
Blockade by nanomolar resveratrol of quantal catecholamine release in chromaffin cells
Upload
independentCategory
view
1download
0
Cathepsin L Colocalizes with Chromogranin A inChromaffin Vesicles to Generate Active Peptides
Nilima Biswas, Juan L. Rodriguez-Flores, Maite Courel, Jiaur R. Gayen,Sucheta M. Vaingankar, Manjula Mahata, Justin W. Torpey, Laurent Taupenot,Daniel T. O’Connor, and Sushil K. Mahata
Departments of Medicine (N.B., J.L.R.-F., M.C., J.R.G., S.M.V., M.M., L.T., D.T.O., S.K.M.), Chemistry and Biochemistry(J.W.T.), and Molecular Genetics (D.T.O.), University of California, San Diego, and Veterans Affairs San DiegoHealthcare System (D.T.O., S.K.M.), La Jolla, California 92093-0838
Chromogranin A (CgA), the major soluble protein in chromaffin granules, is proteolytically pro-cessed to generate biologically active peptides including the catecholamine release inhibitorypeptide catestatin. Here we sought to determine whether cysteine protease cathepsin L (CTSL), anovel enzyme for proteolytic processing of neuropeptides, acts like the well-established serineproteases [prohormone convertase (PC)1/3 or PC2] to generate catestatin by proteolytic processingof CgA. We found that endogenous CTSL colocalizes with CgA in the secretory vesicles of primaryrat chromaffin cells. Transfection of PC12 cells with an expression plasmid encoding CTSL directedexpression of CTSL toward secretory vesicles. Deconvolution fluorescence microscopy suggestedgreater colocalization of CTSL with CgA than the lysosomal marker LGP110. The overexpression ofCTSL in PC12 cells caused cleavage of full-length CgA. CTSL also cleaved CgA in vitro, in time- anddose-dependent fashion, and specificity of the process was documented through E64 (thiol re-agent) inhibition. Mass spectrometry on CTSL-digested recombinant CgA identified a catestatin-region peptide, corresponding to CgA360–373. The pool of peptides generated from the CTSL cleav-age of CgA inhibited nicotine-induced catecholamine secretion from PC12 cells. CTSL processing inthe catestatin region was diminished by naturally occurring catestatin variants, especiallyPro370Leu and Gly364Ser. Among the CTSL-generated peptides, a subset matched those found inthe catestatin region in vivo. These findings indicate that CgA can be a substrate for the cysteineprotease CTSL both in vitro and in cella, and their colocalization within chromaffin granulesin cella suggests the likelihood of an enzyme/substrate relationship in vivo. (Endocrinology150: 3547–3557, 2009)
Chromogranin A (CgA), a 48-kDa acidic glycoprotein, is themajor soluble protein in the catecholamine storage vesicles
and widely distributed in the endocrine cells and neurons (1–3).It is costored and coreleased with catecholamines, neuropep-tides, and ATP by exocytosis from chromaffin cells of the adrenalmedulla as well as sympathetic neurons (4, 5). It controls bio-genesis of secretory granules by targeting secretory proteins tothe granules of regulated pathway (6–9). This glycoprotein alsoserves as a precursor of several biologically active peptides thatare released on exocytotic secretion, such as pancreastatin (hu-man CgA250–301) (10, 11), vasostatin (human CgA1–76) (12),and catecholamine-release inhibitory peptide catestatin (human
CgA352–372) (13–16). During stress, the latter regulates catechol-amine secretion through the autocrine-paracrine-negative feed-back mechanism.
Although CgA is overexpressed in human essential (heredi-tary) hypertension, the plasma concentration of catestatin de-creased in not only the established cases of hypertension but alsonormotensive subjects with a family history of hypertension (17,18). Consistent with human findings, we found that mouse chro-mogranin A gene (Chga) knockout mice are hypertensive andhyperadrenergic (19). Thus, a better understanding of the pro-cessing of catestatin fragment from CgA may provide clues to thedevelopment of hypertension through adrenergic pathway.
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2009 by The Endocrine Societydoi: 10.1210/en.2008-1613 Received November 17, 2008. Accepted April 7, 2009.First Published Online April 16, 2009
Abbreviations: CgA, Chromogranin A protein; CTSL, cathepsin L; LC, liquid chromatog-raphy; MALDI, matrix-assisted laser desorption ionization; MS/MS, tandem mass spec-trometry; m/z, mass to charge ratio; PC, prohormone convertase; TBS-T, tris-buffered salinewith 0.1% Tween 20; TOF, time of flight; WT, wild type.
G E N E R A L E N D O C R I N O L O G Y
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3547
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

Like the other prohormones, CgA contains eight to 10 dibasicresidue sites, which are considered as potential sites for proteo-lytic cleavage (1). Several groups have examined the role of theserine prohormone convertase (PC) in the processing of chro-mogranin/secretogranin family of proteins and neuropeptidesfor generation of bioactive peptides (20–27). Besides the well-established subtilisin-like serine proteases, we reported previ-ously that in vitro digestion of recombinant CgA with anotherserine protease plasmin also generated a 14-amino acid catesta-tin region-specific peptide (28–30) with an IC50 of about 2–3�M. However, the protease(s) responsible for generation of thefull-length 21-mer are yet to be established. The cysteine proteasecathepsin L (CTSL) has recently been implicated as a novel en-zyme for proteolytic processing of neuropeptides (31–33). Tothis end, CTSL has been identified as the cysteine protease com-ponent of purified prohormone thiol protease in the secretoryvesicle (31–34). The function of CTSL in the secretory vesiclesdiffers from that in the lysosomes for protein degradation (35).Several groups reported various roles of CTSL in the secretoryvesicles (36–40). CTSL has also been implicated in multiplephysiological and pathological processes (41–46). In the presentstudy, we characterized the in vitro processing of catestatin re-gion from CgA by CTSL. The findings indicate that CTSL islocalized within chromaffin granules/vesicles and can generateactive catestatin-region fragments by proteolytic cleavage ofCgA. The results may have implications for the development ofhypertension through modulation of catecholamine secretion.
Materials and Methods
Overexpression and purification of recombinantproteins
Recombinant human CgA isoforms, including naturally occurring non-synonymous amino acid replacement variants (Gly364Ser, Pro370Leu,Arg374Gln), were expressed in Escherichia coli and purified as describedpreviously using a C-terminal 6-His affinity tag (30).
Immunoblot analysisThe proteins were separated in a 10% SDS-PAGE (Novex precast gel;
Invitrogen, San Diego, CA) or a 10–20% tricine gel. Rabbit polyclonalantihuman catestatin [1:3000 in tris-buffered saline with 0.1% Tween20 (TBS-T)], goat polyclonal anti-CTSL (1:500 dilution, SC-6498;Santa Cruz Biotechnology, Santa Cruz, CA), and goat polyclonal anti-actin (1:500, SC-1615; Santa Cruz Biotechnology) were used. The mem-brane was incubated with secondary antibody, horseradish peroxidase-conjugated donkey antirabbit (1:3000 in TBS-T), and donkey antigoat(1:3000 in TBS-T) for 1 h. The membrane was then developed by theSupersignal west pico chemiluminescent substrate (Thermo Scientific,Rockford, IL) according to the manufacturer’s protocol.
Digestion of recombinant CgA by CTSLPurified CgA wild-type and variant proteins (10 �M final) were in-
cubated with CTSL (human liver, specific activity: 1.54 U/mg of protein;lot no. 035K2741; catalog no. C6854; Sigma-Aldrich, St. Louis, MO) in400 mM sodium acetate (pH 5.5) with 4 mM EDTA and 8 mM dithio-threitol at 30 C water bath for various periods of time. The reaction wasstopped by the addition of the epoxide (sulfhydryl protease) inhibitorE64 (10 �M).
Matrix-assisted laser desorption (MALDI)/ionizationtime-of-flight (TOF) mass spectrometry
The CTSL digests were acidified in 0.1% trifluoroacetic acid andwere purified by using Zip Tip (Millipore, Billencia, MA). Samples wereanalyzed by mass spectrometry as described previously (30).
