
CK2-defective Arabidopsis plants exhibit enhanced double-strand break repair rates and reduced...
12
CK2-defective Arabidopsis plants exhibit enhanced double-strand break repair rates and reduced survival after exposure to ionizing radiation Jordi Moreno-Romero 1,† , Laia Armengot 1 , M. Mar Marque ` s-Bueno 1 , Anne Britt 2 and M. Carmen Martı´nez 1,* 1 Departament de Bioquı´mica i Biologia Molecular, Universitat Auto ` noma de Barcelona, 08193 Bellaterra, Barcelona, Spain, and 2 Department of Plant Biology, University of California, Davis, CA 95616, USA Received 21 April 2011; revised 28 March 2012; accepted 2 April 2012; published online 11 June 2012. *For correspondence (e-mail [email protected]). † Present address: Department of Plant Biology and Forest Genetics, Uppsala BioCenter, Swedish University of Agricultural Sciences and Linnean Center for Plant Biology, PO Box 7080, SE-75007 Uppsala, Sweden. SUMMARY The multifunctional protein kinase CK2 is involved in several aspects of the DNA damage response (DDR) in mammals. To gain insight into the role of CK2 in plant genome maintenance, we studied the response to genotoxic agents of an Arabidopsis CK2 dominant-negative mutant (CK2mut plants). CK2mut plants were hypersensitive to a wide range of genotoxins that produce a variety of DNA lesions. However, they were able to activate the DDR after exposure to c irradiation, as shown by accumulation of phosphorylated histone H2AX and up-regulation of sets of radio-modulated genes. Moreover, functional assays showed that mutant plants quickly repair the DNA damage produced by genotoxins, and that they exhibit preferential use of non- conservative mechanisms, which may explain plant lethality. The chromatin of CK2mut plants was more sensitive to digestion with micrococcal nuclease, suggesting compaction changes that agreed with the transcriptional changes detected for a number of genes involved in chromatin structure. Furthermore, CK2mut plants were prone to transcriptional gene silencing release upon genotoxic stress. Our results suggest that CK2 is required in the maintenance and control of genomic stability and chromatin structure in plants, and that this process affects several functions, including the DNA damage response and DNA repair. Keywords: protein kinase CK2, Arabidopsis thaliana, DNA damage responses, comet assay, MNAse sensitivity. INTRODUCTION The integrity of DNA is continuously challenged within the cell. To counteract the severe biological consequences of DNA damage, an intricate network of genome surveillance mechanisms, often referred to as DNA damage responses (DDRs), has evolved. Most of the DDR components identified in animals and yeasts have counterparts in plants (Britt, 1996, 1999; Bray and West, 2005; Kimura and Sakaguchi, 2006). However, DNA repair pathways in animals include some components for which homologues have not been found in plants, and some DDR regulators are unique to plants (Britt, 1999; Kimura and Sakaguchi, 2006; Yoshiyama et al., 2009). The initial stages of the DDR in mammals are governed by a pair of closely related protein kinases, termed ataxia telangiectasia mutated (ATM) and ATM- and RAD3-related (ATR). ATM and ATR function as sensors of DNA damage, and control the phosphorylation of histone H2AX in regions close to damaged chromatin (Kinner et al., 2008; Mah et al., 2010). Phosphorylated H2AX (c-H2AX) mediates the forma- tion of DNA damage foci, which are large aggregates of proteins and repair factors that surround the lesion sites (Riches et al., 2008). Plants possess ATM and ATR ortho- logues (Garcia et al., 2003; Culligan et al., 2004). ATM senses double-strand breaks (DSBs), triggering a transcriptional response to ionizing radiation (IR) (Garcia et al., 2003), whereas ATR senses repair intermediates or stalled replica- tion forks and has a prominent role in the UV-induced response (Culligan et al., 2004; Yoshiyama et al., 2009; Furukawa et al., 2010). Both ATR and ATM are involved in IR-induced phosphorylation of H2AX in Arabidopsis, although ATM is responsible for the majority of focus formation in M-phase cells (Friesner et al., 2005). IR-induced DSBs are among the most harmful lesions in DNA, and must therefore be eliminated before chromosome segregation. In ª 2012 The Authors 627 The Plant Journal ª 2012 Blackwell Publishing Ltd The Plant Journal (2012) 71, 627–638 doi: 10.1111/j.1365-313X.2012.05019.x
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of CK2-defective Arabidopsis plants exhibit enhanced double-strand break repair rates and reduced...

