
Plant regeneration from indica rice ( Oryza sativa L.) protoplasts
9
Planta (1989)178:325-333 Planta Springer-Verlag 1989 Plant regeneration from indica rice (Oryza sativa L.) protoplasts Lisa Lee *, Ronald E. Schroll**, Howard D. Grimes, and Thomas K. Hodges*** Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN 47907, USA Abstract. Rice (Oryza sativa L.) plants of the indica cultivar IR54 were regenerated from protoplasts. Conditions were developed for isolating and puri- fying protoplasts from suspension cultures with protoplast yields ranging from 1.106 to 15" 106 via- ble protoplasts/l g flesh weight. Protoplast viabi- lity after purification was generally over 90%. Pro- toplasts were cultured in a slightly modified Kao medium in a Petri plate by placing them onto a Millipore filter positioned on top of a feeder (nurse) culture containing cells from a suspension culture of the japonica rice, Calrose 76. Plating efficiencies of protoplasts ranged from 0.5 to 3.0% ; it was zero in the absence of the nurse cul- ture. Protoplast preparations usually contained no contaminating cells, and when present, the number of cells never exceeded 0.1% of the protoplasts. After three weeks the Millipore filter with callus colonies were transferred off feeder cells and onto a Linsmaier and Skoog-type medium for an addi- tional three weeks. Selected callus colonies that had embryo-like structures were then transferred to regeneration medium containing cytokinins, and regeneration frequencies up to 80% were obtained. Small shoots emerged and were transferred to jars for root development prior to transferring to pots of soil and growing the plants to maturity in growth chambers. Of the cytokinins evaluated, N 6- * Present address: Enichem Americas, Inc., Monmouth Junc- tion, NJ 08852, USA ** Present address: Coastal Plains Soil and Water Conserva- tion, Florence, SC 29502, USA *** To whom correspondence should be addressed Abbreviations: 2,4-D=2,4-dichlorophenoxyacetic acid; pcv= packed cell volume; BAP = N6-benzylaminopurine; FDA = flu- orescein diacetate; FW=fresh weight; IAA=indole-3-acetic acid Media: AA=Muller and Grafe (1978); CPW=Frearson et al. (1973); Kao * = Kao (1977); LS = Linsmaier and Skoog (1965) ; MS=Murashige and Skoog (1962); N6=Chu etal. (1975); PCM = Ludwig et al. (1985) benzylaminopurine was the most effective in pro- moting shoot formation; however, kinetin was also somewhat effective. Regeneration medium could be either an N6 or Murashige and Skoog basal medium. Of 76 plants grown to maturity, 62 were fertile, and the plant heights averaged about three- fourths the height of seed-grown plants. Two other suspension cultures of IR54, one de- veloped from the protoplast callus of the initial IR54 line, and the other developed from callus pro- duced by mature seeds, have yielded protoplasts capable of regenerating plants when using cells of the Calrose 76 suspension as a nurse culture. In addition, protoplasts obtained from three-week- old primary callus of immature embryos of IR54 were capable of regenerating plants when using the same culture conditions. Key words: Cell culture (plant regeneration) - Em- bryogenic cell and callus culture - Feeder layer - Oryza (plant regeneration) - Protoplast (culture) Introduction Regeneration of plants from protoplasts of japoni- ca genotypes of rice have been obtained (Fujimura etal. 1985; Yamada etal. 1986; Abdullah etal. 1986; Toriyama et al. 1986; Kyozuka et al. 1987); however, there have been no reports of success at regenerating rice plants from protoplasts of the in- dica genotypes. It is especially important to regen- erate the indica rice genotypes since these varieties are the principle food source in most of the tropical regions of the world. We have previously developed conditions for regenerating indica rice genotypes from primary callus of IR54 (Koetje et al. 1989). In the present study, we optimized conditions for obtaining high yields of viable protoplasts from primary callus
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Plant regeneration from indica rice ( Oryza sativa L.) protoplasts

Planta (1989)178:325-333 Planta �9 Springer-Verlag 1989
Plant regeneration from indica rice (Oryza sativa L.) protoplasts
Lisa Lee *, Ronald E. Schroll**, Howard D. Grimes, and Thomas K. Hodges*** Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN 47907, USA
Abstract. Rice ( O r y z a sat iva L.) plants of the indica cultivar IR54 were regenerated from protoplasts. Conditions were developed for isolating and puri- fying protoplasts from suspension cultures with protoplast yields ranging from 1.106 to 15" 106 via- ble protoplasts/l g flesh weight. Protoplast viabi- lity after purification was generally over 90%. Pro- toplasts were cultured in a slightly modified Kao medium in a Petri plate by placing them onto a Millipore filter positioned on top of a feeder (nurse) culture containing cells from a suspension culture of the japonica rice, Calrose 76. Plating efficiencies of protoplasts ranged from 0.5 to 3.0% ; it was zero in the absence of the nurse cul- ture. Protoplast preparations usually contained no contaminating cells, and when present, the number of cells never exceeded 0.1% of the protoplasts. After three weeks the Millipore filter with callus colonies were transferred off feeder cells and onto a Linsmaier and Skoog-type medium for an addi- tional three weeks. Selected callus colonies that had embryo-like structures were then transferred to regeneration medium containing cytokinins, and regeneration frequencies up to 80% were obtained. Small shoots emerged and were transferred to jars for root development prior to transferring to pots of soil and growing the plants to maturity in growth chambers. Of the cytokinins evaluated, N 6-
* P r e s e n t addres s : Enichem Americas, Inc., Monmouth Junc- tion, NJ 08852, USA ** P r e s e n t addres s : Coastal Plains Soil and Water Conserva- tion, Florence, SC 29502, USA *** To whom correspondence should be addressed
A b b r e v i a t i o n s : 2,4-D=2,4-dichlorophenoxyacetic acid; pcv= packed cell volume; BAP = N6-benzylaminopurine; FDA = flu- orescein diacetate; FW=fresh weight; IAA=indole-3-acetic acid
M e d i a : AA=Mul l e r and Grafe (1978); CPW=Frearson et al. (1973); Kao * = Kao (1977); LS = Linsmaier and Skoog (1965) ; MS=Murashige and Skoog (1962); N6=Chu etal. (1975); PCM = Ludwig et al. (1985)
benzylaminopurine was the most effective in pro- moting shoot formation; however, kinetin was also somewhat effective. Regeneration medium could be either an N6 or Murashige and Skoog basal medium. Of 76 plants grown to maturity, 62 were fertile, and the plant heights averaged about three- fourths the height of seed-grown plants.
