
Comparative evaluation of trimeric envelope glycoproteins derived from subtype C and B HIV1 R5...
18
Comparative evaluation of trimeric envelope glycoproteins derived from subtype C and B HIV-1 R5 isolates Indresh K. Srivastava a, ⁎ , Elaine Kan a , Yide Sun a , Victoria A. Sharma a , Jimna Cisto a , Brian Burke a , Ying Lian a , Susan Hilt a , Zohar Biron a , Karin Hartog a , Leonidas Stamatatos c,d , R. Holland Cheng b , Jeffrey B. Ulmer a , Susan W. Barnett a a Novartis Vaccines and Diagnostics, Inc., 4560 Horton Street, Emeryville, CA 94608, USA b Molecular and Cellular Biology, University of California at Davis, Davis, CA 95616, USA c Seattle Biomedical Research Institute, University of Washington, Seattle, WA, 98195, USA d Department of Pathobiology, University of Washington, Seattle, WA, 98195, USA Received 27 June 2007; returned to author for revision 19 July 2007; accepted 18 October 2007 Available online 3 December 2007 Abstract We previously reported that an envelope (Env) glycoprotein immunogen (o-gp140ΔV2SF162) containing a partial deletion in the second variable loop (V2) derived from the R5-tropic HIV-1 isolate SF162 partially protected vaccinated rhesus macaques against pathogenic SHIV SF162P4 virus. Extending our studies to subtype C isolate TV1, we have purified o-gp140ΔV2TV1 (subtype C ΔV2 trimer) to homogeneity, performed glycosylation analysis, and determined its ability to bind CD4, as well as a panel of well-characterized neutralizing monoclonal antibodies (mAb). In general, critical epitopes are preserved on the subtype C ΔV2 trimer; however, we did not observe significant binding for the b12 mAb. The molecular mass of subtype C ΔV2 trimer was found to be 450 kDa, and the hydrodynamic radius was found to be 10.87 nm. Our data suggest that subtype C ΔV2 trimer binds to CD4 with an affinity comparable to o-gp140ΔV2SF162 (subtype B ΔV2 trimer). Using isothermal titration calorimetric (ITC) analysis, we demonstrated that all three CD4 binding sites (CD4-BS) in both subtype C and B trimers are exposed and accessible. However, compared to subtype B trimer, the three CD4-BS in subtype C trimer have different affinities for CD4, suggesting a cooperativity of CD4 binding in subtype C trimer but not in subtype B trimer. Negative staining electron microscopy of the subtype C ΔV2 trimer has demonstrated that it is in fact a trimer. These results highlight the importance of studying subtype C Env, and also of developing appropriate subtype C-specific reagents that may be used for better immunological characterization of subtype C Env for developing an AIDS vaccine. © 2007 Elsevier Inc. All rights reserved. Keywords: HIV; Trimer; Subtype C; CD4; Neutralizing antibodies; ITC Introduction Acquired Immunodeficiency Syndrome (AIDS) continues to be a major health problem throughout the world, with approxi- mately 40 million people infected with the human immunode- ficiency virus type 1(HIV-1) and 20 million deaths due to AIDS to date. In the United States, an estimated one million individuals are currently living with HIV, and 40,000 to 80,000 new primary infections occur each year (UNAIDS/WHO, 2004). The situation is continuously deteriorating as a result of the rapid emergence of drug resistance against most of the effective anti-virals. In certain parts of the world, such as Sub-Saharan Africa, India, and China, HIV-1 subtype C is the most prevalent strain. For instance, in Botswana and Swaziland, the prevalence of subtype C HIV infection is estimated to be as high as 35% (UNAIDS/WHO, 2004). Therefore, there is an urgent need for an effective anti-HIV vaccine that can be used either alone as a prophylactic vaccine, or in conjunction with effective anti-virals as a therapeutic vaccine. In the mid 1980s, during the early phase of the AIDS epi- demic, induction of neutralizing antibodies (nAbs) was believed to be sufficient to protect against HIV infection. However, preclinical and early clinical trials in the early 1990s (Mascola et al., 1993, 1996) confirmed that the neutralizing antibody Available online at www.sciencedirect.com Virology 372 (2008) 273 – 290 www.elsevier.com/locate/yviro ⁎ Corresponding author. Fax: +1 510 923 2586. E-mail address: [email protected] (I.K. Srivastava). 0042-6822/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2007.10.022
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative evaluation of trimeric envelope glycoproteins derived from subtype C and B HIV1 R5...

Available online at www.sciencedirect.com
8) 273–290www.elsevier.com/locate/yviro
Virology 372 (200
Comparative evaluation of trimeric envelope glycoproteins derived fromsubtype C and B HIV-1 R5 isolates
Indresh K. Srivastava a,⁎, Elaine Kan a, Yide Sun a, Victoria A. Sharma a, Jimna Cisto a,Brian Burke a, Ying Lian a, Susan Hilt a, Zohar Biron a, Karin Hartog a, Leonidas Stamatatos c,d,
R. Holland Cheng b, Jeffrey B. Ulmer a, Susan W. Barnett a
a Novartis Vaccines and Diagnostics, Inc., 4560 Horton Street, Emeryville, CA 94608, USAb Molecular and Cellular Biology, University of California at Davis, Davis, CA 95616, USAc Seattle Biomedical Research Institute, University of Washington, Seattle, WA, 98195, USA
d Department of Pathobiology, University of Washington, Seattle, WA, 98195, USA
Received 27 June 2007; returned to author for revision 19 July 2007; accepted 18 October 2007Available online 3 December 2007
Abstract
Wepreviously reported that an envelope (Env) glycoprotein immunogen (o-gp140ΔV2SF162) containing a partial deletion in the second variableloop (V2) derived from the R5-tropic HIV-1 isolate SF162 partially protected vaccinated rhesus macaques against pathogenic SHIVSF162P4 virus.Extending our studies to subtype C isolate TV1, we have purified o-gp140ΔV2TV1 (subtype C ΔV2 trimer) to homogeneity, performedglycosylation analysis, and determined its ability to bind CD4, as well as a panel of well-characterized neutralizing monoclonal antibodies (mAb). Ingeneral, critical epitopes are preserved on the subtype CΔV2 trimer; however, we did not observe significant binding for the b12mAb. Themolecularmass of subtype C ΔV2 trimer was found to be 450 kDa, and the hydrodynamic radius was found to be 10.87 nm. Our data suggest that subtype CΔV2 trimer binds to CD4 with an affinity comparable to o-gp140ΔV2SF162 (subtype BΔV2 trimer). Using isothermal titration calorimetric (ITC)analysis, we demonstrated that all three CD4 binding sites (CD4-BS) in both subtype C and B trimers are exposed and accessible. However, comparedto subtype B trimer, the three CD4-BS in subtype C trimer have different affinities for CD4, suggesting a cooperativity of CD4 binding in subtype Ctrimer but not in subtype B trimer. Negative staining electron microscopy of the subtype C ΔV2 trimer has demonstrated that it is in fact a trimer.These results highlight the importance of studying subtype C Env, and also of developing appropriate subtype C-specific reagents that may be usedfor better immunological characterization of subtype C Env for developing an AIDS vaccine.© 2007 Elsevier Inc. All rights reserved.
Keywords: HIV; Trimer; Subtype C; CD4; Neutralizing antibodies; ITC
Introduction
Acquired Immunodeficiency Syndrome (AIDS) continues tobe a major health problem throughout the world, with approxi-mately 40 million people infected with the human immunode-ficiency virus type 1(HIV-1) and 20 million deaths due to AIDSto date. In the United States, an estimated one million individualsare currently living with HIV, and 40,000 to 80,000 new primaryinfections occur each year (UNAIDS/WHO, 2004). The situationis continuously deteriorating as a result of the rapid emergence of
⁎ Corresponding author. Fax: +1 510 923 2586.E-mail address: [email protected] (I.K. Srivastava).
0042-6822/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.virol.2007.10.022
drug resistance against most of the effective anti-virals. In certainparts of the world, such as Sub-Saharan Africa, India, and China,HIV-1 subtype C is the most prevalent strain. For instance, inBotswana and Swaziland, the prevalence of subtype C HIVinfection is estimated to be as high as 35% (UNAIDS/WHO,2004). Therefore, there is an urgent need for an effective anti-HIVvaccine that can be used either alone as a prophylactic vaccine, orin conjunction with effective anti-virals as a therapeutic vaccine.
In the mid 1980s, during the early phase of the AIDS epi-demic, induction of neutralizing antibodies (nAbs) was believedto be sufficient to protect against HIV infection. However,preclinical and early clinical trials in the early 1990s (Mascolaet al., 1993, 1996) confirmed that the neutralizing antibody

274 I.K. Srivastava et al. / Virology 372 (2008) 273–290
responses induced by the monomeric HIV Env vaccines beingtested at that time did not offer the breadth necessary to eitherprevent or control infection with naturally acquired field isolatesof HIV. The focus then changed to the development of vaccinesand regimens that would induce strong cellular responses. Earlypreclinical studies in rhesus macaques had established thecorrelation between strong cytotoxic T-cell (CTL) activity andreduced viral load. Subsequent studies using DNA prime/vectorboost and vector prime/vector boost further strengthened theobservation that strong cellular responses can serve as correlatesof protection from disease development in animal models.However, recent data obtained in a rhesus macaque challengemodel suggest that cellular responses that focus on a singleepitope may not be sufficient to effectively control viral repli-cation for extended periods of time due to the emergence ofescape mutants (Barouch et al., 2003, 2002; Barouch and Letvin,2002; Vogel et al., 2003). It remains to be seen if increasing thebreadth and potency of vaccine-elicited CTL responses will havea positive impact on the outcome of challenge infection. It is nowwidely believed that humoral, cellular, and T-helper cell re-sponses, both at peripheral and mucosal sites, are needed toprovide broad and durable protection against HIV. Conceptually,antibodies would serve as a first line of defense by eithercompletely preventing infection or reducing the virus inocu-lums. Cellular responses would then facilitate the clearance ofHIV-infected CD4+ T-cells that have escaped antibody-mediated neutralization and reduce the severity of the disease.In a proof of concept study, Mascola et al. (2003) have shownthat the induction of cellular responses using DNA vaccines inconjunction with passive transfer of nAbs did not exceed theprotection afforded by passive transfer of nAbs alone. Furtherstudies will be needed to demonstrate any potential advantage ofinducing both potent cellular and humoral responses by aneffective HIV vaccine.
The induction of broadly reactive nAbs against primary HIV-1 strains may prevent HIV infection by blocking the initial stageof infection (Graham, 2000). Although it is difficult to elicitbroadly nAbs against primary HIVor SIV strains in humans andin nonhuman primates (Johnston and Flores, 2001; Parren et al.,2001), it has been shown that antibodies induced by prime-boostvaccine regimens can prevent immunodeficiency virus infectionby inducing effective neutralizing antibody responses. Forexample, priming chimpanzees with replication-competentadenovirus-HIV gp160 recombinants, followed by boostingthem with HIV-1 SF2 gp120 in MF59, elicited high serum titersof antibodies capable of neutralizing homologous and heterol-ogous primary HIV-1 isolates in vitro. This finding correlatedwith in vivo protection of the animals against multiple, intra-venous viral challenges (Robert-Guroff et al., 1998; Zolla-Pazneret al., 1998). Furthermore, Mascola et al. (1999) and Ruprechtet al. (2001) have clearly demonstrated the positive effect of nAbsin passive immunizations upon the outcome of the challengeinfection. Thus, the induction of broadly neutralizing antibodyresponses is important for developing an effective HIV vaccine.
The main question now is how to design immunogens thatcan induce nAbs of the right specificity and affinity to providemeaningful levels of protection against challenge with field
isolates of HIV. We have pursued a modified-envelope HIVvaccine approach based on deleting the second hyper variableregion (V2) from HIV envelope immunogens. This approach isbased on several observations: (i) deletion of the V2 loop re-sulted in 100-fold increased susceptibility of antibody-mediatedneutralization of mutant virus (Stamatatos and Cheng-Mayer,1998a); (ii) deletion of the V2 loop alters the entry of mutantvirus in CD4+ T-cells (Wyatt et al., 1995); (iii) CD4-induciblemAbs – such as 17b and 48d – better recognize V1/V2-deletedenvelopes (Sullivan et al., 1998; Wyatt et al., 1995); and (iv)envelopes without V1 and V2 loops can mediate fusion withCCR5+ cells in the absence of the CD4 receptor (Johnson et al.,2002). Taken together, these observations suggest that the V1and V2 loops may mask conserved regions of the envelopeinvolved in viral entry and susceptibility to neutralization(Johnson et al., 2002; Stamatatos and Cheng-Mayer, 1998b;Sullivan et al., 1998; Wei et al., 2003; Wyatt et al., 1995). Inaddition, removal of the V1 and V2 loops may also eliminateseveral potential N-linked glycosylation sites (NLGS) and ex-pose critical functional epitopes as recently described in the“glycan shield” model proposed by Wei et al. (2003). Ly andStamatatos (2000) have demonstrated that elimination ofspecific NLGS from the V2 loop renders the virus more sus-ceptible to neutralization by various mAbs. Therefore, develop-ment of novel Env immunogens based on V2 loop deletion isrelevant to developing an HIV vaccine. By introducing a partialdeletion of the V2 loop in HIV Env immunogens derived fromthe R5-tropic, subtype B HIV strain SF162, we have altered theimmunogenic properties of HIV Env as demonstrated by theimmunogenicity studies in different animal models (Barnettet al., 2001; Srivastava et al., 2003). Priming rhesus macaqueswith ΔV2SF162-envelope-encoding DNA vaccines, followedby boosting with o-gp140ΔV2SF162, confers partial protectionagainst a pathogenic SHIVSF162P challenge (Barnett et al.,2001; Cherpelis et al., 2001a,b). Quinnan et al. (2005) demon-strated that priming of rhesus macaques with alpha virusexpressing R2 Env, followed by boosting with oligomericprotein, induced antibodies that neutralized diverse primaryisolates. Furthermore, immunized rhesus macaques were pro-tected against the challenge with partially pathogenic SHIVDH12R (Clone 7) as reflected by a reduction in the magnitudeand duration of virus load.
