
Comparison and differentiation of potyvirus isolates and ...
12
Journal of General Virology (1991), 72, 25-36. Printed in Great Britain 25 Comparison and differentiation of potyvirus isolates and identification of strain-, virus-, subgroup-specific and potyvirus group-common epitopes using monoclonal antibodies Ramon Jordan* and John Hammond United States Department of Agriculture, Agricultural Research Service, Florist and Nursery Crops Laboratory, Building 004, Room 208, BARC West, Beltsville Agricultural Research Center, Beltsville, Maryland 20705, U.S.A. A panel of monoclonal antibodies (MAbs) generated against an admixture of 12 potyvirus isolates was used to compare and differentiate diverse potyviruses. Both native and denatured virions of strains of bean yellow mosaic (BYMV), potato virus Y, tobacco etch, pea seed-borne mosaic, iris severe mosaic, iris mild mosaic and asparagus virus- 1 potyviruses were used as immun- ogen and as antigen for screening of the hybridoma cell lines. Thirty cell lines secreting potyvirus-specific antibodies reactive in indirect antigen-coated plate (ACP-) ELISA were selected lbr detailed analysis. All 30 MAbs reacted with at least one strain of BYMV; 11 MAbs reacted with between one and eight of the nine BYMV strains and an additional three MAbs reacted only with isolates within the BYMV subgroup (BYMV, pea mosaic virus and clover yellow vein virus). The remaining 16 MAbs reacted with a BYMV isolate and with at least one of the other 43 potyvirus isolates tested. MAb PTY 1 reacted with all 55 potyvirus isolates tested (representing at least 33 different and distinct aphid-transmissible potyviruses). The poty- virus cross-reactive MAbs generally gave higher reacti- vity values in ACP-ELISA with dissociated virus than with polyclonal antibody-trapped intact virions in triple antibody sandwich ELISA (i.e. were cryptotope- specific). The BYMV strain- and virus-specific MAbs reacted strongly with both types of antigens (i.e. were metatope-specific). At least 25 distinct epitopes (12 cryptotopes and 13 metatopes) could be identified from the MAb-antigen reactivity patterns. The distribution of these epitopes between virus isolates can be used to detect and differentiate potyviruses in infected plant extracts and to examine virus architectures. Some of these epitopes are shared by potyvirus isolates not previously shown to be serologically related. The broad spectrum-reacting MAb PTY 1 recognizes a crypto- tope conserved on all of the aphid-transmissible potyviruses examined and should be a valuable tool for the detection and assay of these potyviruses. Introduction The potyvirus group, named after the type member potato virus Y (PVY), is the largest and economically the most important of the plant virus groups. Members of this group are responsible for important virus diseases affecting all types of vegetable, forage, fruit, ornamental and field crops (Hollings & Brunt, 1981; Milne, 1988). The potyvirus group contains over 160 definitive and possible members (Milne, 1988) which are characterized by having long (12 x 680 to 900 nm) flexuous, filamen- tous particles, containing one molecule of ssRNA of Mr 3"0x 106 to 3"5× 106 enclosed in coat protein composed of up to 2000 subunits of a single capsid polypeptide of Mr 30 × 103 to 37 x 103. All definitive and possible members induce the formation of character- istic cytoplasmic inclusions in infected cells which are serologically unrelated to their respective viral capsid protein. All of the definitive and most of the possible potyviruses are transmitted by aphid vector species but some possible members have fungus, mite or whitefly vectors (Hollings & Brunt, 1981; Milne, 1988). Serological relationships among definitive potyviruses are, however, highly complex and range from very close to intermediate to not detectable and do not always correlate with biological properties (Francki, 1983). Most of the definitive potyviruses have been shown to be serologically related to some degree to at least one other potyvirus but techniques used previously have failed to detect the expected serological relationships between many pairs of potyviruses. Polyclonal antisera show cross-reactivity of varying degrees with closely related viruses or strains of the virus immunogen, and usually fail to cross-react (or do so only very weakly) with more distantly related viruses. Polyclonal antisera produced to denatured potyviruses 0000-9767 © 1991 SGM
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Comparison and differentiation of potyvirus isolates and ...

Journal of General Virology (1991), 72, 25-36. Printed in Great Britain 25
Comparison and differentiation of potyvirus isolates and identification of strain-, virus-, subgroup-specific and potyvirus group-common epitopes using monoclonal antibodies
Ramon Jordan* and John Hammond
United States Department of Agriculture, Agricultural Research Service, Florist and Nursery Crops Laboratory, Building 004, Room 208, BARC West, Beltsville Agricultural Research Center, Beltsville, Maryland 20705, U.S.A.
A panel of monoclonal antibodies (MAbs) generated against an admixture of 12 potyvirus isolates was used to compare and differentiate diverse potyviruses. Both native and denatured virions of strains of bean yellow mosaic (BYMV), potato virus Y, tobacco etch, pea seed-borne mosaic, iris severe mosaic, iris mild mosaic and asparagus virus- 1 potyviruses were used as immun- ogen and as antigen for screening of the hybridoma cell lines. Thirty cell lines secreting potyvirus-specific antibodies reactive in indirect antigen-coated plate (ACP-) ELISA were selected lbr detailed analysis. All 30 MAbs reacted with at least one strain of BYMV; 11 MAbs reacted with between one and eight of the nine BYMV strains and an additional three MAbs reacted only with isolates within the BYMV subgroup (BYMV, pea mosaic virus and clover yellow vein virus). The remaining 16 MAbs reacted with a BYMV isolate and with at least one of the other 43 potyvirus isolates tested. MAb PTY 1 reacted with all 55 potyvirus isolates tested (representing at least 33 different and
distinct aphid-transmissible potyviruses). The poty- virus cross-reactive MAbs generally gave higher reacti- vity values in ACP-ELISA with dissociated virus than with polyclonal antibody-trapped intact virions in triple antibody sandwich ELISA (i.e. were cryptotope- specific). The BYMV strain- and virus-specific MAbs reacted strongly with both types of antigens (i.e. were metatope-specific). At least 25 distinct epitopes (12 cryptotopes and 13 metatopes) could be identified from the MAb-antigen reactivity patterns. The distribution of these epitopes between virus isolates can be used to detect and differentiate potyviruses in infected plant extracts and to examine virus architectures. Some of these epitopes are shared by potyvirus isolates not previously shown to be serologically related. The broad spectrum-reacting MAb PTY 1 recognizes a crypto- tope conserved on all of the aphid-transmissible potyviruses examined and should be a valuable tool for the detection and assay of these potyviruses.
Introduction
The potyvirus group, named after the type member potato virus Y (PVY), is the largest and economically the most important of the plant virus groups. Members of this group are responsible for important virus diseases affecting all types of vegetable, forage, fruit, ornamental and field crops (Hollings & Brunt, 1981; Milne, 1988). The potyvirus group contains over 160 definitive and possible members (Milne, 1988) which are characterized by having long (12 x 680 to 900 nm) flexuous, filamen- tous particles, containing one molecule of ssRNA of Mr 3"0x 106 to 3"5× 106 enclosed in coat protein composed of up to 2000 subunits of a single capsid polypeptide of Mr 30 × 103 to 37 x 103. All definitive and possible members induce the formation of character- istic cytoplasmic inclusions in infected cells which are serologically unrelated to their respective viral capsid
protein. All of the definitive and most of the possible potyviruses are transmitted by aphid vector species but some possible members have fungus, mite or whitefly vectors (Hollings & Brunt, 1981; Milne, 1988).
Serological relationships among definitive potyviruses are, however, highly complex and range from very close to intermediate to not detectable and do not always correlate with biological properties (Francki, 1983). Most of the definitive potyviruses have been shown to be serologically related to some degree to at least one other potyvirus but techniques used previously have failed to detect the expected serological relationships between many pairs of potyviruses. Polyclonal antisera show cross-reactivity of varying degrees with closely related viruses or strains of the virus immunogen, and usually fail to cross-react (or do so only very weakly) with more distantly related viruses.
Polyclonal antisera produced to denatured potyviruses
0000-9767 © 1991 SGM

26 R. Jordan and J. Hammond
(Shepard et al., 1974) or trypsin-treated virions (i.e. trypsin-resistant core particles and trypsin-resistant core protein; Shukla et al., 1988) do, however, show a higher degree of cross-reactivity with different potyviruses. Monoclonal antibodies (MAbs) produced to specific potyvirus members have generally proven to be specific for the immunizing virus or show only limited cross- reactivity with less related potyviruses (Dougherty et al., 1985; Gugerli & Fries, 1983; Hill et al., 1984; Hsu et al., 1988; Sherwood et al., 1987).
A portion of the research in our laboratory is concerned with understanding the molecular biological, biochemical and immunological nature of plant viruses and their gene products and processes of virus-host interactions, including producing reagents and tools for the detection and control of plant viruses. In this latter area there was a need for highly specific serological reagents for the detection and differentiation of various strains and isolates of bean yellow mosaic potyvirus (BYMV) in bulb ornamentals, especially gladiolus. The advent of hybridoma-produced MAb technology has provided the ability to produce homogeneous and biochemically defined immunological reagents with unique and extremely discriminatory abilities. One advantage of MAbs over polyclonal antisera in serologi- cal tests is that MAbs react with a single antigenic determinant (epitope) rather than with many different epitopes and/or antigens. In addition, MAbs specific to unique epitopes can be selected even when impure antigen or antigen mixtures are used as immunogen(s) as long as a target-specific assay is used to select the desired MAbs (Jordan, 1990).
The aims of this study were to produce not only discriminating BYMV-specific MAbs, but also other MAbs with broad-spectrum reactivities with members of the potyvirus group, and to investigate the pattern of antigenic relationships among potyviruses using these MAbs. Previous research with ilarvirus-specific MAbs (Jordan et al., 1985; Jordan & Aebig, 1985) has indicated that virus-specific antigenic determinants are virion surface-located and that ilarvirus group-common epitopes are located within intact virus (exposed only when virions are disrupted). For this study we therefore injected mice with mixtures of native and denatured cytoplasmic inclusions and native (intact) virions and coat protein subunits (dissociated and/or denatured virus) from up to six different isolates of BYMV and six other potyviruses.
