
Experimentally induced subclinical mastitis: are lipopolysaccharide ...

Cloning of hamster osteopontin and expressiondistribution in normal tissues and experimentallyinduced oral squamous-cell carcinoma
Hangqing Jin a, Paloma Valverde b, Jake Chen b,*
Archives of Oral Biology (2006) 51, 236—245
www.intl.elsevierhealth.com/journals/arob
aDepartment of Pediatric Dentistry, University of Texas Health SciencesCenter at San Antonio, San Antonio, TX, USAbDivision of Oral Biology, Tufts University School of Dental Medicine,One Kneeland Street, Boston, MA 02111, USA
Accepted 22 June 2005
KEYWORDSOsteopontin;Hamster;DMBA-inducedcarcinogenesis;Oral squamous-cellcarcinoma
Summary Osteopontin (OPN) is a non-collagenous extracellular matrix (ECM)protein expressed and secreted by several human cancers. This study investigatedthe expression pattern of OPN during development of oral squamous-cell carcinomaby using 7,12-dimethylbenz[a]anthracene (DMBA)-induced squamous-cell carcinomasin buccal pouch of syrian golden hamsters. We first identified the hamster OPN cDNAsequence by screening of a hamster calvariae cDNA library with a rat OPN cDNA probe.The resulting 1449 bp of hamster OPN cDNA led to a deduced protein sequence of 305amino acids containing several putative binding sites to integrins, CD44 receptors,calcium ions and hydroxyapatite, as well as multiple sites for phosphorylation,glycosylation and sulphation. Hamster OPN cDNA was then used as a probe to analyzethe expression of OPN mRNA by Northern blot and in situ hybridization analyses ofnormal and malignant tissues. OPN mRNA was detected in several non-mineralizedtissues as well as in mineralized tissues, but was not present in normal hamster buccalepithelium. DMBA-treated hamster buccal pouches expressed OPN mRNA as early as 4weeks and displayed the highest level of expression at 15 weeks. The specimenstreated with DMBA for 15 weeks exhibited histological features of squamous-cellcarcinoma, presented microcrystalline deposits and showed OPN expression asso-ciated with malignant epithelium and tumor-associated macrophages. To summarize,our results suggest that buccal-pouch carcinogenesis of Syrian golden hamster mayconstitute an excellent experimental model to study the mechanisms by which OPN isassociated with oral cancer pathogenesis, and to validate OPN-based therapeuticapproaches to ameliorate oral cancer progression and metastasis.# 2005 Elsevier Ltd. All rights reserved.
Abbreviations: DMBA, 7,12-dimethylbenz[a]anthracene; ECM, extracellular matrix proteins; OPN, osteopontin* Corresponding author. Tel.: +1 617 636 2729; fax: +1 617 636 0878.E-mail address: [email protected] (J. Chen).
0003–9969/$ — see front matter # 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.archoralbio.2005.06.013

Hamster OPN mRNA expression and oral cancer 237
Introduction
OPN is an ECM protein that plays critical roles in theformation and remodeling of mineralized tissues,1,2
inflammation,2,3 cellular transformation and metas-tasis.4—6
OPN protein sequence is highly modified at thepost-translational level by phosphorylation, glycosy-lation and sulphation.1 It contains several structuraldomains that allow its interaction with cells throughseveral integrin types and CD44 receptors.1,3 Briefly,it has the arginine-glycine-aspartic acid (RGD)domain present in many ECM proteins that is criticalfor binding to avb3, avb1, avb5 or a5b1 integrins.1,3
It also contains binding sites for a9b1 and a4b1integrins7,8 in close proximity to the RGD domain.Furthermore, it harbors one or two heparin-bindingdomains close the carboxyl-terminus of the proteininvolved in the binding to various isoforms of CD44.1,3
In addition to binding to cells, OPN is thought tointeract with hydroxyapatite and calcium ionsthrough aspartate- (D-) rich sequences,1 and to con-tain additional binding domains for calcium ions closeto the carboxyl-terminus of the protein.3 Binding tocalcium and heparin, as well as cleavage of theprotein through its two to three thrombin cleavagesites have also been suggested to regulate interac-tions with integrin-binding domains.3
Even more than 2 decades after its discovery as amarker of epithelial cells transformation,9 there isconsiderable interest in the roles of OPN in tumor-igenesis, both as a marker of malignancy and as acandidate for testing as a prognostic factor.4—6 Tostudy the association between cell transformation,metastasis andOPNexpression in oral cancer, there isa need for an animal model where tumors exhibitsimilar markers than in human and where tumorprogression canbe analyzed reproducibly. Thebuccalpouch of the Syrian hamster is an excellent model toinvestigate oral cancer development, because squa-mous-cell carcinomas induced by the application ofDMBA to the buccal pouch are morphologically andhistologically similar to human tumors,10 and expresssimilar metabolic andmolecularmarkers.11—13 In thisstudy, we have used this animal model to clonehamster OPN cDNA and to analyze OPNmRNA expres-sion pattern in normal tissues and DMBA-induced oralsquamous-cell carcinomas.