In a more recent study, Quinnan and colleagues have clearlydemonstrated that trimeric Env compared to monomeric Env isa potent immunogen (Zhang et al., 2007). They havedemonstrated that gp140R2 immunogen induced antibodiesthat achieved 50% neutralization of 48/48, and 80% neutrali-zation of 43/46 primary strains of diverse HIV-1 subtypestested. The strains tested included members of standard panelsof subtype B and C strains, and other diverse strains known tobe neutralization resistant. The gp120R2 induced antibodiesneutralized only 9/48 of the same strains. Neutralization wasIgG-mediated and HIV-1-specific.
Previously we reported on the engineering and characteriza-tion of soluble trimeric gp140 proteins derived from SF162lacking the V2 loop (Srivastava et al., 2003). Here, we describethe purification and characterization of V2 loop-deleted Env

Fig. 1. Structure and stabilization of HIV-1 subtype C Env in trimericconformation. The linear map of gp120ΔV2 and gp41 of subtype C Env ispresented. The gp120 variable regions (V1 to V5) are indicated as differentshaded squares, and the protease cleavage site is marked with an arrow. All theprotease cleavage site mutations (primary and secondary) that were evaluated forstabilizing subtype C trimers are shown. Effect of these mutations on theexpression and stabilization of subtype C Env in trimeric conformation is alsopresented (T stands for trimer, and M for monomer).
Table 1Development of stable CHO cell clones secreting o-gp140ΔV2TV1
No. of clones Methotrexate concentration (nM) Expression (ng/ml)
588 50 nd84 50 200–40036 100 500–190034 200 500–190021 400 400–140021 800 500–2300
275I.K. Srivastava et al. / Virology 372 (2008) 273–290
immunogen derived from TV1, an R5 dependent subtype Cisolate derived from South Africa. We have previously shownthat TV1 immunogens elicited homologous nAbs, and providedsome breadth against a panel of heterologous subtype B andsubtype C primary isolates (Lian et al., 2005). EM studies in-dicate that the purified protein is in a stable trimeric confor-mation; immunochemical studies indicate that the subtype CΔV2 trimer binds to CD4 at an affinity similar to subtype BΔV2trimers, and it has comparable glycosylation profile to subtype BΔV2 trimer. However, there are differences between subtype Band C ΔV2 trimers in the binding profiles of mAbs. Our studiesreveal differences in the presentation of neutralization epitopeson clade B and clade C gp140 immunogens that could affect theelicitation of neutralizing antibody responses.
Results and discussion
Results
Selection of subtype C HIV envelopeSeveral well-characterized subtype C isolates from South
Africa (Estrelita Van Rensburg, Tygerberg Hospital, Universityof Stellenbosch) were evaluated for their sensitivity toneutralization by homologous and heterologous sera. The Envgenes derived from proviral sequences from six clade C infectedpatients were amplified using primers derived from subtype Cconsensus sequence, and we cloned the PCR product in anexpression vector. The insert was confirmed by restrictionenzyme digest. The Env from six different clones was expressed,and characterized for level of expression in 293 and RD cells andits ability to bind to CD4. Only two clones expressed Env in 293cells to a reasonable level, and the partially purified Env proteinsbound to CD4 in an HPLC-based assay (data not shown). Nodetectable expression of any Env clone was observed in RDcells. Based on expression profile, CD4 binding, and fusogeni-city (Lian et al., 2005), TV1 was selected for further studies.
Analysis of leader sequencesTo optimize the secretion of the expressed Env protein
(gp120), three different leader sequences were evaluated tran-siently for the level of expression of secreted protein: (i) subtype
C specific native leader sequence (Native), (ii) tissue plasmin-ogen activator leader sequence (tPA), and (iii) an optimizedversion of the tPA sequence (tPAopt). Expressed Env proteinswere partially purified, and analyzed by SDS-PAGE, and immu-noprobing. The quantity of Env proteins secreted by these con-structs was relatively similar (data not shown). Therefore, theconstruct with native leader sequence was chosen for furtherevaluation.
Expression and stabilization of o-gp140ΔV2TV1Tomaximize the expression of gp140TV1ΔV2, the gene was
modified. Consistent with our previous observations (Srivastavaet al., 2003, 2002; zur Megede et al., 2000), modified constructsexpressed significantly higher levels of Env protein compared toun-optimized constructs (data not shown). We have previouslyshown that introduction of a single mutation (Arginine to Serine)in the protease cleavage site in HIV-1 Env gene from US4(a subtype B primary isolate) was effective for stabilization oftrimers (Srivastava et al., 2002). However, the same R-to-Smutation was relatively ineffective for stabilization of trimersderived from SF162 (Srivastava et al., 2003), and also for TV1(Fig. 1). Therefore, we evaluated several different combinationsof mutations in the primary (aa494 to 497; REKR) andsecondary (aa486 to 490; KAKRR) protease cleavage sites fortheir relative abilities to stabilize the TV1 Env trimer (Fig. 1).Based upon expression profile, molecular mass and sCD4binding, construct #7 which had a total of five mutations [two inthe primary (E495-S, R497-S) and three in the secondarycleavages site (K488-I, R489-S and R490-S)] was the mosteffective at stabilizing the Env trimer. These mutations wereused to derive stable CHO cell lines secreting o-gp140ΔV2TV1.
Selected clones were grown in presence of methotrexate atconcentrations ranging from 50 nM to 800 nM, andmaintained fora period of 3–4 weeks at one methotrexate concentration beforemoving on to the next drug concentration. Expression of Env wascontinuously monitored using a capture ELISA (Table 1). Basedon the expression profile at the highest methotrexate concentra-tion, the three best clones were selected for further evaluation andadaptation in a 3.5 L perfusion bioreactor. Clone #24Rwas chosenas the cell line for the production of gp140ΔV2TV1, based onlevel of Env expression and growth characteristics of the cells.
Purification of o-gp140ΔV2TV1A three-step strategy was used for efficient purification of
o-gp140ΔV2TV1, involving a capture and two cleaning steps.The Env protein was efficiently captured over a GNA lectincolumn, while most contaminating proteins flowed through. The

Fig. 2. Purification of subtype C Env protein. SDS-PAGE analysis of subtype C Env captured on GNA column (A: lane 2—starting material, lane 3—flow through afterGNA column, lane 4—pool of GNA elutions, lane 5—GNA elution (concentrated), lane 6—flow through after DEAE; lane 7—10× concentrated DEAE flow-through;lane 8—CHAP flow through). Analysis of different concentrations of final preparation of subtype C Env trimer (B: lane 1—2 μg, lane 2—5 μg, lane 3—10 μg, and lane4—molecular weight standard). Final preparation of subtypeCEnv trimer was characterized by immunoprobing using amAb (20-2-C8.5F3) (C: lane 2—gp120SF2 andlane 3—o-gp140ΔV2TV1), and size exclusion-HPLC profile of the purified subtype C Env proteins (D). Trimers (T) and monomers (M) are indicated.
276 I.K. Srivastava et al. / Virology 372 (2008) 273–290
bound Env was eluted with 500mMmethyl mannose pyranoside(MMP). It was then passed over DEAE and CHAP columns,where contaminating proteins bound to the columns, but Envflowed through. The fractions collected from each column wereanalyzed by SDS-PAGE, immunoblot, size exclusion columnchromatography (SEC), and a CD4 binding assay (Fig. 2). TheGNA lectin was very efficient at capturing the Env (N90%), butthe purity was less than 50% (Fig. 2A). After the DEAE column,Env preparation was more than 60–70% pure, and the majorcontaminant was a serum protein (α-2 macroglobulin) (Fig. 2A).However after the final step, i.e. CHAP column, Env was morethan 95% pure (Fig. 2A). The Env preparation was concentratedusing a 100 kDa membrane, buffer was exchanged, and it wasanalyzed over SDS-PAGE (Fig. 2B) where purity was estimatedto be N95% by densitometric scanning (data not shown). Further-more, to determine the proportion of trimer, the purified prep-aration was analyzed over a BioSil 250 SEC (Fig. 2D). Based onthe SEC profile, the purified Env preparation consisted of ap-proximately 70–80% oligomer and 20–30%monomer (Fig. 2D).Therefore, at a final stage of purification, Env trimer was sep-arated from monomer on a higher resolution, pre-calibratedSuperdex-200 column with molecular weight standards of 670,158, 44, 17, and 1.35 kDa (Fig. 3A). We pooled differentfractions constituting different parts of the peak to make fivedifferent pools. These pools were analyzed on PAGE undernative (Fig. 3B) and reduced and denaturing conditions (Fig. 3C).The data suggest that purified Env was in three different confor-mations: (i) larger structures representing the aggregated speciesN670 kDa (pool 1); (ii) trimers N440 kDa (pools 2, 3); and (iii)dimer b440 kDa (pool 4), and monomer N140 kDa (pool 5).However, all of these fractions migrated with an apparentmolecular mass of 140 kDa on reducing and denaturing SDS-
PAGE (Fig. 3C). No change in the electrophoretic mobility ofthese fractions was observed under denaturing conditions com-pared to reducing and denaturing conditions (data not shown)suggesting that Env monomers were noncovalently associatedwith each other to give rise to trimers. To further confirm that thepurified Env is a genuine trimer, and not a disulfide-linkedaggregate, the purified oligomers were treated with increasingconcentrations of β-ME, and the samples were analyzed byHPLC using BioSil 250 column. No change in the mass of theEnv protein was observed as reflected by the retention time upontreatment with increasing β-ME concentration suggesting thatthe trimers were not disulfide-linked aggregates (data notshown). Purified subtype C ΔV2 trimer was recognized as asingle species on westerns blot usingMAb 20-2-C8.5F3 directedagainst the conserved (C4) domain of subtype B gp120 (Fig. 2C)suggesting that there is no major degradation or proteolysisof o-gp140ΔV2TV1, and that the purified preparation washomogeneous.
Biochemical characterization of o-gp140ΔV2TV1
Glycosydic linkage analysis. The nature of the oligosacchar-ides linkages in the purified subtype C ΔV2 trimer wasanalyzed by digesting the Env protein with PNGF, whichreleases asparagine-linked oligosaccharides, with NANase IIand O-glycosidase DS, which release unsubstituted Gal (β1, 3)GalNAc (α1) disaccharides attached to serine or threonine,either alone or in combination. Like o-gp140ΔV2SF162, sub-type C ΔV2 trimer contained predominantly N-linked oligo-saccharides since PNGF treatment appeared to remove all thesugars (Fig. 4A, lanes 3 and 6). Furthermore, we observed thatNANase and O-glycosidase DS had no effect on electrophoretic
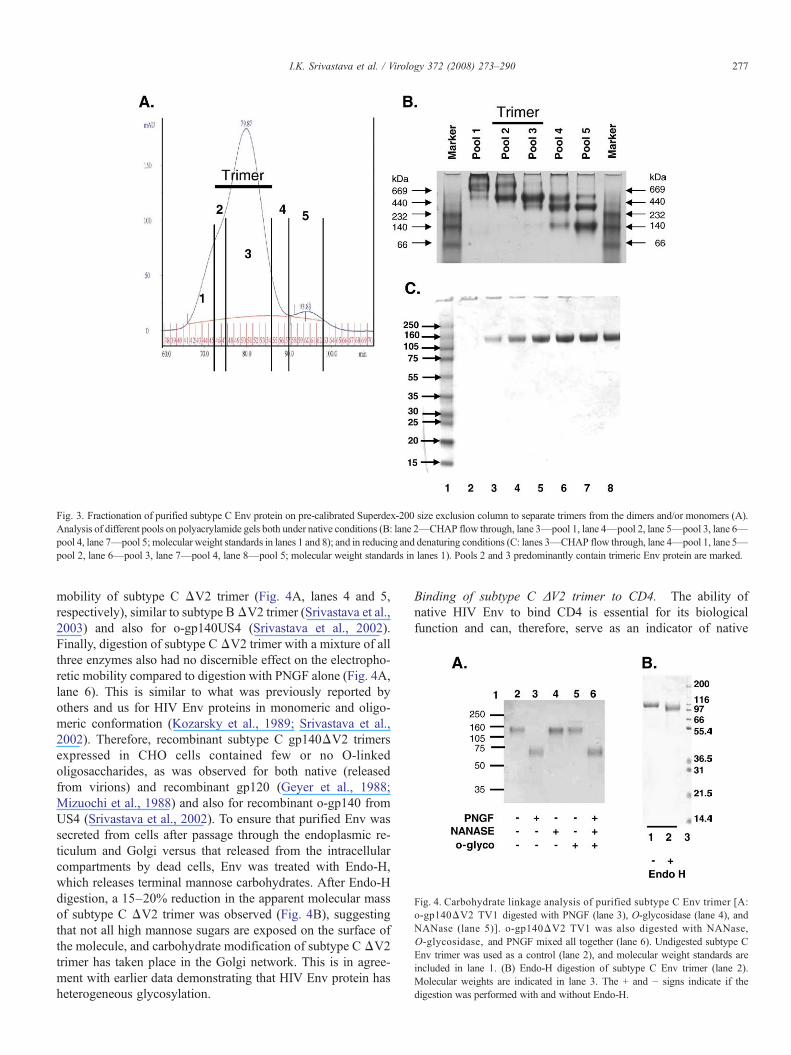
Fig. 4. Carbohydrate linkage analysis of purified subtype C Env trimer [A:o-gp140ΔV2 TV1 digested with PNGF (lane 3), O-glycosidase (lane 4), andNANase (lane 5)]. o-gp140ΔV2 TV1 was also digested with NANase,O-glycosidase, and PNGF mixed all together (lane 6). Undigested subtype CEnv trimer was used as a control (lane 2), and molecular weight standards areincluded in lane 1. (B) Endo-H digestion of subtype C Env trimer (lane 2).Molecular weights are indicated in lane 3. The + and − signs indicate if thedigestion was performed with and without Endo-H.
