
Hairy Root Induction in Helicteres isora L. and Production of Diosgenin in Hairy Roots
Upload
independentCategory
view
2download
0
ORIGINAL PAPER
Cold storage and cryopreservation of hairy root culturesof medicinal plant Eruca sativa Mill., Astragalusmembranaceus and Gentiana macrophylla Pall.
Sheng-Hui Xue Æ Xin-Juan Luo Æ Zhen-Hua Wu ÆHui-Li Zhang Æ Xin-Yu Wang
Received: 21 May 2007 / Accepted: 5 December 2007 / Published online: 18 December 2007
� Springer Science+Business Media B.V. 2007
Abstract To explore the possibility of an effec-
tively long-term preservation of the germplasm of the
HR lines of medicinal plant Astragalus membranac-
eus, Gentiana macrophylla Pall., and Eruca sativa
Mill., both cold storage and cryopreservation
approaches were attempted and compared. After
5-month cold storage on half strength Murashige
and Skoog (1962) (1/2 MS) agar medium (AM), up to
82.9, 75.7, and 100% of the A. membranaceus,
G. macrophylla and E. sativa hairy roots (HRs)
recovered growth, respectively. The survival rates
of A. membranaceus and G. macrophylla HRs sig-
nificantly decreased, whereas that of E. sativa HR
was unchanged with the addition of increased levels
of exogenous abscisic acid (ABA) during cold
storage. Using the encapsulation–vitrification (EV)
method for cryopreservation, the G. macrophylla
HRs died, whereas up to 6 and 73% of the A. mem-
branaceus and E. sativa HRs survived, respectively.
The HR lines evaluated with both methods showed no
significant differences in morphology and growth rate
compared with controls that were not subjected to
preservation methods. These results suggest that cold
storage is a more suitable alternative for the HR lines
of the three studied plant species and that specificity
of plant species have profound effects on the
effectiveness of preservation.
Keywords Astragalus membranaceus �Cryopreservation � Cold storage �Encapsulation–vitrification � Eruca sativa Mill. �Gentiana macrophylla Pall. � Hairy root
Abbreviations
ABA Abscisic acid
AM Agar medium
DMSO Dimethyl sulphoxide
EV Encapsulation–vitrifcation
ED Encapsulation–dehydration
FW Fresh weight
HRs Hairy roots
LN Liquid nitrogen
LM Liquid medium
LS Loading solution
MS Murashinge and Skoog (1962) medium
PVS Plant vitrification solution
Introduction
Hairy roots (HRs), produced from Agrobacterium
rihizogenes infected plant tissue, have been accepted
as a convenient and promising system for producing
interesting secondary metabolites and proteins.
Because HRs usually originate from a single plant
S.-H. Xue � X.-J. Luo � Z.-H. Wu � H.-L. Zhang �X.-Y. Wang (&)
Institute of Cell Biology, School of Life Sciences,
Lanzhou University, Lanzhou 730000, P.R. China
e-mail: [email protected]
123
Plant Cell Tiss Organ Cult (2008) 92:251–260
DOI 10.1007/s11240-007-9329-x

cell infected by A. rhizogenes, they are considered
genetically stable in contrast to callus cultures.
Moreover, the HRs are characterized by stronger
growing potential, and their secondary metabolite
synthesis is not inhibited during the growth phase of
culture, compared with undifferentiated cells. There-
fore, the HRs usually produce secondary compounds
without the loss of concentration frequently observed
during callus or cell suspension cultures (Bourgaud
et al. 1997). To date, the HR lines of more than 100
plant species have been obtained (Georgiev et al.
2007), and many of these were found to exhibit
higher capacity of synthesizing the desired secondary
metabolite than the untransformed roots, combined
with good genetic and biochemical stability (Yoshik-
awa and Furuya 1987; Parr et al. 1988; Flem-
Bonhomme et al. 2004; Kumar et al. 2006).
Astragalus membranaceus, Gentiana macrophylla
Pall. and Eruca sativa Mill. are widely used as
traditional herbal medicines in China. Radix astragali
from the roots of A. membranaceus is rich in
calycosin, polysaccharides, isoflavonoids, c-amino-
butyric acid and various trace elements (Wagner
et al. 1997). Various medicinal properties have been
ascribed to compounds from R. astragali, which may
be utilized as immunostimulants, tonics, diuretics,
antidiabetics, analgesics, expectorants, and sedatives
(Sinclair 1998). The roots of G. macrophylla have
been successfully used to cure viral-induced, neuro-
pathic, respiratory, and cardiovascular diseases
(Zhang et al. 2003) due to the presence of gentiopi-
croside. Gentiopicroside has been found to play
roles in abirritation, defervescence, reducing blood
pressure, antibacterium, antiinflammation, and anti-
rheumatism (Zhang et al. 2003; Yu et al. 2004).
E. sativa contains glucosinolates. The hydrolysates of
glucosinolates, isothiocyanates, have been reported to
inhibit mitosis (Iori et al. 1999) and stimulate apop-
tosis in human tumor cells (Johnson 2002).
In order to explore the possibility of producing
interesting metabolites using HR cultures, we
recently obtained several HR lines from the above
three plant species by infecting their cotyledon or leaf
segments with A. rhizogenes R1000. The HR lines
grew well and showed the traits of the typical HRs.
Polymerase chain reaction (PCR) has demonstrated
that their genomes have been integrated with the rolB
or rolC genes of Ri T-DNA. Moreover, high perfor-
mance liquid chromatography (HPLC) has confirmed
that some of these lines can synthesize gentiopicro-
side or glucosinolates. However, the continuous
subculture is very time-consuming and expensive,
and permits the loss of stability of phenotype and
genotype of plant germplasm. Thus, it is necessary to
develop an effective method for long-term preserva-
tion of the useful HR lines.
