
Contribution of BK channels to action potential repolarisation at minimal cytosolic Ca2+...
15
1 23 Pflügers Archiv - European Journal of Physiology European Journal of Physiology ISSN 0031-6768 Volume 462 Number 4 Pflugers Arch - Eur J Physiol (2011) 462:545-557 DOI 10.1007/s00424-011-0991-9 Contribution of BK channels to action potential repolarisation at minimal cytosolic Ca 2+ concentration in chromaffin cells Ricardo S. Scott, Diego Bustillo, Luis Alcides Olivos-Oré, Inmaculada Cuchillo- Ibañez, Maria Victoria Barahona, Emilio Carbone, et al.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Contribution of BK channels to action potential repolarisation at minimal cytosolic Ca2+...

1 23
Pflügers Archiv - EuropeanJournal of PhysiologyEuropean Journal of Physiology ISSN 0031-6768Volume 462Number 4 Pflugers Arch - Eur J Physiol (2011)462:545-557DOI 10.1007/s00424-011-0991-9
Contribution of BK channels to actionpotential repolarisation at minimalcytosolic Ca2+ concentration inchromaffin cells
Ricardo S. Scott, Diego Bustillo, LuisAlcides Olivos-Oré, Inmaculada Cuchillo-Ibañez, Maria Victoria Barahona, EmilioCarbone, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ION CHANNELS, RECEPTORS AND TRANSPORTERS
Contribution of BK channels to action potentialrepolarisation at minimal cytosolic Ca2+ concentrationin chromaffin cells
Ricardo S. Scott & Diego Bustillo & Luis Alcides Olivos-Oré &
Inmaculada Cuchillo-Ibañez & Maria Victoria Barahona & Emilio Carbone &
Antonio R. Artalejo
Received: 20 June 2011 /Accepted: 20 June 2011 /Published online: 14 July 2011# Springer-Verlag 2011
Abstract BK channels modulate cell firing in excitablecells in a voltage-dependent manner regulated by fluctua-tions in free cytosolic Ca2+ during action potentials. Indeed,Ca2+-independent BK channel activity has ordinarily beenconsidered not relevant for the physiological behaviour ofexcitable cells. We employed the patch-clamp techniqueand selective BK channel blockers to record K+ currentsfrom bovine chromaffin cells at minimal intracellular (about10 nM) and extracellular (free Ca2+) Ca2+ concentrations.Despite their low open probability under these conditions(V50 of +146.8 mV), BK channels were responsible for
more than 25% of the total K+ efflux during the firstmillisecond of a step depolarisation to +20 mV. Moreover,BK channels activated about 30% faster (τ=0.55 ms) thanthe rest of available K+ channels. The other main source offast voltage-dependent K+ efflux at such a low Ca2+ was atransient K+ (IA-type) current activating with V50=−14.2 mV.We also studied the activation of BK currents in responseto action potential waveforms and their contribution toshaping action potentials both in the presence and theabsence of extracellular Ca2+. Our results show that BKchannels activate during action potentials and acceleratecell repolarisation even at minimal Ca2+ concentration,and suggest that they could do so also in the presence ofextracellular Ca2+, before Ca2+ entering the cell facilitatestheir activity.
Keywords BK channel . A-type potassium current . Actionpotential . Chromaffin cell . Single channel recordings
AbbreviationsAP Action potentialAPW Action potential waveformBKi Inactivating BK current or channelBKs Non-inactivating BK current or channelCai
2+ Cytosolic Ca2+
Cao2+ Extracellular Ca2+
ChTx CharybdotoxinHAD Half-amplitude durationIA Fast inactivating potassium currentIbTx Iberiotoxinki Rate constant of inactivationN-methyl-D-glucamine
NMDG
Electronic supplementary material The online version of this article(doi:10.1007/s00424-011-0991-9) contains supplementary material,which is available to authorized users.
R. S. Scott : I. Cuchillo-IbañezDepartamento de Farmacología y Terapéutica,Universidad Autónoma de Madrid,28029 Madrid, Spain
D. Bustillo : L. A. Olivos-Oré :M. V. Barahona :A. R. Artalejo (*)Departamento de Toxicología y Farmacología & InstitutoUniversitario de Investigación en Neuroquímica,Universidad Complutense de Madrid,28040 Madrid, Spaine-mail: [email protected]
E. CarboneDipartimento di Neuroscienze, Università degli Studi di Torino,10125 Torino, Italy
Present Address:R. S. Scott : I. Cuchillo-IbañezInstituto de Neurociencias UMH-CSIC,Campus de San Juan, Sant Joan d’Alacant,Alicante, Spain
Pflugers Arch - Eur J Physiol (2011) 462:545–557DOI 10.1007/s00424-011-0991-9
Author's personal copy

τ Time constantTEA TetraethylammoniumVh Holding potential
Introduction
Upon splanchnic nerve stimulation, chromaffin cells releasecatecholamines, opioids, and ATP to the bloodstream, thuscontributing as the endocrine arm of the sympatheticnervous system to body homeostasis during stressfulsituations [1]. Adenomedullary hormone release is regulat-ed by K+ channels through the control of multiple aspectsof cellular excitability [7, 15]. Five different K+ currentshave been identified in bovine chromaffin cells: a highconductance voltage- and Ca2+-dependent K+ current (BKcurrent; [28, 29]), a delayed rectifier K+ current [29, 39], asmall conductance Ca2+-activated K+ current [2], a Gprotein-activated K+ current [5] and an M-type current[52]. Despite the similarities of chromaffin cells withneurons, a transient A-type K+ current that regulatesinterspike intervals [8, 26] has not been reported in bovinechromaffin cells. A-currents typically activate at subthresh-old voltages and rapidly inactivate. Since chromaffin cells,like many neurons, are able to fire trains of action potentials(APs) [1], this current might be involved in the regulationof the firing frequency during these trains.
The K+ current that has been more thoroughly studied inrelation to action potential firing in chromaffin cells is theBK current. BK channels shape the initial phase ofrepolarisation of Na+- and Ca2+-dependent APs [4, 35,38]. BK channels activate in a voltage-dependent fashion,but the increase of cytosolic Ca2+ (Cai
2+) greatly enhancestheir voltage sensitivity by an allosteric mechanism [19, 20,22]. BK channels are composed of two distinct subunits, αand β, which are arranged in a 1:1 stoichiometry [46]. Eachchannel exists as a tetramer formed by four α subunitseither alone or in paired combinations with β subunits. αSubunits are the pore-forming units whereas β ones areregulatory. While only one α subunit has been identifieduntil now, four types of β subunits (β1-β4) have beenreported [14]. The different β subunits modify the gatingand pharmacological properties of BK channels [6, 14, 16,30, 45, 48, 50, 56]. Thus, channels containing the α subunitalone and those consisting also of β1 or β4 subunitsproduce sustained currents that do not inactivate [31, 53];on the contrary, those formed by the α subunit and β2 orβ3 subunits give rise to inactivating BK currents [53, 55].Likewise, the various β-subunit phenotypes also affect thesensitivity to pore-blockers like charybdotoxin (ChTx) andiberiotoxin (IbTx). Coexpression of α subunit with β2 orβ3 subunits confers a reduced sensitivity to these toxins
whereas BK channels formed by α and β4 subunits areresistant [10, 31, 54].
BK channels from chromaffin cells are kinetically andmolecularly heterogeneous. A functional variant exhibitingrapid inactivation (BKi) and presumably formed by mix-tures of α and β2 or β3 subunits has been wellcharacterised in rat and bovine chromaffin cells [25, 41,42, 55]; this variant coexists with a non-inactivatingcounterpart (BKs), which may be composed by theassociation of α and β4 subunits [9]. Interestingly, BKi
and BKs channels from rat chromaffin cells have beenassigned to different roles in cellular excitability. Thus, cellsexpressing mostly BKi are capable to produce moresustained AP firing than those expressing mainly BKs
channels [24, 42]. This differential behaviour has beenattributed to the more negatively shifted activation range forBKi channels relative to that of BKs channels [44].
