
Can aquaculture help restore and sustain production of giant clams

Pancreatic enzymes sustain systemicinflammation after an initial endotoxinchallengeFlorian Fitzal, MD, Frank A. DeLano, MS, Corey Young, Henrique S. Rosario, MD,Wolfgang G. Junger, PhD, and Geert W. Schmid-Schonbein, PhD, La Jolla, Calif; Vienna, Austria;and Lisbon, Portugal
Background. Sepsis is accompanied by severe inflammation whose mechanism remains uncertain. Werecently demonstrated that pancreatic proteases in the ischemic intestine have the ability to generatepowerful inflammatory mediators that can be detected in the portal vein and in the general circulation.This study was designed to examine several circulatory and inflammatory indices during experimentalendotoxemia and intraintestinal pancreatic protease inhibition.Methods. Immediately after intravenous endotoxin administration, the small intestine was subjected tointraluminal lavage with and without gabexate mesilate, an inhibitor of pancreatic proteases. Shamsand rats without lavage served as controls. Hemodynamics, leukocyte (neutrophil and monocyte), andendothelial cell activation, as well as organ injury in the intestine and the cremaster muscle, wereexamined.Results. After endotoxin administration, control rats developed hypotension, tachycardia, hyperventi-lation, and leukopenia. The intestine and plasma contained mediators that activated leukocytes. Theleukocyte-endothelial interaction within the cremaster muscle microcirculation was enhanced. Endotoxinadministration resulted in elevated interleukin-6 plasma levels. Histologic evidence indicated liver andintestinal injury. In contrast, blockade of pancreatic proteases in the intestinal lumen significantlyimproved hemodynamic parameters and reduced all indices of inflammation in plasma and cell injuryin skeletal muscle microcirculation.Conclusions. Inflammatory mediators derived from the intestine by pancreatic proteases may be involvedin the prolonged inflammatory response and sustain symptoms of sepsis after endotoxin challenge.(Surgery 2003;134:446-56.)
From the Department of Bioengineering, the Whitaker Institute for Biomedical Engineering and Department ofSurgery, UCSD Medical Center, University of California–San Diego, La Jolla, Calif; Department of Surgery,University of Vienna Medical School, and Ludwig Boltzmann Institute for Experimental and ClinicalTraumatology, Vienna, Austria; and Institute of Biochemistry, Faculty of Medicine of Lisbon, Lisbon, Portugal
SYSTEMIC INFLAMMATION (also denoted as systemicinflammatory response syndrome) and sepsis re-main the leading causes of death in intensive careunits. Evidence indicates that the severity ofinflammation as measured by leukocyte (neutro-phil and monocyte) activation in the circulationmay serve as a predictor for survival,1 and that in-
This work was supported by the Max Kade Foundation, and inpart by NIH Grants HL-43026, HL-67825, GM-60475, and byFLAD 460/2000.
Accepted for publication April 12, 2003.
Reprint requests: Florian Fitzal, MD, Department of Surgery,University of Vienna Medical School, Waehringer Guertel 18-20,1090 Vienna, Austria.
� 2003, Mosby, Inc. All rights reserved.
0039-6060/2003/$30.00 + 0
doi:10.1067/S0039-6060(03)00168-5
446 SURGERY
flammatory mediators are produced during sepsis.These inflammatory mediators are detectable inplasma and exhibit a variety of activities which, inaddition to the activation of circulating leukocytes,include suppression of T lymphocyte proliferationand myocardial contraction, as well as several othercell and organ dysfunctions.2-4
The intestine appears to play a central roleduring sepsis.5,6 It generates different inflam-matory mediators,7-10 some of which have beencharacterized (tumor necrosis factor, interleukins,leukotrienes, platelet activating factor), and someof which remain unidentified.9,11
Translocation of intestinal bacteria and endo-toxin has been suggested to be responsible for theprolonged inflammation during sepsis. However,a number of attempts to relate intestinal-derivedendotoxemia to mortality rates in clinical sepsis

SurgeryVolume 134, Number 3
Fitzal et al 447
studies have failed so far.12,13 The intestine maygenerate factors that activate inflammatory cellseven in the absence of endotoxin or bacteria.14-16
Thus, we may have to reexamine our basic hypoth-esis for the pathophysiology of sepsis becauseother inflammatory mediators besides endotoxin,cytokines, or lipid-derived mediators may producecell activation and multiple organ injury.12,17
Recently we demonstrated an important role ofpancreatic digestive enzymes during multiorganfailure following intestinal ischemia. After entryinto the intestinal tissue because of ischemia-induced breakdown of the mucosal barrier, di-gestive enzymes may generate a set of powerfulinflammatory mediators in the intestinal tissue thatactivate leukocytes and endothelial cells in themicrocirculation.18,19 The mediators produced bythe digestive enzymes enhance the oxidative stress,cause myocardial depression, and lead to apoptoticcell death in the tissue parenchyma and rapidmortality.11,20,21 Inhibition of pancreatic proteasesin the lumen of the intestine significantly improvesthe hemodynamics and reduces inflammationduring intestinal ischemia/reperfusion-inducedshock.19,22,23
In endotoxin shock, however, the impact of theinflammatory mediators derived from pancreaticproteases on the cardiovascular system is unex-plored. We hypothesize that sepsis-induced hypo-perfusion of the intestine and elevated permeabilityof the mucosal epithelium may lead to transloca-tion of pancreatic enzymes from the lumen into thewall of the intestine, followed by generation offactors that activate inflammatory cells within thewall. The inflammatory mediators may then enterthe circulation and maintain and enhance inflam-mation during sepsis, even though it may have beentriggered by endotoxin. We examine here oneparticular aspect of this hypothesis. After endotoxinadministration, we study the effect of intraintestinalpancreatic protease inhibition (IPI) on centralcardiopulmonary dynamics, systemic inflamma-tion, the level of inflammatory mediators that causeleukocyte activation, and peripheral muscle paren-chymal cell death.
