
Porin expression in clinical isolates of Klebsiella pneumoniae
Upload
independentCategory
view
0download
0
Response of bacterial isolates from Antarctic shallow sedimentstowards heavy metals, antibiotics and polychlorinated biphenyls
Angelina Lo Giudice • Patrizia Casella •
Vivia Bruni • Luigi Michaud
Accepted: 15 November 2012 / Published online: 27 November 2012
� Springer Science+Business Media New York 2012
Abstract The response of bacterial isolates from Antarctic
sediments to polychlorinated biphenyls (Aroclor 1242
mixture), heavy metal salts (cadmium, copper, mercury and
zinc) and antibiotics (ampicillin, chloramphenicol, kana-
mycin and streptomycin) was investigated. Overall, the
ability to growth in the presence of Aroclor 1242 as a sole
carbon source was observed for 22 isolates that mainly
belonged to Psychrobacter spp. Tolerance to the heavy
metals assayed in this study was in the order of Cd [Cu [ Zn [ Hg and appeared to be strictly related to the
metal concentrations, as determined during previous
chemical surveys in the same area. With regards to anti-
biotic assays, the response of the isolates to the tested
antibiotics ranged from complete resistance to total sus-
ceptibility. In particular, resistances to ampicillin and
chloramphenicol were very pronounced in the majority of
isolates. Our isolates differently responded to the presence
of toxic compounds primarily based on their phylogenetic
affiliation and secondarily at strain level. Moreover, the
high incidence of resistance either to metal or antibiotics,
in addition to the capability to grow on PCBs, confirm that
bacteria are able to cope and/or adapt to the occurrence
pollutants even in low human-impacted environments.
Keywords Heavy metal tolerance � Antibiotic resistance �PCB oxidation
Introduction
In spite of the precautionary measures taken to avoid pol-
lution, Antarctica is not exempt from a moderate anthro-
pogenic contamination. Toxic compounds can reach the
Antarctic environment through long-range transport by
mass flow in the atmosphere and water, contamination due
to improper disposal practices and/or incineration of waste
produced at research bases. This is the case, for example,
of heavy metals, antibiotics and polychlorinated biphenyls
(PCBs) (Montone et al. 2001, 2005; De Souza et al. 2006).
These latter are long-term persistent compounds synthe-
sized as mixtures during the mid-twentieth century and
sold for industrial application. Their use has been regulated
or prohibited since the 1970 s in western nations. Con-
versely, antibiotics and heavy metals are also introduced
through natural processes into the environment.
The low seawater temperature is responsible for reduc-
ing both metabolic (which makes the excretion process less
efficient) and growth rates of organisms, thus promoting
high concentrations of contaminants in the Antarctic biota
and accumulation throughout the food chain. Such con-
centrations are often comparable or even higher than those
from the Northern Hemisphere (Grotti et al. 2008). Con-
trary to heavy metals and PCBs that can have a negative
effect on all forms of life, antimicrobials mainly alter the
microbiosphere by inhibiting microbial activity and disturb
the biogeochemical circulation of biogenesis elements
(Martinez 2009).
Once in the marine environment, all these problematic
contaminants cannot be easily dispersed, thus becom-
ing localized and accumulated in sediments which are the
natural collectors of pollutants (Andrade et al. 2001;
Caroppo et al. 2006). Their fate is then strictly linked to
bacteria, as the most abundant sediment organisms, which
A. Lo Giudice (&) � P. Casella � V. Bruni � L. Michaud
Department of Biological and Environmental Sciences,
University of Messina, Viale Ferdinando Stagno d’Alcontres 31,
98166 Messina, Italy
e-mail: [email protected]
123
Ecotoxicology (2013) 22:240–250
DOI 10.1007/s10646-012-1020-2

represent the first step in the transfer of toxic compounds to
higher trophic levels (Gillan et al. 2005). Bacteria possess
genetic and biochemical capacities for remediation of
heavy metal and PCB pollution. PCBs can be transformed,
despite their recalcitrant nature, into chemical substances
by different microbial metabolic pathways, both aerobic
and anaerobic, facilitating further metabolization. The
biodegradation of certain, usually less chlorinated, PCB
congeners has been reported for several microorganisms
elsewhere in the world (Bedard et al. 1987; Kohler et al.
1988; Dercova et al. 2009). However, to date PCB-
degraders have been rarely isolated from the Antarctic
environment and they derive exclusively from seawater
(Yakimov et al. 1999; De Domenico et al. 2004; Michaud
et al. 2007). Conversely, a single work exists about the
abundance of PCB-degrading bacteria in Antarctic sedi-
ments, but no information on the isolation and identifica-
tion of these PCB-degrading microorganisms was furnished
(Tumeo and Guinn 1997).
Bacteria can adsorb, accumulate (by a passive metabo-
lism-independent or an active metabolism-dependent pro-
cess) and transform heavy metals in most food chains
(De Souza et al. 2006; Haferburg and Kothe 2007). Heavy-
metal resistance has been reported for a number of different
bacterial genera and in some cases it is present together
with antibiotic resistances. Both kinds of resistance are
becoming a global phenomenon, with resistant bacteria that
are ubiquitous and have been also detected in Antarctic
seawater (De Souza et al. 2006).
In this context, the present work was mainly aimed at
analysing the response of bacteria from Antarctic sedi-
ments to heavy metals, antibiotics and PCBs, in order to
provide further insights toward this under-investigated
topic in cold-systems.
Materials and methods
Study area and sample collection
Shallow marine sediment samples were aseptically col-
lected by scuba from two stations in the Terra Nova Bay
(Ross Sea), i.e. Tethys Bay (THS; coordinates: 74�41.
6980S–164�04021400E; depth: 21 m) and Road Bay (ROB;
coordinates: 74�41080.300S–164�07080.300E; depth: 15 m),
which are both close to the Italian Base ‘‘Mario Zucchelli’’.
Bacterial isolation
Sediment samples for bacterial isolation were processed
within approximately 2 h after sampling. Serial dilutions
were prepared (1:10 and 1:100, using filter-sterilized
seawater) and 100 ll of each dilution was spread-plated in
three replicates on Marine Agar 2216 (MA, Difco). Inoc-
ulated plates were incubated in the dark at 4 �C for
1 month. Colonies were randomly isolated from agar
plates, picked and subcultured almost three times under the
same conditions.
