
Potentials for organic agriculture to sustain livelihoods in Tanzania
Upload
independentCategory
view
6download
0
PII S0361-9230(00)00437-8
Cortical networks for working memory and executivefunctions sustain the conscious resting state in man
B. Mazoyer, L. Zago, E. Mellet, S. Bricogne, O. Etard, O. Houde, F. Crivello, M. Joliot, L. Petit andN. Tzourio-Mazoyer*
Groupe d’Imagerie Neurofonctionnelle, UMR6095, CNRS, LEA, Universite de Caen,Universite Paris 5, France
[Accepted 26 October 2000]
ABSTRACT: The cortical anatomy of the conscious resting state(REST) was investigated using a meta-analysis of nine positronemission tomography (PET) activation protocols that dealt withdifferent cognitive tasks but shared REST as a common controlstate. During REST, subjects were in darkness and silence, andwere instructed to relax, refrain from moving, and avoid sys-tematic thoughts. Each protocol contrasted REST to a differentcognitive task consisting either of language, mental imagery,mental calculation, reasoning, finger movement, or spatialworking memory, using either auditory, visual or no stimulusdelivery, and requiring either vocal, motor or no output. A totalof 63 subjects and 370 spatially normalized PET scans wereentered in the meta-analysis. Conjunction analysis revealed anetwork of brain areas jointly activated during conscious RESTas compared to the nine cognitive tasks, including the bilateralangular gyrus, the left anterior precuneus and posterior cingu-late cortex, the left medial frontal and anterior cingulate cortex,the left superior and medial frontal sulcus, and the left inferiorfrontal cortex. These results suggest that brain activity duringconscious REST is sustained by a large scale network of het-eromodal associative parietal and frontal cortical areas, thatcan be further hierarchically organized in an episodic workingmemory parieto-frontal network, driven in part by emotions,working under the supervision of an executive left prefrontalnetwork. © 2001 Elsevier Science Inc.
KEY WORDS: Consciousness, Working memory, Executivefunctions, Resting state, Brain activation, PET.
INTRODUCTION
A large part of our daily mental activities are internalized,id estperformed without external input or motor output, and not goaldirected. During this particular state of consciousness, that is not tobe confounded with arousal or perceptual consciousness, one ismonitoring both somesthesic and vegetative information, such assensations and body position, and experiencing association of freethoughts that deal with the recollection of past experiences, innerspeech, mental images, emotions, planning of future activities, etc.This mental state has been referred to as a Random Episodic SilentThinking (REST) state by previous authors [3], thereby emphasiz-ing the unconstrained nature of this kind of thoughts, as opposed
to a more focused and constrained sort of episodic memory activitywhen one is driven to search into his past history.
Despite the fact that assessing the exact mental content of asubject during REST is by essence difficult and must rely onintrospection, which clearly constitutes a scientific limitation, thereis a considerable interest in the study of the neural bases of theconscious resting state.
First, from a strict cognitive neuroscience point of view, it isimportant to try to establish whether or not a network of brainareas is specifically engaged during this mental state brain. If so,this would provide a strong argument favoring the “computation-al” nature of brain activity during REST [7] and, depending on thelocations of these areas, insights regarding the major cognitiveprocesses involved.
The answer to this question is also of primary importance forcognitive neuroimaging experiments, because conscious rest hasbeen (and is still) widely used in the cognitive neuroimagingcommunity as a reference state to which better controlled cognitiveprocesses are contrasted. This is explicitly the case for positronemission tomography (PET) or functional magnetic resonanceimaging (FMRI) experiments, in which local neural activity duringconscious rest serves as a baseline for assessing hemodynamicvariations during cognitive process execution. To some extent, it isalso implicitly the case for event-related electroencephalographyor magnetoencephalography experiments, because electrical/mag-netic activity is generally compared to a baseline activity measuredover the several tens/hundreds of milliseconds that precede stim-ulus presentation when the subject is in a state that resembles toconscious rest.
Although it is an ill-defined mental state, there are some goodreasons for using conscious rest as a reference state in cognitiveneuroimaging. First, it is applicable in all imaging experiments, asopposed to high level cognitive control tasks that have been usedto test the involvement of a specific cognitive module, with thepotential drawback of masking joint activations in both the refer-ence and the task of interest [24]. As a matter of fact, differencesin the reference conditions have been put forward in order to try tosolve controversies regarding the putative brain network involvedin identical tasks [38]. As such, the conscious resting state canserve as a common reference both within and between laboratory
* Address for correspondence: Dr. Nathalie Tzourio-Mazoyer, Groupe d’Imagerie Neurofonctionnelle, GIP Cyceron, Bd Becquerel BP 5229, 14074 CaenCedex France. Fax:133-231-470-222; E-mail: [email protected]
Brain Research Bulletin, Vol. 54, No. 3, pp. 287–298, 2001Copyright © 2001 Elsevier Science Inc.Printed in the USA. All rights reserved
0361-9230/01/$–see front matter
287
experiments. Second, the conscious resting state does not rely ona given sensory modality for stimulus delivery or on a specific kindof behavioral output. Third, its variability both within and acrosssubjects has been shown to be of the same magnitude than anyother cognitive task [14].
Studies on the cortical anatomy of the resting state have beenfirst relying on the observation of the regional pattern of theglucose metabolic rate (rCMRGlu) or regional cerebral blood flow(rCBF) [14,42,55]. These studies revealed brain areas of higher orlower metabolic/hemodynamic activity, provided figures of theirlocal variability, but did not allow to uncover a specific network ofbrain areas involved during the resting state. A recent PET exper-iment compared rCMRGlu maps in healthy volunteers at REST tothat of patients in a vegetative state [40], showing a widelydistributed network of brain areas having higher neural activityduring REST. A recent FMRI experiment has focused on the issueof the neural bases of the resting state, contrasting REST toauditory perceptual or semantic tasks [7]. A left hemisphere pari-eto-frontal network was found of equal activity during REST andthe semantic task, but of reduced activity during the perceptualtask, leading the authors to propose that mental activity duringREST was of “conceptual processing” nature.
However, this claim remains to be formally demonstrated be-cause it relied on simple contrast analysis (RESTvs. tone, RESTvs. semantic, etc.), concerned task involving the sole auditorymodality, with possible unwanted interference of attentional pro-cess due to scanner noise, and was based on a somewhat limitedsample size, which raises a sensitivity issue. One way of alleviat-ing these limits could be through a meta-analysis of activationprotocols in which a REST condition was used. The idea of usinga meta-analysis to uncover process common to different tasks hasbeen proposed and previously exploited by others [63–66]. In theirapproach, Shulman et al. reanalyzed a set of 9 PET experimentscontrasting passivevs. active visual tasks, using activation mapaveraging in order to uncover processes that generalize acrosstasks. This approach has the potential drawback that it could revealactivation that would not necessarily be common to all protocolsbut rather driven by the strongest ones, with possibly reducedactivity in some others. Actually, conjunction analysis appears asthe optimal statistical approach of choice when trying to identifyjoint activations across protocols [57].
The present study was designed to alleviate some of the con-
cerns raised by previous studies dealing with the neural bases ofthe conscious resting state. It combines the intrinsic power ofmeta-analysis, the specificity of conjunction analysis, and a varietyof cognitive tasks to be contrasted to REST. Accordingly, a set ofnine PET activation protocols performed in our laboratory over thepast 3 years, all including REST as a control task, have beenreanalyzed using conjunction analysis, searching for joint activa-tion during REST as compared to a variety of cognitive tasks,including different stimulus presentation modalities, cognitive pro-cesses, and behavioral outputs.
MATERIALS AND METHODS
Subjects
Sixty-three young healthy male volunteers participated to thestudy (22.06 2.0 years, mean6 SD). All were right-handed asassessed by the Edinburgh questionnaire (88.16 13.7; range,50–100) and were free of brain abnormalities as assessed on theirT1-weighted 3D MRI scan. All subjects gave their informed con-sent. The study was approved by the Basse-Normandie EthicCommittee.
Cognitive Tasks
Each of the 63 subjects participated in one and only one of ninedifferent activation protocols (see Table 1). Each protocol con-sisted of repeated cerebral blood flow mapping with PET during aseries of cognitive tasks, including a resting state condition.