MATERIAL AND METHODS
Twenty-five male Wistar rats (180-220 g; CharlesRiver Breeding Laboratories, Wilmington, Mass)were maintained on a standard rat chow and waterad libitum. All experiments were reviewed andapproved by the University of California San DiegoAnimal Subjects Committee. The rats were fastedovernight. After general anesthesia (60 mg/kg
intramuscular pentobarbital), the left carotid arteryand right jugular vein were cannulated (polyethy-lene tubing, PE-50, Clay Adams, Parsippany, NJ) torecord arterial pressure and heart rate and toadminister supplemental anesthetic agent or fluidsupport (Ringer’s lactate 2 mL/h), respectively. Atracheal tube was inserted to minimize respiratorycomplications. Escherichia coli-derived endotoxin(serotype 127:B8, Sigma Chemical Co, St. Louis,Mo; 4 mg/kg) was administered intravenouslyinto the jugular vein over 5 min. Each animal wasthen observed during the subsequent 5-hourperiod.
Experimental groups. The rats were dividedrandomly into 4 groups (each with 5 rats):
1. SHAM group: sham surgery without endotoxinadministration or intestinal lavage
2. CON group: endotoxin administration withoutintestinal lavage
3. LAV group: endotoxin administration and in-testinal lavage with 450 mL of Krebs-Henseleitbuffer (see the following sections)
4. FOY group: endotoxin administration and in-testinal lavage with 450 mL of Krebs-Henseleitbuffer with 0.38 mmol/L of the proteaseinhibitor gabexate mesilate (FOY; Ono Pharma-ceutical, Tokyo, Japan).
Intestinal lavage and IPI. In the LAV and FOYgroups, a polyethylene tube (PE 250, Clay Adams)was inserted into the duodenal lumen immediatelydistal to the pylorus (fixed with silk 4/0) andanother tube was placed into the terminal ileum topermit free fluid discharge into a collection con-tainer. At the onset of endotoxin administration,the small intestinal lumen was rinsed gently (10mL/min) with 100 mL of Krebs-Henseleit buffersolution (7.7 g NaCl, 0.35 g KCl, 0.29 g CaCl2, 0.3 gMgSO4 per liter of sterile water, 388C, pH 7.6-7.8)without (LAV group) or with FOY for intestinalprotease blockade (FOY group).
Also in the LAV and FOY groups, the smallintestinal lumen was rinsed gently (10 mL/min)every 30 minutes with another 50 mL of buffer(with or without FOY, respectively). Pilot studieswith intravital microscopy showed that intestinallavage did not alter the intestinal microcirculation,either with or without FOY (unpublished data).Blood pressure dropped transiently in some ratsduring the first 5 minutes of lavage, leading toa short period (about 1 minute) of hypotension,with blood pressures of 60 mm Hg. We did notobserve an increase in blood pressure resultingfrom intestinal lavage. In preliminary studieswe examined endotoxin activity by the limulus

SurgerySeptember 2003
448 Fitzal et al
amebocyte lysate activity test (unpublished results)in the intestinal fluid before and after intestinallavage (n = 5 per group). Intestinal lavage with orwithout FOY did not influence endotoxin activity inthe intestinal fluid when compared with prelavagelevels (LAV group, prelavage mean ± SD: 140 ± 179vs postlavage 86 ± 36 EU/mL endotoxin activity;FOY group, prelavage mean ± SD: 66 ± 72 vspostlavage 41 ± 20 EU/mL endotoxin activity).
Cremaster muscle microcirculation. To exam-ine leukocyte and endothelial cell activation in theperipheral microcirculation, the cremaster musclewas exposed for intravital microscopy24 at 120minutes after endotoxin administration. The mus-cle preparation was continuously superfused withKrebs-Henseleit bicarbonate-buffered solution sat-urated with a mixture of 95% N2 and 5% CO2 at378C.
In each muscle, 1 terminal arteriole (30-50 lmdiameter), 3 capillary observation fields (20 to 30capillaries/field), and 3 postcapillary venules (20-30 lm diameter) in the central portion of themuscle were examined with a 320 objective (nu-meric aperture = 0.5) for the capillary fields andthe arterioles and venules with a 340 water-immer-sion objective (numeric aperture = 0.76). Aftera 30-minute adaptation period, the first microvascu-lar bright field and fluorescent images were re-corded with a color CCD camera (model VI470,Optronics, Goleta, Calif) and a videotape recorder(AG6300, Panasonic; Matsushita Electric IndCo, Japan) for offline data analysis. Propidiumiodide (Sigma) was added to the cremaster musclesuperfusate (1 lmol/L final concentration) to labelthe nuclei of dead cells.25 At each time point, theimages of the microvessels were recorded for 1minute; otherwise, the tissue was not illuminated tominimize tissue injury because of the exposure offluorescent light. Images of the muscle microcircu-lation were recorded 180, 240, and 300 minutesafter endotoxin administration.
Microvascular measurements. Red blood cellvelocity (VRBC) was measured online by the cross-correlation technique (Vista Electronics Company,Ramona, Calif). Vessel diameters and lengths as wellas the number of rolling and adherent leukocytes inthe pre-capillary arterioles and in postcapillaryvenules at each time point were determined.Leukocytes that were rolling continuously alongthe vessel wall were counted per unit time.Leukocytes attached firmly to the endothelium fora period of 30 seconds or longer were referred to asadherent cells.
Leukocyte rolling velocity (VWBC) on the endo-thelium in postcapillary venules was measured off-
line from the displacement of individual leukocytesfrom image frame to frame (time duration for 1video frame = 1/30 sec). The leukocyte adhesionindex (LAI) was calculated from the ratio ofVWBC and VRBC in the same postcapillary venulesegment measured during the same time interval.LAI falls between 0 and 1. A decline in LAI resultsfrom an increase in leukocyte-endothelial cellmembrane attachment.26 In each microvascu-lar field, functional capillary density was deter-mined as the fraction of perfused capillaries andthe number of all (perfused and nonperfused)capillaries.