CK2-defective Arabidopsis plants exhibit enhanceddouble-strand break repair rates and reduced survivalafter exposure to ionizing radiation
Jordi Moreno-Romero1,†, Laia Armengot1, M. Mar Marques-Bueno1, Anne Britt2 and M. Carmen Martınez1,*
1Departament de Bioquımica i Biologia Molecular, Universitat Autonoma de Barcelona, 08193 Bellaterra, Barcelona, Spain, and2Department of Plant Biology, University of California, Davis, CA 95616, USA
Received 21 April 2011; revised 28 March 2012; accepted 2 April 2012; published online 11 June 2012.
*For correspondence (e-mail [email protected]).†Present address: Department of Plant Biology and Forest Genetics, Uppsala BioCenter, Swedish University of Agricultural Sciences and Linnean Center for Plant
Biology, PO Box 7080, SE-75007 Uppsala, Sweden.
SUMMARY
The multifunctional protein kinase CK2 is involved in several aspects of the DNA damage response (DDR) in
mammals. To gain insight into the role of CK2 in plant genome maintenance, we studied the response to
genotoxic agents of an Arabidopsis CK2 dominant-negative mutant (CK2mut plants). CK2mut plants were
hypersensitive to a wide range of genotoxins that produce a variety of DNA lesions. However, they were able to
activate the DDR after exposure to c irradiation, as shown by accumulation of phosphorylated histone H2AX
and up-regulation of sets of radio-modulated genes. Moreover, functional assays showed that mutant plants
quickly repair the DNA damage produced by genotoxins, and that they exhibit preferential use of non-
conservative mechanisms, which may explain plant lethality. The chromatin of CK2mut plants was more
sensitive to digestion with micrococcal nuclease, suggesting compaction changes that agreed with the
transcriptional changes detected for a number of genes involved in chromatin structure. Furthermore, CK2mut
plants were prone to transcriptional gene silencing release upon genotoxic stress. Our results suggest that CK2
is required in the maintenance and control of genomic stability and chromatin structure in plants, and that this
process affects several functions, including the DNA damage response and DNA repair.
Keywords: protein kinase CK2, Arabidopsis thaliana, DNA damage responses, comet assay, MNAse sensitivity.
INTRODUCTION
The integrity of DNA is continuously challenged within the
cell. To counteract the severe biological consequences of
DNA damage, an intricate network of genome surveillance
mechanisms, often referred to as DNA damage responses
(DDRs), has evolved. Most of the DDR components identified
in animals and yeasts have counterparts in plants (Britt,
1996, 1999; Bray and West, 2005; Kimura and Sakaguchi,
2006). However, DNA repair pathways in animals include
some components for which homologues have not been
found in plants, and some DDR regulators are unique to
plants (Britt, 1999; Kimura and Sakaguchi, 2006; Yoshiyama
et al., 2009).