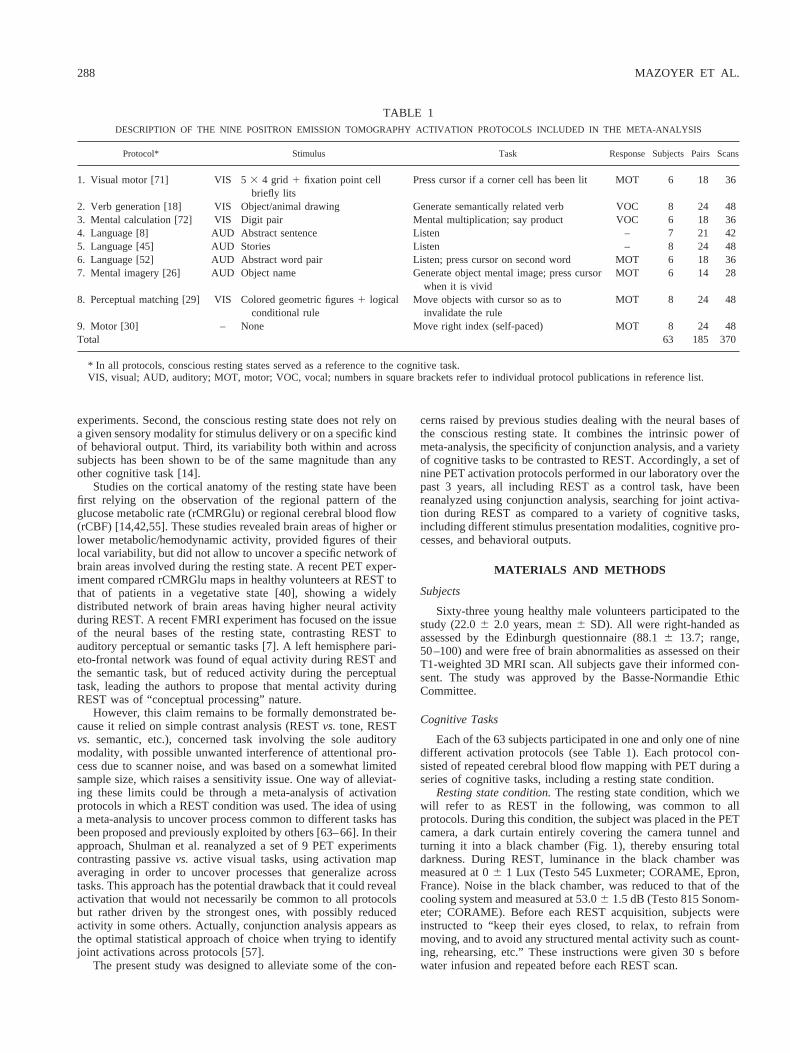
Resting state condition.The resting state condition, which wewill refer to as REST in the following, was common to allprotocols. During this condition, the subject was placed in the PETcamera, a dark curtain entirely covering the camera tunnel andturning it into a black chamber (Fig. 1), thereby ensuring totaldarkness. During REST, luminance in the black chamber wasmeasured at 06 1 Lux (Testo 545 Luxmeter; CORAME, Epron,France). Noise in the black chamber, was reduced to that of thecooling system and measured at 53.06 1.5 dB (Testo 815 Sonom-eter; CORAME). Before each REST acquisition, subjects wereinstructed to “keep their eyes closed, to relax, to refrain frommoving, and to avoid any structured mental activity such as count-ing, rehearsing, etc.” These instructions were given 30 s beforewater infusion and repeated before each REST scan.
TABLE 1DESCRIPTION OF THE NINE POSITRON EMISSION TOMOGRAPHY ACTIVATION PROTOCOLS INCLUDED IN THE META-ANALYSIS
Protocol* Stimulus Task Response Subjects Pairs Scans
1. Visual motor [71] VIS 53 4 grid 1 fixation point cellbriefly lits
Press cursor if a corner cell has been lit MOT 6 18 36
2. Verb generation [18] VIS Object/animal drawing Generate semantically related verb VOC 8 24 483. Mental calculation [72] VIS Digit pair Mental multiplication; say product VOC 6 18 364. Language [8] AUD Abstract sentence Listen – 7 21 425. Language [45] AUD Stories Listen – 8 24 486. Language [52] AUD Abstract word pair Listen; press cursor on second word MOT 6 18 367. Mental imagery [26] AUD Object name Generate object mental image; press cursor
when it is vividMOT 6 14 28
8. Perceptual matching [29] VIS Colored geometric figures1 logicalconditional rule
Move objects with cursor so as toinvalidate the rule
MOT 8 24 48
9. Motor [30] – None Move right index (self-paced) MOT 8 24 48Total 63 185 370
* In all protocols, conscious resting states served as a reference to the cognitive task.VIS, visual; AUD, auditory; MOT, motor; VOC, vocal; numbers in square brackets refer to individual protocol publications in reference list.
288 MAZOYER ET AL.
Upon completion of the PET study, subjects had to answer adetailed questionnaire in order to assess the content of their mentalactivity during the REST conditions. In the questionnaire, thesubjects were asked to subjectively describe the nature of theirmental activity, particularly with respect to the presence of mentalimages, inner speech, memory reminiscences, and somesthesicsensations.
Cognitive tasks.Nine different cognitive tasks, one for eachprotocol, were contrasted to the REST condition: visuomotor, verbgeneration, mental calculation, listening to language stimuli (threeprotocols), visual mental imagery, perceptual matching, and self-paced movement (see Table 1). Stimuli were delivered to thesubjects using either the visual or the auditory modality (fourprotocols each), while responses required finger movement (fiveprotocols) or vocalization (two protocols, see Table 2). Auditorystimuli were recorded on digital tapes and binaurally deliveredusing earphones, while visual stimuli were presented on computerscreen, placed inside the black chamber, that the subject could
view thanks to a mirror attached to the head holder. Responseswere recorded via using either a computer device or a microphone.
Characteristics of the nine cognitive tasks used in the presentstudy have been published elsewhere in some form (see referencesbelow) and will only be sketched here.
FIG. 1. Experimental setup for positron emission tomography (PET) experiments. Black curtains are attached tothe front and rear of the PET camera ensuring total darkness during the conscious resting state condition(luminance in the camera tunnel 06 1 Lux). The black chamber at the PET camera rear can accommodate a PCfor visual stimulus delivery.
TABLE 2NUMBER OF POSITRON EMISSION TOMOGRAPHY ACTIVATION
PROTOCOLS PER STIMULUS/RESPONSE TYPES
Response
Stimulus
Visual Auditory None
Motor 2 2 1Vocal 2 – –None – 2 –
NEURAL BASES OF THE CONSCIOUS RESTING STATE 289

In the visuomotor task (#1) [71], subjects were presented a 435 grid on a computer screen, including a fixation point at its centeron which they were to keep their gaze. Grid cells were succes-sively and randomly lit for 1 s; the subject had to press a buttonwhenever a corner has been lit.
The visual language task (#2) [18] consisted in the presentationof black line drawings of objects (73%) or animals (27%), on awhite computer screen, at a rate of 0.25 Hz. Subjects were re-quested to generate aloud a verb semantically related to the dis-played object/animal drawing.
The calculation task (#3) [72] consisted in simple arithmeticfacts. The subjects had to overtly name the result of the mentalmultiplication of pairs of digits presented on a computer screen atan average rate of 1.25 Hz.
The auditory language tasks consisted in listening to either (1)abstract word pairs presented at a rate of 0.12 Hz (#4) [8], (2)abstract word definitions (#5) [45] consisting in short sentenceslasting 6 s and followed by a 2-s silent delay, or (3) 2-minute longfactual stories (#6) [52]. In all three tasks, subjects were asked toattentively but passively listen to the stimuli that were given in thesubject’s mother tongue.
The mental imagery task (#7) [26] consisted of generating andmaintaining a vivid visual mental image of an object upon hearingits name. Objects’ names were delivered through earphones at a0.1 Hz rate.
During the perceptual matching task (#8) [29], subjects werepresented with 12 colored geometric shapes on a computer screen,and a logical conditional rule such as, for example, “If there is nota red square on the left, then there is a yellow circle on the right”.The task was to move, using the computer mouse, two of theshapes in a two-part box drawn on the screen so as to falsify therule.
Finally, the motor task (#9) [30] consisted in self-paced rightindex flexion and extension that were performed at an average rateof 0.75 Hz.
PET Data Acquisition
The same experimental apparatus and design was used for allPET studies. The subject head was positioned in the PET camerafield of view, immobilization being ensured by means of a customhead holder attached to the camera bed. rCBF was monitored usingPET and oxygen 15 labeled water bolus injection in the left basilicvein. For each rCBF measurement, a single 90-s scan was acquiredusing an ECAT HR1 PET camera (SIEMENS, Erlangen, Ger-many) with septa retracted (3D mode), starting at the arrival of the15O labeled water. Each scan was reconstructed with a 0.5 mm21
cut-off frequency Hanning filter, including correction for attenua-tion (using an acquired 130 Mevents 2D transmission scan), ran-doms and scatter. The time between two PET measurements was8 min.
Image Standardization
rCBF images were spatially normalized using the SPM package(SPM96 version, [23]) to the Montreal Neurological Institute(MNI) template, and filtered using a 12-mm Gaussian filter leadingto a final image smoothness of 12.13 13.6 3 15.1 mm3. Rawcounts volumes were proportionally normalized to 50 ml/min per100 g.
Statistical Analysis
The REST condition was compared to the set of the nine othercognitive tasks using a conjunction analysis [57] as implementedin the SPM package (SPM99 version), searching for brain areas
jointly activated or deactivated during REST as compared to thenine cognitive tasks. Depending on the protocol, two or threereplicates of both the cognitive task and the REST condition wereavailable for each of the 63 subjects, leading to a total of 185 pairsof REST/TASK scans (see Table 1). A multigroup conjunctionanalysis was performed using SPM99, with a significance level setat 0.05 corrected for multiple comparisons.