Tissue samples. Before intestinal lavage and atthe end of the experiment, intestinal fluid was col-lected from the ileal tube and centrifuged at 500 gfor 5 minutes. Aliquots were stored (�708C) untiluse for measurement of protease activity (see thefollowing section). One aliquot of blood (1 mLcontaining 10 U/mL heparin) was collected beforeendotoxin administration, and 1 aliquot at the endof the experiment. The samples were centrifuged(500g for 5minutes) and theplasma stored (�708C)for measurement of leukocyte activation and IL-6levels (see the following section). An additional 0.1mL of heparinized blood was used to determine theleukocyte counts in peripheral blood (UnopetteMicrocollection System, Bekton, NJ).
After euthanasia (5 hours after endotoxinadministration), a segment of the ileum (5 cm)was excised, rinsed in phosphate buffered saline(PBS, 10 mmol/L) and fixed in 3% formalin (24hours). The intestinal segments were embedded inresin, sectioned (1 lm section thickness), stainedwith toluidine blue, and assessed by light micros-copy for morphologic damage (no quantification, 3sections per animal).
Another segment of the terminal ileum (1 g) wasexcised, and all intestinal contents were rinsed. Thesegment was placed into 3 mL of a PBS solution,homogenized, and centrifuged at 1000 g for 10minutes. The supernatants from the homogenatewerecollectedandstored(�708C) for latermeasure-ments of leukocyte activation as pseudopod for-mation (see the following section).
Protease activity assay. Total protease activity inintestinal fluid was measured with a protease assaykit (EnzChek, Molecular Probes, Eugene, Ore).The protease activities detected by this kit includedelastase, trypsin, chymotrypsin and pepsin. Thefluorescence intensity of the kit mixed with thesamples was measured with a fluorometer andenzyme activity was expressed in fluorescence units(FU) with a lower detection limit between 1.0 3
10�3 and 4.4 3 10�5 FU.

SurgeryVolume 134, Number 3
Fitzal et al 449
Leukocyte activation. Supernatant samples ofthe intestinal homogenates and plasma sampleswere mixed with naive human leukocytes (fromasymptomatic volunteers). The inflammatory tri-peptide F-Met-Leu-Phe (10�7 mol/L) and bufferserved as positive and negative controls, respec-tively (not shown). Exposure of the leukocytes leadsto general activation (eg, superoxide formation,degranulation, expression of adhesion molecules,polymerization of actin with pseudopod formation,and mechanical stiffening of the cytoplasm).Pseudopod formation leads to leukocyte entrap-ment in the microcirculation and migration intoextravascular tissue. Thus, as an index of leukocyteactivation, we determined the fraction of cells withpseudopod formation as described earlier.21 Onehundred leukocytes (neutrophils and monocytes)were microscopically examined (1003 objective,numerical aperture 1.4, 103 objective) and cellswith cytoplasmic projections greater than 1 lmwere classified as activated leukocytes.
IL-6 measurements. IL-6 levels of predilutedplasma samples (1:100) were measured with a com-mercial ELISA kit (Quantikine M ELISA; R&DSystems Inc., Minneapolis, Minn). Samples weremeasured in duplicates and values were expressedas ng/mL.
IV administration of gabexate mesylate. Duringintestinal lavage, FOY may be partially absorbedinto the circulation, where it can reduce cytokinelevels,27 leukocyte activation,28 and oxygen-freeradical formation,29 and therefore may influenceparameters of septic shock. To investigate whetherintestinal lavage with FOY affects the hemody-namics by way of intestinal protease inhibition orby direct intravascular systemic actions, 5 rats wereprepared as described previously. Endotoxin ad-ministration (4mg/kg) was followed by 8 FOY bolusadministrations (10mg/kg/h) into the jugular veinevery 30 minutes (FOYiv group). This dose issufficient to reduce leukocyte adhesion and atten-uate ischemia/reperfusion injury.28 Hemodynamicparameters were measured as described previously.
Statistical analysis. Data are presented as mean± standard deviation. Repeated measurements ofprotease activity in intestinal fluids and the plasmalevels of interleukin (IL)-6 were analyzed using thepaired Student t test. TheMann-Whitney Rank Sumtest was used to assess differences in trypsin activitybetween the 2 groups. Repeated measurements ofmicrovascular and hemodynamic parameters in thesame group were compared with repeated analysisof variance (ANOVA). For normally distributeddata, ANOVA was followed by the Tukey’s multiplecomparison correction test (in comparison with
control baseline); for Poisson distributed data, theANOVA on ranks was followed by Dunnett’scorrection test. ANOVA was used to assess di-fferences between groups for microcirculatory,hemodynamic, and IL-6 measurements as well asleukocyte activation at the same time. ANOVA wasfollowed by Tukey’s correction test for normallydistributed data, for Poisson distributed data, andANOVA on ranks was followed by Student NewmanKeuls’ correction test; P < .05 was considered to besignificant with a power of .8.
RESULTS
Intraintestinal pancreatic protease inhibition(IPI). Intraintestinal lavage with FOY served toreduce intestinal serine protease activity (FOYgroup, 92 ± 17 units) compared with its baselinevalue (CON group, 2604 ± 734 units, P < .006) orwith intestinal protease activity after lavage withvehicle (LAV group, 630 ± 441 units, P < .029).
Hemodynamic parameters. Endotoxin adminis-tration in the CON and LAV groups was followed bya significant reduction in mean arterial bloodpressure compared with the baseline and with theSHAM group. In contrast, after an initial drop at 20minutes, the arterial blood pressure in the FOYgroup increased to pre-endotoxemia levels andremained stable thereafter. Throughout the rest ofthe experiment there were no significant differ-ences in femoral pressure between SHAM, FOY,and their respective baseline levels. There were alsono significant differences in pressure between theFOY and LAV groups (Fig 1, A). The FOY groupexhibited a trend (not significant) toward higherarterial pressure than did the LAV group.