Materials and methods
Animals
Outbred 6-week-old male Syrian golden hamsters(Mesocricetus auratus) were purchased from the
National Cancer Institute (Frederick, MD). All pro-cedures using these animals were conducted incompliance with the state regulations, andapproved by the Institutional Animal Care and Useprogram of the University of Texas Health ScienceCenter (San Antonio, TX).
Generation of tumors in hamster buccalpouches
Animals were randomly divided into two groups.In the experimental group, the carcinogen DMBA(Sigma) dissolved in mineral oil (0.5%) was appliedtopically to the left buccal-pouch mucosa with awool brush three times per week. By using poucheson only the left side, the unilateral formation oftumors allowed the experimental animals to eatand swallow normally. The control group was trea-ted with Ringer’s solution over the same timeperiod as the experimental group. To obtain tissuesat different stages of tumor development, animalswere sacrificed at different times (2—15 weeks)after carcinogenesis had been initiated, by per-sonnel blinded to the treatment assignment.During the study, animals were weekly monitoredfor gross abnormalities, including formation oftumors, change of surface color or texture altera-tions. Untreated pouches or Ringer’s-treatedcontrol pouches did not exhibit apparent changes(data not shown). Thickened mucosa with arough surface and whitish granular appearancewas mostly observed in the 4- and 6-week DMBA-treated pouches. In the 15-week DMBA-treatedpouches, solid tumors begun to appear in thepouch mucosa. Normal and tumor tissues werecollected and immediately prepared for histologi-cal evaluation, in situ hybridization or immunohis-tochemistry, and the remainder immediatelyfrozen in liquid nitrogen for subsequent RNA iso-lation.
Isolation of hamster OPN cDNA
An amplified UniZapTM XR Library of adult hamstercalvaria was constructed and plated as previouslydescribed.15 Initial screening of the library wasperformed with a rat OPN cDNA probe labeled with[a-32P]dCTP (>3000 Ci/nmol; Dupont NEN) by usinga T7 Quick Prime kit (Pharmacia) according to theStratagene protocol. Filters were hybridized over-night at 65 8C and then washed in 0.1 � SSC and 0.1%SDS at 65 8C followed by autoradiography for detec-tion of positive clones. Upon replating the positiveclones, secondary screening allowed the identifica-tion of three positive plaques that were selectedand grown to prepare cDNA for sequence analysis.

238 H. Jin et al.
Sequencing was performed by the Center forAdvanced DNA Technologies at the University ofTexas Health Science Center.
Sequence analyses of OPN cDNA andprotein
To translate the hamster OPN cDNA sequence intoprotein sequence, we used ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). DifferentExPaSy proteomics tools available online (http://www.us.expasy.org/) were used to analyze thededuced hamster OPN protein sequence as pre-viously described14 with minor modifications.Briefly, we used PROSITE for identification of pro-tein domains and motifs for post-translationalmodifications. Results obtained with PROSITE wereconfirmed with SignalP (to confirm the location ofthe signal peptide cleavage site), Sulfinator (toconfirm the tyrosine sulfation sites), NetPhos (toconfirm Ser/Thr phosphorylation sites) and NetN-Glyc (to confirm N-glycosylation sites). Putativebinding sites for integrins, calcium or heparinand thrombin cleavage sites, were predicted byextrapolating published functional studies corre-sponding to mouse and human OPN sequences.Multiple alignments of amino acid sequences ofOPN in different species were performed withClustalW.
RNA isolation and Northern blot analysis
Total RNA was isolated with a Micro RNA isolationkit (Stratagene). For Northern blot analyses, 15 mgof each RNA sample from control and experimentalanimals were subjected to electrophoresis in 1%agarose gels containing 2.2 M formaldehyde andthen transferred to nylon membranes by capillarytransfer. Prehybridization of the membranes wasperformed as previously described.15 Hybridizationwas performed at 42 8C with an [a-32P]-dCTP(>3000 Ci/nmol; Dupont NEN)-labeled hamsterOPN cDNA probe prepared from the full-lengthcDNA using a T7 Quick Prime kit (Pharmacia).After hybridization, the membranes were washedas previously described15 and exposed to KodakX-Omat film at �80 8C using two intensifyingscreens.