16S rDNA PCR amplification of bacterial isolates
PCR-amplification of 16S rDNA from bacterial isolates
was carried out under the conditions described earlier
(Michaud et al. 2004). Briefly, a single colony of each
strain was lysed by heating at 95 �C for 10 min. Amplifi-
cation of 16S rDNA was performed with an ABI 9600
thermocycler (PE, Applied Biosystems) using the domain
Bacteria-specific primers 27F (50-AGAGTTTGATCCTGG
CTCAG-30) (spanning positions from 8 to 27 in E. coli
rRNA coordinates) and 1492R (50-CTACGGCTACCTTGT
TACGA-30) (spanning positions from 1492 to 1513 in
E. coli). The reaction mixtures were assembled at 0 �C and
contained 1–10 ng DNA, 10X buffer, 1.5 mM MgCl2,
150 ng of each forward and reverse primer (MWG, Ger-
many), 250 lM dNTP (Polymed, Italy), 0.5 units of
PolyTaq polymerase (Polymed, Italy) and sterile distilled
water to a final volume of 20 ll. The PCR program was as
follows: 3 min at 95 �C, followed by 30 cycles of 1 min at
94 �C, 1 min at 50 �C, 2 min at 72 �C and a final extension
step of 10 min at 72 �C. The expected size of the PCR
product was approximately 1.4 kb. The results of the
amplification reactions were analyzed by agarose gel
electrophoresis (1 %, w/v) in TAE buffer (0.04 M Tris–
acetate, 0.02 M acetic acid, 0.001 M EDTA), containing
1 lg/ml of ethidium bromide.
Sequencing and analysis of 16S rRNA gene
Automated sequencing of 16S rRNA gene from isolates
was carried out by cycle sequencing using the dye termi-
nator method. Sequencing was carried out at the Sequenc-
ing Service of the Macrogen Laboratory (Korea). Next
relatives of isolates were determined by comparison to 16S
rRNA gene sequences in the NCBI GenBank and the
EMBL databases using BLAST, and the ‘‘Seqmatch’’ and
‘‘Classifier’’ programs of the Ribosomal Database Project
II (http://rdp.cme.msu.edu/). All sequences with similarity
C97 % were considered to represent one phylogenetic
group or phylotype and grouped. Selected sequences were
further aligned using the program Clustal W (Thompson
et al. 1994) to the most similar orthologous sequences
retrieved from database. Each alignment was checked
manually, corrected and then analyzed using the neigh-
bour-joining method (Saitou and Nei 1987) according to
the model of Jukes-Cantor distances. Phylogenetic tree was
Response of bacterial isolates 241
123

constructed using the MEGA 5 (Molecular Evolutionary
Genetics Analysis) software (Kumar et al. 1993). The
robustness of the inferred trees was evaluated by 500
bootstrap re-samplings.
Isolates are part of the Italian Collection of Antarctic
Bacteria (CIBAN) of the National Antarctic Museum
(MNA, www.mna.it) ‘‘Felice Ippolito’’ kept in our labo-
ratory at the University of Messina. They are currently
maintained on MA slopes at 4 �C and routinely streaked on
agar plates from tubes every 6 months to control purity and
viability. Antarctic strains are also preserved by freezing
cell suspensions at -80 �C in Marine Broth (MB, Difco) to
which 20 % (v/v) glycerol is added.
The nucleotide sequences of the representative isolates
sequenced in this study have been deposited in the Gen-
Bank database under the accession numbers JX103222–
JX103234.
Bacterial growth in the presence of polychlorinated
biphenyls (PCBs)
Aerobic PCB degradation was screened into the mineral
liquid medium Bushnell Haas (BH; Difco) supplemented
with NaCl (3 %, w/v) and an Aroclor 1242 solution
(100 ppm in dichloromethane) as sole carbon and energy
source (final concentration 0.1 %, v/v) (De Domenico et al.
2004; Michaud et al. 2007). Aroclor 1242 is a mixture of
PCB congeners (ranging from dichloro- to hexachlorobi-
phenyls) made of twelve carbon atoms in the biphenyl
molecule and containing 42 % chlorine by weight.
Before adding the bacterial inoculum, 10 ll of Aroclor
solution were transferred to 100 ml flasks containing
9.5 ml of BH and left in a cabinet until CH2Cl2 had
completely evaporated. The medium was inoculated with
0.5 ml of a freshly prepared bacterial suspension (5 %, v/v)
in 3 % (w/v) NaCl supplemented filter-sterilized distilled
water. The optical density (OD) of the suspension was
adjusted to 1.0 at 600 nm. Cultures were incubated at 4 �C
for 3 weeks on a rotary shaker operated at 100 rpm. The
ability to use PCBs as growth substrates was evaluated
according to the degree of turbidity or the appearance of
cellular flocs in the test tubes. Uninoculated medium was
incubated in parallel as a negative control.
Heavy metal tolerance
Heavy metal tolerance was tested by modifying the method
reported by Chandy (1999). Four metals [i.e. cadmium
(Cd), zinc (Zn), mercury (Hg) and copper (Cu)] were tes-
ted. Metal salt solutions were prepared in filter-sterilized
phosphate buffer saline (PBS) and stored at 4 �C until use
(Selvin et al. 2009). The following concentrations (ppm)
were used: 10, 50, 100, 500, 1,000, 2,500, 5,000, 7,500 and
10,000 ppm. The metals were added as CdCl2 9 2.5 H2O,
HgCl2, CuCl2 9 2H2O and ZnCl2.
Isolates were grown in MB at 4 �C until the OD600nm
was approximately 0.500. Aliquots (100 ll) of each cell
suspension were spread-plated in duplicate on MA plates.
The inoculated plates were considered ready for the
assessment of heavy metal tolerance when the inoculum
was completely absorbed (after 10–15 min). Ten microli-
ters of each metal solution were then spotted on the inoc-
ulated agar plates. Sterile PBS was used as negative
control. Plates were further incubated at 4 �C and exam-
ined for the appearance of well-defined inhibition zone
after 96 h.