Two other suspension cultures of IR54, one de- veloped from the protoplast callus of the initial IR54 line, and the other developed from callus pro- duced by mature seeds, have yielded protoplasts capable of regenerating plants when using cells of the Calrose 76 suspension as a nurse culture. In addition, protoplasts obtained from three-week- old primary callus of immature embryos of IR54 were capable of regenerating plants when using the same culture conditions.
Key words: Cell culture (plant regeneration) - Em- bryogenic cell and callus culture - Feeder layer - O r y z a (plant regeneration) - Protoplast (culture)
Introduction
Regeneration of plants from protoplasts of japoni- ca genotypes of rice have been obtained (Fujimura etal. 1985; Yamada etal. 1986; Abdullah etal. 1986; Toriyama et al. 1986; Kyozuka et al. 1987); however, there have been no reports of success at regenerating rice plants from protoplasts of the in- dica genotypes. It is especially important to regen- erate the indica rice genotypes since these varieties are the principle food source in most of the tropical regions of the world.
We have previously developed conditions for regenerating indica rice genotypes from primary callus of IR54 (Koetje et al. 1989). In the present study, we optimized conditions for obtaining high yields of viable protoplasts from primary callus

326 L. Lee et al. : Regeneration of indica rice from protoplasts
o b t a i n e d f r o m i m m a t u r e e m b r y o s a n d f r o m sus - p e n s i o n ce l l s o f I R 5 4 ( a n i n d i c a g e n o t y p e ) . P l a n t
r e g e n e r a t i o n f r o m r i ce p r o t o p l a s t s w a s o b t a i n e d
f r o m ce l l s u s p e n s i o n s as w e l l as f r o m p r o t o p l a s t s
o f t h e p r i m a r y ca l l u s . O u r p r o c e d u r e i n v o l v e s t h e
u s e o f ce l l s o f a j a p o n i c a g e n o t y p e ( C a l r o s e 76)
as a n u r s e c u l t u r e d u r i n g t h e i n i t i a l s t a g e s o f p r o t o -
p l a s t g r o w t h , t h e n s e l e c t i n g s m a l l c o l o n i e s o f e m -
b r y o g e n i c c a l l u s , a n d t r a n s f e r r i n g th i s c a l l u s t o re-
g e n e r a t i o n m e d i u m c o n t a i n i n g a c y t o k i n i n w h i c h
p r o m o t e d s h o o t d e v e l o p m e n t .
Materials and methods
Plant materials. Seeds of the indica rice (Oryza sativa L.) cv. [R54 were obtained from the International Rice Research Insti- tute (Los Banos, Philippines). Calrose 76 seeds were obtained from Dr. Niel Rutgers, University of California, Davis, USA. Plants were grown in growth chambers as described in Koetje et al. (1989).
Induction of callus and suspension cultures. Immature embryos (5-7 d after panicle extrusion) or mature seeds were surface- sterilized and cultured on either MS (Murashige and Skoog 1962) o r N 6 ( C h u et al. 1975) media as described (Koetje et al. 1989) or on LS medium (Linsmaier and Skoog 1965) containing 3 % (w/v) sucrose, 2.5 rag/12,4-dichlorophenoxyacetic acid (2,4- D), 250 mM tryptophan, and 0.4% (w/v) agarose (BRL; Life Technologies, Gaithersburg, Md., USA).
Several different suspension-culture lines of IR54 were used in this study and they are designated as follows :
IR54-1 (developed from immature embryo callus and the first line to exhibit growth of protoplasts into callus),
IR54-2 (developed from callus obtained from mature seeds),
IR54-3 (developed from protoplast-callus of IR54-1 line), IR54-4 and IR54-5 (younger lines developed from imma-
ture embryo callus). A suspension culture of IR52, developed from young inflorescences, was also used as a nurse culture.
Suspension cell cultures (approx. I g fresh weight (FW)/ 40 ml liquid medium) were initiated by carefully teasing away embryogenic regions from three- to four-week-old primary cal- lus and placing them into liquid medium. The liquid medium contained N6 basal salts and vitamins, 20 mM e-proline, 250 mM tryptophan, 2% sucrose, pH 5.8 prior to autoclaving, and either 4 mg/1 2,4-D for cell line IR54-I or 2 mg/l 2,4-D for the other IR54 cell lines. All suspension cells were cultured on a rotary shaker at 120 rpm in the dark at 25 ~ C. During the establishment of the initial cell suspensions, the medium was changed without removing any callus every ~ 3 d for two to four weeks; subsequently, the cell suspensions were subcul- tured twice each week using 5 ml packed-cell-volume (pcv) per 40 ml of medium for the next three months at which time the cultures had become established or stabilized. Thereafter, the suspensions were subcultured once each week with 5 ml pcv of cells to 40 ml of fresh medium.