In the past decades, cold storage and cryopreser-
vation were widely-explored as better alternatives
for germplasm preservation. Many approaches, e.g.,
slow cooling, vitrification, encapsulation–dehydration
(ED), and encapsulation–vitrification (EV) method,
were developed, and used with varying degrees of
success to preserve plants. The organs/tissues that
were successfully cold stored or cryopreserved mainly
included shoot tips (Pennycooke et al. 2000; Lam-
bardi et al. 2000; Xu et al. 2002), callus (An et al.
2003) and somatic embryo (Ford et al. 2000; Shiota
et al. 1999). However, only a few studies have
evaluated preservation for HRs (Benson and Hamill
1991; Teoh 1996; Yoshimatsu et al. 1996; Phunchin-
dawn et al. 1997; Hirata et al. 2002), and results
varied by plant species and methods used. The
purpose of the present work was to develop a protocol
for effective, long-term preservation of our HR lines
of A. membranaceus, G. macrophylla, and E. sativa.
Materials and methods
Plant material and culture condition
The HR cultures of Astragalus membranaceus,
Gentiana macrophylla Pall. and Eruca sativa Mill.
were established by transforming their cotyledon or
leaf explants with A. rhizogenes R1000, following
which the cultures were stored for *2 years via
regular subculture on hormone free 1/2 MS agar
medium (AM) with 0.088 M sucrose at 25�C in the
dark in 3 week intervals. The HR lines grew well and
had no visible morphological abnormalities.
Cold storage of the HR lines of A. membranaceus,
G. macrophylla, and E. sativa
Fresh HR tips *1-cm long were excised from the HR
cultures grown for 10 days on hormone free 1/2 MS
AM with 0.088 M sucrose, and transferred onto the
252 Plant Cell Tiss Organ Cult (2008) 92:251–260
123

same medium supplied with 0, 0.5 mg l-1 or,
1.0 mg l-1 ABA, respectively. After storage for
5 months at 4�C in the dark, they were retransferred
onto fresh hormone free 1/2 MS AM and cultured at
25�C in the dark. Two weeks later, their survival rates
(%), expressed as the ratio of survival HRs to total
cold-stored HRs, were evaluated. The HR cultures
stored for 5 months on the same medium at 25�C
were used as a control.
Cryopreservation of the HR lines
of A. membranaceus, G. macrophylla,
and E. sativa
The encapsulation–vitrification (EV) method (Hirai
and Sakai 1999) was employed, with minor modifi-
cations. Briefly, the HR tips excised from the HR
cultures grown for 10 days on hormone free 1/2 MS
AM were suspended in 2% (w/v) sodium alginate
solution supplied with 0.1, 0.3, or 0.5 M sucrose.
Then they were dropped in 50 mM CaCl2 solution
with 0.1, 0.3, or 0.5 M sucrose to be encapsulated
with a pipette. Afterwards, (i) the encapsulated HRs
were pre-cultured for 1 and 3 days at 25�C in 1/2 MS
liquid medium (LM) containing an identical level of
sucrose as that used during encapsulation, and then
vitrified for 60 min at 25�C with plant vitrification
solution 3 (PVS3) (Nishizawa et al. 1993), 50% (w/v)
glycerol and 50% (w/v) sucrose, with a change after
30 min (Protocol 1); (ii) the encapsulated HRs were
pre-cultured for 3 days in 1/2 MS LM with 0.3 M
sucrose at 25�C, treated for 20 min at 25�C in MS
LM containing 2 M glycol and 0.4 M sucrose
(loading solution, LS), and then vitrified for 60 min
at 25�C in PVS3 with a change after 30 min (Protocol
2); or (iii) the encapsulated HRs were pre-cultured for
3 days at 25�C in 1/2 MS LM with 0.3 M sucrose, LS
treated for 20 min at 25�C, and then vitrified for
0–120 min in PVS 3 or PVS2 (Protocol 3). PVS2
consisted of 30% glycerol (w/v), 15% (w/v) ethylene
glycol, 15% (w/v) dimethyl sulfoxide and 0.4 M
sucrose in MS medium (pH 5.8) (Sakai et al. 1990).
After vitrified, the encapsulated HRs were trans-
ferred into 2-ml-cryotubes and stored in liquid
nitrogen (LN). Three days later, they were thawed
in water bath at 40�C for 2 min, and then incubated
for 30 min in MS LM with 1.2 M sucrose in a 50-ml
conical flask 25�C with a change every 10 min.
Afterwards, they were transferred onto 1/2 MS AM
with 0.088 M sucrose and cultured at 25�C in the
dark. Two weeks later, their survival rates were
evaluated following the method above.
Analysis of growing potential of the HR cultures
after cold or cryopreservation
For appraising the effects of cold storage and
cryopreservation on the growing potential of the
HR lines, we inoculated the HR lines of the above
three plant species, which were subjected to cold
storage or cryopreservation and recovered growth,
onto 1/2 MS AM. After cultured at 25�C for 15 days,
their fresh weight (FW) was measured. Each bottle
was initially inoculated with five tips 1-cm long,
weighing *6 mg (E. sativa), 7 mg (G. macrophy-
lla), and 10 mg (A. membranaceus), respectively.
The growing potentials of the HR lines were
indicated by the increment of the HR lines, being
defined as the ratios of the final fresh weight (FW) to
the initial FW. The HR lines not subjected to cold
storage and cryopreservation were taken as control.
In addition, we inoculated the HR lines of
E. sativa and G. macrophylla, which were subjected
to cold storage or cryopreservation and recovered
growth, into 30 ml of 1/2 MS LM in 150-ml conical
flasks. Then they were shake-cultured at 25�C at
100 rpm and weighed every 5 days. Finally, their
growth curves were drawn. Each bottle was initially
inoculated with 30 HRs 3-cm long with branches,
weighing totally about 180 mg (E. sativa) and
310 mg (G. macrophylla), respectively.
All the media and solutions used were autoclaved
at 121�C for 20 min and all manipulations were
performed under strictly sterile conditions.