In this report, we describe the binding properties of 30 strain-specific, virus-specific, subgroup-specific and po- tyvirus cross-reactive MAbs to intact and dissociated virus. These antibodies have been used to identify at least 25 distinct coat protein epitopes, to differentiate diverse potyviruses, to demonstrate serological relation-
ships not previously recognized, and to begin a detailed analysis of potyvirus structure. Preliminary reports have been published (Jordan & Hammond, 1986, 1988).
Methods
Virus isolates. The potyvirus isolates used, and their acronyms and sources are listed in Table 1. Most of the isolates were tested as purified virus preparations but some were tested only as virus-infected sap extracts (see Results). BYMV G, CYVV Pratt, MDMV A, MDMV B, PMV 204-1, SCMV D and StaVY isolates were tested as purified virus as supplied by the respective donors and BCMV, LMV and PRSV were tested only as sap extracts from lyophilized tissue, as supplied by American Type Culture Collection (ATCC; L. McDaniel). Virus and cytoplasmic inclusions (CI) from most of the other isolates were purified by one standard method (Hammond & Lawson, 1988) from a variety of hosts (pea, turnip, celery, tobacco, squash, soybean, Iris and Belamcanda).
Antigen preparation. Five forms of virus and CI antigens were used for immunization and/or in serological tests: native virus (virus), LiC1- dissociated virus (subunit), pH 9.5-denatured virus (pH-protein), 0.2% SDS-denatured virus (SDS-protein) and 2% SDS, 5% 2-mercaptoeth- anol-denatured virus (SDS/MCE-protein). The subunit antigen was prepared from purified virus particles by freezing them in 3 M-LiC1 for at least 16 h at - 2 0 °C (Francki & McLean, 1968). After thawing, intact virus particles and RNA were removed by centrifugation at 160000g for 1-5 h and the soluble coat protein subunit was diluted in the appropriate buffer and used immediately or stored at - 2 0 °C. The pH-protein antigen was prepared from purified virus or CI by heating at 56 to 60°C for 15min in 0-04M-Tris pH9.5. The SDS-protein antigen was prepared from purified virus (or CI) by heating at 56 to 60 °C for 15 min in 0-2% SDS. The SDS/MCE-protein was prepared from purified virus (or CI) by boiling for 5 rain in 2% SDS, 5% 2- mercaptoethanol. For immunization, SDS/MCE-protein was precipi- tated with 10 volumes acetone at - 2 0 ° C and resuspended in Tris- buffered saline (TBS; 20 mM-Tris-HC1 pH 7.5, 0.15 M-NaCI).
Immunization and hybridoma production. Each of two female BALB/c mice was immunized with admixtures of native and denatured virus and CI from up to 12 serologically related and unrelated potyvirus isolates by six intraperitoneal injections over a six month period. The first injection (day 0) contained 50 ~tg each native virus and CI from BYMV GDD in TBS (emulsified in Freund's complete adjuvant). The second injection (day 22) contained 50 ~.g each pH-protein from virus and CI from BYMV GDD in TBS [emulsified in Freund's incomplete adjuvant (FICA)]. The third injection (day 120) contained between 1 and 30 lag each pH-protein from virus and CI from BYMV GDD, IMMV and ISMV, and between 2 and 15 ~tg each native virus and CI from BYMV Wa Tulip, BYMV G82-18, PVY and AV-1 in TBS-FICA (150 ~tg total). The fourth injection (day 125) contained between 2 and 50 p.g each pH-protein from virus and CI from the same seven isolates used in injection 3, plus PSbMV and TEV, in TBS-FICA (170 lag total). The fifth injection (day 147) contained between 1 and 30 lag each native virus and SDS/MCE-protein and CI from the same nine isolates used in injection 4 in TBS only (270 ~tg total). The final injection (day 198) contained between 1 and 70 lag each pH-protein from virus and CI from the same nine isolates used in injections 4 and 5, plus BYMV Goldfield, BYMV 49er and BYMV Ideal A, in TBS only (400 lag total).
Five days after the final immunization (day 203), one mouse was killed, the spleen was removed, the splenocytes were fused with P3/NS1/1-AG4-1 myeloma cells ( 'NSI') with 45% polyethylene glycol as the fusogen and cultured at low cell density in RPMI 1640 medium

Potyv irus epitopes 27
T a b l e 1. Potyvirus isolates used in this s tudy
Virus Acronym/isolate Source/donor*
Bean yellow mosaic wrus Bean yellow mosaic wrus Bean yellow mosaic wrus Bean yellow mosaic wrus Bean yellow mosaic wrus Bean yellow mosaic virus Bean yellow mosaic wrus Bean yellow mosaic virus Bean yellow mosaic wrus Bean yellow mosaac v~rus Bean yellow mosaic virus Asparagus virus 1 Bean common mosaic virus Carnation vein mottle virus Celery mosaic virus Cowpea aphid-borne mosaic virus Clover yellow vein virus Iris mild mosaic virus Iris severe mosaic virus Lettuce mosaic virus Maize dwarf mosaic virus Maize dwarf mosaic virus Papaya ringspot virus Pea mosaic virus Pea mosaic virus Pea seed-borne mosaic virus Pepper mottle virus Plum pox virus Plum pox virus Pokeweed mosaic virus Potato virus A Potato virus Y Potato virus Y Soybean mosaic virus Statice virus Y Sugarcane mosaic virus Sweet potato feathery mottle virus Sweet potato feathery mottle virus Sweet potato latent virus Tobacco etch virus Tobacco etch virus Tobacco vein mottle virus Tulip breaking virus Tulip chlorotic blotch virus Turnip mosaic virus Turnip mosaic virus Vallota mosaic virus Watermelon mosaic virus II Zucchini yellow mosaic virus
BYMV GDD BYMV G82-18 BYMV G82-29 BYMV 49er BYMV Sierra Snow BYMV Goldfield BYMV Ice Cap BYMV Ideal A BYMV Masd. 2 BYMV Wa Tulip BYMV G AV-1 N BCMV PV25 CVMV CelMV PV374 CpAbMV CYVV Pratt IMMV ISMV LMV PV63 MDMV A MDMV B PRSV-W1 PV380 PMV 204-1 PMV PV89 PSbMV PVI84 PeMV NC165 PPV PV286 PPV PoMV PVA PVY-3 PVY-C SMV PV94 StaVY SCMV D SPFMV Common SPFMV lbadan SPLV TEV NAT TEV PV69 TVMV TBV TCBV TuMV PVI34 TuMV Linc 2 VaMV WMV II ZYMV
Gladiolus, Utah, U.S.A. Gladiolus, Ore., U.S.A. Gladiolus, Ore., U.S.A. Gladiolus, Ca., U.S.A. Gladiolus, Ca., U.S.A. Gladiolus, Ca., U.S.A. Gladiolus, Fla., U.S.A. Iris, Wash., U.S.A. Orchid, Pa., U.S.A. Tulip, Wash., U.S.A. R. H. Lawson, Md., U.S.A. G. Mink, Wash., U.S.A. ATCC, Md., U.S.A. Dianthus, Md., U.S.A. ATCC, Md., U.S.A. G. Thottappilly, Nigeria R. H. Lawson, Md., U.S.A. Iris, Wash., U.S.A. Iris, Wash., U.S.A. ATCC, Md., U.S.A. A. G. Gillaspie, Md., U.S.A. A. G. Gillaspie, Md., U.S.A. ATCC, Md., U.S.A. R. H. Lawson, Md., U.S.A. ATCC, Md., U.S.A. ATCC, Md,, U.S.A. J. W. Moyer, N.C., U.S.A. ATCC, Md., U.S.A. S. Hurtt, Md., U.S.A. Pokeweed, Md., U.S.A. R. W. Goth, Md., U.S.A. R. W. Goth, Md., U.S.A. V. D. Damsteegt, Md., U.S.A. ATCC, Md., U.S.A. R. H. Lawson, Md., U.S.A. A. G. Gillaspie, Md., U.S.A. J. W. Moyer, N.C., U.S.A. G. Thottappilly, Nigeria J. W. Moyer, N.C., U.S.A. W. G. Dougherty. Fla., U.S.A. ATCC, Md., U.S.A. K. M. Franklin, Ken., U.S.A. A. F. L. M. Derks, Netherlands W. P. Mowat, United Kingdom ATCC, Md., U.S.A. Tulip, Wash., U.S.A. Vallota, Md., U.S.A. J. Poulston, Ca., U.S.A. J. A. Dodds, Ca., U.S.A.
* Potyviruses, except CVMV, identified only by host, were collected by J. Hammond.
containing 15~ foetal bovine serum and HAT medium, essentially as described previously (Jordan et al., 1989; Jordan, 1990).
