
Corticotropin-Releasing Factor (CRF) and the Urocortins Differentially Regulate Catecholamine...
16
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/6605544 Corticotropin-Releasing Factor (CRF) and the Urocortins Differentially Regulate Catecholamine Secretion in Human and... Article in Endocrinology · May 2007 DOI: 10.1210/en.2006-0967 · Source: PubMed CITATIONS 30 READS 31 12 authors, including: Some of the authors of this publication are also working on these related projects: Neurotrophin receptors role on adult neurogenesis View project Vasileios Minas Wirral University Teaching Hospital NHS Fo… 38 PUBLICATIONS 745 CITATIONS SEE PROFILE Ioannis Charalampopoulos Medical School, University of Crete 72 PUBLICATIONS 1,541 CITATIONS SEE PROFILE Maria Lambropoulou Democritus University of Thrace 101 PUBLICATIONS 657 CITATIONS SEE PROFILE Achilleas Gravanis University of Crete 179 PUBLICATIONS 4,318 CITATIONS SEE PROFILE All content following this page was uploaded by Maria Lambropoulou on 01 December 2016. The user has requested enhancement of the downloaded file. All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.
Transcript of Corticotropin-Releasing Factor (CRF) and the Urocortins Differentially Regulate Catecholamine...
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/6605544
Corticotropin-ReleasingFactor(CRF)andtheUrocortinsDifferentiallyRegulateCatecholamineSecretioninHumanand...
ArticleinEndocrinology·May2007
DOI:10.1210/en.2006-0967·Source:PubMed
CITATIONS
30
READS
31
12authors,including:
Someoftheauthorsofthispublicationarealsoworkingontheserelatedprojects:
NeurotrophinreceptorsroleonadultneurogenesisViewproject
VasileiosMinas
WirralUniversityTeachingHospitalNHSFo…
38PUBLICATIONS745CITATIONS
SEEPROFILE
IoannisCharalampopoulos
MedicalSchool,UniversityofCrete
72PUBLICATIONS1,541CITATIONS
SEEPROFILE
MariaLambropoulou
DemocritusUniversityofThrace
101PUBLICATIONS657CITATIONS
SEEPROFILE
AchilleasGravanis
UniversityofCrete
179PUBLICATIONS4,318CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyMariaLambropoulouon01December2016.
Theuserhasrequestedenhancementofthedownloadedfile.Allin-textreferencesunderlinedinblue
arelinkedtopublicationsonResearchGate,lettingyouaccessandreadthemimmediately.

Corticotropin-Releasing Factor (CRF) and the UrocortinsDifferentially Regulate Catecholamine Secretion inHuman and Rat Adrenals, in a CRF ReceptorType-Specific Manner
E. Dermitzaki, C. Tsatsanis, V. Minas, E. Chatzaki, I. Charalampopoulos, M. Venihaki, A. Androulidaki,M. Lambropoulou, J. Spiess, E. Michalodimitrakis, A. Gravanis, and A. N. Margioris
Departments of Clinical Chemistry-Biochemistry (E.D., C.T., M.V., A.A., A.N.M.), Pharmacology (V.M., I.C., A.G.), andForensic Sciences (E.M.), School of Medicine, University of Crete, Heraklion GR-710 03, Crete, Greece; Departments ofPharmacology (E.C.) and Histology-Embryology (M.L.), School of Medicine, Democritus University of Thrace,Alexandroupolis, GR-68 100, Greece; Department of Neuroscience (J.S.), J. Burns School of Medicine, University of Hawaii,Honolulu, Hawaii 96822
Corticotropin-releasing factor (CRF) affects catecholamineproduction both centrally and peripherally. The aim of thepresent work was to examine the presence of CRF, its relatedpeptides, and their receptors in the medulla of human and ratadrenals and their direct effect on catecholamine synthesisand secretion. CRF, urocortin I (UCN1), urocortin II (UCN2),and CRF receptor type 1 (CRF1) and 2 (CRF2) were present inhuman and rat adrenal medulla as well as the PC12 pheo-chromocytoma cells by immunocytochemistry, immunofluo-rescence, and RT-PCR. Exposure of dispersed human and ratadrenal chromaffin cells to CRF1 receptor agonists inducedcatecholamine secretion in a dose-dependent manner, an ef-fect peaking at 30 min, whereas CRF2 receptor agonists sup-
pressed catecholamine secretion. The respective effects wereblocked by CRF1 and CRF2 antagonists. CRF peptides affectedcatecholamine secretion via changes of subplasmaliminal ac-tin filament polymerization. CRF peptides also affected cat-echolamine synthesis. In rat chromaffin and PC12 cells, CRF1and CRF2 agonists induced catecholamine synthesis via ty-rosine hydroxylase. However, in human chromaffin cells, ac-tivation of CRF1 receptors induced tyrosine hydroxylase,whereas activation of CRF2 suppressed it. In conclusion, itappears that a complex intraadrenal CRF-UCN/CRF-receptorsystem exists in both human and rat adrenals controlling cat-echolamine secretion and synthesis. (Endocrinology 148:1524–1538, 2007)
THE CATECHOLAMINERGIC SYSTEM and the hypo-thalamus-pituitary-adrenal axis comprise the two ma-
jor adaptation mechanisms to stress. The two axes interact atseveral levels to coordinate their response to stressful stimuli.Over the years, multiple reports have provided evidence thata complex corticotropin-releasing factor (CRF)-based systemexists within the adrenal glands coordinating the two stressaxes at a peripheral, intraadrenal level. Indeed, CRF, uro-cortin I (UCN1), urocortin II (UCN2) (in humans stresscopin-related peptide), urocortin III (in humans stresscopin), theirreceptors CRF receptor type 1 (CRF1), CRF receptor type 2(CRF2), and the decoy receptor CRF-binding protein havebeen reported to be present in whole human, rodent, bovine,and canine adrenals (1–11). However, there is no work study-ing the CRF system in the adrenal medulla, as a whole,vis-a-vis the distribution of each particular CRF peptide and
their receptors as well as their effects on catecholamine se-cretion and synthesis. The information available so far isfragmented and involves either rodent or human adrenals(12, 13).
We have previously shown that CRF stimulates catechol-amine production in adrenal chromaffin cells (14–16). CRFelevates adenylate cyclase activity in rat adrenal membranesand cAMP in bovine chromaffin cells (3), whereas UCN1elevates cAMP in PC12 cells (17). The significance of CRFpeptides on adrenal medulla has been demonstrated in glu-cocorticoid-supplemented CRF null mice in which tyrosinehydroxylase (TH), the rate-limiting enzyme in catecholaminesynthesis, and phenylethanolamine N-methyltransferase(PNMT), the enzyme catalyzing the conversion of norepi-nephrine (NE) to epinephrine (E), are lower, compared withCRF (�/�) mice (18). Furthermore, after immobilizationstress, the expression of PNMT does not appear to increasein CRF null mice to the same extent as in CRF(�/�) mice(19). Similar data have been reported in CRF1 null mice inwhich the levels of E are almost half that of wild-type miceof the same strain, whereas PNMT mRNA are even lower atalmost 25% (20). ACTH treatment of CRF1 null mice im-proves but does not completely restore E and PNMT mRNAlevels. Interestingly, the morphology of chromaffin cells inCRF1 null mice appears to be dramatically different, com-pared with that of the wild-type, exhibiting depletion of
First Published Online December 28, 2006Abbreviations: AMPT, l-2-Methyl-3-(-4hydroxyphenyl)-alanine;
CRF, corticotropin-releasing factor; CRF1, CRF receptor type 1; CRF2,CRF receptor type 2; DA, dopamine; DAPI, 4�,6-diamidino-2-phenylin-dole; E, epinephrine; FCS, fetal calf serum; HS, horse serum; NE, nor-epinephrine; NSD-1015, 3-(hydrazinomethyl)-phenol hydrochloride;PNMT, phenylethanolamine N-methyltransferase; TBS, Tris and NaCl;TH, tyrosine hydroxylase; UCN1, urocortin I; UCN2, urocortin II.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/07/$15.00/0 Endocrinology 148(4):1524–1538Printed in U.S.A. Copyright © 2007 by The Endocrine Society
doi: 10.1210/en.2006-0967
1524

epinephrine-storing secretory granules, a finding not com-pletely restored by ACTH (20). Finally, UCN1 has also beenshown to induce TH expression in PC12 cells via the cAMP/protein kinase A pathway (17).
The aim of the present work was first to map the distri-bution of CRF peptides and their respective receptors insections of human and rat adrenals and subsequently exam-ine the role of each component of the system in catechol-amine synthesis and secretion, in vitro. For this purpose weused human cadaveric adrenals from traffic accident victims,fresh rat adrenals, and the PC12 cell line. Selective and non-selective CRF1 and CRF2 receptor agonists and antagonistswere used to examine their effects on catecholamine secre-tion, actin filament dynamics, and TH expression (21).
Materials and MethodsReagents and antibodies
Rat/human recombinant CRF was purchased from Tocris (Ellisville,MO). Cortagine (nonpeptide CRF1 antagonist), antisauvagine-30 (syn-thetic CRF2 antagonist), and mouse UCN2 (CRF2 agonist) were providedby Dr. J. Spiess (J. Burns School of Medicine, University of Hawaii,Honolulu, HI), whereas antalarmin (nonpeptide CRF1 antagonist) wasprovided by Dr. G. P. Chrousos (National Institute of Child Health andHuman Development, National Institutes of Health). The pharmaco-logical tyrosine hydroxylase inhibitor, l-2-methyl-3-(-4hydroxyphenyl)-alanine (AMPT) and L-aromatic amino acid decarboxylase inhibitor,3-(hydrazinomethyl)-phenol hydrochloride (NSD-1015) were pur-chased from Sigma-Aldrich Corp. (St. Louis, MO). Mouse monoclonalantibodies against TH and actin were obtained from Chemicon (Te-mecula, CA). The antisera used for CRF, UCN1, and UCN2 proteindetection were purchased from Phoenix Pharmaceuticals Inc. (Belmont,CA). The antisera used for CRF receptor protein detection in humansections, the anti-CRF1 (4467a-CRF1) and anti-CRF2 (2064a-CRF2), werekindly provided by Dr. D. Grigoriadis (Neurocrine Bioscience Inc. SanDiego, CA) (22, 23), whereas the antisera used for CRF receptors proteindetection (sc-12381 and sc-20550) in both human and rat sections werepurchased from Santa Cruz Biotechnology (Santa Cruz, CA). Secondaryantibodies used were antimouse fluorescein isothiocyanate-labeled con-jugated, antimouse tetramethylrhodamine isothiocyanate conjugated,and antimouse horseradish peroxidase conjugated and were all pur-chased from Chemicon. Bradford Coomassie Brilliant Blue G-250 wasobtained from Bio-Rad Laboratories, Inc. (Hercules, CA), and nitrocel-lulose membranes for Western blotting were purchased from Millipore(Bedford, MA). Immunoreactive bands were visualized with an en-hanced chemiluminescence kit from PerkinElmer Life Sciences (Nor-walk, CT). Collagenase A, DNase I, collagen, poly-l-lysine, phallacidin,and trypsin were all obtained from Sigma. All sterile tissue apparatuswere obtained from Corning (Corning, NY). RPMI 1640, DMEM-F12,l-glutamine, fetal calf serum (FCS), penicillin/streptomycin, and horseserum (HS) were purchased from Invitrogen (Carlsbad, CA). All otherchemicals and reagents were obtained from Sigma, if not statedotherwise.