Protoplast isolation and culture. Several parameters were evalu- ated in order to obtain high yields of viable protoplasts from rice cells. The final procedure used was as follows, and this was the procedure for the optimization experiments except as indicated in the figure legends. About 0.75-1 g FW of primary callus or suspension cells were incubated in 10 ml enzyme solu-
tion that contained 1% (w/v) cellulase "Onozuka '" RS (Yakult Honsha Co., Tokyo, Japan), 0.1% (w/v) Pectolyase Y-23 (Se- ishin Pharmaceutical Co., Tokyo, Japan), in CPW salts (Frear- son et al. 1973), 0.4 M mannitol, and 5 mM 2-(N-morpholin- o)ethanesulfonic acid (MES), pH 6.0. The cells were digested for 4 h on a 40-rpm shaker in the dark at 25 ~ C. The mixture was filtered through a 30-gm nylon mesh (Tetko Co., Elmsford, N.Y., USA), washed with two volumes of the CPW medium, and centrifuged at 80.g for 15 min. The protoplasts were then suspended in 1 ml of CPW medium, layered onto 5 ml of 0.6 M sucrose, and centrifuged at 40.g for 10 min. The floating and purified protoplasts (containing less than 0.1% cells) were col- lected, washed with CPW medium, and suspended in a proto- plast culture medium (Kao*) which was slightly modified from Kao (1977) by including 50 mg/1 arginine, 0.5 rag/1 2,4-D, and 7% (w/v) glucose. The osmolality of this medium was 459 m m o l . k g - 1 Fluorescein diacetate (FDA) (ICN Pharma- ceuticals, Cleveland, Oh., USA) was used to determine the via- bility of the protoplasts (Widholm 1972). Cell contamination was determined by staining cell walls with Calcofluor White (Polysciences, Warrington, Pa., USA) (Galbraith 1981).
Prior to protoplast isolation, the suspension cell cultures were transferred to either fresh N6 medium as described above, or to an MS or an amino acid (AA) medium (Muller and Grafe 1978) containing 2% sucrose and 2 mg/1 2,4-D and using 0.5 or 2.0 pcv per 40 ml of liquid.
Protoplasts were cultured using a feeder-layer method origi- nally developed for maize (Ludwig et al. 1985; Kamo et al. 1987) but with the following modifications. Protoplasts in Kao* medium (0.2 ml) were plated at a density of 5.10 s .ml-1 onto a 0.8-gm-pore Millipore filter (Millipore Co., Bedford, Ma., USA) which was placed onto feeder cells imbedded in agarose. The feeder-cell layer was prepared according to Kamo et al. (1987) and consisted of 1.5 ml pcv of 4-d-old suspension cells placed into 20 ml Kao* medium containing 0.5 mg/1 2,4-D with 0.8% (w/v) agarose (Sea Plaque LGT; FMC Co., Rockland, Me., USA), and 5 ml of this mixture was transferred to a Petri plate (60 mm diameter, 15 mm high). The feeder cells were ob- tained from one of the following sources: a Calrose 76 cell suspension that was maintained in AA medium, two IR54 cell suspensions (IR54-4, IR54-5) that were maintained in N 6 medi- um, and an IR52 cell suspension (initiated from callus obtained from young inflorescences, S. Yang and T.K. Hodges, personal communication) that was maintained in LS medium. The Cal- rose 76 cell suspension was initiated with callus obtained from immature embryos, and it was maintained originally on an N6 medium with 2 mg/1 2,4-D. After 22 months, this culture was transferred to AA medium in order to improve protoplast yields, and it is currently maintained in the AA medium.
The plated protoplast cultures were incubated in the dark at 25 ~ C. After three weeks, the plating efficiency was deter- mined by counting the number of colonies that were at least 0.1 mm in diameter (with the aid of 8 x magnification of a dis- secting microscope) and dividing the number of colonies by the number of protoplasts plated onto each filter. The filters were then transferred to 25 ml fresh LS basal medium contain- ing 2% (w/v) sucrose, 0.5 mg/1 2,4-D, and 0.4% (w/v) agarose (BRL) in Petri plates (t00 mm diameter, 15 mm high) and cul- tured in the dark at 25 ~ C. Three weeks later, callus colonies 1-2 mm in diameter with embryo-like structures were trans- ferred to regeneration medium containing either MS basal salts or N6 basal salts medium supplemented with 3% (w/v) sucrose, 0.4% agarose, 0.03 pM naphthalene-l-acetic acid, and various cytokinins at a concentration of 9 gM in Petri plates (100 mm, 25 mm). Five colonies (about 1-2 mm in diameter) were plated onto each plate. Plates were kept in the dark for one week and were then transferred to light chambers (75 p.mol, m - 2. s - 1

L. Lee et al. : Regeneration of indica rice from protoplasts 327
~ ' 6 . 0
5 . 0
4 . 0
03
m 2.0 13,. 0 s
13.
~3
5~
1.0
0.0
0.0 0 .2 0 .4 0 .6 0 .8
Mannitol Conc. (M)
I O0
80
6 0 v
55 40
2O
Fig. 1. The effect of mannitol concentration on protoplast yield and viability in IR54 indica rice. Embryo-derived, primary cal- lus was grown on MS medium supplemented with 2 rag/1 2,4-D for three weeks. Callus (approx. 1 g) was teased apart and placed into 10 ml of enzyme solution, pH 5.6, (see Materials and methods) at various mannitol concentrations for 4 h. Proto- plasts were then isolated as described in Materials and methods, and viability was determined by FDA staining
photosynthetic photon fluence rate) with 16 h-photoperiods at 25 ~ C. Plantlets with shoots 5 cm or longer were then trans- ferred into jars (100 mm high, 60 mm in diameter) containing 0.8% (w/v) agar with MS basal salts and 3% (w/v) sucrose to promote root development. Regenerated plants (> 10 cm in length) were then transferred to pots containing soil and placed in a growth chamber (day temperature 30 ~ C, night temperature 24 ~ C, with a 10-h photoperiod) and grown as described for seed-grown plants (Koetje et al. 1989).