Liquid chromatography (LC) and tandem massspectrometry (MS/MS) analysis
LC-MALDI was acquired on a Tempo LC MALDI spotter (AppliedBiosystems/MDS Sciex, Foster City, CA) using a Chromolith CapRodmonolithic capillary column (150 � 0.1 mm) (EMD; VWR Interna-tional, West Chester, PA). LC-MALDI spots were acquired at a rate of4 sec/spot from 8 to 16 min. A plate model and default calibration wereperformed for reflector-positive mode as well as a default calibration for2 kV-positive modes on the 4800 MALDI TOF/TOF analyzer (AppliedBiosystems). Reflector-positive spectra were obtained by acquiring 500shots from mass to charge ratio (m/z) 1000–4000 amu (atomic massunits). MS/MS spectra were acquired in 2 kV-positive modes with col-lision-induced dissociation (CID) on at resolution 200 full width at halfmaximum; for 3000 shots. The MS/MS data were analyzed by GlobalProteome server 3.0 (Applied Biosystems) and subjected to database searchusing Mascot 2.1.1 (www.matrixscience.com). Data were searched againstthe Swissprot database (ftp://ftp.ncbi.nlm.nih.gov/blast/db/FASTA/) con-taining 237,168 sequences, in which had been inserted the recombinantsequences of interest. The search results indicated that individual ion scoresgreater than 42 indicate identity or extensive homology (P � 0.05). Onlysignificant scores for the peptides defined by a Mascot probability analysis99% or greater confidence interval were listed.
Peptide synthesisPeptides were synthesized at a 10- to 100-�mol scale using 9-fluore-
nylmethoxycarbonyl protection chemistry and purified to greater than80% homogeneity by reverse-phase HPLC on a C18 column.
Secretagogue-stimulated catecholamine releaseNorepinephrine secretion was assayed in [3H]norepinephrine-la-
beled PC12 cells as described previously (47). The cells were treated withnicotine (60 �M) in secretion medium, either alone or in combinationwith recombinant CgA (1.5 �M), full length, and digested with CTSL.Digestion reaction (1 h) contained CgA (10 �M) and CTSL (0.3 �M).After digestion, peptides were purified by C18 cartridge (Waters, Milford,MA), lyophilized, and suspended in water. For a dose response, fiveascending doses (0.075, 0.15, 0.3, 0.9, and 1.5 �M), equivalent of di-gested CgA, were used. 3[H]norepinephrine in the supernatants and celllysates were measured by liquid scintillation counting, and the resultswere expressed as percent secretion: [amount released/(amount released� amount in cell lysate)] � 100. Net secretion is secretagogue-stimulatedrelease minus basal release.
Cell culture and transient transfectionRat pheochromocytoma PC12 cells were grown as described previously
(47). Cells were transfected with pcDNA3.1 (vector) or pcDNA3.1-pre-proCTSL (a kind gift from Dr. Vivian Hook, Department of Medicine,University of California, San Diego) by using SuperFect transfection re-agent as per the manufacturer’s protocol (QIAGEN, Valencia, CA).Transfected cells were cultured on poly-L-lysine-coated coverslips. Cellswere fixed at 48 h for immunofluorescence and were harvested at 72 hof posttransfection for protein analysis.
Primary culture of rat chromaffin cellsYoung female rats were killed to collect adrenal medulla. The med-
ullary tissues were digested following a published protocol (http://www.natureprotocols.com) in Hanks’ balanced salt solution containing col-lagenase type I, BSA, deoxyribonuclease I, and hyaluronidase I-S. Cells
3548 Biswas et al. CTSL in Generation of Catestatin Variants Endocrinology, August 2009, 150(8):3547–3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

were grown in DMEM with 10% fetal bovine serum and cultured for 3 dfor immunocytochemistry.
Immunofluorescence for subcellular localizationCells were washed with PBS and fixed with 2.5% paraformaldehyde
for 20 min at room temperature. Fixed cells were permeabilized for 10min with 0.5% Triton X-100 in PBS, incubated for 5 min with 150 mM
glycine in PBS, and exposed for 30 min to 5% BSA in PBS. Cells were thenincubated for 30 min at room temperature with either goat polyclonalanti-CTSL (1:100; Santa Cruz Biotechnology) and rabbit polyclonal an-ti-CgA (1:1500) (48) or goat polyclonal anti-CTSL (1:100; Santa CruzBiotechnology) and rabbit polyclonal LGP110 (1:100) (9) in PBS con-taining 2% BSA. Cells were subsequently washed and incubated for 30min with a Alexa Fluor 594-conjugated donkey antigoat IgG (1:350;Invitrogen) and Alexa Fluor 488-conjugated donkey antirabbit at 1:250;(Invitrogen) together with 1 �g/ml Hoechst 33342 in PBS containing 1%BSA. Coverslips were washed with PBS, mounted in SlowFade antifadekit (Invitrogen). Three-dimensional images were captured on a Delta-Vision deconvolution microscopy system operated by SoftWoRx soft-ware (Applied Precision, Issaquah, WA), using oil immersion objectives(�60, numerical aperture 1.4) as described previously (49).
Statistical analysisBiochemical data are expressed as the mean � SEM. Curve fitting was
accomplished in the program Kaleidagraph (Synergy Software, Reading,PA). Multiple comparisons were made using one-way ANOVA fol-lowed by Bonferroni’s post hoc test. Statistical significance was con-cluded at P � 0.05.
Results
Colocalization in vivo of endogenous CTSL with CgA inprimary rat chromaffin cells
To determine the functional interactions between CgA andCTSL, the intracellular distribution of these two proteins wereexamined in primary cultures of rat chromaffin cells (Fig. 1A).Consistent with the localization of secretory vesicle, deconvolu-tion microscopy revealed colocalization of CgA with CTSL asevidenced by discrete and punctuate immunostaining (Fig. 1A).
Overexpression of CTSL in PC12 cells and its effect onproteolytic processing of CgA
The subcellular localization of CTSL was examined in PC12cells transfected with an expression plasmid encoding prepro-CTSL (Fig. 1B). The cells were stained with either CgA and CTSL(Fig. 1B, upper panel) or CTSL and lysosomal type 1 integralmembrane glycoprotein LGP110 (Fig. 1B, lower panel). CTSLshowed colocalization with CgA, similar results in primary cul-tures of rat chromaffin cells. LGP110, as seen by green fluores-cence, showed very little overlap with CTSL. The estimated co-localization (by Pearson’s correlation coefficient, scaled from �1to �1) of CgA with CTSL in primary cultures of rat chromaffincells (Fig. 1A) and PC12 cells (Fig. 1B, upper panel) were 0.7690and 0.5972, respectively. The overlap distribution betweenCTSL (red) and LGP110 (green) was substantially lower, withPearson’s correlation coefficient of 0.1089 (Fig. 1B, lowerpanel). PC12 cells were transfected with either the vectorpcDNA3 or pcDNA3-prepro-CTSL to test the involvement ofCTSL in CgA processing. Overexpression of CTSL (Fig. 1C, leftpanel) reduced the levels of both full-length CgA and a 24-kDa
fragment (Fig. 1B, middle panel). A minor 40-kDa catestatin-positive fragment appeared in CTSL-overexpressing cells, com-pared with the control (Fig. 1B, middle panel). In parallel, purifiedrecombinant CgA (10 �M) was digested with various concentra-tions of CTSL (0.06, 0.03, and 0.015 �M) and subjected to immu-noblot with anticatestatin antibody for identification of catestatinregion-specific peptides (Fig. 1C, right panel). In the lane marked-Enz, recombinant CgA was incubated without the enzyme, andimmunoblotting clearly showed the position of undigested full-length CgA; with prolonged exposure, lower-molecular-weightCgA fragments were not observed in the absence of enzyme (resultsnot shown). On the other hand, CTSL-digested CgA samplesshowed five catestatin-positive fragments, besides undigested CgA(Fig. 1C, lanes 2–4). CgA underwent less digestion on exposure tolow protease concentration, as evidenced by the presence of morefull-length protein, shown in lanes 3 and 4, compared with lane 2(Fig. 1C). Catestatin antibody recognized a series of peptides rang-ing from 10 to 50 kDa (Fig. 1C, right panel). The 11-kDa fragmentwas observed only at lower enzyme concentrations (0.03 and 0.015�M; lanes 3 and 4, right panel), suggesting its further processing athigher protease concentrations. The other fragments of 22, 24, 34,and 40 kDa were prominent throughout all conditions. The frag-ments of 24 and 40 kDa were noticed in the transfected cells as wellas in the in vitro digestions (Fig. 1C, middle and right panel). Thus,CTSL generated catestatin immunopositive peptides in CTSL over-expressed PC12 cells that resemble the in vitro digestion of recom-binant CgA.