The physiological relevance of BK channels has beennormally related to their sensitivity to Cai
2+ rises. Althoughit is well established that BK channels can be maximallyactivated by extreme depolarisations even when Cai
2+ isbuffered to low nanomolar levels [9, 11, 19, 20, 22, 50, 55],the relative contribution of this current to the total K+ effluxand, therefore, its relevance for the electrical behaviour ofthe cells at minimal Cai
2+ has not been investigated. Here,we explore the biophysical properties of two K+ currents(BK and A-type) and investigate the role of BK channels inmodulating AP shape from bovine chromaffin cells in theabsence of a Cai
2+ rise. Our results indicate that BKchannels accelerate repolarisation of APs under thoseconditions and suggest that they could do so also in thepresence of extracellular Ca2+, before Ca2+ entering the cellfacilitates their activity.
Methods
Isolation and culture of bovine chromaffin cells
Bovine chromaffin cells were isolated from adrenal glandsof adult cows and kept in culture following standardmethods [32, 49]. Cells were plated in 1 cm-diameter glasscoverslips at a density of 5×104 cells per coverslip andwere used for 1–4 days after isolation.
Electrophysiological methods and data analysis
Pipettes of approximately 3 MΩ were fabricated fromborosilicate glass (Kimax 51, Witz Scientific, Holland, OH,USA) with a Narishige PP83 puller (Japan) and subse-quently fire-polished and coated with dental wax. An EPC-9 patch-clamp amplifier and PULSE/PATCHMASTERsoftware (HEKA Elektronic, Lambrecht, Germany) were
546 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

used for data acquisition and stimuli generation. Data wereacquired through a combination of the high time resolutionPULSE/PATCHMASTER software and the lower timeresolution X-Chart plug-in module to PULSE software. Inthe high resolution mode, conventional whole-cell orperforated-patch currents or voltages were filtered at2 kHz and sampled at 8.33 kHz (120 μs), whereas currentsfrom excised-patches were filtered at 2.9 kHz and sampledat a frequency of 10 kHz. Low frequency acquisition was at3 Hz irrespective of the recording configuration. Forvoltage-clamp experiments, after breaking-in or aftermembrane-patch perforation had been stabilised, cells wereheld at a voltage (Vh) of −70 mV, and those with leakagecurrents larger than 20 pAwere rejected. Series resistancesin the whole-cell and perforated-patch configurations werealways kept below 10 and 25 MΩ, respectively, and werecompensated by 90%. A p/n protocol for subtracting leakand capacitive currents was routinely applied except for tailcurrent measurements.
Drugs were applied onto the cell under investigation bymeans of a gravity-driven perfusion system with fiveindependent lines controlled by electronic valves (The LeeCompany, Westbrook, CO, USA). This system allows theexchange of the medium surrounding a cell in less than 1 s.Experiments were conducted at room temperature (∼25°C).
Activation curves were built by using tail-currentmeasurements upon repolarisation from different potentials.To avoid capacitive transient contamination, monoexponen-tial functions starting 500 μs after the beginning of therepolarisation were fit to the deactivation phase of thecurrents. Peak–current amplitudes were extrapolated to thebeginning of the repolarising step from the resultingexponential fits and subsequently plotted against membranepotential during the activation pulse. The followingBoltzmann equation was then fitted to the plotted data:
I=Imax ¼ 1= 1þ exp V50 � Vmð Þ=K½ � ð1Þ
where V50 is the voltage at which tail current amplitude (I)is half of its maximum (Imax), Vm is the membranepotential, and K is the reciprocal of the slope at theinflexion point of the activation curve.
To generate steady-state inactivation curves, a double-pulse Cole-Moore protocol was used and the currentresponses to the second pulse plotted against the potentialof the first-conditioning pulse. The following Boltzmannequation was fitted to that relation:
I=Imax ¼ 1þ exp V50 � Vcð Þ=K½ ��1 ð2Þwhere Vc is the potential during the conditioning pulse, V50
is the voltage at which conductance is inactivated by 50%(with respect to the response elicited in the absence of a
conditioning pulse), and K is the reciprocal of the slope atthe inflexion point of the inactivation curve. The fitting wasalways performed by the least-squares method.
The minimum number of active channels in outside-outpatches (n) was estimated as:
n ¼ n0=p ð3Þ
where n’ is the mean of the maximum number of currentlevels observed over tens of consecutive sweeps at a givenpotential and p is the BK channel open probability at thesame potential, obtained from the activation vs. voltagerelationship [18].
Ensemble averages of BK and non-BK channel activityfrom patches containing mixtures of K+ channels wereperformed by (1) identifying BK channel unitary currentsfrom sweeps showing unequivocal large BK channelopenings, (2) disregarding openings with current ampli-tudes lower than those of BK channels and plotting theminimum number of points required to describe anidealised BK channel activity, (3) interpolating amongthese points to generate traces representing idealised BKchannel openings, and (4) subtracting these traces fromactual traces to get the non-BK channel activity. Averagesof both idealised (BK currents) and subtracted (non-BKcurrents) traces reflect the relative contribution of voltage-dependent BK current and non-BK current to total K+
current.The following AP characteristics were measured: (1)
peak, defined as the most positive value recorded during theAP; (2) amplitude, the peak less the resting membranepotential, the latter being defined as the value of themembrane potential recorded 10 ms prior to the onset of adepolarising current injection, and (3) half-amplitudeduration (HAD), the duration for which AP amplitudeexceeds 50% of its amplitude. For all cells, average valuesfor all parameters in control and after drug treatment werecalculated from 3 to 15 individual APs.
Solutions
The standard external solution had the following composi-tion (in millimolars)—140 NaCl, 2.8 KCl, 2 MgCl2, tenglucose, and ten HEPES; pH 7.3, adjusted with NaOH. Tomeasure K+ tail currents, an extracellular solution of thefollowing composition was used—140 KCl, 2 MgCl2, and10 HEPES; pH 7.3, adjusted with KOH. K+ currentsactivated by action potential waveforms (APWs) wereisolated by employing a standard external solution modifiedby replacing Na+ with N-methyl-D-glucamine (NMDG).The pipette’s solution always contained (in millimolars)—140 KCl and 10 HEPES; pH 7.3, adjusted with KOH.BAPTA (10 mM) was also added to this solution to buffer
Pflugers Arch - Eur J Physiol (2011) 462:545–557 547
Author's personal copy

Cai2+ to around 10 nM [29]. In some experiments, 10 mM
ethylene-bis(oxyethylenenitrilo)tetraacetic acid (EGTA), in-stead of BAPTA, was added to the internal solution, withoutproducing obvious differences. The internal solution usedfor perforated-patch recordings was the same as forconventional whole-cell experiments except that itcontained amphotericin B (0.30.5 mg×mL−1). Amphoter-icin B was prepared as previously described [49].
Chemicals
Dulbecco modified Eagle’s medium, foetal bovine serum,penicillin, and streptomycin were purchased from Gibco(Madrid, Spain). Bovine serum albumin, 5-Fl-2′-deoxyur-idine, cytosine arabinoside, L-leucine methyl ester, dime-thylsulphoxide, 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES), EGTA, 1,2-bis(2-aminophe-noxy)ethane-N,N,N′,N′-tetraacetic acid (BAPTA), NMDG,amphotericin B, and tetraethylammonium (TEA) were fromSigma-Aldrich (Spain); IbTx and ChTx were from PeptideInstitute (Osaka, Japan). Collagenase A from Clostridiumhistolyticum was obtained from Boehringer-Mannheim(Barcelona, Spain), Percoll from Pharmacia (Barcelona,Spain), and paxilline from Tocris Biosciences (Bristol, UK).Other reagents were from Panreac (Barcelona, Spain).