Results
Protoplast isolation from primary callus and suspen- sion-culture cells. Initial experiments on the isola- tion of protoplasts from primary callus grown on N 6 medium using our standard culture conditions (Koetje et al. 1989) resulted in yields of only 1.104 to 5-104 protoplasts/gFW, which was too low to carry out substantial experiments related to growth of the protoplasts. Therefore, experimental condi- tions were sought that would produce at least 106 viable protoplasts/gFW. Hydrolyzing enzymes (cellulase and pectinase) used for removing cell walls were suspended in three different media (MS, PCM, CPW) and the highest yields of viable proto- plasts were obtained with CPW (data not pre- sented). The osmotic potential of the medium con- taining these enzymes in CPW was adjusted with mannitol, and although this adjustment was ex- tremely important, a broad concentration range (0.3-0.6 M) was suitable for stabilizing the proto- plasts (Fig. 1). Higher yields of protoplasts of maize were obtained when callus was grown on MS medium (Imbrie-Milligan and Hodges 1986),
N6- AGAR 0.0285
MS- . ~ 0.797 AGAR (63%)
N6- 1 0.104 AGAROSE (79%)
AGAROSE (81%) i | |
0 1 2
Viable Protoplasts (xl0 6 ) /gFW Fig. 2. The effect of basal medium (MS or N6) and solidifying agent (agar or agarose) on protoplast yield and viability in IR54 indica rice. Embryo-derived, primary callus was grown on either MS-Agar (1.5%), MS-Agarose (0.4%), N6-Agar (1.5%), or N6-Agarose (0.4%) for three weeks. All four treat- ments included 2 mg/1 2,4-D. Callus (approx. 1 g) was teased apart, placed into 10 ml of enzyme solution, pH 5.6, containing 0.4 M mannitol, and incubated for 4 h. Protoplasts were then isolated as described in Materials and methods, and viability was determined by FDA staining. The numbers without paren- theses refer to the protoplast yield; the numbers in parentheses indicate the viability (%) observed in each treatment
v 6.0 1 oo
5.5 90
5.0 80 "i"
• 4 . 5 ">"
r 7 0 4 0
3.5 60
~- 3.0 2 50 r
2.5 . ) , , . , .~ .~ . ~ , ~ , = .~ . t , , . 40 0 2 4 6 8 10 12 14 16 18 20 22 24
Time ( h )
Fig. 3. The effect of incubation time of the callus with the en- zyme solution on protoplast yield and viability in indica rice. Embryo-derived, primary callus was grown on MS medium supplemented with 2 mg/1 2,4-D and 0.4% agarose. Callus (ap- prox. 1 g) was teased apart, placed into 10 ml of enzyme solu- tion, pH 5.6, containing 0.4 M mannitol, and incubated for 4 h. Protoplasts were then isolated as described in Materials and methods, and viability was determined by FDA staining
and this was also true for rice (Fig. 2). Also, note that the yield of viable protoplasts was much high- er when the cells were grown on agarose as com- pared to agar. The length of time the callus tissue was digested with the cell-wall-hydrolyzing en- zymes was critical for obtaining both high yields and high viability of the protoplasts (Fig. 3). Four

328 L. Lee et al. : Regeneration of indica rice from protoplasts
[ ] v 7.0 LL
r 6.0
b 5.o
4.0 C/3
~. 3.0 o o n 2.0
1.0
100
9O
80 --= e ~
. m > o~
7O
60 > 5.2 5.6 6.0 6.4 6.8 7.2
pH of Enzyme Solution
Fig. 4. The effect of initial pH of the enzyme solution oll proto- plast yield and viability in 1R54 indica rice. Embryo-derived, primary callus was grown on MS medium supplemented with 2 rag/1 2,4-D and 0.4% agarose. Callus (approx. 1 g) was teased apart and placed into 10 ml of enzyme solution containing 0.4 M mannitol and incubated for 4 h. The initial pH of the enzyme solution was varied by either KOH or HC1 titration. Protoplasts were then isolated as described in Materials and methods, and viability was determined by FDA staining
hours of digestion was chosen as the standard time for subsequent experiments. We also found that the pH of the enzyme solution was very critical (Fig. 4). A slight deviation from pH 6.0 caused marked reduction in both the yield and viability of protoplasts. Taken together, these experimental conditions increased the yields of viable proto- plasts from the initial values of 1 .104- 5.104 pro- toplasts/gFW of primary callus up to about 5. 106--7.106 viable protoplasts/gFW. The proto- plast viabilities in subsequent experiments was gen- erally greater than 90%.
Protoplast yields from cells grown in N 6 liquid suspensions were initially quite low. However, when the above experimental conditions were ap- plied to several different suspension culture lines, even when grown in N 6 medium (see next section), protoplast yields varied from 1.106 to 15.10 6 via- ble protoplasts/gFW. These yields were sufficiently high that large-scale protoplast-culturing studies could be undertaken.
Growth and regeneration of cell suspensions. Several IR54 cell suspensions were established using callus obtained from either immature embryos (IR54-1, IR54-4, IR54-5) or mature seeds (IR54-2). The sus- pensions were initiated from clusters of callus that contained embryo-like structures. The growth rates of the suspension cultures were slow, with doubling times of 3-7 d for different cultures. Regeneration of plants from IR54-1 and IR54-2 suspension cul- tures were evaluated when the cultures were ten
months old. Both cultures contained cell clusters of several sizes ranging from less than 10 cells per cluster to more than 100 cells per cluster. These cells were highly cytoplasmic and contained small vacuoles. Clusters of 1-2 mm in diameter were se- lected from the suspension cultures and subcul- tured onto agar-solidified MS medium containing 0.5 mg/1 2,4-D in Petri plates (five calli per plate). After three weeks the calli were transferred to fresh MS medium lacking 2,4-D. About one-third of the calli cultured produced plants with an average of six plants per callus piece.