Hybridoma selection and initial screening assays. Hybridomas produc- ing potyvirus antigen-specific antibodies were detected by indirect antigen-coated plate (ACP-) EL1SA (AI Moudallal et al., 1984; procedure 1) beginning 10 days after fusion. Flexible PVC microtitre plates (Dynatech) were coated (100~tl per well) with one of four different antigen preparations, each containing a 1 : 1 mixture of native and SDS-protein antigens in TBS, by incubation for 2 to 3 h at 20 to 25 °C. Antigen preparation A consisted of an admixture of virus antigens from BYMV GDD, BYMV Goldfield, BYMV 49er, BYMV G82-18, BYMV Ideal A, BYMV Wa Tulip, PSbMV, SMV, TEV and
PVY (4.8 ~tg antigen per well). Antigen preparation B consisted of an admixture of CI antigens from the same potyvirus isolates used in A (2 ~tg antigen per well). Antigen preparation C consisted of an admixture of virus and CI antigens from ISMV, IMMV and AV-I (2.4 ~g antigen per well). Antigen preparation D contained protein material from uninfected pea, tobacco, Iris and Belamcanda plants prepared in the same manner as the viral immunogen preparations (5 ~tg antigen per well). Final SDS concentrations in all cases were less than 0.004~. After antigen coating, the wells were washed three times with TBS containing 0.05~ Tween 20. The remaining binding sites on the plastic were saturated by incubation (1 hr, 23 °C) with TBS containing 1% dry milk and 0.5% bovine serum albumin (BSA). This was replaced with a l : 5 to 1 : 10 dilution [in TBS, 0' 1% dry milk, 0-05 %

28 R. Jordan and J. Hammond
BSA (TBS-B)] of hybridoma culture supernatant. After overnight incubation at 6 °C the wells were washed as above and antigen-specific antibodies were detected with a goat anti-mouse IgG, IgA, IgM alkaline phosphatase conjugate (Kirkegaard and Perry) at 125 ng/ml in TBS-B (3 h, 23 °C). The plates were washed again as above and substrate (1 mg/ml p-nitrophenyl phosphate in 0.1 M-diethanolamine, 1 mM-Mg2CI pH9.8) was added. Absorbance values (A40s) were measured in a microplate ELISA reader after between 1 and 4 h at 23 °C and again after subsequent overnight incubation at 6 °C. Growth medium and cell culture medium from NSI myeloma cells were used as negative controls in the assays, and immune serum and ascites fluids from the mouse used for cell fusion was used as a positive control.
Potyvirus antigen-specific antibody-producing hybridoma cell lines were cloned to single cells by limiting dilution (Coller & Coller, 1983). The class and subclass of the MAbs were identified by Ouchterlony immunodiffasion and indirect ELISA with rabbit antisera specific for mouse heavy and light chains (Litton Bionetics).
Antigen specificity and cross-reactivity assays. The antigen specificity of the MAbs was tested in ACP-ELISA and triple antibody sandwich (TAS-) ELISA. The antigens for these assays were purified intact virus, subunit, SDS-protein and SDS/MCE-protein prepared from BYMV GDD as described above. For ACP-ELISA, polystyrene (PS) microtitre plates were coated by incubation (2 to 3 h, 23 °C) with virus antigen diluted in 0.05 ra-sodium carbonate-bicarbonate buffer (coat- ing buffer) at pH 9.6 (2 gg/ml, 100 ~tl per well). For TAS-ELISA (A1 Moudallal et al., 1984; procedure 3), PVC plates were first incubated (16 h, 6 °C) with polyclonal rabbit antibodies (1 : 800 in TBS) produced against BYMV GDD (Hammond & Lawson, 1988) before blocking with TBS, 1% dry milk, 0.5% BSA and subsequent incubation (3 h, 23 °C) with virus antigens diluted in TBS. All subsequent ELISA steps (culture supernatants, anti-mouse conjugate and substrate) were as described above.
The cross-reactivities of the MAbs with the different purified potyvirus isolates were determined in ACP-ELISA on PS plates using intact virus and/or SDS-protein diluted in coating buffer as described above. Plant samples were prepared by extracting fresh leaf tissue in coating buffer containing 2% polyvinylpyrrolidone (PVP; Mr 40000) (10 ml/g leaf), incubating for 1 to 2 h at 20 °C, centrifuging at 10000g for 10 min, and then making a 10-fold dilution of the supernatant in coating buffer for use in ACP-ELISA on PS plates as described (final dilution, l : 100). Sap extracts from lyophilized leaf tissue from the ATCC (BYMV GDD, BCMV, LMV, PRSV and PPV; generously provided by Larry McDaniel) were prepared by extracting the contents of a vial (0.02 to 0.05 g) in 1.2 ml coating buffer, 2% PVP, clarifying as above and then making a 20-fold dilution in coating buffer for analysis by ACP-ELISA as above (final dilution, 1:480 to 1:1200).
All of the selected MAbs were used (in duplicate) at the same time against each test antigen in a minimum of three independent assays.
Twenty four cell lines produced antibodies reactive with CI antigen preparat ions only and the remaining 49 cell lines produced antibodies reactive to virus antigen preparations. The 24 CI-react ive hybr idomas and six of the virus-reactive hybr idomas did not remain stable through propagat ion, cryostorage and subsequent clon- ing. Culture supernatants f rom the remaining 43 hybri- doma cell lines, designated P T Y MAbs, were evaluated in a number of different kinds of serological assays. The character izat ion o f 30 o f these P T Y M A b s and their use in potyvirus antigenic analysis will be presented here.
Antigen specificity assays
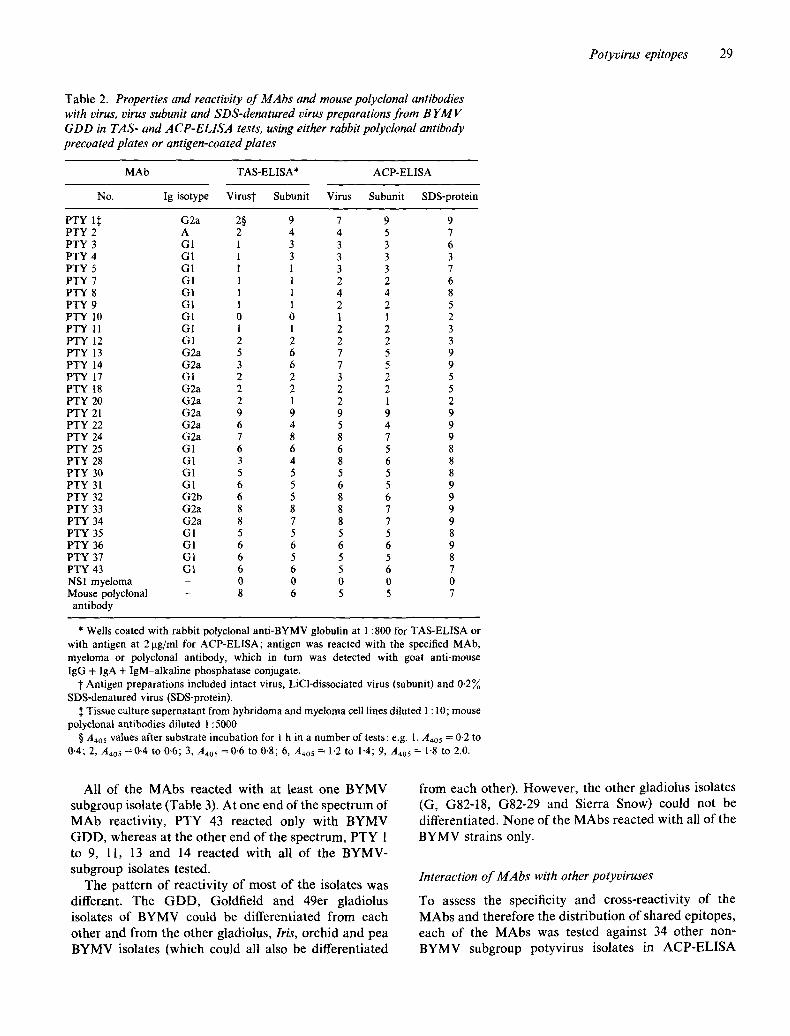
The ability of the P T Y M A b s to detect epitopes on intact virus, virus coat protein subunit or detergent-denatured virus preparat ions was tested in A C P - and TAS-ELISA. The results of isotype analysis of the PTY M A b s and their reactivities with the different virus preparat ions are presented in Table 2. Most M A b s detected intact B Y M V G D D trapped by rabbit polyclonal an t ibody in TAS- ELISA. M A b s P T Y 13 to 43 generally were more reactive with ant ibody- t rapped virus and coat protein subunit than PTY 1 to 12. All MAbs had equal or greater reactivity with viral antigens coated directly onto P S microti tre plates in A C P - E L I S A than with ant ibody- t rapped antigens. In these tests, P T Y 1 to 12 gave A4os values three- to ninefold greater with plate- trapped subunit or SDS-protein than with an t ibody- t rapped intact virus, whereas the other 19 M A b s reacted equally well with all ant igen preparat ions in bo th types of tests. Most o f the M A b s had greater reactivi ty with SDS- protein than with virus-subunit and/or intact virus. React ions of the M A b s with pH-prote in and S D S / M C E - protein were similar to SDS-protein reactions (data not shown). In other tests, where compar isons were made between reactions o f M A b s with intact virus diluted in TBS for ant igen-coat ing of P V C plates versus dilution in coating buffer for incubat ion on PS plates, the coating buffer-PS combina t ion generally produced higher A4os values for all o f the MAbs.
Results
Production of hybridomas
Of the 1728 wells into which the products o f the fusion were plated, 767 contained hybr idoma colonies. In the initial screening, 73 hybr idomas produced antibodies reactive with at least one of the viral ant igen prepara- tions A, B or C, but showed no reactivity with control preparat ion D. Another 21 hybr idomas produced antibo- dies reactive with all four preparat ions and were not tested further.
Interaction of MAbs with B Y M V subgroup isolates
Based on both molecular hybridizat ion and double- ant ibody sandwich ELISA, the bean yellow mosaic potyvirus subgroup contains three members , (BYMV, C Y V V and PMV), each having many strains (Barnett et al., 1987). Since an addit ional aim of this study was to produce serological reagents for the identification and differentiation o f strains of B Y M V isolated f rom various bulb crops (such as gladiolus, tulip and Iris), twelve BYMV-subgroup isolates were tested in A C P - E L I S A using the P T Y MAbs.