Statistical analysis
Results are expressed as means±standard error of the mean(SEM). Statistical differences between means were assessedby the Student’s t test. A p value<0.05 was taken as thelimit of significance.
Results
Multiple voltage-activated K+ currents with differentinactivation kinetics and pharmacological sensitivitiesare expressed in bovine chromaffin cells
We have studied K+ currents from bovine chromaffin cellsdialysed in the whole-cell configuration with an internalsolution containing 10 mM BAPTA and superfused with abath solution devoid of added Ca2+ (minimal Cai
2+). Underthese conditions, the occupancy of Ca2+ binding sites in BKchannels is thought to be negligible [20]. Step depolarisa-tions from Vh=−70 mV elicited partially inactivatingoutward currents (Fig. 1a, left panel) that were attenuatedby ChTx (150 nM; Fig. 1a, right panel, and b), a selectiveBK channel blocker [21]. The remaining ChTx-resistantcomponent was mostly due to a fast inactivating K+ current(henceforth, named IA) that, at potentials positive to +5 mV,was progressively masked by a slowly inactivating current
(mean rate constant of inactivation, ki, ∼10 s−1; Fig. 1a,right panel, and c). These results suggest the existence of atleast three types of voltage-activated K+ currents in bovinechromaffin cells differing in their inactivation kinetics andsensitivity to ChTx: (1) a ChTx-sensitive current, namelythe BK current, (2) a fast inactivating ChTx-resistantcurrent that we have designated as IA, and (3) a non- orslowly inactivating ChTx-resistant current that couldcorrespond, at least in part, to a delayed rectifier K+ current[39].
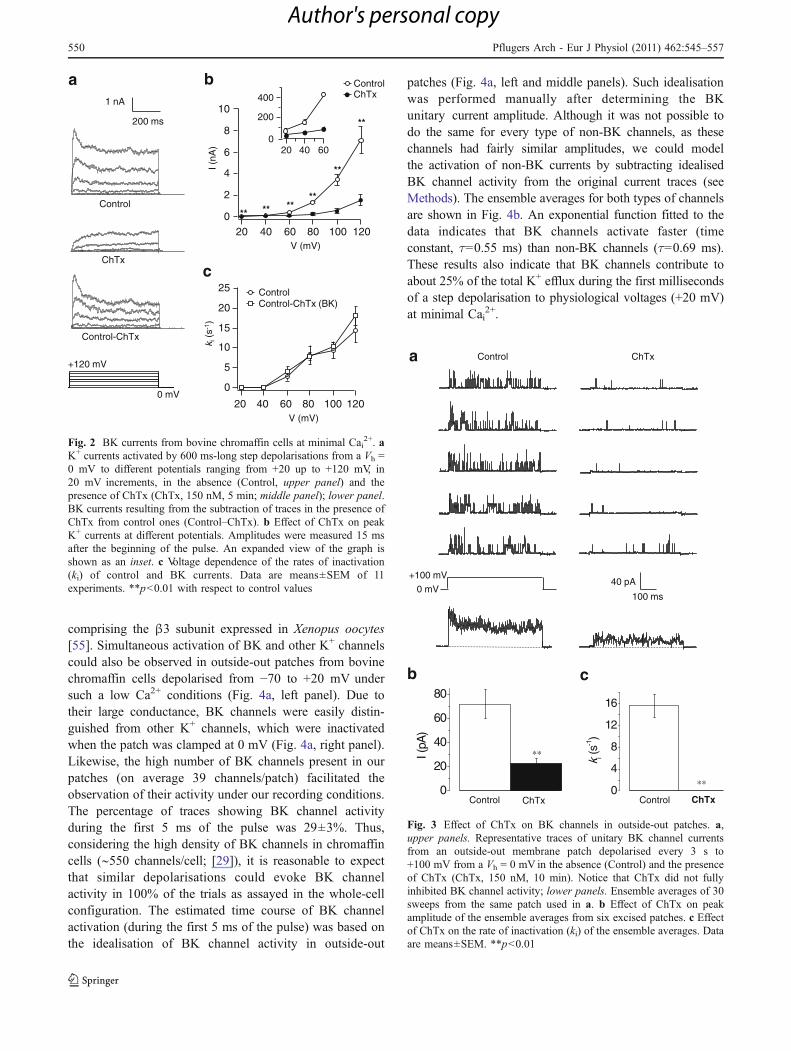
To record BK currents in isolation, cells were clamped at0 mV for over 2 min to inactivate the delayed rectifierK+ current [39, 47] and IA. Step depolarisations from aVh = 0 mV activated outward currents that were stronglyblocked by 150 nM ChTx (80%; Fig. 2a, b). The currentcomponent sensitive to ChTx, likely the BK current,exhibited voltage-dependent inactivation (Fig. 2c). Due tothe lack of inactivation of ChTx-resistant currents, the rateof inactivation of K+ currents in control conditions wassimilar to that of the ChTx-sensitive component at allvoltages, suggesting that, under these conditions, BKcurrent inactivation kinetics can be studied in the absenceof channel blockers (Fig. 2c). The ChTx-resistant currentcomponent could be attributed to either delayed rectifier K+
channels still active despite a sustained depolarisation at0 mV, or to non-inactivating BK channels insensitive to thetoxin. The latter possibility was favoured by experimentsemploying paxilline, a selective BK channel blocker [40]that inhibits both fast and slowly inactivating BK channelsin rat chromaffin cells [27]. Supplemental Fig. 1A showsrepresentative recordings of the effects of paxilline (1 μM)on K+ currents elicited by step-like depolarisation fromeither a Vh = −70 or 0 mV in two chromaffin cellspredominantly expressing non-inactivating BK channels.In the cell held at −70 mV, paxilline treatment disclosed fastinactivating K+ currents similar to those observed in cellsincubated with ChTx. On the other hand, paxilline fullyabolished the K+ currents when holding the cell at 0 mV,suggesting that the ChTx-resistant currents activated fromthat Vh are mostly carried by non-inactivating BK channels(Suppl. Fig 1B). To further support this, we studied theeffect of ChTx on unitary BK channel currents recordedfrom outside-out patches (Fig. 3). Multiple BK channelswere easily detected and identified by their large conduc-tance (∼190 pS with 140 mM Ki
+ and 2.8 mM Ko+) [28,
29]. Moreover, BK channels were the only channel typeobserved in more than 95% of outside-out patches analysed(n=50). The ensemble averages of unitary BK currentsduring test pulses to +100 mV from Vh=0 mV showed clearinactivating time courses that were abolished after additionof ChTx (150 nM) to the external side of the patches. Thetoxin reduced BK channel open probability by ∼75% andunmasked a non-inactivating ChTx-resistant BK current
548 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

component (Fig. 3a, c). Interestingly, the contribution ofthis non-inactivating and ChTx-resistant component tothe averaged BK current in excised patches (∼25%;Fig. 3b) is similar to that observed in whole-cell record-ings of BK current at the same voltages (Fig. 2b).Altogether, our results point toward the coexistence inbovine chromaffin cells of inactivating and non-inactivating BK channels with differential sensitivity toChTx and suggest that most of the ChTx-resistant currentrecorded in the whole-cell configuration from Vh=0 mVcan be attributed to non-inactivating BK channels (BKs
channels). Interestingly, the adrenal medulla expresses theβ4 subunit that produces non-inactivating and ChTx-resistant BK currents [9].
The effects of TEA on K+ currents were also consistentwith those observed in the presence of ChTx. ExtracellularTEA produced a concentration-dependent reduction of K+
currents evoked by long depolarising pulses (Suppl. Fig.2A and B) and caused pronounced changes in their rate ofinactivation (Suppl. Fig. 2C). In this respect, TEA(30 mM) accelerated the inactivation in cells depolarisedfrom Vh=−70 mV to potentials positive to +40 mV, thussuggesting a preferential block of BK current with respectto IA (Suppl. Fig. 2C, left panel). At variance, TEA(30 mM) slowed the inactivation of K+ currents activatedfrom Vh=0 mV (Suppl. Fig. 2C, right panel), possiblybecause it predominantly affects BKi channels, the onlyinactivating channels active at that Vh [29].