Statistical analysis
All the experiments above were repeated in triplicate,
each containing a minimum of three samples. The
final survival rates and increment of the HR lines
were represented by the means with standard devi-
ation (SD) of the three experiments. The differences
between various treatments were determined by
Duncan’s Multiple Range Test (P B 0.05) using
SPSS 10.0 software. The growth curves were drawn
with Excel software (Microsoft, Redmond, WA).
Plant Cell Tiss Organ Cult (2008) 92:251–260 253
123
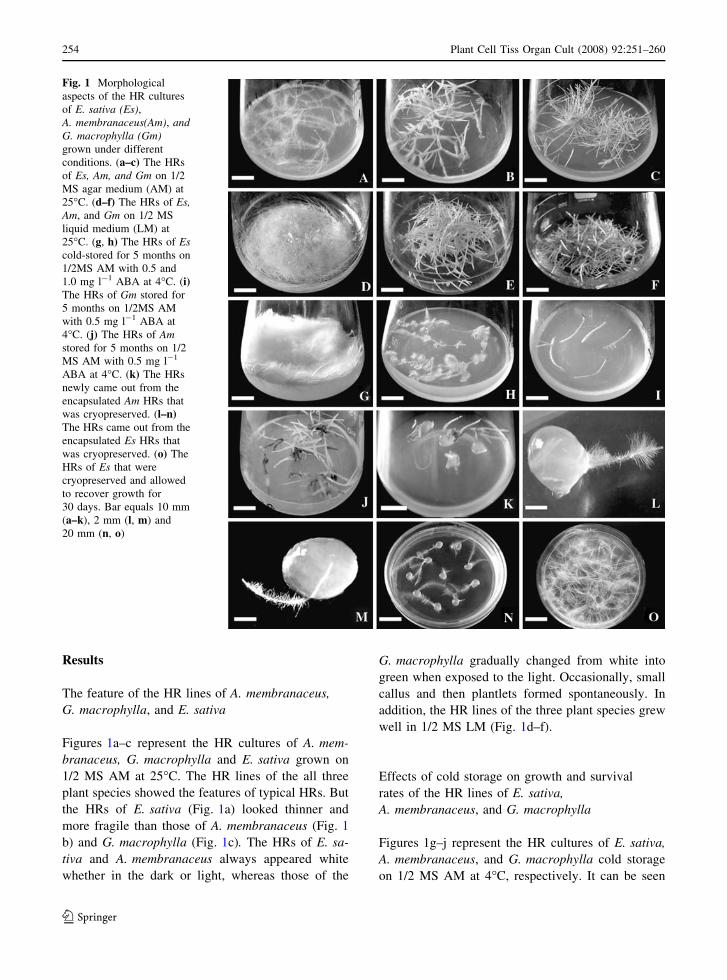
Results
The feature of the HR lines of A. membranaceus,
G. macrophylla, and E. sativa
Figures 1a–c represent the HR cultures of A. mem-
branaceus, G. macrophylla and E. sativa grown on
1/2 MS AM at 25�C. The HR lines of the all three
plant species showed the features of typical HRs. But
the HRs of E. sativa (Fig. 1a) looked thinner and
more fragile than those of A. membranaceus (Fig. 1
b) and G. macrophylla (Fig. 1c). The HRs of E. sa-
tiva and A. membranaceus always appeared white
whether in the dark or light, whereas those of the
G. macrophylla gradually changed from white into
green when exposed to the light. Occasionally, small
callus and then plantlets formed spontaneously. In
addition, the HR lines of the three plant species grew
well in 1/2 MS LM (Fig. 1d–f).
Effects of cold storage on growth and survival
rates of the HR lines of E. sativa,
A. membranaceus, and G. macrophylla
Figures 1g–j represent the HR cultures of E. sativa,
A. membranaceus, and G. macrophylla cold storage
on 1/2 MS AM at 4�C, respectively. It can be seen
Fig. 1 Morphological
aspects of the HR cultures
of E. sativa (Es),A. membranaceus(Am), andG. macrophylla (Gm)grown under different
conditions. (a–c) The HRs
of Es, Am, and Gm on 1/2
MS agar medium (AM) at
25�C. (d–f) The HRs of Es,Am, and Gm on 1/2 MS
liquid medium (LM) at
25�C. (g, h) The HRs of Escold-stored for 5 months on
1/2MS AM with 0.5 and
1.0 mg l-1 ABA at 4�C. (i)The HRs of Gm stored for
5 months on 1/2MS AM
with 0.5 mg l-1 ABA at
4�C. (j) The HRs of Amstored for 5 months on 1/2
MS AM with 0.5 mg l-1
ABA at 4�C. (k) The HRs
newly came out from the
encapsulated Am HRs that
was cryopreserved. (l–n)The HRs came out from the
encapsulated Es HRs that
was cryopreserved. (o) The
HRs of Es that were
cryopreserved and allowed
to recover growth for
30 days. Bar equals 10 mm
(a–k), 2 mm (l, m) and
20 mm (n, o)
254 Plant Cell Tiss Organ Cult (2008) 92:251–260
123

that very limited growth was still maintained with
the HR lines of all the three plant species. The HRs
of E. sativa (Fig. 1g, h) appeared thin but dense, with
a white coloring, while those of A. membranaceus
(Fig. 1j) and G. macrophylla (Fig. 1i) appeared
strong rare and had white or beige coloring. In
addition, it was found that the HR cultures stored on
the medium without ABA universally grew better
than those on the medium with ABA and that the HR
system of E. sativa grew better than those of other
plants on the medium. These results were also
supported by the quantitative analysis of their fresh
weigh (FW) (Fig. 2).
Table 1 shows the survival rates of the HR lines of
the studied three plant species after 5 months in cold
storage. On the medium either with or without ABA,
all HRs of E. sativa recovered and showed high
survival (100%). However, the survival rates of the
HR lines of A. membranaceus and G. macrophylla
were only 82.9% and 75.7% at maximums, respec-
tively, being significantly less than that of E. sativa.