After endotoxin administration, the heart rate(Fig 1, B) and respiratory rate (Fig 1, C) increasedsignificantly in the CON, LAV, and FOY groups at60 minutes when compared with the SHAM group.At 180 minutes after endotoxemia, the heart andrespiratory rate of the LAV and CON groupremained elevated significantly when comparedwith SHAMs. The FOY group had lower values thanthe CON group (P < .004), which were not signifi-cantly different from the SHAM values. At 300minutes, the FOY group had lower heart and res-piratory rates than the LAV and CON groups(P < .002).
Leukocyte counts. Endotoxemia induced a sig-nificant leukopenia in venous blood of all groups at300 minutes (CON: 2410 ± 650 cells/mm3; LAV:1380 ± 620 cells/mm3; FOY: 3010 ± 2390 cells/mm3) compared with the SHAM group (7400 ±1610 cells/mm3; P < .001).

SurgerySeptember 2003
450 Fitzal et al
Fig 1. Cardiopulmonary parameters in rats measured during 300 minutes after endotoxin administration (4 mg/kgintravenously). The groups are: SHAM, CON (controls with endotoxin administration), LAV (endotoxin administrationand intestinal lavage with buffer), FOY (endotoxin administration and intestinal lavage with the protease inhibitor FOY,0.37 mM). Hypotension (A), tachycardia (B), and hyperventilation (C) were attenuated by intraintestinal proteaseinhibition. Significant differences at P < .05 between LAV compared with its baseline (*); FOY compared with its baseline(y); CON compared with its baseline and SHAM (�); CON compared with its baseline (§); CON, LAV, FOY compared withSHAM (k); LAV compared with FOY (P < .002) (P); FOY compared with CON (P < .004) (#); and LAV compared withSHAM (P < .004) (**); n = 5 rats in each group.

SurgeryVolume 134, Number 3
Fitzal et al 451
Fig 2. Systemic cell activation. Leukocyte and endothelial cell activation in the cremaster muscle in postcapillary venulesmeasured by intravital microscopy at 180, 240, and 300 minutes after endotoxin administration. Groups are the same asin Fig 1. A, Number of rolling leukocytes. B, Number of adherent leukocytes to endothelial cells. C, Leukocyte adhesionindex (LAI = VWBC/VRBC) as indicator for endothelial/leukocyte activation and interaction. D, The number ofpropidium iodide (PI) positive parenchymal cremaster cells as marker for peripheral cell death. Data are presented asmean ± SD. *P < .05 compared with SHAM at the same time point. yP < .05 FOY compared with CON and LAV at thesame time point. �P < .05 when compared with 180 minutes after endotoxemia.
Microvascular leukocyte kinetics. The numberof rolling leukocytes (Fig 2, A) in postcapillaryvenules of the CON and LAV groups at 240 minutesand 300 minutes (P < .004) after endotoxin ad-ministration were increased compared with theSHAM group. In the FOY group, the number ofrolling leukocytes did not differ from that of theSHAMs. FOY attenuated the reduction of rollingleukocytes in the CON and LAV groups at 240 and300 minutes after endotoxemia (P < .004).
In the CON and LAV groups, the centralleukopenia (see previous) was accompanied by anincreased number of leukocytes adhering to post-capillary venules in the microcirculation (Fig 2, B).The number of adherent leukocytes at 180 minutesafter endotoxemia was elevated (P < .003 vs SHAMgroup) and remained high in the CON and LAVgroups. However, in animals receiving FOY lavage,their number decreased already at 240 minutes and
was reduced significantly at 300 minutes afterendotoxin administration (P < .001 compared withthe CON and LAV groups).
The increased leukocyte membrane adhesion inCON and LAV animals was also detected bya decreased LAI at 300 minutes after endotoxinadministration (Fig 2, C) (P < .01 vs values at 180minutes). In the FOY group, the effect wasattenuated at 300 minutes after endotoxin applica-tion (P < .008 vs CON and LAV group).
In some arterioles, rolling and adherent leuko-cytes were observed in CON and LAV rats at 240and 300 minutes after endotoxemia (Table I). Wedid not observe rolling or adherent leukocytes inarterioles of FOY animals.
Microvascular cell death and capillary occlu-sion. Cell death in the cremaster tissue was almostundetectable until 4 hours after endotoxin admin-istration (Fig 2, D). But in the CON and LAV

SurgerySeptember 2003
452 Fitzal et al
Table I. Microvascular parameters in cremaster muscle at different times after endotoxin administration(4 mg/kg IV)
Functional capillary density
Cap/field 180 min 240 min 300 min
SHAM 0.86 ± 0.05 0.65 ± 0.11 0.71 ± 0.16CON 0.69 ± 0.24 0.51 ± 0.33 0.31 ± 0.19*y
LAV 0.45 ± 0.24 0.46 ± 0.28 0.25 ± 0.09*
FOY 0.47 ± 0.30 0.62 ± 0.31 0.62 ± 0.23
Rolling leukocytes in arterioles
Cells/min 180 min 240 min 300 min
SHAM 0 ± 0 0 ± 0 0 ± 0CON 4 ± 9 13 ± 16 11 ± 12LAV 0 ± 0 9 ± 10 19 ± 26FOY 0 ± 0 0 ± 0 0 ± 0
Adherent leukocytes in arterioles
Cells/mm2 180 min 240 min 300 min
SHAM 0 ± 0 0 ± 0 0 ± 0CON 4 ± 8 4 ± 6 7 ± 10LAV 0 ± 0 2 ± 2 4 ± 5FOY 0 ± 0 0 ± 0 0 ± 0
Functional capillary density = the fraction of not-perfused capillaries in the cremaster muscle in 3 different tissue regions (area about 400 lm 3
400 lm). The number of rolling leukocytes in precapillary arterioles passing a given observation point and counted over 30 seconds (cells/min). Thenumber of adherent leukocytes (without movement for at least 30 seconds) per vessel area measured in an arteriolar vessel segment length of 100 lm(cells/mm2). Mean ± SD are shown.*P < .05 compared with SHAM and FOY at the same time point.yP < .05 compared with value at 180 minutes in the same group.
groups, cell death was increased at 300 minutescompared with the respective values at 180 minutes(P < .001) or the values in the SHAM group(P < .001). The majority of the propidium iodide-positive cells were in the extravascular tissue. At 300minutes after endotoxin administration, cell deathin the cremaster muscle tissue of the FOY groupwas reduced compared with the CON and LAVgroups (P < .001) (Fig 2, D).