Potyvirus epitopes 29
Table 2. Properties and reactivity of MAbs and mouse polyclonal antibodies with virus, virus subunit and SDS-denatured virus preparations from B YMV GDD in TAS- and ACP-ELISA tests, using either rabbit polyclonal antibody precoated plates or antigen-coated plates
MAb TAS-ELISA* ACP-ELISA
No. Ig isotype Virust Subunit Virus Subunit SDS-protein
PTY 1~ G2a 2§ 9 7 9 9 PTY 2 A 2 4 4 5 7 PTY 3 G1 1 3 3 3 6 PTY 4 G1 1 3 3 3 3 PTY 5 GI 1 1 3 3 7 PTY 7 G1 1 1 2 2 6 PTY 8 G1 1 1 4 4 8 PTY 9 GI 1 1 2 2 5 PTY 10 G1 0 0 1 1 2 PTY 11 GI 1 1 2 2 3 PTY 12 GI 2 2 2 2 3 PTY 13 G2a 5 6 7 5 9 PrY 14 G2a 3 6 7 5 9 PTY I7 GI 2 2 3 2 5 PTY 18 G2a 2 2 2 2 5 PTY 20 G2a 2 1 2 1 2 PTY 21 G2a 9 9 9 9 9 PTY 22 G2a 6 4 5 4 9 PTY 24 G2a 7 8 8 7 9 PTY 25 G1 6 6 6 5 8 PTY 28 G1 3 4 8 6 8 PTY 30 G1 5 5 5 5 8 PTY 31 G1 6 5 6 5 9 PTY 32 G2b 6 5 8 6 9 PTY 33 G2a 8 8 8 7 9 PTY 34 G2a 8 7 8 7 9 PTY 35 G1 5 5 5 5 8 PTY 36 G1 6 6 6 6 9 PTY 37 G1 6 5 5 5 8 PTY 43 G1 6 6 5 6 7 NS1 myeloma - 0 0 0 0 0 Mouse polyclonal - 8 6 5 5 7
antibody
* Wells coated with rabbit polyclonal anti-BYMV globulin at 1:800 for TAS-ELISA or with antigen at 2 ~tg/ml for ACP-ELISA; antigen was reacted with the specified MAb, myeloma or polyclonal antibody, which in turn was detected with goat anti-mouse IgG + IgA + IgM-alkaline phosphatase conjugate.
J" Antigen preparations included intact virus, LiCl-dissociated virus (subunit) and 0.2% SDS-denatured virus (SDS-protein).
:~ Tissue culture supernatant from hybridoma and myeloma cell lines diluted 1 : 10; mouse polyclonal antibodies diluted 1:5000.
§ Aa0 s values after substrate incubation for 1 h in a number of tests: e.g. 1, A4o5 = 0.2 to 0.4; 2, A4o5 =0-4 to 0.6; 3, A405 =0.6 to 0.8; 6, A4o5 = 1.2 to 1-4; 9, A405 = 1.8 to 2.0.
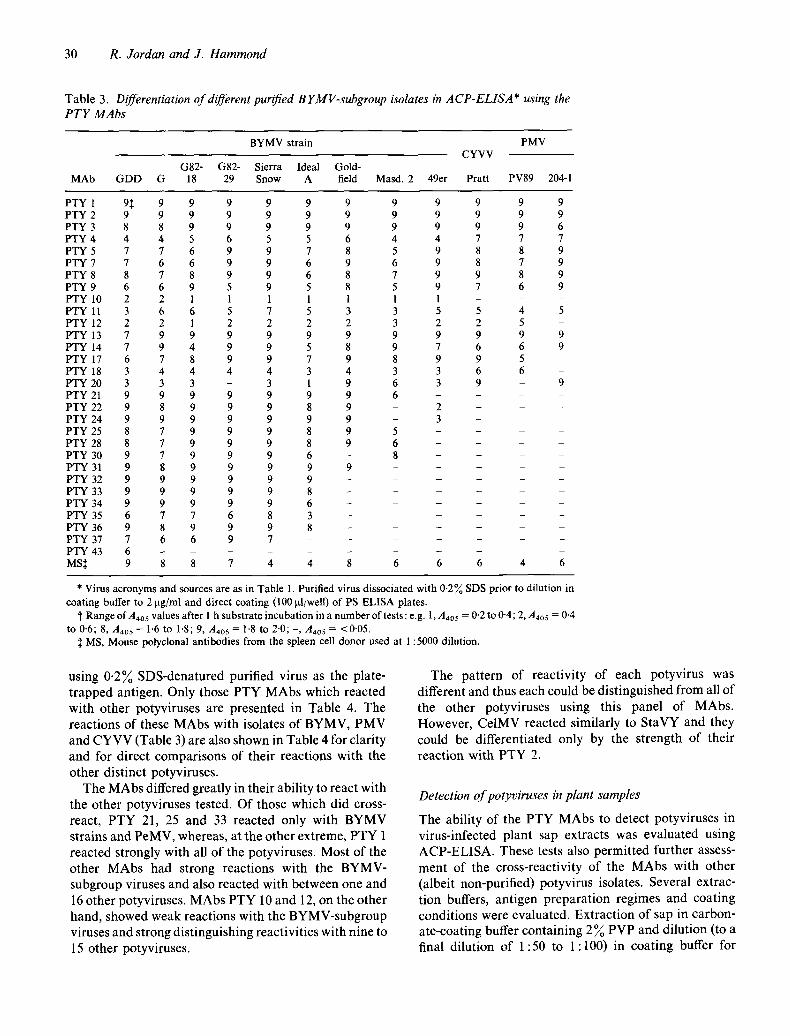
Al l o f t he M A b s r e a c t e d w i t h a t l eas t o n e B Y M V
s u b g r o u p i so la te ( T a b l e 3). A t one end o f t he s p e c t r u m o f
M A b reac t iv i ty , P T Y 43 r e a c t e d o n l y w i t h B Y M V
G D D , w h e r e a s at t he o t h e r e n d o f t he s p e c t r u m , P T Y 1
to 9, 11, 13 a n d 14 r e a c t e d w i t h all o f t he B Y M V -
s u b g r o u p i so la tes tes ted.
T h e p a t t e r n o f r e a c t i v i t y o f m o s t o f t he i so la tes was
d i f fe ren t . T h e G D D , G o l d f i e l d a n d 49er g lad io lus
i so la tes o f B Y M V could be d i f f e r e n t i a t e d f r o m e a c h
o t h e r and f r o m t h e o t h e r g lad io lus , Iris, o r c h i d a n d p e a
B Y M V isola tes ( w h i c h cou ld all a lso be d i f f e r e n t i a t e d
f r o m e a c h other) . H o w e v e r , t he o t h e r g l ad io lus i so la tes
(G, G82-18 , G82-29 and S ie r ra Snow) cou ld no t be
d i f fe ren t i a ted . N o n e o f the M A b s r e a c t e d w i t h all o f the
B Y M V s t ra ins only.
Interaction of MAbs with other potyviruses
T o assess the spec i f ic i ty a n d c ros s - r eac t i v i t y o f the
M A b s a n d t h e r e f o r e the d i s t r i b u t i o n o f s h a r e d ep i topes ,
e a c h o f t he M A b s was t es ted aga ins t 34 o t h e r non-
B Y M V s u b g r o u p p o t y v i r u s i so la tes in A C P - E L I S A
30 R. Jordan and J. Hammond
T a b l e 3. Differentiation of different purified B YMV-subgroup isolates in ACP-ELISA* using the PTY MAbs
BYMV strain PMV CYVV
G82- G82- Sierra Ideal Gold- MAb GDD G 18 29 Snow A field Masd. 2 49er Pratt PV89 204-1
PTY 1 9:~ 9 9 9 9 9 9 9 9 9 9 9 PTY 2 9 9 9 9 9 9 9 9 9 9 9 9 PTY 3 8 8 9 9 9 9 9 9 9 9 9 6 PTY 4 4 4 5 6 5 5 6 4 4 7 7 7 PTY 5 7 7 6 9 9 7 8 5 9 8 8 9 PTY 7 7 6 6 9 9 6 9 6 9 8 7 9 PTY 8 8 7 8 9 9 6 8 7 9 9 8 9 PTY 9 6 6 9 5 9 5 8 5 9 7 6 9 PTY 10 2 2 1 1 1 1 1 1 1 - - - PTY 11 3 6 6 5 7 5 3 3 5 5 4 5 PTY 12 2 2 1 2 2 2 2 3 2 2 5 - PTY 13 7 9 9 9 9 9 9 9 9 9 9 9 PTY 14 7 9 4 9 9 5 8 9 7 6 6 9 PTY 17 6 7 8 9 9 7 9 8 9 9 5 - PTY 18 3 4 4 4 4 3 4 3 3 6 6 - PTY 20 3 3 3 - 3 1 9 6 3 9 - 9 PTY 21 9 9 9 9 9 9 9 6 . . . . PTY 22 9 8 9 9 9 8 9 - 2 - - - PTY 24 9 9 9 9 9 9 9 - 3 - - - PTY 25 8 7 9 9 9 8 9 5 . . . . PTY 28 8 7 9 9 9 8 9 6 . . . . PTY 30 9 7 9 9 9 6 - 8 - - - PTY 31 9 8 9 9 9 9 9 . . . . PTY 32 9 9 9 9 9 9 . . . . . . PTY 33 9 9 9 9 9 8 . . . . . PTY 34 9 9 9 9 9 6 . . . . . . PTY 35 6 7 7 6 8 3 . . . . . . PTY 36 9 8 9 9 9 8 . . . . . . PTY 37 7 6 6 9 7 . . . . . . . PTY 43 6 . . . . . . . . . MS~ 9 8 8 7 4 4 8 6 6 6 4 6
* Virus acronyms and sources are as in Table 1. Purified virus dissociated with 0-2% SDS prior to dilution in coating buffer to 2 ~tg/ml and direct coating (100 p.l/well) of PS ELISA plates.
t Range of A405 values after I h substrate incubation in a number of tests: e.g. 1, A4o5 = 0.2 to 0-4; 2, A~o5 = 0.4 to 0.6; 8, A405 = 1.6 to 1.8; 9, Aa0s = 1.8 to 2-0; -, Aaos = <0-05.