Antibiotic resistance
The susceptibility tests to antibiotics were carried out on
MA plates supplemented with chloramphenicol (25 and
50 ppm), ampicillin (range 50 and 100 ppm), streptomycin
(25, 250, 350 and 500 ppm) and kanamycin (25, 150 and
250 ppm). Inoculated plates were incubated at 4 �C for
96 h.
Data elaboration
Results were converted to a binary matrix and used to
calculate a Jaccard similarity matrix. The matrix was then
analyzed by non-metric multidimensional scaling (nMDS)
by using the software Primer-6 (Plymouth Marine Labo-
ratory, Roborough, United Kingdom). nMDS is a multi-
variate ordination method transforming complex data into a
new format by constructing a new set of variables.
This procedure condenses data to one point in a two-
dimensional plane so that highly similar data are plotted
close together.
Results
Phylogenetic affiliation of Antarctic isolates
A total of 98 strains were isolated from MA plates: 72 were
from the site ROB and 26 from THS, respectively. The
phylogenetic affiliation of isolates is reported in Fig. 1.
Only one representative isolate per phylotype is reported.
Based on the results of the 16S rDNA sequencing isolates
mainly belonged to the Gammaproteobacteria (65.3 % of
total isolates), followed by the CF group of Bacteroidetes
(24.5 %), Actinobacteria (9.2 %) and Alphaproteobacteria
(1.0 %). Comparative sequence analysis indicated that
isolates were closely related to already known and/or pre-
viously isolated bacteria (16S rRNA similarity, C97 %).
242 A. Lo Giudice et al.
123
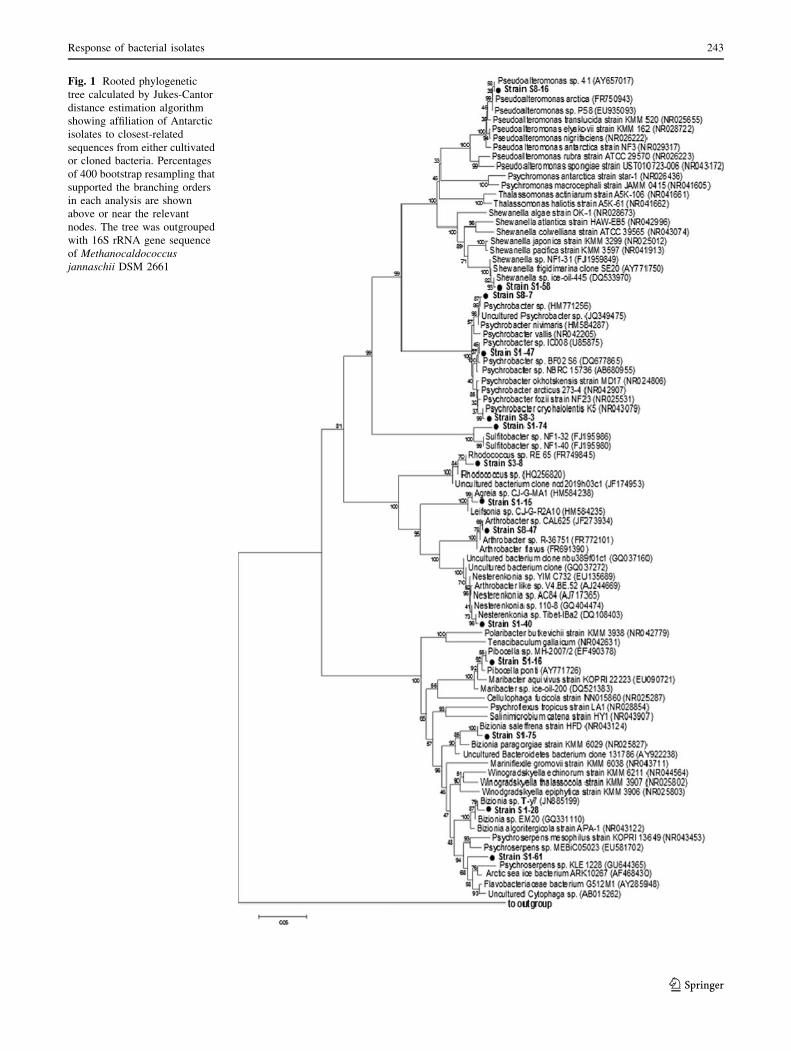
Fig. 1 Rooted phylogenetic
tree calculated by Jukes-Cantor
distance estimation algorithm
showing affiliation of Antarctic
isolates to closest-related
sequences from either cultivated
or cloned bacteria. Percentages
of 400 bootstrap resampling that
supported the branching orders
in each analysis are shown
above or near the relevant
nodes. The tree was outgrouped
with 16S rRNA gene sequence
of Methanocaldococcusjannaschii DSM 2661
Response of bacterial isolates 243
123
The Gammaproteobacteria clustered in two main groups
(Fig. 1). The first one branched in two sub-clusters formed
by Pseudoalteromonas (strain S8-16 representing 12
isolates) and Shewanella (strain S1-58 representing three
isolates) affiliates, respectively. The larger group included
isolates that were strongly related to the genus Psychrob-
acter (strains S8-7, S1-47 and S8-3 representing 49 isolates).
The next abundant group, the CF group of Bacteroidetes,
formed a single cluster that included representatives of
the genera Bizionia (nine isolates represented by the strains
S1-28 and S1-75), Pibocella (three isolates represented by
the strain S1-16) and Psychroserpens (12 isolates repre-
sented by the strain S1-61).
The Actinobacteria clustered in two groups. In particu-
lar, the first one included exclusively isolates that were
strongly related to the genus Rhodococcus (strain S3-8).
Isolates in the second group branched in two clusters
formed by Agreia (five isolates represented by the strain
S1-15) and Arthrobacter/Nesterenkonia (one and two
isolates represented by the strains S8-47 and S1-40,
respectively). Finally, the sole affiliate (strain S1-74) to the
Alphaproteobacteria shared the highest degree of sequence
identity with a Sulphitobacter sp.