Functional capillary density in the cremastermuscle was reduced in the CON and LAV groups at300 minutes after endotoxin administration whencompared with the SHAM rats (P < .001) (Table I).At 300 minutes, the FOY group had an improvedcapillary perfusion with greater average functionalcapillary density than in the CON and LAV groups(P < .001).
IL-6 plasma levels. IL-6 levels at 300 minutesafter endotoxemia were elevated significantly inCON and LAV rats compared with their respectivebaseline values before endotoxin administration(Fig 3). In the FOY group, this effect was attenuated(P < .009 vs CONandLAVgroups). The FOY group,however, still showed a significant increase in theIL-6 levels at 300 minutes when compared withthe corresponding values before endotoxemia.
Leukocyte activation. The intestinal homoge-nates of CON and LAV animals 300 minutes afterendotoxin administration activated naive humanleukocytes (P < .002 vs SHAM) (Fig 4, A). A similarpattern was observed with plasma of the CON andLAV groups (P < .001 vs SHAM) (Fig 4, B). In theFOY group, the level of leukocyte activation pro-duced by the intestinal homogenates (Fig 4, A) andby the plasma (Fig 4, B) were attenuated (P < .002vs the CON and LAV groups), but remained higherthan in the SHAM group.
Intestinal homogenates from SHAM animals,which were prepared after rinsing the intestinewith either Krebs-Henseleit buffer or with FOY,did not activate human leukocytes (4 ± 2 acti-vated leukocytes/100 cells and 4 ± 1 activatedleukocytes/100 cells). FOY alone did not activatehuman leukocytes (3 ± 2 activated leukocytes/100cells). After intestinal lavage with either Krebs-Henseleit buffer or with FOY, the lavage fluiddid not activate human leukocytes (15 ± 12 acti-vated leukocytes/100 cells and 18 ± 6 activatedleukocytes/100 cells).
Intestinal tissue structure. In the LAV and CONgroups, endotoxin administration resulted in com-parable intestinal tissue damage with vacuolated

SurgeryVolume 134, Number 3
Fitzal et al 453
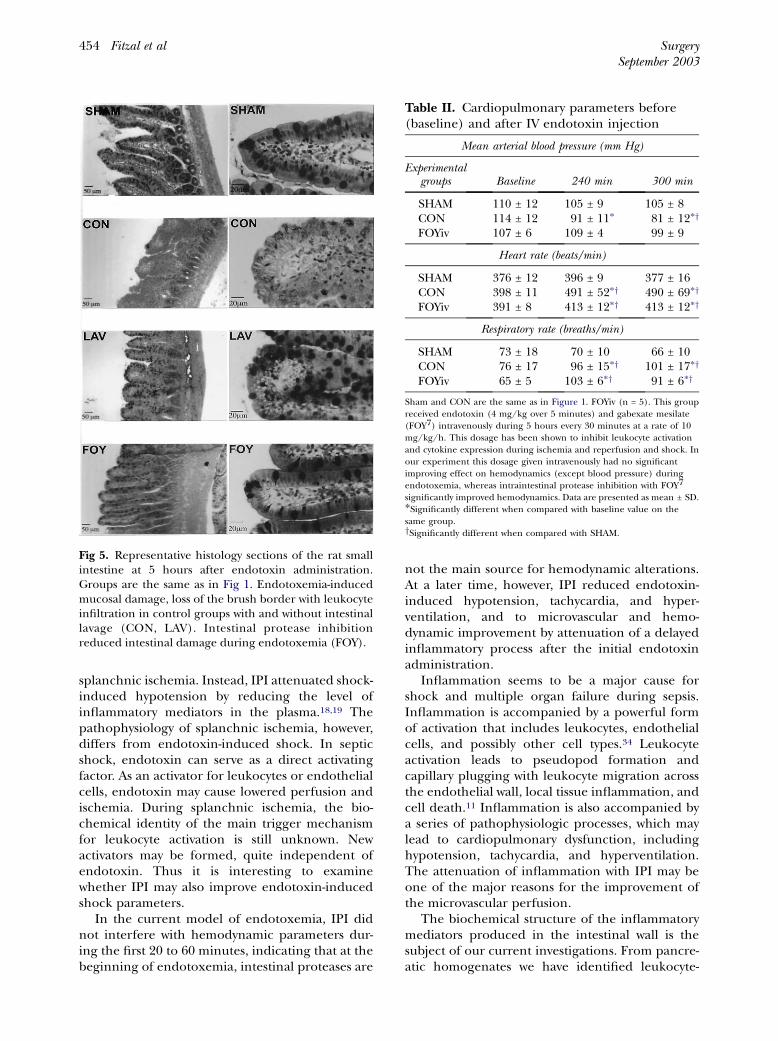
epithelial cells at the tip of the villi, extrusion/sloughing of the epithelial cells, mild edema in thelamina propria, and some loss of adhesion betweenthe epithelial cells. We also observed loss of thebrush border cells in those rats with early formationof villus digestion after endotoxemia. Damage inthe CON and LAV groups was noted only in theupper half of the villi, and no damage was observedfrom the crypts to the serosa. The FOY groupseemed to exhibit less intestinal damage when com-pared with the LAV and CON group. The FOY ratshad only mild extrusion of epithelial cells (Fig 5).
