod1p (Fig. 1b).
Next, it was investigated the level of intracellular
oxidation as well as the accumulation of oxidized
Fig. 1 Sod1p is required for CR-mediated CLS extension. Life
span of non-CR (a) or CR cells (b). Cells were grown in SD 2 %
(a) or SD 0.5 % medium (b), until the middle of exponential
growth phase and washed with water. For life span measure-
ments, these cultures were then transferred to water and
incubated at 37 �C/160 rpm. Cellular viability was measured
by standard dilution plate counts and expressed as the
percentage of the colony-forming units at time 0 h of incubation
in water. Values are mean ± SD of three independent
experiments
Table 1 Catalase activity
Strains U/mg ptn
WT (BY4741) 1.3 ± 0.1
sod1 1.5
WT CTT1 38.4 ± 0.5
sod1 CTT1 24.1 ± 1.0
Catalase activity was determined on the basis of H2O2
consumption as described in ‘‘Materials and methods’’
section. The results represent the mean ± standard deviation
of at least three independent experiments
Biogerontology
123

proteins and lipids during incubation of cells in water
(Fig. 2). Higher levels of protein oxidation or lipid
peroxidation accelerate the progress of aging and
neurodegenerative diseases (Radak et al. 2011). To
quantify the production of ROS, we used the DCFDA
assay. Intracellular fluorescence due to dichlorofluores-
cin oxidation has been generally considered as a
representation of cellular ROS concentration (Bar
2002), especially for measuring hydrogen peroxide
concentration (Carter et al. 1994). As can be seen in
Fig. 2a, ROS accumulation was higher in the mutant,
being more evident after 72 h in water. The accumula-
tion of ROS caused a high increase in the levels of protein
carbonyl groups in both strains but in sod1 mutant the
increase was much more impressive (Fig. 2b). Sod1p
deficiency also led to a higher increase in lipid oxidation
(Fig. 2c). Among the parameters used to analyze the
oxidative damage, the level of carbonylation showed the
highest increase in response to aging. It is known that
cells degrade oxidized proteins within hours and days,
whereas lipid peroxidation products are detoxified
within minutes (Dalle-Donnea et al. 2003). Thus, in
accordance with Agarwal et al. (2005) that correlated the
increased life span of calorie-restricted yeasts with the
induction of expression of scavengers, including SOD1,
by CR, our results demonstrated that calorie-restricted
cells of sod1 mutant strain died much sooner than WT,
which seems to be related to a higher accumulation of
oxidative damages to lipids and proteins.
The majority of ROS production occurs in the
electron transport chain of the mitochondria (Sheu
et al. 2006). In this process, one molecule of oxygen
receives four electrons and then can be reduced to
water, but some of the electrons escape to form ROS. In
higher eukaryotes, superoxide is thought to be gener-
ated when electrons are transferred to and from
ubiquinone, specifically at complex I and complex
III. S. cerevisiae lacks complex I, but has NADH
dehydrogenases located at the mitochondrial inner
membrane space able to catalyze the partial reduction
of oxygen to superoxide in this compartment (Herrero
et al. 2008). To maintain redox homeostasis, multiple
enzyme defense system, like superoxide dismutases
and catalases enzymes, is present in the cells. By the
action of Sod, superoxide, which stays in the compart-
ment in which it was generated, is rapidly converted to
oxygen and hydrogen peroxide. According to litera-
Fig. 2 Oxidative stress in calorie-restricted cells, measured as
fold increase in fluorescence (intracellular oxidation) (a),
protein carbonylation (b) or lipid peroxidation (c). Cells were
grown in SD 0.5 % medium, harvested in mid-log growth phase
and then stressed by incubating them in water at 37 �C/160 rpm.