Anatomical localization of activation/deactivation foci was per-formed using anatomical landmarks of the single subject MRInormalized to MNI template provided by SPM; in order to com-pare our results with that of previous studies, each focus was alsoassigned a Brodmann’s area (BA) number based on its stereotacticcoordinates and the Talairach atlas [67]. Conjunction maps ob-served in the present study were compared to that of a previouslypublished similar meta-analysis [66] and to two other studiesdealing with the conscious resting state [7,40], using the Euclidiandistance between corresponding foci in the Talairach space as anindex of similarity.
RESULTS
Behavioral Observations
Among the 63 subjects that were included in the study, 41properly answered the questionnaire dealing with their RESTcondition mental activity. Overall, a majority of subjects (56%)reported that mental events were partly dealing with autobio-graphic reminiscences, either recent or ancient, consisting of fa-miliar faces, scenes, dialogs, stories, melodies, etc.
Inner speech and mental images were the most frequentlyreported types of mental events during REST. Ranking the occur-rence of such events on a three-level scale (Absent, Rare, Fre-quent; Table 3), we found that mental images (respectively innerspeech) occurred in 83% (respectively 73%) of the subjects. How-ever, there was no association between the occurrences of thesetwo types of events (x2 5 4.16,p 5 0.38,df 5 4) meaning thatsubjects did not engage systematically in one type of mentalactivity during the REST condition. No systematic difference wasobserved between the groups of subjects of the various protocolsregarding the occurrence of mental events during REST, althoughthere was a much larger proportion of subjects reporting frequentmental imagery activity during REST in the group of subjects ofthe perceptual matching protocol (87%) as compared to the othergroups (23%) (p , 0.001, chi-square test). Finally, some subjectsreported discomfort or light pain in the back of their head (58%) orin their left arm (22%).
Conjunction Analysis of PET Data
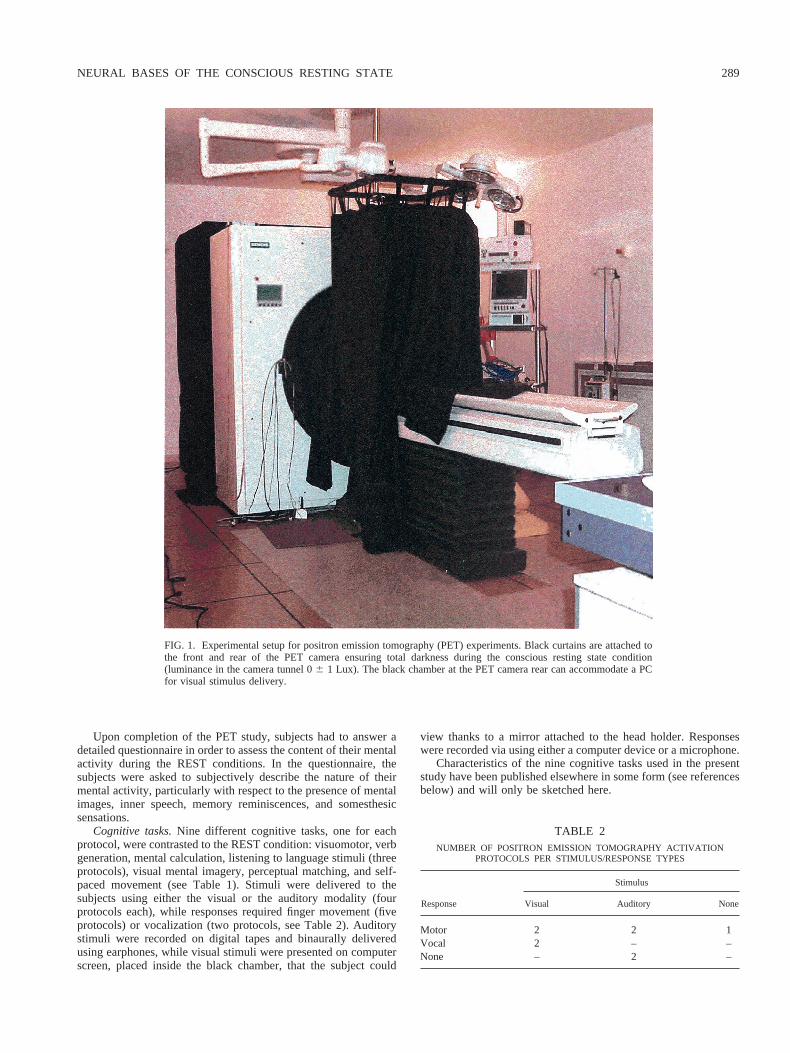
Brain areas jointly “activated” during REST as compared tothe nine cognitive tasks.As shown in Table 4 and Fig. 2, a network
TABLE 3OBSERVED OCCURRENCES OF REPORTED MENTAL IMAGERY AND
INNER SPEECH MENTAL ACTIVITIES DURING THE CONSCIOUSRESTING CONDITION IN 41 SUBJECTS
Mental Imagery
Inner Speech
None Rare Frequent
None 1 3 3 7Rare 4 7 10 21Frequent 6 4 3 13Total 11 14 16 41
x2 5 4.16,df 5 4, p 5 0.38.
290 MAZOYER ET AL.
of brain areas, mainly localized in the left hemisphere, was foundjointly activated during REST. The larger activation focus wasfound in the left posterior cingulate cortex (BA 30 and 31),extending caudally to the anterior precuneus (BA 7) and paracen-tral lobule (BA 5). In addition, bilateral activations were found atthe junction of the middle occipital gyrus and the angular gyrus(BA 19 and 39). Foci of activations were also detected in the right
post-central gyrus (BA 2), at the level of the hand/arm represen-tation.
All other foci of activation were found in the left frontal lobe,including the upper parts of the superior and middle frontal gyrus(BA 6/8), the median superior frontal (BA 10) and anterior cin-gulate (BA 32) cortex, the inferior frontal gyrus (BA 45/46), thelower parts of the superior and middle frontal gyrus (BA 10/11).
FIG. 2. Three-dimensional renderings of statistical parametric conjunction maps showing brain areas ofhigher normalized cerebral flow during conscious resting state than in any of the nine cognitive tasks includedin the protocol. Left: front view, Right: rear view. Conjunction maps were generated at a 0.001 statistical(uncorrected for multiple comparisons) for display purposes.
TABLE 4ACTIVATION CLUSTERS DURING CONSCIOUS RESTING STATE* AS COMPARED TO THE NINE TASKS INCLUDED IN THE META-ANALYSIS
k Anatomical localization BA x y z Z pcorrected
18 L superior occipital/angular 19, 39 252 274 18 6.63 0.00014 L superior occipital 19 240 282 34 6.04 0.00023 R angular 39 46 268 26 6.03 0.000
R superior occipital/angular 19, 39 48 272 16 5.48 0.005149 L posterior cingulate 31 28 236 42 6.78 0.000
L paracentral lobule/anterior precuneus 5, 7 28 240 60 5.66 0.002L paracentral lobule 5 24 234 56 5.49 0.005
3 L posterior cingulate 30 26 250 26 5.17 0.02373 L superior frontal sulcus 6 220 30 58 6.01 0.000
L superior frontal 8 218 40 48 5.59 0.003L middle frontal 8 224 24 52 5.40 0.008
5 L middle frontal 8 232 22 42 5.60 0.00341 L inferior frontal (triangularis) 45, 46 246 36 18 5.86 0.00141 L median superior frontal 10 24 58 12 6.17 0.00060 L anterior cingulate 32 0 38 22 5.68 0.002
L anterior cingulate 32, 10 24 46 24 5.37 0.0092 L anterior cingulate 32 24 50 26 5.08 0.0354 L middle frontal sulcus 10 246 48 0 5.09 0.0335 L orbital superior frontal 11 226 36 210 5.50 0.0051 L superior frontal 10 218 64 10 5.06 0.038
73 R postcentral 2 24 234 64 6.61 0.000R postcentral 5 32 240 60 5.29 0.013
12 R postcentral 2 44 224 34 5.55 0.0041 R rolando sulcus 4 24 222 66 5.14 0.027
* p # 0.05 corrected for multiple comparisons.L, left; R, right; BA, Brodmann’s area; x, y, z stereotactic coordinates;k 5 activation cluster size in voxels.