Fast activation of BK channels in outside-out patches
The masking of IA by BK current shown in Fig. 1 isindicative of a fast activation of BK channels even atminimal Cai
2+, as it has also been reported for BK channels
Control ChTx
50 ms1 nA
Control
cb
a
0
10
20
30
40
50
60 ControlChTx
i(s
k-1)
V (mV)
ChTx
200 ms
2 nA
0 mV+130 mV
-70 mV
-50 0 50 100 150 -50 0 50 100 150
0
2
4
6
8
10
12 ControlChTx
I (nA
)
V (mV)
* * ****
**
**
**
****
** ****
*
*
Fig. 1 Voltage-activated K+ currents in bovine chromaffin cells atminimal Cai
2+. a Typical experiment showing K+ currents activated by600 ms-long step depolarisations from a Vh = −70 mV to differentpotentials ranging from −50 mV to +130 mV in the absence (Control,left panel) and the presence of ChTx (150 nM for 5 min; ChTx, rightpanel). b Effect of ChTx on peak current amplitudes at the differenttest potentials. c Voltage dependence of the rate of K+ currentinactivation (ki) in the absence and the presence of ChTx. The inset
shows in an expanded scale the initial portion of current responsesevoked by voltage pulses to −10 mV up to +50 mV. ChTx discloses afast inactivating K+ current, IA, which is apparent between −40 and+100 mV; notice that in the absence of the toxin, IA is masked atvoltages positive to +5 mV by a non- or slowly inactivating K+
current. Cells were dialysed with a solution containing 10 mMBAPTA and superfused with a solution devoid of added Ca2+. Data aremeans±SEM of 21 experiments. *p<0.05, **p<0.01
Pflugers Arch - Eur J Physiol (2011) 462:545–557 549
Author's personal copy
comprising the β3 subunit expressed in Xenopus oocytes[55]. Simultaneous activation of BK and other K+ channelscould also be observed in outside-out patches from bovinechromaffin cells depolarised from −70 to +20 mV undersuch a low Ca2+ conditions (Fig. 4a, left panel). Due totheir large conductance, BK channels were easily distin-guished from other K+ channels, which were inactivatedwhen the patch was clamped at 0 mV (Fig. 4a, right panel).Likewise, the high number of BK channels present in ourpatches (on average 39 channels/patch) facilitated theobservation of their activity under our recording conditions.The percentage of traces showing BK channel activityduring the first 5 ms of the pulse was 29±3%. Thus,considering the high density of BK channels in chromaffincells (∼550 channels/cell; [29]), it is reasonable to expectthat similar depolarisations could evoke BK channelactivity in 100% of the trials as assayed in the whole-cellconfiguration. The estimated time course of BK channelactivation (during the first 5 ms of the pulse) was based onthe idealisation of BK channel activity in outside-out
patches (Fig. 4a, left and middle panels). Such idealisationwas performed manually after determining the BKunitary current amplitude. Although it was not possible todo the same for every type of non-BK channels, as thesechannels had fairly similar amplitudes, we could modelthe activation of non-BK currents by subtracting idealisedBK channel activity from the original current traces (seeMethods). The ensemble averages for both types of channelsare shown in Fig. 4b. An exponential function fitted to thedata indicates that BK channels activate faster (timeconstant, τ=0.55 ms) than non-BK channels (τ=0.69 ms).These results also indicate that BK channels contribute toabout 25% of the total K+ efflux during the first millisecondsof a step depolarisation to physiological voltages (+20 mV)at minimal Cai
2+.
0
4
8
12
16
k i (s-1
)
0
20
40
60
80
I (pA
)
a
b c
40 pA
100 ms
+100 mV
0 mV
ChTxControl
**
**
**
Control ChTx Control ChTx
Fig. 3 Effect of ChTx on BK channels in outside-out patches. a,upper panels. Representative traces of unitary BK channel currentsfrom an outside-out membrane patch depolarised every 3 s to+100 mV from a Vh = 0 mV in the absence (Control) and the presenceof ChTx (ChTx, 150 nM, 10 min). Notice that ChTx did not fullyinhibited BK channel activity; lower panels. Ensemble averages of 30sweeps from the same patch used in a. b Effect of ChTx on peakamplitude of the ensemble averages from six excised patches. c Effectof ChTx on the rate of inactivation (ki) of the ensemble averages. Dataare means±SEM. **p<0.01
8
10200 ms
1 nA
0 mV
+120 mV
6
4
2
012010080604020
** ** ****
**
**
400
200
0604020
25
20
15
10
5
0
12010080604020
ControlControl-ChTx (BK)
k i(s
-1)
Control
ChTx
Control-ChTx
V (mV)
V (mV)
I (nA
)
ControlChTx
a
c
b
Fig. 2 BK currents from bovine chromaffin cells at minimal Cai2+. a
K+ currents activated by 600 ms-long step depolarisations from a Vh =0 mV to different potentials ranging from +20 up to +120 mV, in20 mV increments, in the absence (Control, upper panel) and thepresence of ChTx (ChTx, 150 nM, 5 min; middle panel); lower panel.BK currents resulting from the subtraction of traces in the presence ofChTx from control ones (Control–ChTx). b Effect of ChTx on peakK+ currents at different potentials. Amplitudes were measured 15 msafter the beginning of the pulse. An expanded view of the graph isshown as an inset. c Voltage dependence of the rates of inactivation(ki) of control and BK currents. Data are means±SEM of 11experiments. **p<0.01 with respect to control values
550 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

Voltage dependence of K+ currents activation at minimal Cai2+
Tail current measurements using symmetrical K+ solutions(140 mM) were performed to study the voltage dependenceof K+ currents activation from bovine chromaffin cells. Toisolate BK currents, cells were clamped at 0 mV andsubjected to 10 ms step depolarisations to increasingvoltages from +35 to +215 mV by steps of 15 mV (Suppl.
Fig. 3A, left panel). A Boltzmann function fit to tail currentamplitudes on return to −70 mV gave a V50 of +146.8 and aK of 31.7 mV (Suppl. Fig. 3A, right panel). To obtain theactivation curve of IA, tail currents were generated in thepresence of IbTx (150 nM), another specific blocker of BKcurrents [21]. The current resistant to IbTx should mainlycorrespond to IA because delayed rectifier K+ channelsactivate too slowly to significantly contribute to whole-
0 1 2 3 4 50
1
2
3
4BK
non-BK
time (ms)
2 ms
10 pAVh= -70 mV Vh= 0 mVBK non-BK
a
b
Fig. 4 Fast activation of BKchannels at minimal Cai
2+ inoutside-out patches. a left panel.Representative current traces(15 ms-long) of multichannelactivity in an outside-out patchdepolarised to +20 mV from aVh = −70 mV. BK channels areeasily distinguishable amongother K+ channels due to theirlarge conductance; middle pan-el. Idealised BK channel open-ings and non-BK channelactivity (resulting from the sub-traction of idealised BK channelopenings from the original cur-rent traces; see “Methods”) dur-ing the first 5 ms of the stepdepolarisation; right panel.Recordings of channel activityfrom the same representativepatch held at a 0 mV. Notice thatmost of non-BK channels areinactivated at this Vh. b Ensem-ble averages (20–30 sweeps) foridealised (BK) and subtracted(non-BK) current traces from sixdifferent patches during the first5 ms of depolarisation. An ex-ponential function was fitted toaverage data from each class ofchannels (τBK=0.55 ms andτnon-BK=0.69 ms)
Pflugers Arch - Eur J Physiol (2011) 462:545–557 551
Author's personal copy

cell K+ currents during such a brief depolarisations [29,39]. IbTx reduced by about 60% the amplitude of tailcurrents (Suppl. Fig. 3B, left panel), in good agreementwith the effect of ChTx in excised patches and whole-cellcurrents in asymmetric K+ (Fig. 1). Fitting a Boltzmannfunction to tail current amplitudes in the presence of IbTxgave a V50 of −14.2 mV, and a K of 11.9 mV (Suppl. Fig. 3B,right panel), in the range of typical A-type K+ currents [26].