Oxidative damages were analyzed over time. The results were
expressed as a ratio between fluorescence, lipid peroxidation or
protein carbonylation of stressed cells and control situation
(time zero of incubation in water) of each strain. The results
represent the mean ± SD of at least three independent
experiments. *p \ 0.05 (sod1 vs. WT)
Biogerontology
123

ture, superoxide dismutase is a major antioxidant in
yeast stationary phase, playing a very important role in
its survival (Longo et al. 1996). Stationary phase is
characterized by nutrient limitation, no cell division
and mitochondrial respiration. The fast loss of viability
of cells lacking Sod1p observed under normal but not
low aeration levels suggests that Sod2p is able to
prevent migration of ROS from the mitochondria to the
cytoplasm only under lower oxygen levels (Longo
et al. 1996). Peroxide produced by superoxide
dismutases must be fully reduced to water through
catalase or peroxidases to avoid production of hydro-
xyl radical, the most reactive and dangerous radicals.
Superoxide also undergoes spontaneous dismutation
albeit at a slower rate. However, the quantity of
peroxide produced from superoxide in this way is two-
fold higher (Fernandes et al. 2007). However, the
differences between the rates of conversion of the
enzymatic and non-enzymatic reactions, high rates of
peroxide production could be achieved from high
concentrations of superoxide (as under deficiency of
Sod1p) in proper intracellular conditions (pH, metal
concentration). Furthermore, the high sensitivity of the
sod1 mutant to oxidative stress is now largely attrib-
uted to its inability to assemble iron-sulfur proteins,
leading to high intracellular level of free iron, trigger-
ing to a higher possibility of Fenton reactions occur
(Srinivasan et al. 2000). Thus, next, we investigated if
catalase overexpression, one of the enzymes respon-
sible for the conversion of peroxide to water and
oxygen, might overcome Sod1p deficiency, allowing
sod1 mutants enjoy the benefits of CR. Yeast cells
overexpressing cytosolic catalase (WT CTT1 and sod1
CTT1) were obtained by introducing pPGK-CTT1 into
WT and sod1 strains. To test CTT1 overexpression, all
strains were grown in SD 0.5 % medium until mid-log
phase, a growth condition that increases life span in
water. As expected, catalase activity was very low and
practically the same in both WT and sod1 strains
(specific activity around 1 U/mg protein). However, in
sod1 CTT1 and WT CTT1 strains, catalase activities
were 20 to 30-fold higher than sod1 and WT stains,
respectively, confirming that cells possessing pPGK-
CTT1 overexpress CTT1 (Table 1).
b Fig. 3 Effect of CTT1 overexpression on life span (a), protein
carbonylation (b) or lipid peroxidation (c). WT CTT1 and sod1
CTT1 cells were grown in SD 0.5 % medium up to mid-log
growth phase, washed and stressed by shifting them to water at
37 �C/160 rpm. Cellular viability was measured by standard
dilution plate counts and expressed as the percentage of the
colony-forming units at time 0 h of incubation in water.
Oxidative damages were analyzed over time. The results were
expressed as protein carbonylation and lipid peroxidation of
stressed cells and control situation (time zero of incubation in
water) of each strain. Protein carbonylation was detected by a
slot blot using an anti-DNP antibody. The levels of lipid
peroxidation were determined through TBARS method and the
results were expressed as a ratio between the level of lipid
peroxidation of stressed cells and control situation (time zero of
stress—incubation in water) of each strain. Values are
mean ± SD of three independent experiments
Biogerontology
123
According to Fig. 3a, when catalase was overex-
pressed, calorie-restricted cells of sod1 and WT strains
showed similar survival rates and levels of oxidative
damage during incubation in water. The life span
showed by sod1 CTT1 strain was higher than its parental
sod1 (Fig. 1b). However, overexpression of CTT1 only
improve longevity of calorie-restricted WT cells after
48 h of incubation in water. After 24 h, 35 % of WT
CTT1 cells were still alive (Fig. 3a) whereas its parental
strain WT showed a survival rate twofold higher
(Fig. 1b). Recently, several findings indicate that ROS
may serve as essential signaling molecules that promote
metabolic health and longevity (Ristow and Schmeisser
2011). Conditions involved with ROS generation, such
as heat and oxidative mild-stresses, lead to Hsf activa-
tion (Liu and Thiele 1996) and Yap1 nuclear localiza-
tion (Rodrigues-Pousada et al. 2010). It is well known
that increased ROS levels are detrimental. However,
lowering ROS levels below the homeostatic set point
may interrupt the physiological role of oxidants in
cellular processes such as induction of defense systems
against stress. Consistently, an excess of antioxidant, as
possible occurred with WT CTT1 at the beginning of
incubation in water, might prevent these ROS signals
and interfere with the life-span-extending capabilities of
CR. Only after 48 h in a non-dividing condition, when
high levels of oxidative damage had accumulated
(Fig. 2), CTT1 overexpression was suitable to counter-
act and regulate overall ROS levels to maintain
physiological homeostasis. Corroborating the idea of
the role of ROS as second messengers involved in signal
transduction, it was recently shown that deficiency in
peroxisomal catalase extended chronological lifespan of
yeast by inducing elevated levels of hydrogen peroxide,
which, in turn, activated superoxide dismutases (Mesq-
uita et al. 2010). As can be seen in Fig. 3b and C,
overexpressing CTT1 sod1 strain pre-grown under CR
also showed lower increases in levels of protein
carbonyl groups and lipid peroxidation during incuba-
tion in water than its parental strain sod1 (Fig. 2b, c).