Bacterial growth in the presence of polychlorinated
biphenyls (PCBs)
A total of 22 isolates (18 Gammaproteobacteria, three
Actinobacteria and one Bacteroidetes) were able to grow
into the mineral salt medium amended with Aroclor 1242. In
particular, PCB-oxidizing bacteria were mainly affiliated to
Psychrobacter spp. (16 isolates, namely S1-6, S1-30, S1-33,
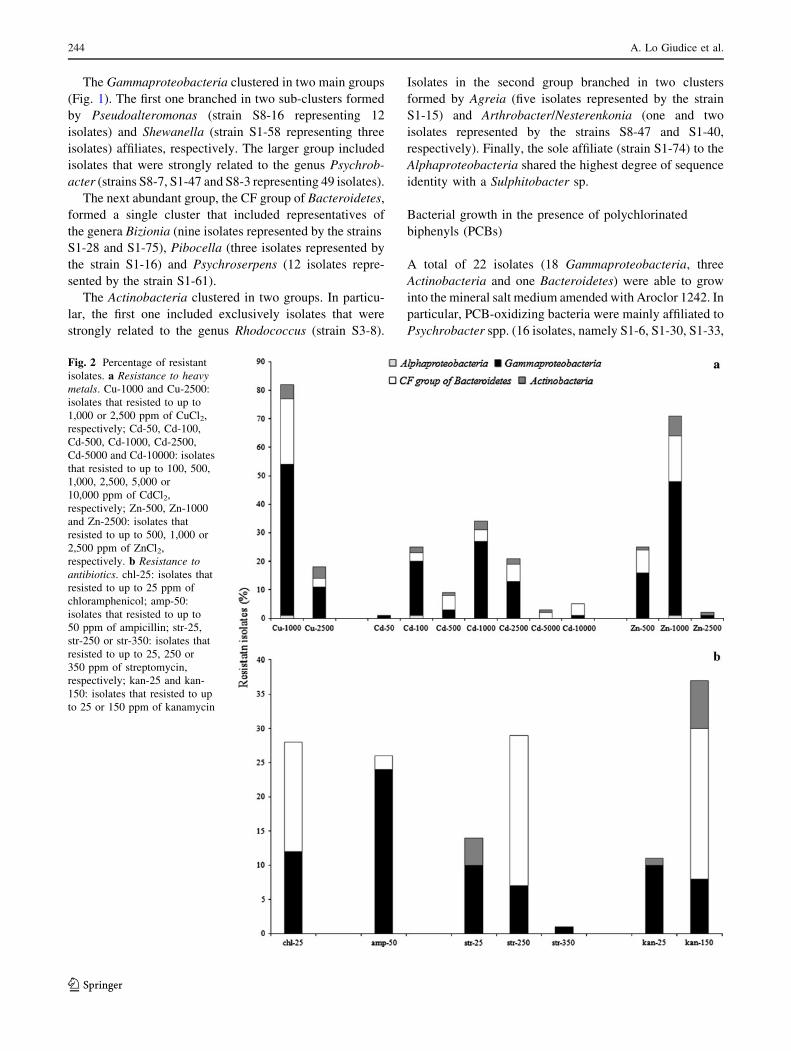
Fig. 2 Percentage of resistant
isolates. a Resistance to heavymetals. Cu-1000 and Cu-2500:
isolates that resisted to up to
1,000 or 2,500 ppm of CuCl2,
respectively; Cd-50, Cd-100,
Cd-500, Cd-1000, Cd-2500,
Cd-5000 and Cd-10000: isolates
that resisted to up to 100, 500,
1,000, 2,500, 5,000 or
10,000 ppm of CdCl2,
respectively; Zn-500, Zn-1000
and Zn-2500: isolates that
resisted to up to 500, 1,000 or
2,500 ppm of ZnCl2,
respectively. b Resistance toantibiotics. chl-25: isolates that
resisted to up to 25 ppm of
chloramphenicol; amp-50:
isolates that resisted to up to
50 ppm of ampicillin; str-25,
str-250 or str-350: isolates that
resisted to up to 25, 250 or
350 ppm of streptomycin,
respectively; kan-25 and kan-
150: isolates that resisted to up
to 25 or 150 ppm of kanamycin
244 A. Lo Giudice et al.
123
S1-41, S1-47, S1-60, S3-2, S3-4, S8-2, S8-3, S8-5, S8-7,
S8-12, S8-14, S8-42, S8-43) with Pseudoalteromonas,
Shewanella, Bizionia, Nesterenkonia, Arthrobacter and Rho-
dococcus that were represented by a single isolate each (namely
S8-16, S1-58, S1-67, S1-21, S8-47 and S3-8, respectively).
Heavy metal and antibiotic tolerance
All strains were strongly sensitive to HgCl2 as they did not
grow even at the lowest concentration that was tested for
such salt (10 ppm). Conversely, as it is shown in Fig. 2a,
isolates generally tolerated up to 1,000 ppm of the
remaining three metals. More in detail, the heavy metal
tolerance per genus is shown in Table 1. In particular,
members of the genera Psychroserpens (among the CFB
group of Bacteroidetes), Psychrobacter and Pseudoalte-
romonas (among the Gammaproteobacteria) were among
those strains that tolerated higher metal concentrations.