IV administration of FOY versus IPI. The FOYivgroup exhibited no attenuation of tachycardia andhyperventilation at 300 minutes after endotoxinadministration (Table II). Only the blood pressurewas slightly improved compared with the CONgroup but remained significantly reduced com-pared with SHAM animals. Compared with theFOY group, the FOYiv group had a higher respi-ratory rate at 300 minutes and higher heart rate at240 and 300 minutes after endotoxemia (P < .003).
DISCUSSION
Although the possibility that pancreatic pro-teases may play a role in shock has been men-tioned in the past,30 no evidence for or againstthis hypothesis has been advanced. There weresuggestions that proteases may be involved inshock-induced mucosal injury.31,32 Lefer and hiscollaborators observed a relationship betweenpancreatic proteases and a myocardial depressantactivity,33 suggesting that intestinal proteases may
Fig 3. Plasma interleukin-6 levels in femoral arterialblood measured 300 minutes after intravenous endo-toxin administration (4 mg/kg). Groups are the same asin Fig 1. *P < .05 compared with baseline values beforeendotoxin application. yP < .05 compared with FOY atthe same time point.
be involved in the development of cardiac in-sufficiency during shock.
Recently, we obtained direct evidence thatpancreatic homogenates alone as well as severalabdominal organ homogenates mixed with trypsinor chymotrypsin—but not without them—generatepotent inflammatory mediators, which, amongseveral pathophysiologic activities, also cause leu-kocyte activation. The generation of the inflamma-tory mediator from these organ homogenates canbe attenuated by nonspecific protease inhibition.21
The pancreatic homogenates exhibit myocardialdepressor activity.20 Intravenous administration ofa potent protease inhibitor did not restore bloodpressure to normal levels during shock induced by
Fig 4. Neutrophil activation measured by pseudopodformation of naive human leukocytes mixed with (A)intestine homogenates, and (B) plasma before and after300 minutes of intravenous endotoxin administrationinto rats. The increase in activating factors at 240 and 300minutes after endotoxin administration was significantlyreduced by intraintestinal pancreatic protease inhibitionwith FOY. *P < .05 compared with SHAM. yP < .05 com-pared with FOY at the same time point.
SurgerySeptember 2003
454 Fitzal et al
splanchnic ischemia. Instead, IPI attenuated shock-induced hypotension by reducing the level ofinflammatory mediators in the plasma.18,19 Thepathophysiology of splanchnic ischemia, however,differs from endotoxin-induced shock. In septicshock, endotoxin can serve as a direct activatingfactor. As an activator for leukocytes or endothelialcells, endotoxin may cause lowered perfusion andischemia. During splanchnic ischemia, the bio-chemical identity of the main trigger mechanismfor leukocyte activation is still unknown. Newactivators may be formed, quite independent ofendotoxin. Thus it is interesting to examinewhether IPI may also improve endotoxin-inducedshock parameters.
In the current model of endotoxemia, IPI didnot interfere with hemodynamic parameters dur-ing the first 20 to 60 minutes, indicating that at thebeginning of endotoxemia, intestinal proteases are
Fig 5. Representative histology sections of the rat smallintestine at 5 hours after endotoxin administration.Groups are the same as in Fig 1. Endotoxemia-inducedmucosal damage, loss of the brush border with leukocyteinfiltration in control groups with and without intestinallavage (CON, LAV). Intestinal protease inhibitionreduced intestinal damage during endotoxemia (FOY).
not the main source for hemodynamic alterations.At a later time, however, IPI reduced endotoxin-induced hypotension, tachycardia, and hyper-ventilation, and to microvascular and hemo-dynamic improvement by attenuation of a delayedinflammatory process after the initial endotoxinadministration.
Inflammation seems to be a major cause forshock and multiple organ failure during sepsis.Inflammation is accompanied by a powerful formof activation that includes leukocytes, endothelialcells, and possibly other cell types.34 Leukocyteactivation leads to pseudopod formation andcapillary plugging with leukocyte migration acrossthe endothelial wall, local tissue inflammation, andcell death.11 Inflammation is also accompanied bya series of pathophysiologic processes, which maylead to cardiopulmonary dysfunction, includinghypotension, tachycardia, and hyperventilation.The attenuation of inflammation with IPI may beone of the major reasons for the improvement ofthe microvascular perfusion.
The biochemical structure of the inflammatorymediators produced in the intestinal wall is thesubject of our current investigations. From pancre-atic homogenates we have identified leukocyte-
Table II. Cardiopulmonary parameters before(baseline) and after IV endotoxin injection
Mean arterial blood pressure (mm Hg)
Experimentalgroups Baseline 240 min 300 min
SHAM 110 ± 12 105 ± 9 105 ± 8CON 114 ± 12 91 ± 11* 81 ± 12*y
FOYiv 107 ± 6 109 ± 4 99 ± 9
Heart rate (beats/min)
SHAM 376 ± 12 396 ± 9 377 ± 16CON 398 ± 11 491 ± 52*y 490 ± 69*y
FOYiv 391 ± 8 413 ± 12*y 413 ± 12*y
Respiratory rate (breaths/min)
SHAM 73 ± 18 70 ± 10 66 ± 10CON 76 ± 17 96 ± 15*y 101 ± 17*y
FOYiv 65 ± 5 103 ± 6*y 91 ± 6*y
Sham and CON are the same as in Figure 1. FOYiv (n = 5). This groupreceived endotoxin (4 mg/kg over 5 minutes) and gabexate mesilate(FOY7) intravenously during 5 hours every 30 minutes at a rate of 10mg/kg/h. This dosage has been shown to inhibit leukocyte activationand cytokine expression during ischemia and reperfusion and shock. Inour experiment this dosage given intravenously had no significantimproving effect on hemodynamics (except blood pressure) duringendotoxemia, whereas intraintestinal protease inhibition with FOY7
significantly improved hemodynamics. Data are presented as mean ± SD.*Significantly different when compared with baseline value on thesame group.ySignificantly different when compared with SHAM.