These result leads us to conclude that overexpression of
catalase is able to overcome the deficiency of Sod1p in
the mechanism in which CR prolongs life span.
Saccharomyces cerevisiae shows features of apop-
totic death during chronological aging, as a form of
removal of damaged cells (Herker et al. 2004). To
investigate whether apoptosis might contribute to the
lower longevity shown by sod1 cells, we analyzed two
apoptotic markers, caspase activity and externalization
of phosphatidylserine. In yeast, phosphatidylserine
externalization can be detected by FITC labeled annexin
V staining (Gomes et al. 2008). Concomitantly, cells
must be checked for membrane integrity by incubation
with propidium iodide. Caspases (Cysteine-requiring
Aspartate protease) are a family of proteases that
mediate cell death and are important to process of
apoptosis. Homologues of metazoan caspases have been
identified on the yeast genome and shown to be
implicated in hydrogen peroxide induced apoptosis
(Fabrizio and Longo 2008). Caspase 3 (also referred to
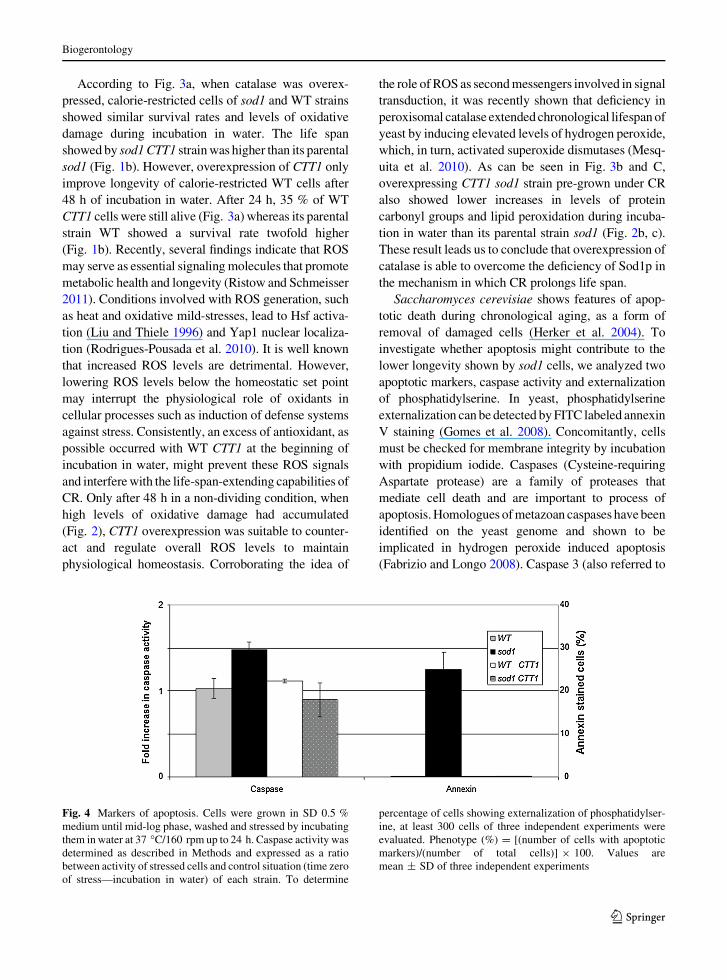
Fig. 4 Markers of apoptosis. Cells were grown in SD 0.5 %
medium until mid-log phase, washed and stressed by incubating
them in water at 37 �C/160 rpm up to 24 h. Caspase activity was
determined as described in Methods and expressed as a ratio
between activity of stressed cells and control situation (time zero
of stress—incubation in water) of each strain. To determine
percentage of cells showing externalization of phosphatidylser-
ine, at least 300 cells of three independent experiments were
evaluated. Phenotype (%) = [(number of cells with apoptotic
markers)/(number of total cells)] 9 100. Values are
mean ± SD of three independent experiments
Biogerontology
123

as CPP32, Yama and apopain) is a member of the CED-
3 subfamily of caspases and plays a central role in
mediating nuclear apoptosis including chromatin con-
densation and DNA fragmentation. According to our
results, apoptosis was induced earlier during incubation
in water in cells lacking Sod1p (Fig. 4). Caspase activity
was not induced in WT but increased 50 % in sod1
yeasts. In addition, more than 20 % of sod1 cells were
stained with annexin V, what was not observed in WT.