Dose- and time-dependent, specific cleavage of CgAin vitro
Purified recombinant CgA protein (10 �M) was incubated withvarying concentrations of CTSL (Fig. 2A, upper panel) to assess thedegree of proteolytic processing. CTSL readily digested the recom-binant CgA, and the extent of digestion was dose dependent. Themajor products produced were having apparent molecular massesranging from 8 to 10 kDa to 65 kDa. At 0.03 and 0.015 �M CTSL,digestionwaspartialasevidencedbythepresenceoffull-lengthCgArunning as a 70-kDa protein (Fig. 2A, lanes 6 and 7, upper panel).When recombinant CgA (10 �M) was digested with CTSL (0.15�M) for various periods of time (0–60 min), it suggests a time-dependentdegradationofCgAbyCTSL(Fig.2A, lowerpanel).Thesequentially produced major products have apparent molecularmasses of 10, 11, 12.5, 16, 18, 20, 27, 34, 43, 50, and 55 kDa asdetected by SDS-PAGE. Among the generated peptides, the frag-ments with 10–11 kDa were prominent at all the time points. CgAwas not processed to smaller peptides in either the absence of CTSL(Fig. 2A, lower panel, lane 9) or presence of a general cysteine pro-tease inhibitor E64 (Fig. 2A, lower panel, lane 8), documenting thespecificity of CTSL toward processing of CgA.
Identification of different catestatin region-specificpeptides generated from CTSL digestion of CgA
We have shown previously that the recombinant wild-type(WT) CgA is cleaved easily by plasmin, generating a very stable14-mer peptide (CgA360–373) (30). In the present study, we di-gested recombinant CgA protein (10 �M) with CTSL (0.3 �M for15 min) to verify whether CTSL can generate the catestatin re-
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3549
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

gion-specific peptide. The digestion products were subjected toLC-MS/MS analyses and the peptides generated in the catestatinregion, with greater than 99% confidence interval, are listed in
supplemental Table 1, published as supple-mental data on The Endocrine Society’sJournals Online web site at http://endo.endojournals.org. Thirteen peptides weregenerated from the N-terminal part of the ca-testatin region, ranging from CgA332–359,with a common stretch of sequence, viz.,QEEEEDNRDSSMK (supplemental Table1). In addition, the following peptides weregenerated by CTSL: nine peptides in the pan-creastatindomain,11peptides inthevasosta-tin region, nine peptides in the WE14 do-main, four peptides in the GE25 domain, andfour peptides in the chromacin region (sup-plemental Table 2, A–E).
Digestion of CgA was also attempted ata lower dose of CTSL (0.03 �M). MALDI-TOF analysis of the digests revealed thegeneration of internal catestatin frag-ments, CgA360–366, CgA367–374, CgA365–373,CgA358–366, CgA365–374, CgA360–373,CgA360–374, and CgA358–373 (Fig. 2B).Among those, peptides with mass 840, 1545and 1701 were also generated from plasmindigestion of CgA (30). We have shown pre-viously that the internal peptides CgA360–373
andCgA360–374 actasnicotinicantagonist tocatecholamine secretion with IC50 of 2.4 and3.0 �M, respectively (30). The generation ofthese active peptides in CTSL digestion im-plicatesCgAasasubstrateofCTSL.Thepep-tides with mass 1027 (CgA365–373), 1143(CgA358–366), 1183 (CgA365–374), and 1848(CgA358–373) were generated exclusivelywith CTSL digestion, suggesting additionalcleavage specificity for CTSL. Two pep-tides in the C-terminal part (CgA374–394,mass 2409, and CgA374–393, mass 2253)and one peptide in the N-terminal part(CgA336–359, mass 2883) of catestatin werealso generated. In the control experiments,when CTSL or CgA alone were incubated for15minat30C,nodetectablepeaksinm/zrange800-4000 were produced (results not shown).
Cleavage of synthetic catestatin-region peptides (CgA360–380 orCgA333–379) by CTSL
To explore the catestatin-region diges-tion pattern within CgA by CTSL, we usedtwo synthetic peptides as substrates: a short21-amino acid synthetic peptide, CgA360–380,with one dibasic site (Arg373Arg374) and alonger 47-amino acid synthetic peptide,
CgA333–379, which spans the entire catestatin region includingits two dibasic sites (Lys338Arg339 and Arg373Arg374). Eachpeptide (10 �M) was digested with CTSL and then subjected to
FIG. 1. In vivo role of CTSL for the generation of catestatin-related peptide. A, Subcellular localization ofendogenous CTSL and CgA in chromaffin granules or secretory vesicles of primary rat chromaffin cells. Cellswere examined by deconvolution microscopy. A series of xy optical sections along the z-axis was acquiredwith increments of 0.2 �m using �60 oil immersion objectives. The data set was processed to generatethree-dimensional (3D) or representative xy section. Colocalization of CgA (green) and CTSL (red) wasshown by overlay of the images, which is illustrated by yellow fluorescence. B, Intracellular distribution ofCgA and the endosomal/lysosomal marker LGP110. PC12 cells were transfected with pcDNA3.1-CTSL andprocessed for immunocytochemistry using rabbit polyclonal anti-CgA primary and antirabbit IgG-Alexa Fluor488-conjugated secondary (green fluorescence). CTSL was detected with goat polyclonal anti-CTSL andantigoat IgG-Alexa Fluor 594-conjugated secondary (red fluorescence; upper panel). In the lower panel, cellswere stained with rabbit polyclonal anti-Lgp110 (green) and goat polyclonal anti-CTSL (red). The nucleiwere visualized with Hoechst 33342 (blue). Scale bar, 5 �m. C, Overexpression of CTSL and processing ofCgA in cella. PC12 cells were transfected with pcDNA3.1 (lane marked V) or pcDNA3.1-CTSL (lane markedCTSL). Three days after transfection, cells were harvested, lysed, and subjected to 10–20% tricine gel.Proteins were then transferred to polyvinyl difluoride membrane and analyzed by Western blot usinganticatestatin antibody. Catestatin immunoreactive bands were shown in the middle panel. A parallel blotwas stained with anti-CTSL and antiactin antibody (left panel). In the right panel, purified humanrecombinant CgA (10 �M) was digested with 0.06, 0.03, and 0.015 �M of CTSL and subjected toimmunoblot with anticatestatin antibody. In lane 1 of right panel, synthetic catestatin (CgA352–372) wasloaded. Molecular size markers (Bio-Rad prestained broad range) were used in lane marked M. The lanewith -Enz represents zero enzyme control.
3550 Biswas et al. CTSL in Generation of Catestatin Variants Endocrinology, August 2009, 150(8):3547–3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

MALDI-TOF analysis. Analysis of the mass spectrometric datarevealed generation of the following eight peptides CgA360 –366
(ARAYGFR, m/z 840), CgA374 –380 (RGWRPSS, m/z 845),CgA367–374 (GPGPQLRR, m/z 880), CgA365–373 (FRGPG-PQLR, m/z 1027), CgA365–374 (FRGPGPQLRR, m/z 1183),CgA360–373 (ARAYGFRGPGPQLR, m/z 1545), and CgA367–380
(GPGPQLRRGWRPSS, m/z 1550) and CgA365–380 (FRGPG-PQLRRGWRPSS, m/z 1853) from the short monodibasic cates-tatin synthetic peptide (Fig. 3A and Table 1). The peptides with m/z840, 880, 1027, 1183, and 1545 were also generated from thefull-length recombinant protein, thus reinforcing the cleavage spec-ificity of CTSL. The digestion of the long catestatin peptide(CgA333–379) peptide with the flanking dibasic residues was at-tempted with very low concentration of CTSL (0.7 nM) and forbetter sensitivity theMALDI-TOFanalysiswas carriedout in linearmode from 1000 to 6000 Da range (Fig. 3B). The presence of un-digested peptide of (m/z 5496) was evident and indicated the partialcleavage of the synthetic substrate. The following peptides were de-duced from the mass analysis: CgA367–379 (m/z 1464), CgA360–373
(m/z 1546), CgA340–354 (m/z 1768), CgA360–379 (m/z 2286),CgA357–378/CgA358–379 (m/z 2590), CgA341–363 (m/z 2748),CgA333–359 (m/z 3227), CgA333–366 (m/z 4049), and CgA333–373
(m/z 4755) (Table 2).Active catestatin fragment CgA360–373 was generated by
CTSL digestion from both full-length CgA and the long cates-tatin region-spanning synthetic peptide CgA333–379, with m/z
about 1546 in each case; thus, CTSL’s cleavage preference withinthe catestatin region was similar in both CgA and a syntheticlocal region (Figs. 2B and 3).