The initial stages of the DDR in mammals are governed by
a pair of closely related protein kinases, termed ataxia
telangiectasia mutated (ATM) and ATM- and RAD3-related
(ATR). ATM and ATR function as sensors of DNA damage,
and control the phosphorylation of histone H2AX in regions
close to damaged chromatin (Kinner et al., 2008; Mah et al.,
2010). Phosphorylated H2AX (c-H2AX) mediates the forma-
tion of DNA damage foci, which are large aggregates of
proteins and repair factors that surround the lesion sites
(Riches et al., 2008). Plants possess ATM and ATR ortho-
logues (Garcia et al., 2003; Culligan et al., 2004). ATM senses
double-strand breaks (DSBs), triggering a transcriptional
response to ionizing radiation (IR) (Garcia et al., 2003),
whereas ATR senses repair intermediates or stalled replica-
tion forks and has a prominent role in the UV-induced
response (Culligan et al., 2004; Yoshiyama et al., 2009;
Furukawa et al., 2010). Both ATR and ATM are involved in
IR-induced phosphorylation of H2AX in Arabidopsis,
although ATM is responsible for the majority of focus
formation in M-phase cells (Friesner et al., 2005). IR-induced
DSBs are among the most harmful lesions in DNA, and must
therefore be eliminated before chromosome segregation. In
ª 2012 The Authors 627The Plant Journal ª 2012 Blackwell Publishing Ltd
The Plant Journal (2012) 71, 627–638 doi: 10.1111/j.1365-313X.2012.05019.x

eukaryotes, DSBs can be repaired by two major pathways:
homologous recombination (HR) and non-homologous
DNA end joining (NHEJ). HR is a high-fidelity mechanism
that uses unbroken, homologous sequences to template
repair of DSBs. It is mostly used to repair DSB breaks due to
DNA replication, using sister chromatid information (Orel
et al., 2003). In contrast, NHEJ does not require significant
sequence homology for DSB repair, and is a prominent
mechanism in eukaryotes (Lieber, 2010). In plants, four
major non-homologous recombination pathways have been
described: (i) canonical non-homologous end joining
(C-NHEJ), which is Ku-dependent, (ii) alternative end joining
(A-EJ or A-NHEJ), which is more error-prone and uses
microhomologies for recombination (also called MMEJ), (iii)
the back-up pathway (B-NHEJ), which is Ku-independent
and involves Parp1, Xrcc1 and DNA ligase III, and (iv) an
unidentified pathway that confers severe genomic instability
(Charbonnel et al., 2011). A hierarchical organization of
these pathways during post-S phase has been proposed in
Arabidopsis (Charbonnel et al., 2011).
DNA damage responses are tightly linked to chromatin
structure. In plants, mutations in replication-coupled chro-
matin assembly factor or chromatin assembly factor 1 (CAF-
1) result in sensitivity to DNA damage, de-repression of
transcriptional silencing, and genome instability (Takeda
et al., 2004; Kirik et al., 2006; Schonrock et al., 2006). Chro-
matin is also a major factor in the regulation of the HR
pathway, as exemplified by different Arabidopsis mutants,
such as fas1-4, which is defective in the p150 subunit of CAF-
1, mim, which lacks a chromatin structural component
related to the structural maintenance of chromosomes
(SMC) family (Mengiste et al., 1999; Kirik et al., 2006), and
bru1, which encodes a nuclear protein of unknown function
(Takeda et al., 2004).
Protein kinase CK2 is an evolutionarily conserved Ser/Thr
kinase involved in a wide variety of cellular functions
(Issinger, 1993; Allende and Allende, 1995; Meggio and
Pinna, 2003), including the DDR (Loizou et al., 2004). Inter-
estingly, most DDR-related CK2 substrates in mammals are
proteins that facilitate access to DNA or form a scaffold to
support complexes involved in chromatin remodelling.
Examples include HP1b, a heterochromatin-binding protein
that facilitates access of DDR factors (Ayoub et al., 2008,
2009), XRCC1, a scaffold protein that recruits the machinery
for single-strand break (SSB) repair (Loizou et al., 2004;
Parsons et al., 2010) and is also involved in DSB repair in
Arabidopsis (Charbonnel et al., 2010), and MDC1, an adaptor
protein that interacts with the MRN (MRE11–RAD50–NBS1)
complex involved in DSB repair (Chapman and Jackson,
2008; Melander et al., 2008; Spycher et al., 2008).
Here we have used an inducible dominant-negative allele
of CK2 to investigate the role of this protein kinase in
damage tolerance and DNA repair in the higher plant
Arabidopsis. We demonstrate that lack of CK2 activity
confers hypersensitivity to a variety of genotoxic agents,
although CK2-defective plants displayed normal levels of IR-
induced histone H2AX phosphorylation and are able to
activate a transcriptional response. Our results suggest that
mutant plants show preferential use of non-conservative
pathways for DNA repair. Moreover, changes in chromatin
structure and de-repression of transcriptional gene silencing
suggest that CK2 is required in the maintenance of chroma-
tin structure, sustaining the viability of plant cells despite the
deleterious effects of genotoxic agents.