However, CTT1 overexpression abolishes the premature
induction of apoptosis observed in sod1 strain, which
might explain the increased longevity of sod1 CTT1.
When sod1 cells were engineered to overexpress CTT1,
oxidative damage decreased, delaying the induction of
apoptosis and the onset of morbidity.
Conclusion
Our results confirmed that CR augments ROS defense
in S. cerevisiae, used as a model in this work, by a
Sod1p dependent mechanism. On the other hand, the
overexpression of CTT1 was able to overcome the
absence of Sod1p. In fact, the increase of life span
occurred because this overexpression might play an
important role in the cell membrane protection,
decreasing the oxidative damages and delaying the
induction of apoptotic process. On the order hand, it is
important to observe that the excess of catalase can be
harmful for the cell, getting the cell to an unbalanced
scenario between antioxidant defense system and
ROS, deregulating the physiological homeostasis.
Acknowledgments This work was supported by grants from
CAPES and CNPq.
References
Adamis PDB, Panek AD, Eleutherio E (2007) Vacuolar com-
partmentation of the cadmium-glutathione complex pro-
tects Saccharomyces cerevisiae from mutagenesis. Toxicol
Lett 173:1–7
Aebi H (1964) Catalase in vitro. Methods Enzymol 105:114–118
Agarwal S, Sharma S, Agrawal V, Roy N (2005) Caloric
restriction augments ROS defense in S. cerevisiae by a
Sir2p independent mechanism. Free Radic Res. 39:55–62
Bar G (2002) The quantitative measurement of H2O2 generation in
isolated mitochondria. J Bioenerg Biomemb. 34:227–233
Baur JA (2010) Resveratrol, sirtuins, and the promise of a DR
mimetic. Mech Aging Dev. 131:261–269
Botstein D, Fink GR (2011) Yeast: an experimental organism
for 21st Century biology. Genetics 189:695–704
Carmona-Gutierrez D, Buttner S (2014) The many ways to age
for a single yeast cell. Yeast 31(8):289–298
Carter WO, Narayanan PK, Robinson JP (1994) Intracellular
hydrogen peroxide and superoxide anion detection in
endothelial cells. J Leukoc Biol 55:253–258
Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kos-
matka KJ, Beasley TM, Allison DB, Cruzen C, Simmons
HA, Kemnitz JW, Weindruch R (2009) Caloric restriction
delays disease onset and mortality in rhesus monkeys.