In addition, the survival rates of the HR lines of both
A. membranaceus and G. macrophylla significantly
decreased with increased ABA concentration. Also it
was found that the control HR lines of E. sativa and
A. membranaceus stored for 5 months at 25�C on
medium with or without ABA, all died. However, for
the HRs of G. macrophylla, the HRs grown on
medium with ABA all died, but the HRs on medium
without ABA exhibited a 100% survival rate.
Effects of sucrose concentrations and times
during pre-culture on cryopreservation of the HR
lines of E. sativa, A. membranaceus,
and G. macrophylla
After being pre-cultured in 1/2 MS LM with different
concentrations of sucrose at 25�C for 1 or 3 days
followed by PVS3-vitrification and LN storage (Pro-
tocol 1), all HR cultures of E. sativa recovered more or
less (from 6.6% to 67.8%) under any of the conditions
given in Table 2. The effect of Condition 5, 3-day pre-
culture in medium with 0.3 M sucrose, was optimal,
with a survival rate up to 67.8%. But for the HRs of
A. membranaceus, only those subjected to 3-day pre-
culture in medium with 0.3 M sucrose (Condition 5)
were able to survive, and had a very low survival rate
(about 6%). The HR line from G. macrophyll did not
survive under any of the given conditions in Table 2.
Effects of pre-culture and LS treatment
on cryopreservation of the HR lines of E. sativa,
A. membranaceus, and G. macrophylla
When pre-cultured for 3 days in 1/2 MS LM with
0.3 M sucrose, LS-treated for 20 min, PVS3-vitrified
for 60 min at 25�C, and then stored in LN (Protocol
2), 72.7% of the HR cultures of E. sativa survived
Fig. 2 The increment of fresh weight (FW) of the HR lines of
E. sativa, A. membranaceus, and G. macrophylla during cold
storage on MS agar medium with different concentrations of
ABA at 4�C. The increments are indicated as the final–initial
FW (mg)
Table 1 Effects of ABA concentrations and temperatures
during storage on survival rates of the HR lines of E. sativa,A. membranaceus and G. macrophylla
HR cultures ABA (mg l-1) Survival rates (% ± SD)*
4�C 25�C
E. sativa 0 100f 0a
0.5 100f 0a
1.0 100f 0a
G. macrophyll 0 75.7 ± 6.1e 100f
0.5 63.4 ± 4.7d 0a
1.0 18.4 ± 2.3b 0a
A. membranaceus 0 82.9 ± 4.0e 0a
0.5 46.5 ± 5.0c 0a
1.0 41.4 ± 2.0c 0a
* All the HRs was stored for 5 months at either 4 or 25�C.
Values represent the means with standard deviation (SD) of
three independent experiments. Each experiment consisted of
three repeats, each repeat contained 10 HRs at least. The
different letters following the values means a significant
difference between each two values (P B 0.05) according to
Duncan’s Multiple Range Test
Plant Cell Tiss Organ Cult (2008) 92:251–260 255
123
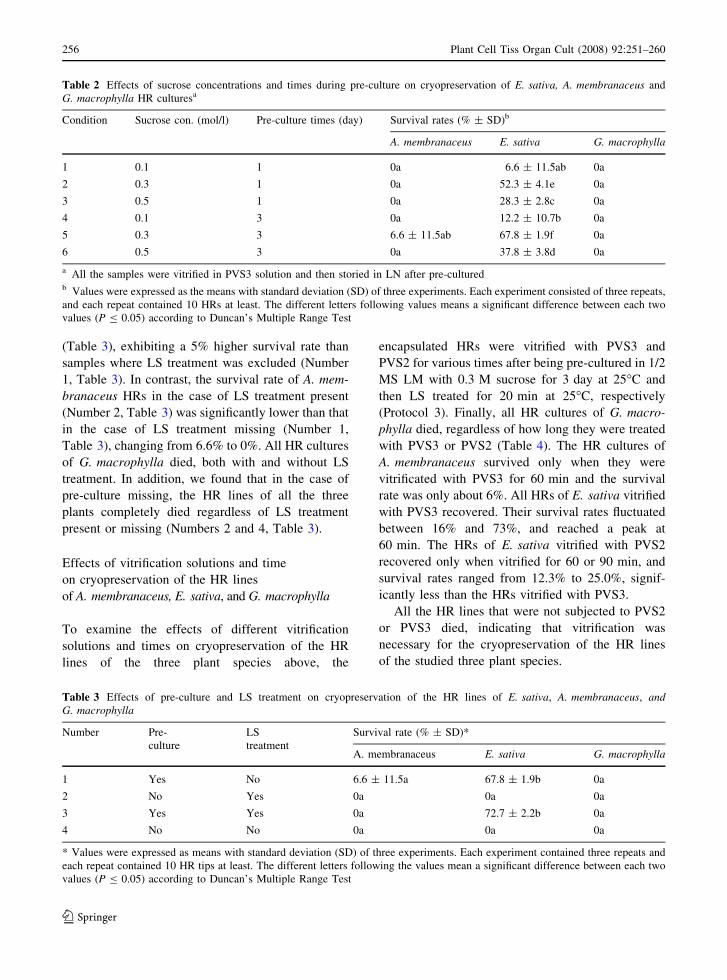
(Table 3), exhibiting a 5% higher survival rate than
samples where LS treatment was excluded (Number
1, Table 3). In contrast, the survival rate of A. mem-
branaceus HRs in the case of LS treatment present
(Number 2, Table 3) was significantly lower than that
in the case of LS treatment missing (Number 1,
Table 3), changing from 6.6% to 0%. All HR cultures
of G. macrophylla died, both with and without LS
treatment. In addition, we found that in the case of
pre-culture missing, the HR lines of all the three
plants completely died regardless of LS treatment
present or missing (Numbers 2 and 4, Table 3).