The concentration of 1,000 ppm was considered as a
threshold in order to individuate more resistant strains
(41; listed in Table 2). Tolerance patterns were generally
similar within the same species, though they often
Table 1 Heavy metal tolerance
per genus (the unidentified
alphaproteobacterium S1-74
was not included in the table)
Genus (no. of isolates) Heavy metal concentration (ppm)
50 100 500 1,000 2,500 5,000 10,000
CdCl2
Psychrobacter (49) 1 15 1 19 13 0 0
Pseudoalteromonas (12) 0 3 2 6 0 0 1
Shewanella (3) 0 1 0 2 0 0 0
Bizionia (9) 0 2 2 2 3 0 0
Pibocella (3) 0 1 0 1 0 1 0
Psychroserpens (12) 0 0 3 1 3 1 4
Clavibacter (5) 0 1 1 2 1 0 0
Arthrobacter (1) 0 0 0 0 1 0 0
Nesterenkonia (2) 0 1 0 1 0 0 0
Rhodococcus (1) 0 0 0 0 0 1 0
Total (97) 1 24 9 34 21 3 5
CuCl2
Psychrobacter (49) 0 0 0 41 8 0 0
Pseudoalteromonas (12) 0 0 0 10 2 0 0
Shewanella (3) 0 0 0 2 1 0 0
Bizionia (9) 0 0 0 8 1 0 0
Pibocella (3) 0 0 0 3 0 0 0
Psychroserpens (12) 0 0 0 12 0 0 0
Clavibacter (5) 0 0 0 4 1 0 0
Arthrobacter (1) 0 0 0 1 0 0 0
Nesterenkonia (2) 0 0 0 0 2 0 0
Rhodococcus (1) 0 0 0 0 1 0 0
Total (97) 0 0 0 81 16 0 0
ZnCl2
Psychrobacter (49) 0 0 12 37 0 0 0
Pseudoalteromonas (12) 0 0 2 9 1 0 0
Shewanella (3) 0 0 2 1 0 0 0
Bizionia (9) 0 0 4 5 0 0 0
Pibocella (3) 0 0 2 1 0 0 0
Psychroserpens (12) 0 0 2 10 0 0 0
Clavibacter (5) 0 0 1 4 0 0 0
Arthrobacter (1) 0 0 0 1 0 0 0
Nesterenkonia (2) 0 0 0 1 1 0 0
Rhodococcus (1) 0 0 0 1 0 0 0
Total (97) 0 0 25 70 2 0 0
Response of bacterial isolates 245
123
differed among species in the same genus. The highest
tolerated concentration of CuCl2 and ZnCl2 generally
resulted 2,500 ppm (16 and two strains, respectively),
whereas CdCl2 was also tolerated up to 5,000 and
10,000 ppm, even if by few strains (that were mainly
affiliated to Psychroserpens sp.). Multiple metal-resis-
tance was exhibited by only six strains, i.e. Psychrobacter
spp. S8-3 and S8-25, Pseudoalteromonas sp. S1-17,
Clavibacter sp. S1-37, Rhodococcus sp. S3-8 and Nest-
erenkonia sp. S1-40.
Table 2 Strains that tolerated heavy metal salts over the threshold concentration of 1,000 ppm (A: 2,500 ppm; B: 5,000 ppm; C: 10,000 ppm)
Phylum or class Next relative by GenBank alignment (AN, organism) Strain Metal salt
CuCl2 ZnCl2 CdCl2
Gammaproteobacteria PSU85875, Psychrobacter sp. IC008 S1-12 A
PSU85875, Psychrobacter sp. IC008 S1-47 A
HM771256, Psychrobacter sp. 4Dc S1-3 A
HM771256, Psychrobacter sp. 4Dc S1-43 A
HM771256, Psychrobacter sp. 4Dc S8-14 A
HM771256, Psychrobacter sp. 4Dc S1-41 A
HM771256, Psychrobacter sp. 4Dc S1-30 A
HM771256, Psychrobacter sp. 4Dc S1-45 A
HM771256, Psychrobacter sp. 4Dc S3-4 A
HM771256, Psychrobacter sp. 4Dc S8-12 A
HM771256, Psychrobacter sp. 4Dc S8-46 A
HM771256, Psychrobacter sp. 4Dc S8-5 A
DQ521392, Psychrobacter sp. ice-oil-471 S1-4 A
DQ521392, Psychrobacter sp. ice-oil-471 S1-34 A
DQ521392, Psychrobacter sp. ice-oil-471 S1-39 A
DQ521392, Psychrobacter sp. ice-oil-471 S8-29 A
DQ521392, Psychrobacter sp. ice-oil-471 S8-32 A
DQ521392, Psychrobacter sp. ice-oil-471 S8-25 A A
DQ521392, Psychrobacter sp. ice-oil-471 S8-3 A A
AY657017, Pseudoalteromonas sp. 41 S1-17 A A
AY657017, Pseudoalteromonas sp. 41 S8-17 C
AY657017, Pseudoalteromonas sp. 41 S8-20 A
DQ533970, Shewanella sp. ice-oil-445 S1-49 A
Bacteroidetes FJ889663, Bizionia sp. NF1-21 S1-53 A
FJ889663, Bizionia sp. NF1-21 S1-75 A
AY694003, B. algoritergicola strain APA-1 S1-66 A
AY694003, B. algoritergicola strain APA-1 S1-67 A
AY771777, Pibocella ponti clone SE92 S1-10 B
GU644365, Psychroserpens sp. KLE1228 S1-54 C
GU644365, Psychroserpens sp. KLE1228 S1-55 A
GU644365, Psychroserpens sp. KLE1228 S1-8 C
GU644365, Psychroserpens sp. KLE1228 S1-13 C
GU644365, Psychroserpens sp. KLE1228 S1-27 C
GU644365, Psychroserpens sp. KLE1228 S1-52 B
GU644365, Psychroserpens sp. KLE1228 S1-11 A
GU644365, Psychroserpens sp. KLE1228 S1-72 A
Actinobacteria EU374910, Clavibacter sp. AP29 S1-37 A A
AF134184, Arthrobacter agilis strain LV7 S8-47 A
DQ108403, Nesterenkonia sp. Tibet-IBa2 S1-21 A
DQ108403, Nesterenkonia sp. Tibet-IBa2 S1-40 A A
GQ153645, Rhodococcus sp. ANT-2400 S9 S3-8 A B
246 A. Lo Giudice et al.
123

Results from the antibiotic tolerance test are reported in
Fig. 2b. A total of 64 strains were resistant to at least one of
the tested antibiotics. Chloramphenicol and ampicillin
showed a strong inhibition effect against Antarctic isolates,
even at low concentrations (25–50 ppm), with resistant
strains that were 28 and 26, respectively. Overall, the
remaining two antibiotics were tolerated by a higher
number of isolates with the majority of them that were
resistant to kanamycin up to 150 ppm (37 out of 47
kanamycin-resistant strains) and streptomycin up to
250 ppm (29 out of 45 strains).
With regards to the phylogenetic affiliation, the unique
alphaproteobacterium (isolate S1-74) was susceptible to all
the tested antibiotics. Resistance to streptomycin and
kanamycin was predominantly observed for all bacteria
that belonged to the CF group of Bacteroidetes¸ in addition
to Pseudoalteromonas isolates among Gammaproteobac-
teria with Pseudoalteromonas sp. S1-46 that was resistant
to streptomycin up 350 ppm (Table 3). Multiple antibiotic-
resistance was exhibited by forty-five strains (Table 4).
Resistance patterns often differed within the same species.