Protoplast culture. Protoplasts were obtained from the IR54-1 suspension line in high yields (using the optimized protocol for protoplast isolation from primary callus) when the cells were main- tained on N6 medium or if they were subcultured into either AA or MS media for 6 d prior to proto- plast isolation. The protoplast yields ranged from 1 . 1 0 6 t o 3 . / 0 6 / g F W f o r cells cultured in N 6 medi- um, from / . / 0 6 tott5.106/gFW for cells on AA medium, and from 1.106 to 6" i06/gFW for cells on MS medium. Protoplast size ranged from 5 to 20 pm in diameter (Fig. 5 b). For different proto- plast isolations, the final preparations obtained after flotation on a pad of sucrose contained either no contaminating cells (with walls) or less than 0.1% cells.
Plating efficiencies for protoplasts of the IR54- / line were determined after three weeks of culture (Table 1) and in the presence or absence of differ- ent cell lines as feeders. With Calrose 76 as the feeder cells, protoplast plating efficiencies for IR54-1 ranged from 0.5 to 3.0% when the suspen- sion cultures were of different ages, and when they had been cultured in different media for 6 d prior to protoplast isolation. The particular media and cell density at the time of the last subculture prior to protoplast isolation did not appear to make any consistent difference in plating efficiency when cells of Calrose 76 (Fig. 5 a) were the feeder (Ta- ble 1). In some experiments, the effectiveness of Calrose 76 as the nurse culture varied from flask to flask. The basis for this variation is currently being evaluated, but it does not appear to be relat- ed to rates of growth or changes in cluster sizes. When IR54-/ was cultured in AA medium, both IR54-4 and IR52 functioned initially as effective nurse cultures (plating efficiencies of 0.5%) but these nurse cultures did not sustain the growth of the IR54-1 protoplasts back into callus. In subse- quent studies, however, the IR52 suspension cul- ture (an indica genotype) was found to be capable of supporting IR54-1 protoplasts to grow into cal-

L. Lee et al. : Regeneration of indica rice from protoplasts 329
Fig. 5. a Clusters of Calrose 76 japonica-rice cell suspensions that were used for nurse cells in the protoplast culture studies. b Freshly isolated protoplasts from IR54-1 indica-rice suspension cells, e Callus obtained from protoplasts of IR54-1 grown on a filter for four weeks after transfer to LS medium (seven weeks after isolation), d Embryo-like structures on callus obtained from protoplasts of IR54-1 cell suspensions, e Protoplast-derived callus from IR54-1 cell suspensions with a shoot breaking through a coleoptile
lus and to generate plants. In separate experiments it was found that IR54-1 was ineffective as a feeder culture for its own protoplasts. It should be noted that protoplast growth back into callus of an indi- ca genotype (IR54-1) was most effective when a japonica genotype (Calrose 76) was the feeder. These results illustrate the strict necessity for an appropriate nurse culture in order for protoplasts of the IR54-1 line to grow into callus.
Calli derived from protoplasts were easily seen with the unaided eye on the filter about four weeks after isolation, and a lawn of callus was formed about seven weeks after isolation (Fig. 5c). Many
embryo-like structures were visible on the callus (Fig. 5 d). Calli containing these embryo-like struc- tures were selected and transferred to regeneration media. Shoot-like structures appeared about two to three weeks after transferring the callus to regen- eration medium and into light (Figs. 5 e, 7 a).
Regeneration of IR54-! plants from protoplast-de- rived-callus; importance of cytokinins. Ten (or fif- teen for the MS-2 treatment) pieces of calli con- taining embryo-like structures (Fig. 5d) were se- lected and transferred to regeneration media con- taining different cytokinins (Fig. 6). The regenera-

330 L. Lee et al. : Regeneration of indica rice from protoplasts
Table 1. Effect of feeder (nurse) cells on the plating efficiency of rice protoplasts obtained from IR54-1 suspension cells when the culture was 7, 10, or 12 months old. Data are protoplast plating efficiency (%)
Feeder cells IR54-1 age (months)
IR54-1 suspension media 6 d prior to protoplast isolation
N6_5a AA-2 AA-0.5 MS-2 MS-0.5
None 7 0 0 0 0 0 Catrose 76 7 2.5 0.5 3.0 3.0 1.3
10 2.6 0.5 0.9 0.7 0.8 12 1.4 1.7 1.6 1.3 1.3
IR54-4 7 <0.1 <0.01 0.5 <0.1 <0.01 IR52 7 < 0.001 0.5 0.5 <0.1 < 0.01 IR54-5 7 < 0.01 < 0.01 < 0.1 0 0