Fig. 3. Fractionation of purified subtype C Env protein on pre-calibrated Superdex-200 size exclusion column to separate trimers from the dimers and/or monomers (A).Analysis of different pools on polyacrylamide gels both under native conditions (B: lane 2—CHAP flow through, lane 3—pool 1, lane 4—pool 2, lane 5—pool 3, lane 6—pool 4, lane 7—pool 5; molecular weight standards in lanes 1 and 8); and in reducing and denaturing conditions (C: lanes 3—CHAP flow through, lane 4—pool 1, lane 5—pool 2, lane 6—pool 3, lane 7—pool 4, lane 8—pool 5; molecular weight standards in lanes 1). Pools 2 and 3 predominantly contain trimeric Env protein are marked.
277I.K. Srivastava et al. / Virology 372 (2008) 273–290
mobility of subtype C ΔV2 trimer (Fig. 4A, lanes 4 and 5,respectively), similar to subtype BΔV2 trimer (Srivastava et al.,2003) and also for o-gp140US4 (Srivastava et al., 2002).Finally, digestion of subtype CΔV2 trimer with a mixture of allthree enzymes also had no discernible effect on the electropho-retic mobility compared to digestion with PNGF alone (Fig. 4A,lane 6). This is similar to what was previously reported byothers and us for HIV Env proteins in monomeric and oligo-meric conformation (Kozarsky et al., 1989; Srivastava et al.,2002). Therefore, recombinant subtype C gp140ΔV2 trimersexpressed in CHO cells contained few or no O-linkedoligosaccharides, as was observed for both native (releasedfrom virions) and recombinant gp120 (Geyer et al., 1988;Mizuochi et al., 1988) and also for recombinant o-gp140 fromUS4 (Srivastava et al., 2002). To ensure that purified Env wassecreted from cells after passage through the endoplasmic re-ticulum and Golgi versus that released from the intracellularcompartments by dead cells, Env was treated with Endo-H,which releases terminal mannose carbohydrates. After Endo-Hdigestion, a 15–20% reduction in the apparent molecular massof subtype C ΔV2 trimer was observed (Fig. 4B), suggestingthat not all high mannose sugars are exposed on the surface ofthe molecule, and carbohydrate modification of subtype C ΔV2trimer has taken place in the Golgi network. This is in agree-ment with earlier data demonstrating that HIV Env protein hasheterogeneous glycosylation.
Binding of subtype C ΔV2 trimer to CD4. The ability ofnative HIV Env to bind CD4 is essential for its biologicalfunction and can, therefore, serve as an indicator of native
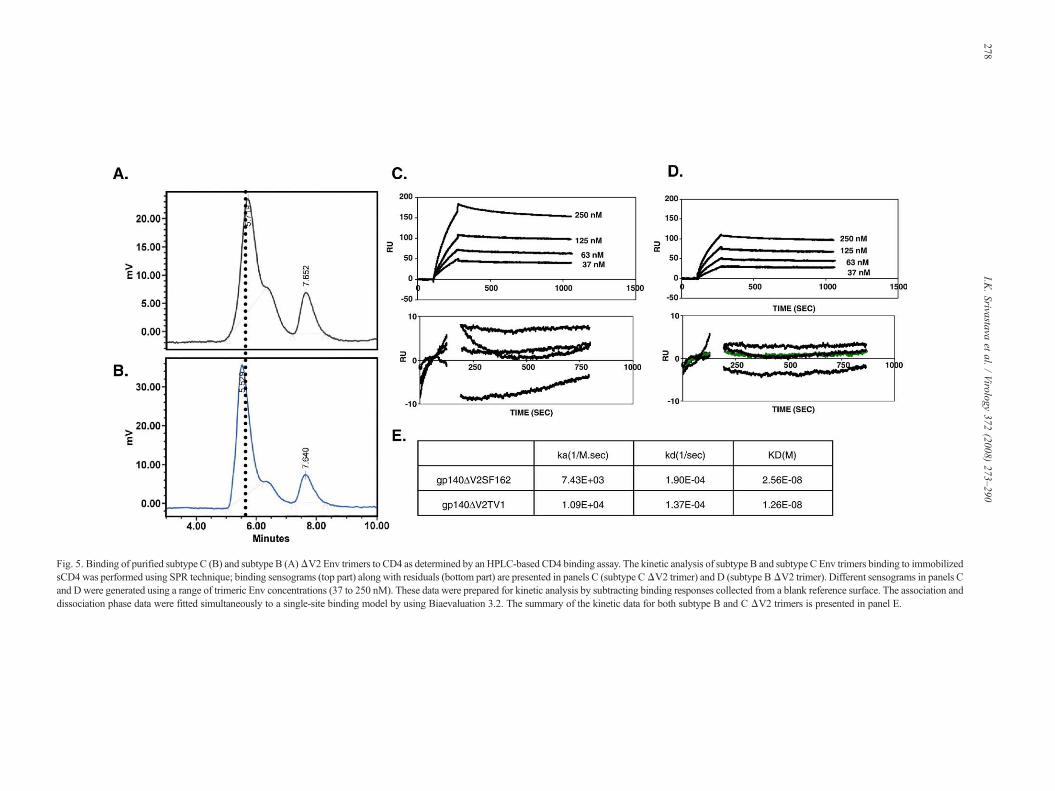
Fig. 5. Binding of purified subtype C (B) and subtype B (A)ΔV2 Env trimers to CD4 as determined by an HPLC-based CD4 binding assay. The kinetic analysis of subtype B and subtype C Env trimers binding to immobilizedsCD4 was performed using SPR technique; binding sensograms (top part) along with residuals (bottom part) are presented in panels C (subtype CΔV2 trimer) and D (subtype BΔV2 trimer). Different sensograms in panels Cand D were generated using a range of trimeric Env concentrations (37 to 250 nM). These data were prepared for kinetic analysis by subtracting binding responses collected from a blank reference surface. The association anddissociation phase data were fitted simultaneously to a single-site binding model by using Biaevaluation 3.2. The summary of the kinetic data for both subtype B and C ΔV2 trimers is presented in panel E.
278I.K
.Srivastava
etal.
/Virology
372(2008)
273–290

279I.K. Srivastava et al. / Virology 372 (2008) 273–290
structure and functionality of recombinant Env preparations(Scandella et al., 1993). Using an HPLC-based receptor-bindingassay (Srivastava et al., 2002), subtype C ΔV2 Env was shownto bind to CD4 (Fig. 5B) similar to theΔV2 trimer derived fromSF162 (Fig. 5A). However, the retention time for subtype CΔV2 trimer was shorter compared to subtype B ΔV2 trimer,suggesting that subtype C ΔV2 trimer is larger compared tosubtype B ΔV2 trimer. The kinetic constants observed for ΔV2trimers derived from TV1 and SF162 are presented in Figs. 5C–E. The calculated on rates forΔV2TV1 andΔV2SF162 trimerswere 1.09E+04 and 7.43E+03, respectively. In addition, thedissociation rate for subtype C ΔV2 trimer was slightly slowercompared to subtype B ΔV2 trimer. These data suggest tatΔV2TV1 trimer has higher binding affinity to sCD4 thanΔV2SF162 trimer.
Isothermal titration calorimetric analysis. Since isothermaltitration calorimetric (ITC) analysis is a very powerful tech-nique not only for the strength of interaction, but also fordetermining the stoichiometry of receptor ligand binding, ITCwas used to determine the stoichiometry of Env trimer and CD4.The data are presented in Fig. 6. For the data analysis, it was
Fig. 6. Thermodynamic properties of subtype C ΔV2 trimer were determined using isand the heat of mixing and dilution was subtracted from the final data. Panel A represeeach injection (power: time). The area under each spike is proportional to the heat conmole of injected ligand (envelope) as a function of molar ratio. For the trimeric Envs,The solid line represents the best nonlinear fit to the experimental data. The kinetic datactive sites in Env for CD4 (N) are presented in panel C. In panel D, the kinetic da
assumed that if all the CD4 binding sites in the trimer wereaccessible, then the number of active sites (N) would be equal to3, whereas for monomer the N would be equal to 1. The calcu-lated numbers of CD4 binding sites for the trimer were found tobe 2.79 and 0.965 for the trimer and monomer, respectively(Fig. 6C), suggesting that N90% of the Env molecules werefunctionally active in terms of their ability to bind CD4, and allthree CD4 binding sites in the purified subtype C trimers areaccessible. We have also calculated the affinity of each bindingsite in Env trimer for CD4. It was quite interesting to note thatall three CD4 binding sites in the subtype C ΔV2 trimer haddifferent KDs, suggesting cooperativity in binding (Fig. 6D).This is in contrast to subtype BΔV2 trimer, where all three CD4binding sites had relatively similar KDs (Sharma et al., 2006).
Immunochemical characterization of o-gp140ΔV2TV1The structural integrity of subtype C Env was further eval-
uated by surface immunoprobing with a panel of mAbs ofdefined epitope specificities (IgGCD4, b12, 2F5, 4E10, 447D,391-95d, G34, 17b, 4.8d 2.3E, LF17), using Surface PlasmonResonance assay (SPR), capture ELISA, and Far Western assay.The mAb IgGCD4, which binds to the CD4-binding site,
othermal calorimetric analysis (ITC). All titrations were performed in duplicate,nts the raw data depicting an endothermic reaction, where heat is consumed aftersumed by each injection of Env into CD4. Panel B represents integrated areas perthe number of binding sites cannot be determined based on the values on X-axis.a such as entropy (ΔS), enthalpy (ΔH), affinity of interaction (K), and calculatedta for each binding site in Env for CD4 are presented.
280 I.K. Srivastava et al. / Virology 372 (2008) 273–290
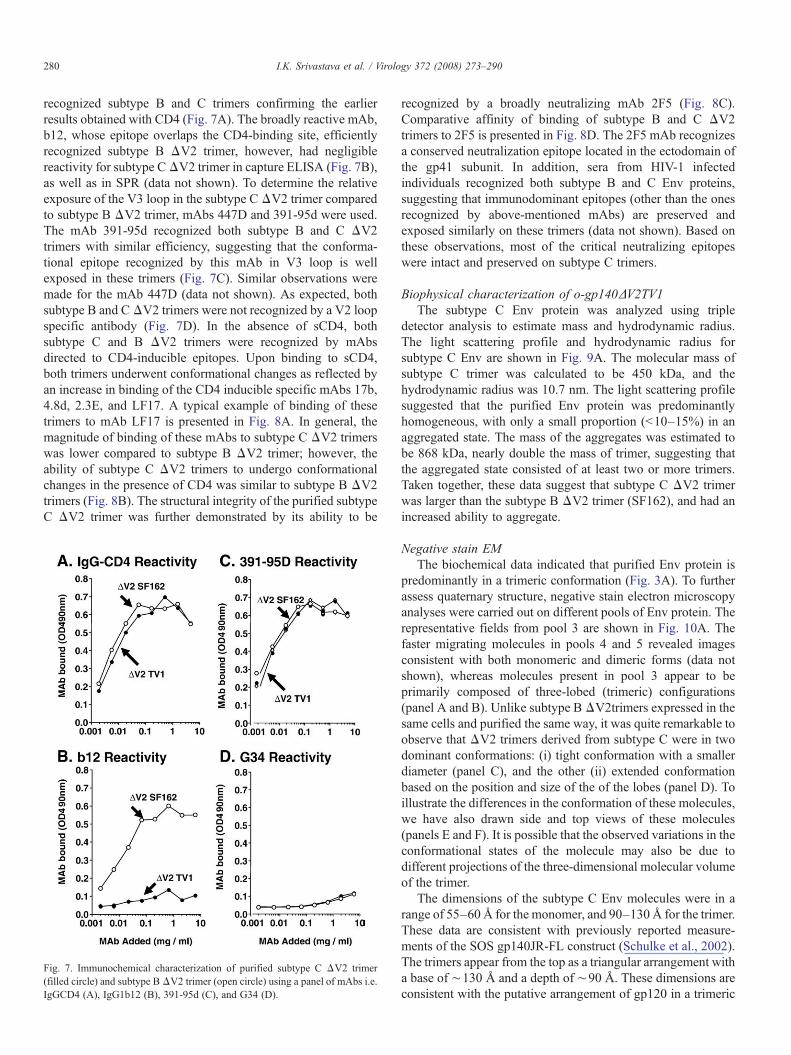
recognized subtype B and C trimers confirming the earlierresults obtained with CD4 (Fig. 7A). The broadly reactive mAb,b12, whose epitope overlaps the CD4-binding site, efficientlyrecognized subtype B ΔV2 trimer, however, had negligiblereactivity for subtype CΔV2 trimer in capture ELISA (Fig. 7B),as well as in SPR (data not shown). To determine the relativeexposure of the V3 loop in the subtype C ΔV2 trimer comparedto subtype B ΔV2 trimer, mAbs 447D and 391-95d were used.The mAb 391-95d recognized both subtype B and C ΔV2trimers with similar efficiency, suggesting that the conforma-tional epitope recognized by this mAb in V3 loop is wellexposed in these trimers (Fig. 7C). Similar observations weremade for the mAb 447D (data not shown). As expected, bothsubtype B and CΔV2 trimers were not recognized by a V2 loopspecific antibody (Fig. 7D). In the absence of sCD4, bothsubtype C and B ΔV2 trimers were recognized by mAbsdirected to CD4-inducible epitopes. Upon binding to sCD4,both trimers underwent conformational changes as reflected byan increase in binding of the CD4 inducible specific mAbs 17b,4.8d, 2.3E, and LF17. A typical example of binding of thesetrimers to mAb LF17 is presented in Fig. 8A. In general, themagnitude of binding of these mAbs to subtype C ΔV2 trimerswas lower compared to subtype B ΔV2 trimer; however, theability of subtype C ΔV2 trimers to undergo conformationalchanges in the presence of CD4 was similar to subtype B ΔV2trimers (Fig. 8B). The structural integrity of the purified subtypeC ΔV2 trimer was further demonstrated by its ability to be
Fig. 7. Immunochemical characterization of purified subtype C ΔV2 trimer(filled circle) and subtype BΔV2 trimer (open circle) using a panel of mAbs i.e.IgGCD4 (A), IgG1b12 (B), 391-95d (C), and G34 (D).
recognized by a broadly neutralizing mAb 2F5 (Fig. 8C).Comparative affinity of binding of subtype B and C ΔV2trimers to 2F5 is presented in Fig. 8D. The 2F5 mAb recognizesa conserved neutralization epitope located in the ectodomain ofthe gp41 subunit. In addition, sera from HIV-1 infectedindividuals recognized both subtype B and C Env proteins,suggesting that immunodominant epitopes (other than the onesrecognized by above-mentioned mAbs) are preserved andexposed similarly on these trimers (data not shown). Based onthese observations, most of the critical neutralizing epitopeswere intact and preserved on subtype C trimers.