Science 325:201–204
Dalle-Donnea I, Rossib R, Giustarinib D, Milzania A, Colombo
R (2003) Protein carbonyl groups as biomarkers of oxi-
dative stress. Clin Chim Acta 329:23–38
de Moraes LMP, Astolfi-filho S, Oliver SG (1995) Development
of yeast strains for the efficient utilisation of starch: eval-
uation of constructs that express—amylase and glucoam-
ylase separately or as bifunctional fusion proteins. Appl
Microbiol Biotechnol 43:1067–1076
del Valle LG (2011) Oxidative stress in aging: theoretical out-
comes and clinical evidences in humans. Biomed Aging
Pathol 1:1–7
Demir AB, Koc A (2010) Assessment of chronological lifespan
dependent molecular damages in yeast lacking mitochon-
drial antioxidant genes. Biochem Biophys Res Commun
400:106–110
Fabrizio P, Longo VD (2003) The chronological life span of
Saccharomyces cerevisiae. Aging Cell 2:73–81
Fabrizio P, Longo VD (2008) Chronological aging-induced
apoptosis in yeast. Biochim Biophys Acta 1783:1280–1285
Fernandes PN, Mannarino SC, Silva CG, Pereira MD, Panek AD,
Eleutherio ECA (2007) Oxidative stress response in
eukaryotes: effect of glutathione, superoxide dismutase and
catalase on adaptation to peroxide and menadione stresses in
Saccharomyces cerevisiae. Redox Rep 12:236–244
Fukai T, Ushio-Fukai M (2011) Superoxide dismutases: role in
redox signaling, vascular function, and diseases. Antioxid
Redox Signal 15:1583–1606
Gietz D, St Jean A, Woods RA, Schiestl RH (1992) Improved
method for high efficiency transformation of intact yeast
cells. Nucleic Acids Res 20:1425–1435
Gomes DS, Pereira MD, Panek AD, Andrade LR, Eleutherio E
(2008) Apoptosis as a mechanism for removal of mutated
cells of Saccharomyces cerevisiae: the role of Grx2 under
cadmium exposure. Biochim Biophys Acta 1780:160–166
Harman D (2006) Free radical theory of aging: an update
increasing the functional life span. Ann NY Acad Sci
1067:1–12
Harris N, Bachler M, Costa V, Mollapour M, Moradas-Ferreira
P, Piper P (2005) Overexpressed Sod1pacts either to reduce
or to increase the life spans and stress resistance of yeast,
depending on whether it is Cu(2?)-deficient or an active
Cu, Zn-superoxide dismutase. Aging Cell 4:41–52
Herker E, Jungwirth H, Lehmann KA, Maldener C, Frohlich
KU, Wissing S, Buttner S, Fehr M, Sigrist S, Madeo F
(2004) Chronological aging leads to apoptosis in yeast.
J Cell Biol 164:501–507
Herrero E, Ros J, Bellı G, Cabiscol E (2008) Redox control and
oxidative stress in yeast cells. Biochim Biophys Acta
1780:1217–1235
Huberts DEW, Gonzalez J, Lee SS, Litsios A, Hubmann G, Wit
EC, Heinemann EC (2014) Calorie restriction does not
Biogerontology
123

elicit a robust extension of replicative lifespan in Saccha-
romyces cerevisiae. PNAS 111:11727–11731
Kaeberlein M (2010) Lessons on longevity from budding yeast.
Nature 464:513–519
Khurana V, Lindquist S (2010) Modelling neurodegeneration in
Saccharomyces cerevisiae: why cook with baker’s yeast?