In situ hybridization andimmunohistochemical analyses
Preparation of OPN riboprobes and in situ hybridiza-tion analysis were performed by methods previouslydescribed.15,16 Immunohistochemistry was per-formed with a monoclonal mouse anti-cytokeratin
concentrate antibody (Zymed Laboratories), abroad spectrum monoclonal antibody cocktail ofclones A1 and A3 that reacts with cytokeratins 10,14/15, 16 and 19. Sections were processed forimmunohistochemical analysis with a Histostain-SPkit (Zymed Laboratory) as previously described.17
Results
Isolation and sequence characterization ofhamster OPN cDNA and protein
Screening of a hamster calvariae cDNA library with arat OPN cDNA probe, followed by sequencing ofthree independent positive plaques, led to theidentification of hamster OPN cDNA (Fig. 1A). The1449-bp hamster OPN cDNA sequence included 80 bpof 50-untranslated region, 915 bp of open-readingframe and 454 bp of 30-untranslated region. Onetypical polyadenylation site and a stretch of ade-nines were also found at the 30-untranslated region.The ATG corresponding to the first methionine of thededuced hamster OPN protein sequence of 305amino acids, conformed with the Kozak consensussequence for adequate mammalian initiationcodons.18 Analysis of the hamster OPN proteinsequence with the program PROSITE, revealed thepresence of 16 amino acids signal sequence, two N-glycosylation sites, one sulphation site and multiplephosphorylation sites for casein kinase II or proteinkinase C (Fig. 1A). In addition, this software recog-nized a D-rich domain for potential binding to hydro-xyapatite and calcium ions and the RGD domain(Fig. 1A). To identify additional domains and motifsin the hamster OPN protein sequence, we extrapo-lated published information of functional analysis ofOPN protein sequence in other species.1,3,8 Thus,non-RGD integrin-binding sites, and calcium- orheparin-binding sites were also predicted for ham-ster OPN protein as summarized in Fig. 1B. In addi-tion, two out of the three thrombin cleavage sitesdescribed for mouse OPN were also found in thehamster orthologue (Fig. 1B).
We next performed the alignment of hamster OPNamino acid sequence with that corresponding toseven OPN orthologues with the program ClustalW.As shown in Fig. 2, there was a high degree ofsequence conservation across species of the major-ity of the domains depicted in Fig. 1B, with someexceptions. For example, the calcium-bindingdomain was missing in the OPN sequences of cowand sheep, and the binding site for a4b1ELVTDFPTDLPAT previously described for humanOPN,8 was only conserved in the pig and orangutanorthologues (Fig. 2).

Hamster OPN mRNA expression and oral cancer 239
Figure 1 Hamster OPN cDNA, deduced protein sequence and predicted domain distribution. (A) Hamster OPN cDNAand protein. The nucleotide sequence of hamster OPN cDNA has been deposited with EMBL under Accession numberAJ890352. Numbers on the left indicate nucleotide number. The first ATG corresponding to the first methionine is shownin bold, the stop codon for translation is indicated with an asterisk and potential polyadenylation sites at the 30
untranslated region are double underlined. Putative N-glycosylation sites are single underlined, protein kinase Cphosphorylation sites are double underlined, casein kinase II phosphorylation sites are shown in gray background and atyrosine sulphation site is shown in black background. The signal peptide cleavage site is indicated with two arrowsbetween amino acids 16 and 17, the aspartate-rich sequence is shown between brackets and the RGD domain is dottedunderlined. (B) Predicted domain distribution. Information relative to domains and motifs predicted with onlineprogramswas combinedwith extrapolated information from published functional studies of human ormouse OPN (showreferences here). Domains are shown as boxes except for the heparin-binding domains which are also underlined. Thefirst and last amino acid of each domain are indicated outside the boxes. Thrombin cleavage sites are indicated witharrows.