Voltage dependence of K+ currents inactivationat minimal Cai
2+
To study the voltage dependence of BK current inactiva-tion, we held the cells at 0 mV and measured the K+
currents at +105 mVafter a pre-pulse of variable amplitudethat was long enough to induce steady-state inactivation ofBK channels (600 ms). Pre-pulse amplitudes were from +15to +105 mV, in 15 mV step increments (Fig. 5a). Only theinactivating component of the current elicited by the testpulse was considered to construct the steady-state inactiva-tion curve. A Boltzmann function fitted to the data gave aV50 of +45.2 mVand a K of −15.5 mV (Fig. 5c), in goodagreement with previously reported values of steady-stateBK channel inactivation in rat chromaffin cells at minimalCai
2+ [9]. Importantly, the little steady-state inactivation ofBK channels at 0 mV validate the use of this potential as Vh
to isolate BK currents. The steady-state inactivation of IAwasstudied in the presence of ChTx (150 nM) by usingdepolarising pre-pulses of increasing voltages from Vh=−90to +15 mV, followed by a test pulse to +20 mV (Fig. 5b). ABoltzmann function with V50 of −42.8 and K of −7.1 mVwas fitted to the experimental data (Fig. 5c).
The time course of recovery from inactivation of IA and BKcurrent was also studied (Suppl. Fig. 4). BK currents wereelicited by 600 ms-long variably spaced paired pulses to+100 mV from a Vh = 0 mV. Ratios between peak currentresponses to pulse pairs were plotted against intervalsbetween pulses. A double exponential function fitted to thedata gave two rate constants (kfast=23 s−1 and kslow=0.77 s−1), reflecting two different kinetic components withsimilar amplitudes (Suppl. Fig. 4A). The recovery frominactivation of IAwas characterised by using paired pulsesto +20 mV from Vh=−70 mV and was also described by adouble exponential function with a kfast=27 s−1 accountingfor 85% of the recovery and kslow=1.84 s−1 (Suppl. Fig. 4B).
BK channels contribute to shape APs in the absenceof extracellular Ca2+
Our results show that BK channels activated during step-like pulses to +20 mV contribute by about 25% to fast K+
outward current at minimal Cai2+. We therefore investigated
BK current activation during APWs in the nominal absence
of extracellular Ca2+ (Cao2+). APs were recorded under
perforated-patch conditions and then used as APWs tostimulate chromaffin cells. APWs activated outwardlydirected currents that reached their maximal amplitude nearthe peak of the AP (Fig. 6). These currents were partiallysensitive to either IbTx (150 nM) or paxilline (1 μM). Withrespect to total K+ current, the peak amplitude of the currentcomponent sensitive to IbTx and paxilline was 36.8±
+105 mV
0 mV
a
c
1 nA
200 ms
-100 -70 -40 -10 20 50 80 110 140
0.00
0.25
0.50
0.75
1.00 IAIBK
I nor
mal
ized
V (mV)
b
-90 mV
+15 mV+20 mV
200 pA
200 ms
Fig. 5 Steady-state inactivation of IA and BK current at minimal Cai2+.
a BK currents were activated by 600 ms-long conditioning pulses todifferent potentials from a Vh = 0 mV, which were immediately followedby 600 ms pulses to +105 mV (see adjoining protocol). b A similarstimulation paradigm was used to study inactivation of IA (Vh =−90 mV; test pulse to +20 mV; see adjoining protocol). c Steady-stateinactivation curves of IA and BK current. Only the inactivatingcomponent of the current responses to the second pulse was considered.Data were normalised with respect to currents elicited by test pulses to+20 or +105 mVapplied from the corresponding Vh (0 or −90 mV).Boltzmann functions were fitted to the experimental data and gave thefollowing values: V50=+45.25 mVand K=−15.55 mV for BK current,and V50=−42.83 mVand K=−7.13 mV for IA. Data are means±SEM ofseven cells for each type of current
552 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

4.6% (n=14 cells) and 44.4±5.2% (n=6 cells), respec-tively, thus suggesting that BK currents are largelyactivated during APs in the absence of Cao
2+ (Fig. 6a,b). Interestingly, the current component sensitive to BKchannel blockers was an early one as expected from thefast activation of BK channels as compared with other K+
channels expressed in chromaffin cells.Next, we investigated whether BK channels play any role
in controlling the shape of electrically induced APs in theabsence of Cao
2+. For comparison purposes, we resolved firstto record APs under perforated-patch conditions both in thepresence (2 mM) and the absence of Cao
2+ (Fig. 7a).Compared with control conditions (2 mM Cao
2+), APs inthe absence of Cao
2+ showed a slower repolarisation (half-amplitude duration, HAD, of 4.34±0.19 ms in the presenceof Cao
2+ and 6.46±0.37 ms in the absence of Cao2+; n=23
and 18 cells, respectively, p<0.0001). Such a difference wasparticularly evident during the second half of the depolarisa-tion presumably because the absence of the inward Ca2+
currents during late stages of the AP diminished theactivation of BK channels (Suppl. Fig. 5B). Treatment withIbTx (150 nM) or paxilline (1 μM) in the absence of Cao
2+
produced a delay in repolarization immediately after the APpeak, thus giving rise to APs with longer HADs (5.55±
0.68 ms before and 6.77±2.3 ms after IbTx; n=7 cells, p<0.05; 7.03±0.34 ms before and 8.1±0.39 ms after paxilline;n=11 cells, p<0.01). The effect of BK channel blockers onAP duration was nevertheless more pronounced in thepresence of Cao
2+ (3.85±0.24 ms before and 5.25±0.35 msafter IbTx; n=12 cells, p<0.001; 4.87±0.21 ms before and8.23±0.41 ms after paxilline; n=11 cells, p<0.0001; Fig. 7a,b). We did not observe any consistent effect of either IbTx orpaxilline on AP peak. IbTx reduced AP peak in cells bathedin the absence of Cao
2+ (29.09±5.21 mV before and 23.76±5.52 mV after IbTx; n=7 cells, p<0.05) whereas paxillineproduced the opposite effect but only in cells superfusedwith a Ca2+-containing solution (40.97±2.23 mV before and47.51±3.95 mV after paxilline; n=11 cells, p<0.05). Webelieve these effects are not related to actions on BKchannels and are most likely attributable to changes inresting membrane potential and Na+ channel availability[27]. Altogether, these data point to a marked contribution ofBK channels to AP repolarisation both in the presence andthe absence of Ca2+ entry. Indeed, data from Pancrazio et al.suggested that a significant fraction of BK current cellsactivates before the onset of Ca2+ current during an APW inbovine chromaffin cells [35]. Our results using APWs tostimulate the cells in the presence of 2 mM Cao
2+ are fully
10 ms
50 mV
400 pA
400 pA 200 pA
200 pA
APW Control IbTXControl-IbTx (BK)
APW Control PaxillineControl-Paxilline (BK)
a b Fig. 6 Effect of IbTx andpaxilline on K+ currents inducedby action potential waveforms inthe absence of Cao
2+. a Upperpanel. Perforated-patch record-ings of K+ currents (black trace)induced by a fast action poten-tial waveform (APW; dashedline) from a chromaffin cellbathed in a free Ca2+ solution.Blue trace depicts the currentremaining after the administra-tion of IbTx (150 nm, 5 min).Red trace results from subtract-ing the blue trace (in the pres-ence of IbTx) from the blackone (Control), hencecorresponding to the BK cur-rent; lower panel. Effect of IbTxon K+ currents evoked by abroader APW from the samecell. Dotted lines denote the0 mV level. b. Similar to a, butshowing representative resultsobtained with paxilline (1 μM,10 min). Notice that BK channelcurrents activate early during theAPW irrespective of APduration
Pflugers Arch - Eur J Physiol (2011) 462:545–557 553
Author's personal copy

consistent with this (Suppl. Fig. 5A). Under these conditions,APWs-activated K+ currents often exhibited two separatedcomponents. Furthermore, treatment with BK channel block-ers allowed identifying also two peaks in the BK channel-mediated current: an early peak occurring during the initialupswing of the APW and a delayed one that coincided intime with the surge of Ca2+ entry (Suppl. Fig. 5A, B).Interestingly, the relative size of this second phase of BKcurrent increases as AP duration increases with the ensuedprolongation of Ca2+ entry and further accumulation of Cai
2+.These results additionally support the notion that during an
AP some BK channels could be activated even before Ca2+
entering the cell drastically increases their opening probability.