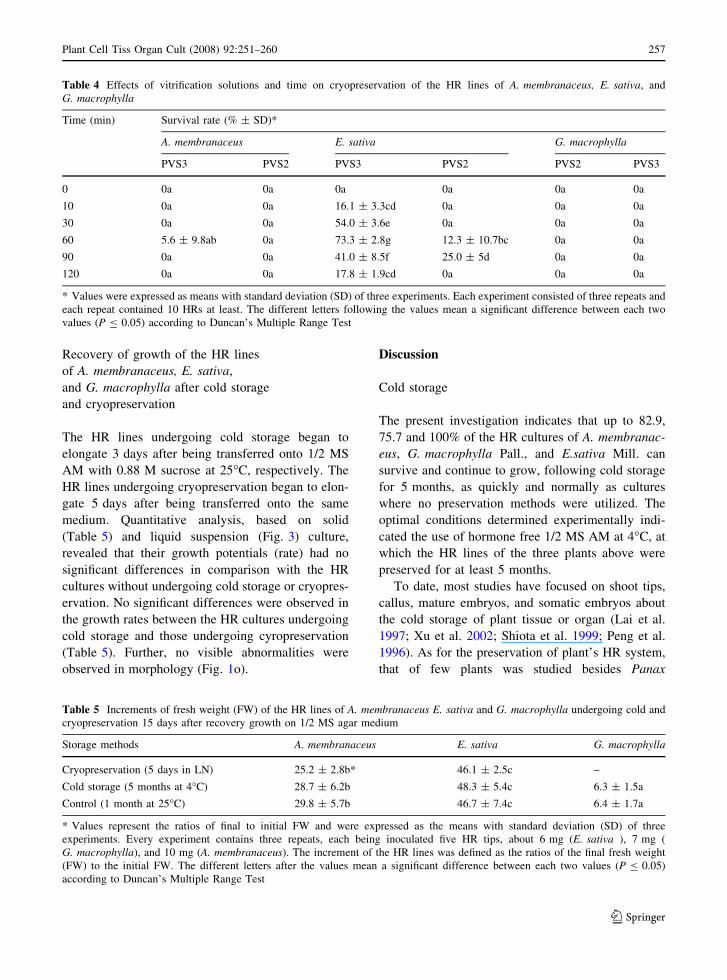
Effects of vitrification solutions and time
on cryopreservation of the HR lines
of A. membranaceus, E. sativa, and G. macrophylla
To examine the effects of different vitrification
solutions and times on cryopreservation of the HR
lines of the three plant species above, the
encapsulated HRs were vitrified with PVS3 and
PVS2 for various times after being pre-cultured in 1/2
MS LM with 0.3 M sucrose for 3 day at 25�C and
then LS treated for 20 min at 25�C, respectively
(Protocol 3). Finally, all HR cultures of G. macro-
phylla died, regardless of how long they were treated
with PVS3 or PVS2 (Table 4). The HR cultures of
A. membranaceus survived only when they were
vitrificated with PVS3 for 60 min and the survival
rate was only about 6%. All HRs of E. sativa vitrified
with PVS3 recovered. Their survival rates fluctuated
between 16% and 73%, and reached a peak at
60 min. The HRs of E. sativa vitrified with PVS2
recovered only when vitrified for 60 or 90 min, and
survival rates ranged from 12.3% to 25.0%, signif-
icantly less than the HRs vitrified with PVS3.
All the HR lines that were not subjected to PVS2
or PVS3 died, indicating that vitrification was
necessary for the cryopreservation of the HR lines
of the studied three plant species.
Table 2 Effects of sucrose concentrations and times during pre-culture on cryopreservation of E. sativa, A. membranaceus and
G. macrophylla HR culturesa
Condition Sucrose con. (mol/l) Pre-culture times (day) Survival rates (% ± SD)b
A. membranaceus E. sativa G. macrophylla
1 0.1 1 0a 6.6 ± 11.5ab 0a
2 0.3 1 0a 52.3 ± 4.1e 0a
3 0.5 1 0a 28.3 ± 2.8c 0a
4 0.1 3 0a 12.2 ± 10.7b 0a
5 0.3 3 6.6 ± 11.5ab 67.8 ± 1.9f 0a
6 0.5 3 0a 37.8 ± 3.8d 0a
a All the samples were vitrified in PVS3 solution and then storied in LN after pre-culturedb Values were expressed as the means with standard deviation (SD) of three experiments. Each experiment consisted of three repeats,
and each repeat contained 10 HRs at least. The different letters following values means a significant difference between each two
values (P B 0.05) according to Duncan’s Multiple Range Test
Table 3 Effects of pre-culture and LS treatment on cryopreservation of the HR lines of E. sativa, A. membranaceus, andG. macrophylla
Number Pre-
culture
LS
treatment
Survival rate (% ± SD)*
A. membranaceus E. sativa G. macrophylla
1 Yes No 6.6 ± 11.5a 67.8 ± 1.9b 0a
2 No Yes 0a 0a 0a
3 Yes Yes 0a 72.7 ± 2.2b 0a
4 No No 0a 0a 0a
* Values were expressed as means with standard deviation (SD) of three experiments. Each experiment contained three repeats and
each repeat contained 10 HR tips at least. The different letters following the values mean a significant difference between each two
values (P B 0.05) according to Duncan’s Multiple Range Test
256 Plant Cell Tiss Organ Cult (2008) 92:251–260
123
Recovery of growth of the HR lines
of A. membranaceus, E. sativa,
and G. macrophylla after cold storage
and cryopreservation
The HR lines undergoing cold storage began to
elongate 3 days after being transferred onto 1/2 MS
AM with 0.88 M sucrose at 25�C, respectively. The
HR lines undergoing cryopreservation began to elon-
gate 5 days after being transferred onto the same
medium. Quantitative analysis, based on solid
(Table 5) and liquid suspension (Fig. 3) culture,
revealed that their growth potentials (rate) had no
significant differences in comparison with the HR
cultures without undergoing cold storage or cryopres-
ervation. No significant differences were observed in
the growth rates between the HR cultures undergoing
cold storage and those undergoing cyropreservation
(Table 5). Further, no visible abnormalities were
observed in morphology (Fig. 1o).