Biophysical characterization of o-gp140ΔV2TV1The subtype C Env protein was analyzed using triple
detector analysis to estimate mass and hydrodynamic radius.The light scattering profile and hydrodynamic radius forsubtype C Env are shown in Fig. 9A. The molecular mass ofsubtype C trimer was calculated to be 450 kDa, and thehydrodynamic radius was 10.7 nm. The light scattering profilesuggested that the purified Env protein was predominantlyhomogeneous, with only a small proportion (b10–15%) in anaggregated state. The mass of the aggregates was estimated tobe 868 kDa, nearly double the mass of trimer, suggesting thatthe aggregated state consisted of at least two or more trimers.Taken together, these data suggest that subtype C ΔV2 trimerwas larger than the subtype B ΔV2 trimer (SF162), and had anincreased ability to aggregate.
Negative stain EMThe biochemical data indicated that purified Env protein is
predominantly in a trimeric conformation (Fig. 3A). To furtherassess quaternary structure, negative stain electron microscopyanalyses were carried out on different pools of Env protein. Therepresentative fields from pool 3 are shown in Fig. 10A. Thefaster migrating molecules in pools 4 and 5 revealed imagesconsistent with both monomeric and dimeric forms (data notshown), whereas molecules present in pool 3 appear to beprimarily composed of three-lobed (trimeric) configurations(panel A and B). Unlike subtype BΔV2trimers expressed in thesame cells and purified the same way, it was quite remarkable toobserve that ΔV2 trimers derived from subtype C were in twodominant conformations: (i) tight conformation with a smallerdiameter (panel C), and the other (ii) extended conformationbased on the position and size of the of the lobes (panel D). Toillustrate the differences in the conformation of these molecules,we have also drawn side and top views of these molecules(panels E and F). It is possible that the observed variations in theconformational states of the molecule may also be due todifferent projections of the three-dimensional molecular volumeof the trimer.
The dimensions of the subtype C Env molecules were in arange of 55–60 Å for the monomer, and 90–130 Å for the trimer.These data are consistent with previously reported measure-ments of the SOS gp140JR-FL construct (Schulke et al., 2002).The trimers appear from the top as a triangular arrangement witha base of ∼130 Å and a depth of ∼90 Å. These dimensions areconsistent with the putative arrangement of gp120 in a trimeric
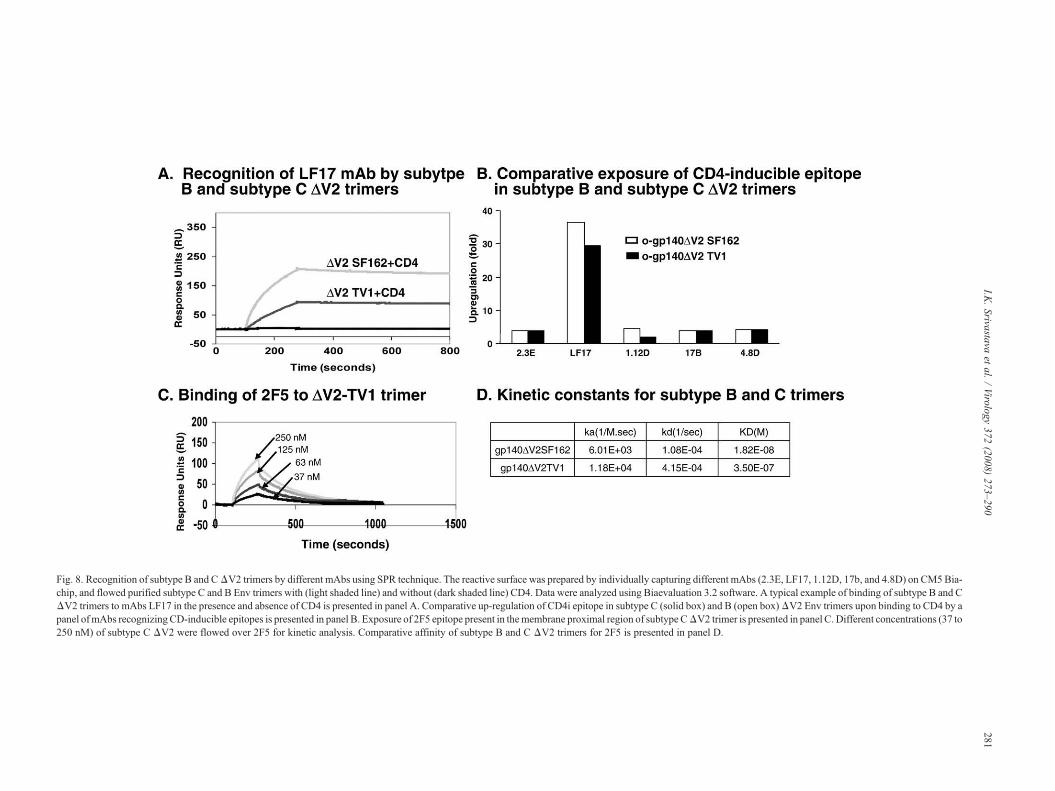
Fig. 8. Recognition of subtype B and CΔV2 trimers by different mAbs using SPR technique. The reactive surface was prepared by individually capturing different mAbs (2.3E, LF17, 1.12D, 17b, and 4.8D) on CM5Bia-chip, and flowed purified subtype C and B Env trimers with (light shaded line) and without (dark shaded line) CD4. Data were analyzed using Biaevaluation 3.2 software. A typical example of binding of subtype B and CΔV2 trimers to mAbs LF17 in the presence and absence of CD4 is presented in panel A. Comparative up-regulation of CD4i epitope in subtype C (solid box) and B (open box)ΔV2 Env trimers upon binding to CD4 by apanel ofmAbs recognizingCD-inducible epitopes is presented in panel B. Exposure of 2F5 epitope present in themembrane proximal region of subtype CΔV2 trimer is presented in panel C. Different concentrations (37 to250 nM) of subtype C ΔV2 were flowed over 2F5 for kinetic analysis. Comparative affinity of subtype B and C ΔV2 trimers for 2F5 is presented in panel D.
281I.K
.Srivastava
etal.
/Virology
372(2008)
273–290
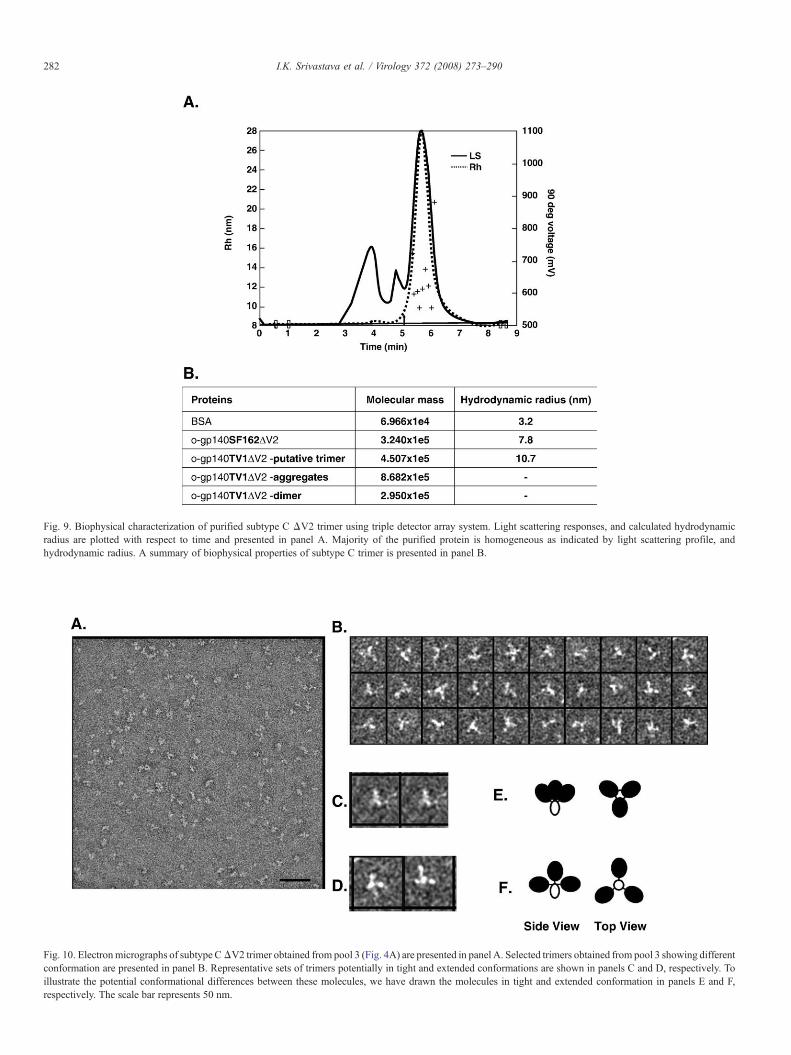
Fig. 9. Biophysical characterization of purified subtype C ΔV2 trimer using triple detector array system. Light scattering responses, and calculated hydrodynamicradius are plotted with respect to time and presented in panel A. Majority of the purified protein is homogeneous as indicated by light scattering profile, andhydrodynamic radius. A summary of biophysical properties of subtype C trimer is presented in panel B.
Fig. 10. Electronmicrographs of subtype CΔV2 trimer obtained from pool 3 (Fig. 4A) are presented in panel A. Selected trimers obtained from pool 3 showing differentconformation are presented in panel B. Representative sets of trimers potentially in tight and extended conformations are shown in panels C and D, respectively. Toillustrate the potential conformational differences between these molecules, we have drawn the molecules in tight and extended conformation in panels E and F,respectively. The scale bar represents 50 nm.
282 I.K. Srivastava et al. / Virology 372 (2008) 273–290

283I.K. Srivastava et al. / Virology 372 (2008) 273–290
conformation (Chen et al., 2005). The mass measurementsestimated from the STEM micrographs (data not shown)indicated the presence of mono-disperse protein solutions, show-ing well-behaved Gaussian distributions corresponding to a massof 113 kDa for the monomers and 353 kDa for the trimers (with-out the water molecules). A larger mass distribution was ob-served for the trimers (with a standard deviation of ∼8% of themean for the trimers versus ∼5% for the monomer), which maybe due to a smaller amount of aggregates or a lower proportion ofheterogeneity in the carbohydrate moieties in the trimers.