a The IR54-1 cell line was routinely maintained in N 6 medium. In this experiment, cells were cultured for 6 d in different media (N6, AA, or MS), and the cell densities were varied at subculture by using 0.5, 2.0, or 5.0 ml pcv per 40 ml of liquid media
8O
60
40 o
20
a Regeneration Media [ ] MSB [, a~ I [ ] MSK
�9 oN. . o o N6-5 AA-2 AA-0.5 MS-2
Suspension Media MS-0,5
g
z
15
10
c U Regeneration Media
[ ] MSB
"~ [ ] MSZ I ' L [ ] MSK
|o~ d oo. N6-5 AA-2 AA-0.5 MS-2 MS-0,5
Suspension Media
25
g 20
15 k~
g �9 = 10
5 o er
Regeneration Media �9 N6B [ ] N6Z [ ] N6K
0 o ~ ~ i oo o o o N6-5 AA-2 AA-0.5 MS-2 MS-0.5 MS-0.5
Suspension Media
15 Regeneration Media
~ d ~ N6B N6Z 10Ii
5
R Z 0 0 0 0 0 . 0 0 . 0
N6-5 AA-2 AA-0.5 MS-2 Suspension Media
Fig. 6a-d. Effect of different cytokinins in the regeneration media on the frequency of plant regeneration of rice protoplasts isolated from IR54-1 suspension cells grown for 6 d in five different media. Protoplasts were isolated in CPW medium and subsequently cultured in Kao* medium�9 The N6-5 suspension medium represent suspension cells subcultured to N6 medium with 5 ml pcv to 40 ml of liquid medium�9 For regeneration, colonies were selected that possessed embryo-like structures and transferred to either MS or N6 media with the cytokinins. Ten (or 15 for MS-2 treatment) calli were plated in each regeneration medium from each suspension-medium treatment. Calrose 76 was used as nurse cells, a Frequency of plant regeneration (% of selected protoplast-calli that produced plants) on MS media with either B (benzylaminopurine), Z (zeatin), or K (kinetin) at 2 mg/1. b Frequency of plant regeneration (% of selected protoplast calli that produced plants) on N6 media with B, Z, or K at 2 mg/l. e Number of plants regenerated from 10 (or i5, see above) protopiast-derived calli on MS media with various cytokinins, fl Number of plants regenerated from 10 (or 15, see above) protoplast-derived calli on N6 media with various cytokinins. These results were from a single experiment
tion media contained either N6 or MS media with either benzylaminopurine (MS-B, N6-B), zeatin (MS-Z, N6-Z), or kinetin (MS-K, N6-K). The number of calli that produced plants was deter- mined (Fig. 6 a, b) as well as the number of plants
that were produced by the calli (Fig. 6c, d). Al- though there were some exceptions, regeneration frequency of calli was generally higher with MS (Fig. 6a) than with N6 (Fig. 6b) as the regenera- tion medium. Similarly, the number of plants re-
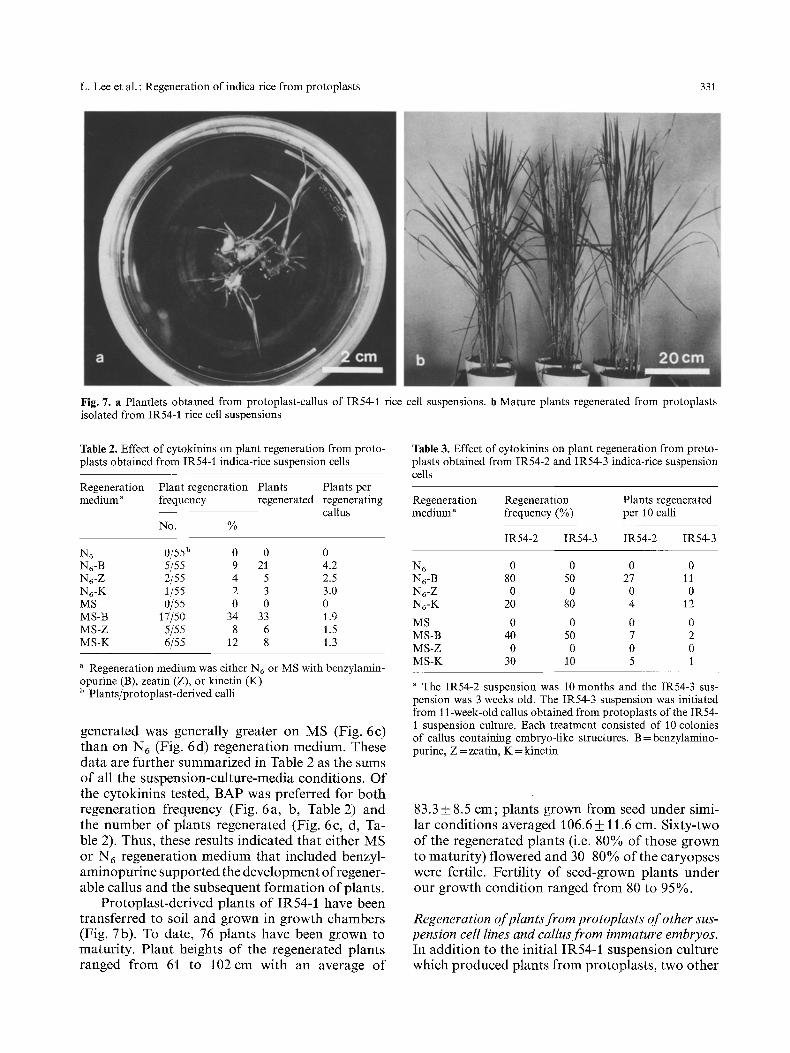
L. Lee et al. : Regeneration of indica rice from protoplasts 331
Fig. 7. a Plantlets obtained from protoplast-callus of IR54-1 rice cell suspensions, b Mature plants regenerated from protoplasts isolated from IR54-1 rice cell suspensions
Table 2. Effect of cytokinins on plant regeneration from proto- plasts obtained from IR54-1 indica-rice suspension cells
Table 3. Effect of cytokinins on plant regeneration from proto- plasts obtained from IR54-2 and IR54-3 indica-rice suspension cells
Regeneration Plant regeneration Plants Plants per medium" frequency regenerated regenerating Regeneration Regeneration Plants regenerated
callus medium a frequency (%) per 10 calli No. %
IR54-2 IR54-3 IR54-2 IR54-3 N~ 0/55 b 0 0 0 N6-B 5/55 9 21 4.2 N 6 0 0 0 0 N~-Z 2/55 4 5 2.5 N6-B 80 50 27 11 N6-K 1/55 2 3 3.0 N6-Z 0 0 0 0 MS 0/55 0 0 0 N6-K 20 80 4 12 MS-B 17/50 34 33 1.9 MS 0 0 0 0 MS-Z 5/55 8 6 1.5 MS-B 40 50 7 2 MS-K 6/55 12 8 1.3 MS-Z 0 0 0 0
MS-K 30 10 5 1 a Regeneration medium was either N 6 o r MS with benzylamin-
opurine (B), zeatin (Z), or kinetin (K) ~ The IR54-2 suspension was 10 months and the IR54-3 sus- b Plants/protoplast-derived calli pension was 3 weeks old. The IR54-3 suspension was initiated
from ll-week-old callus obtained from protoplasts of the IR54- I suspension culture. Each treatment consisted of 10 colonies of callus containing embryo-like structures. B = benzylamino- purine, Z = zeatin, K = kinetin
generated was generally greater on MS (Fig. 6c) than on N6 (Fig. 6d) regeneration medium. These data are further summarized in Table 2 as the sums of all the suspension-culture-media conditions. Of the cytokinins tested, BAP was preferred for both regeneration frequency (Fig. 6a, b, Table 2) and the number of plants regenerated (Fig. 6c, d, Ta- ble 2). Thus, these results indicated that either MS or N6 regeneration medium that included benzyl- aminopurine supported the development of regener- able callus and the subsequent formation of plants.