Suppression of catecholamine release by CTSL-generated CgA fragments
Wehaveshownpreviously thatplasmingeneratedCgApeptidesinhibited nicotine-stimulated catecholamine release (30). To deter-mine the potency of the peptides generated by the CTSL digestion,we performed norepinephrine secretion assay in 3H-norepineph-rine-loaded PC12 cells. Recombinant CgA (10 �M) was digestedwith CTSL (0.3 �M), and the resulting peptides were purified witha C18 cartridge. The nicotine-stimulated norepinephrine secretionwas then tested in the presence of undigested CgA, digested CgA, orsyntheticCgA360–373 (Fig.4A, left panel),whichcauseddifferentialinhibition of nicotine-evoked catecholamine release: 20.87% byundigested CgA, 96.79% by CTSL-generated peptide mixture, and74.32% by the synthetic catestatin peptide (CgA360–373). TheCTSL-generated peptide mixtures abolished nicotine-evoked cate-cholamine secretionwithan IC50 of0.105 �M (Fig.4B, rightpanel).
Digestion of CgA containing naturally occurringcatestatin variants by CTSL
In our previous communication, we have shown differentialcleavage of human catestatin variants by plasmin (30). In thepresent study, we sought to determine whether CTSL is also
FIG. 2. Proteolysis of purified recombinant CgA by CTSL. A, Dose dependence (upper panel). Recombinant human CgA (10 �M) was incubated with different doses(0.6, 0.3, 0.15, 0.06, 0.03, 0.015 �M; lanes 2–7, upper panel) of CTSL for 15 min at 30 C. Lower panel, Time dependence. CgA (10 �M) was incubated with 0.15 �M ofCTSL and aliquots were taken at different time intervals. The reaction products were subjected to SDS-PAGE on a 10% gel and stained with Coomassie blue. Molecularsize markers (precision plus protein unstained standards) were used in lane marked M. E64 (CTSL inhibitor) specificity controls are also shown. The lane with-Enz representszero enzyme control. B, Analysis of peptides generated by CTSL. CgA (10 �M/reaction) was digested with 0.03 �M of CTSL for 15 min at 30 C. The reaction product wassubjected to MALDI-TOF. Potential fragments generating from the catestatin region as identified by protein prospector program were listed in the table below.
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3551
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

capable of displaying differential cleavage pattern to human ca-testatin variants. Therefore, the following purified proteins CgA-WT, CgA-Gly364Ser, CgA-Pro370Leu, and CgA-Arg374Gln at
10 �M were subjected to proteolysis by CTSL (0.03 �M) and theresulting peptides were analyzed by SDS-PAGE (Fig. 5A) andMALDI-mass spectrometry (Fig. 5B).
On SDS-PAGE, the digestion pattern, as evidenced by theimmunoblot with catestatin antibody, appeared qualitativelycomparable and quantitatively different for the four proteins.Several catestatin-positive fragments were detected with mass11, 26, 34, and 40 kDa (Fig. 5A). In all four proteins, the detectedlowest-molecular-mass catestatin-positive fragment was about11 kDa that differed quantitatively: Gly364Ser (9.67% com-pared with WT), Pro370Leu (70.5% compared with WT), andArg374Gln (80.6% compared with WT) (Fig. 5A).
MALDI-TOF revealed the generation of short internalfragments with m/z 840 (CgA360–366), 880 (CgA367–374),1027 (CgA365–373), 1143 (CgA358–366), 1183 (CgA365–374),1318 (CgA362–373), 1545 (CgA360–373), 1567 (CgA349–361), and1848 (CgA358–373) from the wild-type CgA (Fig. 5B). Digestionof CgA-Gly364Ser did not generate the small catestatin peptide(CgA360–373; expected m/z �1576); instead, two internal frag-ments (CgA360 –366 and CgA367–374) within the functional ca-testatin domain were generated (Fig. 5B). Consistent with our
TABLE 1. Digestion of catestatin-region synthetic peptideCgA360–380 by CTSL
Mass (m/z)submitted
Mass (m/z)matched Sequence
840.4344 840.4475 _ARAYGFR_G845.4232 845.4377 R_RGWRPSS_880.5095 880.5112 F_RGPGPQLR_R or
R_GPGPQLRR_G orG_PGPQLRRG_W
1027.5590 1027.5796 G_FRGPGPQLR_R1183.6744 1183.6807 G_FRGPGPQLRR_G1545.8408 1545.8397 _ARAYGFRGPGPQLR_R1550.8342 1550.8299 R_GPGPQLRRGWRPSS_1853.9360 1853.9994 G_FRGPGPQLRRGWRPSS_
Catestatin region synthetic peptide was digested with CTSL for 15 min at 30 C,and the reaction products were analyzed by MALDI-TOF. Possible fragmentsgenerated in the catestatin region are listed in the table. CgA360–380 was 10�M/reaction, and CTSL was 0.015 �M. hCgA360–380 synthetic peptide:ARAYGFRGPGPQLRRGWRPSS.
FIG. 3. Digestion of catestatin-region synthetic peptides by CTSL. Catestatin region synthetic peptides, CgA360–380 (A) or CgA333–379 (B) (10 �M/reaction), weredigested with 0.015 �M and 0.68 nM CTSL, respectively, for 15 min at 30 C, and the reaction products were analyzed by MALDI-TOF in reflectron mode for A and linearmode for B.
3552 Biswas et al. CTSL in Generation of Catestatin Variants Endocrinology, August 2009, 150(8):3547–3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

plasmin digestion, CTSL digestion of CgA-Pro370Leu alsofailed to generate functional catestatin peptide (CgA360 –373;expected m/z �1561) (Fig. 5B). Despite having a mutationwithin the dibasic site (Arg373Arg374), the CTSL digestionpattern of CgA-Arg374Gln was comparable with WT-CgA(Fig. 5B).
Thus, CTSL displayed differential proteolytic digestionpattern in human variants of catestatin, and the generation ofsmall catestatin fragments (such as CgA360 –373) was impairedin the Gly364Ser and Pro370Leu versions of CgA.
Discussion
OverviewThe biosynthesis of the physiologically active catestatin pep-
tide (CgA352–372) requires a proteolytic processing of its CgAprecursor within the chromaffin granules/vesicles of adrenal me-
dulla or large dense core vesicles of endocrine or neuroendocrinecells. But the mechanism and the specific proteases involved inthe processing of the prohormone precursors have so far beenincompletely understood. We have shown previously that plas-min, a major fibrinolytic enzyme, cleaved CgA in vitro to pro-duce a catestatin fragment that inhibited nicotine-stimulatednorepinephrine secretion from PC12 cells (30). The involvementof the serine proteases PC1/PC3 and PC2 in the proteolytic deg-radation of CgA have already been demonstrated (20–27). Re-cent findings revealed that CTSL is present in the secretory ves-icles and is responsible for the processing of the neuropeptideenkephalin and neuropeptide Y (31–33). Overexpression ofCTSL in PC12 cells reduced the abundance of full-length CgA,indicating substrate-like behavior of the latter (Fig. 1C). In ad-dition, the generated catestatin immunopositive peptides inCTSL-overexpressing PC12 cells resembled in vitro digestionproducts of recombinant CgA (Fig. 1C). Subcellular colocaliza-tion of CgA with CTSL in secretory granules also implicated
the functional relationship between thesetwo proteins (Fig. 1, A and B). CathepsinB, D, and K are secreted from thyroid ep-ithelial cells in a regulated pathway andinvolved in extracellular prohormoneprocessing (50 –52). Although CTSL is de-scribed as lysosomal protease, our immu-nofluorescence data in sympathochro-maffin PC12 cells indicated substantialcolocalization of CTSL with the secretoryprotein CgA. This is consistent with theobservation in mouse pituitary, in whichCTSL showed partial colocalization withthe lysosomal membrane-associated pro-tein 1 but more extensive colocalizationwith adrenocorticotropic hormone, �-endor-phin, and �-melanocyte stimulating hormone(33). Previously we demonstrated very littleoverlap in localization between CgA andthe late endosome/lysosome marker pro-tein LGP110 in the A35C cells (9).