Discussion
The HIV Env glycoprotein is the target of broadly cross-reactive nAbs and will be an important component of asuccessful HIV vaccine. Therefore, it is important to engineerthe most relevant Env protein conformation for inclusion in apotential HIV vaccine. It is well documented that gp120monomer elicits high titer anti-env binding antibodies (Barnettet al., 1997; Haigwood et al., 1992; Mascola et al., 1999; Steimerand Haigwood, 1991) that neutralize T-cell adapted viruses, butare limited in their ability to neutralize primary isolates (Mascolaet al., 1996; Moore et al., 1995; VanCott et al., 1995; Wrin andNunberg, 1994). Furthermore, the antibodies induced by gp120react more strongly to monomer as compared to oligomer(Broder et al., 1994; VanCott et al., 1995). The observation thatbroadly reactive nAbs, such as IgG1b12, 2G12, and 2F5,generally have stronger affinity for the native envelope (trimer)than for monomeric gp120 or gp41 (Fouts et al., 1997; Robenet al., 1994) provides, at least conceptually, the impetus toevaluate trimeric Env glycoproteins as vaccine candidates(Binley et al., 2000; Iyer et al., 2007; Pancera et al., 2005;Quinnan et al., 2005; Sanders et al., 2000, 2002; Srivastava et al.,2003, 2002; Yang et al., 2000a,b, 2002, 2001; Zhang et al.,2007). So far, the data support this approach, as oligomers haveshown to be more effective in inducing strong antibodyresponses to conformational epitopes (Earl et al., 1994; VanCottet al., 1995; Yang et al., 2001). We have reported that a solubleoligomeric gp140 immunogen derived from the envelope ofSF162 elicited cross-reactive nAbs in rabbits and macaques(Barnett et al., 2001). In addition, we have demonstrated that theanimals immunized with o-gp140ΔV2 SF162 were partiallyprotected after pathogenic challenge with SHIVSF162P4(Barnettet al., 2001). Finally, we have also shown that, in intranasalprime and intramuscular boost regimen, subtype B trimeric Env-based vaccine induced strong antibodies that neutralized heter-ologous subtype B primary isolates (Srivastava et al., 2003;Vajdy et al., 2004). In addition, Quinnan and colleagues inducedpotent antibodies that neutralized diverse primary isolates usingan alpha prime and oligomeric protein boost strategy. Further-more, immunized rhesus macaques were protected against thepartially pathogenic SHIVDH12R (Clone 7) as reflected by thereduced magnitude and duration of viral load (Quinnan et al.,2005). More recently, Zhang et al. compared the immunogenic-ity of oligomeric and monomeric Env in rabbits. They haveclearly demonstrated that oligomeric Env is a better and morepotent immunogen compared to monomer. Rabbits immunized
with oligomeric Env induced antibodies that achieved 50%neutralization of 48/48, and 80% neutralization of 43/46 primarystrains of diverse HIV-1 subtypes tested. The strains testedincluded members of standard panels of subtype B and C strains,and other diverse strains known to be neutralization resistant.However, the gp120R2 induced antibodies that neutralized 9/48of the same strains (Zhang et al., 2007). These resultsdemonstrate that induction of truly broad spectrum nAbs is anachievable goal in HIV-1 vaccine development. We haverecently extended these studies on trimeric Env to subtype C,which is more prevalent in Sub-Saharan Africa and Asia. Basedon various biochemical, immunological, and physical criteria,trimeric subtype C Env proteins were similar in most aspects tosubtype B Env. In proof of concept immunogenicity studies inrabbits and monkeys, we have shown that ΔV2 trimers derivedfrom TV1 induced antibodies that neutralized some heterolo-gous isolates (Lian et al., 2005) and hence hold promise asvaccine candidates.
To elicit functional antibody responses, oligomeric HIV Envimmunogens need to be stabilized to retain their trimericconformation upon delivery. Several approaches are currentlybeing used: (1) elimination of the protease cleavage sitebetween gp120 and gp41 (Broder et al., 1994; Earl et al.,1994, 1991, 2001); (2) introduction of intramolecular disulfidebond formation (SOS) between gp120 and gp41 (Binley et al.,2000; Farzan et al., 1998; Sanders et al., 2000; Schulke et al.,2002); (3) in addition to the SOS modifications, introduction ofan isoleucine-to-proline substitution at position 559 in the N-terminal heptad repeat region of gp41 (SOSIP) (Iyer et al., 2007;Sanders et al., 2002); (4) extension of the gp41 coiled coilregion by the addition of GCN4 (a transcription factor thatnormally forms stable homotrimers) sequences, either alone orin conjunction with added cysteines between gp120 and gp41(Farzan et al., 1998; Yang et al., 2000a,b); and (5) addition of atrimerization domain from the C terminus of bacteriophage T4fibritin (Yang et al., 2002). We have used the protease cleavagesite mutation strategy to create stable trimers from subtype B(Srivastava et al., 2003, 2002), and now from subtype C.Consistent with our previous work on SF162, we observed thatseveral mutations in the primary and secondary cleavage sitesare required for creating stable subtype C trimers. It appears thatsequence variation, glycosylation, size of variable loops,expression levels, and the type of protein expression systemmay all play roles in stabilizing the HIV Env glycoprotein inoligomeric conformation (Yang et al., 2000a,b). Therefore,optimal strategies for the generation of stable, soluble HIVenvelope oligomers from a given isolate or clade will need to bedetermined experimentally on a case-by-case basis.
The binding of HIV Env protein to CD4, an obligatory stepin viral replication (Dalgleish et al., 1984; Klatzmann et al.,1984), is dependent upon conformation (Lasky et al., 1987;McDougal et al., 1986) and glycosylation (Barr et al., 1987; Liet al., 1993; Papandreou et al., 1996). Evidence so far suggeststhat N-linked glycosylation is important for the formation ofdisulfide bonds which indirectly facilitate functional confor-mation and binding to CD4 (Fennie and Lasky, 1989; Fenouilletet al., 1989; Leonard et al., 1990; Li et al., 1993). The V2 loop

284 I.K. Srivastava et al. / Virology 372 (2008) 273–290
deletion that also involved the deletion of a glycosylation site atposition 186 (Ly and Stamatatos, 2000; Maezawa et al., 1983)did not affect the ability of o-gp140ΔV2TV1 to bind to sCD4,suggesting that the purified protein was in its correct nativeconformation. The apparent affinity, kinetics, and thermody-namics of the CD4-Env interaction were studied by SPR andITC. The dissociation constant (KD) for subtype C Env trimerwas in nanomolar range and not significantly different from theKD of subtype B trimer (Sharma et al., 2006; Srivastava et al.,2003). To determine if all three CD4 binding sites in a trimer areaccessible for CD4 binding, we performed ITC analysis.Consistent with our previous data for subtype B (Sharmaet al., 2006), all three CD4 binding sites in subtype C trimerwere accessible to CD4. However, what is unique about subtypeC trimers is that the three CD4 binding sites have differentaffinities, suggesting a co-cooperativity for CD4 binding. Usinggp120-GCN4 trimers, Pancera et al. (2005) have shown thatonly two out of the three CD4 binding sites were accessible ingp120 trimers, and the third binding site was exposed only upondeletion of variable loops 1 and 2. It is possible that access to allthree binding sites may be dependent upon the orientation of themonomers, which is controlled by gp41. It appears that thefolding of trimers created by a heterologous trimerizationdomain such as GCN4 (Pancera et al., 2005) is different fromthat of the trimers created by native gp41 sequence (Sharmaet al., 2006; Srivastava et al., 2003), as reflected by the acces-sibility of the CD4 binding sites. Our data generated using SPR,ITC, and mAb binding suggest that both subtype B and C Envproteins underwent significant molecular rearrangement uponbinding to CD4. Similar findings were reported by Stites et al.(1997). It is important to emphasize that thermodynamic param-eters are highly dependent on the experimental conditions suchas pH, temperature, buffer, etc.; therefore it is generally difficultto compare thermodynamic properties of similar molecules ob-served from different studies. Nevertheless, apparent affinitiesmeasured by SPR, as well as enthalpies and Gibbs free energiescalculated from ITC data for subtype B and C trimers, wereremarkably consistent with previous results. In those experi-ments, gp120 derived from a different isolate was produced ininsect cells and was used under different experimental condi-tions. Myszka et al. (2000) reported that full-length gp120expressed in Drosophila bound to CD4 with an affinity of22 nM and an apparent Gibbs free energy of −9.5 kcal/mol.Similar data were obtained for subtype B V2 loop-deleted tri-mers (Sharma et al., 2006), and also for subtype C trimers(datanot shown).
As mentioned previously, glycosylation of the Env is notdirectly involved in CD4 binding; however, it provides a func-tional conformation to facilitate CD4 binding. As a measure ofproper glycosylation, we demonstrated that the purified subtypeC Env trimers contained predominantly N-linked oligosacchar-ides, similar to monomeric and trimeric derived from othersubtypes (Kozarsky et al., 1989; Srivastava et al., 2002). Fur-thermore, we found that there are three kinds of polysaccharidesassociated with Env. We purified and sequenced two poly-saccharides, and found that they were of terminal mannose andcomplex types. Similar observations were made of Env trimers
derived from subtype B isolates such as US4 and SF162, andconsistent to what is reported for native and recombinant Envproteins.
There is no consensus regarding the structure and the mass ofrecombinant HIV Env glycoproteins. Differences in theobserved mass of putative trimers may be because they werederived from different isolates, produced under different condi-tions, and characterized by different techniques (Broder et al.,1994; Earl et al., 1990, 1991; Grundner et al., 2002; Schulkeet al., 2002; Yang et al., 2000a, 2002). In the present study, weperformed an in-depth biophysical characterization of thepurified subtype C protein by using triple detector analysis andnegative stain EM. Combined, these methods strongly supportthe conclusion that the purified Env protein was predominantlyin a trimeric conformation. As shown earlier, we found that themonomeric Env proteins had higher Rh values of 5.07 nm forgp120US4; and 4.33 nm for gp120SF162ΔV2 (Srivastava et al.,2003, 2002). In contrast, Env glycoprotein trimers were tightlyfolded structures, as indicated by smaller than expected Rh
values: 8.4 nm for o-gp140US4 and 7.6 nm for subtype B ΔV2trimer (Srivastava et al., 2003, 2002), and 10.87 nm for subtypeC trimer. It is quite surprising to observe that cleaved subtype Atrimers produced using SOSIP approach have a size of 12–13 nm which is very similar to what we observed for uncleavedsubtype C V2 loop-deleted trimers (Iyer et al., 2007). These dataare consistent with earlier observations that there are significantdifferences in the structure and folding of monomer and trimer(Moore et al., 1994; Wyatt et al., 1998). The reported molecularmass for transiently expressed Env protein oligomers derivedfrom JR-FL, YU2, and HxBc2 is smaller than expected (400–410 kDa) (Binley et al., 2000; Yang et al., 2000a), and themolecular mass of the oligomer derived from JR-FL usingSOSIP approach is considerably larger than expected (Grundneret al., 2002; Sanders et al., 2002). More recently, Jeffs et al.(2004) have purified and characterized Env oligomers derivedfrom different primary isolates, and it was quite puzzling to notethat, with the exception of Env proteins derived from subtype Fand O, all the other Envs derived from other subtypes (A, B, C,and D) were aggregated. Therefore, it is not trivial to purify Envproteins in correct structure and conformation.
To further evaluate subtype C Env structure, we performeddeep-stain EM analysis and found the trimers to be in two basicconformations, which were characterized as open and closed.This is in contrast to our previous observations with subtype BEnv, which appeared to be in a single, closed conformation(Srivastava et al., 2003). It is possible that a more relaxed or openconformation, such as the one we observed for subtype C Env,may provide more access thereby inducing a broader spectrumof antibody responses.
We have determined the structural integrity of purifiedsubtype C trimer using several mAbs of known epitope speci-ficity such as those near or on the CD4-binding site (recognizedby mAbs IgG1b12 and CD4-IgG2), the CD4-inducible site thatoverlaps the co-receptor binding site (recognized by mAb 17band 4.8d, 2.3 E, LF17), gp41 (2F5), a carbohydrate-dependentepitope on gp120 (recognized by 2G12), and V3 loop epitopes(recognized by mAbs 447D and 391-95d). Interestingly,

285I.K. Srivastava et al. / Virology 372 (2008) 273–290
qualitative and quantitative differences were noted for subtype Cversus subtype B Env (Srivastava et al., 2003). Trimers derivedfrom subtypes B and C bound to CD4 and mAb IgG-CD4;however, subtype C trimers showed no binding to b12 in captureELISA assay and had 20-fold reduced binding to b12 byBiacore. Similarly, Jeffs et al. (2004) did not observe binding ofsubtype C Env to b12. Is it because the epitope recognized byb12 was not preserved on subtype C trimers, or because the b12epitope is different on subtype C compared to subtype B? Wehave aligned the amino acid residues involved in b12 bindingbetween subtype B and subtype C isolates, and observed that 5out of 15 residues in subtype C are different compared to subtypeB. If the difference in b12 binding between subtype B and Ctrimeric V2 Env can be attributed to this difference, remains to bedetermined. It has been observed by David Montefiori (personalcommunication) and Julie Overbaugh (personal communica-tion) that the concentration of b12 required to neutralize TV1virus is at least a log higher than that required for SF162 virus.These data are consistent to the fact that b12 epitope is notcompletely preserved in subtype C isolate compared to subtypeB. In general, the magnitude of binding of mAbs directed againstthe CD4i epitopes was lower for subtype C trimer compared tosubtype B trimer either in the presence or absence of CD4.However, the fold increase in the mAbs directed against CD4-inducible epitopes observed in presence of CD4 was similar forsubtype B and C trimers, suggesting that subtype C trimer alsounderwent structural rearrangement. The other broadly neutral-izing epitopes such as those recognized by mAbs 2F5 and 4E10were exposed and preserved in subtype C Env trimers. However,these epitopes appear to have been lost or not preserved in Envderived from another subtype C isolate BA29 (Jeffs et al., 2004).Whether this was due to differences between the TV1 and BA29strains or due to aggregation of the BA29 Env preparation re-mains to be determined.
Taken together, the data demonstrate that a V2-deleted form ofEnv trimer derived from theTV1 subtypeC strain can be producedin – and purified from – CHO cells. The resulting preparation ispredominantly trimerwithmany similar biophysical, biochemical,and immunological characteristics to subtype B Env protein.Furthermore, it has induced nAb responses in rabbits, consistentwith our previously reported immunogenicity studies in rabbitsand rhesusmacaques (Lian et al., 2005). These data emphasize thepotential role of trimeric Envs for vaccine applications.