NEURAL BASES OF THE CONSCIOUS RESTING STATE 291
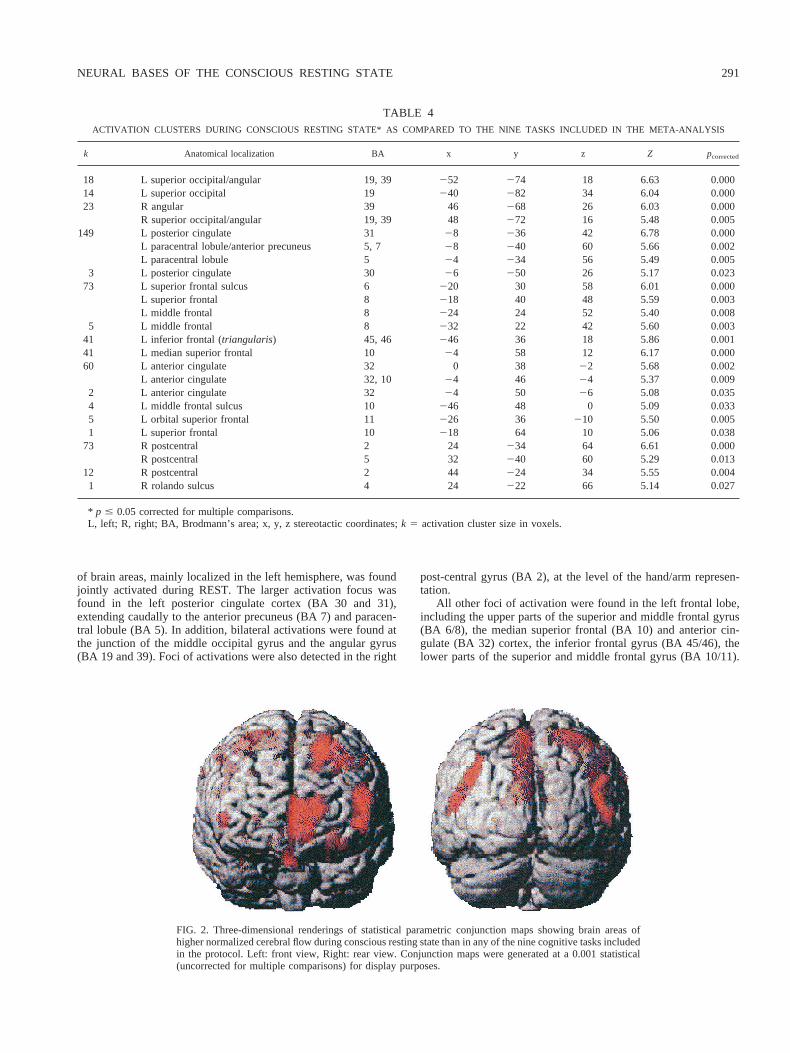
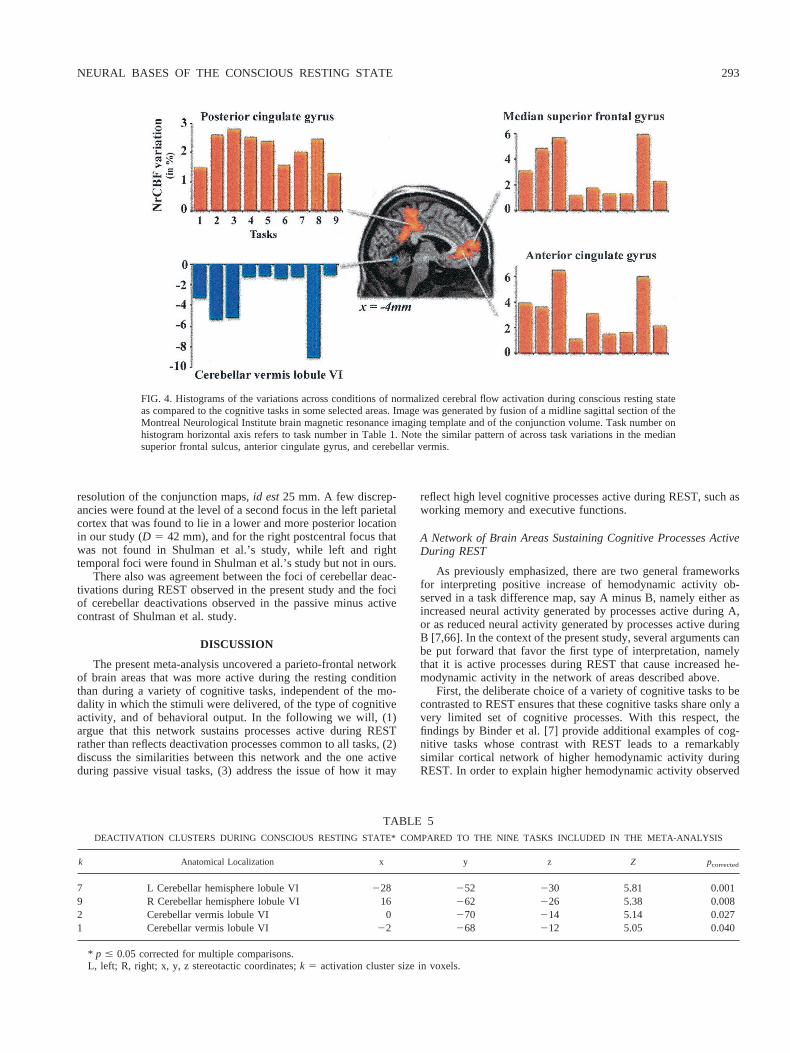
Figures 3 and 4 demonstrates the consistency of these activationsacross the different protocols, although in two areas, namely themedian superior frontal and anterior cingulate gyrus, four tasksinduced more variations as compared to REST than the five others.Interestingly, these four tasks share visual stimulus presentation asa common feature.
Brain areas jointly “deactivated” during REST as compared tothe nine cognitive tasks.As shown in Table 5 and Fig. 4, we foundonly a restricted set of brain areas with lower NrCBF duringREST, all located in the lobule VI (according to [62]) of bothcerebellum hemispheres and the vermis. Note again on Fig. 4 thatthe amplitude of cerebellar deactivation during REST varied be-tween protocols, the largest ones being observed during the samefour tasks involving visual stimulus presentation.
Comparison with other studies dealing with the consciousresting state.As shown in Table 6, a good agreement was observedwith the results reported in the FMRI study of REST by Binder etal. [7]. Congruent foci of activation were found at the level of theposterior cingulate cortex, angular gyrus, left anterior cingulate,dorsolateral prefrontal, and orbitofrontal cortex. Most discrepan-cies came from the absence of detection in the FMRI study of focireported in our study at the level of both the median frontal cortex,the inferior frontal sulcus, and right rolandic cortex. In addition,
angular gyrus activations were found more symmetrical in thepresent study than in the FMRI study, while the parahippocampalfocus found in the Binder et al. study did not have a counterpart inours. Finally, there was no report of hemodynamic variations in thecerebellum during REST in the FMRI study, which could beexplained by the limited axial field of view of the FMRI acquisi-tion.
Laurey et al.’s results were also in remarkable agreement withours at the level of the posterior cingulate, left occipito-parietal,and left dorsolateral prefrontal [40]. However, additional increasedglucose metabolism during REST as compared to the vegetativestate were also described in the right prefrontal cortex, and bilat-erally in the temporal cortex. It is worth noticing that the RESTactivation conjunction map obtained at a less stringent threshold(0.001 uncorrected) shows a right dorsolateral prefrontal focusconsistent with that of Laurey et al. study (see Fig. 2).
Comparison with Shulman et al. meta-analysis.Table 7 sum-marizes the comparison between our meta-analysis and that ofShulman et al. Overall, a good agreement was observed in terms ofnumber and location of activation foci found either during REST(present study) or visual passive tasks [66]. Posterior cingulate, leftand right occipito-parietal, left frontal, anterior cingulate, andcerebellar foci found in the two studies were within twice the
FIG. 3. Histograms of the variations across conditions of normalized cerebral flow activation duringconscious resting state as compared to the cognitive tasks in some selected areas. Images were generatedby fusion of axial sections of the Montreal Neurological Institute brain magnetic resonance imagingtemplate and of the conjunction volume at the same level. Task number on histogram horizontal axis refersto task number in Table 1.
292 MAZOYER ET AL.
resolution of the conjunction maps,id est25 mm. A few discrep-ancies were found at the level of a second focus in the left parietalcortex that was found to lie in a lower and more posterior locationin our study (D 5 42 mm), and for the right postcentral focus thatwas not found in Shulman et al.’s study, while left and righttemporal foci were found in Shulman et al.’s study but not in ours.
There also was agreement between the foci of cerebellar deac-tivations during REST observed in the present study and the fociof cerebellar deactivations observed in the passive minus activecontrast of Shulman et al. study.
DISCUSSION
The present meta-analysis uncovered a parieto-frontal networkof brain areas that was more active during the resting conditionthan during a variety of cognitive tasks, independent of the mo-dality in which the stimuli were delivered, of the type of cognitiveactivity, and of behavioral output. In the following we will, (1)argue that this network sustains processes active during RESTrather than reflects deactivation processes common to all tasks, (2)discuss the similarities between this network and the one activeduring passive visual tasks, (3) address the issue of how it may
reflect high level cognitive processes active during REST, such asworking memory and executive functions.
A Network of Brain Areas Sustaining Cognitive Processes ActiveDuring REST
As previously emphasized, there are two general frameworksfor interpreting positive increase of hemodynamic activity ob-served in a task difference map, say A minus B, namely either asincreased neural activity generated by processes active during A,or as reduced neural activity generated by processes active duringB [7,66]. In the context of the present study, several arguments canbe put forward that favor the first type of interpretation, namelythat it is active processes during REST that cause increased he-modynamic activity in the network of areas described above.