Human and rat primary adrenal chromaffin cell cultures
Cadaver human adrenals were obtained from traffic accident victimsfrom our Forensic Medicine Department. The glands were taken ac-cording to the standard protocols and processed within 12 h of death.The University of Crete Ethics Committee for human studies approvedthe procedure and use of specimen. Rat adrenals were obtained fromadult female Sprague-Dawley rats. The experimental protocols wereapproved by the Laboratory Animal Care Committee and the PublicVeterinary Department of the Heraklion prefecture, Crete, Greece. Inbrief, human or rat adrenal medulla was cut in small pieces and incu-bated in freshly prepared dispersion medium containing 1 mg/ml col-lagenase A and 0.1 mg/ml DNase I (human adrenals) or 2 mg/mlcollagenase A, 0.1 mg/ml DNase I, and 0.125% trypsin (rat adrenals) fora maximum of 1 h in water bath. Then human or rat cell suspensionswere incubated in 0.146 m NH4Cl for 1–2 min on ice for the eliminationof erythrocytes. Human chromaffin cells were cultured in DMEM-F12
supplemented with 10 mm l-glutamine, 4.5 mm glucose, 15 mm HEPES,100 U/ml penicillin, 0.1 mg/ml streptomycin, 10% HS, and 2.5% FCS,whereas rat chromaffin cells were cultured in DMEM-F12 supplementedwith 10 mm l-glutamine, 3.7 g/liter sodium bicarbonate, 4.5 mm glucose,15 mm HEPES, 100 U/ml penicillin, 0.1 mg/ml streptomycin, and 20%FCS in collagen-coated plates for 2 d. One day before each experiment,the initial culture media were changed with serum-free media supple-mented with 0.1% BSA. Cells were then stimulated with CRF, UCN1,UCN2, or cortagine at various time intervals.
PC12 rat pheochromocytoma cell culture
PC12 cells were obtained from two sources: Dr. M. Greenberg (Chil-dren’s Hospital, Boston, MA) and the American Type Culture Collection(Manassas, VA). Cells were grown in RPMI 1640 containing 10 mml-glutamine, 15 mm HEPES, 100 U/ml penicillin, 0.1 mg/ml strepto-mycin, 10% HS, and 5% FCS at 5% CO2 and 37 C. One day before eachexperiment, the initial culture media were changed with serum-freemedia supplemented with 0.1% BSA.
Immunocytoshemistry
Immunohistochemistry was performed on paraffin-embedded tissuesections (4–6 �m), which had been routinely fixed in 4% bufferedformalin. Paraffin sections were deparaffinized in xylene and rehy-drated in graded alcohol to TBS [50 mm Tris, 150 mm NaCl (pH 7.4)]. Theslides were micro waved for 3 � 4 min in 10 mm citrate, washed in TBS,incubated for 10 min with 3% H2O2, and blocked for 10 min with PowerBlock (BioGenex, San Ramon, CA). Slides were then incubated for 75 minwith anti-CRF1 (4467a; 1:1500) or anti-CRF2 (2064a; 1:1500) diluted in10% normal rabbit serum in PBS. Alternatively, slides were incubatedwith anti-CRF1 (sc-12381; 1:50) or anti-CRF2 (sc-20550; 1:50) diluted in 2%BSA in TBS overnight at 4 C. For the detection of CRF peptides or TH,slides were incubated for 60 min with anti-CRF (H-019–06; 1:100), anti-UCN1 (H-019–14; 1:100), anti-UCN2 (H-019–24; 1:50), or anti-TH(MAB318; 1:100), all diluted in TBS containing 2% BSA. For preabsorp-tion studies, primary antibodies were incubated with or without therespective blocking peptides overnight at 4 C. Subsequently all theantibodies used were then washed with TBS, incubated with biotinyl-ated secondary antigoat antibody (BioGenex) for 20 min at room tem-perature, and finally streptavidin-conjugated horseradish peroxidasefor 20 min at room temperature. Peroxidase-staining reaction was per-formed with 3,3�-diaminobenzidine tetrahydrochloride (3 min) andstopped in tap water. Sections from all tissues were counterstained inMayer’s hematoxylin (1 min) and covered with coverslips. In negativecontrols, primary antibodies were omitted. Tissues were photographedwith a digital camera (Nikon, Tokyo, Japan) under a microscope (Olym-pus Tokyo, Japan).
Confocal laser-scanning microscopy
Cells were seeded into 8-well chamber slides coated with poly-l-lysine. At the end of each treatment period, cells were fixed by incu-bation in 3.7% formaldehyde and permeabilized by 0.2% Triton X-100before blocking. Subsequently cells were incubated with rhodamineconjugated to phalloidin (Molecular Probe, Inc., Eugene, OR). The cov-erslips were analyzed using a confocal laser-scanning module (LeicaCorp., Lasertechnik, Heidelberg, Germany) attached to an inverted mi-croscope (IM35; Carl Zeiss, Oberkochen, Germany) equipped with anargon-krypton ion laser. Confocal images were acquired using a 63/1.25oil immersion objective and CLSM software (Leica).
Immunofluorescence
PC12 cells were grown in chamber slides. The cells were fixed andpermeabilized by acetone followed by 0.01% Triton X-100 before block-ing. For the detection of CRF receptors in the membrane section, cellswere not permeabilized before blocking. Subsequently cells were incu-bated with primary antibodies for 1 h. For preabsorption studies, pri-mary antibodies were incubated with or without the respective blockingpeptides overnight at 4 C. Finally, cells were incubated with secondaryantibodies and mounted with antifade reagent containing 4�,6-dia-midino-2-phenylindole (DAPI) for nuclei visualization and covered withcoverslips. The slides were photographed under an Olympus micro-scope with a Leica camera.
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1525

Measurement of catecholamines
Cells were grown in six-well plates, coated with poly-l-lysine (PC12cells) or collagen (human and rat chromaffin cells). After stimulation, theconcentration of endogenous catecholamines in the culture media wasmeasured by HPLC; catecholamines were extracted by alumina columnsas per the manufacturer’s protocol (Chromsystems, Munich, Germany).Subsequently the extracts were injected into HPLC (Agilent, Palo Alto,CA) equipped with an electrochemical CLC 100 detector (Chromsys-tems). The signals were recorded in an analog digital converter (HP-35900 C; Hewlett-Packard, Boblingen, Germany) connected to a com-puter. The concentration of endogenous catecholamines in the culturemedia was also measured by a highly sensitive RIA (TriCat RIA,RE29395; IBL Immuno Biological Laboratories, Hamburg, Germany)used as previously described (24). Cells were harvested and sonicatedfor quantification of total cellular proteins as previously described (21).Data are presented as nanograms catecholamines per milligram of pro-tein. No significant difference was found between the mean concentra-tions of catecholamines measured by the HPLC method and RIA.
RT-PCR analysis for CRF receptors
Total RNA from cells was extracted and processed for the PCR asdescribed previously (24). Each cycle consisted of 30 sec at 94 C, 60 secat 55 C, and 60 sec at 72 C for 35 cycles to detect the product at theexponential phase of the amplification. The primers for rat CRF1 were5�-AGGCGGGATCCAGGCAGTAGAGA-3� (sense) and 5�-TCCCGG-TAGCCATTGTTTGTCGTG-3� (antisense), whereas the primers for ratCRF2 were 5�-CTGGTGGCTGCTTTCCTGGTTTTC-3� (sense) and 5�-ATGGGGCCCTGGTAGATGTAGTCC-3� (antisense). Oligonucleotideswere synthesized by MWG-Biotech (Ebersberg, Germany). Ten micro-liters of the amplified product were separated on a 2% agarose gel andvisualized by ethidium bromide staining.
Real-time PCR for TH
For quantitation of rat TH mRNA, cDNA was synthesized and pro-cessed as described previously (24). For quantitation of human THmRNA, cDNA was synthesized and processed as described for rat THmRNA with some modifications: each cycle consisted of 40 sec at 94 C,40 sec at 60 C, and 40 sec at 72 C for a maximum of 40 cycles. Noby-products were present in the reaction as indicated by the dissociationpattern provided at the end of the reaction and agarose gel electro-phoresis (data not shown). The primers for human TH were 5�-CCCT-GGTTCCCAAGAAAAGT-3� (sense) and 5�-GCGTGGTGTAGACCTC-CTTC-3� (antisense).
Western blot analysis
PC12 cell lysates were electrophoresed through a 12% sodium do-decyl sulfate-polyacrylamide gel and transferred to nitrocellulose mem-branes as described previously (25). Membranes were processed ac-cording to standard Western blotting procedures. To detect proteinlevels, membranes were incubated with anti-TH antibody and thenexposed to X-Omat AR films (Kodak, Rochester, NY). A PC-based ImageAnalysis program was used to quantify the intensity of each band(Image Analysis, Inc., Ontario, Canada). To normalize for protein con-tent, the blots were stripped and stained with antiactin antibody; theconcentration of TH was normalized vs. actin.
Statistical analysis
For the statistical evaluation of our data, we used ANOVA, post hoccomparison of means followed by two multiple comparison tests: Fish-er’s least significance difference and the Newman-Keuls test. For dataexpressed as percent changes, compared with control values, we usedthe nonparametric Kruskal-Wallis test for several independent samples.
ResultsMapping of CRF peptides and their receptors in human andrat adrenal medulla.
Immunohistochemical mapping was performed in sec-tions of six nonpathological human and eight nonpathologi-
cal rat adrenals using antisera against TH, CRF, UCN1,UCN2, CRF1, and CRF2 as described in Materials and Methods.Figure 1A (TH panel) depicts chromaffin cells in humansections stained with TH antibody. The adrenal cortex servedas control and did not stain with the TH antibody as ex-pected. UCN2 and the CRF2 receptor were mainly expressedin the adrenal cortex (Fig. 1A, UCN2 and CRF2 panels), CRFwas abundantly expressed in both the adrenal medulla andcortex (Fig. 1A, CRF panel), whereas UCN1 and the CRF1
receptor were mainly expressed in the adrenal medulla (Fig.1A, UCN1 and CRF1 panels). It should be noted that thespecificity of the antibodies used was tested by preincubationwith the respective peptides as described in Materials andMethods. Preabsorbed antibodies did not stain (Fig. 1A, lastrow).
In the rat, the chromaffin cells positive for TH (Fig. 1B, THpanel) were also positive for UCN2, CRF, CRF1, and CRF2
(Fig. 1B, UCN2, CRF, CRF1, and CRF2 panels). UCN1 wasexpressed throughout the adrenal medulla, although the in-tensity of staining fluctuated between individual chromaffincells (Fig. 1B, UCN1 panel). Indeed, several clusters of chro-maffin cells exhibited a dense brown color, whereas othersshowed a lesser density of staining but nevertheless positive,compared with the control sections exposed to either noantiserum (Fig. 1B, NEG panel) or to preabsorbed antibodies(Fig. 1B, last row). As a result, all cells in the adrenal medullaappeared to be positive for UCN1, although with some fluc-tuation in staining, most probably due to the blocking effectof locally produced UCN1. The immunohistochemical datafrom human and rat adrenals are summarized in Table 1.