240 H. Jin et al.
Figure 2 Sequence alignment of hamster OPN protein and closest orthologues. The amino acid sequences of OPN fromhamster, cow, sheep, pig, human, orangutan, rat and mouse were aligned with the program ClustalW. The consensus lineobtained by ClustalW analyses means: (*) identical or conserved residues in all sequences in the alignment; (:) indicates

Hamster OPN mRNA expression and oral cancer 241
Figure 3 Expression of hamster OPN mRNA in normal tissues. (A) Northern hybridization of OPN mRNA expression wasperformed with total RNA prepared from several hamster tissues from 1-day-old male hamsters using 32P-labeled hamstercDNA as a probe. The relative amount of RNA loaded in each line is indicated by the ethidium bromide staining of the 18 Sand 28 S ribosomal RNA subunits shown in the panel beneath the gel. OPN expression was seen in mandible, brain, kidney,skin and stomach. (B) In situ hybridization analyses with OPN riboprobe in hamster tibia. In situ hybridization using a cRNAprobe for hamster OPN was performed on sections of the tibia from a 1-day-old male hamster. Bright (B) and dark (C) fieldmicrographs are illustrated. Hypertrophic cartilage cells (arrowheads) in the growth plate showed a strong hybridizationsignal whereas fibroblastic cells in the perichondrium (pc) lacked of signal. Magnification �250.
Tissue specific expression of hamster OPNmRNA in mineralized and non-mineralizedtissues
To determine the expression profile of hamster OPNin normal tissues, we performed Northern blot ana-lysis of total RNA isolated from different tissues byusing the full-length hamster OPN cDNA as a probe(Fig. 3A). A single 1.5-kb band corresponding tohamster OPN mRNA, was clearly observed in themandibular bone sample, used as positive control, aswell as in several non-mineralized tissues includingbrain, kidney, muscle, skin and stomach (Fig. 3A).
Further analyses of OPN mRNA expression by insitu hybridization on hamster tibial bone tissuesections, showed that hypertrophic cartilage cellsin the growth plate strongly hybridized to the OPNriboprobe whereas cells in the perichondrium,essentially composed of collagen fibers and fibro-blasts, lacked of hybridization signal (Fig. 3B and C).In situ hybridization in other normal hamster tissueswere similar (data not shown) to that in our previousstudies for rat and porcine OPN.19—21
conserved substitutions; (.) indicates semi-conserved substitand calcium-binding domains are shown as D-RICH, HEP and CAdomains are dotted underlined and single underlined, respdescribed formouse OPN (RG, RS and RS, shown in dark backgrothe hamster orthologue. The putative a4b1 binding site debackground) is absent in the hamster OPN sequence.
Expression of hamster OPN in buccal-pouch tumor tissue
In the next set of experiments, we performed North-ern blot hybridization analysis with OPN cDNA probeof normal buccal mucosa and DMBA-treated buccalmucosa. As shown in Fig. 4A, normal buccal mucosadid not express OPN mRNA, whereas buccal mucosaof animals treated with DMBA expressed OPN mRNAlevels that increased with the exposure time. After15 weeks of topical administration of DMBA on thehamster buccal pouch mucosa, multiple tumorousnodules were clearly noticeable in each animal(Fig. 4B and C). Tumors were well differentiated,with cellular polymorphism, hyperchromatism andnumerous keratin pearls confirming histologicallytheir identity as squamous-cell carcinomas as pre-viously described.15