:~ MS, Mouse polyclonal antibodies from the spleen cell donor used at 1:5000 dilution.
u s i n g 0 . 2 % S D S - d e n a t u r e d p u r i f i e d v i r u s as t h e p l a t e -
t r a p p e d a n t i g e n . O n l y t h o s e P T Y M A b s w h i c h r e a c t e d
w i t h o t h e r p o t y v i r u s e s a r e p r e s e n t e d in T a b l e 4. T h e
r e a c t i o n s o f t h e s e M A b s w i t h i so l a t e s o f B Y M V , P M V
a n d C Y V V ( T a b l e 3) a re a l so s h o w n in T a b l e 4 fo r c l a r i t y
a n d fo r d i r e c t c o m p a r i s o n s o f t h e i r r e a c t i o n s w i t h t h e
o t h e r d i s t i n c t p o t y v i r u s e s .
T h e M A b s d i f f e r ed g r e a t l y in t h e i r a b i l i t y to r e a c t w i t h
t h e o t h e r p o t y v i r u s e s t e s t ed . O f t h o s e w h i c h d i d c ross -
r eac t , P T Y 21, 25 a n d 33 r e a c t e d o n l y w i t h B Y M V
s t r a i n s a n d P e M V , w h e r e a s , a t t h e o t h e r e x t r e m e , P T Y 1
r e a c t e d s t r o n g l y w i t h all o f t h e p o t y v i r u s e s . M o s t o f t h e
o t h e r M A b s h a d s t r o n g r e a c t i o n s w i t h t h e B Y M V -
s u b g r o u p v i r u s e s a n d a lso r e a c t e d w i t h b e t w e e n o n e a n d
16 o t h e r p o t y v i r u s e s . M A b s P T Y 10 a n d 12, o n t h e o t h e r
h a n d , s h o w e d w e a k r e a c t i o n s w i t h t h e B Y M V - s u b g r o u p
v i r u s e s a n d s t r o n g d i s t i n g u i s h i n g r e a c t i v i t i e s w i t h n i n e to
15 o t h e r p o t y v i r u s e s .
T h e p a t t e r n o f r e a c t i v i t y o f e a c h p o t y v i r u s w a s
d i f f e r e n t a n d t h u s e a c h c o u l d b e d i s t i n g u i s h e d f r o m all o f
t h e o t h e r p o t y v i r u s e s u s i n g t h i s p a n e l o f M A b s .
H o w e v e r , C e l M V r e a c t e d s i m i l a r l y to S t a V Y a n d t h e y
c o u l d b e d i f f e r e n t i a t e d o n l y b y t h e s t r e n g t h o f t h e i r
r e a c t i o n w i t h P T Y 2.
Detection of potyviruses in plant samples
T h e a b i l i t y o f t h e P T Y M A b s to d e t e c t p o t y v i r u s e s in
v i r u s - i n f e c t e d p l a n t s ap e x t r a c t s w a s e v a l u a t e d u s i n g
A C P - E L I S A . T h e s e t e s t s a l so p e r m i t t e d f u r t h e r assess -
m e n t o f t h e c r o s s - r e a c t i v i t y o f t h e M A b s w i t h o t h e r
( a l b e i t n o n - p u r i f i e d ) p o t y v i r u s i so la tes . S e v e r a l e x t r a c -
t i o n buf fe r s , a n t i g e n p r e p a r a t i o n r e g i m e s a n d c o a t i n g
c o n d i t i o n s w e r e e v a l u a t e d . E x t r a c t i o n o f s a p in c a r b o n -
a t e - c o a t i n g b u f f e r c o n t a i n i n g 2 % P V P a n d d i l u t i o n ( to a
f ina l d i l u t i o n o f 1 : 5 0 to 1 : 1 0 0 ) in c o a t i n g b u f f e r for

Ta
ble
4
. D
iffer
entia
tion
of d
iver
se p
otyv
irus
es i
n A
CP
-EL
ISA
* us
ing
poty
vira
l cr
oss-
reac
tive
PT
Y M
Abs
PT
Y
MA
b
Po
tyv
iru
st
l 2
3 4
5 7
8 9
I0
tl
12
13
20
21
25
33
M
S:~
BY
MV
§
++
+
++
+
++
+
+
++
+
++
+
++
+
++
(+
) +
+
+
++
(+
) +
++
+
++
+
++
+
++
C
YV
V
++
+
++
+
++
+
+
++
+
+
++
+
+
+
++
(+
) +
++
+
+
- -
- +
+
PM
VII
+
++
+
++
+
++
+
++
+
++
+
++
+
++
+
++
-
++
(+
) +
++
+
+
- -
- +
+
IMM
V
++
+
++
+
++
+
++
+
+
+
+
-
- +
.
..
..
+
+
PV
Y-3
+
+
+
+
+
+
(+
) (+
) +
+
(+
) +
+
(+
) .
..
..
.
+
+
Pe
M¥
+
++
-
++
-
- +
++
+
- +
+
- +
++
+
++
+
+
TV
MV
+
++
-
++
+
..
..
+
++
+
++
+
+
++
.
..
.
+
TE
V¶
+
++
+
++
-
++
+
+
+
++
+
+
+
+
..
..
.
+
TC
BV
+
++
+
++
-
++
+
+
++
+
+
++
+
+
+
.
..
.
++
T
uM
V*
*
++
+
++
+
+
++
+
+
++
+
+
+
(+)
..
..
+
+
Tu
MV
Lin
c2
+
++
+
+
- +
+
++
+
+
++
-
(+)
..
..
+
+
PS
bM
V
++
+
++
+
++
+
..
..
.
++
.
..
..
+
++
T
BV
+
++
+
++
+
+
..
..
+
-
+
- (+
) -
- -
+
SM
V
+
+
+
- (+
) .
..
.
+
- -
(+)
- -
- +
P
oM
V
++
+
++
+
+
- -
- +
-
- +
-
(+)
- -
- +
++
M
DM
V
B
+
+
+
+
+
+
+
+
+
- +
+
(+
) (+
) .
..
..
.
+
MD
MV
A
+
+
+
-
+
(+)
..
..
..
..
..
(+
) S
CM
VD
+
++
+
+
++
+
+
..
..
+
+
.
..
.
+
SP
FM
VI-
'~
+
+
+
+
+
(+)
+
..
..
..
..
.
+
Sta
VY
~:~
+
++
+
++
+
+
..
..
..
..
..
..
+
C
elM
V
++
+
++
+
+
..
..
..
..
..
..
A
V-1
+
+
+
-
(+)
+
- -
- +
.
..
.
+
Cp
Ab
MV
+
+
+
-
+
+
..
..
.
+
..
..
..
(+
) Z
YM
V
+
+
+
- +
+
.
..
.
(+)
..
..
+
+
* In
dir
ect
EL
ISA
usi
ng
2 M
g/m
l pu
rifi
ed p
oty
vir
us
dis
soci
ated
wit
h 0
.2 %
SD
S p
rio
r to
dil
uti
on
in
ca
rbo
na
te b
uff
er a
nd
dir
ect
coat
ing
of
PS
EL
ISA
pla
tes.
Re
lati
ve
EL
ISA
A~0
5 v
alu
es (
1 to
3 h
su
bst
rate
in
cub
atio
n):
+
+
+
, >
1.
2; +
+
, 0
.6 t
o 1
.2;
+,
0.2
to
0'6
; (+
),
> 0
.5 a
fter
5 t
o 1
6 h
sub
stra
te i
ncu
bat
ion
an
d n
eg
ati
ve
co
ntr
ols
<0
.1;
, <
0.0
5.
t" V
iru
s a
cro
ny
ms
and
so
urc
es a
re a
s in
Ta
ble
1.
:~ M
S,
Mo
use
po
lycl
on
al a
nti
bo
die
s fr
om
th
e sp
leen
cel
l d
on
or
use
d a
t 1
:50
00
dil
uti
on
.
§ R
epre
sen
ts n
ine
stra
ins
of
BY
MV
(f
rom
Ta
ble
3).
II Rep
rese
nts
tw
o s
trai
ns
of
PM
V
(fro
m T
ab
le 3
).
¶ R
epre
sen
ts f
ou
r st
rain
s o
f T
EV
, in
clu
din
g T
EV
-HA
T
an
d T
EV
-se
ve
re s
trai
ns.
** R
epre
sen
ts s
even
str
ain
s o
f T
uM
V.
1"t
Rep
rese
nts
tw
o s
trai
ns
of
SP
FM
V.
:~:~
Rep
rese
nts
tw
o s
trai
ns
of
Sta
VY
.