First, the deliberate choice of a variety of cognitive tasks to becontrasted to REST ensures that these cognitive tasks share only avery limited set of cognitive processes. With this respect, thefindings by Binder et al. [7] provide additional examples of cog-nitive tasks whose contrast with REST leads to a remarkablysimilar cortical network of higher hemodynamic activity duringREST. In order to explain higher hemodynamic activity observed
FIG. 4. Histograms of the variations across conditions of normalized cerebral flow activation during conscious resting stateas compared to the cognitive tasks in some selected areas. Image was generated by fusion of a midline sagittal section of theMontreal Neurological Institute brain magnetic resonance imaging template and of the conjunction volume. Task number onhistogram horizontal axis refers to task number in Table 1. Note the similar pattern of across task variations in the mediansuperior frontal sulcus, anterior cingulate gyrus, and cerebellar vermis.
TABLE 5DEACTIVATION CLUSTERS DURING CONSCIOUS RESTING STATE* COMPARED TO THE NINE TASKS INCLUDED IN THE META-ANALYSIS
k Anatomical Localization x y z Z pcorrected
7 L Cerebellar hemisphere lobule VI 228 252 230 5.81 0.0019 R Cerebellar hemisphere lobule VI 16 262 226 5.38 0.0082 Cerebellar vermis lobule VI 0 270 214 5.14 0.0271 Cerebellar vermis lobule VI 22 268 212 5.05 0.040
* p # 0.05 corrected for multiple comparisons.L, left; R, right; x, y, z stereotactic coordinates;k 5 activation cluster size in voxels.
NEURAL BASES OF THE CONSCIOUS RESTING STATE 293
in a conjunction of REST minus task contrast, a cognitive processcommon to all tasks but REST should generate decreased neuralactivity during the task. Transmodal inhibition due to focal atten-tion could be one such process and there have been multiplereports in the literature of deactivations in auditory or somato-sensory areas during visual tasks [28,53,64], or in the associativevisual cortex during somato-sensory tasks [33]. However,transmodal inhibition effects seem to be restricted to primarycortices and thus cannot be at the origin of the network of jointactivations observed during REST because, with the exception ofthe right rolandic area, this network includes no primary or uni-modal associative areas. Regarding the right rolandic area activa-tion during REST, one can hypothetize that subjects were likely topay more attention to external somato-sensory inputs during RESTthan during stimulus-response driven tasks. Recalling that all oursubjects had a catheter for water infusion in their left arm, one canreasonably postulate that increased rCBF during REST in the rightpostcentral region could be due to a shift of attention away fromthis somato-sensory input. However, one could also argue that it isan active attentional process during REST that causes this rCBFincrease, as it has also been shown that focal attention on periph-eral somatosensory stimulation enhances activity in the corre-sponding cortex [48].
Second, use of a conjunction analysis to pool activation mapsacross protocols guarantees that when increased, rCBF is increasedin all REST minus task comparisons, whereas averaging mapsacross protocols, as was performed in a similar meta-analysis [66],could reveal activation during REST that would not necessarily becommon to all protocols but rather driven by the strongest ones,
with possibly reduced activity during REST in some others (see,e.g., Figs. 2–5 of [66]).
Third, the network active at REST observed in the presentstudy remarkably overlaps the network of areas of reduced metab-olism during the vegetative state (VS, [40]) as compared to aconscious resting condition in normal volunteers. Considering thatVS is by definition a disorder of consciousness, that affects bothexternal and self awareness, this finding strongly supports theproposal that the network active at REST found in the presentstudy closely reflects cognitive process at work during REST, suchas the free generation, association, and monitoring of consciousthoughts. In the same vein, a recent study on propofol induced lossof consciousness in humans demonstrated that it was associatedwith reduction of neural activity in a subset of areas belonging tothe network active at REST of the present study, namely thebilateral parieto-occipital junction, precuneus, posterior cingulate,and left orbitofrontal cortex [20].
A Common Network of Brains Areas Active During REST andPassive Tasks
We observed striking similarities between the network of brainareas of higher blood flow during REST of the present study andthe network of areas reported to be activated during passive visualconditions as compared to active ones [66], a finding which hasbeen also reported by others [7]. Indeed, despite several method-ological differences between the two meta-analyses (sample size,spatial normalization and statistical procedure) the only majormismatch between the two networks concerned temporal cortexfoci found in the meta-analysis of passive minus active visual
TABLE 6COMPARISON OF ACTIVATION FOCI OBSERVED DURING CONSCIOUS RESTING STATE (REST) IN THE PRESENT STUDY WITH: (1) FOCI OF
DECREASED METABOLISM DURING VEGETATIVE STATE AS COMPARED TO REST IN HEALTHY CONTROLS (LEFT COLUMNS [40], (2) FOCI OFFUNCTIONAL MAGNETIC RESONANCE IMAGING SIGNAL ACTIVATION DURING REST AS COMPARED TO TONE LISTENING (RIGHT COLUMNS, [7])
Laurey et al. Study D (mm) Present Study D (mm) Binder et al. Study
L31 22, 240, 36 8.0 L31/7 28, 236, 42L31 26, 250, 26 6.5 L23/31 29, 255, 24
5.2 R23/31 9,253, 23L39/19 250, 262, 30 17.0 L39 252, 274, 18 12.6 L39 243, 270, 26L40 264, 234, 36
R39 46,268, 26R2 24,234, 64
L2 244, 24, 42L8 236, 14, 40 9.1 L8 232, 22, 42 14.5 L8/9/10 220, 30, 44L6 232, 26, 56 9.1 L8 224, 24, 52L9 216, 56, 26 18.5 L10 24, 58, 12L/47 256, 34, 0 17.2 L10 246, 48, 0
L32 0, 38,22 11.0 L12/32 29, 34,27L10 228, 56, 20 23.7 L46 246, 36, 18 11.0 L45 251, 26, 14
L11 226, 36,210 8.2 L11 226, 28,28R8 14, 38, 52R46 46, 32, 30R10 30, 52, 24R6 42, 12, 54L21 266, 244, 26L37 262, 260, 28R22 48,256, 16
L36 226, 235, 212
Foci are identified by their stereotactic coordinates and Brodmann area number according to the Talairach atlas. Cells in grey indicate mismatch betweenthe studies.D, euclidian distance between corresponding foci in the Talairach space, L, left; R, right.
294 MAZOYER ET AL.
conditions. This discrepancy could be due to enhanced visualattention during active visual tasks generating transmodal inhibi-tion in the auditory cortex [53].
In their discussion, Shulman et al. [66] could not settle betweenalternative interpretations of their results, namely whether higherblood flow during passive visual tasks as compared to active visualtasks were due to active processes during the passive tasks or tocommon inhibitory processes during the active tasks. The similar-ities between the passive visual conditions and REST networksfavors the first interpretation and leads to postulate that this net-work sustains cognitive processes that are active during both thepassive visual tasks and REST. Among possible candidates, Shul-man et al. proposed that unconstrained verbal thought, monitoringof the external environment, body image, and emotional statecould be at work during passive visual conditions: all these pro-cesses are likely to be also at work during the REST conditions asimplemented both in the present study and in the Binder et al.FMRI study [7].
A Network Sustaining Working Memory and ExecutiveFunctions During REST
Actually, based on the introspective questionnaires of our sub-jects regarding the content of their mind during REST, one mustadmit that processes at work during REST go well beyond uncon-strained verbal thoughts, but also include generation and manipu-lation of mental images, reminiscence of past experiences based onepisodic memory, and making plans. Moreover, in order to followthe instructions of the REST condition (see Materials and Methodssection), subjects had to recruit inhibitory process in order both torefrain from moving and to avoid structured mental activities,especially those related to the cognitive task of the protocol theyparticipated in. All these cognitive processes are components ofworking memory and executive systems, known to be sustained byhigh-order associative heteromodal and paralimbic areas [25,46].
As a matter of fact, the network active during REST as well asduring passive visual conditions exclusively includes such corti-ces. The high-order nature of these areas led previous authors toconsider “conceptual processing” as the main cognitive process atwork during REST, and more specifically semantic processing [7].This interpretation was based on the observation that, when com-pared to the same tone matching task, REST and a semantic taskelicited hemodynamic activity increases of similar amplitudes in acommon network of brain areas. However, the present study showsthat hemodynamic activity in this network is more active duringREST than during several tasks included in the conjunction anal-ysis that did require semantic processing. This apparent discrep-ancy between the two studies could be due to a lack of sensitivityof the FMRI study, which could be due either to a limited samplesize or to a threshold effect, and led us to propose an alternativeinterpretation of our findings. Before proceeding to this, and es-pecially to a tentative segregation of the network activated atREST in separate sub-networks sustaining different cognitivecomponents, it seems worthwhile to emphasize that such an inter-pretation will be largely putative both because of the ill controllednature of the REST state and of the observational nature of theexperimental design. However, the large corpus of cognitive neu-roimaging data that is available today allows one to use inductivereasoning for observational data interpretation.
Within this framework, it is noteworthy that retrosplenial areas,namely the posterior cingulate cortex and precuneus, are known tobe involved in episodic memory [70] (for review see [13,59]), acognitive function that has psychological commonality with RESTas emphasized by others [3]. Retrosplenial activations have indeedbeen reported in a large variety of episodic memory tasks, wouldthey be based on explicit visual material [22,58], requesting theretrieval of material including a visual memory component [3,21,50,54,69], based on verbal material [1,2,4,9,27,32,68], or irrespec-tive of the imagery content of the material to be retrieved [39].