0
5
10
15
20
25
Basal Nicotine (60µM)
Nic+
CTSLdigestedCgA mix(1.5µM)
Nic+
SyntheticCgA
360-373(10µM)
Nic+
FullLengthCgA
(1.5µM)
Net
3 H-N
orep
inep
hrin
e re
leas
e (%
)
A
p<0.001
p<0.001
p<0.01
0
20
40
60
80
100
120
-0.5 0 0.5 1 1.5 2CTSL digested CgA (µM)
Net
3 H-N
orep
inep
hrin
e re
leas
e(%
of c
ontro
l)
IC50
~0.105 µM
Stimulus: Nicotine (60 µM)
B
FIG. 4. Effect of CTSL-digested peptides on secretagogue-stimulated norepinephrine secretion by PC12cells. A, PC12 cells were stimulated with 60 �M nicotine (Nic) in the absence (basal) or presence ofundigested CgA (1.5 �M), CTSL-digested CgA (1.5 �M), or synthetic catestatin peptide (CgA360–373; 10�M). After 30 min incubation, the cells and media were analyzed for norepinephrine secretion. In control,the net norepinephrine release (100%) was measured in the presence of 60 �M nicotine alone. B, Dose-dependent inhibition of CTSL digests on secretion assay was shown.
TABLE 2. Digestion of catestatin-region synthetic peptide CgA333–379 by CTSL
Mass (m/z)Average Sequence Residues of CgA
1464.6 R_GPGPQLRRGWRPS_ 367–3791546.4 R_ARAYGFRGPGPQLR_R or F_RARAYGFRGPGPQL_R 360–373 or 359–3721767.9 R_LEGQEEEEDNRDSSM_K 340–3542286.5 R_ARAYGFRGPGPQLRRGWRPS_ 360–3792590.1 L_SFRARAYGFRGPGPQLRRGWRP_S or
S_FRARAYGFRGPGPQLRRGWRPS_357–378 or 358–379
2747.6 E_EEDNRDSSMKLSFRARAYGFRGPG_P orL_EGQEEEEDNRDSSMKLSFRARAY_G
346–369 or 341–363
3227.6 _ELTAEKRLEGQEEEEDNRDSSMKLSFR_A 333–3594049.9 _ELTAEKRLEGQEEEEDNRDSSMKLSFRARAYGFR_G 333–3664755.7 _ELTAEKRLEGQEEEEDNRDSSMKLSFRARAYGFRGPGPQLR_R 333–3735496.1 _ELTAEKRLEGQEEEEDNRDSSMKLSFRARAYGFRGPGPQLRRGWRPS_ 333–379
Catestatin region synthetic peptide was digested with CTSL for 15 min at 30 C, and the reaction products were analyzed by MALDI-TOF. Possible fragments generatedin the catestatin region are listed in the table. CgA333–379 was 10 �M/reaction, and CTSL was 0.68 nM. hCgA333–379 synthetic peptide:ELTAEKRLEGQEEEEDNRDSSMKLSFRARAYGFRGPGPQLRRGWRPS.
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3553
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

Cleavage in vitroIn the present communication, we undertook biochemical
approaches coupled with mass spectrometry to characterize theproteolytic digestion of CgA with CTSL. At the outset, we useda three-step purification procedure to obtain highly purified pro-teins, and the in vitro proteolysis study showed that the humanrecombinant CgA proteins were processed readily in a time- anddose-dependent fashion (Fig. 2A). At high protease concentra-tions, a series of N-terminal catestatin fragments were generated(supplemental Table 1). Earlier we identified a minimal activecore of bovine catestatin, consisting of the N-terminal 15-aminoacid residues (bovine CgA344–358) within the catestatin (bovineCgA344–364) region (15) using synthetic N- and C-terminal andbidirectional deletion peptides.
Correspondence of CTSL cleavage sites to previouslyestablished sites in CgA in vivo
In the catestatin region, previous analyses in bovine chromaf-fin granule peptides identified cleavage at R3312L332 of bovineCgA (53); we found cleavage at the corresponding human siteR3392L340 (Table 2 and Fig. 6B). Likewise, previously deter-mined cleavage at bovine CgA R3582G359 corresponds to ourhuman CgA R3662G367, and bovine CgA R3532G354 corre-sponds to our human CgA R3612A362. In previously determinedsites of human CgA cleavage in pheochromocytoma (54), siteR3612A362 is also found in our CTSL digests (Fig. 6B), whereas
L3722R373 would be expected after carboxypeptidase digestionof the peptide resulting from our R3732R374 cleavage (Fig. 6B).
We obtained additional fragments from in vitro digestion,which were not described previously. It is conceivable that post-translational modifications of the purified protein are not thesame as native protein, in which case different conformationsmay influence accessibility to proteases. We also used a widerange of CTSL concentrations to digest CgA; some of those con-ditions may not replicate the in vivo substrate to enzyme ratioand might have resulted in generation of additional fragments.Additional in vivo proteomic analyses along with the in vitrodigestion data will be necessary to clarify this point.
Previous studies on mice with targeted ablation of the CTSLgene demonstrated decreased processing of ACTH, �-endor-phin, �-MSH, and proneuropeptide Y in brain (33, 55).
Consensus CTSL cleavage site in CgAThe WebLogo evaluation (56) of CTSL-generated peptides
from CgA indicates that this protease does not have a strong,unequivocal preference for basic residues at the cleavage site(P1-P1�), although it does prefer leucine at the P2 subsite (Fig.6A). Whereas a preference of CTSL for aromatic and hydropho-bic residues upstream of the cleavage site has been reported pre-viously (57–59), we did observe such a preference in CgA. In theabsence of Leu at the P2 or P3 position, CTSL did show a pro-pensity toward positively charged residues lysine and arginine at
FIG. 5. Digestion of CgA with catestatin variants by CTSL. A, Immunoblots. WT and mutant CgA (10 �M/reaction) containing the catestatin variants was digested with0.03 �M CTSL for 15 min and subjected to separation on a 10–20% tricine gel. Proteins were then transferred to polyvinyl difluoride membrane, and immunoblottedwith anticatestatin antibody. In the last lane, synthetic catestatin (CgA352–372) was loaded. Molecular size markers (Biorad prestained broad range) were used in lanemarked M. B, MALDI-TOF. CTSL digests (same as A) were subjected to MALDI-TOF analysis.