Immunofluorescence analysis confirmed the presence ofUCN2, CRF, UCN1 (Fig. 2A), and their receptors CRF1 andCRF2 in PC12 cells (Fig. 2, B and C). Again, PC12 cells ex-posed to preabsorbed antibodies showed no staining for eachrespective peptide (data not shown). Finally, the CRF1 (26)and CRF2 receptor transcripts were also detected by RT-PCRanalysis of PC12 cells (Fig. 2D).
Effects of CRF1 and CRF2 agonists and antagonists oncatecholamine secretion from dispersed human and ratadrenal chromaffin and PC12 cells
Selective activation of CRF1 receptors resulted in inductionof catecholamine secretion from dispersed human and ratadrenal chromaffin and PC12 cells, an effect peaking at 30min and lasting for around 2 h. Conversely, selective acti-vation of CRF2 receptors resulted in suppression of catechol-amine secretion.
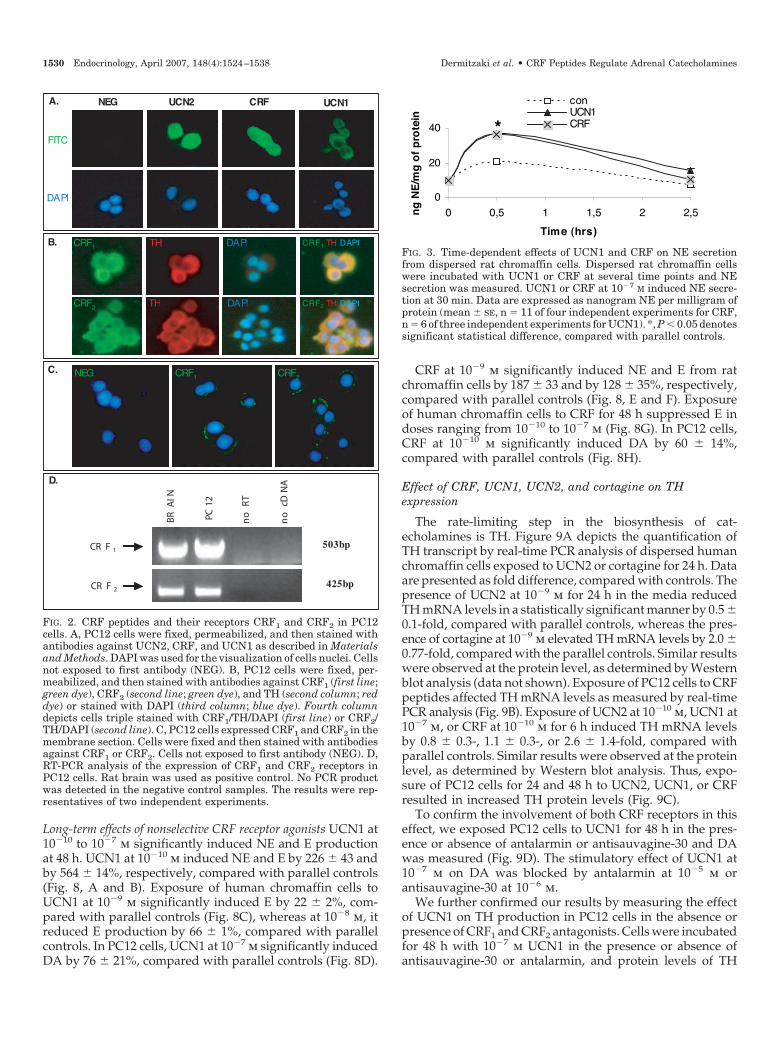
To determine the optimal time point for the acute effect ofCRF peptides, rat chromaffin cells were stimulated with theCRF1 and CRF2 receptor agonists CRF or UCN1 at 10�7 m andNE release was measured (Fig. 3). More specifically, expo-sure of rat chromaffin cells to CRF or UCN1 at 10�7 m for 30min induced NE secretion to 36.8 � 3.1 ng NE per milligramof protein or 37.3 � 4.4 ng NE per milligram of protein,compared with 20.9 � 0.5 ng NE per milligram of protein ofcontrol cells, respectively. Time-response experiments werealso performed for CRF and UCN1 on E (human and ratchromaffin cells) or dopamine (DA) (PC12 cells) secretion.
1526 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines

The resulting curves were similar to those observed for NE(rat chromaffin cells), and they are not shown here.
Effect of selective CRF receptor ligands on catecholaminesecretion
Dispersed rat and human chromaffin and PC12 cells wereexposed to UCN2, a specific endogenous ligand for the CRF2receptor or cortagine, a specific synthetic CRF1 agonist, atconcentrations ranging from 10�10 to 10�7 m. Catecholamines
were measured at 30 min, the time point found earlier torepresent their peak effect on time-response curves.
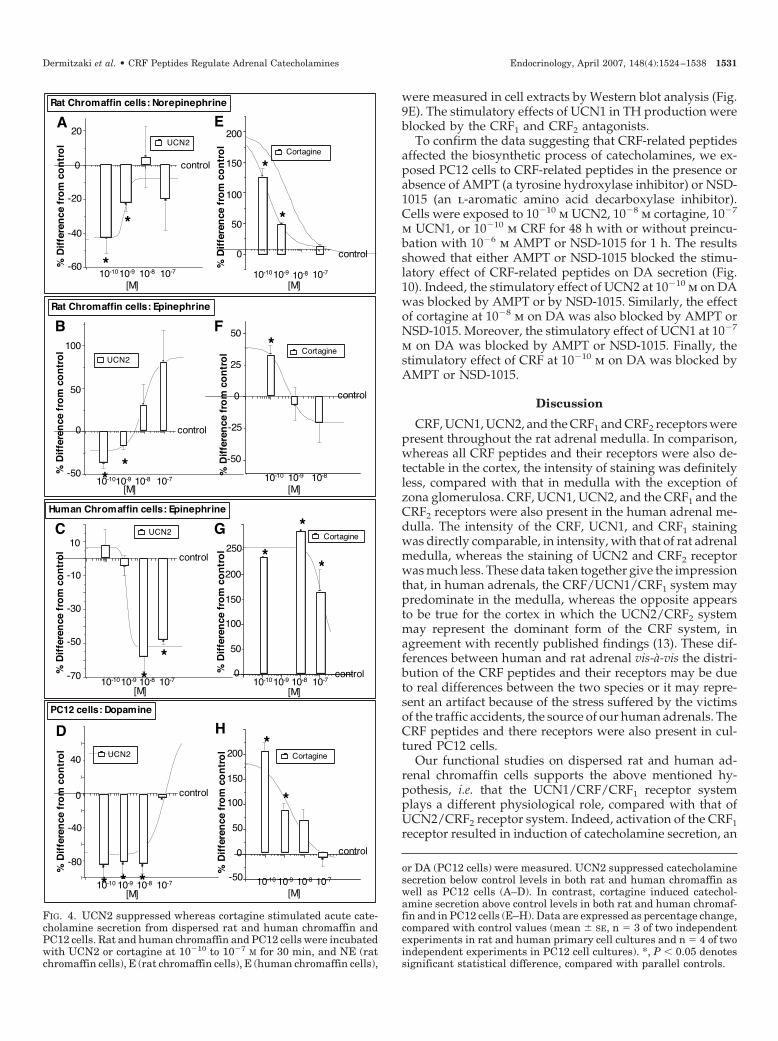
CRF2 agonist. Figure 4, A and B, depict the inhibitory effectof UCN2 on NE and E secretion in rat chromaffin cells.Exposure of rat chromaffin cells to UCN2 at 10�10 m for 30min suppressed NE secretion by 44 � 9% and E secretion by38 � 7%, compared with parallel controls. Moreover, expo-sure of human chromaffin cells to UCN2 in doses ranging
FIG. 1. Distribution of CRF-related pep-tides and their receptors CRF1 and CRF2 inhuman and rat adrenal medulla. Chromaf-fin cells were identified by anti-TH stain-ing. A, Human adrenal sections were pre-pared and then stained with anti-TH,UCN2, CRF, UCN1, CRF1, or CRF2 as de-scribed in Materials and Methods; TH-pos-itive cells were exclusively localized in themedulla (first line; TH). CRF2 and its en-dogenous agonist UCN2 were mainly ex-pressed in the cortex (second and sixthlines; UCN2 and CRF2), whereas CRF wasabundantly expressed throughout theadrenals (third line; CRF). CRF1 and itsendogenous agonist UCN1 were mainly ex-pressed in the medulla (fourth and fifthlines; UCN1 and CRF1). Sections exposedto antibodies preabsorbed with their cor-responding peptides revealed no specificstaining (seventh line). B, Rat adrenal sec-tions. TH-positive cells were exclusively lo-calized in the medulla (first line; TH). Sec-tions not exposed to primary antibodies(first line; NEG). UCN2, CRF, UCN1,CRF1, and CRF2 were strongly expressedin the medulla (second to sixth lines; UCN2,CRF, UCN1, CRF1, and CRF2). Sectionsexposed to antibodies preabsorbed withtheir corresponding peptides revealed nospecific staining (seventh line). Photos in A(column a), A (seventh line), and B (columna) were magnified �40, whereas photos inA (columns b–d), B (columns b–d), and B(seventh line) were magnified �250. Med,Medulla; retic, reticularis; fasc, fasciculate;glom, glomerulosa; caps, capsule.
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1527

from 10�8 to 10�7 m for 30 min significantly suppressed Esecretion below control levels (Fig. 4C). Furthermore, expo-sure of PC12 cells to UCN2 in doses ranging from 10�10 to10�8 m for 30 min significantly suppressed DA secretionbelow control levels (Fig. 4D).
CRF1 agonist. Figure 4, E and F, depict the effect of cortagineon NE and E secretion in rat chromaffin cells. Cortagine at10�10 m for 30 min induced NE secretion by 121 � 14% andE secretion by 32 � 8%, compared with parallel controls.Human chromaffin cells had the same response to cortagine
as rat in all doses tested, i.e. it induced E secretion in astatistically significant manner (Fig. 4G). Furthermore, ex-posure of PC12 cells to cortagine in doses ranging from 10�10
to 10�8 m for 30 min significantly induced DA secretionabove control levels (Fig. 4H).