TABLE 7COMPARISON OF ACTIVATION AND DEACTIVATION FOCI OBSERVED IN SHULMAN’S METANALYSIS [66] AND IN THE PRESENT STUDY
Shulman et al. [66] Study D (mm) Present Study
Activation foci during control state1. L31 25, 249, 40 13.5 L31/7 28, 236, 422. L40 253, 239, 42 42.0 L39 252, 274, 183. L39/19 245, 267, 36 15.9 L19 240, 282, 344. R40 45,257, 34 13.6 R39 46,268, 26
R2 24,234, 645. L8 227, 27, 40 7.3 L8 232, 22, 426. L8/9 211, 41, 42 9.2 L8 218, 40, 487. R8/9 5, 49, 368. L9 215, 55, 26 18.0 L10 24, 58, 129. L10 219, 57, 8 15.5 L10 24, 58, 12
10. L32/10 21, 47,24 3.1 L32/10 24, 246, 2411. L10/47 233, 45,26 14.6 L10 246, 48, 0
19.8 L46 246, 36, 1812. L32 3, 31,210 11.0 L32 0, 38,2213. L20 249, 219, 21814. R34/28 21,29, 218Deactivations foci during control state (cerebellum)
L lobule VI 223, 261, 214 19.0 L lobule VI 228, 252, 230Vermis lobule VI 23, 269, 28 6.7 Vermis lobule VI 0,270, 214R lobule VI 33,263, 218 18.8 R lobule VI 16,262, 226
Foci are identified by their stereotactic coordinates and Brodmann area number according to the Talairach atlas. Cells in grey indicate mismatch betweenthe 2 studies.D, euclidian distance in the Talairach space between corresponding foci; L, left; R, right.
NEURAL BASES OF THE CONSCIOUS RESTING STATE 295

Actually, the analysis of the REST questionnaires shows that thesubjects of the present study preferentially reported autobiographicepisodes. With this respect, it is worth noticing that the recall ofautobiographic episodic memory, that includes a greater emotionalcontent than non-autobiographic retrieval, has been reported toinduce larger activation of the posterior cingulate cortex [19]. Inaddition, a recent review of the neuroimaging literature hasstressed the involvement of the retrosplenial regions in emotionalprocessing and provided strong arguments for a specific role of theposterior cingulate (BA 30) as a mediator of the interaction be-tween emotion and episodic memory [41]. The hypothesis that theposterior cingulate is implicated in salient emotion monitoring isfurther supported by the fact that other regions dealing in emo-tional processing are activated during REST, in particular theorbital frontal cortex.
Regarding the precuneus, one should emphasize that it is itsmost anterior part that was found activated during REST. Anantero-posterior functional segregation within the precuneus hasalready been suggested by Buckner et al. [10], who reporteddeactivation of its anterior portion during various cognitive taskswhen compared to REST or passive conditions, whereas the pos-terior precuneus had been repeatedly reported as involved inconscious effortful recollection of episodes [10]. The present studyconfirms the activation during REST of the anterior precuneus and,given its anatomical and functional proximity with the posteriorprecuneus and posterior cingulate, leads us to postulate that itcould be part of a network specifically involved in the free willedrecall, as opposed to constraint or effortful recall, of episodicmemory items present during REST.
In the same vein, activations located at the junction between theangular and the middle occipital gyri have been described, to ourknowledge, only during the REST condition as compared to pic-ture [10] and verbal [2] recall tasks. These cortical areas, which aredifferent but in the vicinity of occipital areas involved duringvisual mental imagery [43,44,58], are singular in that they do notbelong to the unimodal associative cortex, such as those involvedin visual mental imagery, but rather to the high-order heteromodalassociative cortex [47,51]. Together with the anterior precuneus,they appear to constitute a network specific for the recall andmaintenance of multimodal thoughts, such as mental images, innerspeech, etc., from episodic and long term memory through freeassociation, under the control of the prefrontal network of regionsevidenced in the present study.
As a matter of fact, working memory load from long termmemory and manipulation of thoughts are known to be sustainedby hierarchically organized and reverberating fronto-parietal re-ciprocal connections [25]. During REST, we observed a highhierarchical level network constituted with left prefrontal, bilateralangular gyri, and retrosplenial regions. The claim that this set ofareas does constitute a network is supported by neuroanatomicalstudies in the rhesus monkey that have shown long fibers projec-tion from the prefrontal cortex to the posterior cingulate cortex, aswell as reciprocal connections through the occipito-frontal fascic-ulus between the parieto-occipital junction (corresponding to theangular gyrus in humans) and the prefrontal cortices [51]. Withinthis large scale cortical network, heteromodal dorsolateral andparalimbic medial prefrontal cortices co-activation corresponds tothe integration of mental events and to the monitoring of emotionsand motivational resources [46], the orbital frontal region being, inparticular, known to be devoted to emotional and motivationalprocessing [5,6].
Meanwhile, recent data from neuroimaging studies have dem-onstrated that, although it seems difficult to dissociate the workingmemory from the manipulation of data in working memory [11],the left middle and inferior frontal gyrus are in charge of manip-
ulation and inhibition of material held on line [15,16,56]. Inparticular, the left inferior frontal gyrus, together with the anteriorcingulate, has been reported to be specifically implicated in theinhibitory control of interference during go-no go tasks [36,37,49],working memory tasks [17,31] and reasoning tasks[29]. The im-plication of an inhibitory control network is related, as statedabove, to the inhibition of movements, and of mental activitiesrelated to the cognitive task of the protocol they participated in, asit was explicitly stated by the REST condition instructions.
Finally, the leftward asymmetry of prefrontal activations foundduring REST can be brought together with a recent model forepisodic memory retrieval, the “generate-recognize” model, as-signing specialization for recall to the left prefrontal cortex (BA45, 46), and for recognition to its right hemisphere homologue[12,60]. This model is consistent with the results of the presentstudy, because, in absence of external stimuli, the generationprocess dominates the episodic memory recollection during REST.
Joint Deactivations During REST
A final point of discussion on the results of the present studyconcerns the joint deactivations observed in both cerebellar hemi-spheres. Considering the variety of tasks used in our meta-analysisand the absence of cerebellar hypometabolism during both thevegetative state [40] and propofol induced loss of consciousness[20], the most likely explanation for joint cerebellar deactivationsin REST minus task contrasts is that they reflect common pro-cesses active during the tasks.
Based on the results of a recent study, a first candidate could beinternal timing because it appears to be associated with cerebellaractivations at locations very close to that of the present study [34].However, although one may consider that internal timing is anintrinsic component of cognitive activity, explicit instruction oftiming was not given to the subjects but in the finger movementtask.
Another candidate could be the executive component proper toeach task because it has been demonstrated that cerebellar hemi-spheres are concerned with executive language, visuospatial, ormnemonic functions [61]. In particular, the protocols showing thelarger activations during the tasks as compared to REST are thosecontaining a major memory component (see Fig. 4): recall for theverb generation and the arithmetical facts tasks, maintenance of therule in working memory for the visuomotor and perceptual match-ing tasks.
However, it should be underlined that these protocols also sharevisual stimulus presentation as a common feature that requiresboth coordination of the voluntary eye movements and spatialattention, two processes that are under the control of a networkincluding the lateral zone of the cerebellum to which belongs thelobule VI [35].
CONCLUSION
The conscious resting state in humans is sustained by a largescale network of heteromodal associative parietal and frontal cor-tical areas, that can be further hierarchically organized in anepisodic working memory parieto-frontal network, driven in partby emotions, working under the supervision of an executive pre-frontal network. The bilateral angular gyrus and the anterior pre-cuneus, parts of the parieto-frontal sub-network, appears to bespecifically involved during REST and may reflect the recall andmaintenance of multimodal thoughts through free associationwhich characterizes this mental state.
296 MAZOYER ET AL.

ACKNOWLEDGEMENTS
The authors are indebted to the Cyceron Cyclotron (P. Lochon, O.Tirel) and PET camera staff (V. Beaudoin, G. Perchey) for their invaluablehelp in acquiring the PET activation studies. The authors would also liketo thank A. Mazard for her help in some of data acquisition. Parts of thiswork have been supported by grants from the GIS Sciences de la Cogni-tion.
This work has been presented in part at the 5th International Confer-ence on Functional Mapping of the Human Brain (Dusseldorf, Germany,June 1999).
REFERENCES
1. Andreasen, N. C.; O’Leary, D. S.; Arndt, S.; Cizadlo, T.; Hurtig, R.;Rezai, K.; Watkins, G. L.; Pontos, L. L. B.; Hichwa, R. Short-term andlong-term verbal memory: A positron emission tomography study.Proc. Natl. Acad. Sci. USA 92:5111–5115; 1995.
2. Andreasen, N. C.; O’Leary, D.; Arndt, S.; Cizadlo, T.; Rezai, K.;Watkins, G. L.; Boles Ponto, L. L.; Hichwa, R. PET studies ofmemory: Novel and practised free recall of complex narratives. Neu-roimage 2:284–295; 1995.