Protoplast-derived plants of IR54-1 have been transferred to soil and grown in growth chambers (Fig. 7b). To date, 76 plants have been grown to maturity. Plant heights of the regenerated plants ranged from 61 to 102 cm with an average of
83.3 _+ 8.5 cm; plants grown from seed under simi- lar conditions averaged 106.6 + 11.6 cm. Sixty-two of the regenerated plants (i.e. 80% of those grown to maturity) flowered and 30 80% of the caryopses were fertile. Fertility of seed-grown plants under our growth condition ranged from 80 to 95%.
Regeneration of plants from protoplasts of other sus- pension cell lines and callus from immature embryos. In addition to the initial IR54-1 suspension culture which produced plants from protoplasts, two other

332 L. Lee et al. : Regeneration of indica rice from protoplasts
Table 4. Plant regeneration from protoplasts obtained from calli" from immature embryo of IR54 indica rice
Regeneration Calli plated Calli Regeneration Plants Plants regenerated medium forming plants frequency (%) regenerated per callus
N6-B 25 16 64 87 5.4 MS-B 25 16 64 71 4.4
a Immature embryos were plated on LS medium supplemented with 2% sucrose and 0.5 mg/1 2,4-D and protoplasts were isolated from three-week-old embryogenic calli. Regeneration media were N6 or MS containing benzylaminopurine (B)
IR54 suspension lines - IR54-2 (initiated with seed callus) and IR54-3 (initiated with protoplast-callus of IR54-1) - have been developed that are capable of producing protoplasts that regenerate into ma- ture plants. These cell cultures were grown in Ne medium. Protoplast callus of both IR54-2 and IR54-3 was regenerable when cultured on either N6 or MS medium (Table 3). The IR54-2 line re- quired cytokinins in the regeneration media, and like the IR54-1 line, BAP was the most effective for both regeneration frequency and number of plants produced. For the IR54-3 cells, both BAP and kinetin supported plant regeneration.
Protoplasts obtained from embryogenic pieces of three-week-old primary callus of IR54 immature embryos were also capable of regenerating plants. Although protoplast plating efficiency was only 0.3%, 64% of the callus clusters selected and placed onto regeneration medium (either N6 or MS with BAP) produced plants (Table 4). The number of plants per regenerating callus was 5.4 with N6-B regeneration medium and 4.4 with MS-B regenera- tion medium (Table 4).
Discussion
We succeeded in obtaining regeneration of mature, fertile plants from protoplasts of the indica rice cultivar IR54. These results were obtained using protoplasts isolated from three different cell sus- pension lines and from three-week-old primary cal- lus obtained from immature embryos. The suspen- sion cultures were developed using callus from im- mature embryos (IR54-1), mature seeds (IR54-2), and from protoplast calli (IR54-3, derived from IR54-/). Plant regeneration from rice protoplasts of japonica rice cultivars has been reported pre- viously (Fujimura et al. 1985; Yamada et al. /986; Abdullah et al. 1986; Toriyama et al. 1986; Kyo- zuka et al. /987); it is now possible to regenerate fertile plants from protoplasts of IR54, an indica variety. A combination of factors was necessary to overcome the difficulties of protoplast isolation and culture (resynthesis of cell walls, initiation of and sustained cell divisions), and finally plant re-
generation from these indica-rice cells. First, the primary callus (Koetje et al. 1989) and the cell sus- pensions were capable of plant regeneration at the time of protoplast isolations. Although this may not be absolutely essential (Abdullah et al. /986), it would seem to be a desirable situation. Second, we were able to obtain conditions that allowed for the release of reasonably large numbers of viable protoplasts (Figs. 1-4) which made protoplast-cul- ture experiments possible. Third, use of the feeder- layer technique in protoplast culture was found to be essential (Table 1). The source (both geno- type and cell line) of nurse cells in our feeder layer proved to be critical in supporting the formation and subsequent growth of colonies from proto- plasts. Only suspension cells of a line of Calrose 76 (a japonica-type rice) and of one IR52 (indica) line were effective as nurse cells. The Calrose 76 suspension cells also promoted callus formation from protoplasts of other japonica rice cultivars, including itself (Calrose 76) and Taipei 309 (data not shown). Fourth, selection of small (1-2 mm in diameter) embryogenic calli at an early stage of callus formation (Fig. 5 d) and transferring to re- generation medium was also important. Finally, the use of a cytokinin, especially N6-benzylamino - purine, in the regeneration medium was found to be most critical (Tables 2, 3; Fig. 6). Thus, a com- bination of several factors was important for ob- taining regenerated plants from protoplasts of IR54.
Some other conditions were not so critical. For example, growing the IR54-1 line in three different liquid media CN~, MS, AA) for 6 d prior to proto- plast isolation did not eliminate the regeneration ability of the cells (Table 1). This illustrates some resiliency of the cells at least during a short period. In this regard it is important also to note that plant regeneration from protoplasts of japonica geno- types grown in several different liquid media (Fuji- mura et al. 1985; Yamada etal. 1986; Abdullah et al. 1986; Toriyama et al. 1986; Kyozuka et al. /987) have been reported.