Effects of nonselective CRF receptor ligands oncatecholamine secretion
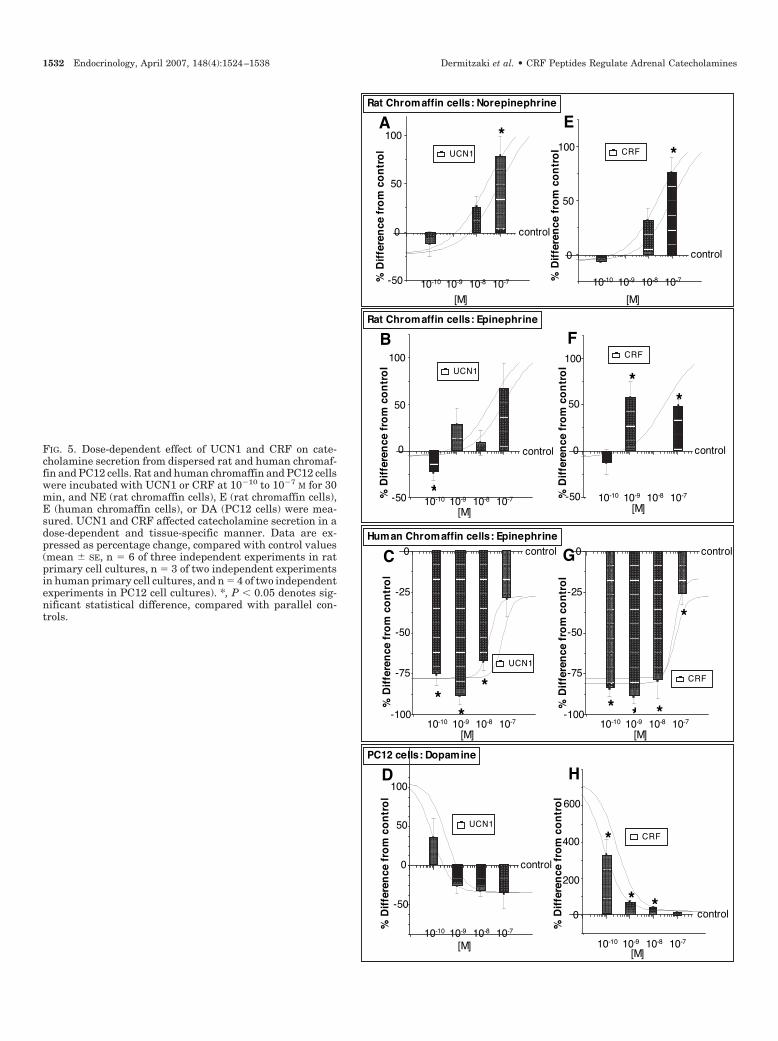
Exposure of rat chromaffin cells to UCN1 at 10�7 m sig-nificantly induced NE secretion at 30 min by 78 � 3%, com-
FIG, 1. Continued
1528 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines

pared with parallel controls (Fig. 5A). Furthermore, exposureof rat chromaffin cells to UCN1 at 10�10 m significantly sup-pressed E secretion by 24 � 8%, compared with parallelcontrols, whereas at 10�7 m, it induced E secretion by 78 �4%, compared with parallel controls (Fig. 5B). In humanchromaffin cells, UCN1 at all doses tested significantly re-duced E secretion (Fig. 5C). In PC12 cells, UCN1 at 10�8 msignificantly reduced DA secretion by 32 � 8%, comparedwith parallel control cells (Fig. 5D).
CRF at a concentration of 10�9 to 10�7 m had a stimulatoryeffect on catecholamine secretion from rat chromaffin cells(Fig. 5, E and F). Human chromaffin cells responded slightlydifferently, compared with rat chromaffin cells, to CRF.Hence, CRF at all doses tested significantly reduced E se-cretion below control levels from human chromaffin cells(Fig. 5G). In PC12 cells, CRF at 10�9 m induced significantlyDA secretion by 68 � 11%, compared with parallel controlcells (Fig. 5H).
Effect of CRF receptor antagonists
To confirm the specificity of each receptor signal, we ex-posed PC12 cells to CRF receptor agonists in the absence orpresence of antalarmin (a CRF1 antagonist) or antisauvag-ine-30 (a CRF2 antagonist) for 30 min, and DA secretion wasmeasured (Fig. 6A). Indeed, the inhibitory effect of UCN2 at10�10 m on DA secretion was blocked by antisauvagine-30 at10�8 m and not antalarmin at 10�8 m. The stimulatory effectof cortagine at 10�8 m on DA secretion was blocked byantalarmin at 10�6 m and not antisauvagine-30 at 10�6 m. Theeffect of UCN1 at 10�8 m on DA secretion was blocked byantisauvagine-30 at 10�6 m and not antalarmin at 10�6 m. Thestimulatory effect of CRF at 10�8 m on DA secretion wasblocked by both antalarmin at 10�6 m and antisauvagine-30at 10�6 m. Similar results were obtained in primary rat andhuman chromaffin cells (data not shown).
CRF1 ligands suppressed actin filament polymerization,whereas CRF2 ligands induced it
Exocytosis of catecholamines stored in secretory granulesdepends on the depolymerization and reorganization of sub-membrane actin filaments (21). Treatment of dispersed ratadrenal chromaffin cells with phallacidin, an actin filamentstabilizer, inhibited the acute stimulatory effect of CRF on E
secretion (Fig. 6B). Immunofluorescence analysis was per-formed to visualize actin filaments and determine their po-lymerization status. PC12 cells were treated with the agonistsCRF, cortagine, UCN1, and UCN2 at 10�8 m for 30 min, fixed,permeabilized, and then stained with rhodamine-phalloidinfor the visualization of filamentous actin as described inMaterials and Methods (Fig. 6C). Additional cells exposed tovehicles were also prepared and stained with rhodamine-phalloidin (Fig. 6C, con). Cells exposed to vehicles showeda continuously dense staining restricted to the subplasmale-mmal area beneath the cell membrane. In contrast, cells ex-posed to CRF or cortagine showed a broken subplasmale-mmal ring of actin filaments with gap areas of filamentousactin disassembly (Fig, 6C, CRF, cortagine panels). More-over, cells exposed to UCN1 or UCN2 showed an enhancedfilamentous actin assembly either in subplasmalemmal areabeneath the cell membrane or to the cytoplasmic section (Fig,6C, UCN1, UCN2 panels). These results suggest that expo-sure of PC12 cells to CRF or cortagine at 10�8 m for 30 mincaused actin filament depolymerization, whereas exposureof PC12 cells to UCN1 or UCN2 at 10�8 m caused inductionof filamentous actin formation.
CRF receptor ligands affect catecholamine synthesis and theTH transcript and protein levels
Long-term effects of selective CRF receptor agonists
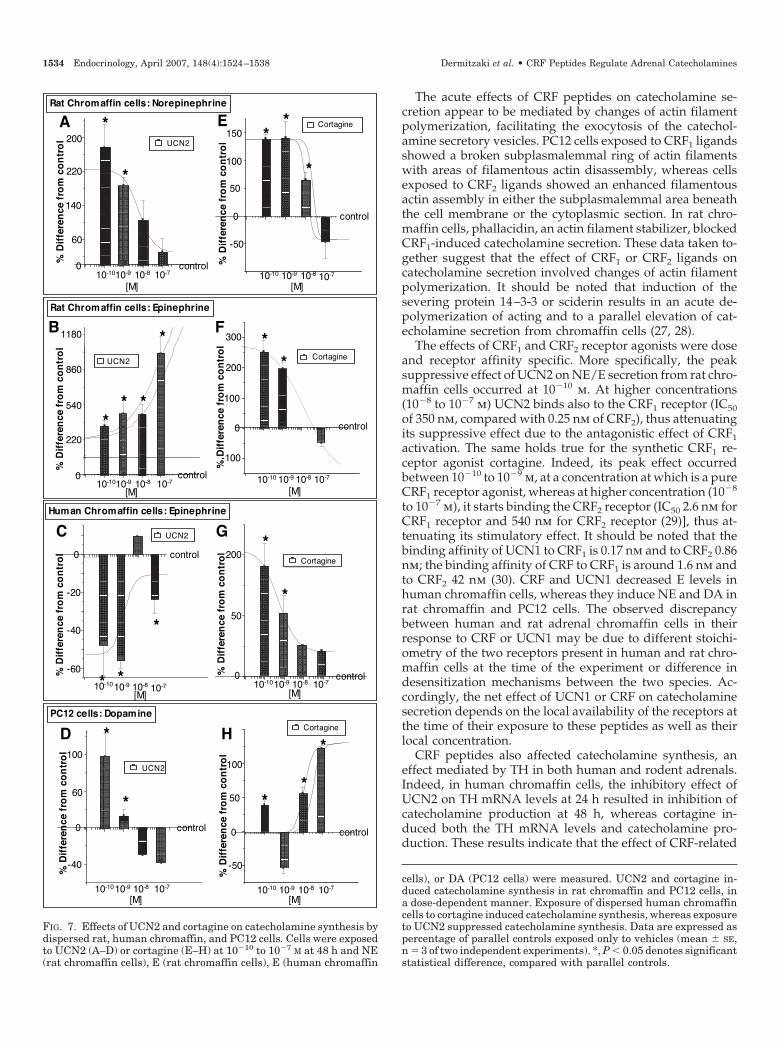
CRF2 agonist. Exposure of rat chromaffin cells to the pre-dominantly CRF2 agonist UCN2 at 10�9 m for 48 h stimulatedNE by 187 � 19% and E by 461 � 120%, compared withparallel controls (Fig. 7, A and B). Exposure of human chro-maffin cells to UCN2 at 10�9 m reduced E by 55 � 4%,compared with controls (Fig. 7C). Exposure of PC12 cells toUCN2 for 48 h induced DA in doses ranging from 10�10 to10�9 m (Fig. 7D).
CRF1 agonist. Cortagine at 10�9 m induced NE and E by139 � 30, and 188 � 21%, respectively, compared with par-allel controls (Fig. 7, E and F). Exposure of human chromaffincells to cortagine at 10�9 m induced E by 50 � 15%, comparedwith parallel controls (Fig. 7G). Exposure of PC12 cells tocortagine at 10�10 m induced DA by 40 � 16%, comparedwith parallel controls (Fig. 7H).
TABLE 1. Distribution of CRF-related peptides and their receptors in the medulla and cortex of human and rat adrenals
MedullaCortex
Reticularis Fasciculata Glomerulosa
HumanUCN2 � ��� � ��CRF �� �� �� ��UCN1 ��� � � �CRF1 ��� � � �CRF2 � �� �� �
RatUCN2 ��� �� �� �CRF ��� � �� ���UCN1 ��� � �� ���CRF1 ��� � �� ��CRF2 ��� � �� ��
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1529
Long-term effects of nonselective CRF receptor agonists UCN1 at10�10 to 10�7 m significantly induced NE and E productionat 48 h. UCN1 at 10�10 m induced NE and E by 226 � 43 andby 564 � 14%, respectively, compared with parallel controls(Fig. 8, A and B). Exposure of human chromaffin cells toUCN1 at 10�9 m significantly induced E by 22 � 2%, com-pared with parallel controls (Fig. 8C), whereas at 10�8 m, itreduced E production by 66 � 1%, compared with parallelcontrols. In PC12 cells, UCN1 at 10�7 m significantly inducedDA by 76 � 21%, compared with parallel controls (Fig. 8D).
CRF at 10�9 m significantly induced NE and E from ratchromaffin cells by 187 � 33 and by 128 � 35%, respectively,compared with parallel controls (Fig. 8, E and F). Exposureof human chromaffin cells to CRF for 48 h suppressed E indoses ranging from 10�10 to 10�7 m (Fig. 8G). In PC12 cells,CRF at 10�10 m significantly induced DA by 60 � 14%,compared with parallel controls (Fig. 8H).