In situ hybridization analyses with OPN riboprobeon sections of tumor tissues indicated that trans-formed squamous cells of the hamster buccalpouch tumor expressed OPN mRNA at markedlyincreased level than normal tissue (Fig. 5A and B).
utions. Aspartate-rich sequence, heparin-binding domain-BINDING, respectively. RGD and SLAYGLR integrin-bindingectively. Of the three putative thrombin cleavage sitesund), only the first two are conserved or semi-conserved inscribed for human OPN (ELVTDFPTDLPAT shown in gray
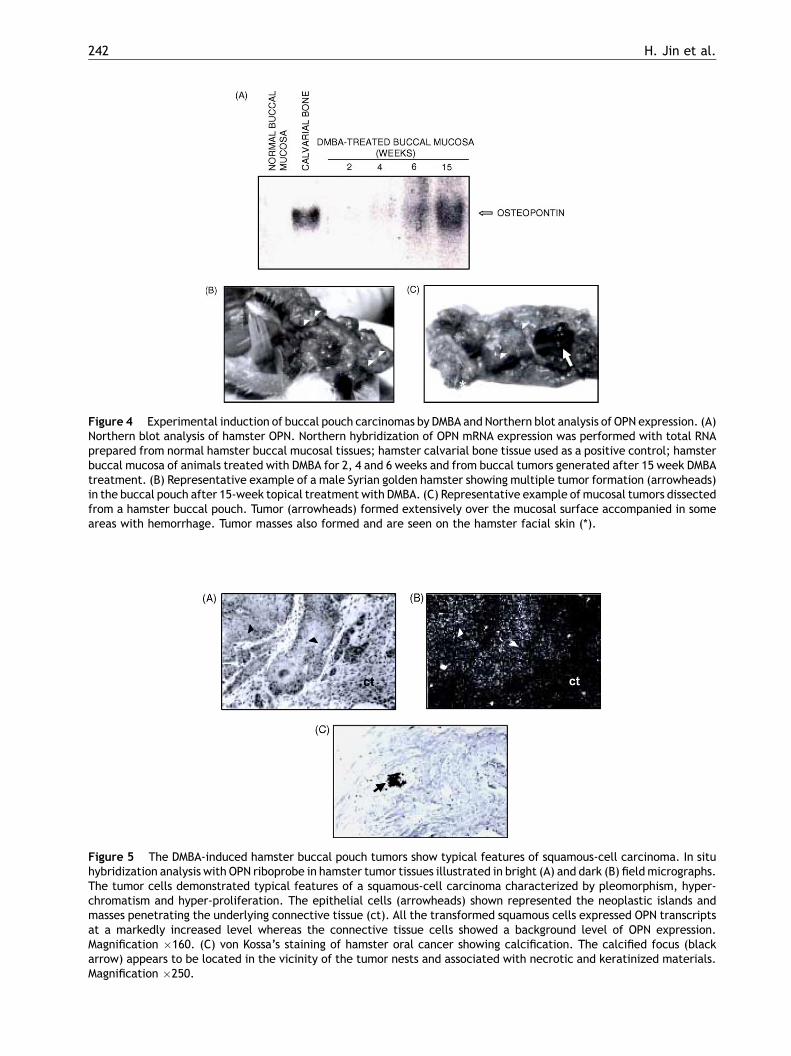
242 H. Jin et al.
Figure 4 Experimental induction of buccal pouch carcinomas by DMBA and Northern blot analysis of OPN expression. (A)Northern blot analysis of hamster OPN. Northern hybridization of OPN mRNA expression was performed with total RNAprepared from normal hamster buccal mucosal tissues; hamster calvarial bone tissue used as a positive control; hamsterbuccal mucosa of animals treated with DMBA for 2, 4 and 6 weeks and from buccal tumors generated after 15 week DMBAtreatment. (B) Representative example of a male Syrian golden hamster showing multiple tumor formation (arrowheads)in the buccal pouch after 15-week topical treatment with DMBA. (C) Representative example of mucosal tumors dissectedfrom a hamster buccal pouch. Tumor (arrowheads) formed extensively over the mucosal surface accompanied in someareas with hemorrhage. Tumor masses also formed and are seen on the hamster facial skin (*).
Figure 5 The DMBA-induced hamster buccal pouch tumors show typical features of squamous-cell carcinoma. In situhybridization analysis with OPN riboprobe in hamster tumor tissues illustrated in bright (A) and dark (B) fieldmicrographs.The tumor cells demonstrated typical features of a squamous-cell carcinoma characterized by pleomorphism, hyper-chromatism and hyper-proliferation. The epithelial cells (arrowheads) shown represented the neoplastic islands andmasses penetrating the underlying connective tissue (ct). All the transformed squamous cells expressed OPN transcriptsat a markedly increased level whereas the connective tissue cells showed a background level of OPN expression.Magnification �160. (C) von Kossa’s staining of hamster oral cancer showing calcification. The calcified focus (blackarrow) appears to be located in the vicinity of the tumor nests and associated with necrotic and keratinized materials.Magnification �250.

Hamster OPN mRNA expression and oral cancer 243
Figure 6 Expression of OPN in monocytes and neoplasic epithelial cells of hamster buccal tumor tissue. Bright (A) anddark (B) field micrographs for OPN in situ hybridization are illustrated. Expression of OPN mRNA can be seen in bothepithelium (e) andmonocytes (arrowheads) by in situ hybridization. Magnification�200. (C and D) Photomicrographs ofin situ hybridization of OPN of a different area of the hamster buccal pouch tumor tissue. Malignant tumor epithelialcells (e) express OPN hybridization signals. Inflammatory cell infiltration can be seen in the adjacent areas. Strongsignals for OPN are observed in the mononucleated histiocytes (arrowheads). The signal intensity in individualmonocytes exceeds that expressed in malignant squamous cells (e). Bright (C) and dark (D) field micrographs areillustrated. Magnification �200. (E) Immunohistochemical staining using an antibody against keratin that showsimmunoreaction in the malignant epithelial cells (e) but not in the cells of the monocyte/macrophage lineage(arrowheads). Magnification �200.