Discussion
Cold storage
The present investigation indicates that up to 82.9,
75.7 and 100% of the HR cultures of A. membranac-
eus, G. macrophylla Pall., and E.sativa Mill. can
survive and continue to grow, following cold storage
for 5 months, as quickly and normally as cultures
where no preservation methods were utilized. The
optimal conditions determined experimentally indi-
cated the use of hormone free 1/2 MS AM at 4�C, at
which the HR lines of the three plants above were
preserved for at least 5 months.
To date, most studies have focused on shoot tips,
callus, mature embryos, and somatic embryos about
the cold storage of plant tissue or organ (Lai et al.
1997; Xu et al. 2002; Shiota et al. 1999; Peng et al.
1996). As for the preservation of plant’s HR system,
that of few plants was studied besides Panax
Table 4 Effects of vitrification solutions and time on cryopreservation of the HR lines of A. membranaceus, E. sativa, and
G. macrophylla
Time (min) Survival rate (% ± SD)*
A. membranaceus E. sativa G. macrophylla
PVS3 PVS2 PVS3 PVS2 PVS2 PVS3
0 0a 0a 0a 0a 0a 0a
10 0a 0a 16.1 ± 3.3cd 0a 0a 0a
30 0a 0a 54.0 ± 3.6e 0a 0a 0a
60 5.6 ± 9.8ab 0a 73.3 ± 2.8g 12.3 ± 10.7bc 0a 0a
90 0a 0a 41.0 ± 8.5f 25.0 ± 5d 0a 0a
120 0a 0a 17.8 ± 1.9cd 0a 0a 0a
* Values were expressed as means with standard deviation (SD) of three experiments. Each experiment consisted of three repeats and
each repeat contained 10 HRs at least. The different letters following the values mean a significant difference between each two
values (P B 0.05) according to Duncan’s Multiple Range Test
Table 5 Increments of fresh weight (FW) of the HR lines of A. membranaceus E. sativa and G. macrophylla undergoing cold and
cryopreservation 15 days after recovery growth on 1/2 MS agar medium
Storage methods A. membranaceus E. sativa G. macrophylla
Cryopreservation (5 days in LN) 25.2 ± 2.8b* 46.1 ± 2.5c –
Cold storage (5 months at 4�C) 28.7 ± 6.2b 48.3 ± 5.4c 6.3 ± 1.5a
Control (1 month at 25�C) 29.8 ± 5.7b 46.7 ± 7.4c 6.4 ± 1.7a
* Values represent the ratios of final to initial FW and were expressed as the means with standard deviation (SD) of three
experiments. Every experiment contains three repeats, each being inoculated five HR tips, about 6 mg (E. sativa ), 7 mg (
G. macrophylla), and 10 mg (A. membranaceus). The increment of the HR lines was defined as the ratios of the final fresh weight
(FW) to the initial FW. The different letters after the values mean a significant difference between each two values (P B 0.05)
according to Duncan’s Multiple Range Test
Plant Cell Tiss Organ Cult (2008) 92:251–260 257
123

(Yoshimatsu et al. 1996). The HR line of Panax was
reported to have survived for 4 months on hormone
free MS AM at 4�C, with 100% survival rate. It can
be seen the survival rate of E. sativa HRs is identical
to that of Panax HRs. The survival rates of the HR
lines of A. membranaceus and G. macrophylla,
82.9% and 75.7%, are not as high as that of E. sativa
and Panax, but are not excessively low. So it is
feasible to preserve the HR lines of the three plants
above using our present protocol.
Effect of ABA on cold storage
of A. membranaceus, G. macrophylla,
and E. sativa HRs
ABA is a hormone regulating plant seed germination
and development that responds to various environ-
ment stresses (Finkelstein et al. 2002). Previous work
has demonstrated that ABA enhances plant tolerance
to low temperature, drought, wounds, or pathogens,
when exogenously applied. The shoot of garlic was
successfully preserved on B5 medium with 0.3 mg l-
1 6-BA, 0.1 mg l-1 NAA and 10 mg l-1 ABA at 0�C
for a year (Xu et al. 2002). The carrot somatic
embryos undergoing 10-5 M ABA-treatment and
then desiccation were found to be able to survive
for 169 weeks at -25�C and at least 24 weeks at 5�C,
respectively (Shiota et al. 1999). Neither medium
with or without exogenously added ABA had any
effect on the survival rate of the HR lines of E. sativa
in the present work. For the HR lines of both
A. membranaceus and G. macrophylla, adding ABA
significantly reduced survival rates. This result is
different from the above author’s results, but not
inconsistent with other literature. For example, the
mature embryos of Citrus microcarpa were found to
survive on 1/2 MS medium with 5 mg l-1 chloroch-
oline chloride (CCC) at 25�C for a year without
subculture, with 100% survival rate. However, when
15 mg l-1 ABA was added, its survival rate dropped
sharply to 51.2% (Lai et al. 1997). These differences
can be explained by the disagreement of the plant
organ/tissues and/or media used by different authors.
It is possible that the effect of ABA differs with the
specificity of plant species, tissue/organ and/or
medium composition.