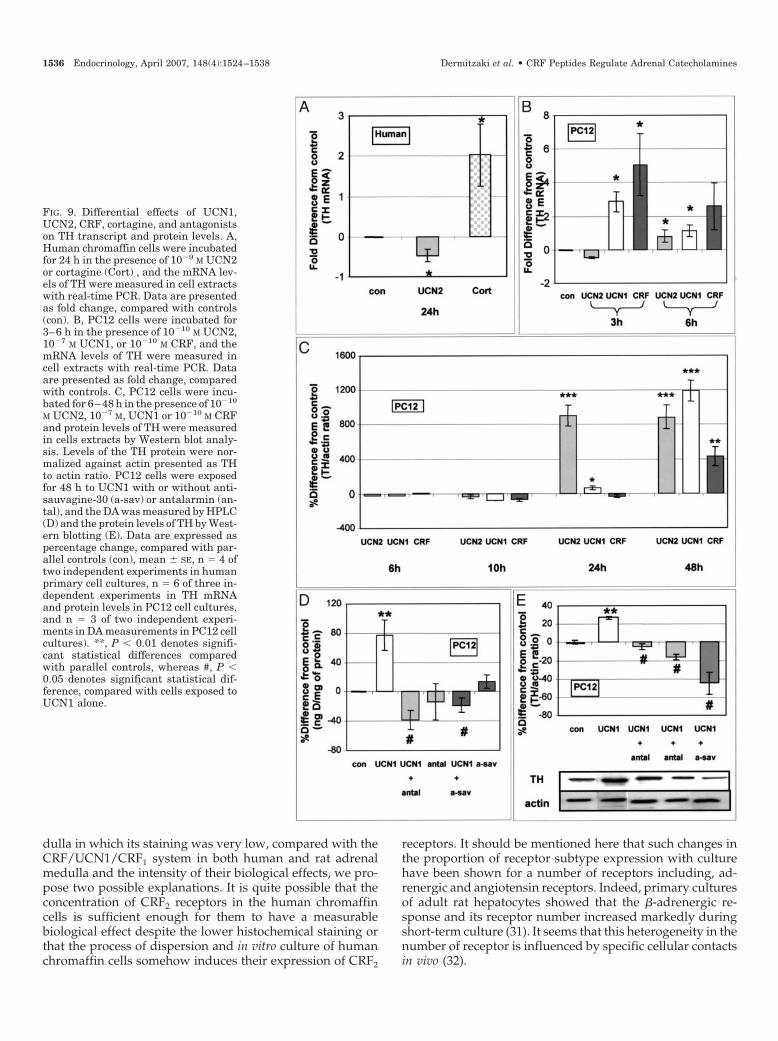
Effect of CRF, UCN1, UCN2, and cortagine on THexpression
The rate-limiting step in the biosynthesis of cat-echolamines is TH. Figure 9A depicts the quantification ofTH transcript by real-time PCR analysis of dispersed humanchromaffin cells exposed to UCN2 or cortagine for 24 h. Dataare presented as fold difference, compared with controls. Thepresence of UCN2 at 10�9 m for 24 h in the media reducedTH mRNA levels in a statistically significant manner by 0.5 �0.1-fold, compared with parallel controls, whereas the pres-ence of cortagine at 10�9 m elevated TH mRNA levels by 2.0 �0.77-fold, compared with the parallel controls. Similar resultswere observed at the protein level, as determined by Westernblot analysis (data not shown). Exposure of PC12 cells to CRFpeptides affected TH mRNA levels as measured by real-timePCR analysis (Fig. 9B). Exposure of UCN2 at 10�10 m, UCN1 at10�7 m, or CRF at 10�10 m for 6 h induced TH mRNA levelsby 0.8 � 0.3-, 1.1 � 0.3-, or 2.6 � 1.4-fold, compared withparallel controls. Similar results were observed at the proteinlevel, as determined by Western blot analysis. Thus, expo-sure of PC12 cells for 24 and 48 h to UCN2, UCN1, or CRFresulted in increased TH protein levels (Fig. 9C).
To confirm the involvement of both CRF receptors in thiseffect, we exposed PC12 cells to UCN1 for 48 h in the pres-ence or absence of antalarmin or antisauvagine-30 and DAwas measured (Fig. 9D). The stimulatory effect of UCN1 at10�7 m on DA was blocked by antalarmin at 10�5 m orantisauvagine-30 at 10�6 m.
We further confirmed our results by measuring the effectof UCN1 on TH production in PC12 cells in the absence orpresence of CRF1 and CRF2 antagonists. Cells were incubatedfor 48 h with 10�7 m UCN1 in the presence or absence ofantisauvagine-30 or antalarmin, and protein levels of TH
CRF1 TH DAPI CRF1 TH DAPI
CRF2 TH DAPI CRF2 TH DAPI
B.
C. CRF1 CRF2
CR F 1
CR F 2
503bp
425bp
BR
AIN
PC12
no
RT
no
cD
NA
NEG
CRF UCN1
FITC
A.
DAPI
UCN2
D.
NEG
FIG. 2. CRF peptides and their receptors CRF1 and CRF2 in PC12cells. A, PC12 cells were fixed, permeabilized, and then stained withantibodies against UCN2, CRF, and UCN1 as described in Materialsand Methods. DAPI was used for the visualization of cells nuclei. Cellsnot exposed to first antibody (NEG). B, PC12 cells were fixed, per-meabilized, and then stained with antibodies against CRF1 (first line;green dye), CRF2 (second line; green dye), and TH (second column; reddye) or stained with DAPI (third column; blue dye). Fourth columndepicts cells triple stained with CRF1/TH/DAPI (first line) or CRF2/TH/DAPI (second line). C, PC12 cells expressed CRF1 and CRF2 in themembrane section. Cells were fixed and then stained with antibodiesagainst CRF1 or CRF2. Cells not exposed to first antibody (NEG). D,RT-PCR analysis of the expression of CRF1 and CRF2 receptors inPC12 cells. Rat brain was used as positive control. No PCR productwas detected in the negative control samples. The results were rep-resentatives of two independent experiments.
0
20
40
0 0,5 1 1,5 2 2,5
Time (hrs)
ng
NE
/mg
of
pro
tein
conUCN1CRF*
FIG. 3. Time-dependent effects of UCN1 and CRF on NE secretionfrom dispersed rat chromaffin cells. Dispersed rat chromaffin cellswere incubated with UCN1 or CRF at several time points and NEsecretion was measured. UCN1 or CRF at 10�7 M induced NE secre-tion at 30 min. Data are expressed as nanogram NE per milligram ofprotein (mean � SE, n � 11 of four independent experiments for CRF,n � 6 of three independent experiments for UCN1). *, P � 0.05 denotessignificant statistical difference, compared with parallel controls.
1530 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines
were measured in cell extracts by Western blot analysis (Fig.9E). The stimulatory effects of UCN1 in TH production wereblocked by the CRF1 and CRF2 antagonists.
To confirm the data suggesting that CRF-related peptidesaffected the biosynthetic process of catecholamines, we ex-posed PC12 cells to CRF-related peptides in the presence orabsence of AMPT (a tyrosine hydroxylase inhibitor) or NSD-1015 (an l-aromatic amino acid decarboxylase inhibitor).Cells were exposed to 10�10 m UCN2, 10�8 m cortagine, 10�7
m UCN1, or 10�10 m CRF for 48 h with or without preincu-bation with 10�6 m AMPT or NSD-1015 for 1 h. The resultsshowed that either AMPT or NSD-1015 blocked the stimu-latory effect of CRF-related peptides on DA secretion (Fig.10). Indeed, the stimulatory effect of UCN2 at 10�10 m on DAwas blocked by AMPT or by NSD-1015. Similarly, the effectof cortagine at 10�8 m on DA was also blocked by AMPT orNSD-1015. Moreover, the stimulatory effect of UCN1 at 10�7
m on DA was blocked by AMPT or NSD-1015. Finally, thestimulatory effect of CRF at 10�10 m on DA was blocked byAMPT or NSD-1015.
Discussion
CRF, UCN1, UCN2, and the CRF1 and CRF2 receptors werepresent throughout the rat adrenal medulla. In comparison,whereas all CRF peptides and their receptors were also de-tectable in the cortex, the intensity of staining was definitelyless, compared with that in medulla with the exception ofzona glomerulosa. CRF, UCN1, UCN2, and the CRF1 and theCRF2 receptors were also present in the human adrenal me-dulla. The intensity of the CRF, UCN1, and CRF1 stainingwas directly comparable, in intensity, with that of rat adrenalmedulla, whereas the staining of UCN2 and CRF2 receptorwas much less. These data taken together give the impressionthat, in human adrenals, the CRF/UCN1/CRF1 system maypredominate in the medulla, whereas the opposite appearsto be true for the cortex in which the UCN2/CRF2 systemmay represent the dominant form of the CRF system, inagreement with recently published findings (13). These dif-ferences between human and rat adrenal vis-a-vis the distri-bution of the CRF peptides and their receptors may be dueto real differences between the two species or it may repre-sent an artifact because of the stress suffered by the victimsof the traffic accidents, the source of our human adrenals. TheCRF peptides and there receptors were also present in cul-tured PC12 cells.
Our functional studies on dispersed rat and human ad-renal chromaffin cells supports the above mentioned hy-pothesis, i.e. that the UCN1/CRF/CRF1 receptor systemplays a different physiological role, compared with that ofUCN2/CRF2 receptor system. Indeed, activation of the CRF1receptor resulted in induction of catecholamine secretion, an
or DA (PC12 cells) were measured. UCN2 suppressed catecholaminesecretion below control levels in both rat and human chromaffin aswell as PC12 cells (A–D). In contrast, cortagine induced catechol-amine secretion above control levels in both rat and human chromaf-fin and in PC12 cells (E–H). Data are expressed as percentage change,compared with control values (mean � SE, n � 3 of two independentexperiments in rat and human primary cell cultures and n � 4 of twoindependent experiments in PC12 cell cultures). *, P � 0.05 denotessignificant statistical difference, compared with parallel controls.