RESULTS
Arabidopsis plants depleted of CK2 activity are hypersensi-
tive to genotoxins
In previous studies, we demonstrated that an inducible
dominant-negative allele of CK2 (CK2mut) could be suc-
cessfully used to deplete CK2 activity in Arabidopsis plants
(Moreno-Romero et al., 2008; Marques-Bueno et al., 2011).
Here we have used the same mutant to study the involve-
ment of CK2 in plant DDRs. We performed transient induc-
tions of the transgene (typically treatment of 5-day-old
seedlings with dexamethasone for 48 h), followed by
genotoxic treatment on the 7th day. Unless otherwise indi-
cated, uninduced CK2mut plants were used as controls. Our
results show that mutant seedlings are hypersensitive to IR
at 100 Gy (Figure 1a), exhibiting anthocyanin accumulation,
cotyledon necrosis and developmental arrest. The effects
are specifically due to accumulation of CK2mut protein, as
dexamethasone treatment had no effects on c-irradiated
wild-type or atm plants. The Arabidopsis atm mutant is a
well-known IR-hypersensitive mutant that is defective in the
DDR response. Moreover, CK2mut plants were hypersensi-
tive to IR when dexamethasone treatment was performed
before or after c irradiation (Figure 1b). As Arabidopsis
mutants affected in DNA repair processes show defects in
root growth under genotoxic stress (Garcia et al., 2003;
Culligan et al., 2006), we measured daily root growth in post-
IR CK2mut seedlings. Root growth was permanently
arrested upon 80 Gy treatment in CK2mut seedlings
(Figure 1c, left; note that CK2mut roots were initially shorter,
as previously described by Moreno-Romero et al., 2008),
whereas the arrest was transient in Arabidopsis wild-type.
Moreover, root viability appeared highly compromised in
CK2mut plants (Figure 1c, right).
We have recently shown that depletion of CK2 activity
alters polar auxin transport (Marques-Bueno et al., 2011). In
order to ascertain whether the IR hypersensitivity was an
indirect effect of auxin transport impairment, we treated
CK2mut and control seedlings with either indole-3-acetic
acid (IAA) or N-1-naphtylphtalamic acid (NPA) (an inhibitor
of polar auxin transport) prior to IR exposure. Quantification
of plant fresh weight in post-IR 20-day-old plants showed
that the IR hypersensitivity of CK2mut seedlings was not
628 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

reversed by exogenous IAA (Figure S1), in contrast to other
CK2mut phenotypes previously reported by Marques-Bueno
et al. (2011). Moreover, NPA slightly affected the growth of
post-irradiated control plants, but did not have deleterious
effects. Altogether, these data led us to conclude that the
hypersensitivity phenotype shown by CK2mut plants is not
an indirect consequence of impaired auxin transport.
We also tested the sensitivity of CK2mut plantlets to UV-C
and methyl methanesulfonate (MMS). CK2mut plants were
hypersensitive to UV-C radiation, showing significant
growth inhibition at 30 000 J m)2 (Figure 1d), and to MMS
over a range of 25–100 ppm (Figure 1e).
Taken together, these results strongly suggest that CK2 is
required to successfully recover from different genotoxic
treatments in Arabidopsis. The genotoxins tested are known
to produce different kinds of lesions into DNA, and thus CK2
may act upstream of different signalling pathways necessary
for DNA repair. Alternatively, CK2 may be required at multiple
points or may play a more general role in plant survival.
For the subsequent studies, we focused on IR-induced
responses using c irradiation. IR-induced responses have
been well characterized in Arabidopsis wild-type, facilitating
their study in mutant genotypes. Moreover, c irradiation has
a DSB production ratio (number of DSBs/base pair versus
dose) that is similar in all eukaryotes (Su, 2006).