3. Andreasen, N. C.; O’Leary, D. S.; Cizadlo, T.; Arndt, S.; Rezai, K.;Watkins, G. L.; Boles Ponto, L. L.; Hichwa, R. D. Remembering thepast: Two facets of episodic memory explored with positron emissiontomography. Am. J. Psychiatry 152:1576–1585; 1995.
4. Andreasen, N. C.; O’Leary, D. S.; Cizadlo, T.; Arndt, S.; Rezai;Watkins, G. L.; Boles Ponto, L. L.; Hichwa, R. D. II. PET studies ofmemory: Novel versus practiced free recall of word lists. Neuroimage2:296–305; 1995.
5. Bechara, A.; Damasio, H.; Damasio, A. R. Emotion, decision makingand the orbitofrontal cortex. Cereb. Cortex 10:295–307; 2000.
6. Bechara, A.; Damasio, H.; Tranel, D.; Anderson, S. W. Dissociation ofworking memory from decision making within the human prefrontalcortex. J. Neurosci. 18:428–437; 1998.
7. Binder, J. R.; Frost, J. A.; Hammeke, T. A.; Bellgowan, P. S.; Rao,S. M.; Cox, R. W. Conceptual processing during the conscious restingstate: A functional MRI study. J. Cogn. Neurosci. 11:80–93; 1999.
8. Bricogne, S.; Mellet, E.; Mazoyer, N.; Zago, L.; Beaudoin, V.;Perchey, G.; Tirel, O.; Denis, M.; Mazoyer, B. Mental navigationwithin an environment learned from reading. Neuroimage 9:S372;1999.
9. Buckner, R. L.; Petersen, S. E.; Ojemann, G. J.; Miezin, F. M.; Squire,L. R.; Raichle, M. E. Functional anatomical studies of explicit andimplicit memory retrieval tasks. J. Neurosci. 15:12–29; 1995.
10. Buckner, R. L.; Raichle, M. E.; Miezin, F. M.; Petersen, S. E. Func-tional anatomic studies of memory retrieval for auditory words andvisual pictures. J. Neurol. Sci. 16:6219–6235; 1996.
11. Bunge, S. A.; Klingberg, T.; Jacobsen, R. B.; Gabrieli, J. D. E. Aresource model of the neural basis of executive working memory.Proc. Natl. Acad. Sci. USA 97:3573–3578; 2000.
12. Cabeza, R.; Anderson, N. D.; Kester, J.; Lennartsson, E. R.; McIntosh,A. R. Involvement of prefrontal regions on episodic retrieval: Evi-dence for a generate-recognize asymmetry model. Brain Cogn.; inpress.
13. Cabeza, R.; Nyberg, L. Imaging cognition II: An empirical review of275 PET and fMRI studies. J. Cogn. Neurosci. 12:1–47; 2000.
14. Cameron, O. G.; Modell, J. G.; Hichwa, R. D.; Agranoff, B. W.;Koeppe, R. A. Changes in sensory-cognitive input: Effects on cerebralblood flow. J. Cereb. Blood Flow Metab. 10:38–42; 1990.
15. Clark, C. R.; Egan, G. F.; McFarlane, A. C.; Morris, P.; Weber, D.;Sonkkilla, C.; Marcina, J.; Tochon-Danguy, H. J. Updating workingmemory for words: A PET activation study. Hum. Brain Mapp. 9:42–54; 2000.
16. Collette, F.; Salmon, E.; VanderLinden, M.; Chicherio, C.; Belleville,S.; Degueldre, C.; Delfiore, G.; Franck, G. Regional brain activityduring tasks devoted to the central executive of working memory.Cogn. Brain Res. 7:411–417; 1999.
17. D’Esposito, M.; Postle, B. R.; Jonies, J.; Smith, E. E. The neuralsubstrate and temporal dynamics of interference effects in workingmemory as revealed by event-related functional MRI. Proc. Natl.Acad. Sci. USA 96:7514–7519; 1999.
18. Etard, O.; Mellet, E.; Papathanassiou, D.; Benali, K.; Houde´, O.;Mazoyer, B.; Tzourio-Mazoyer, N. Picture naming without Broca andWernicke’s area. Neuroreport 11:617–621; 1999.
19. Fink, G. R.; Markowitsch, H. J.; Reinkemeier, M.; Bruckbauer, T.;Kessler, J.; Heiss, W.-D. Cerebral representation of one’s own past:Neural networks involved in autobiographical memory. J. Neurosci.16:4275–4282; 1996.
20. Fiset, P.; Paus, T.; Daloze, T.; Plourde, G.; Meuret, P.; Bonhomme, V.;Hajj-Ali, N.; Backman, S. B.; Evans, A. C. Brain mechanisms ofpropofol-induced loss of consciousness in humans: A positron emis-sion tomographic study. J. Neurosci. 19:5506–5513; 1999.
21. Fletcher, P. C.; Frith, C. D.; Baker, S. C.; Shallice, T.; Frackowiak,R. S. J.; Dolan, R. J. The mind’eye—Precuneus activation in memory-related imagery. Neuroimage 2:195–200; 1995.
22. Fletcher, P. C.; Shallice, T.; Frith, C. D.; Frackowiak, R. S. J.; Dolan,R. J. Brain activity during memory retrieval. The influence of imageryand semantic cueing. Brain 119:1587–1596; 1996.
23. Friston, K. J.; Ashburner, J.; Frith, C. D.; Poline, J.-B.; Heather, J. D.;Frackowiak, R. S. J. Spatial registration and normalization of images.Hum. Brain Mapp. 2:165–189; 1995.
24. Friston, K. J.; Price, C. J.; Fletcher, P.; Moore, C.; Frackowiak,R. S. J.; Dolan, R. J. The trouble with cognitive subtraction. Neuro-image 4:97–104; 1996.
25. Fuster, J. M. Linkage at the top. Neuron 21:1223–1224; 1998.26. Ghaem, O.; Mellet, E.; Crivello, F.; Tzourio, N.; Mazoyer, B.;
Berthoz, A.; Denis, M. Mental navigation along memorized routesactivates the hippocampus, precuneus, and insula. Neuroreport 8:739–744; 1997.
27. Grasby, P. M.; Frith, C. D.; Friston, K. J.; Bench, C.; Frackowiak,R. S. J.; Dolan, R. J. Functional mapping of brain areas implicated inauditory-verbal memory function. Brain 116:1–20; 1993.
28. Haxby, J. V.; Horwitz, B.; Ungerleider, L. G.; Maisog, J. M.; Pietrini,P.; Grady, C. L. The functional organization of human extrastiatecortex: A PET-rCBF study of selective attention to faces and locations.J. Neurosci. 14:6336–6353; 1994.
29. Houde´, O.; Zago, L.; Mellet, E.; Moutier, S.; Pineau, A.; Mazoyer, B.;Tzourio-Mazoyer, N. Shifting from the perceptual brain to the logicalbrain: The impact of cognitive inhibition training. J. Cogn. Neurosci.12:721–728; 2000.
30. Joliot, M.; Papathanassiou, D.; Mellet, E.; Quinton, O.; Mazoyer, N.;Courtheoux, P.; Mazoyer, B. FMRI and PET of self-paced fingermovement: Comparison of intersubject stereotaxic averaged data.Neuroimage 10:430–477; 1999.
31. Jonides, J.; Smith, E. E.; Marshuetz, C.; Koeppe, R. A.; Reuter-Lorenz, P. A. Inhibition in verbal working memory revealed by brainactivation. Proc. Natl. Acad. Sci. USA 95:8410–8413; 1998.
32. Kapur, S.; Craik, F. I. M.; Tulving, E.; Wilson, A. A.; Houle, S.;Brown, G. M. Neuroanatomical correlates of encoding in episodicmemory: Levels of processing effect. Proc. Natl. Acad. Sci. USA91:2008–2011; 1994.
33. Kawashima, R.; O’Sullivan, B. T.; Roland, P. E. Positron emissiontomography studies of cross-modality inhibition in selective atten-tional tasks: Closing the “mind’s eye”. Proc. Natl. Acad. Sci. USA92:5969–5972; 1995.
34. Kawashima, R.; Okuda, J.; Umetsu, A.; Sugiura, M.; Inoue, K.; Su-zuki, K.; Tabuchi, M.; Tsukiura, T.; Narayan, S. L.; Nagasaka, T.;Yanagawa, I.; Fujii, T.; Takahashi, S.; Fukuda, H.; Yamadori, A.Human cerebellum plays an important role in memory-timed fingermovement: An fMRI study. J. Neurophysiol. 83:1079–1087; 2000.
35. Kim, Y. H.; Gitelman, D. R.; Nobre, A. C.; Parrish, T. B.; LaBar,K. S.; Mesulam, M. M. The large-scale neural network for spatialattention displays multifunctional overlap but differential asymmetry.Neuroimage 9:269–277; 1999.
36. Konishi, S.; Nakajima, K.; Uchida, I.; Kameyama, M.; Nakahara, K.;Sekihara, K.; Miyashita, Y. Transient activation of inferior prefrontalcortex during cognitive set shifting. Nat. Neurosci. 1:80–84; 1998.