Calcification in hamster oral cancer tissue wasdetected by von Kossa’s staining. Calcified foci wereidentified near the tumor nests and appeared to beassociated with necrotic and keratinized materials(Fig. 5C).
Similar to the expression pattern of bone sialo-protein mRNA,15 tumor masses including thosepenetrating through the basement membrane wereall reactive to the OPN riboprobe (Fig. 6A and B).Interestingly, mononucleated histiocytes alsoexpressed OPN mRNA. These monocytes scatteredamong the inflammatory cells beneath the tumorcell layers expressed high levels of OPN mRNA.Importantly, the signal intensity in individual mono-cytes exceeded that expressed in malignant squa-mous cells. Expression of OPN mRNA in bothmalignant epithelium and monocytes in a differentarea of the hamster buccal tumor tissue is alsoshown in Fig. 6C and D at higher magnification. To
rule out the possibility that the scattered mononu-cleated cells in connective tissue might containindividual invading epithelial cells, immunohisto-chemical staining with an anti-cytokeratin antibodywas performed in the serial sections (Fig. 6E). Ourresults showed a negative reaction of these cellswith the anti-keratin antibody, whereas the neo-plastic epithelium stained strongly, indicating thelack of epithelial origin of these cells. Based upontheir morphological features and location, thesecells appear to belong to themonocyte/macrophagelineage.
Discussion
As part of our studies, we have isolated the hamsterOPN cDNA and predicted the domains and motifs ofthe deduced protein sequence. Comparison of the

244 H. Jin et al.
hamster OPN sequence with that of some of itsclosest orthologues indicates high degree of con-servation in several of its domains and motifs,including the signal sequence, the D-rich domainand the RGD-binding domain. The domain SVVYGLRdescribed in human OPN for binding to a9b1 anda4b1 integrins7,8 is present in hamster, rat or mouseOPN with substitution of residues VV for amino acidsLA. Such conservative substitution is not expectedto affect integrin binding affinity, since only some ofthe last four amino acids of SVVYGLR domain havebeen shown to be required for a9b1 adhesion prop-erties.7 On the other hand, the ELVTDFPTDLPAT-containing domain described for human OPN as anadditional a4b1 binding site8 is not present in ham-ster OPN or in the sequences of other rodent ortho-logues. Based upon the domain distribution ofhamster OPN, multiple mechanisms may explainits involvement in tumor progression and malig-nancy as previously suggested for human OPN.4,5
Thus, secretion of OPN into the local tumor micro-environment may promote tumor cell adhesion andmigration by binding integrins and CD44 receptors.Through its ability to bind several extracellularmatrix proteins and metalloproteases, OPN maymodulate certain cell—extracellular matrix interac-tions as well as facilitate tumor cell infiltration andmetastasis. OPN may also regulate tumor cell phe-notype by regulating intracellular signaling path-ways through autocrine/paracrine mechanisms.OPN can also act as an adapter molecule mediatingthe attachment of cancer cells to mineralized tis-sues through its hydroxyapatite and cell-attach-ment sequences.
The pattern of expression of hamster osteopontinmRNA in normal tissues appear to be consistent withthat previously described by other investigators inother species.1,3,5 Thus, constitutive OPN expres-sion has been previously detected in osteoclasts,osteoblasts, epithelial cells from several tissues(breast, kidney and skin), nerve cells, vascularsmooth muscle cells and endothelial cells.1,3 Onthe other hand, OPN expression has been shownto be induced in T lymphocytes, epidermal cells,bone cells, macrophages and cancer cells in remo-deling processes including inflammation, ischemia-reperfusion, bone resorption and tumor progres-sion.1,3,5 Consistent with its role in tumor progres-sion, we have found that OPN mRNA is expressed insquamous-cell carcinomas induced in the hamstercheek pouch by exposure to DMBA carcinogen,whereas normal buccal pouch epithelium does notexpress significant levels of OPN mRNA. As thisexperimental model of carcinogenesis is well char-acterized and highly reproducible, it provides asystem in which the expression of OPN can be