Discussion
In the present study, we have characterised voltage-gated K+
currents from cultured bovine chromaffin cells when bothsides of their plasma membranes were exposed to minimalCa2+ concentrations (no added external Ca2+ and Cai
2+
buffered to about 10 nM). Under these conditions, the
a
b
Fig. 7 Effect of IbTx on actionpotentials in the presence andthe absence of Cao
2+. a Upperpanel. Effect of IbTx (150 nM,5 min) on action potentialsinduced by a brief constant-current injection (20 pA, 20 ms)in chromaffin cells in the pres-ence (2 mM) or the nominalabsence of Cao
2+; lower panel.Depict similar experiments asshown in upper panel but paxil-line (1 μM, 10 min) instead ofIbTx was employed. AP record-ings were performed underperforated-parch conditions.Notice that both IbTx and pax-illine widens the AP by delayingrepolarisation. Dotted lines de-note the 0 mV level. b Summaryof the effects of IbTx and paxil-line on AP half-amplitude dura-tion (HAD). Data are expressedas percent change with respectto control values in the absenceof BK channel blockers. Dataare means±SEM of results col-lected from the number of cellsspecified within parenthesis. *p<0.05, **p<0.01, ***p<0.001
554 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

occupancy of Ca2+-binding sites responsible for the activa-tion of BK channels is thought to be negligible [20]. Twomain conclusions arise from our results: (1) like manyneurones, chromaffin cells are endowed with fast-inactivating IA-like K+ currents, (2) even at minimal Cai,
2+
BK current significantly contribute to the K+ efflux evokedby voltage depolarisations to physiological potentials (from0 to +40 mV).
The transient, IA-type, K+ current
A transient outward, IA-type, K+ current activating between
−70 and +30 mV was apparent in the presence of ChTx andnanomolar Cai
2+. This current might have previouslypassed unnoticed in whole-cell recordings because underphysiological Ca2+ concentrations BK currents could havemasked it. IA from bovine chromaffin cells behaves as arapidly inactivating current with a τ of about 25 ms in thevoltage range from 0 to +60 mV (Fig. 1c). Furthermore,recovery from inactivation is also fast, taking about 100 msto develop by 80%. IA had a moderate sensitivity to TEAand was resistant to high concentrations (150 nM) of ChTx.The ion channel responsible for IA could correspond to theinactivating FK channel of 18 pS identified by Marty andNeher [29]. Similarly to the transient current describedhere, FK current maximally activates between +20 and+40 mV and becomes inactivated in about 60 ms. Atvariance, the channel underlying IA would differ from thefast-inactivating but TEA-resistant (40 mM) channel clonedby Garcia-Guzman et al. from a bovine chromaffin cellcDNA library [13].
IA may contribute to the regulation of chromaffin cellexcitability during trains of APs. Due to the partial overlap(from −65 to −30 mV) of the voltage ranges for activation andsteady-state inactivation, IA would be steadily activatedbetween consecutive APs, thus acting as a damper on thedevelopment of interspike depolarisations and thereby reduc-ing the frequency of firing. This frequency-encoding role isplayed also because of the inactivation properties of IA, whichinactivates during the spike depolarisation but quicklyrecovers over the hyperpolarisation phase that immediatelyfollows each AP. IA can be of importance to chromaffin cellphysiology not only by determining AP firing rates in a waythat reflects the stimulus intensity (e.g., acetylcholine outputfrom splanchnic nerve terminals) but also by imposing anupper frequency limit so that the exhaustion of the readilyreleasable pool of secretory vesicles is prevented [17, 49].
Physiological relevance of BK channels in the absenceof Ca2+ entry
The high ionic conductance of BK channels (∼190 pS) andtheir high density in the membrane of bovine chromaffin
cells (550 channels per cell; [29]) can explain thesignificant fraction of K+ efflux carried by BK channels atminimal Cai
2+. Hence, despite the low open probability ofany individual BK channel under those conditions, the largenumber of BK channels expressed in a typical chromaffincell together with their large single-channel current atpositive potentials makes it possible that whole-cell BKcurrents of appreciable magnitude are effectively activated.
Throughout this study, we have used a Vh=0 mV toisolate BK currents. A number of experimental evidencessupport the suitability of this Vh for that purpose: (1) at thatpotential, only two out of 50 outside-out membrane patchesshowed unitary currents arising from channels different tothe BK ones; moreover, these other channels contributedminimally to the ensemble average current in those twopatches; (2) the relative density of ChTx-resistant BKs
channels in outside-out patches was similar to the fractionalcontribution of ChTx-resistant BKs current to the whole-cell outward K+ current (Figs. 2 and 3), thus suggesting thatBK channels (BKi+BKs) underlie most of the outwardcurrent at the whole-cell level; (3) paxilline, a BK channelblocker that inhibits both BKi and BKs channels in rat andmouse chromaffin cells [27, 40, 51], abolished whole-cellK+ currents activated from Vh=0 mV (Suppl. Fig. 1); (4)although IbTx eliminates a noticeable fraction of K+
currents elicited by voltage steps from −70 to 0 mV (Suppl.Fig. 2B), the toxin-sensitive component represents onlyabout 2% of the maximal BK current available from Vh=0 mV (Suppl. Fig. 2A, B).
The voltage-dependence of BK current activation variesamong cell types, presumably due to the expression ofsplice variants of pore-forming α subunits and a variety ofregulatory β subunits [6, 54, 55]. So, α and β subunitsfrom human myometrium cells coexpressed in X. oocytesshow a V50 of +220 mV when Cai
2+ varies from 10 to100 nM [50]; in contrast, BK channels in rat supraopticneurones show a V50 of +80 mVat a similar Cai
2+ [44]. BKcurrents from bovine chromaffin cells activate with a V50
of +147 mV in 10 nM Cai2+ (Suppl. Fig. 2A). This value is
similar to that reported for BK channels from ratchromaffin cells (V50 of +135 mV; [9]), which are knownto express α as well as β2 and β3 subunits [53, 55, 56].