32 R. Jordan and J. Hammond
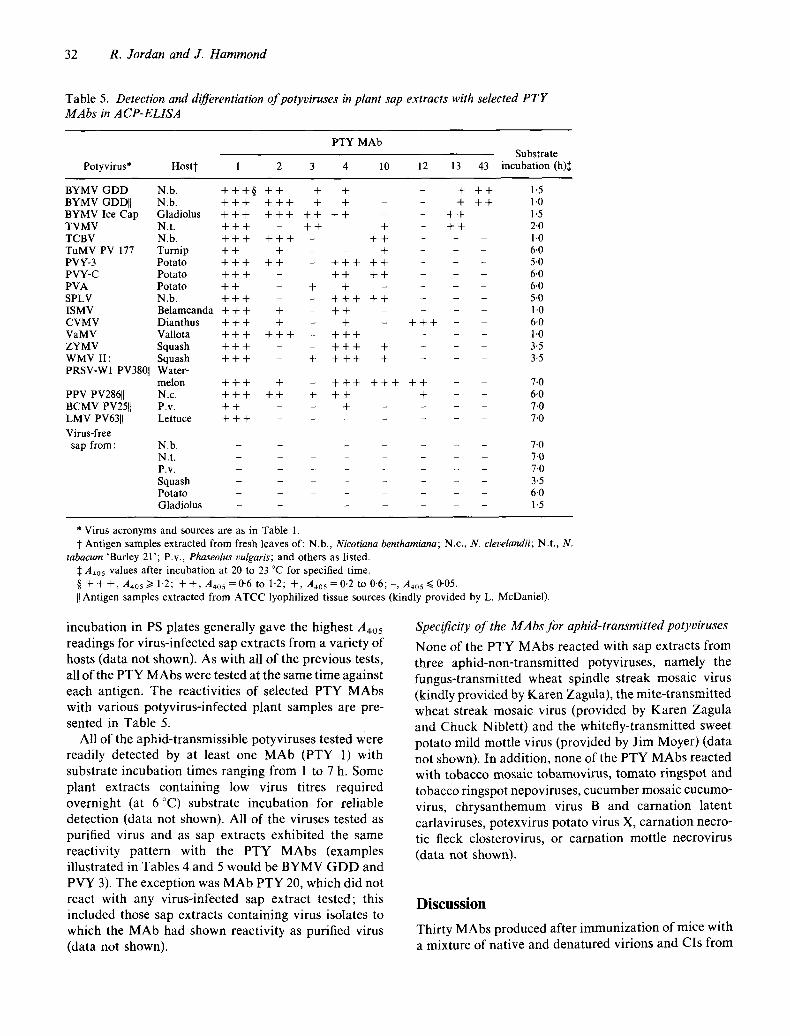
T a b l e 5. Detection and differentiation of potyviruses in plant sap extracts with selected P T Y MAbs in ACP-ELISA
PTY MAb Substrate
Potyvirus* Host~ 1 2 3 4 10 12 13 43 incubation (h):[:
B Y M V G D D N.b. + + + § + + + + - + + + 1.5 BYMV GDD[] N.b. + + + + + + + + - - + + + 1.0 B Y M V I c e C a p Gladiolus + + + + + + + + + + - - + + 1.5 TVMV N.t. + + + - + + + - + + 2-0 TCBV N.b. + + + + + + - + + - - 1.0 TuMV PV 177 Turnip + + + - - + - 6.0 PVY-3 Potato + + + + + - + + + + + - - 5-0 PVY-C Potato + + + - - + + + + - - 6.0 PVA Potato + + - + + - - 6.0 SPLV N.b. + + + - - + + + + + - - 5-0 ISMV Belamcanda + + + + + + . . . . 1.0 CVMV Dianthus + + + + + - + + + - - 6-0 VaMV Vallota + + + + + + + + + - - 1-0 ZYMV Squash + + + - + + + + - 3.5 WMV II: Squash + + + - + + + + + - - 3.5 PRSV-WI PV38011 Water-
melon + + + + + + + + + + + + - - 7.0 PPV PV28611 N.c. + + + + + + + + + - 6.0 BCMV PV25[[ P.v. + + - + . . . . 7.0 LMV PV6311 Lettuce + + + . . . . . . 7.0 Virus-free
sap from: N.b. - . . . . . 7.0 N.t. - . . . . 7-0 P.v. - . . . . . . . 7.0 Squash . . . . . . . . 3.5 Potato . . . . . . . 6.0 Gladiolus . . . . . . . 1.5
* Virus acronyms and sources are as in Table 1. ~" Antigen samples extracted from fresh leaves of: N.b., Nicotiana benthamiana; N.c., N. clevelandii; N.t., N.
tabacum 'Burley 21'; P.v., Phaseolus vulgaris; and others as listed. A~05 values after incubation at 20 to 23 °C for specified time.
§ + + + , A4os ~> 1.2; + + , A~0s =0.6 to 1.2; + , A405 = 0,2 to 0.6; -, A405 ~< 0-05. II Antigen samples extracted from ATCC lyophilized tissue sources (kindly provided by L. McDaniel).
i n c u b a t i o n in PS p l a t e s g e n e r a l l y g a v e t h e h i g h e s t A4o 5
r e a d i n g s fo r v i r u s - i n f e c t e d s a p e x t r a c t s f r o m a v a r i e t y o f
h o s t s ( d a t a n o t s h o w n ) . A s w i t h all o f t h e p r e v i o u s t es t s ,
al l o f t h e P T Y M A h s w e r e t e s t e d a t t h e s a m e t i m e a g a i n s t
e a c h a n t i g e n . T h e r e a c t i v i t i e s o f s e l e c t e d P T Y M A b s
w i t h v a r i o u s p o t y v i r u s - i n f e c t e d p l a n t s a m p l e s a r e p r e -
s e n t e d in T a b l e 5.
Al l o f t h e a p h i d - t r a n s m i s s i b l e p o t y v i r u s e s t e s t e d w e r e
r ead i l y d e t e c t e d b y a t l e a s t o n e M A b ( P T Y 1) w i t h
s u b s t r a t e i n c u b a t i o n t i m e s r a n g i n g f r o m 1 to 7 h. S o m e
p l a n t e x t r a c t s c o n t a i n i n g low v i r u s t i t r e s r e q u i r e d
o v e r n i g h t (a t 6 °C) s u b s t r a t e i n c u b a t i o n for r e l i a b l e
d e t e c t i o n ( d a t a n o t s h o w n ) . Al l o f t h e v i r u s e s t e s t e d as
p u r i f i e d v i r u s a n d as s a p e x t r a c t s e x h i b i t e d t h e s a m e
r e a c t i v i t y p a t t e r n w i t h t h e P T Y M A b s ( e x a m p l e s
i l l u s t r a t e d in T a b l e s 4 a n d 5 w o u l d b e B Y M V G D D a n d
P V Y 3). T h e e x c e p t i o n w a s M A b P T Y 20, w h i c h d i d n o t
r e a c t w i t h a n y v i r u s - i n f e c t e d s a p e x t r a c t t e s t e d ; t h i s
i n c l u d e d t h o s e s ap e x t r a c t s c o n t a i n i n g v i r u s i so l a t e s to
w h i c h t h e M A b h a d s h o w n r e a c t i v i t y as p u r i f i e d v i r u s
( d a t a n o t s h o w n ) .
Specificity of the MAbs for aphid-transmitted potyviruses
N o n e o f t he P T Y M A b s r e a c t e d w i t h s a p e x t r a c t s f r o m
t h r e e a p h i d - n o n - t r a n s m i t t e d p o t y v i r u s e s , n a m e l y t h e
f u n g u s - t r a n s m i t t e d w h e a t s p i n d l e s t r e a k m o s a i c v i r u s
( k i n d l y p r o v i d e d by K a r e n Z a g u l a ) , t h e m i t e - t r a n s m i t t e d
w h e a t s t r e a k m o s a i c v i ru s ( p r o v i d e d b y K a r e n Z a g u l a
a n d C h u c k N i b l e t t ) a n d t h e w h i t e f l y - t r a n s m i t t e d s w e e t
p o t a t o m i l d m o t t l e v i r u s ( p r o v i d e d b y J i m M o y e r ) ( d a t a
n o t s h o w n ) . I n a d d i t i o n , n o n e o f t h e P T Y M A b s r e a c t e d
w i t h t o b a c c o m o s a i c t o b a m o v i r u s , t o m a t o r i n g s p o t a n d
t o b a c c o r i n g s p o t n e p o v i r u s e s , c u c u m b e r m o s a i c c u c u m o -
v i rus , c h r y s a n t h e m u m v i r u s B a n d c a r n a t i o n l a t e n t
c a r l a v i r u s e s , p o t e x v i r u s p o t a t o v i r u s X , c a r n a t i o n n e c r o -
t ic f leck c l o s t e r o v i r u s , o r c a r n a t i o n m o t t l e n e c r o v i r u s
( d a t a n o t s h o w n ) .
Discussion
T h i r t y M A b s p r o d u c e d a f t e r i m m u n i z a t i o n o f m i c e w i t h
a m i x t u r e o f n a t i v e a n d d e n a t u r e d v i r i o n s a n d C I s f r o m

Potyvirus epitopes 33
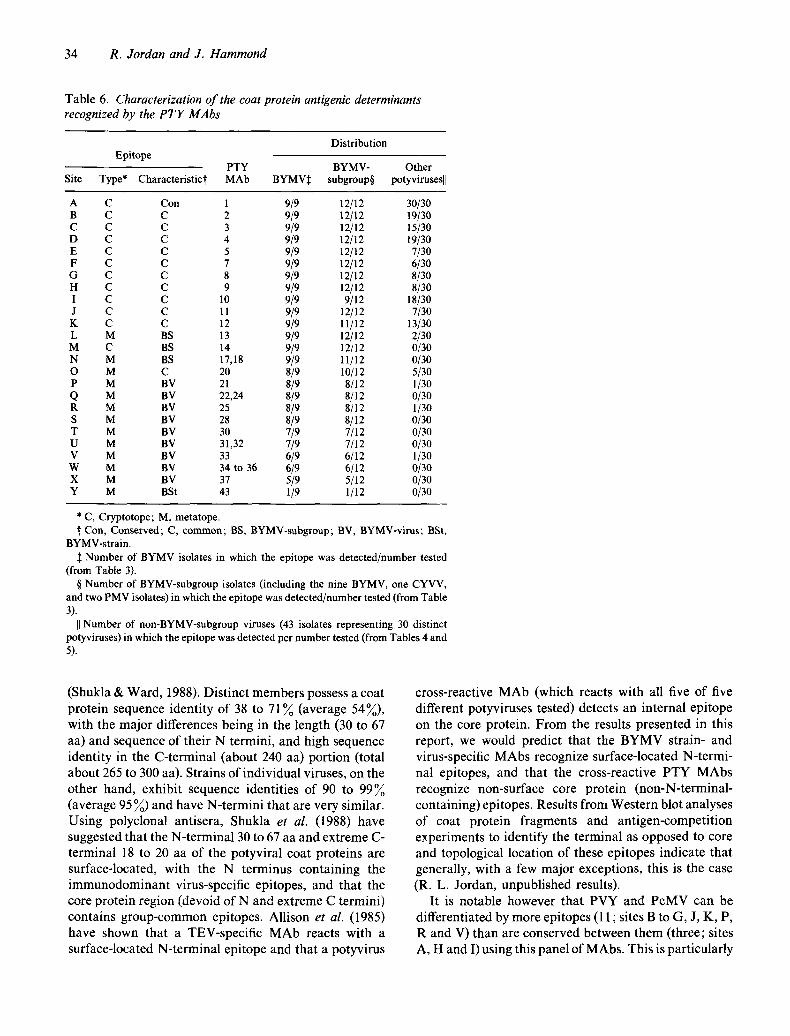
up to 12 different potyvirus isolates have been partially characterized and used to examine potyvirus antigenic relationships. Epitopes specific to BYMV strains, BYMV, the BYMV subgroup, or common to many distinct potyviruses were identified using these antibodies.