SurgeryVolume 134, Number 3
Fitzal et al 455
activating factors with hydrophilic and hydro-phobic characteristics and a range of molecularweights, some of them less than 3kD.4,21 Proteasesthemselves may be less involved in activation ofleukocytes,21 supporting the hypothesis thatproteases have more of a role as producers ofinflammatory mediators. That IPI inhibits theformation of inflammatory mediators supportsthe hypothesis that such mediators may be pre-dominantly produced within the wall of the in-testine by pancreatic digestive enzymes. IPI alsoreduced endotoxin-induced microvascular leuko-cyte and endothelial cell activation, parenchymalcell death, and the production of IL-6.
Sepsis is accompanied by intestinal ischemiaand mucosal cell injury.35-38 Elevation of themucosal epithelial permeability permits entry ofpancreatic enzymes into the interstitium of theintestinal wall. Entry of enzymes into the sub-mucosal interstitium may lead to generation ofinflammatory mediators by proteolytic cleavagewithin the intestinal wall. Our findings support thishypothesis, because IPI served to reduce the gener-ation of activating factors in the small intestineas well as mucosal damage during endotoxemia.Additional aspects of this hypothesis remain to beexamined.
It may be argued that intestinal endotoxin orintestine-derived cytokines may be blocked by IPI.Lavage of the intestine was less effective in pre-venting the systemic inflammation than was lavagecombined with protease inhibition. We investigatedthe relation between IPI, endotoxin activity, andtumor necrosis factor alpha (TNF-a) levels duringintestinal ischemia-induced shock (unpublishedresults). IPI did not reduce intestinal endotoxinactivity. The levels of TNF-a in intestinal homog-enates during shock were identical in IPI andcontrol animals. Endotoxin activity and TNF-alevels did not correlate with the amount ofleukocyte activation either in the intestine or inthe plasma. Taken together, this evidence suggeststhat intestinal endotoxin and intestine-derivedTNF-a may be less involved in the mechanisms bywhich IPI attenuates shock.
We hypothesize that intestinal absorption of FOYand inhibition of leukocyte activation and adhe-sion,39 as well as reduction of cytokine levels,40 mayin part account for the improved hemodynamicsand decreased microvascular inflammation in thecurrent experiments. However, IV-administeredFOY at a dose known to reduce leukocyte activa-tion28 did not improve hemodynamics duringsepsis in our model. Only endotoxin-inducedhypotension was partially attenuated in this group,
possibly as a result of the inhibitory effect of FOYon the constitutive and inducible nitric oxidesynthase.41 Our results do not clarify whether FOYleaking into the extraintestinal tissue may, in part,have accounted for the effects seen in the FOYgroup. Further studies are required with otherprotease inhibitors.
Intestine-derived oxygen radicals may be influ-enced by IPI. Recent data demonstrated that IPIdid not affect either xanthine dehydrogenase toxanthine oxidase conversion or xanthine oxidaseactivity in the ischemic intestine. The addition ofallopurinol to the lavage fluid during IPI did notincrease the beneficial effect of IPI even thoughallopurinol inhibited the xanthine oxidase activityin the ischemic intestine.19 Systemic leukocyteactivation and shock may be related more closelyto protease activity than to the action of oxygen freeradicals derived from xanthine oxidase.
In conclusion, we provide here the first evidenceto suggest that, in addition to endotoxin, intestinalproteases may be involved in the generation ofintestine-derived inflammatory mediators in theplasma. These mediators may sustain systemic cellactivation and produce organ injury after anendotoxin challenge. Intraintestinal pancreaticprotease blockade may have clinical utility in septicshock patients.
The authors wish to thank Mr William H. Loomis,UCSD Department of Surgery, for technical help withthe IL-6 measurements, and Dr Tony Hugli, La JollaInstitute for Molecular Medicine, for his thoughtfulsuggestions regarding this study. FOY� was a gift fromOno Pharmaceutical.
REFERENCES
1. Mazzoni MC, Schmid-Schobein GW. Mechanisms andconsequences of cell activation in the microcirculation.Cardiovasc Res 1996;32:709-19.
2. Parillo JE, Burch C, Shelhamer JH, Parker MM, Natanson C,Schuette W. A circulating myocardial depressing substancein humans with septic shock. J Clin Invest 1985;76:1539-53.
3. Haglund U. Systemic mediators released from the gut incritical illness. Crit Care Med 1993;21:S15-S18.
4. Schmid-Schonbein GW, Kistler EK, Hugli TE. Mechanismsfor cell activation and its consequences for biorheology andmicrocirculation: Multiple-organ failure in shock. Biorheol2001;38:185-201.
5. Yang S, Koo DJ, Chaudry IH, Wang P. The important role ofthe gut in initiating the hyperdynamic response duringearly sepsis. J Surg Res 2000;89:31-7.
6. Evans WE, Darin JC. Effect of enterectomy in endotoxinshock. Surgery 1966;60:1026-9.
7. Hassoun HT, Kone BC, Mercer DW, Moody FG, WeisbrodtHW, Moore FA. Post-injury multiple organ failure: the roleof the gut. Shock 2001;15:1-10.

SurgerySeptember 2003
456 Fitzal et al
8. Tamion F, Richard V, Lyoumi S, et al. Gut ischemia andmesenteric synthesis of inflammatory cytokines afterhemorrhagic or endotoxic shock. Am J Physiol 1997;273:G314-21.
9. Adams CA, Xu DZ, Deitch EA. Factors larger than 100kD inposthemorrhagic shock mesenteric lymph are toxic forendothelial cells. Surgery 2001;129:351-62.
10. Deitch EA, Adams C, Lu Q, Xu DZ. A time course study ofthe protective effect of mesenteric lymph duct ligation onhemorrhagic shock-induced pulmonary injury and the toxiceffects of lymph from shocked rats on endothelial cellmonolayer permeability. Surgery 2001;129:39-47.
11. Schmid-Schonbein GW, Hugli TE, Kistler EB, Sofianos A,Mitsuoka H. Pancreatic enzymes and microvascular cellactivation in multiorgan failure. Microcirc 2001;8:5-14.