Nat Rev 11:436–449
Liu XD, Thiele DJ (1996) Oxidative stress induced heat shock
factor phosphorylation and HSF-dependent activation of
yeast metallothionein gene transcription. Genes Dev
10:592–603
Longo VD, Gralla EB, Valentine JS (1996) Superoxide dismutase
activity is essential for stationary phase survival in Sac-
charomyces cerevisiae. Mitochondrial production of toxic
oxygen species in vivo. J Biol Chem 271:12275–12280
Mannarino SC, Amorim MA, Pereira MD, Moradas-Ferreira P,
Panek AD, Costa V, Eleutherio EC (2008) Glutathione is
necessary to ensure benefits of calorie restriction during
aging in Saccharomyces cerevisiae. Mech Aging Dev
129:700–705
Mannarino SC, Vilela LF, Brasil AA, Aranha JN, Moradas-
Ferreira P, Pereira MD, Costa V, Eleutherio EC (2011)
Requirement of glutathione for Sod1 activation during
lifespan extension. Yeast 28:19–25
Mattison JA, Roth GS, Beasley TM, Tilmont EM, Handy AM,
Herbert RL, Longo DL, Allison DB, Young JE, Bryant M,
Barnard D, Ward WF, Qi W, Ingram DK, de Cabo R (2012)
Impact of caloric restriction on health and survival in
rhesus monkeys from the NIA study. Nature 489:318–321
Mecocci P, Polidori MC, Troiano L, Cherubini A, Cecchetti R,
Pini G, Straatman M, Monti D, Stahl W, Sies H, Franceschi
C, Senin U (2000) Plasma antioxidants and longevity: a
study on healthy centenarians. Free Radic Biol Med
28:1243–1248
Mesquita A, Weinberger M, Silva A, Sampaio-Marques B,
Almeida B, Leao C, Costa V, Rodrigues F, Burhans WC,
Ludovico P (2010) Caloric restriction or catalase inacti-
vation extends yeast chronological lifespan by inducing
H2O2 and superoxide dismutase activity. Proc Natl Acad
Sci USA 107:15123–15128
Miao L, St. Clair DK (2009) Regulation of superoxide dismu-
tase genes: implications in diseases. Free Radic Biol Med
47:344–356
Mirisola MG, Braun RJ, Petranovic D (2013) Approaches to
study yeast cell aging and death. FEMS Yeast Res
14:109–118
Ozbay B, Dulger H (2002) Lipid peroxidation and antioxidant
enzymes in Turkish population: relation to age, gender,
exercise, and smoking. Tohoku J Exp Med 197:119–124
Parrella E, Longo VD (2008) The chronological life span of
Saccharomyces cerevisiae to study mitochondrial dys-
function and disease. Methods 46(4):256–262
Pereira MD, Herdeiro RS, Fernandes PN, Eleutherio E, Panek
AD (2003) Targets of oxidative stress in yeast sod mutants.
Biochim Biophys Acta 1620:245–251
Piper P (2006) Long-lived yeast as a model for aging research.
Yeast 23:215–226
Poljsak B (2011) Strategies for reducing or preventing the
generation of oxidative stress. Oxid Med Cell Longev
2011:194586
Radak Z, Zhao Z, Goto S, Koltai E (2011) Age-associated
neurodegeneration and oxidative damage to lipids, proteins
and DNA. Mol Asp Med 32:305–315
Reverter-Branchat G, Cabiscol E, Tamarit J, Sorolla MA,
Angeles de la Torre M, Ros J (2007) Chronological and
replicative life-span extension in Saccharomyces cerevi-
siae by increased dosage of alcohol dehydrogenase 1.
Microbiology 153:3667–3676
Ribaric S (2012) Diet and aging. Oxid Med Cell Longev
2012:741468
Ristow M, Schmeisser S (2011) Extending life span by increasing
oxidative stress. Free Radic Biol Med 51:327–336
Rodrigues-Pousada C, Menezes RA, Pimentel C (2010) The Yap
family and its role in stress response. Yeast 27:245–258
Sambrook J, Maniatis T, Fritsch EF (1989) Molecular cloning: a
laboratory manual. Cold Spring Harbor Laboratory Press,
New York
Sheu SS, Nauduri D, Anders MW (2006) Targeting antioxidants
to mitochondria: a new therapeutic direction. Biochim
Biophys Acta 1762:256–265
Shimokawa I, Chiba T, Yamaza H, Komatsu T (2008) Lon-
gevity genes: insights from calorie restriction and genetic
longevity models. Mol Cells 30:427–435
Srinivasan C, Liba A, Imlay JA, Valentine JS, Gralla EB (2000)
Yeast lacking superoxide dismutase(s) show elevated lev-
els of ‘‘free iron’’ as measured by whole cell electron
paramagnetic resonance. J Biol Chem 275:29187–29192
Steels EL, Learmonth RP, Watson K (1994) Stress tolerance and
membrane lipid unsaturation in Saccharomyces cerevisiae
grown aerobically or anaerobically. Microbiology
140:569–576
Stickland LH (1951) The determination of small quantities of
bacteria by means of the Biuret reaction. J Gen Microbiol
5:698–703
Biogerontology
123




