It was also possible to regenerate plants from protoplasts derived directly from three- to four-

L. Lee et al. : Regeneration of indica rice from protoplasts 333
week-old primary callus of immature embryos of IR54. Conditions required for protoplast growth were similar to those developed for the protoplasts from suspension cultures. The capability of proto- plasts of primary callus of immature embryos to regenerate plants is obviously of considerable in- terest because of the ease of initiating primary cal- lus as compared to the difficulty, time, and labor involved in initiating and maintaining cell suspen- sions. In addition, less time in culture produces less chance for somaclonal variation and loss of totipotency.
The cytokinin requirement for plant regenera- tion from embryogenic calli of IR54-1 protoplasts is interesting because cytokinins are frequently as- sociated with regeneration phenomena involving organogenesis. However, prior to the formation of plantlet structures, embryo-like structures were ob- served on the protoplast calli, and the high fre- quencies of these structures must, in part at least, account for the high regeneration frequencies found. Hormone-free regeneration media resulted in the appearance of a few green spots on the pro- toplast calli, but these calli did not produce plants. This finding is consistent with the report of Kyo- zuka et al. (1987) that cytokinins (kinetin and zea- tin) at 2 or 5 mg/1) (9 gM) concentrations stimu- lated plant regeneration from protoplast calli de- rived from some japonica rice cultivars; however, cytokinin was apparently not required for regener- ation from protoplasts of other japonica lines (Ab- dullah et al. 1986; Kyozuka et al. 1987). The posi- tive cytokinin effect on regeneration might consist in balancing the endogenous IAA levels present in cells and tissue since the IAA precursor, trypto- phan, was present in the media used in the present study.
Production of transgenic japonica-rice plants has recently been reported (Toriyama et al. 1988; Zhang and Wu 1988), and it should also be possible now to genetically engineer the indica cultivars by means of transformation of the protoplasts and regeneration of fertile plants.
The technical assistance of Michelle Brady in this research was greatly appreciated. The research was supported by the Rocke- feller Foundation. This is Journal Paper No. 11785 from the Purdue Agricultural Experiment Station.
References Abdullah, R., Cocking, E.C., Thompson, J.A. (1986) Efficient
plant regeneration from rice protoplasts through somatic embryogenesis. Bio/Technology 4, 1087-1090
Chu, C.C., Wang, C.C., Sun, C.S., Hus, C., Yin, K.C., Chu, C.Y. (1975) Establishment of an efficient medium for anther culture of rice through comparative experiments on the ni- trogen sources. Sci. Sin. 18, 659 668
Fujimura, T., Sakurai, M., Akagi, H., Negishi, T., Hirose, A. (1985) Regeneration of rice plants from protoplasts. Plant Tissue Cult. Lett. 2, 74-75
Frearson, E.M., Power, J.B., Cocking, E.C. (1973) The isola- tion, culture and regeneration of Petunia leaf protoplasts. Dev. Biol. 33, 130-137
Galbraith, D.W. (1981) Microfluorimetric quantitation of cellu- lose biosynthesis by plant protoplasts using calcofluor white. Physiol. Plant. 53, 111 116
Imbrie-Milligan, C.W., Hodges, T.K. (1986) Microcallus for- mation from maize protoplasts prepared from embryogenic callus. Planta 168, 395-401
Kamo, K.K., Chang, K.L., Lynn, M.E., Hodges, T.K. (1987) Embryogenic callus formation from maize protoplasts. Planta 172, 245-251
Kao, K.N. (1977) Chromosomal behavior in somatic hybrids of soybean - Nicotiana glauca. Mol. Gen. Genet. 150, 225 230
Koetje, D.S., Grimes, H.D., Wang, Y.C., Hodges, T.K. (1989) Regeneration of indica rice (Oryza sativa L.) from primary callus derived from immature embryos. J. Plant Physiol., in press
Kyozuka, J., Hayashi, Y., Shimamoto, K. (1987) High fre- quency plant regeneration from rice protoplasts by novel nurse culture methods. Mol. Gen. Genet. 206, 408-413
Linsmaier, E.M., Skoog, F. (1965) Organic growth factor re- quirements of tobacco tissue cultures. Physiol. Plant. 18, 100-127
Ludwig, S.R., Somers, D.A., Petersen, W.L., Pohlman, R.F., Zarowitz, M.A., Gengenbach, B.G., Messing, J. (1985) High frequency callus formation from maize protoplasts. Theor. Appl. Genet. 71, 344-350
Muller, A.J., Grafe, R. (1978) Isolation and characterization of cell lines of Nicotiana tabacum lacking nitrate reductase. Mol. Gen. Genet. 161, 67-76
Murashige, T., Skoog, F. (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15, 473-497
Toriyama, K., Hinata, K., Sasaki, T. (1986) Haploid and dip- loid plant regeneration from protoplasts of anther callus in rice. Theor. Appl. Genet. 73, 16-19
Toriyama, K., Arimoto, Y., Uchimiya, H., Hinata, K. (1988) Transgenic rice plants after direct gene transfer into proto- plasts. Bio/Technology 6, 1072-1074
Widholm, J.M. (1972) The use of fluorescein diacetate and phenosafranine for determining viability of cultured plant cells. Stain Technol. 47, 189 194
Yamada, Y., Yang, Z.Q., Tang, D.T. (1986) Plant regeneration from protoplast-derived callus of rice (Oryza sativa L.) Plant Cell Rep. 4, 85-88
Zhang, H.M., Yang, H., Reeh, E.L., Golds, T.J., Davis, A.S., Mulligan, B.J., Cocking, E.C., Davey, M.R. (1988) Trans- genic rice plants produced by electroporation-mediated plasmid uptake into protoplasts. Plant Cell Rep 7, 379 384
Received 2 December 1988; accepted 3 March 1989




