Our results also indicate that BK channels largelycontribute to the total K+ efflux during the first millisecondof a square pulse to physiological potentials (+20 mV) atminimal Cai
2+. Moreover, we have shown that BK channelsactivate significantly during APs and shorten their durationin the absence of Ca2+ entry. This raises the possibility of asizeably voltage-dependent action of these channels in theinitial AP repolarization also under conditions of physiolog-ical Ca2+. The time window required for a strictly voltage-dependent BK channel-mediated modulation of membranepotential during an AP in the presence of Cao
2+ starts during
Pflugers Arch - Eur J Physiol (2011) 462:545–557 555
Author's personal copy

the rapid depolarising phase (phase 0) of the AP and isexpected to end by the time that Ca2+ enters through voltage-activated Ca2+ channels and reaches its binding sites on BKchannels. In bovine chromaffin cells, Ca2+ currents elicitedunder voltage-clamp by pre-recorded APW starts to activatesoon after the peak of the AP (Suppl. Fig. 5B) [35], as insome neurons [3, 23]. The fact that virtually everymembrane patch from chromaffin cells contains BK channelssuggests that these channels are distributed rather homoge-neously along the cell surface [36]. Additionally, although ithas been found that BK channels and Ca2+ channels are co-localised in the frog neuromuscular junction [37] and thecalyx synapse from the chick ciliary ganglion [43], inchromaffin cells, Ca2+ channels and BK channels are weaklyco-localised (mean distance between channels >160 nm) [36]except for the Cav1.3 L-type and fast-inactivating BKchannels of mouse chromaffin cells, which are more tightlycoupled (mean distance between channels 50 and 160 nm)[27]. The latter consideration, however, might not applydirectly to bovine chromaffin cells, which express amuch smaller proportion of BKi channels (∼18% inbovine vs. ∼58% in the rat) [25] and of L-type Ca2+
channels (∼15% of whole-cell Ca2+ current in bovine vs.45–50% in mouse and rat) [12] than their rodent counter-parts. Hence, in the case of a single action potentialoriginating from a non-spontaneously active bovinechromaffin cell, it is unlikely that most BK channels couldinitially sense Ca2+ ions entering the cell if only a few Ca2+
channels are opened before the peak of the AP and Ca2+ ionsdiffusing from a Ca2+ channel barely reach a few hundredsof nanomolars at the inner side of BK channels [33, 34]. Ourexperimental data support the notion of a two-phaseactivation of BK currents during APs under physiologicalCao
2+ conditions. The initial phase of activation would occurin an essentially Ca2+-independent manner and would resultfrom both voltage depolarisation during the upswing of APand the concomitant increase in driving force for K+ outflow.The second, delayed phase of activation, would be aconsequence of the increased Cai
2+ brought about by Ca2+
entry during the late stages of AP, which will drasticallypromote K+ efflux despite the progressive repolarization. Asa major conclusion, our results suggest that BK channelsmay indeed influence the shape of AP without the need oflarge depolarisations or micromolar changes in Cai
2+ andpropose a role for these channels on AP early repolarisationbefore AP-elicited Ca2+ entry takes place, especially atsubregions of the membrane where BK channels are locatedfar from Ca2+ channels [27]. This would occur particularlyduring single or low frequency (<0.1–1 Hz) AP firing in cellsheld at rather negative resting potentials, while contribution ofmicromolar Ca2+ would be determinant at higher frequency(3–10 Hz) or in spontaneously firing and more depolarizedchromaffin cells. Under these conditions, Ca2+-entry during
interspikes would nevertheless be sufficient to activate anadequate fraction of BK channels that contribute to the APamplitude, width and after-hyperpolarisation.
Acknowledgments We thank Dr. Antonio G. García for thegenerous supply of chromaffin cell cultures, and Drs. Paola Imbriciand Arnaud Ruiz for their valuable comments on the manuscript.Useful discussions with Drs. Juan Antonio Gilabert and Guido Ulateare also greatly acknowledged. This work was supported by grantsfrom the Spanish Ministerio de Ciencia e Innovación (BFU2005-06034; Spanish Ion Channel Initiative (SICI) grant CSD2008-00005)to A.R.A. and (RYC-2009-03979 and SAF2010-20604) to R.S.S.
References
1. Artalejo AR (1995) Electrical properties of adrenal chromaffincells. In: H. Scherübl and Hescheler J (ed) The electrophysiologyof neuroendocrine cells. CRC Press Inc, pp. 259–299.
2. Artalejo AR, Garcia AG, Neher E (1993) Small-conductanceCa(2+)-activated K+ channels in bovine chromaffin cells.Pflügers Arch 423:97–103
3. Borst JG, Sakmann B (1998) Calcium current during a singleaction potential in a large presynaptic terminal of the ratbrainstem. J Physiol 506:143–157
4. Butler A, Tsunoda S, McCobb DP, Wei A, Salkoff L (1993) mSlo,a complex mouse gene encoding “maxi” calcium-activatedpotassium channels. Science 261:221–224
5. Cannon SD, Wilson SP, Walsh KB (1994) A G protein-activatedK+ current in bovine adrenal chromaffin cells: possible regulatoryrole in exocytosis. Mol Pharmacol 45:109–116
6. Chang CP, Dworetzky SI, Wang J, Goldstein ME (1997)Differential expression of the alpha and beta subunits of thelarge-conductance calcium-activated potassium channel: implica-tion for channel diversity. Brain Res Mol Brain Res 45:33–40
7. Coetzee WA, Amarillo Y, Chiu J, Chow A, Lau D, McCormack T,Moreno H, Nadal MS, Ozaita A, Pountney D, Saganich M, deVega-Saenz ME, Rudy B (1999) Molecular diversity of K+
channels. Ann N YAcad Sci 868:233–2858. Connor JA (1980) The fast K+ channel and repetitive firing. In:
Neurosciences CSHRit (ed) Molluscan nerve cells: From biophysicsto behaviour, pp. 126–133. Cold Spring Harbor Laboratory Press,Woodbury NY
9. Ding JP, Lingle CJ (2002) Steady-state and closed-state inactivationproperties of inactivating BK channels. Biophys J 82:2448–2465
10. Ding JP, Li ZW, Lingle CJ (1998) Inactivating BK channels in ratchromaffin cells may arise from heteromultimeric assembly ofdistinct inactivation-competent and noninactivating subunits.Biophys J 74:268–289
11. Dopico AM, Widmer H, Wang G, Lemos JR, Treistman SN (1999)Rat supraoptic magnocellular neurones show distinct largeconductance, Ca2+-activated K+ channel subtypes in cell bodiesversus nerve endings. J Physiol 519:101–114
12. Garcia AG, Garcia-De-Diego AM, Gandia L, Borges R, Garcia-Sancho J (2006) Calcium signaling and exocytosis in adrenalchromaffin cells. Physiol Rev 86:1093–1131
13. Garcia-Guzman M, Calvo S, Cena V, Criado M (1992) Molecularcloning and permanent expression in a neuroblastoma cell line ofa fast inactivating potassium channel from bovine adrenalmedulla. FEBS Lett 308:283–289
14. Ghatta S, Nimmagadda D, Xu X, O’Rourke ST (2006) Large-conductance, calcium-activated potassium channels: structural andfunctional implications. Pharmacol Ther 110:103–116
556 Pflugers Arch - Eur J Physiol (2011) 462:545–557
Author's personal copy

15. Gonzalez-Garcia C, Ceña V, Keiser HR, Rojas E (1993) Catechol-amine secretion induced by tetraethylammonium from culturedbovine adrenal chromaffin cells. Biochim Biophys Acta 1177:99–105