correlated with the initiation and progression oforal carcinogenesis. In human, OPN protein has beendetected in a significant percentage of premalig-nant and malignant oral lesions, but not in normaloral epithelium.22 Indeed, OPN was localized in 52%of dysplasias and 54% of carcinomas in situ, suggest-ing that OPN expression may represent an earlyevent in tumor progression. However, in the samestudy OPN expression was also detected in 75% ofhyperplasias, suggesting that OPN expression maybe associated with benign changes not directlyrelated to malignancy. In our experimental model,we have also detected OPN mRNA expression in 4-and 6-week DMBA-treated specimens that exhibitfeatures of premalignant oral epithelium, includingthickened mucosa with a rough surface and whitishgranular appearance (data not shown). However,the highest level of OPN expression was found in15-week DMBA-treated specimens, coinciding withthe development of squamous-cell carcinomas inthe hamster buccal pouch. It is currently unknown,whether the high proportion of human premalignantoral lesions expressing OPN protein could indicate apropensity for malignant progression or could berelated to prognosis. In a number of solid neoplasms,including breast, gastric and brain tumors, OPN hasbeen shown to be a prognostic factor for tumorprogression and survival.4,5
In situ hybridization analysis of DMBA-inducedoral carcinomas indicated that hamster OPN mRNAwas highly expressed by either malignant epitheliumor macrophages intimately associated with tumorcells. The presence of OPN mRNA in macrophageswas particularly pronounced at the edge of thetumors (i.e. the tumor/stroma interface) and inareas of tumor necrosis. Similar OPN mRNA expres-sion pattern was previously described in humancarcinomas of the kidney and endometrium,23
whereas carcinomas of other tissues (i.e. breast,pancreas or lung) only exhibited significant expres-sion of OPN in infiltrated macrophages.4,5,23 Thedifferential pattern of expression of OPN in thedifferent tumor types, may lead to differentialeffects of OPN in determining their malignantpotential, since OPN produced by tumor cells andby macrophages was previously suggested to playdifferent roles.24 Thus, macrophage-derived OPNwas demonstrated to function as a chemoattrac-tant, being associated with a reduced tumor bur-den, whereas tumor-derived OPN inhibitedmacrophage function and enhanced survival oftumor cells. In fact, high levels of plasma OPN havebeen shown to correlate with the risk of tumorrelapse and death in patients with head and necksquamous-cell carcinomas.25 However, the effectsof OPN in tumor progression and metastasis in other

Hamster OPN mRNA expression and oral cancer 245
tumors may depend not only of its expression levelsand pattern expression, but also on its post-transla-tional modifications and the levels of expression ofintegrins and CD44 receptors. Thus, ovarian tumorsof low malignant potential under-express the avb3integrin, to which OPN can bind, which account forthe lack of responsiveness to malignancy-promotingeffects of OPN, even in the presence of higher levelsof OPN than in benign ovarian tissue.4,5
Several OPN-expressing oral squamous-cell carci-noma-derived cell lines, including UMSCC-1,22 SASand BSC-OF26 have been developed, and studies invitro with these cell lines may help elucidate theroles of OPN in oral transformed epithelium. Indeed,downregulation of OPN with specific antisense oli-gonucleotides in BSC-OF cells has been recentlyshown to decrease invasiveness in vitro.26 Ourdemonstration that OPN is expressed in hamsterbuccal-pouch carcinomas but not in normal buccalpouch provides a useful in vivo model which can beused to further characterize the involvement of thisprotein in development of oral cancer and to designtherapeutic approaches to ameliorate oral cancerpathogenesis by specifically downregulating OPNexpression or by blocking OPN binding propertiesspecifically.
Acknowledgments
This study was supported by a Smokeless TobaccoResearch Council grant (0472) and NIH/NIDCR grantsDE11088 and DE14537 to Dr. J. Chen.
References
1. Sodek J, Ganss B, McKee MD. Osteopontin. Crit Rev Oral BiolMed 2000;11:279—303.
2. Denhardt DT, Giachelli CM, Rittling SR. Role of osteopontin incellular signaling and toxicant injury. Annu Rev PharmacolToxicol 2001;41:723—49.
3. O’Regan A, Berman JS. Osteopontin: a key cytokine in cell-mediated and granulomatous inflammation. Int J Exp Pathol2000;81:373—90.
4. Furger KA, Menon RK, Tuck AB, Bramwell VHC, Chambers AF.The functional and clinical roles of osteopontin in cancer andmetastasis. Curr Mol Med 2001;1:621—32.
5. Rittling SR, Chambers AF. Role of osteopontin in tumourprogression. Br J Cancer 2004;90:1877—81.
6. Wai PY, Kuo PC. The role of osteopontin in tumormetastasis. JSurg Res 2004;121:228—41.
7. Yokosaki Y, Matsuura N, Sasaki T, Murakami I, Schneider H,Higashiyama S, et al. The integrin a9b1 binds to a novelrecognition sequence (SVVYGLR) in the thrombin-cleavedamino-terminal fragment of osteopontin. J Biol Chem1999;274:36328—34.