Materials and methods
Cloning full length or V2-modified subtype C HIV envelopevariants
The sequences encoding the open reading frame of theectodomain of the Env protein from the HIV-1 TV1ΔV2 isolateswere codon-modified as described elsewhere (Haas et al., 1996;zur Megede et al., 2000), and constructed synthetically as a2.1 kb EcoRI–XbaI DNA fragment. In order to stabilize theoligomeric structure of the encoded gp140 protein, we in-troduced a series of mutations in the primary (REKR) and sec-ondary protease (KAKRR) cleavage sites in the Env polypeptide
(Earl and Moss, 1993). The details of these mutations arepresented in Fig. 1. The synthetic PCR products were digestedwith EcoRI/XhoI and cloned into the pCMVIII vector for proteinproduction. The pCMVIII vector contains the CMV immediateearly enhancer and promoter, a polylinker for multiple cloningsites, a bovine growth hormone polyA termination signal, akanamycin resistance gene, and an E. coli origin of replication.In addition, it contains Mdhfr (Dihydrofolate reductase) andNeomycin resistance genes that allow for stable transfection,selection and amplification of the gene of interest in ChineseHamster Ovary (CHO) cells. Similar methods were used to clonea consensus envelope gene (E0975) derived from publishedsubtype C sequences from Botswana.
Transient transfection
Plasmids encoding the various gp140 forms of the TV1 envwere transiently transfected into 293T cells, and gp140 proteinexpression was quantitated 48 to 72 hour post-transfection in celllysates and in supernatants. The supernatants were used forpartial purification of the Env protein from different clones.These semi-purified subtype C Envs were screened for theirability to bind CD4, and to determine levels of expression bySDS-PAGE analysis and quantitative immunoprobing.
Development of stable CHO cell lines secreting o-gp140ΔV2TV1
Stable CHO cell lines secreting gp140ΔV2TV1were derivedusing DG-44 cells that have a double deletion mutation in theDHFR gene, thus making the cell line dependent on the additionof hypoxanthine, glycine, and thymidine to the growth media,following the experimental protocol described elsewhere(Srivastava et al., 2002).
Production of o-gp140ΔV2TV1
The three highest expressing gp140ΔV2TV1-CHO cellclones were used to seed 3 L bioreactors (one for each clone)for adaptation to low serum containing media (0.5%) and celldensity and perfusion rate that facilitated the maximumexpression of gp140ΔV2TV1. Bioreactors were monitoreddaily for cell density, pH, dCO2, dO2, glucose, lactate,glutamine, and ammonia concentrations. The structure, confor-mation, and expression levels of secreted gp140ΔV2TV1 weremonitored weekly. From the three bioreactor-adapted clones, thebest producer clone was used to seed a 12.5 L bioreactor forproduction run. At the end of the run, collected media wasconcentrated 20-fold through a 100-kDa-membrane filter andstored at −80 °C in presence of 1 mM EDTA and 1 mM EGTA.
Purification of o-gp140ΔV2TV1
The o-gp140ΔV2TV1 was purified from the concentratedCHO cell supernatant using a lectin capture, and two ion ex-change steps as described elsewhere in detail (Srivastava et al.,2003, 2002). All the column fractions were analyzed on poly-acrylamide gels (PAGE; native, and reduced and denatured

286 I.K. Srivastava et al. / Virology 372 (2008) 273–290
conditions) and also in a CD4 receptor-binding assay (seebelow). The fractions showing strong CD4 binding activity werepooled and fractionated on a 16×90 mm Superdex-200column(Pharmacia, Sweden) equilibrated with 10 mM NaCitratecontaining 300 mM NaCl. Fractions containing o-gp140ΔV2were pooled, concentrated using a Stir cell (Millipore, Inc.,Bedford, MA), and stored frozen at −80 °C. During the puri-fication process, fractions were analyzed by PAGE both underreducing and denaturing and under native conditions followingstandard methods. Gels were stained with Coomassie brilliantblue or processed for immunoblotting.
Biophysical characterization of o-gp140ΔV2TV1
The molecular mass (Mw), hydrodynamic radius (Rh), andintrinsic viscosity (IV) of o-gp140ΔV2TV1 were determinedusing a triple detector array system (TDA 300, Viscotek Corp.,Houston, TX) in conjunction with a pre-calibrated gel filtrationHPLC column (Bio Sil SEC-250; BioRad Laboratories, Hercules,CA) using Alliance 2690 HPLC system (Waters Corporation,Milford, MA). Approximately 100 μg of the purified protein(gp140ΔV2TV1) was analyzed in 20 mM NaH2PO4, 2 mMNa2HPO4 buffer at pH7.4 at the flow rate of 1ml/min. The signalsfrom light scattering, viscometer, and refractive index detectorswere collected and analyzed using EZ Pro software package(Viscotek Corp., Houston, TX).
CD4 binding assay
To determine the capability of purified envelope protein to bindCD4, we used an HPLC-based assay using fluorescinated CD4 asdescribed elsewhere in detail (Srivastava et al., 2002). PurifiedsCD4 was labeled with amine-reactive succinimidyl esters ofcarboxyfluorescein following the instructions provided by manu-facturer (Molecular Probes, Inc., Eugene, OR). Approximately1μg of purifiedHIVEnv protein gp120wasmixedwith 0.33 μg offluorescinated-CD4 (equal molar concentrations) in a reactionvolume of 60 μl using 2× phosphate-buffered saline at pH 7.4.After a 15 min incubation at the RT, 50 μl of this sample wasinjected onto a Bio Sil SEC-250 (Bio-Rad laboratories, Hercules,CA) gel filtration high performance liquid chromatography(HPLC) column using Alliance 2690 HPLC system (WatersCorporation, Milford, MA). Samples were run in 20 mMNaH2PO4, 2 mM Na2HPO4,and 400 mM ammonium sulfatebuffer, pH6.0, at the flow rate of 1ml/min. The fluorescence profilewas monitored at 490 nm using a 996 fluorescent detector andMillennium software package (Waters Corporation, Milford, MA).
Glycosylation profile analysis
The glycosylation profile of the purified proteins wasassessed by four methods as described elsewhere in detail(Srivastava et al., 2002): first, enzymatic deglycosylation of thepurified HIV-Env proteins was performed using reagents fromBioRad Research Laboratories, Hercules, CA (catalog # 170-6500). Briefly, to detect N-linked and O-linked oligosacchar-ides, 25–100 μg of the purified protein was treated separately
with Peptidyl N-glycosidase F (PNGF) (2.5 U/ml; at 37 °C for1 h); and NANase II and O-glycosidase (1 U/ml; at 37 °C for1 h). At the end of the incubation period, samples were diluted insample buffer containing 0.1% SDS and β-mercaptoethanol andanalyzed by SDS-PAGE. In addition, we have also performedEndo-H digestion of the purified protein following manufac-turer's protocol (Boehringer Mannheim).
Immunochemical characterization of o-gp140ΔV2TV1
The binding of well-characterized HIV-Env specific mAbs,all derived from clade B HIV-1-infected patients, was performedby a capture ELISA. Subtype C ΔV2 trimers were captured byits gp41 portion via the mAb D50 that recognizes a linear gp41peptide (Earl et al., 1997) as described previously (Stamatatosand Cheng-Mayer, 1995). Briefly, ELISA plates were coatedwith these respective antibodies at a concentration of 10 μg/mland o-gp140ΔV2 from subtypes B and C was captured. Thecaptured proteins were incubated with serial dilutions of variousmAbs (0.01 mg/ml to 10 mg/ml) for 2 hour at RT. Plates werewashed, and specifically bound antibodies were detected usinganti-mouse IgG coupled to alkaline phosphatase as describedearlier (Srivastava et al., 2003, 2002).
Surface plasmon resonance assay
Surface Plasmon Resonance assays were performed using aBIACORE 3000 optical biosensor system (Biacore AB,Uppsala,Sweden) with simultaneous monitoring of relevant flow cells. Toperform the kinetic study of the binding of subtype CΔV2 trimerand subtype B ΔV2 trimer to sCD4, sCD4 was immobilizedusing carbodiimide coupling onto a low charge CM3 sensor chipto attain a response of 2000 RU. Using PBS (pH 7.4) with 0.05%Tween 20, association was assessed by passing subtype C ΔV2trimer and subtype B ΔV2 trimer over the CD4 in different flowcells at a flow rate of 20–30 μl/min. The concentration range forsubtype C ΔV2 trimer and subtype B ΔV2 trimer was 15 to500 mM. We have used Bovine Serum Albumin (BSA) as nega-tive control. For kinetic analysis, Biosensor data were preparedby subtracting binding responses obtained for BSA from thespecific responses. The association and dissociation phase datawere fitted simultaneously to a single-site binding model byusing the nonlinear data analysis software BIAevaluation 3.2.
mAbs binding assay
To perform the binding of HIV Env to different mAbs such asb12, 2F5, 4E10, 2G12, 17b, 48d, 23E, and LF17, these antibodieswere immobilized on to CM5 sensor chip using amine couplingreaction to attain approximately 1000 RUs, and the assay wasperformed under the experimental conditions described as above.
Isothermal titration calorimetric (ITC) analysis
Isothermal titration calorimetry was performed on a VP-ITC(Microcal LLC, Northampton, MA) using a 1.4 ml cell, and a0.25 ml syringe. Proteins were dialyzed extensively in PBS

287I.K. Srivastava et al. / Virology 372 (2008) 273–290
containing 0.005% Tween and concentrated using a stir cell(Millipore Inc.) prior to loading. Three to five millimolar CD4was loaded in the cell, with 18 to 30 mM envelope protein in thesyringe. Titrations were performed using 15 μl injections with a5 min delay between each injection. All runs were adjusted forheat of dilution and mixing by subtracting a blank run ofenvelope titrated into buffer only. Binding isotherms obtainedwere auto-fitted for a floating number of binding sites usingOrigin® software (MicroCal LLC, Northampton,MA). Fits weregenerated using the Origin® 7.0 software parameter “ligand incell” option; thus, generated figures for the trimers do not reflectthe n values on the X-axis due to the software, which does not re-calculate the molar ratio on the figure but does reflect the changein the fit obtained (MicroCal LLC, Northampton, MA).
Immunoelectron microscopy
EM analysis of HIV Env derived from subtype C in differentconformations such as trimer, dimer, and monomer wasperformed by negative staining as previously described(Srivastava et al., 2003). The trimeric (pool 3; Fig. 4A), dimeric(pool 4; Fig. 4A), and monomeric (pool 5; Fig. 4A) proteinsobtained after the final preparative sizing column were diluted inborate-buffered saline, affixed to carbon membranes, stainedwith 1% uranyl formate, and mounted on copper grids foranalysis. Electron micrographs were recorded at a nominalmagnification of 100,000× at 100 kVon a JEOL JEM 1200EXelectron microscope. Measurements were made using imageanalysis software Image-Pro Plus (http://www.mediacy.com/).Fifty or more particles from fractions corresponding to theputative monomer/dimer and trimer of gp140ΔV2 TV1 weremeasured and analyzed statistically.
Acknowledgments
We thank Dr. Rino Rappuoli, Vice President and Head ofVaccines Research for his support and encouragement. We alsothankDrs. SusanEngelbrecht andEstrelita Janse vanRensburg forvirus isolates and outstanding collaboration in subtype C vaccinedevelopment; Dennis Burton, Susan Zolla-Pazner, James Robin-son, Michael Gorny, Herman Katinger, and Gabriela Steigler forproviding mAbs for the structural characterization of the purifiedproteins; Mark Wininger and Karen Matsuoka for their help incarrying out large-scale cell cultures; and Nelle Cronen and KCEgan, for excellent editorial and administrative support. This workreceived financial support provided by NIH CRDA (NO-AI-25473, TO#1, NO-AI-95367, TO#3) and D&DT contract (NO-1-AI-05396). Research in Drs. Stamatatos and Cheng's labo-ratories was supported by investigator-initiated NIH researchgrants RO1 AI47708 (LS) and UC Discovery grant (RHC).
References
Barnett, S.W., Rajasekar, S., Legg, H., Doe, B., Fuller, D.H., Haynes, J.R.,Walker, C.M., Steimer, K.S., 1997. Vaccination with HIV-1 gp120 DNAinduces immune responses that are boosted by a recombinant gp120 proteinsubunit. Vaccine 15 (8), 869–873.
Barnett, S.W., Lu, S., Srivastava, I., Cherpelis, S., Gettie, A., Blanchard, J.,Wang, S., Mboudjeka, I., Leung, L., Lian, Y., Fong, A., Buckner, C., Ly, A.,Hilt, S., Ulmer, J.,Wild, C.T.,Mascola, J.R., Stamatatos, L., 2001. The abilityof an oligomeric human immunodeficiency virus type 1 (HIV-1) envelopeantigen to elicit neutralizing antibodies against primary HIV-1 isolates isimproved following partial deletion of the second hypervariable region.J. Virol. 75 (12), 5526–5540.
Barouch, D.H., Letvin, N.L., 2002. Viral evolution and challenges in thedevelopment of HIV vaccines. Vaccine 20 (Suppl 4), A66–A68.
Barouch, D.H., Kunstman, J., Kuroda, M.J., Schmitz, J.E., Santra, S., Peyerl, F.W.,Krivulka, G.R., Beaudry, K., Lifton, M.A., Gorgone, D.A., Montefiori, D.C.,Lewis, M.G., Wolinsky, S.M., Letvin, N.L., 2002. Eventual AIDS vaccinefailure in a rhesus monkey by viral escape from cytotoxic T lymphocytes.Nature 415 (6869), 335–339.
Barouch, D.H., Kunstman, J., Glowczwskie, J., Kunstman, K.J., Egan, M.A.,Peyerl, F.W., Santra, S., Kuroda, M.J., Schmitz, J.E., Beaudry, K., Krivulka,G.R., Lifton, M.A., Gorgone, D.A., Wolinsky, S.M., Letvin, N.L., 2003.Viral escape from dominant simian immunodeficiency virus epitope-specificcytotoxic T lymphocytes in DNA-vaccinated rhesus monkeys. J. Virol. 77(13), 7367–7375.