Cortagine.N UCN2
% D
iffe
ren
ce f
rom
co
ntr
ol
10-9 10-8 10-7
[M]
*
*
UCN2
-40
0
-60
20
control
10-10 10-1010-9 10-8 10-7
[M]
0
100
200
Cortagine
*
*50
150
control
% D
iffe
ren
ce f
rom
co
ntr
ol
A ERat Chromaffin cells: Norepinephrine
-20
Cortagine.E UCN2
BRat Chromaffin cells: Epinephrine
F
10-1010-9 10-8 10-7
[M]
**
UCN2
control
% D
iffe
ren
ce f
rom
co
ntr
ol
-50
0
50
10050
10-10 10-9 10-8
[M]
-50
0
Cortagine*
-25
25
control
% D
iffe
ren
ce f
rom
co
ntr
ol
C
UCN2
GCHuman Chromaffin cells: Epinephrine
10-1010-9 10-8 10-7
[M]*
*
UCN2
control
-50
-10
10
-30
-70% D
iffe
ren
ce f
rom
co
ntr
ol
100
200
10-1010-9 10-8 10-7
[M]
0
Cortagine
*
*
*
50
150
250
control% D
iffe
ren
ce f
rom
co
ntr
ol
Cortagine.D UCN2
0
0
0
0
0
D HPC12 cells: Dopamine
-80
0
10-10 10-9 10-8 10-7
[M]
40
* **
UCN2
-40
control
% D
iffe
ren
ce f
rom
co
ntr
ol
10-10 10-9 10-8 10-7
[M]
Cortagine*
*
control0
50
100
150
200
-50
% D
iffe
ren
ce f
rom
co
ntr
ol
FIG. 4. UCN2 suppressed whereas cortagine stimulated acute cate-cholamine secretion from dispersed rat and human chromaffin andPC12 cells. Rat and human chromaffin and PC12 cells were incubatedwith UCN2 or cortagine at 10�10 to 10�7 M for 30 min, and NE (ratchromaffin cells), E (rat chromaffin cells), E (human chromaffin cells),
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1531
FBRat Chromaffin cells: Epinephrine
10-10 10-9 10-8 10-7
[M]-50
50
100UCN1
*
0 control
% D
iffe
ren
ce f
rom
co
ntr
ol
% D
iffe
ren
ce f
rom
co
ntr
ol
10-10 10-9 10-8 10-7
[M]
50
100 CRF
*
0 control
-50
*
*
Rat Chromaffin cells: Norepinephrine
10-10 10-9 10-8 10-7
[M]
-50
50
100
UCN1
0 control
% D
iffe
ren
ce f
rom
co
ntr
ol
% D
iffe
ren
ce f
rom
co
ntr
ol
10-10 10-9 10-8 10-7
[M]
50
100 CRF *
0 control
A E
controlCHuman Chromaffin cells: Epinephrine
10-10 10-9 10-8 10-7
[M]
-100
-50
-25
UCN1
*
-75
control
% D
iffe
ren
ce f
rom
co
ntr
ol
0
*
*
10-10 10-9 10-8 10-7
[M]
-100
-50
-25
CRF
*
-75
% D
iffe
ren
ce f
rom
co
ntr
ol
0
* *
*
G
10-10 10-9 10-8 10-7
[M]
0
50 UCN1
-50
control
% D
iffe
ren
ce f
rom
co
ntr
ol
100
10-10 10-9 10-8 10-7
[M]
0
400 CRF
200
control
% D
iffe
ren
ce f
rom
co
ntr
ol
600
*
*
DPC12 cells: Dopamine
H
*
FIG. 5. Dose-dependent effect of UCN1 and CRF on cate-cholamine secretion from dispersed rat and human chromaf-fin and PC12 cells. Rat and human chromaffin and PC12 cellswere incubated with UCN1 or CRF at 10�10 to 10�7 M for 30min, and NE (rat chromaffin cells), E (rat chromaffin cells),E (human chromaffin cells), or DA (PC12 cells) were mea-sured. UCN1 and CRF affected catecholamine secretion in adose-dependent and tissue-specific manner. Data are ex-pressed as percentage change, compared with control values(mean � SE, n � 6 of three independent experiments in ratprimary cell cultures, n � 3 of two independent experimentsin human primary cell cultures, and n � 4 of two independentexperiments in PC12 cell cultures). *, P � 0.05 denotes sig-nificant statistical difference, compared with parallel con-trols.
1532 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines
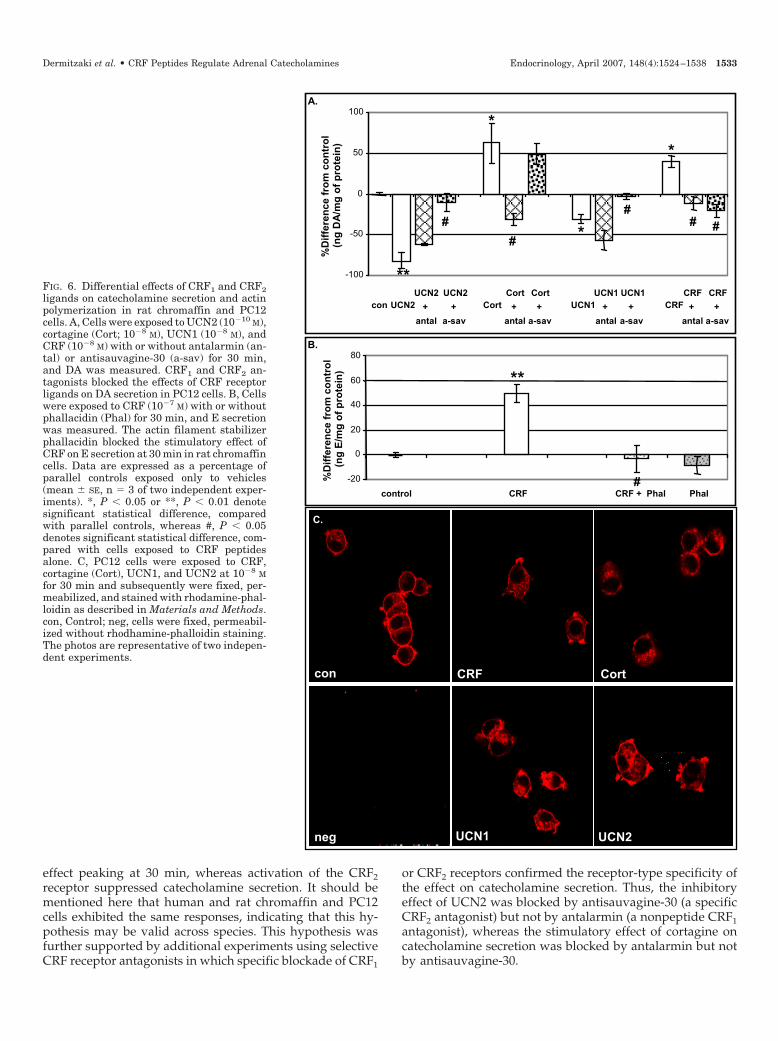
effect peaking at 30 min, whereas activation of the CRF2receptor suppressed catecholamine secretion. It should bementioned here that human and rat chromaffin and PC12cells exhibited the same responses, indicating that this hy-pothesis may be valid across species. This hypothesis wasfurther supported by additional experiments using selectiveCRF receptor antagonists in which specific blockade of CRF1
or CRF2 receptors confirmed the receptor-type specificity ofthe effect on catecholamine secretion. Thus, the inhibitoryeffect of UCN2 was blocked by antisauvagine-30 (a specificCRF2 antagonist) but not by antalarmin (a nonpeptide CRF1antagonist), whereas the stimulatory effect of cortagine oncatecholamine secretion was blocked by antalarmin but notby antisauvagine-30.
-100
-50
0
50
100
cnereffiD
%e
from
clortno
D gn(A
/mg
ofp
etor)ni
con CortCort
+antal
Cort+
a-sav
CRFCRF
+antal
CRF+
a-sav
UCN1UCN1
+antal
UCN1+
a-sav
UCN2+
a-sav
UCN2+
antal
UCN2
*
# ## *
#
**
#
A.
*
-20
0
20
40
60
80
control CRF CRF + Phal Phal
cnereffiD
%e
from
clortno
gm/E gn(
foetorpi
)n **
#
B.
neg
CRF Cortcon
UCN1 UCN2
C.
FIG. 6. Differential effects of CRF1 and CRF2ligands on catecholamine secretion and actinpolymerization in rat chromaffin and PC12cells. A, Cells were exposed to UCN2 (10�10 M),cortagine (Cort; 10�8 M), UCN1 (10�8 M), andCRF (10�8 M) with or without antalarmin (an-tal) or antisauvagine-30 (a-sav) for 30 min,and DA was measured. CRF1 and CRF2 an-tagonists blocked the effects of CRF receptorligands on DA secretion in PC12 cells. B, Cellswere exposed to CRF (10�7 M) with or withoutphallacidin (Phal) for 30 min, and E secretionwas measured. The actin filament stabilizerphallacidin blocked the stimulatory effect ofCRF on E secretion at 30 min in rat chromaffincells. Data are expressed as a percentage ofparallel controls exposed only to vehicles(mean � SE, n � 3 of two independent exper-iments). *, P � 0.05 or **, P � 0.01 denotesignificant statistical difference, comparedwith parallel controls, whereas #, P � 0.05denotes significant statistical difference, com-pared with cells exposed to CRF peptidesalone. C, PC12 cells were exposed to CRF,cortagine (Cort), UCN1, and UCN2 at 10�8 Mfor 30 min and subsequently were fixed, per-meabilized, and stained with rhodamine-phal-loidin as described in Materials and Methods.con, Control; neg, cells were fixed, permeabil-ized without rhodhamine-phalloidin staining.The photos are representative of two indepen-dent experiments.
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1533
The acute effects of CRF peptides on catecholamine se-cretion appear to be mediated by changes of actin filamentpolymerization, facilitating the exocytosis of the catechol-amine secretory vesicles. PC12 cells exposed to CRF1 ligandsshowed a broken subplasmalemmal ring of actin filamentswith areas of filamentous actin disassembly, whereas cellsexposed to CRF2 ligands showed an enhanced filamentousactin assembly in either the subplasmalemmal area beneaththe cell membrane or the cytoplasmic section. In rat chro-maffin cells, phallacidin, an actin filament stabilizer, blockedCRF1-induced catecholamine secretion. These data taken to-gether suggest that the effect of CRF1 or CRF2 ligands oncatecholamine secretion involved changes of actin filamentpolymerization. It should be noted that induction of thesevering protein 14–3-3 or sciderin results in an acute de-polymerization of acting and to a parallel elevation of cat-echolamine secretion from chromaffin cells (27, 28).
The effects of CRF1 and CRF2 receptor agonists were doseand receptor affinity specific. More specifically, the peaksuppressive effect of UCN2 on NE/E secretion from rat chro-maffin cells occurred at 10�10 m. At higher concentrations(10�8 to 10�7 m) UCN2 binds also to the CRF1 receptor (IC50of 350 nm, compared with 0.25 nm of CRF2), thus attenuatingits suppressive effect due to the antagonistic effect of CRF1activation. The same holds true for the synthetic CRF1 re-ceptor agonist cortagine. Indeed, its peak effect occurredbetween 10�10 to 10�9 m, at a concentration at which is a pureCRF1 receptor agonist, whereas at higher concentration (10�8
to 10�7 m), it starts binding the CRF2 receptor (IC50 2.6 nm forCRF1 receptor and 540 nm for CRF2 receptor (29)], thus at-tenuating its stimulatory effect. It should be noted that thebinding affinity of UCN1 to CRF1 is 0.17 nm and to CRF2 0.86nm; the binding affinity of CRF to CRF1 is around 1.6 nm andto CRF2 42 nm (30). CRF and UCN1 decreased E levels inhuman chromaffin cells, whereas they induce NE and DA inrat chromaffin and PC12 cells. The observed discrepancybetween human and rat adrenal chromaffin cells in theirresponse to CRF or UCN1 may be due to different stoichi-ometry of the two receptors present in human and rat chro-maffin cells at the time of the experiment or difference indesensitization mechanisms between the two species. Ac-cordingly, the net effect of UCN1 or CRF on catecholaminesecretion depends on the local availability of the receptors atthe time of their exposure to these peptides as well as theirlocal concentration.
CRF peptides also affected catecholamine synthesis, aneffect mediated by TH in both human and rodent adrenals.Indeed, in human chromaffin cells, the inhibitory effect ofUCN2 on TH mRNA levels at 24 h resulted in inhibition ofcatecholamine production at 48 h, whereas cortagine in-duced both the TH mRNA levels and catecholamine pro-duction. These results indicate that the effect of CRF-related
cells), or DA (PC12 cells) were measured. UCN2 and cortagine in-duced catecholamine synthesis in rat chromaffin and PC12 cells, ina dose-dependent manner. Exposure of dispersed human chromaffincells to cortagine induced catecholamine synthesis, whereas exposureto UCN2 suppressed catecholamine synthesis. Data are expressed aspercentage of parallel controls exposed only to vehicles (mean � SE,n � 3 of two independent experiments). *, P � 0.05 denotes significantstatistical difference, compared with parallel controls.