12. Hardaway R. A review of septic shock. Am Surg 2000;66:22-9.
13. Zeni F, Freeman B, Natanson C. Anti-inflammatory thera-pies to treat sepsis and septic shock. A reassessment. CritCare Med 1997;25:1095-100.
14. Mainous MR, Ertel W, Chaudry IH, Deitch EA. The gut:a cytokine generating organ in systemic inflammation?Shock 1995;4:193-9.
15. Deitch EA, Xu D, Franko L, Ayala A, Chaudry IH. Evidencefavoring the role of the gut as a cytokine-generating organin rats subjected to hemorrhagic shock. Shock 1994;1:141-6.
16. Koike K, Moore EE, Moore FA, Read RA, Carl VS, BanerjeeA. Gut ischemia/reperfusion produces lung injury inde-pendent of endotoxin. Crit Care Med 1994;22:1438-44.
17. Bone RC. Why sepsis trials fail. JAMA 1996;276:565-6.18. Mitsuoka H, Kistler EB, Schmid-Schonbein GW. Generation
of in vivo activating factors in the ischemic intestine bypancreatic enzymes. PNAS 2000;97:1772-7.
19. Mitsuoka H, Kistler EB, Schmid-Schonbein GW. Proteaseinhibition in the intestinal lumen: Attenuation of inflam-mation and early indicators of multiple organ failure inshock. Shock 2002;17:205-9.
20. Kistler EB, Lefer AM, Hugli TE, Schmid-Schonbein GW.Plasma activation during splanchnic arterial occlusionshock. Shock 2000;14:30-4.
21. Kistler EB, Hugli TE, Schmid-Schonbein GW. The pancreasas a source of cardiovascular cell activating factors. Micro-circ 2000;7:183-92.
22. Mitsuoka H, Schmid-Schonbein GW. Mechanisms forblockade of in vivo activator production in the ischemicintestine and multi-organ failure. Shock 2000;14:522-7.
23. Fitzal F, De Lano F, Young C, Rosario HS, Schmid-Schonbein GW. Intestinal pancreatic protease inhibitionattenuates intestinal-ischemia-induced shock by reducingcell activation and gut injury. J Vasc Res 2002;39:320-9.
24. Baez S. An open cremaster muscle preparation for the studyof blood vessels by in vivo microscopy. Microvas Res1973;5:384-94.
25. Suematsu M, DeLano FA, Poole D, Engler RL, Miyasaka M,Zweifach BW, Schmid-Schonbein GW. Spatial and temporal
correlation between leukocyte behavior and cell injury inpostischemic rat skeletal muscle. Lab Invest 1994;70:684-95.
26. Schmid-Schonbein GW, Skalak R, Simon SI, Engler RL. Theinteraction between leukocytes and endothelium in-vivo.Proc NY Acad Sci 1988;516:348-61.
27. Aosasa S, Ono S, Mochizzuki H, Tsujimoto H, Ueno C,Matsumoto A. Mechanism of the inhibitory effect ofprotease inhibitor on tumor necrosis factor alpha pro-duction of monocytes. Shock 2001;15:101-5.
28. Harada N, Okajima K, Mushimoto S. Gabexate mesylate,a synthetic protease inhibitor, reduces ischemia/reperfu-sion injury of rat liver by inhibiting leukocyte activation. CritCare Med 1999;27:1958-64.
29. Ohashi I, Nishijima J, Murata A, et al. Inhibitory effect ofa synthetic protease inhibitor (gabexate mesylate) in therespiratory burst oxidase in human neutrophils. J Biochem1995;118:1001-6.
30. Bounous G. Mucosal injury in low flow states. Can J Surg1974;17:434.
31. Montgomery A, Borgstrom A, Haglund U. Pancreaticproteases and intestinal mucosal injury after ischemia andreperfusion in the pig. Gastronterology 1992;102:216-22.
32. Bounous G. Pancreatic proteases and oxygen-derived freeradicals in acute ischemic enteropathy. Surgery 1985;99:92-4.
33. Lefer AM, Spath JA. Pancreatic hypoperfusion and theproduction of a myocardial depressant factor in hemor-rhagic shock. Ann Surg 1974;179:868-76.
34. Nieuwenhuijzen GAP, Haskel Y, Lu Q, et al. Macrophageelimination increases bacterial translocation and gut-origin-septicemia, but attenuates symptoms and mortality rate ina model of systemic inflammation. Ann Surg 1993;218:791-9.
35. Turnbull RG, Talbot JA, Hamilton SM. Hemodynamicchanges in gut barrier function in sequential hemorrhagicand endotoxic shock. Crit Care Med 1995;38:705-12.
36. Oldner A, Goiny M, Rudehill A, Ungerstedt U, Sollevi A.Tissue hypoxanthine reflects gut vulnerability in porcineendotoxin shock. Crit Care Med 1999;27:790-7.
37. Smithies M, Yee TH, Jackson L, Beale R, Bihari D.Protecting the gut and the liver in the critically ill: effectsof dopexamine. Crit Care Med 1994;22:789-95.
38. McCarthy Pastores S, Katz DP, Kvetan V. Splanchnicischemia and gut mucosal injury in sepsis and the multipleorgan dysfunction syndrome. Am J Gastroenterol 1996;91:1697-710.
39. Lu SP, Tsai CC, Shau WY, et al. Effects of gabexate mesilate(FOY) on ischemia-reperfusion-induced acute lung injuryin dogs. J Surg Res 1999;87:152-63.
40. Malyon AD, Husein M, Weiler-Mithoff EM. How manyprocedures to make a breast? Br J Plast Surg 2001;54:227-31.
41. Colasanti M, Persichini T, Venturini G, Menegatti E, LauroGM, Ascenzi P. Effect of gabexate mesylate (FOY), a drugfor serine proteinase-mediated diseases on the nitric oxidepathway. Biochem Biophys Res Commun 1998;246:453-6.
Copyright © 2022 FDOKUMEN




