8. Bayless KJ, Davis GE. Identification of dual a4b1integrinbinding sites within a 38 amino acid domain in the N-terminal
thrombin fragment of human osteopontin. J Biol Chem2001;276:13483—9.
9. Senger DR,Wirth DF, Hynes RO. Transformedmammalian cellssecrete specific proteins and phosphoproteins. Cell 1979;16:885—93.
10. Gimenez-Conti IB, Slaga TJ. The hamster cheek pouch carci-nogenesis model. J Cell Biochem 1993;17:83—90.
11. Chang KW, Sarraj S, Lin SC, Tsai PI, Solt D. p53 expression, p53and Ha-ras mutation and telomerase activation during nitro-samine-mediated hamster pouch carcinogenesis. Carcino-genesis 2000;21:1441—51.
12. Chen YK, Hsuen SS, Lin LM. Immunohistochemical demon-stration of p73 protein in the early stages of DMBA-inducedsquamous-cell carcinogenesis in hamster buccal pouch. ArchOral Biol 2002;47:695—9.
13. Balasenthil S, Rao KS, Nagini S. Altered cytokeratin expres-sion during chemoprevention of hamster buccal pouch carci-nogenesis by S-allylcysteine. Polish J Pharmacol 2003;55:793—8.
14. Valverde P. Cloning, expression and mapping of hWW45, anovel WW domain-containing gene. Biochem Biophys ResCommun 2000;276:990—8.
15. Sasaguri K, Ganss B, Sodek J, Chen JK. Expression of bonesialoprotein in mineralized tissues of tooth and bone inbuccal-pouch carcinomas of Syrian golden hamsters. ArchOral Biol 2000;45:551—62.
16. Chen J, Jin H, Ranly DM, Sodek J, Boyan BD. Altered expres-sion of bone sialoproteins in Vitamin D-deficient rBSP2.7Luctransgenic mice. J Bone Miner Res 1999;14:221—9.
17. Chen J, Aufdemorte TB, Jiang H, Liu AR, ZhangW, Thomas HF.Neoplastic odontogenic epithelial cells express bone sialo-protein. Histochem J 1998;30:1—6.
18. Kozak M. Interpreting cDNA sequences: some insights fromstudies on translation. Mamm Genome 1996;7:563—74.
19. Chen J, McCulloch CAG, Sodek J. Bone sialoproteins in devel-oping porcine dental tissues: cellular expression and com-parison of tissue localization with osteopontin andosteonectin. Arch Oral Biol 1993;38:241—9.
20. Chen J, Singh K, Mukherjee BB, Sodek J. Developmentalexpression of osteopontin (OPN) in rat tissues: evidencefor a role of OPN in bone formation and resorption. Matrix1993;13:113—23.
21. Chen J, McKee M, Nanci A, Sodek J. Message RNA expressionand ultrastructural distribution of bone sialoprotein (BSP)and osteopontin (OPN) in fetal porcine calvarial bone. His-tochem J 1994;26:67—78.
22. Devoll RE, Li W, Woods KV, Pinero GJ, Butler WT. Osteopontin(OPN) distribution in premalignant and malignant lesions oforal epithelium and expression in cell lines derived fromsquamous cell carcinoma of the oral cavity. J Oral Pathol1999;28:97—101.
23. Brown LF, Papadopoulos-Sergiou A, Berse B, Manseau EJ,Tognazzi K, Perruzzi CA, et al. Osteopontin expression anddistribution in human carcinomas. Am J Pathol 1994;145:610—23.
24. Crawford HC, Matrisian LM, Liaw L. Distinct roles of osteo-pontin in host defense activity and tumor survival duringsquamous cell carcinoma progression in vivo. Cancer Res1998;58:5206—15.
25. Le QT, Sutphin PD, Raychaudhuri S, Yu SC, Terris DJ, Lin HS,et al. Identification of osteopontin as a prognostic plasmamarker for head and neck squamous cell carcinomas. ClinCancer Res 2003;9:59—67.
26. Muramatsu T, Shima K, Ohta K, Kizaki H, Ro Y, Kohno Y, et al.Inhibition of osteopontin expression and function in oralcancer cell lines by antisense oligonucleotides. Cancer Lett2005;217:87—95.
Copyright © 2022 FDOKUMEN










![[The role of osteopontin in cardiovascular diseases]](https://static.fdokumen.com/doc/165x107/6345a46803a48733920b74c5/the-role-of-osteopontin-in-cardiovascular-diseases.jpg)