Cortagine.E
BRat Chromaffin cells: Epinephrine
F300
10-10 10-9 10-8
[M]
-100
100
Cortagine
*
0
200
control
% D
iffe
ren
ce f
rom
co
ntr
ol
UCN2
10-1010-9 10-8 10-7
[M]
**
UCN2
control
% D
iffe
ren
ce f
rom
co
ntr
ol
0
220
540
860
1180 *
*
*
10-7
A ERat Chromaffin cells: Norepinephrine
UCN2
% D
iffe
ren
ce f
rom
co
ntr
ol
10-9 10-8 10-7
[M]
*
*
UCN2
140
0
60
200
control10-10
220
Cor
10-10 10-9 10-8 10-7
[M]
-50
50
150Cortagine
**
0
100
control%
Dif
fere
nce
fro
m c
on
tro
l
*
UCN2
GCHuman Chromaffin cells: Epinephrine
10-1010-9 10-8 10-7
[M]* *
UCN2
control
-60
-20
0
-40
% D
iffe
ren
ce f
rom
co
ntr
ol
*
Cortagine200
10-1010-9 10-8 10-7
[M]
0
Cortagine
*
*50
control% D
iffe
ren
ce f
rom
co
ntr
ol
Cortagine.D
D HPC12 cells: Dopamine
10-10 10-9 10-8 10-7
[M]
Cortagine
**
control
-50
0
50
100
% D
iffe
ren
ce f
rom
co
ntr
ol
UCN2100
10-1010-9 10-8 10-7
[M]
0
UCN2
*
*60
control
% D
iffe
ren
ce f
rom
co
ntr
ol
-40
*
FIG. 7. Effects of UCN2 and cortagine on catecholamine synthesis bydispersed rat, human chromaffin, and PC12 cells. Cells were exposedto UCN2 (A–D) or cortagine (E–H) at 10�10 to 10�7 M at 48 h and NE(rat chromaffin cells), E (rat chromaffin cells), E (human chromaffin
1534 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines
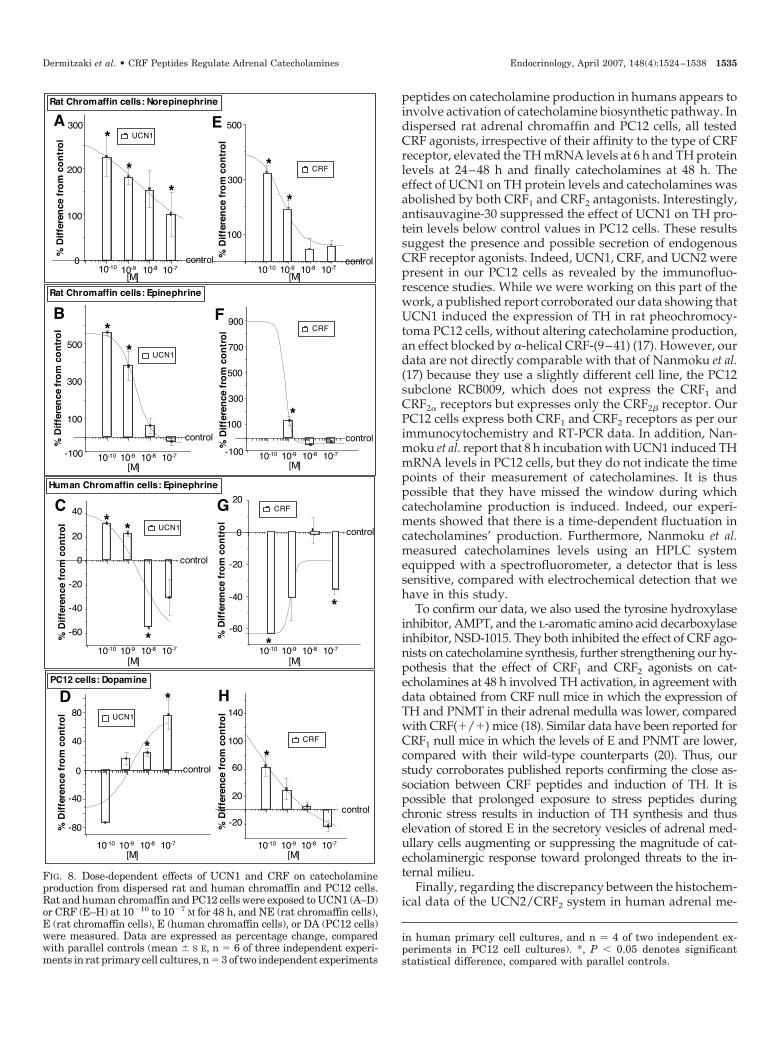
peptides on catecholamine production in humans appears toinvolve activation of catecholamine biosynthetic pathway. Indispersed rat adrenal chromaffin and PC12 cells, all testedCRF agonists, irrespective of their affinity to the type of CRFreceptor, elevated the TH mRNA levels at 6 h and TH proteinlevels at 24–48 h and finally catecholamines at 48 h. Theeffect of UCN1 on TH protein levels and catecholamines wasabolished by both CRF1 and CRF2 antagonists. Interestingly,antisauvagine-30 suppressed the effect of UCN1 on TH pro-tein levels below control values in PC12 cells. These resultssuggest the presence and possible secretion of endogenousCRF receptor agonists. Indeed, UCN1, CRF, and UCN2 werepresent in our PC12 cells as revealed by the immunofluo-rescence studies. While we were working on this part of thework, a published report corroborated our data showing thatUCN1 induced the expression of TH in rat pheochromocy-toma PC12 cells, without altering catecholamine production,an effect blocked by �-helical CRF-(9–41) (17). However, ourdata are not directly comparable with that of Nanmoku et al.(17) because they use a slightly different cell line, the PC12subclone RCB009, which does not express the CRF1 andCRF2� receptors but expresses only the CRF2� receptor. OurPC12 cells express both CRF1 and CRF2 receptors as per ourimmunocytochemistry and RT-PCR data. In addition, Nan-moku et al. report that 8 h incubation with UCN1 induced THmRNA levels in PC12 cells, but they do not indicate the timepoints of their measurement of catecholamines. It is thuspossible that they have missed the window during whichcatecholamine production is induced. Indeed, our experi-ments showed that there is a time-dependent fluctuation incatecholamines’ production. Furthermore, Nanmoku et al.measured catecholamines levels using an HPLC systemequipped with a spectrofluorometer, a detector that is lesssensitive, compared with electrochemical detection that wehave in this study.
To confirm our data, we also used the tyrosine hydroxylaseinhibitor, AMPT, and the l-aromatic amino acid decarboxylaseinhibitor, NSD-1015. They both inhibited the effect of CRF ago-nists on catecholamine synthesis, further strengthening our hy-pothesis that the effect of CRF1 and CRF2 agonists on cat-echolamines at 48 h involved TH activation, in agreement withdata obtained from CRF null mice in which the expression ofTH and PNMT in their adrenal medulla was lower, comparedwith CRF(�/�) mice (18). Similar data have been reported forCRF1 null mice in which the levels of E and PNMT are lower,compared with their wild-type counterparts (20). Thus, ourstudy corroborates published reports confirming the close as-sociation between CRF peptides and induction of TH. It ispossible that prolonged exposure to stress peptides duringchronic stress results in induction of TH synthesis and thuselevation of stored E in the secretory vesicles of adrenal med-ullary cells augmenting or suppressing the magnitude of cat-echolaminergic response toward prolonged threats to the in-ternal milieu.
Finally, regarding the discrepancy between the histochem-ical data of the UCN2/CRF2 system in human adrenal me-
Rat Chromaffin cells: Norepinephrine
A
10-10 10-9 10-8 10-7
[M]
0
200
300UCN1*
100
control
% D
iffe
ren
ce f
rom
co
ntr
ol
**
% D
iffe
ren
ce f
rom
co
ntr
ol
10-10 10-9 10-8 10-7
[M]
300
500
CRF*
100
control
*
E
Rat Chromaffin cells: Epinephrine
B
10-10 10-9 10-8 10-7
[M]
-100
300
500UCN1
*
100
control
% D
iffe
ren
ce f
rom
co
ntr
ol
*
F
% D
iffe
ren
ce f
rom
co
ntr
ol
10-10 10-9 10-8 10-7
[M]
500
700
CRF
100
control
-100
*300
900
Human Chromaffin cells: Epinephrine
G
10-10 10-9 10-8 10-7
[M]
-60
-20
0
CRF
*-40
control
% D
iffe
ren
ce f
rom
co
ntr
ol
20
*
C
10-10 10-9 10-8 10-7
[M]
-60
0
20UCN1*
-40
control
% D
iffe
ren
ce f
rom
co
ntr
ol
40
*
*
-20
H
10-10 10-9 10-8 10-7
[M]
0
40
UCN1
-40
control
% D
iffe
ren
ce f
rom
co
ntr
ol 80
-80
*
*D
10-10 10-9 10-8 10-7
[M]
-20
60
CRF
20
control
% D
iffe
ren
ce f
rom
co
ntr
ol
100
*
140
PC12 cells: Dopamine
FIG. 8. Dose-dependent effects of UCN1 and CRF on catecholamineproduction from dispersed rat and human chromaffin and PC12 cells.Rat and human chromaffin and PC12 cells were exposed to UCN1 (A–D)or CRF (E–H) at 10�10 to 10�7 M for 48 h, and NE (rat chromaffin cells),E (rat chromaffin cells), E (human chromaffin cells), or DA (PC12 cells)were measured. Data are expressed as percentage change, comparedwith parallel controls (mean � S E, n � 6 of three independent experi-ments in rat primary cell cultures, n � 3 of two independent experiments
in human primary cell cultures, and n � 4 of two independent ex-periments in PC12 cell cultures). *, P � 0.05 denotes significantstatistical difference, compared with parallel controls.
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1535
dulla in which its staining was very low, compared with theCRF/UCN1/CRF1 system in both human and rat adrenalmedulla and the intensity of their biological effects, we pro-pose two possible explanations. It is quite possible that theconcentration of CRF2 receptors in the human chromaffincells is sufficient enough for them to have a measurablebiological effect despite the lower histochemical staining orthat the process of dispersion and in vitro culture of humanchromaffin cells somehow induces their expression of CRF2
receptors. It should be mentioned here that such changes inthe proportion of receptor subtype expression with culturehave been shown for a number of receptors including, ad-renergic and angiotensin receptors. Indeed, primary culturesof adult rat hepatocytes showed that the �-adrenergic re-sponse and its receptor number increased markedly duringshort-term culture (31). It seems that this heterogeneity in thenumber of receptor is influenced by specific cellular contactsin vivo (32).