3554 Biswas et al. CTSL in Generation of Catestatin Variants Endocrinology, August 2009, 150(8):3547–3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from
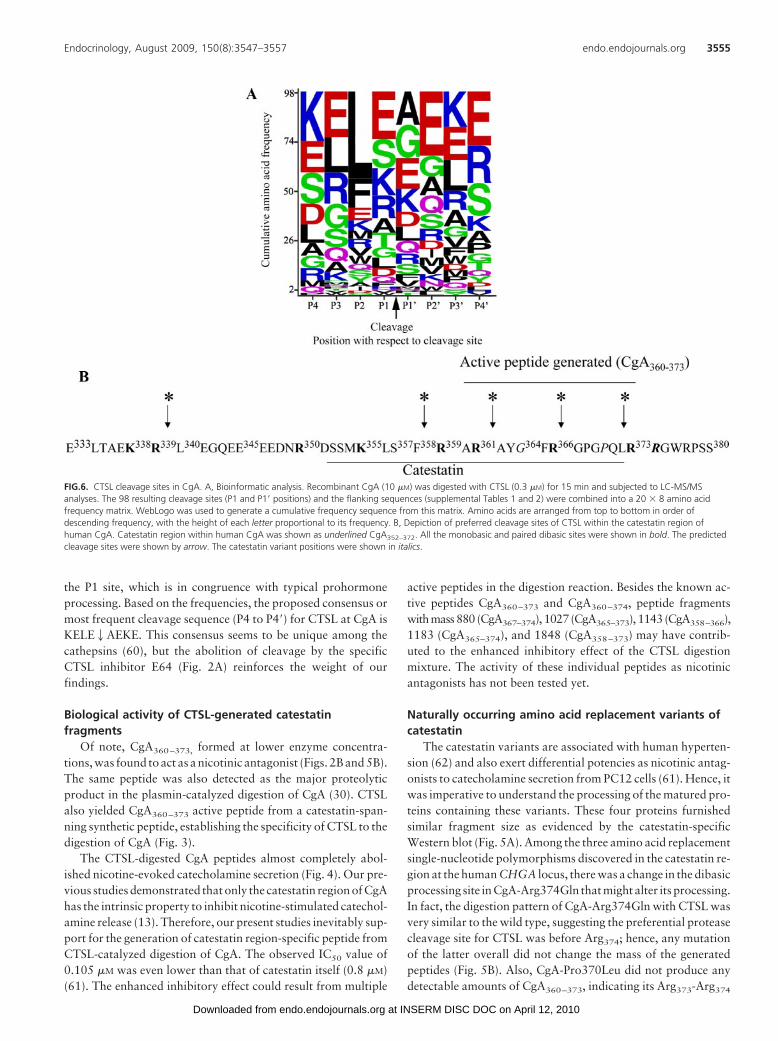
the P1 site, which is in congruence with typical prohormoneprocessing. Based on the frequencies, the proposed consensus ormost frequent cleavage sequence (P4 to P4�) for CTSL at CgA isKELE2AEKE. This consensus seems to be unique among thecathepsins (60), but the abolition of cleavage by the specificCTSL inhibitor E64 (Fig. 2A) reinforces the weight of ourfindings.
Biological activity of CTSL-generated catestatinfragments
Of note, CgA360–373, formed at lower enzyme concentra-tions, was found to act as a nicotinic antagonist (Figs. 2B and 5B).The same peptide was also detected as the major proteolyticproduct in the plasmin-catalyzed digestion of CgA (30). CTSLalso yielded CgA360–373 active peptide from a catestatin-span-ning synthetic peptide, establishing the specificity of CTSL to thedigestion of CgA (Fig. 3).
The CTSL-digested CgA peptides almost completely abol-ished nicotine-evoked catecholamine secretion (Fig. 4). Our pre-vious studies demonstrated that only the catestatin region of CgAhas the intrinsic property to inhibit nicotine-stimulated catechol-amine release (13). Therefore, our present studies inevitably sup-port for the generation of catestatin region-specific peptide fromCTSL-catalyzed digestion of CgA. The observed IC50 value of0.105 �M was even lower than that of catestatin itself (0.8 �M)(61). The enhanced inhibitory effect could result from multiple
active peptides in the digestion reaction. Besides the known ac-tive peptides CgA360–373 and CgA360–374, peptide fragmentswith mass 880 (CgA367–374), 1027 (CgA365–373), 1143 (CgA358–366),1183 (CgA365–374), and 1848 (CgA358–373) may have contrib-uted to the enhanced inhibitory effect of the CTSL digestionmixture. The activity of these individual peptides as nicotinicantagonists has not been tested yet.
Naturally occurring amino acid replacement variants ofcatestatin
The catestatin variants are associated with human hyperten-sion (62) and also exert differential potencies as nicotinic antag-onists to catecholamine secretion from PC12 cells (61). Hence, itwas imperative to understand the processing of the matured pro-teins containing these variants. These four proteins furnishedsimilar fragment size as evidenced by the catestatin-specificWestern blot (Fig. 5A). Among the three amino acid replacementsingle-nucleotide polymorphisms discovered in the catestatin re-gion at the human CHGA locus, there was a change in the dibasicprocessing site in CgA-Arg374Gln that might alter its processing.In fact, the digestion pattern of CgA-Arg374Gln with CTSL wasvery similar to the wild type, suggesting the preferential proteasecleavage site for CTSL was before Arg374; hence, any mutationof the latter overall did not change the mass of the generatedpeptides (Fig. 5B). Also, CgA-Pro370Leu did not produce anydetectable amounts of CgA360–373, indicating its Arg373-Arg374
FIG.6. CTSL cleavage sites in CgA. A, Bioinformatic analysis. Recombinant CgA (10 �M) was digested with CTSL (0.3 �M) for 15 min and subjected to LC-MS/MSanalyses. The 98 resulting cleavage sites (P1 and P1� positions) and the flanking sequences (supplemental Tables 1 and 2) were combined into a 20 � 8 amino acidfrequency matrix. WebLogo was used to generate a cumulative frequency sequence from this matrix. Amino acids are arranged from top to bottom in order ofdescending frequency, with the height of each letter proportional to its frequency. B, Depiction of preferred cleavage sites of CTSL within the catestatin region ofhuman CgA. Catestatin region within human CgA was shown as underlined CgA352–372. All the monobasic and paired dibasic sites were shown in bold. The predictedcleavage sites were shown by arrow. The catestatin variant positions were shown in italics.
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3555
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

bond cleavage was inefficient (Fig. 5B). In particular, both plas-min and CTSL proteases were unable to generate the catestatinfragment (CgA360–373) from CgA-Pro370Leu variant. In otherwords, CgA-Pro370Leu variant displayed resistance to cleavageby both plasmin and CTSL. The digestion pattern of CgA-Gly364Ser by CTSL was different from that of the wild-typeprotein (Fig. 5B), in which the formation of catestatin-positive11-kDa fragment and CgA360–373 short peptide was less efficient(Fig. 5, A and B). The processing sites at Arg3592 and Arg3732in CgA-Gly364Ser were similar to that of wild-type, as evidencedfrom the generation of internal peptides CgA360–366, CgA365–373,and CgA365–374, but the generation of 14-amino acid active frag-ment (CgA360–373) could not be substantiated. It is possible thatchange of glycine to serine induced more internal cleavage, gen-erating short fragments. The digestion patterns of the full-lengthprotein and synthetic peptides delineated the preferred cleavagesite of CTSL are in and around the catestatin region, as summa-rized in Fig. 6B. Unlike plasmin, the digestion of CgA with abroad range of CTSL concentrations did not offer preferentialgeneration of any particular peptide. The internal cleavage atSer3572, Arg3592, 2Gly364, and Arg3662 were apparent inwild-type and the catestatin mutants. The formation of CgA360–373
within a narrow window of substrate to protease ratio suggesteda complex proteolytic cleavage of CgA by CTSL. The concen-tration of the thiol protease in the secretory vesicles may be quitelow (63), and under that substrate to enzyme ratio, in lieu of theinternal sites, other sites may be preferably cleaved to generatethe longer catestatin fragments. The endogenous inhibitors ofgeneral cysteine protease might also play a role to direct thecleavage specificity.
ConclusionsTaken together, our results indicate that CgA could be a sub-
strate of CTSL, and the catestatin nonsynonymous variantsshowed differential processing toward CTSL digestion. Furtherexperiments will be designed to verify the proteolytic profiling ofthe proteins using a different protease and/or a combinationof proteases to better understand the development of humanhypertension.
Acknowledgments
Wearegrateful toProfessorElizabethA.Komives (DepartmentofChem-istry and Biochemistry, University of California, San Diego) for helpingus in MALDI-TOF and MS/MS experiments.
Address all correspondence and requests for reprints to: Sushil K.Mahata, Ph.D., Department of Medicine (0838), University of Cali-fornia, San Diego, School of Medicine and Veterans Affairs San DiegoHealthcare System, 9500 Gilman Drive, La Jolla, California 92093-0838. E-mail: [email protected] (http://medicine.ucsd.edu/hypertension).
This work was supported by National Institutes of Health GrantsR29 DA011311 and R01 DA011311 (to S.K.M.), DK 60702 (toD.T.O.), P01 HL58120 and U01 HL69758 (to S.K.M. and D.T.O.); andgrants from the Department of Veterans Affairs (to S.K.M. and D.T.O.).