Cryopreservation
The approaches used to cryopreserve plant tissues
and organs mainly include slow cooling (SC),
vitrification (V), encapsulation–dehydration (ED),
and encapsulation–vitrification (EV) method, with
the latter two methods appearing to produce more
effective results. The EV method was employed in
the present work. The core of EV method is to first
encapsulate the plant tissue/organ and later vitrificate
with PVS before LN-storage. To improve the survival
rate of the HR lines, we added auxiliary treatments,
mainly including the pre-culture for different periods
of days in MS LM with sucrose of different
concentration, LS treatment and vitrification at dif-
ferent lengths of time (in min) with different PVS. It
was indicated that pre-culture was essential for the
preservation of E. sativa and A. membranaceus HRs,
but completely ineffective on that of G. macrophylla
HRs. LS treatment was valid for the preservation the
E. sativa HRs, whereas completely ineffective on the
HRs of other two plants.
Fig. 3 The growth curves of the HR lines of E. sativa (a) and
G. macrophylla (b) with and without undergoing cold storage
and cryopreservation in 1/2 MS liquid medium at 25�C. The
values were expressed as the fresh weight (mg) on different
days after culture. Each bottle was inoculated with 30 HRs 3-
cm long with branches, 180 mg (E. sativa ) and 310 mg
(G. macrophylla), respectively. Since none of G. macrophyllaHRs survived following cryopreservation under any given
condition, there is no growth curve of G. macrophyllaundergoing cryopreservation
258 Plant Cell Tiss Organ Cult (2008) 92:251–260
123

PVS2 is a vitrification solution most widely used in
cryopreservation of plant organs/tissues. In the pres-
ent experiment, however, it was found completely
ineffective on the preservation of G. macrophylla and
A. membranaceu HRs, but had a small positive effect
on the storage of E. sativa HRs. PVS3 was completely
ineffective on the preservation of G. macrophylla
HRs, slightly effective for A. membranaceus HRs and
significantly valid for E. sativa HRs (Table 4). The
optimal protocol for E. sativa HRs was 3-day pre-
culture in 1/2MS LM with 0.3 M sucrose at 25�C ?LS treatment for 20 min at 25�C ? PVS3 vitrification
for 60 min at 25�C, with 73% survival rate. The
optimal protocol for A. membranaceus HRs was 3-
day pre-culture in 1/2 MS LM with 0.3 M sucrose at
25�C ? PVS3 vitrification for 60 min at 25�C, with
6% survival rate. The HRs of G. macrophylla, did not
recover under any of the given conditions.
To date, the HRs of only Beta vulgaris, Nicotiana
rustica (Benson and Hamill 1991), Artemisia annua
(Teoh et al. 1996), Panax (Yoshimatsu et al. 1996),
Armoracia rusticana (Phunchindawan et al. 1997)
and Vinca minor (Hirata et al. 2002) have been
successfully cryopreserved, to our knowledge. It was
reported that 60% of Armoracia rusticana HRs was
recovered following cryopreservation with EV
method (Phunchindawan et al. 1997). This value is
smaller than in E. sativa HRs.
The relationship between plant HR’s preservation
and species
In the present work, the survival rate of the HRs of
E. sativa following cold-storage or cryopreservation
was always found to be significantly higher than that
of G. macrophylla and A. membranaceus HRs. The
HRs of G. macrophylla always exhibited lower
survival rates than that of the other two plants and
exhibited a 0% survival rate after cryopreservation.
Also, the response of the HR lines of the three plants
to LS treatment and PVS solution were different from
each other during the cryopreservation of the three
plants studied, E. sativa and A. membranaceus were
reported to be highly-resistant to drought (Tewari
et al. 1995; Sun et al. 2005; Guo et al. 1987), and
E. sativa. G. macrophylla is completely nonresistant
to drought. The plants exhibiting high drought-
resistance generally are capable of stronger osmotic
regulation. This suggests questions as to whether the
relatively high survival rates of E. sativa and
A. membranaceus HRs, following cryopreservation,
are linked with their stronger drought resistance.
In conclusion, the effect of cold and cryopreser-
vation of A. membranaceus, G. macrophylla, and
E. sativa HR lines were compared in the present
study. The HRs of the three plant species survived
after cold storage, with survival rate up to 82.9, 75.7
and 100%, respectively. But with EV cryopreserva-
tion method, only the E. sativa and A. membranaceus
HR lines remained alive, with 73 and 6% survive
rate. The HR lines of the three plant species with cold
storage or cryopreservation grew comparably to that
of controls without preservation in both solid and
liquid medium. These results suggest cold storage is a
better alternative for the long-term preservation of the
HR lines of the three plant species. In addition, cold
storage is simple, convenient, time-saving and low-
cost compared with EV cryopreservation. However, it
remains to be determined whether this approach
preserves stable germplasms of the HR lines with
respect to phenotype and molecular biology.
Acknowledgments We would like to show our thanks to
Ph.D. Michael Kirberger in the Department of Chemistry and
Max Oginsky in the Department of Biology, Georgia State
University for spending precious time to revise our paper.
References
An CS, Wang XD, Yuan XF, Zhao B, Wang YC (2003)
Optimization of cryopreservation of Artemisia annua L.
callus. Biotechnol Lett 25:35–38
Benson EE, Hamill JD (1991) Cryopreservation and post
freezing molecular and biosynthetic stability in trans-
formed roots of Beta vulgaris and Nicotiana rustica. Plant
Cell Tiss Organ Cult 24:163–172
Bourgaud F, Bouque V, Gontier E, Guckert A (1997) Hairy
root cultures for the production of secondary metabolites.
AgBiotech News Inf 9:205–208
Finkelstein RR, Gampala S, Rock CD (2002) Abscisic acid
signaling in seeds and seedlings. The Plant Cell Suppl
S15–S45
Flem-Bonhomme Le V, Laurain-Mattar D, Fliniaux MA
(2004) Hairy root induction of Papaver somniferum var.
album, a difficult-to-transform plant, by A. rhizogenesLBA 9402. Planta 218:890–893
Ford CS, Jones NB, Van Staden J (2000) Cryopreservation and
plant regeneration from somatic embryos of Pinus patula.