Discussion
In the present study, we report for the first time on the
response of bacteria from Antarctic shallow sediments
towards heavy metals, antibiotics and PCBs, in order to
further contribute to the understanding of the role played
by microbial communities, as the initial step in most food
chains, in the transfer of toxic compounds to higher trophic
levels (De Souza et al. 2006). Such contaminants have been
detected in the Antarctic sedimentary compartment, also
in the Terra Nova Bay area (Fuoco et al. 1994, 1995;
Giordano et al. 1999; Dalla Riva et al. 2004). However, to
the best of our knowledge, their effect on indigenous
bacteria has been never investigated.
Interestingly, the nMDS analysis that was computed on
the entire data set from the screening showed that bacterial
isolates clustered according to their phylogenetic affiliation
(Fig. 3). In fact, only few Gammaproteobacteria isolates
were resistant to heavy metal concentrations higher than
1,000 ppm, whereas they were strongly represented among
antibiotic-resistant and/or PCB-oxidizing isolates. In con-
trast, the CF group of Bacteroidetes seemed to be more
resistant to the tested heavy metals. The comparison of the
antibiotic and heavy metal resistance patterns at genus
level revealed some distinctive features. For example,
members of the genus Psychroserpens (among the CF
group of Bacteroidetes) generally tolerated higher con-
centrations of the cadmium salt than the other genera,
whereas the genera Psychrobacter and Pseudoalteromonas
generally better tolerated copper and zinc. With respect to
the antibiotic resistance, the genera Psychrobacter,
Pseudoalteromonas, Shewanella and Bizionia showed the
widest range of resistance as they grew also in the presence
of ampicillin. Among Actinobacteria, all isolates that
belonged to the genera Nesterenkonia, Clavibacter and
Arthrobacter were resistant to kanamycin up to 150 ppm.
The observed differences in antibiotic and heavy metal
resistance/tolerance patterns can be dependent on the
ecology and physiology of the bacterial isolates, indicating
different modes and mechanisms of resistance/tolerance
acquisition, as it was suggested by Vaz-Moreira et al.
(2011). Additionally, such results indicate that distinctive
resistance/tolerance patterns could be related to the phy-
logenetic affiliation. Thus the dissemination of heavy metal
tolerance and antibiotic resistance may possess great rele-
vance for the population dynamics.
However, growth patterns often highly differed among
isolates in the same species, thus appearing to be more
likely strain- rather than species-specific. This finding
suggests a possible process of resistance/tolerance acqui-
sition or loss.
As it was stated in the introduction, information on
PCB-degrading cold-adapted microorganisms remain quite
scarce and, in particular, data regarding Antarctica are
limited to few investigations which demonstrated their occur-
rence in the water column at Terra Nova Bay (Yakimov
et al. 1999; De Domenico et al. 2004; Michaud et al. 2007).
Table 3 Antibiotic tolerance per genus (the unidentified alphapro-
teobacterium S1-74 was not included in the table)
Genus (no. of resistant
isolates)
Antibiotic concentration (ppm)
Chl
25
Amp
50
Str
25
Str
250
Str
350
Kan
25
Kan
150
Psychrobacter (18 out of
49)
4 15 4 2 0 6 1
Pseudoalteromonas (11
out of 12)
7 8 4 4 1 3 6
Shewanella (3 out of 3) 1 1 2 1 0 1 1
Bizionia (9 out of 9) 3 2 0 9 0 0 9
Pibocella (3out of 3) 3 0 0 3 0 0 3
Psychroserpens(11 out of 12)
11 0 0 11 0 0 11
Clavibacter
(5 out of 5)
0 0 1 0 0 0 5
Arthrobacter(1 out of 1)
0 0 0 0 0 0 1
Nesterenkonia(2 out of 2)
0 0 2 0 0 0 2
Rhodococcus(1 out of 1)
0 0 1 0 0 1 0
Total (64 out of 97) 29 26 14 30 1 11 39
Strains that were resistant to almost one antibiotic are considered
Response of bacterial isolates 247
123

The isolation of PCB-oxidizing bacteria from Antarctic
sediments is here reported for the first time. Interestingly,
the percentage of bacteria that were able to grow into the
mineral salt medium amended with Aroclor 1242 was
consistently higher than that previously determined for
bacteria from seawater (21.4 vs 7.1 %; Michaud et al.
2007). This finding suggests that the sedimentary microbial
community may be more adapted to the presence of PCBs
than that inhabiting the water column, due to the accu-
mulation of PCBs in sediments.
The phylogenetic analysis revealed that the PCB-
oxidizers that were affiliated to Psychrobacter, Pseudoal-
teromonas, Arthrobacter and Rhodococcus species were
strongly related (97–99 % of similarity) to PCB-degrading
Table 4 Antibiotic resistance patterns of Antarctic isolates
Phylum or
class*
Strain (s) Next relative by GenBank alignment (AN,
organism)
Chl**
(ppm)
Amp**
(ppm)
Str** (ppm) Kan**
(ppm)