Disclosure Summary: The authors have nothing to disclose.
References
1. Winkler H, Fischer-Colbrie R 1992 The chromogranins A and B: the first 25years and future perspectives. Neuroscience 49:497–528
2. Taupenot L, Harper KL, O’Connor DT 2003 Mechanisms of disease: thechromogranin-secretogranin family. N Engl J Med 348:1134–1149
3. Montero-Hadjadje M, Vaingankar S, Elias S, Tostivint H, Mahata SK, AnouarY 2008 Chromogranins A and B and secretogranin II: evolutionary and func-tional aspects. Acta Physiol (Oxf) 192:309–324
4. Takiyyuddin MA, Cervenka JH, Hsiao RJ, Barbosa JA, Parmer RJ, O’ConnorDT 1990 Chromogranin A. Storage and release in hypertension. Hypertension15:237–246
5. Takiyyuddin MA, Cervenka JH, Sullivan PA, Pandian MR, Parmer RJ,Barbosa JA, O’Connor DT 1990 Is physiologic sympathoadrenal catechol-amine release exocytotic in humans? Circulation 81:185–195
6. Kelly RB 1985 Pathways of protein secretion in eukaryotes. Science 230:25–327. Kim T, Tao-Cheng JH, Eiden LE, Loh YP 2001 Chromogranin A, an “on/off”
switch controlling dense-core secretory granule biogenesis. Cell 106:499–5098. Kim T, Zhang CF, Sun Z, Wu H, Loh YP 2005 Chromogranin A deficiency in
transgenic mice leads to aberrant chromaffin granule biogenesis. J Neurosci25:6958–6961
9. Courel M, Rodemer C, Nguyen ST, Pance A, Jackson AP, O’connor DT,Taupenot L 2006 Secretory granule biogenesis in sympathoadrenal cells: iden-tification of a granulogenic determinant in the secretory prohormone chro-mogranin A. J Biol Chem 281:38038–38051
10. Tatemoto K, Efendiæ S, Mutt V, Makk G, Feistner GJ, Barchas JD 1986Pancreastatin, a novel pancreatic peptide that inhibits insulin secretion. Nature324:476–478
11. Cadman PE, Rao F, Mahata SK, O’Connor DT 2002 Studies of the dysgly-cemic peptide, pancreastatin, using a human forearm model. Ann NY Acad Sci971:528–529
12. Aardal S, Helle KB, Elsayed S, Reed RK, Serck-Hanssen G 1993 Vasostatins,comprising the N-terminal domain of chromogranin A, suppress tension inisolated human blood vessel segments. J Neuroendocrinol 5:405–412
13. Mahata SK, O’Connor DT, Mahata M, Yoo SH, Taupenot L, Wu H, Gill BM,Parmer RJ 1997 Novel autocrine feedback control of catecholamine release. Adiscrete chromogranin A fragment is a noncompetitive nicotinic cholinergicantagonist. J Clin Invest 100:1623–1633
14. Mahata SK, Mahata M, Parmer RJ, O’Connor DT 1999 Desensitization ofcatecholamine release: the novel catecholamine release-inhibitory peptide ca-testatin (chromogranin A344–364) acts at the receptor to prevent nicotinic cho-linergic tolerance. J Biol Chem 274:2920–2928
15. Mahata SK, Mahata M, Wakade AR, O’Connor DT 2000 Primary structureand function of the catecholamine release inhibitory peptide catestatin (chro-mogranin A344–364): Identification of amino acid residues crucial for activ-ity. Mol Endocrinol 14:1525–1535
16. Mahata SK 2004 Catestatin—the catecholamine release inhibitory peptide: astructural and functional overview. Curr Med Chem Immun Endocr MetabAgents 4:221–234
17. TakiyyuddinMA,ParmerRJ,KailasamMT,Cervenka JH,Kennedy B,ZieglerMG, Lin MC, Li J, Grim CE, Wright FA, et al 1995 Chromogranin A in humanhypertension. Influence of heredity. Hypertension 26:213–220
18. O’Connor DT, Kailasam MT, Kennedy BP, Ziegler MG, Yanaihara N, ParmerRJ 2002 Early decline in the catecholamine release-inhibitory peptide cates-tatin in humans at genetic risk of hypertension. J Hypertens 20:1335–1345
19. Mahapatra NR, O’Connor DT, Vaingankar SM, Hikim AP, Mahata M, RayS, Staite E, Wu H, Gu Y, Dalton N, Kennedy BP, Ziegler MG, Ross J, MahataSK 2005 Hypertension from targeted ablation of chromogranin A can berescued by the human ortholog. J Clin Invest 115:1942–1952
20. Benjannet S, Leduc R, Adrouche N, Falgueyret JP, Marcinkiewicz M, SeidahNG, Mbikay M, Lazure C, Chretien M 1987 Chromogranin B (secretograninI), a putative precursor of two novel pituitary peptides through processing atpaired basic residues. FEBS Lett 224:142–148
21. Hoflehner J, Eder U, Laslop A, Seidah NG, Fischer-Colbrie R, Winkler H 1995Processing of secretogranin II by prohormone convertases: importance of PC1in generation of secretoneurin. FEBS Lett 360:294–298
22. Eskeland NL, Zhou A, Dinh TQ, Wu H, Parmer RJ, Mains RE, O’Connor DT1996 Chromogranin A processing and secretion: specific role of endogenousand exogenous prohormone convertases in the regulated secretory pathway.J Clin Invest 98:148–156
23. Seidah NG, Day R, Marcinkiewicz M, Chretien M 1998 Precursor converta-ses: an evolutionary ancient, cell-specific, combinatorial mechanism yieldingdiverse bioactive peptides and proteins. Ann NY Acad Sci 839:9–24
24. Laslop A, Weiss C, Savaria D, Eiter C, Tooze SA, Seidah NG, Winkler H 1998
3556 Biswas et al. CTSL in Generation of Catestatin Variants Endocrinology, August 2009, 150(8):3547–3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from

Proteolytic processing of chromogranin B and secretogranin II by prohormoneconvertases. J Neurochem 70:374–383
25. Seidah NG, Prat A 2002 Precursor convertases in the secretory pathway, cy-tosol and extracellular milieu. Essays Biochem 38:79–94
26. Doblinger A, Becker A, Seidah NG, Laslop A 2003 Proteolytic processing ofchromogranin A by the prohormone convertase PC2. Regul Pept 111:111–116
27. Scamuffa N, Calvo F, Chretien M, Seidah NG, Khatib AM 2006 Proproteinconvertases: lessons from knockouts. FASEB J 20:1954–1963
28. Parmer RJ, Mahata M, Gong Y, Mahata SK, Jiang Q, O’Connor DT, Xi XP,Miles LA 2000 Processing of chromogranin A by plasmin provides a novelmechanism for regulating catecholamine secretion. J Clin Invest 106:907–915
29. Jiang Q, Taupenot L, Mahata SK, Mahata M, O’Connor DT, Miles LA,Parmer RJ 2001 Proteolytic cleavage of chromogranin A (CgA) by plasmin:selective liberation of a specific bioactive CgA fragment that regulates cate-cholamine release. J Biol Chem 276:25022–25029
30. Biswas N, Vaingankar SM, Mahata M, Das M, Gayen JR, Taupenot L, TorpeyJW, O’Connor DT, Mahata SK 2008 Proteolytic cleavage of human chromo-granin a containing naturally occurring catestatin variants: differential pro-cessing at catestatin region by plasmin. Endocrinology 149:749–757
31. Yasothornsrikul S, Greenbaum D, Medzihradszky KF, Toneff T, Bundey R,Miller R, Schilling B, Petermann I, Dehnert J, Logvinova A, Goldsmith P,Neveu JM, Lane WS, Gibson B, Reinheckel T, Peters C, Bogyo M, Hook V2003 Cathepsin L in secretory vesicles functions as a prohormone-processingenzyme for production of the enkephalin peptide neurotransmitter. Proc NatlAcad Sci USA 100:9590–9595
32. Hwang SR, Garza C, Mosier C, Toneff T, Wunderlich E, Goldsmith P, HookV 2007 Cathepsin L expression is directed to secretory vesicles for enkephalinneuropeptide biosynthesis and secretion. J Biol Chem 282:9556–9563