One MAb, PTY 43, was specific to an epitope present on only one of the nine BYMV isolates used (BYMV GDD). Ten MAbs (PTY 22, 24, 28, 30, 31, 32 and 34 to 37) were specific to epitopes shared by between five and eight of the nine BYMV isolates, whereas MAbs PTY 14, 17 and 18 were specific to epitopes common to 11 or 12 of the 12 BYMV subgroup isolates (which include BYMV, CYVV and PMV strains) (Table 3). Three MAbs (PTY 21, 25 and 33) react with three unique epitopes shared between specific isolates of BYMV and PeMV. MAb PTY 13 detects an additional epitope shared between the 12 BYMV-subgroup isolates and TVMV and TCBV. The remaining 12 MAbs (PTY 1 to 5, 7 to 12, and 20) react with epitopes present on a BYMV-subgroup isolate, as well as on at least one of the
other potyvirus isolates tested (Tables 4 and 5). PTY 1 reacted with an epitope occurring on all 55 potyviruses isolates (representing 33 distinct viruses) presented here; in results to be presented elsewhere it has reacted with over 100 isolates, representing 40 distinct potyviruses.
The type of antigenic determinants recognized by the PTY MAbs can tentatively be determined from the results of the antigen specificity tests using intact and disrupted/denatured virions in the ACF- and TAS- ELISA tests. The BYMV strain- and virus-specific MAbs (including the BYMV-PeMV cross-reacting MAbs) PTY 21 to 43 generally reacted equally well with intact virions, virus subunits and denatured virus, indicating that these MAbs probably recognize antigenic determinants, metatopes, exposed on the virion surface that are also found on depolymerized subunits. The remaining subgroup-specific and cross-reactive PTY MAbs reacted best with plate-trapped (disrupted) virus, subunit and denatured virus coat protein, suggesting that these MAbs recognize antigenic sites, cryptotopes, found not on the virion surface but rather on coat protein subunit sites found within the intact virion. None of the MAbs appears to react with neotopes, sites found only on intact virions. MAb PTY 20 was the only antibody that did not react with both purified virus and virus-infected sap extracts. This antibody apparently reacts with an antigenic determinant that has been modified and/or exposed during virus purification. A similar situation has been reported for a PVA-specific MAb (Boonekamp & Pomp, 1986).
--Antigens that exhibit similar or identical reactivity patterns with a group of test antibodies can be grouped together and, further, can be differentiated from those
groups of antigens that show markedly different reacti- vity patterns. The different patterns of reactivity of the different potyviruses using this panel of MAbs (Tables 3, 4 and 5) indicate that most of the epitopes detected are distinct. Approximately 25 different potyviral coat protein epitopes can be defined based on their antigen reactivity patterns with the 30 PTY MAbs and the 55 different potyvirus isolates (Table 6). PTY MAbs 17 and 18, 22 and 24, 31 and 32, and 34, 35 and 36 define four epitopes, whereas the remaining 21 PTY MAbs each define unique antigenic determinants.
The MAb-antigen reactivity patterns presented (Tables 3 to 5) indicate a wide range of serological interrelatedness among the tested potyvirus isolates. The results presented here indicate serological relationships not detected in previous studies with polyclonal antisera, and many not reported in prior studies with cross- reactive MAbs (Dougherty et al., 1985; Hsu et al., 1988). For example, Mowat (1985) found no relationship between TCBV and TBV, TEV or PVY, or between TBV and TEV. In this study eight epitopes (A, B, E, F to I, and K) were found to be common to TCBV,TEV and PVY, with epitopes A, B, I and K also being found on TBV (Tables 4 and 6). Purcifull et al. (1975) reported that PeMV was unrelated to TuMV, whereas we found four epitopes common to PeMV and TuMV (A, H, I and K). Sun et al. (1974) found no relationship between TVMV and PVY, TEV, CYVV or SMV. Data illustrated in Tables 4 and 6 show two epitopes (A and I) common to all five of these viruses; epitopes A and J common to TVMV, TEV, PVY and CYVV; epitopes A and K shared by TVM¥, TEV and CYVV; and epitopes A and L present in both TVMV and CYVV.
However, no conclusions can be drawn from this work as to the closeness of the serological relationships among the viruses studied. The range and degrees of cross- reactions found with this panel of MAbs do, however, support the observation that the serological relationships among the aphid-transmitted potyviruses are indeed numerous and complex (HoUings & Brunt, 1981; Milne, 1988).
No obvious correlations were observed between epitopes conserved between different viruses and the host ranges of the viruses. For example, five viruses naturally infecting solanaceous hosts (PeMV, PVA, PVY, TEV and TVMV) share only the epitope recog- nized by PTY 1. Similarly, six viruses naturally infecting legumes (BYMV, CpAbMV, CYVV, PMV, PSbMV and SMV) share only the group-conserved PTY 1 epitope, although the BYMV-subgroup viruses (BYMV, CYVV and PMV) share several metatopes and cryptotopes.
Coat protein amino acid (aa) sequence data (now available for 17 strains of eight distinct potyviruses) have been used to differentiate distinct viruses and strains
34 R. Jordan and J. Hammond
Table 6. Characterization of the coat protein antigenic determinants recognized by the P T Y MAbs
Epitope Distribution
PTY BYMV- Other Site Type* Characteristict MAb BYMV:~ subgroup§ potyvirusesll
A C Con 1 9/9 12/12 30/30 B C C 2 9/9 12/12 19/30 C C C 3 9/9 12/12 15/30 D C C 4 9/9 12/12 19/30 E C C 5 9/9 12/12 7/30 F C C 7 9/9 12/12 6/30 G C C 8 9/9 12{12 8/30 H C C 9 9/9 12/12 8/30 I C C 10 9/9 9/12 18/30 J C C 11 9/9 12/12 7/30 K C C 12 9/9 11/12 13/30 L M BS 13 9/9 12/12 2/30 M C BS 14 9/9 12/12 0/30 N M BS 17,18 9/9 11/12 0/30 O M C 20 8/9 10/12 5/30 P M BV 21 8/9 8/12 1/30 Q M BV 22,24 8/9 8/12 0/30 R M BV 25 8/9 8/12 1/30 S M BV 28 8/9 8/12 0/30 T M BV 30 7/9 7/12 0/30 U M BV 31,32 7/9 7/12 0/30 V M BV 33 6/9 6/12 1/30 W M BV 34to36 6/9 6/12 0/30 X M BV 37 5/9 5/12 0/30 Y M BSt 43 1/9 1/12 0/30
* C, Cryptotope; M, metatope. ~" Con, Conserved; C, common; BS, BYMV-subgroup; BV, BYMV-virus; BSt,
BYMV-strain. :~ Number of BYMV isolates in which the epitope was detected/number tested
(from Table 3). § Number of BYMV-subgroup isolates (including the nine BYMV, one CYVV,
and two PMV isolates) in which the epitope was detected/number tested (from Table 3).
I] Number of non-BYMV-subgroup viruses (43 isolates representing 30 distinct potyviruses) in which the epitope was detected per number tested (from Tables 4 and 5).
(Shukla & Ward, 1988). Distinct members possess a coat protein sequence identity of 38 to 71 ~ (average 54~), with the major differences being in the length (30 to 67 aa) and sequence of their N termini, and high sequence identity in the C-terminal (about 240 aa) portion (total about 265 to 300 aa). Strains of individual viruses, on the other hand, exhibit sequence identities of 90 to 99~ (average 95 ~) and have N-termini that are very similar. Using polyclonal antisera, Shukla et al. (1988) have suggested that the N-terminal 30 to 67 aa and extreme C- terminal 18 to 20 aa of the potyviral coat proteins are surface-located, with the N terminus containing the immunodominant virus-specific epitopes, and that the core protein region (devoid of N and extreme C termini) contains group-common epitopes. Allison et al. (1985) have shown that a TEV-specific MAb reacts with a surface-located N-terminal epitope and that a potyvirus
cross-reactive MAb (which reacts with all five of five different potyviruses tested) detects an internal epitope on the core protein. From the results presented in this report, we would predict that the BYMV strain- and virus-specific MAbs recognize surface-located N-termi- nal epitopes, and that the cross-reactive PTY MAbs recognize non-surface core protein (non-N-terminal- containing) epitopes. Results from Western blot analyses of coat protein fragments and antigen-competition experiments to identify the terminal as opposed to core and topological location of these epitopes indicate that generally, with a few major exceptions, this is the case (R. L. Jordan, unpublished results).
It is notable however that PVY and PeMV can be differentiated by more epitopes (11 ; sites B to G, J, K, P, R and V) than are conserved between them (three; sites A, H and I) using this panel of MAbs. This is particularly

Potyvirus epitopes 35
surprising as several authors have suggested that PeMV is actually a strain of PVY, based on coat protein amino acid and 3' non-coding sequence identities (Shukla & Ward, 1988; Shukla et al., 1988b; van der Vlugt et al., 1989; Robaglia et al., 1989; Turpen, 1989; Frenke 1 et al., 1989). The reason for such differentiation by these MAbs is open to speculation but is probably not closely related to host range or vector specificity as eight of the 11 differentiating epitopes are cryptotopes located in the core protein region of the coat protein, where there is the greatest amino acid sequence identity between potyviruses.