Barr, P.J., Steimer, K.S., Sabin, E.A., Parkes, D., George-Nascimento, C.,Stephans, J.C., Powers, M.A., Gyenes, A., Van Nest, G.A., Miller, E.T., et al.,1987. Antigenicity and immunogenicity of domains of the humanimmunodeficiency virus (HIV) envelope polypeptide expressed in the yeastSaccharomyces cerevisiae. Vaccine 5 (2), 90–101.
Binley, J.M., Sanders, R.W., Clas, B., Schuelke, N., Master, A., Guo, Y.,Kajumo, F., Anselma, D.J., Maddon, P.J., Olson, W.C., Moore, J.P., 2000. Arecombinant human immunodeficiency virus type 1 envelope glycoproteincomplex stabilized by an intermolecular disulfide bond between the gp120and gp41 subunits is an antigenic mimic of the trimeric virion-associatedstructure. J. Virol. 74 (2), 627–643.
Broder, C.C., Earl, P.L., Long, D., Abedon, S.T., Moss, B., Doms, R.W., 1994.Antigenic implications of human immunodeficiency virus type 1 envelopequaternary structure: oligomer-specific and -sensitive monoclonal anti-bodies. Proc. Natl. Acad. Sci. U. S. A. 91 (24), 11699–11703.
Chen, B., Vogan, E.M., Gong, H., Skehel, J.J., Wiley, D.C., Harrison, S.C.,2005. Structure of an unliganded simian immunodeficiency virus gp120core. Nature 433 (7028), 834–841.
Cherpelis, S., Jin, X., Gettie, A., Ho, D.D., Barnett, S.W., Shrivastava, I.,Stamatatos, L., 2001a. DNA-immunization with a V2 deleted HIV-1 envelopeelicits protective antibodies in macaques. Immunol. Lett. 79 (1–2), 47–55.
Cherpelis, S., Srivastava, I., Gettie, A., Jin, X., Ho, D.D., Barnett, S.W.,Stamatatos, L., 2001b. DNAvaccination with the human immunodeficiencyvirus type 1 SF162DeltaV2 envelope elicits immune responses that offerpartial protection from simian/human immunodeficiency virus infection toCD8(+) T-cell-depleted rhesus macaques. J. Virol. 75 (3), 1547–1550.
Dalgleish, A.G., Beverley, P.C., Clapham, P.R., Crawford, D.H., Greaves, M.F.,Weiss, R.A., 1984. The CD4 (T4) antigen is an essential component of thereceptor for the AIDS retrovirus. Nature 312 (5996), 763–767.
Earl, P.L., Moss, B., 1993. Mutational analysis of the assembly domain of theHIV-1 envelope glycoprotein. AIDS Res. Hum. Retrovir. 9 (7), 589–594.
Earl, P.L., Doms, R.W., Moss, B., 1990. Oligomeric structure of the humanimmunodeficiency virus type 1 envelope glycoprotein. Proc. Natl. Acad.Sci. U. S. A. 87 (2), 648–652.
Earl, P.L., Koenig, S., Moss, B., 1991. Biological and immunological propertiesof human immunodeficiency virus type 1 envelope glycoprotein: analysis ofproteins with truncations and deletions expressed by recombinant vacciniaviruses. J. Virol. 65 (1), 31–41.
Earl, P.L., Broder, C.C., Long, D., Lee, S.A., Peterson, J., Chakrabarti, S.,Doms, R.W., Moss, B., 1994. Native oligomeric human immunodeficiencyvirus type 1 envelope glycoprotein elicits diverse monoclonal antibodyreactivities. J. Virol. 68 (5), 3015–3026.
Earl, P.L., Broder, C.C., Doms, R.W., Moss, B., 1997. Epitope map of humanimmunodeficiency virus type 1 gp41 derived from 47 monoclonal antibodiesproduced by immunization with oligomeric envelope protein. J. Virol. 71(4), 2674–2684.
Earl, P.L., Sugiura, W., Montefiori, D.C., Broder, C.C., Lee, S.A., Wild, C.,Lifson, J., Moss, B., 2001. Immunogenicity and protective efficacy of

288 I.K. Srivastava et al. / Virology 372 (2008) 273–290
oligomeric human immunodeficiency virus type 1 gp140. J. Virol. 75 (2),645–653.
Farzan, M., Choe, H., Desjardins, E., Sun, Y., Kuhn, J., Cao, J., Archambault,D., Kolchinsky, P., Koch, M., Wyatt, R., Sodroski, J., 1998. Stabilization ofhuman immunodeficiency virus type 1 envelope glycoprotein trimers bydisulfide bonds introduced into the gp41 glycoprotein ectodomain. J. Virol.72 (9), 7620–7625.
Fennie, C., Lasky, L.A., 1989. Model for intracellular folding of the humanimmunodeficiency virus type 1 gp120. J. Virol. 63 (2), 639–646.
Fenouillet, E., Clerget-Raslain, B., Gluckman, J.C., Guetard, D., Montagnier, L.,Bahraoui, E., 1989. Role of N-linked glycans in the interaction between theenvelope glycoprotein of human immunodeficiency virus and its CD4cellular receptor. Structural enzymatic analysis. J. Exp. Med. 169 (3),807–822.
Fouts, T.R., Binley, J.M., Trkola, A., Robinson, J.E., Moore, J.P., 1997.Neutralization of the human immunodeficiency virus type 1 primary isolateJR-FL by human monoclonal antibodies correlates with antibody binding tothe oligomeric form of the envelope glycoprotein complex. J. Virol. 71 (4),2779–2785.
Geyer, H., Holschbach, C., Hunsmann, G., Schneider, J., 1988. Carbohydratesof human immunodeficiency virus. Structures of oligosaccharides linked tothe envelope glycoprotein 120. J. Biol. Chem. 263 (24), 11760–11767.
Graham,B.S., 2000. Clinical Trials ofHIVVaccines, pp. 207–221.HIVDatabase:http://hiv-web.lanl.gov/content/hiv-db/REVIEWS/GRAHAM2000/Graham.html.
Grundner, C., Mirzabekov, T., Sodroski, J., Wyatt, R., 2002. Solid-phaseproteoliposomes containing human immunodeficiency virus envelopeglycoproteins. J. Virol. 76 (7), 3511–3521.
Haas, J., Park, E.C., Seed, B., 1996. Codon usage limitation in the expression ofHIV-1 envelope glycoprotein. Curr. Biol. 6 (3), 315–324.
Haigwood, N.L., Nara, P.L., Brooks, E., Van Nest, G.A., Ott, G., Higgins, K.W.,Dunlop, N., Scandella, C.J., Eichberg, J.W., Steimer, K.S., 1992. Native butnot denatured recombinant human immunodeficiency virus type 1 gp120generates broad-spectrum neutralizing antibodies in baboons. J. Virol. 66,172–182.
Iyer, S.P., Franti, M., Krauchuk, A.A., Fisch, D.N., Ouattara, A.A., Roux, K.H.,Krawiec, L., Dey, A.K., Beddows, S., Maddon, P.J., Moore, J.P., Olson, W.C.,2007. Purified, proteolytically mature HIV type 1 SOSIP gp140 envelopetrimers. AIDS Res. Hum. Retrovir. 23 (6), 817–828.
Jeffs, S.A., Goriup, S., Kebble, B., Crane, D., Bolgiano, B., Sattentau, Q., Jones,S., Holmes, H., 2004. Expression and characterisation of recombinantoligomeric envelope glycoproteins derived from primary isolates of HIV-1.Vaccine 22 (8), 1032–1046.
Johnson, W.E., Morgan, J., Reitter, J., Puffer, B.A., Czajak, S., Doms, R.W.,Desrosiers, R.C., 2002. A replication-competent, neutralization-sensitivevariant of simian immunodeficiency virus lacking 100 amino acids ofenvelope. J. Virol. 76 (5), 2075–2086.
Johnston, M.I., Flores, J., 2001. Progress in HIV vaccine development. Curr.Opin. Pharmacol. 1 (5), 504–510.
Klatzmann, D., Champagne, E., Chamaret, S., Gruest, J., Guetard, D., Hercend,T., Gluckman, J.C., Montagnier, L., 1984. T-lymphocyte T4 moleculebehaves as the receptor for human retrovirus LAV. Nature 312 (5996),767–768.
Kozarsky, K., Penman,M., Basiripour, L., Haseltine,W., Sodroski, J., Krieger,M.,1989. Glycosylation and processing of the human immunodeficiency virustype 1 envelope protein. J. Acquir. Immune Defic. Syndr. 2 (2), 163–169.
Lasky, L.A., Nakamura, G., Smith, D.H., Fennie, C., Shimasaki, C., Patzer, E.,Berman, P., Gregory, T., Capon, D.J., 1987. Delineation of a region of thehuman immunodeficiency virus type 1 gp120 glycoprotein critical forinteraction with the CD4 receptor. Cell 50 (6), 975–985.
Leonard, C.K., Spellman, M.W., Riddle, L., Harris, R.J., Thomas, J.N., Gregory,T.J., 1990. Assignment of intrachain disulfide bonds and characterization ofpotential glycosylation sites of the type 1 recombinant human immunode-ficiency virus envelope glycoprotein (gp120) expressed in Chinese hamsterovary cells. J. Biol. Chem. 265 (18), 10373–10382.
Li, Y., Luo, L., Rasool, N., Kang, C.Y., 1993. Glycosylation is necessary for thecorrect folding of human immunodeficiency virus gp120 in CD4 binding.J. Virol. 67 (1), 584–588.
Lian, Y., Srivastava, I., Gomez-Roman, V.R., Zur Megede, J., Sun, Y., Kan, E.,Hilt, S., Engelbrecht, S., Himathongkham, S., Luciw, P.A., Otten, G., Ulmer,J.B., Donnelly, J.J., Rabussay, D., Montefiori, D., van Rensburg, E.J.,Barnett, S.W., 2005. Evaluation of envelope vaccines derived from theSouth African subtype C human immunodeficiency virus type 1 TV1 strain.J. Virol. 79 (21), 13338–13349.
Ly, A., Stamatatos, L., 2000. V2 loop glycosylation of the humanimmunodeficiency virus type 1 SF162 envelope facilitates interaction ofthis protein with CD4 and CCR5 receptors and protects the virus fromneutralization by anti-V3 loop and anti-CD4 binding site antibodies. J. Virol.74 (15), 6769–6776.
Maezawa, S., Hayashi, Y., Nakae, T., Ishii, J., Kameyama, K., Takagi, T., 1983.Determination of molecular weight of membrane proteins by the use of low-angle laser light scattering combined with high-performance gel chroma-tography in the presence of a non-ionic surfactant. Biochim. Biophys. Acta747 (3), 291–297.
Mascola, J.R., Mathieson, B.J., Zack, P.M., Walker, M.C., Halstead, S.B.,Burke, D.S., 1993. Summary report: workshop on the potential risks ofantibody-dependent enhancement in human HIV vaccine trials. AIDS Res.Hum. Retrovir. 9 (12), 1175–1184.
Mascola, J.R., Snyder, S.W., Weislow, O.S., Belay, S.M., Belshe, R.B.,Schwartz, D.H., Clements, M.L., Dolin, R., Graham, B.S., Gorse, G.J.,Keefer, M.C., McElrath, M.J., Walker, M.C., Wagner, K.F., McNeil, J.G.,McCutchan, F.E., Burke, D.S., 1996. Immunization with envelope subunitvaccine products elicits neutralizing antibodies against laboratory-adaptedbut not primary isolates of human immunodeficiency virus type 1. TheNational Institute of Allergy and Infectious Diseases AIDS VaccineEvaluation Group. J. Infect. Dis. 173 (2), 340–348.
Mascola, J.R., Lewis, M.G., Stiegler, G., Harris, D., VanCott, T.C., Hayes, D.,Louder, M.K., Brown, C.R., Sapan, C.V., Frankel, S.S., Lu, Y., Robb, M.L.,Katinger, H., Birx, D.L., 1999. Protection of Macaques against pathogenicsimian/human immunodeficiency virus 89.6PD by passive transfer ofneutralizing antibodies. J. Virol. 73 (5), 4009–4018.
Mascola, J.R., Lewis, M.G., VanCott, T.C., Stiegler, G., Katinger, H., Seaman,M., Beaudry, K., Barouch, D.H., Korioth-Schmitz, B., Krivulka, G.,Sambor, A., Welcher, B., Douek, D.C., Montefiori, D.C., Shiver, J.W.,Poignard, P., Burton, D.R., Letvin, N.L., 2003. Defining the protectiveantibody response for HIV-1. Curr. Mol. Med. 3 (3), 209–216.
McDougal, J.S., Nicholson, J.K., Cross, G.D., Cort, S.P., Kennedy, M.S.,Mawle, A.C., 1986. Binding of the human retrovirus HTLV-III/LAV/ARV/HIV to the CD4 (T4) molecule: conformation dependence, epitope mapping,antibody inhibition, and potential for idiotypic mimicry. J. Immunol. 137(9), 2937–2944.
Mizuochi, T., Spellman, M.W., Larkin, M., Solomon, J., Basa, L.J., Feizi, T.,1988. Structural characterization by chromatographic profiling of theoligosaccharides of human immunodeficiency virus (HIV) recombinantenvelope glycoprotein gp120 produced in Chinese hamster ovary cells.Biomed. Chromatogr. 2 (6), 260–270.