25 50 25 250 350 25 150
GAM S1-29 PSU85875, Psychrobacter sp. IC008 X
S1-35 PSU85875, Psychrobacter sp. IC008 X
S1-45 HM771256, Psychrobacter sp. 4Dc X X X X
S8-7, S8-42, S8-46 HM771256, Psychrobacter sp. 4Dc X X X
S8-43 HM771256, Psychrobacter sp. 4Dc X X
S1-9 HM771256, Psychrobacter sp. 4Dc X X
S1-38, S8-44, S3-3,
S3-4, S8-2, S8-14
HM771256, Psychrobacter sp. 4Dc X
S3-2 HM771256, Psychrobacter sp. 4Dc X
S8-34 DQ521392, Psychrobacter sp. iceoil471 X X X X
S1-19 DQ521392, Psychrobacter sp. iceoil471 X
S8-29 DQ521392, Psychrobacter sp. iceoil471 X
S1-46 AY657017, Pseudoalteromonas sp. 41 X X X X
S8-13, S8-16,
S8-20, S8-24
AY657017, Pseudoalteromonas sp. 41 X X X X
S8-38 AY657017, Pseudoalteromonas sp. 41 X X X X
S8-17 AY657017, Pseudoalteromonas sp. 41 X X X
S1-14, S1-17 AY657017, Pseudoalteromonas sp. 41 X X
S1-22, S8-1 AY657017, Pseudoalteromonas sp. 41 X
S1-49 DQ533970, Shewanella sp. iceoil445 X X X X
S1-68 DQ533970, Shewanella sp. iceoil445 X X
S1-58 DQ533970, Shewanella sp. iceoil445 X
CFB S1-53 FJ889663, Bizionia sp. NF121 X X X X
S1-57, S1-75 FJ889663, Bizionia sp. NF121 X X
S1-44, S1-56 AY694003, B. algoritergicola strain APA1 X X X
S1-66 AY694003, B. algoritergicola strain APA1 X X X
S1-28, S1-67, S1-71 AY694003, B. algoritergicola strain APA1 X X
S1-10, S1-26, S1-16 AY771777, Pibocella ponti clone SE92 X X X
S1-13, S1-27, S1-54,
S1-55, S1-61, S1-63,
S1-11,
S1-52, S1-51, S1-72
GU644365, Psychroserpens sp. KLE1228 X X X
ACT S1-37 EU374910, Clavibacter sp. AP29 X X
S1-15, S3-7, S3-12 EU374910, Clavibacter sp. AP29 X
S1-21, S1-40 DQ108403, Nesterenkonia sp. TibetIBa2 X X
S8-47 AF134184, Arthrobacter agilis strain LV7 X
S3-8 GQ153645, Rhodococcus sp. ANT2400 S9 X X
* GAM, Gammaproteobacteria; CFB, CF group of Bacteroidetes; ACT, Actinobacteria
** Chl, chloramphenicol; Amp, ampicillin; Str, streptomycin; Kan, kanamycin
248 A. Lo Giudice et al.
123

bacteria that were isolated form Antarctic seawater during
previous expeditions at Terra Nova Bay (Yakimov et al.
1999; Michaud et al. 2007). Thus such bacteria could be
autochthonous and shared between the water column and
sediment compartments. Conversely, as far as we know, no
report exists about the ability of cold-adapted bacteria
belonging to the genera Shewanella, Bizionia and Nest-
erenkonia to utilize PCBs, as we observed through the
present study.
Microorganisms can develop resistance in the growing
presence of toxic compounds in the environment. The most
common resistance is to metals and antibiotics, which can
be a result of bio-essentiality or abuse of the metals, and/or
antibiotics. In the present study, multi-resistance either to
antibiotics or metals was seldom observed, thus confirming
results previous reported by De Souza et al. (2006) for
bacteria from Antarctic seawater. Tolerance to the four
heavy metals assayed in this study was in the order of
Cd [ Cu [ Zn [ Hg and appeared to be strictly related to
the metal concentrations, as determined during chemical
surveys in the same area. Concentrations of mercury in the
Terra Nova Bay sediments are among the lowest ever
reported for coastal marine environments (Bargagli et al.
1998). This feature could explain the very high suscepti-
bility to mercury of our Antarctic isolates which are
probably not adapted to the presence of mercury in their
surrounding environment. For the same reason, a strong
tolerance was recorded in the present study towards cad-
mium. In fact, cadmium content in sediments of the Terra
Nova Bay generally results considerably higher than
background typical concentrations due to the presence of
biogenic calcareous debris, where Cd tends to be enriched,
or to the Cd uptake by phytoplankton and the subsequent
discard with detritus and faecal pellets (Ianni et al. 2010).
Finally, a moderate tolerance was observed towards copper
and zinc whose concentrations in the Terra Nova Bay
generally resulted similar to those regarded as background
in most marine coastal areas (Bargagli et al. 1996).
With regards to antibiotic assays, the response of the
isolates to the tested antibiotics ranged from complete
resistance to total susceptibility. In particular, resistances to
ampicillin and chloramphenicol were very pronounced in
the majority of isolates, confirming previous results for
Antarctic marine bacteria (De Souza et al. 2006; Lo Giu-
dice et al. 2010, 2012) Antibiotic resistance was not
expected at so high frequency as tested bacteria were from
an environment where pollution from both chemicals and
drugs is quite scarce. So, it is possible that acquired anti-
biotic resistance traits could become widespread even in
environments where both the exposure to antibiotics and
the anthropic pressure are minimal.
In conclusion, physiological responses by bacteria from
Antarctic sediment towards PCBs, heavy metals and anti-
biotics were investigated. Antarctic bacteria responded
differently to the presence of such toxic compounds pri-
marily based on their phylogenetic affiliation and second-
arily at strain level. Moreover, the high incidence of
resistance either to metal or antibiotics, in addition to the
capability to grow on PCBs, confirm that bacteria are able
to cope and/or adapt to the occurrence pollutants even in
low human-impacted environments. Thus, the study of the
microbial responses to anthropogenic stress could furnish
useful information on the contaminant exposure of a cer-
tain ecosystem and, therefore, an overall picture of envi-
ronmental health. A genome-wide transcriptional level
analysis will be a focus of future research to reveal more
in-depth detail in the bacterial responses to such kind of
pollutants.
Acknowledgments This research was supported by grants from
PNRA (Programma Nazionale di Ricerche in Antartide), Italian
Ministry of Education and Research (Research Project PNRA
2004/1.6), and from MNA (Museo Nazionale dell’Antartide).
References
Andrade S, Poblet A, Scagliola M, Vodopivez C, Curtosi A, Pucci A,
Marcovecchio J (2001) Distribution of heavy metals in surface
sediments from an Antarctic marine ecosystem. Environ Monit
Assess 66:147–158
Bargagli R, Nelli L, Ancora S, Focardi S (1996) Elevated cadmium
accumulation in marine organisms from Terra Nova Bay
(Antarctica). Polar Biol 16:13–520
Bargagli R, Monaci F, Sanchez-Hernandez JC, Cateni D (1998)
Biomagnification of mercury in an Antarctic marine coastal food
web. Mar Ecol Progr Ser 169:65–76
Bedard DL, Wagner RE, Brennan MJ, Haberl ML, Brown JF Jr
(1987) Extensive degradation of Aroclors and environmental
Fig. 3 Non-metric multidimensional scaling (nMDS) ordination of
the obtained results. Plain line represents groups of 80 % similarity
(Jaccard). Asterisk, Alphaproteobacteria; Filled circle, Gammaprote-obacteria; Square, CFB group of Bacteroidetes; Rectangle,
Actinobacteria
Response of bacterial isolates 249
123

transformed polychlorinated biphenyls by Alcaligenes eutrophusH850. Appl Environ Microbiol 53:1094–1102
Caroppo C, Stabili L, Aresta M, Corinaldesi C, Danovaro R (2006)
Impact of heavy metals and PCBs on marine picoplankton.