33. Funkelstein L, Toneff T, Hwang SR, Reinheckel T, Peters C, Hook V 2008Cathepsin L participates in the production of neuropeptide Y in secretoryvesicles, demonstrated by protease gene knockout and expression. J Neuro-chem 106:384–391
34. Hook V, Funkelstein L, Lu D, Bark S, Wegrzyn J, Hwang SR 2008 Proteasesfor processing proneuropeptides into peptide neurotransmitters and hor-mones. Annu Rev Pharmacol Toxicol 48:393–423
35. Kirschke H, Barrett AJ, Rawlings ND 1995 Proteinases 1: lysosomal cysteineproteinases. Protein Profile 2:1581–1643
36. Docherty K, Carroll R, Steiner DF 1983 Identification of a 31,500 molecularweight islet cell protease as cathepsin B. Proc Natl Acad Sci USA 80:3245–3249
37. Docherty K, Hutton JC, Steiner DF 1984 Cathepsin B-related proteases in theinsulin secretory granule. J Biol Chem 259:6041–6044
38. Kukor Z, Mayerle J, Kruger B, Toth M, Steed PM, Halangk W, Lerch MM,Sahin-Toth M 2002 Presence of cathepsin B in the human pancreatic secretorypathway and its role in trypsinogen activation during hereditary pancreatitis.J Biol Chem 277:21389–21396
39. Neves FA, Duncan KG, Baxter JD 1996 Cathepsin B is a prorenin processingenzyme. Hypertension 27:514–517
40. Jutras I, Reudelhuber TL 1999 Prorenin processing by cathepsin B in vitro andin transfected cells. FEBS Lett 443:48–52
41. McCoy K, Gal S, Schwartz RH, Gottesman MM 1988 An acid protease se-creted by transformed cells interferes with antigen processing. J Cell Biol 106:1879–1884
42. Goretzki L, Schmitt M, Mann K, Calvete J, Chucholowski N, Kramer M,Gunzler WA, Janicke F, Graeff H 1992 Effective activation of the proenzymeform of the urokinase-type plasminogen activator (pro-uPA) by the cysteineprotease cathepsin L. FEBS Lett 297:112–118
43. Kasai M, Shirasawa T, Kitamura M, Ishido K, Kominami E, Hirokawa K 1993Proenzyme from of cathepsin L produced by thymic epithelial cells promotesproliferation of immature thymocytes in the presence of IL-1, IL-7, and anti-CD3 antibody. Cell Immunol 150:124–136
44. Stypmann J, Glaser K, Roth W, Tobin DJ, Petermann I, Matthias R, MonnigG, Haverkamp W, Breithardt G, Schmahl W, Peters C, Reinheckel T 2002Dilated cardiomyopathy in mice deficient for the lysosomal cysteine peptidasecathepsin L. Proc Natl Acad Sci USA 99:6234–6239
45. Yang M, Zhang Y, Pan J, Sun J, Liu J, Libby P, Sukhova GK, Doria A,Katunuma N, Peroni OD, Guerre-Millo M, Kahn BB, Clement K, Shi GP
2007 Cathepsin L activity controls adipogenesis and glucose tolerance. NatCell Biol 9:970 –977
46. Li W, Kornmark L, Jonasson L, Forssell C, Yuan XM 2009 Cathepsin L issignificantly associated with apoptosis and plaque destabilization in humanatherosclerosis. Atherosclerosis 202:92–102
47. Mahata M, Mahata SK, Parmer RJ, O’Connor DT 1996 Vesicular monoaminetransport inhibitors. Novel action at calcium channels to prevent catechol-amine secretion. Hypertension 28:414–420
48. Taupenot L, Remacle JE, Helle KB, Aunis D, Bader MF 1995 Recombinanthuman chromogranin A: expression, purification and characterization of theN-terminal derived peptides. Regul Pept 56:71–88
49. Courel M, Vasquez MS, Hook VY, Mahata SK, Taupenot L 2008 Sorting ofthe neuroendocrine secretory protein secretogranin II into the regulated se-cretory pathway: role of N- and C-terminal �-helical domains. J Biol Chem283:11807–11822
50. Brix K, Linke M, Tepel C, Herzog V 2001 Cysteine proteinases mediate ex-tracellular prohormone processing in the thyroid. Biol Chem 382:717–725
51. Colombo B, Longhi R, Marinzi C, Magni F, Cattaneo A, Yoo SH, Curnis F,Corti A 2002 Cleavage of chromogranin A N-terminal domain by plasminprovides a new mechanism for regulating cell adhesion. J Biol Chem 277:45911–45919
52. Tepel C, Bromme D, Herzog V, Brix K 2000 Cathepsin K in thyroid epithelialcells: sequence, localization and possible function in extracellular proteolysisof thyroglobulin. J Cell Sci 113 Pt 24:4487–4498
53. Metz-Boutigue MH, Garcia-Sablone P, Hogue-Angeletti R, Aunis D 1993Intracellular and extracellular processing of chromogranin A. Determinationof cleavage sites. Eur J Biochem 217:247–257
54. Orr DF, Chen T, Johnsen AH, Chalk R, Buchanan KD, Sloan JM, Rao P, ShawC 2002 The spectrum of endogenous human chromogranin A-derived peptidesidentified using a modified proteomic strategy. Proteomics 2:1586–1600
55. Funkelstein L, Toneff T, Mosier C, Hwang SR, Beuschlein F, Lichtenauer UD,Reinheckel T, Peters C, Hook V 2008 Major role of cathepsin L for producingthe peptide hormones ACTH, �-endorphin, and �-MSH, illustrated by pro-tease gene knockout and expression. J Biol Chem 283:35652–35659
56. Crooks GE, Hon G, Chandonia JM, Brenner SE 2004 WebLogo: a sequencelogo generator. Genome Res 14:1188–1190
57. Yasothornsrikul S, Aaron W, Toneff T, Hook VY 1999 Evidence for theproenkephalin processing enzyme prohormone thiol protease (PTP) as a mul-ticatalytic cysteine protease complex: activation by glutathione localized tosecretory vesicles. Biochemistry 38:7421–7430
58. Puzer L, Cotrin SS, Alves MF, Egborge T, Araujo MS, Juliano MA, Juliano L,Bromme D, Carmona AK 2004 Comparative substrate specificity analysis ofrecombinant human cathepsin V and cathepsin L. Arch Biochem Biophys 430:274–283
59. Abboud-Jarrous G, Atzmon R, Peretz T, Palermo C, Gadea BB, Joyce JA,Vlodavsky I 2008 Cathepsin L is responsible for processing and activation ofproheparanase through multiple cleavages of a linker segment. J Biol Chem283:18167–18176
60. Beynon RJ, Bond JS 1989 Proteolytic enzymes: a practical approach. NewYork: IRL Press at Oxford University Press
61. Mahata SK, Mahata M, Wen G, Wong WB, Mahapatra NR, Hamilton BA,O’Connor DT 2004 The catecholamine release-inhibitory “catestatin” frag-ment of chromogranin A: naturally occurring human variants with differentpotencies for multiple chromaffin cell nicotinic cholinergic responses. MolPharmacol 66:1180–1191
62. Rao F, Wen G, Gayen JR, Das M, Vaingankar SM, Rana BK, Mahata M,Kennedy BP, Salem RM, Stridsberg M, Abel K, Smith DW, Eskin E, Schork NJ,Hamilton BA, Ziegler MG, Mahata SK, O’Connor DT 2007 Catecholaminerelease-inhibitory peptide catestatin [chromogranin A(352–372)]: naturallyoccurring amino acid variant Gly364Ser causes profound changes in humanautonomic activity and alters risk for hypertension. Circulation 115:2271–2281
63. Krieger TJ, Hook VY 1991 Purification and characterization of a novel thiolprotease involved in processing the enkephalin precursor. J Biol Chem 266:8376–8383
Endocrinology, August 2009, 150(8):3547–3557 endo.endojournals.org 3557
at INSERM DISC DOC on April 12, 2010 endo.endojournals.orgDownloaded from
Copyright © 2022 FDOKUMEN




