The central and C-terminal portions of the BYMV GDD coat protein, whose gene has been cloned, sequenced and expressed as a fusion protein in Escheri- chia call (Hammond & Hammond, 1989), have consider- able identity with other sequenced potyvirus coat proteins. MAb PTY 1 and several of the cross-reactive PTY MAbs react with the bacterially expressed protein (Hammond & Hammond, 1989; R. L. Jordan & J. Hammond, unpublished results). Comparison of amino acid sequences and MAb cross-reactivities with purified virus and deletion mutants of the cloned coat protein gene are currently being used as another means of defining the epitopes with which the MAbs react.
Several of the PTY MAbs seem potentially useful for routine virus detection and identification. For example, MAbs PTY 22, 28, 30, 31, 36, 37 and 43 could be used for the detection and discrimination of strains of BYMV in gladiolus, iris and orchids. Admixtures of MAbs PTY 14, 17, 18 and one or more of the BYMV-specific MAbs could be useful in the detection of a wide rang e of BYMV-subgroup viruses in a variety of plant species. At the other extreme of reaction specificity, the broad spectrum reactivity of MAb PTY 1 makes it an extremely valuable tool in quarantine, diagnostic and screening applications for the detection of aphid- transmissible potyviruses.
Many of these MAbs could also be useful as probes in studies concerning coat protein structure-function analysis, including for example virus-helper compo- nent-vector interactions, coat protein-protein or pro- tein-RNA interactions, and in defining specific protein determinants involved in coat protein-mediated virus cross-protection.
We wish to acknowledge gratefully and thank Mary Ann Guaragna for excellent technical assistance, Peggy Hall for typing the manu- script, Larry McDaniel and Larry Sikora for manuscript review and V. D. Damsteegt, A. F. L. M. Derks, J. A. Dodds, W. G. Dougherty, K. M. Franklin, A. G. Gillaspie, R, W. Garb, S. Hurtt, R. H. Lawson, L. McDaniel, G. Mink, J. W. Mayer, W. P. Mowat, T. P. Pirone, J. Poulson and G. Thottapilly for providing virus isolates. The use of trade, firm, or corporation names in this article does not imply the endorsement or approval by the USDA-ARS of any product to the exclusion of others that may be suitable.
References ALLISON, R. F., DOUGHERTY, W. G., PARKS, T. D., WILLIS, L.,
JOHNSTON, R. E., KELLY, M. & ARMSTRONG, F. B. (1985). Biochemical analysis of the capsid protein gene and capsid protein of tobacco etch virus: N-terminal amino acids are located on the virion's surface. Virology 147, 309-316.
AL MOUDALLAL, Z., ALTSCHUH, D., BRIAND, J. P. & VAN REGENMOR- TEL, M. H. V. (1984). Comparative sensitivity of different ELISA procedures for detecting monoclonal antibodies. Journal of Immuno- logical Methods 68, 35-43.
BARNETI, O. W., RANDLES, J. W. & BURROWS, P. M. (1987). Relationships among Australian and North American isolates of the bean yellow mosaic potyvirus subgroup. Phytopathology 77, 791-799.
BOONEKAMP, P. M. & POMP, H. (1986). Problems concerning the production of monoclonal antibodies for plant diagnostic purposes. Acta Horticulturae 177, 103-109.
COLLER, H. A. 8£ COLLER, B. S. (1986). Poisson statistical analysis of repetitive subcloning by the limiting dilution technique as a way of assessing hybridoma monoclonality. Methods in Enzymology 121, 412-417.
DOUGHERTY, W. G., WILLIS, L. d~ JOHNSON, R. E. (1985). Topographic analysis of tobacco etch virus capsid protein epitopes. Virology 144, 66-72.
FRANCKI, R. I. B. (1983). Current problems in plant virus taxonomy. In A Critical Appraisal of Viral Taxonomy, pp. 63-104. Edited by R.E.F. Matthews. Boca Raton: CRC Press.
FRANCKI, R. ]. B. & MCLEAN, G. D. (1968). Purification of potato virus X and preparation of infectious ribonucleic acid by degradation with lithium chloride. Australian Journal of Biological Sciences 21, 1311- 1318.
FRENKEL, M. J., WARD, C. W. & SHUKLA, D. D. (1989). The use of 3' non-coding nucleotide sequences in the taxonomy of potyviruses: application to watermelon mosaic virus 2 and soybean mosaic virus- N. Journal of General Virology 70, 2775-2783.
GUGERLI, P. & FRIES, P. (1983). Characterization of monoclonal antibodies to potato virus Y and their use for virus detection. Journal of General Virology 64, 2471-2477
HAMMOND, J. & HAMMOND R. W. (1989). Molecular cloning, sequencing and expression in Escherichia call of the bean yellow mosaic virus coat protein gene. Journal of General Virology 70, 1961- 1974.
HAMMOND, J. & LAWSON, R. H. (1988). An improved purification procedure for preparing potyviruses and cytoplasmic inclusions from the same tissue. Journal of Virological Methods 20, 203-217.
HILL, E. K., HILL, J. H. & DURAND, D. P. (1984). Production of monoclonal antibodies to viruses in the potyvirus group: use in radioimmunoassay. Journal of General Virology 65, 525-532.
HOLLINGS, M. & BRUNT, A. A. (1981). Potyviruses. In Handbook of Plant Virus Infections and Comparative Diagnosis, pp.732-807. Edited by E. Kurstak. Amsterdam: Elsevier.
HSU, H. T., FRANSSEN, J. M., VAN DER HULST, C. T. C., DERKS, A. F. L. M. & LAWSON, R. H. (1988). Factors affecting selection of epitope specificity of monoclonal antibodies to tulip breaking potyviruses. Phytopathology 78, 1337-1340.
JORDAN, R. L. (1990). Strategy and techniques for the production of monoclonal antibodies. In Serological Detection and Identification of Plant Viral and Bacterial Pathogens. Edited by R. O. Hampton & E. Ball. St Paul, Minnesota: APS Press (in press).
JORDAN, R. L. & AEBIG, J. A. (1985). Evaluating the specific immunoreactivities of monoclonal antibodies to two plant viruses. Acta Horticulturae 164, 385-393.
JORDAN, R. L. & HAMMOND, I. (1986). Analysis of antigenic specificity of monoclonal antibodies to several potyviruses. Phytopathology 76, 1091.
JORDAN, R. L. & HAMMOND, J. (1988). Epitope specificity of strain-, virus-, subgroup-specific and potyvirus group cross-reactive mono- clonal antibodies. Phytopathology 78, 1600.
JORDAN, R. L., AEBIG, J. A. & Hsu, H. T. (1985). Reactivities of apple mosaic virus (ApMV)/Prunus necrotic ringspot virus (NRSV) monoclonal antibodies with ApMV, NRSV and other ilarviruses. Phytopathology 75, 1353.

36 R. Jordan and J. Hammond
JORDAN, R. L., KONAI, M., LEE, I.-M. & DAVIS, R. E. (1989). Species- specific and cross-reactive monoclonal antibodies to the spiroplas- mas Spiroplasma citri and S. kunkelli. Phytopathology 79, 880-887.
MILNE, R. G. (1988). Taxonomy of the rod-shaped filamentous viruses. In The Plant Viruses, vol. 4, The Filamentous Plant Viruses, pp. 3-50. Edited by R. G. Milne. New York: Plenum Press.
MOWAT, W. P. (1985). Tulip chlorotic blotch virus, a second potyvirus causing tulip flower break. Annals of Applied Biology 106, 65-73.
PURCIFULL, n. E., ZITTER, T. A. & HIEBI~RT, E. (1975). Morphology, host range, and serological relationships of pepper mottle virus. Phytopathology 65, 559-562.
ROBAGLIA, C., DURAND-TARDIF, i . , TRONCHET, i . , BOUDAZlN, G., ASTIER-MANIFACIER, S. & CASSE-DELBART, F. (1989). Nucleotide sequence of potato virus Y (N strain) genomic RNA. Journal of General Virology 70, 935-947.
SHEPARD, J. F., SECOR, G. A. & PURCIFULL, O. T. (1974). Immunochemical cross-reactivity between the dissociated capsid proteins of PVY group plant viruses. Virology .58, 464-475.
SHERWOOD, J. L., SANBORN, M. R. & KEYSER, G. C. (1987). Production of monoclonal antibodies to peanut mottle virus and their use in enzyme-linked immunosorbent assay and dot-immunobinding assay. Phytopathology 77, 1158-1161.
SHUKLA, D. D. & WARD, C. W. (1988). Amino acid sequence homology of coat proteins as a basis for identification and classification of the potyvirus group. Journal of Generul Virology 69, 2703-2710.
SHUKLA, D. D., STRIKE, P. M., TRACY, S. L., GOUGI~, K. H. & WARD, C. W. (1988a). The N and C termini of the coat proteins of potyviruses are surface-located and the N terminus contains the major virus-specific epitopes. Journal of General Virology 69, 1497- 1508.
SHUKLA, D. D., THOMAS, J. E., MCKERN, N. M., TRAC¥, S. C. & WARD, C. W. (1988b). Coat protein of potyviruses. IV. Comparison of biological properties, serological relationships, and coat protein amino acid sequences of four strains of potato virus Y. Archives of Virology 102, 207-219.
SUN, M. K. C., GOODING, G. V., PIRONE, T. P. & TOLIN, S. A. (1974). Properties of tobacco vein-mottling virus, a new pathogen of tobacco. Phytopathology 64, 1133-1136.
TURPEN, T. (1989). Molecular cloning of a potato virus Y genome: nucleotide sequence homology in non-coding regions of potyviruses. Journal of General Virology 70, 1951-1960.
VAN DER VLUGT, R., ALLEFS, S., DE HAAN, P. & GOLDBACH, R. (1989). Nucleotide sequence of the T-terminal region of potato virus yN RNA. Journal of General Virology 70, 229-233.
(Received 25 June 1990; Accepted 22 October 1990)




