37. Konishi, S.; Nakajima, K.; Uchida, I.; Sekihara, K.; Miyashita, Y.No-go dominant brain activity in human inferior prefrontal cortexrevealed by functional magnetic resonance imaging. Eur. J. Neurosci.10:1209–1213; 1998.
38. Kosslyn, S. M.; Thompson, W. L.; Kim, I. J.; Alpert, N. M. Topo-
NEURAL BASES OF THE CONSCIOUS RESTING STATE 297

graphical representations of mental images in primary visual cortex.Nature 378:496–498; 1995.
39. Krause, B. J.; Schmidt, D.; Mottaghy, F. M.; Taylor, J.; Halsband, U.;Herzog, A.; Tellmann, L.; Mu¨ller-Gartner, H. W. Episodic retrievalactivates the precuneus irrespective of the imagery content of wordpair associates. A PET study. Brain 122:255–263; 1999.
40. Laureys, S.; Goldman, S.; Phillips, C.; Van Bogaert, P.; Aerts, J.;Luxen, A.; Franck, G.; Maquet, P. Impaired effective cortical connec-tivity in vegetative state: Preliminary investigations using PET. Neu-roimage 9:377–382; 1999.
41. Maddock, R. J. The retrosplenial cortex and emotion: New insightsfrom functional neuroimaging of the human brain. TINS 22:310–316;1999.
42. Mazziotta, J. C.; Phelps, M. E.; Halgren, E. Local cerebral glucosemetabolic response to audiovisual stimulation and deprivation: Studiesin human subjects with positron CT. Hum. Neurobiol. 2:11–23; 1983.
43. Mellet, E.; Petit, L.; Mazoyer, B.; Denis, M.; Tzourio, N. Reopeningthe mental imagery debate: Lessons from functional anatomy. Neuro-image 8:129–139; 1998.
44. Mellet, E.; Tzourio, N.; Crivello, F.; Joliot, M.; Denis, M.; Mazoyer,B. Functional anatomy of spatial mental imagery generated fromverbal instructions. J. Neurosci. 16:6504–6512; 1996.
45. Mellet, E.; Tzourio, N.; Denis, M.; Mazoyer, B. Cortical anatomy ofmental imagery of concrete nouns based on their dictionnary defini-tion. Neuroreport 9:803–808; 1998.
46. Mesulam, M. M. From sensation to cognition. Brain 121:1013–1052;1998.
47. Mesulam, M. M. Principles of behavioral neurology. Philadelphia:F. A. Davis Company; 1985.
48. Meyer, E.; Ferguson, S. S. G.; Zatorre, R. J.; Alivisatos, B.; Marrett,S.; Evans, A. C.; Hakim, A. M. Attention modulates the somatosen-sory cerebral blood flow response to vibrotactile stimulation as mea-sured by positron emission tomography. Ann. Neurol. 29:440–443;1991.
49. Nobre, A. C.; Coull, J. T.; Frith, C. D.; Mesulam, M. M. Orbito-frontalcortex is activated during breaches of expectation in tasks of visualattention. Nat. Neurosci. 2:11–12; 1999.
50. Owen, A. M.; Milner, B.; Petrides, M.; Evans, A. C. Memory forobject features versus memory for object location: A positron-emissiontomography study of encoding and retrieval processes. Proc. Natl.Acad. Sci. USA 93:9212–9217; 1996.
51. Pandya, D. N.; Yeterian, E. H. Architecture and connections of corticalassociation areas. In: Peters, A.; Jones, E. D., eds. Cerebral cortex.New York: Plenum Press; 1985:3–61.
52. Papathanassiou, D.; Etard, O.; Mellet, E.; Zago, L.; Mazoyer, B.;Tzourio-Mazoyer, N. A common language network for comprehensionand production: A contribution to the definition of language epicentreswith PET. Neuroimage 11:347–357; 2000.
53. Petit, L.; Dubois, S.; Tzourio, N.; Dejardin, S.; Crivello, F.; Michel, C.;Etard, O.; Denise, P.; Roucoux, A.; Mazoyer, B. PET study of thehuman foveal fixation system. Hum. Brain Mapp. 8:28–43; 1999.
54. Petit, L.; Orssaud, C.; Tzourio, N.; Crivello, F.; Berthoz, A.; Mazoyer,B. Functional anatomy of a prelearned sequence of horizontal saccadesin humans. J. Neurosci. 16:3714–3726; 1996.
55. Phelps, M. E.; Mazziotta, J. C.; Kuhl, D. E.; Nuwer, M.; Packwood, J.;Metter, J.; Engel J. Jr. Tomographic mapping of human cerebralmetabolism: Visual stimulation and deprivation. Neurology 31:517–529; 1981.
56. Postle, B. R.; Berger, J. S.; D’Esposito, M. Functional neuroanatomi-cal double dissociation of mnemonic and executive control processescontributing to working memory performance. Proc. Natl. Acad. Sci.USA 96:12959–12964; 1999.
57. Price, C. J.; Friston, K. J. Cognitive conjunction: A new approach tobrain activation experiments. Neuroimage 5:261–270; 1997.
58. Roland, P. E.; Gulya´s, B. Visual memory, visual imagery, and visualrecognition of large field patterns by the human brain: Functionalanatomy by positron emission tomography. Cereb. Cortex 1:79–93;1995.
59. Rudge, P.; Warrington, E. K. Selective impairment of memory andvisual perception in splenial tumours. Brain 114(Pt. 1B):349–360;1991.
60. Rugg, M. D.; Fletcher, P. C.; Allan, K.; Frith, C. D.; Frackowiak, R. S.;Dolan, R. J. Neural correlates of memory retrieval during recognitionmemory and cued recall. Neuroimage 8:262–273; 1998.
61. Schmahmann, J. D. From movement to thought: Anatomic substratesof the cerebellar contribution to cognitive processing. Hum. BrainMapp. 4:174–198; 1996.
62. Schmahmann, J. D.; Doyon, J.; McDonald, D.; Holmes, C.; Lavoie,K.; Hurwitz, A. S.; Kabani, N.; Toga, A.; Evans, A.; Petrides, M.Three-dimensional MRI atlas of the human cerebellum in proportionalstereotaxic space. Neuroimage 10:233–260; 1999.
63. Shulman, G. L.; Corbetta, M.; Buckner, R. L.; Fiez, J. A.; Miezin,F. M.; Raichle, M. E.; Petersen, S. E. Common blood flow changesacross visual tasks .I. Increases in subcortical structures and cerebel-lum but not in nonvisual cortex. J. Cogn. Neurosci. 9:624–647; 1997.
64. Shulman, G. L.; Corbetta, M.; Buckner, R. L.; Raichle, M. E.; Fiez,J. A.; Miezin, F. M.; Petersen, S. E. Top-down modulation of earlysensory cortex. Cereb. Cortex 7:193–206; 1997.
65. Shulman, G. L.; Corbetta, M.; Fiez, J. A.; Buckner, R. L.; Miezin,F. M.; Raichle, M. E.; Petersen, S. E. Searching for activations thatgeneralize over tasks. Hum. Brain Mapp. 5:317–322; 1997.
66. Shulman, G. L.; Fiez, J. A.; Corbetta, M.; Buckner, R. L.; Miezin, F.;Raichle, M. E.; Petersen, S. E. Common blood flow changes acrossvisual tasks: II. Decreases in cerebral cortex. J. Cogn. Neurosci.9:648–663; 1997.
67. Talairach, J.; Tournoux, P. Co-planar stereotaxic atlas of the humanbrain 3-dimensional proportional system: An approach to cerebralimaging. Stuttgart, New York: Thieme; 1988.
68. Tulving, E.; Kapur, S.; Markowitsch, H. J.; Craik, F. I. M.; Habib, R.;Houle, S. Neuroanatomical correlates of retrieval in episodic memory:Auditory sentence recognition. Proc. Natl. Acad. Sci. USA 91:2012–2015; 1994.
69. Tulving, E.; Markowitsch, H. J.; Craik, F. I. M.; Habib, R.; Houle, S.Novelty and familiarity activations in PET studies of memory encod-ing and retrieval. Cereb. Cortex 6:71–79; 1996.
70. Valenstein, E.; Bowers, D.; Verfaellie, M.; Heilman, K. M.; Day, A.;Watson, R. T. Retrosplenial amnesia. Brain 110(Pt. 6):1631–1646;1987.
71. Zago, L.; Chavoix, C.; Mazoyer, N.; Mellet, E.; Beaudoin, V.; Lochon,P.; Eustache, F.; Mazoyer, B.; Baron, J.-C. Functional anatomy ofvisuo-spatial working memory using PET. Neuroimage 6:S931; 1999.
72. Zago, L.; Pesenti, M.; Mellet, E.; Bricogne, S.; Seron, X.; Beaudouin,V.; Lochon, P.; Mazoyer, B.; Mazoyer, N. Functional anatomy ofsimple and complex mental calculation using PET. Neuroimage 13:314–327; 2001.
298 MAZOYER ET AL.
Copyright © 2022 FDOKUMEN




