Plant Cell Rep 19:610–615
Georgiev MI, Pavlov I, Bley T (2007) Hairy root type plant
in vitro systems as sources of bioactive substances. Appl
Microbiol Biot 74:1175–1185
Plant Cell Tiss Organ Cult (2008) 92:251–260 259
123

Guo BZ, Zhang HZ, Pan JT, Yang YC, Wu ZL, He TN, Zhou
LH, Huang RF (1987) Flora of Qinghai economic plants,
Section I. Qinghai People’s Press, Xining, China, pp 312–
314
Hirai D, Sakai A (1999) Cryopreservation of in vitro-grown
meristems of potato (Solanum tuberosum L.) by encap-
sulation–vitrification. Potato Res 42:153–160
Hirata K, Mukai M, Goda S, Ishio-kinugasa M, Yoshida K,
Sakai A, Miyamoto K (2002) Cryopreservation of hairy
root cultures of Vinca minor (L.) by encapsulation–
dehydration. Biotechnol Lett 24:371–376
Iori R, Bernadi R, Gueyrard D, Rollin P, Palmieri S (1999)
Formation of glucoraphanin by chemoselective oxidation
of natural glucoerucin: a chemoenzymatic route to sulfo-
raphane. Bioorg Med Chem Lett 9:1047–1048
Johnson IT (2002) Glucosinolates in the human diet: bio-
availability and implications for health. Phytochem Rev
1:183–188
Kumar V, Satyanarayana KV, Sarala IS, Indu EP, Giridhar P,
Chandrashekar A, Ravishankar GA (2006) Stable trans-
formation and direct regeneration in Coffea canephoraPex. Fr. by Agrobacterium rhizogenes mediated transfor-
mation without hairy-root phenotype. Plant Cell Rep
25:214–222
Lai ZX, Chen ZG, He BZ, Lin SQ (1997) In vitro germplasm
preservation of Citrus microcarpa Bge. embryo. Crop
Variety Resour 4:44–46
Lambardi M, Fabbri A, Caccavale A (2000) Cryopreservation
of white poplar (Populus alba L.) by vitrification of in
vitro-grown shoot tips. Plant Cell Rep 19:213–218
Murashige T, Skoog F (1962) A revised medium for rapid
growth and bioassays with tobacco tissue cultures. Plant
Physiol 15:473–497
Nishizawa S, Sakai A, Amano Y, Matsuzawa T (1993) Cryo-
preservation of asparagus (Asparagus officinalis L.)
embryonic suspension cells and subsequent plant regen-
eration by vitrification. Plant Sci 91:67–73
Parr AJ, Peerless AC, Hamill JD, Walton NJ, Robins RJ,
Rhodes MJC (1988) Alkaloid production by transformed
root cultures of Catharanthus roseus. Plant Cell Rep
7:309–312
Peng MZ, Deng JS, Gan L (1996) Preservation of Avocadoin vitro under low temperature. J Fruit Sci 13:96–98
Pennycooke JC, Towill LE (2000) Cryopreservation of shoot
tips from in vitro plants of sweet potato [Ipomoea batatas(L.) Lam.] by vitrification. Plant Cell Rep 19:733–737
Phunchindawan M, Hirata K, Sakai A, Miyamoto K (1997)
Cryopreservation of encapsulated shoot primordia
induced in horseradish (Armoracia rusticana) hairy root
cultures. Plant Cell Rep 16:469–473
Sakai A, Kobayashi S, Oiyama I (1990) Cryopreservation of
nucellar cells of Naval orange (Colocasia esculenta (L.)
Schott) by vitrification. Plant Cell Rep 9:30–33
Shiota H, Tachibana K, Watabe K, Kamada H (1999) Suc-
cessful long-term preservation of abscisic-acid-treated and
desiccated carrot somatic embryos. Plant Cell Rep
18:749–753
Sinclair S (1998) Chinese herbs: a clinical review of Astrag-alus, Ligusticum, and Schizandrae. Altern Med Rev
3:338–344
Sun WC, Guan CY, Meng YX, Liu ZG, Zhang T, Li X , Yang
SZ, Ling LJ, Chen SY, Zeng XC, Wang HL (2005)
Intergeneric crosses between Eruca sativa Mill. and
Brassica Species. Acta Agronomica Sinica 31:36–42
Teoh KH, Weathers PJ, Cheetham RD, Walcerz DB (1996)
Cryopreservation of transformed (hairy) roots of Artemi-sia annua. Cryobiology 33:106–117
Tewari JP, Bansal VK, Stringam GR, Thiagarajah MR (1995)
Reaction of some wild and cultivated Eruca accessions
against Leptosphaeria maculans. Can J Plant Pathol
17:362–363
Wagner H, Bauer R, Xiao PG, Chen JM, Michler G (1997)
Radix astragali (Huang Qi). Chinese Drug Monographs
Anal 1:1–17
Xu PW, Qu SS, Liu HY, Zhang J, Sun JB, Huang BY (2002) A
preliminary study on in vitro conservation of the garlic
germplasm resources in China. Scientia Agricultura Si-
nica 35(3):314–319
Yoshikawa T, Furuya T (1987) Saponin production by cultures
of Panax ginseng transformed with Agrobacterium rhiz-ogenes. Plant Cell Rep 6:449–453
Yoshimatsu K, Yamaguchi H, Shimomura K (1996) Traits of
Panax ginseng hairy root after cold storage and cryo-
preservation. Plant Cell Rep 15:555–560
Yu FR, Li RD, Wang R (2004) Inhibitory effects of the Gen-tiana macrophylla (Gentianaceae) extract on rheumatoid
arthritis of rats. J Ethnopharmacol 95:77–81
Zhang XL, Jin L, Liu LS (2003) Progress on research of
Gentiana macrophylla Pall. in the resent decade. J Tradit
Chin Med 9:62–63
260 Plant Cell Tiss Organ Cult (2008) 92:251–260
123
Copyright © 2022 FDOKUMEN




