CK2 expression and activity do not significantly change
after exposure to IR
The CK2a and b subunits are encoded by two small multi-
gene families. The Arabidopsis nuclear genome contains
four genes for the catalytic subunit (a) and four genes for the
regulatory subunit (b) (Salinas et al., 2006). The IR-induced
expression changes of CK2-encoding genes were very small,
never reaching twofold, whereas TSO2 (encoding the small
subunit of ribonucleotide reductase, which was used as a
positive control) was highly over-expressed under the same
conditions (Figure S2a). We also analyzed CK2 expression
data from the AtGenExpress project, which compiles Ara-
bidopsis data from ATH1 GeneChip arrays (Kilian et al.,
2007). CK2a and b transcript levels showed no significant
changes in Arabidopsis plants (18-day-old plantlets, Col-0
ecotype) treated with 1.5 lg ml)1 bleomycin (a radiomi-
metic) plus 22 lg ml)1 mitomycin C (a cross-linking agent)
(b)
(c)
atmwt CK2mut–D
ex
+D
ex
0 200 Gy50 100
–D
ex
+γ
+D
ex
+γ
+γ
+D
ex
0
1
2
3
4
0 1 2 3 4 5 6 7 8 9
Days post IR
Daily
rootgro
wth
(mm
/day)
+Dex
–Dex
(a)–D
ex
+D
ex
(d)
–D
ex
+D
ex
(e)
0 10 000 15 000 30 000 J/m2
0 25 50 100 ppm
Figure 1. Hypersensitivity of CK2mut plants to genotoxins.
(a–c) Responses to ionizing radiation (IR). Five-day-old seedlings were treated
with dexamethasone (Dex) for 48 h and then c-irradiated.
(a) Phenotype of 21-day-old seedlings after receiving 100 Gy.
(b) Dose–response experiment. IR was applied either after dexamethasone
treatment (+Dex +c) or before dexamethasone treatment (+c +Dex).
(c) Daily root growth (left panel), and cell death detected by Evan’s blue
staining (right panel) after 80 Gy IR. Scale bar = 0.3 mm.
(d) Dose–response to UV-C. Experiments were performed as in (a).
(e) Dose–response to methanesulfonate (MMS).
Images show 14-day-old seedlings.
CK2 in DNA damage responses in Arabidopsis 629
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

(Figure S2b). Although the plant ages and genotoxins used
in the array experiments were different from those em-
ployed in our work, both set of data support the idea that
CK2 is not transcriptionally regulated in response to DNA-
damaging agents. This is consistent with data showing that
mammalian CK2 activity is regulated mainly at a post-
translational level (Filhol and Cochet, 2009). However, no
significant changes of CK2 activity were found in c-irradiated
Arabidopsis wild-type plantlets (Figure S2c).
IR-induced global changes of gene expression in CK2mut
seedlings
Plants subjected to genotoxic stress become impaired in a
wide number of cellular functions. Particularly well known is
the IR-induced transcriptional burst in Arabidopsis plants,
affecting a large number of genes (Chen et al., 2003; Nagata
et al., 2005; Culligan et al., 2006; Kim et al., 2007; Ricaud
et al., 2007). We analyzed transcript profiles of c-irradiated
mutant plants (100 Gy) using Affymetrix ATH1 chips. The
study was performed at 1.5 h post-IR, as transcript radio-
modulation in Arabidopsis occurs as an early wave after IR
and lasts approximately 3 h, with only 10% of the genes still
showing changes 5 h post-IR (Ricaud et al., 2007). The
experimental design and comparative analysis performed
are summarized in Figure 2(a). Pairwise comparisons
revealed different numbers of affected genes depending on
the variable analyzed (Figure 2a, bottom). For instance,
induction of the transgene (pairwise comparison number 2,
‘CK2mut effect’) produced the widest changes, affecting
6614 sequences (P < 0.001) out of the 22 746 sequences
present in the array (after subtracting the changes obtained
in plants transformed with the empty vector, shown in
comparison number 1). IR-induced transcriptional changes
in control plants (pairwise comparison number 3, ‘gamma
effect’) correlated well with those reported in the literature
for Arabidopsis wild-type plants (Table S1a).