Moore, J.P., Sattentau, Q.J., Wyatt, R., Sodroski, J., 1994. Probing the structureof the human immunodeficiency virus surface glycoprotein gp120 with apanel of monoclonal antibodies. J. Virol. 68 (1), 469–484.
Moore, J.P., Cao, Y., Qing, L., Sattentau, Q.J., Pyati, J., Koduri, R., Robinson, J.,Barbas, C.F., Burton, D.R., Ho, D.D., 1995. Primary isolates of humanimmunodeficiency virus type 1 are relatively resistant to neutralization bymonoclonal antibodies to gp120, and their neutralization is not predicted bystudies with monomeric gp120. J. Virol. 69 (1), 101–109.
Myszka, D.G., Sweet, R.W., Hensley, P., Brigham-Burke, M., Kwong, P.D.,Hendrickson, W.A., Wyatt, R., Sodroski, J., Doyle, M.L., 2000. Energeticsof the HIV gp120-CD4 binding reaction. Proc. Natl. Acad. Sci. U. S. A. 97(16), 9026–9031.
Pancera,M., Lebowitz, J., Schon,A., Zhu, P., Freire, E., Kwong, P.D., Roux,K.H.,Sodroski, J., Wyatt, R., 2005. Soluble mimetics of human immunodeficiencyvirus type 1 viral spikes produced by replacement of the native trimerizationdomain with a heterologous trimerization motif: characterization and ligandbinding analysis. J. Virol. 79 (15), 9954–9969.
Papandreou, M.J., Idziorek, T., Miquelis, R., Fenouillet, E., 1996. Glycosylationand stability of mature HIV envelope glycoprotein conformation undervarious conditions. FEBS Lett. 379 (2), 171–176.

289I.K. Srivastava et al. / Virology 372 (2008) 273–290
Parren, P.W., Marx, P.A., Hessell, A.J., Luckay, A., Harouse, J., Cheng-Mayer,C., Moore, J.P., Burton, D.R., 2001. Antibody protects macaques againstvaginal challenge with a pathogenic R5 simian/human immunodeficiencyvirus at serum levels giving complete neutralization in vitro. J. Virol. 75(17), 8340–8347.
Quinnan Jr., G.V., Yu, X.F., Lewis,M.G., Zhang, P.F., Sutter, G., Silvera, P., Dong,M., Choudhary, A., Sarkis, P.T., Bouma, P., Zhang, Z., Montefiori, D.C.,Vancott, T.C., Broder, C.C., 2005. Protection of rhesus monkeys againstinfection with minimally pathogenic simian-human immunodeficiency virus:correlations with neutralizing antibodies and cytotoxic Tcells. J. Virol. 79 (6),3358–3369.
Roben, P., Moore, J.P., Thali, M., Sodroski, J., Barbas, C.F., Burton, D.R., 1994.Recognition properties of a panel of human recombinant Fab fragments tothe CD4 binding site of gp120 that show differing abilities to neutralizehuman immunodeficiency virus type 1. J. Virol. 68 (8), 4821–4828.
Robert-Guroff, M., Kaur, H., Patterson, L.J., Leno, M., Conley, A.J., McKenna,P.M., Markham, P.D., Richardson, E., Aldrich, K., Arora, K., Murty, L.,Carter, L., Zolla-Pazner, S., Sinangil, F., 1998. Vaccine protection against aheterologous, non-syncytium-inducing, primary human immunodeficiencyvirus. J. Virol. 72 (12), 10275–10280.
Ruprecht, R.M., Hofmann-Lehmann, R., Smith-Franklin, B.A., Rasmussen, R.A.,Liska, V., Vlasak, J., Xu, W., Baba, T.W., Chenine, A.L., Cavacini, L.A.,Posner, M.R., Katinger, H., Stiegler, G., Bernacky, B.J., Rizvi, T.A., Schmidt,R., Hill, L.R., Keeling, M.E., Montefiori, D.C., McClure, H.M., 2001.Protection of neonatal macaques against experimental SHIV infection byhuman neutralizing monoclonal antibodies. Transfus. Clin. Biol. 8 (4),350–358.
Sanders, R.W., Schiffner, L., Master, A., Kajumo, F., Guo, Y., Dragic, T., Moore,J.P., Binley, J.M., 2000. Variable-loop-deleted variants of the humanimmunodeficiency virus type 1 envelope glycoprotein can be stabilized byan intermolecular disulfide bond between the gp120 and gp41 subunits.J. Virol. 74 (11), 5091–5100.
Sanders, R.W., Vesanen, M., Schuelke, N., Master, A., Schiffner, L.,Kalyanaraman, R., Paluch, M., Berkhout, B., Maddon, P.J., Olson, W.C.,Lu, M., Moore, J.P., 2002. Stabilization of the soluble, cleaved, trimericform of the envelope glycoprotein complex of human immunodeficiencyvirus type 1. J. Virol. 76 (17), 8875–8889.
Scandella, C.J., Kilpatrick, J., Lidster, W., Parker, C., Moore, J.P., Moore, G.K.,Mann, K.A., Brown, P., Coates, S., Chapman, B., et al., 1993. Nonaffinitypurification of recombinant gp120 for use in AIDS vaccine development.AIDS Res. Hum. Retrovir. 9 (12), 1233–1244.
Schulke, N., Vesanen, M.S., Sanders, R.W., Zhu, P., Lu, M., Anselma, D.J.,Villa, A.R., Parren, P.W., Binley, J.M., Roux, K.H., Maddon, P.J., Moore, J.P.,Olson, W.C., 2002. Oligomeric and conformational properties of aproteolytically mature, disulfide-stabilized human immunodeficiency virustype 1 gp140 envelope glycoprotein. J. Virol. 76 (15), 7760–7776.
Sharma, V.A., Kan, E., Sun, Y., Lian, Y., Cisto, J., Frasca, V., Hilt, S.,Stamatatos, L., Donnelly, J.J., Ulmer, J.B., Barnett, S.W., Srivastava, I.K.,2006. Structural characteristics correlate with immune responses induced byHIV envelope glycoprotein vaccines. Virology 352, 131–144.
Srivastava, I.K., Stamatatos, L., Legg, H., Kan, E., Fong, A., Coates, S.R.,Leung, L., Wininger, M., Donnelly, J.J., Ulmer, J.B., Barnett, S.W., 2002.Purification and characterization of oligomeric envelope glycoprotein froma primary r5 subtype B human immunodeficiency virus. J. Virol. 76 (6),2835–2847.
Srivastava, I.K., Stamatatos, L., Kan, E., Vajdy, M., Lian, Y., Hilt, S., Martin, L.,Vita, C., Zhu, P., Roux, K.H., Vojtech, L., D, C.M., Donnelly, J., Ulmer, J.B.,Barnett, S.W., 2003. Purification, characterization, and immunogenicity of asoluble trimeric envelope protein containing a partial deletion of the V2 loopderived from SF162, an R5-tropic human immunodeficiency virus type 1isolate. J. Virol. 77 (20), 11244–11259.
Stamatatos, L., Cheng-Mayer, C., 1995. Structural modulations of the envelopegp120 glycoprotein of human immunodeficiency virus type 1 upon oligo-merization and differential V3 loop epitope exposure of isolates displayingdistinct tropism upon virion-soluble receptor binding. J. Virol. 69 (10),6191–6198.
Stamatatos, L., Cheng-Mayer, C., 1998a. An envelope modification that rendersa primary, neutralization-resistant clade B human immunodeficiency virus
type 1 isolate highly susceptible to neutralization by sera from other clades.J. Virol. 72, 7840–7845.
Stamatatos, L., Cheng-Mayer, C., 1998b. An envelope modification that rendersa primary, neutralization-resistant clade B human immunodeficiency virustype 1 isolate highly susceptible to neutralization by sera from other clades.J. Virol. 72 (10), 7840–7845.
Steimer, K.S., Haigwood, N.L., 1991. Importance of conformation on theneutralizing antibody response to HIV-1 gp120. Biotechnol. Ther. 2 (1–2),63–89.
Stites, W.E., Pantaleo, G., Koup, R.A., 1997. Protein–protein interactions:interface structure, binding thermodynamics, and mutational analysiscorrelates of immune protection in HIV-1 infection: what we know,what we don't know, what we should know. Chem. Rev. 97 (5),1233–1250.
Sullivan,N., Sun, Y., Binley, J., Lee, J., Barbas III, C.F., Parren, P.W., Burton, D.R.,Sodroski, J., 1998. Determinants of human immunodeficiency virus type 1envelope glycoprotein activation by soluble CD4 and monoclonal antibodies.J. Virol. 72 (8), 6332–6338.
UNAIDS/WHO, 2004. 2004 Report on the Global AIDS Epidemic.Vajdy, M., Singh, M., Kazzaz, J., Soenawan, E., Ugozzoli, M., Zhou, F.,
Srivastava, I., Bin, Q., Barnett, S., Donnelly, J., Luciw, P., Lourdes, L.,Montefiori, D., O'Hagan, D., 2004.Mucosal and systemic anti-HIVresponsesin rhesus macaques following combinations of intra-nasal and parenteralimmunizations. AIDS Res. Hum.Retrovir. 20 (11), 1269–1281.
VanCott, T.C., Bethke, F.R., Burke, D.S., Redfield, R.R., Birx, D.L., 1995. Lackof induction of antibodies specific for conserved, discontinuous epitopes ofHIV-1 envelope glycoprotein by candidate AIDS vaccines. J. Immunol. 155(8), 4100–4110.
Vogel, T.U., Reynolds, M.R., Fuller, D.H., Vielhuber, K., Shipley, T., Fuller, J.T.,Kunstman, K.J., Sutter, G., Marthas, M.L., Erfle, V., Wolinsky, S.M., Wang,C., Allison, D.B., Rud, E.W., Wilson, N., Montefiori, D., Altman, J.D.,Watkins, D.I., 2003. Multispecific vaccine-induced mucosal cytotoxic Tlymphocytes reduce acute-phase viral replication but fail in long-term controlof simian immunodeficiency virus SIVmac239. J. Virol. 77 (24),13348–13360.
Wei, X., Decker, J.M., Wang, S., Hui, H., Kappes, J.C., Wu, X., Salazar-Gonzalez, J.F., Salazar, M.G., Kilby, J.M., Saag, M.S., Komarova, N.L.,Nowak, M.A., Hahn, B.H., Kwong, P.D., Shaw, G.M., 2003. Antibodyneutralization and escape by HIV-1. Nature 422 (6929), 307–312.
Wrin, T., Nunberg, J.H., 1994. HIV-1MN recombinant gp120 vaccine serum,which fails to neutralize primary isolates of HIV-1, does not antagonizeneutralization by antibodies from infected individuals. Aids 8 (11),1622–1623.
Wyatt, R., Moore, J., Accola, M., Desjardin, E., Robinson, J., Sodroski, J., 1995.Involvement of the V1/V2 variable loop structure in the exposure of humanimmunodeficiency virus type 1 gp120 epitopes induced by receptor binding.J. Virol. 69 (9), 5723–5733.
Wyatt, R., Kwong, P.D., Desjardins, E., Sweet, R.W., Robinson, J.,Hendrickson, W.A., Sodroski, J.G., 1998. The antigenic structure of theHIV gp120 envelope glycoprotein. Nature 393 (6686), 705–711.
Yang, X., Farzan, M., Wyatt, R., Sodroski, J., 2000a. Characterization ofstable, soluble trimers containing complete ectodomains of humanimmunodeficiency virus type 1 envelope glycoproteins. J. Virol. 74 (12),5716–5725.
Yang, X., Florin, L., Farzan, M., Kolchinsky, P., Kwong, P.D., Sodroski, J.,Wyatt, R., 2000b. Modifications that stabilize human immunodeficiencyvirus envelope glycoprotein trimers in solution. J. Virol. 74 (10),4746–4754.
Yang, X., Wyatt, R., Sodroski, J., 2001. Improved elicitation of neutral-izing antibodies against primary human immunodeficiency viruses bysoluble stabilized envelope glycoprotein trimers. J. Virol. 75 (3),1165–1171.
Yang, X., Lee, J., Mahony, E.M., Kwong, P.D., Wyatt, R., Sodroski, J., 2002.Highly stable trimers formed by human immunodeficiency virus type 1envelope glycoproteins fused with the trimeric motif of T4 bacteriophagefibritin. J. Virol. 76 (9), 4634–4642.
Zhang, P.F., Cham, F., Dong, M., Choudhary, A., Bouma, P., Zhang, Z., Shao, Y.,Feng, Y.R., Wang, L., Mathy, N., Voss, G., Broder, C.C., Quinnan Jr., G.V.,

290 I.K. Srivastava et al. / Virology 372 (2008) 273–290
2007. Extensively cross-reactive anti-HIV-1 neutralizing antibodies inducedby gp140 immunization. Proc. Natl. Acad. Sci. U. S. A. 104 (24),10193–10198 (Electronic publication 2007 May 31).
Zolla-Pazner, S., Lubeck, M., Xu, S., Burda, S., Natuk, R.J., Sinangil, F.,Steimer, K., Gallo, R.C., Eichberg, J.W., Matthews, T., Robert-Guroff, M.,1998. Induction of neutralizing antibodies to T-cell line-adapted and primary
human immunodeficiency virus type 1 isolates with a prime-boost vaccineregimen in chimpanzees. J. Virol. 72 (2), 1052–1059.
zur Megede, J., Chen, M.C., Doe, B., Schaefer, M., Greer, C.E., Selby, M., Otten,G.R., Barnett, S.W., 2000. Increased expression and immunogenicity ofsequence-modified human immunodeficiency virus type 1 gag gene. J. Virol.74 (6), 2628–2635.




