16. Hanner M, Schmalhofer WA, Munujos P, Knaus HG, KaczorowskiGJ, Garcia ML (1997) The beta subunit of the high-conductancecalcium-activated potassium channel contributes to the high-affinityreceptor for charybdotoxin. Proc Natl Acad Sci USA 94:2853–2858
17. Heinemann C, von Rüden L, Chow RH, Neher E (1993) A two-step model of secretion control in neuroendocrine cells. PflügersArch 424:105–112
18. Horn R (1991) Estimating the number of channels in patchrecordings. Biophys J 60:433–439
19. Horrigan FT, Aldrich RW (2002) Coupling between voltage sensoractivation, Ca2+ binding and channel opening in large conduc-tance (BK) potassium channels. J Gen Physiol 120:267–305
20. Horrigan FT, Cui J, Aldrich RW (1999) Allosteric voltage gatingof potassium channels I. Mslo ionic currents in the absence of Ca(2+). J Gen Physiol 114:277–304
21. Kaczorowski GJ, Garcia ML (1999) Pharmacology of voltage-gated and calcium-activated potassium channels. Curr Opin ChemBiol 3:448–458
22. Latorre R, Morera FJ, Zaelzer C (2010) Allosteric interactions andthe modular nature of the voltage- and Ca2+-activated (BK)channel. J Physiol 588:3141–3148
23. Llinas R, Sugimori M, Simon SM (1982) Transmission bypresynaptic spike-like depolarization in the squid giant synapse.Proc Natl Acad Sci USA 79:2415–2419
24. Lovell PV, McCobb DP (2001) Pituitary control of BK potassiumchannel function and intrinsic firing properties of adrenalchromaffin cells. J Neurosci 21:3429–3442
25. Lovell PV, James DG, McCobb DP (2000) Bovine versus ratadrenal chromaffin cells: big differences in BK potassium channelproperties. J Neurophysiol 83:3277–3286
26. Malin SA, Nerbonne JM (2001) Molecular heterogeneity of thevoltage-gated fast transient outward K+ current, I(Af), in mam-malian neurons. J Neurosci 21:8004–14
27. Marcantoni A, Vandael DH, Mahapatra S, Carabelli V, Sinnegger-Brauns MJ, Striessnig J, Carbone E (2010) Loss of Cav1.3channels reveals the critical role of L-type and BK channelcoupling in pacemaking mouse adrenal chromaffin cells. JNeurosci 30:491–504
28. Marty A (1981) Ca-dependent K channels with large unitaryconductance in chromaffin cell membranes. Nature 291:497–500
29. Marty A, Neher E (1985) Potassium channels in cultured bovineadrenal chromaffin cells. J Physiol 367:117–141
30. McManus OB, Helms LM, Pallanck L, Ganetzky B, Swanson R,Leonard RJ (1995) Functional role of the beta subunit of highconductance calcium-activated potassium channels. Neuron14:645–50
31. Meera P, Wallner M, Toro L (2000) A neuronal beta subunit(KCNMB4) makes the large conductance, voltage- and Ca2+ -activated K+ channel resistant to charybdotoxin and iberiotoxin.Proc Natl Acad Sci USA 97:5562–5567
32. Moro MA, Lopez MG, Gandia L, Michelena P, Garcia AG (1990)Separation and culture of living adrenaline- and noradrenaline-containing cells from bovine adrenal medullae. Anal Biochem185:243–248
33. Muller A, Kukley M, Uebachs M, Beck H, Dietrich D (2007)Nanodomains of single Ca2+ channels contribute to actionpotential repolarisation in cortical neurons. J Neurosci 27:483–95
34. Naraghi M, Neher E (1997) Linearized buffered Ca2+ diffusion inmicrodomains and its implications for calculation of [Ca2+] at themouth of a calcium channel. J Neurosci 17:6961–6973
35. Pancrazio JJ, Johnson PA, Lynch C 3rd (1994) A major role forcalcium-dependent potassium current in action potential repolar-isation in adrenal chromaffin cells. Brain Res 668:246–251
36. Prakriya M, Lingle CJ (2000) Activation of BK channels in ratchromaffin cells requires summation of Ca(2+) influx frommultiple Ca(2+) channels. J Neurophysiol 84:1123–1135
37. Robitaille R, Garcia ML, Kaczorowski GJ, Charlton MP (1993)Functional colocalization of calcium and calcium-gated potassiumchannels in control of transmitter release. Neuron 11:645–55
38. Rudy B (1988) Diversity and ubiquity of K channels. Neurosci-ence 25:729–49
39. Sala S, Soria B (1991) Inactivation of Delayed Potassium Currentin Cultured Bovine Chromaffin Cells. Eur J Neurosci 3:462–472
40. Sanchez M, McManus OB (1996) Paxilline inhibition of thealpha-subunit of the high-conductance calcium-activated potassi-um channel. Neuropharmacology 35:963–968
41. Solaro CR, Lingle CJ (1992) Trypsin-sensitive, rapid inactivationof a calcium-activated potassium channel. Science 257:1694–1698
42. Solaro CR, Prakriya M, Ding JP, Lingle CJ (1995) Inactivatingand noninactivating Ca(2+)- and voltage-dependent K+ current inrat adrenal chromaffin cells. J Neurosci 15:6110–6123
43. Sun XP, Schlichter LC, Stanley EF (1999) Single-channelproperties of BK-type calcium-activated potassium channels at acholinergic presynaptic nerve terminal. J Physiol 518:639–651
44. Sun L, Xiong Y, Zeng X, Wu Y, Pan N, Lingle CJ, Qu A, Ding J(2009) Differential regulation of action potentials by inactivatingand noninactivating BK channels in rat adrenal chromaffin cells.Biophys J 97:1832–1842
45. Tanaka Y, Meera P, Song M, Knaus HG, Toro L (1997) Molecularconstituents of maxi KCa channels in human coronary smoothmuscle: predominant alpha+beta subunit complexes. J Physiol502:545–557
46. Torres YP, Morera FJ, Carvacho I, Latorre R (2007) A marriage ofconvenience: beta-subunits and voltage-dependent K+ channels. JBiol Chem 282:24485–24489
47. Twitchell WA, Rane SG (1993) Opioid peptide modulation of Ca(2+)-dependent K+ and voltage-activated Ca2+ currents in bovine adrenalchromaffin cells. Neuron 10:701–709
48. Uebele VN, Lagrutta A, Wade T, Figueroa DJ, Liu Y, McKenna E,Austin CP, Bennett PB, Swanson R (2000) Cloning and functionalexpression of two families of beta-subunits of the large conductancecalcium-activated K+ channel. J Biol Chem 275:23211–23218
49. Ulate G, Scott SR, Gonzalez J, Gilabert JA, Artalejo AR (2000)Extracellular ATP regulates exocytosis in inhibiting multiple Ca(2+)channel types in bovine chromaffin cells. Pflügers Arch 439:304–314
50. Valverde MA, Rojas P, Amigo J, Cosmelli D, Orio P, BahamondeMI, Mann GE, Vergara C, Latorre R (1999) Acute activation ofMaxi-K channels (hSlo) by estradiol binding to the beta subunit.Science 285:1929–1931
51. Vandael DH, Marcantoni A, Mahapatra S, Caro A, Ruth P, ZuccottiA, Knipper M, Carbone E (2010) Ca(v)1.3 and BK channels fortiming and regulating cell firing. Mol Neurobiol 42:185–198
52. Wallace DJ, Chen C, Marley PD (2002) Histamine promotesexcitability in bovine adrenal chromaffin cells by inhibiting an M-current. J Physiol 540:921–939
53. WallnerM,Meera P, Toro L (1999)Molecular basis of fast inactivationin voltage and Ca2+-activated K+ channels: a transmembrane beta-subunit homolog. Proc Natl Acad Sci USA 96:4137–4142
54. Xia XM, Ding JP, Lingle CJ (1999) Molecular basis for theinactivation of Ca2+- and voltage-dependent BK channels inadrenal chromaffin cells and rat insulinoma tumor cells. JNeurosci 19:5255–5264
55. Xia XM, Ding JP, Zeng XH, Duan KL, Lingle CJ (2000)Rectification and rapid activation at low Ca2+ of Ca2+-activated,voltage-dependent BK currents: consequences of rapid inactiva-tion by a novel beta subunit. J Neurosci 20:4890–4903
56. Zeng XH, Ding JP, Xia XM, Lingle CJ (2001) Gating propertiesconferred on BK channels by the beta3b auxiliary subunit in theabsence of its NH(2)- and COOH termini. J Gen Physiol 117:607–628
Pflugers Arch - Eur J Physiol (2011) 462:545–557 557
Author's personal copy




