FIG. 9. Differential effects of UCN1,UCN2, CRF, cortagine, and antagonistson TH transcript and protein levels. A,Human chromaffin cells were incubatedfor 24 h in the presence of 10�9 M UCN2or cortagine (Cort) , and the mRNA lev-els of TH were measured in cell extractswith real-time PCR. Data are presentedas fold change, compared with controls(con). B, PC12 cells were incubated for3–6 h in the presence of 10�10 M UCN2,10�7 M UCN1, or 10�10 M CRF, and themRNA levels of TH were measured incell extracts with real-time PCR. Dataare presented as fold change, comparedwith controls. C, PC12 cells were incu-bated for 6–48 h in the presence of 10�10
M UCN2, 10�7 M, UCN1 or 10�10 M CRFand protein levels of TH were measuredin cells extracts by Western blot analy-sis. Levels of the TH protein were nor-malized against actin presented as THto actin ratio. PC12 cells were exposedfor 48 h to UCN1 with or without anti-sauvagine-30 (a-sav) or antalarmin (an-tal), and the DA was measured by HPLC(D) and the protein levels of TH by West-ern blotting (E). Data are expressed aspercentage change, compared with par-allel controls (con), mean � SE, n � 4 oftwo independent experiments in humanprimary cell cultures, n � 6 of three in-dependent experiments in TH mRNAand protein levels in PC12 cell cultures,and n � 3 of two independent experi-ments in DA measurements in PC12 cellcultures). **, P � 0.01 denotes signifi-cant statistical differences comparedwith parallel controls, whereas #, P �0.05 denotes significant statistical dif-ference, compared with cells exposed toUCN1 alone.
1536 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines

In conclusion, our data suggest that a complex in-tramedullary CRF-UCN/CRF receptor system exists inboth human and rat adrenals, controlling catecholaminesynthesis and secretion. Activation of either the CRF1 orCRF2 receptors results in opposing effects on catechol-amine secretion. Thus, the local concentration of CRF pep-tides in the adrenals, and perhaps also in the brain, mayaffect catecholamine secretion and synthesis under a com-plex mechanism that depends on the ad hoc concentrationof ligands and receptor availability. Our data, validated inprimary human and rat chromaffin cells, highlight a po-tential difference between primates and rodents in thelocal regulation of catecholamine secretion.
Acknowledgments
Received July 19, 2006. Accepted December 20, 2006.Address all correspondence and requests for reprints to: Andrew N.
Margioris, Department of Clinical Chemistry-Biochemistry, School ofMedicine, University of Crete, Heraklion GR-710 03, Crete, Greece. E-mail: [email protected].
This work was supported by the Hellenic Secretariat for Research andTechnology (�ENE 01E386) and a grant from Medicon Hellas Co.
Disclosure Statement: E.D., C.T., V.M., E.C., I.C., M.V., A.A., M.L., J.S.,E.M., A.G., and A.N.M. have nothing to declare.
References
1. Suda T, Tomori N, Tozawa F, Mouri T, Demura H, Shizume K 1984 Distri-bution and characterization of immunoreactive corticotropin-releasing factorin human tissues. J Clin Endocrinol Metab 59:861–866
2. Hashimoto K, Murakami K, Hattori T, Niimi M, Fujino K, Ota Z 1984Corticotropin-releasing factor (CRF)-like immunoreactivity in the adrenal me-dulla. Peptides 5:707–711
3. Dave JR, Eiden LE, Eskay RL 1985 Corticotropin-releasing factor binding toperipheral tissue and activation of the adenylate cyclase-adenosine 3�,5�-mono-phosphate system. Endocrinology 116:2152–2159
4. Udelsman R, Harwood JP, Millan MA, Chrousos GP, Goldstein DS, Zim-lichman R, Catt KJ, Aguilera G 1986 Functional corticotropin releasing factorreceptors in the primate peripheral sympathetic nervous system. Nature 319:147–150
5. Aguilera G, Millan MA, Hauger RL, Catt KJ 1987 Corticotropin-releasingfactor receptors: distribution and regulation in brain, pituitary, and peripheraltissues. Ann NY Acad Sci 512:48–66
6. Bruhn TO, Engeland WC, Anthony EL, Gann DS, Jackson IM 1987 Corti-cotropin-releasing factor in the dog adrenal medulla is secreted in response tohemorrhage. Endocrinology 120:25–33
7. Willenberg HS, Bornstein SR, Hiroi N, Path G, Goretzki PE, ScherbaumWA, Chrousos GP 2000 Effects of a novel corticotropin-releasing-hormone
receptor type I antagonist on human adrenal function. Mol Psychiatry5:137–141
8. Chatzaki E, Margioris AN, Gravanis A 2002 Expression and regulation ofcorticotropin-releasing hormone binding protein (CRH-BP) in rat adrenals.J Neurochem 80:81–90
9. Takahashi K, Totsune K, Saruta M, Fukuda T, Suzuki T, Hirose T, Imai Y,Sasano H, Murakami O 2005 Expression of urocortin 3/stresscopin in humanadrenal glands and adrenal tumors. Peptides 27:178–182
10. Mazzocchi G, Malendowicz LK, Rebuffat P, Gottardo G, Nussdorfer GG1997 Neurotensin stimulates CRH and ACTH release by rat adrenal medullain vitro. Neuropeptides 31:8–11
11. Oki Y, Iwabuchi M, Masuzawa M, Watanabe F, Ozawa M, Iino K, To-minaga T, Yoshimi T 1998 Distribution and concentration of urocortin, andeffect of adrenalectomy on its content in rat hypothalamus. Life Sci 62:807– 812
12. Yamauchi N, Otagiri A, Nemoto T, Sekino A, Oono H, Kato I, Yanaihara C,Shibasaki T 2005 Distribution of urocortin 2 in various tissues of the rat.J Neuroendocrinol 17:656–663
13. Fukuda T, Takahashi K, Suzuki T, Saruta M, Watanabe M, Nakata T, SasanoH 2005 Urocortin 1, urocortin 3/stresscopin, and corticotropin-releasing factorreceptors in human adrenal and its disorders. J Clin Endocrinol Metab 90:4671–4678
14. Venihaki M, Gravanis A, Margioris AN 1997 Comparative study betweennormal rat chromaffin and PC12 rat pheochromocytoma cells: productionand effects of corticotropin-releasing hormone. Endocrinology 138:698 –704
15. Venihaki M, Ain K, Dermitzaki E, Gravanis A, Margioris AN 1998 KAT45,a noradrenergic human pheochromocytoma cell line producing corticotropin-releasing hormone. Endocrinology 139:713–722
16. Margioris AN, Venihaki M, Gravanis A 2001 Interleukin (IL)-1 family ofcytokines and corticotropin-releasing hormone (CRH) in the adrenal gland.Adrenal disorders. Totowa, NJ: Humana Press
17. Nanmoku T, Takekoshi K, Fukuda T, Isobe K, Shibuya S, Kawakami Y 2005Urocortin stimulates tyrosine hydroxylase activity via the cAMP/protein ki-nase a pathway in rat pheochromocytoma PC12 cells. Neurosci Lett 382:124–127
18. Kubovcakova L, Sabban EL, Kvetnansky R, Krizanova O 2002 Comparativestudy of catecholamine synthesizing enzymes in adrenal medulla of CRHknock-out mice, their CRH (�/�) mates and Sprague-Dawley rats. EndocrRegul 36:107–113
19. Kubovcakova L, Tybitanclova K, Sabban EL, Majzoub J, Zorad S, Vietor I,Wagner EF, Krizanova O, Kvetnansky R 2004 Catecholamine synthesizingenzymes and their modulation by immobilization stress in knockout mice. AnnNY Acad Sci 1018:458–465
20. Yoshida-Hiroi M, Bradbury MJ, Eisenhofer G, Hiroi N, Vale WW, NovotnyGE, Hartwig HG, Scherbaum WA, Bornstein SR 2002 Chromaffin cell func-tion and structure is impaired in corticotropin-releasing hormone receptortype 1-null mice. Mol Psychiatry 7:967–974
21. Dermitzaki E, Gravanis A, Venihaki M, Stournaras C, Margioris AN 2001Opioids suppress basal and nicotine-induced catecholamine secretion viaa stabilizing effect on actin filaments. Endocrinology 142:2022–2031
22. Chatzaki E, Crowe PD, Wang L, Million M, Tache Y, Grigoriadis DE 2004CRF receptor type 1 and 2 expression and anatomical distribution in the ratcolon. J Neurochem 90:309–316
23. Chatzaki E, Murphy BJ, Wang L, Million M, Ohning GV, Crowe PD, PetroskiR, Tache Y, Grigoriadis DE 2004 Differential profile of CRF receptor distri-bution in the rat stomach and duodenum assessed by newly developed CRFreceptor antibodies. J Neurochem 88:1–11
FIG. 10. Effect of CRF-related peptides on thepathway of catecholamine biosynthesis. Cellswere exposed to 10�10 M UCN2, 10�8 M cortagine(Cort), 10�7 M UCN1, or 10�10 M CRF with orwithout AMPT at 10�6 M or NSD-1015 (NSD) at10�6 M for 48 h. The effects of CRF-related pep-tides were completely blocked by either AMPT orNSD-1015. Data are expressed as a percentagechange, compared with parallel controls exposedonly to vehicles (mean � SE, n � 6 of three inde-pendent experiments). **, P � 0.01 denotes sig-nificant statistical difference, compared with par-allel controls, whereas #, P � 0.05 denotessignificant statistical difference, compared withcells exposed to CRF peptides alone.
Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines Endocrinology, April 2007, 148(4):1524–1538 1537

24. Charalampopoulos I, Dermitzaki E, Vardouli L, Tsatsanis C, Stournaras C,Margioris AN, Gravanis A 2005 Dehydroepiandrosterone sulfate and allo-pregnanolone directly stimulate catecholamine production via induction oftyrosine hydroxylase and secretion by affecting actin polymerization. Endo-crinology 146:3309–3318
25. Laemmli UK 1970 Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227:680–685
26. Dermitzaki E, Tsatsanis C, Charalampopoulos I, Androulidaki A, AlexakiVI, Castanas E, Gravanis A, Margioris AN 2005 Corticotropin-releasing hor-mone activates protein kinase C in an isoenzyme-specific manner. BiochemBiophys Res Commun 327:828–836
27. Roth D, Burgoyne RD 1995 Stimulation of catecholamine secretion fromadrenal chromaffin cells by 14-3-3 proteins is due to reorganisation of thecortical actin network. FEBS Lett 374:77–81
28. Zhang L, Rodriguez Del Castillo A, Trifaro JM 1995 Histamine-evoked chro-maffin cell scinderin redistribution, F-actin disassembly, and secretion: in the
absence of cortical F-actin disassembly, an increase in intracellular Ca2� failsto trigger exocytosis. J Neurochem 65:1297–1308
29. Tezval H, Jahn O, Todorovic C, Sasse A, Eckart K, Spiess J 2004 Cortagine,a specific agonist of corticotropin-releasing factor receptor subtype 1, is anx-iogenic and antidepressive in the mouse model. Proc Natl Acad Sci USA101:9468–9473
30. Jahn O, Tezval H, van Werven L, Eckart K, Spiess J 2004 Three-amino acidmotifs of urocortin II and III determine their CRF receptor subtype selectivity.Neuropharmacology 47:233–242
31. Nakamura T, Tomomura A, Noda C, Shimoji M, Ichihara A 1983 Acquisitionof a �-adrenergic response by adult rat hepatocytes during primary culture.J Biol Chem 258:9283–9289
32. Hansson E 1985 Primary cultures from defined brain areas; effects of seedingtime on the development of �-adrenergic- and dopamine-stimulated cAMP-activity during cultivation. Brain Res 353:187–192
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
1538 Endocrinology, April 2007, 148(4):1524–1538 Dermitzaki et al. • CRF Peptides Regulate Adrenal Catecholamines



