Environ Toxicol 21:541–551
Chandy JP (1999) Heavy metal tolerance in chromogenic and non-
chromogenic marine bacteria from Arabian Gulf. Environ Monit
Assess 59:321–330
Dalla Riva S, Abelmoschi ML, Magi E, Soggia F (2004) The utilization
of the Antarctic environmental specimen bank (BCAA) in
monitoring Cd and Hg in an Antarctic coastal area in Terra Nova
Bay (Ross Sea - Northern Victoria Land). Chemosphere 56:59–69
De Domenico M, Lo Giudice A, Michaud L, Saitta M, Bruni V (2004)
Diesel oil and PCB-degrading bacteria isolated from Antarctic
seawater (Terra Nova Bay, Ross Sea). Polar Res 23:141–146
De Souza M-J, Nair S, Loka Bharathi PA, Chandramohan D (2006)
Metal and antibiotic-resistance in psychrotrophic bacterial from
Antarctic marine waters. Ecotoxicology 15:379–384
Dercova K, Seligova J, Dudasova H, Mikulasova M, Silharova K,
Tothova L, Hucko P (2009) Characterization of the bottom
sediments contaminated with polychlorinated biphenyls: evalua-
tion of ecotoxicity and biodegradability. Intl Biodet Biodegr 63:
440–449
Fuoco R, Colombini MP, Abete C (1994) Determination of
polychlorobiphenyls in environmental samples from Antarctica.
Int J Environ Anal Chem 55:15–25
Fuoco R, Colombini MP, Abete C, Carignani S (1995) Polychloro-
biphenyls in sediment, soil and sea water samples from
Antarctica. Int J Environ Anal Chem 61:309–318
Gillan DC, Danis B, Pernet P, Joly G, Dubois P (2005) Structure of
sediment-associated microbial communities along a heavy-metal
contamination gradient in the marine environment. Appl Environ
Microbiol 71:679–690
Giordano R, Lombardi G, Ciaralli L, Beccaloni E, Sepe A, Ciprotti
M, Costantini S (1999) Major and trace elements in sediments
from Terra Nova Bay, Antarctica. Sci Total Environ 227:29–40
Grotti M, Soggia F, Lagomarsino C, Dalla Riva S, Goessler W,
Francesconi KA (2008) Natural variability and distribution of
trace elements in marine organisms from Antarctic coastal
environments. Antarc Sci 20:39–51
Haferburg G, Kothe E (2007) Microbes and metals: interactions in the
environment. J Basic Microbiol 47:453–467
Ianni C, Magi E, Soggia F, Rivaro P, Frache R (2010) Trace metal
speciation in coastal and off-shore sediments from Ross Sea
(Antarctica). Microchem J 96:203–212
Kohler HPE, Kohler-Staub D, Focht DD (1988) Cometabolism of
polychlorinated biphenyls: enhanced transformation of Aroclor 1254
by growing bacterial cells. Appl Environ Microbiol 54:1940–1945
Kumar S, Tamura K, Nei M (1993) Mega: molecular evolutionary
genetics analysis, version 1.02. The Pennsylvania State Univer-
sity, University Park, Pa 16802
Lo Giudice A, Casella P, Caruso C, Mangano S, Bruni V, De
Domenico M, Michaud L (2010) Occurrence and characteriza-
tion of psychrotolerant hydrocarbon–oxidizing bacteria from
surface seawater along the Victoria Land coast (Antarctica).
Polar Biol 33:929–943
Lo Giudice A, Caruso C, Mangano S, Bruni V, De Domenico M,
Michaud L (2012) Marine bacterioplankton diversity and
community composition in an Antarctic coastal environment.
Microb Ecol 63:210–223
Martinez JL (2009) Environmental pollution by antibiotics and by
antibiotic resistance determinants. Environ Poll 157:2893–2902
Michaud L, Di Cello F, Brilli M, Fani R, Lo Giudice A, Bruni V
(2004) Biodiversity of cultivable Antarctic psychrotrophic
marine bacteria isolated from Terra Nova Bay (Ross Sea).
FEMS Microbiol Lett 230:63–71
Michaud L, Di Marco G, Bruni V, Lo Giudice A (2007) Biodegra-
dative potential and characterization of psychrotolerant poly-
chlorinated biphenyl-degrading marine bacteria isolated from a
coastal station in the Terra Nova Bay (Ross Sea, Antarctica).
Mar Poll Bull 54:1754–1761
Montone RC, Taniguchi S, Weber RR (2001) Polychlorinated
biphenyls in marine sediments of Admiralty Bay, King George
Island, Antarctica. Mar Pollut Bull 42:611–614
Montone RC, Taniguchi S, Boian C, Weber RR (2005) PCBs and
chlorinated pesticides (DDTs, HCHs and HCB) in the atmo-
sphere of the southwest Atlantic and Antarctic oceans. Mar Poll
Bull 50:778–786
Saitou N, Nei M (1987) The neighbor-joining method: a new method
for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Selvin J, Shanmugha PS, Kiran GS, Thangavelua T, Bai NS (2009)
Sponge-associated marine bacteria as indicators of heavy metal
pollution. Microbiol Res 164:352–363
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucl Acids Res 22:4673–4680
Tumeo MA, Guinn DA (1997) Evaluation of bioremediation in cold
regions. J Cold Reg Engin 11:221–231
Vaz-Moreira I, Nunes OC, Manaia CM (2011) Diversity and
antibiotic resistance patterns of Sphingomonadaceae isolates
from drinking water. Appl Environ Microbiol 77:5697–5706
Yakimov MM, Giuliano L, Bruni V, Scarfı S, Golyshin PN (1999)
Characterization of Antarctic hydrocarbon-degrading bacteria
capable of producing bioemulsifiers. New Microbiol 22:249–259
250 A. Lo Giudice et al.
123
Copyright © 2022 FDOKUMEN




