Genes showing IR-induced transcriptional changes in
control and mutant seedlings were grouped into GO func-
tional categories, depicted in Figure 2(b) (‘gamma effect’
and ‘CK2mut + gamma effect’, respectively). The number of
genes within each category is shown in Table S1a. There are
four shared categories, but the number of genes within them
and/or their statistical significance differ between the two
conditions. Ninety genes were present in both datasets
(‘CK2mut + gamma effect’ and ‘gamma effect’), 89 showing
positive correlation (59 up-regulated and 30 down-regulated
in both cases) and one showing negative correlation (up-
regulated in ‘CK2mut + gamma effect’ and down-regulated
in ‘gamma effect’) (Figure 2c, left, and Table S1b). The IR-
induced fold changes in expression were lower in mutant
than control plants (see the slope of the correlation plot in
Figure 2c, right), and this was confirmed by RT-PCR for
some of these genes (Figure S3). A time-course study
showed that gene under-expression in the mutant was due
to weaker induction not induction delay (Figure 2d).
To gain insight into the nature of the IR-induced tran-
scriptional response, we further analyzed the genes specif-
ically involved in DNA repair processes. One hundred and
fifty genes have a known or predicted function in DNA repair
(http://www.uea.ac.uk/~b270/repair.htm). Some are radio-
modulated (rapidly IR-induced) (Schonrock et al., 2006;
Dohmann et al., 2008), although the majority are not clearly
induced after DNA damage (http://bar.utoronto.ca/ or http://
www.weigelworld.org/resources/microarray/AtGenExpress).
DNA repair genes with significant changes in our experi-
mental conditions (ANOVA, P value <0.001) were classified
into DNA repair pathways. Many of those genes were up- or
down-regulated in non-irradiated CK2mut seedlings,
although the fold changes were rather modest (Figure 2e
and Table S1c). In particular, several of the genes involved in
HR appeared to be transcriptionally repressed, but no major
changes were found in the genes of the NHEJ pathways.
Analysis of array data from other authors (Menges et al.,
2005) showed that genes involved in HR are cell cycle-
regulated, with peaks of expression at S phase, whereas
genes involved in NHEJ pathways show steady-state expres-
sion levels during the cell cycle (Figure S4a). We have
previously demonstrated that CK2mut gene expression
leads to cell-cycle arrest at G1/S (Moreno-Romero et al.,
2008), which agrees with the expression profile of cell
Figure 2. Transcript profiling of IR-induced responses in CK2mut seedlings.
(a) Experimental design and scatter graphs. Pairwise comparisons are numbered 1 to 4; the corresponding scatter graphs show the number of up-regulated genes
(red) and down-regulated genes (green). ANOVA P value <0.001.
(b) BiNGO gene ontology (GO) enrichment analysis of IR-induced responses in dexamethasone-treated CK2mut plants (right) and untreated CK2mut plants (left).
Output categories (P value <0.05) are represented by coloured circles: red/pink, up-regulated genes; green, down-regulated genes (bi-colour circles include both
over- and under-expressed genes). Circle sizes are proportional to the number of genes. Circle perimeters represent the results of comparing the corresponding
expression data in each condition with those in dexamethasone-treated non-irradiated CK2mut plants (‘CK2mut effect’). Unmarked perimeters mean that the sub-
categories were absent in ‘CK2mut effect’, and black, white or dashed perimeters mean relative over-expression, under-expression or both, respectively. The GO
categories were subsequently grouped into higher-order functions (pale blue areas), which are indicated in bold when present in the two conditions, in italics when
present only in ‘gamma effect’, or underlined when present only in ‘CK2mut + gamma effect’.
(c) Venn diagrams and correlation plot obtained by comparison between datasets 3 and 4.
(d) Validation of transcriptional changes by semi-quantitative RT-PCR. Values were normalized to those of the EF1A (at5g60390) gene. Values are means � SD of
three biological triplicates. a.u., arbitrary units.
(e) Fold-change analysis of DNA repair genes. Genes showing significant changes (ANOVA, P value <0.001) were classified into different DNA repair pathways and
pairwise comparisons were performed [shown in (a) as 2 (+Dex), 3 (–Dex +c) and number 4 (+Dex +c)].
630 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638
Administrator
Resaltado
Administrator
Resaltado
Administrator
Resaltado

MPG At3g12040
TagI - E. coliAt5g44680
At1g75090
At1g80850
APE1 At3g48420
RAD51 At5g20850
DMC1 At3g22880
RecA – E. coli At3g10140
BRCA1 At4g21070
At1g04020
RAD50 At2g31970
BLM At1g10930
RecQ – E. coli At4g35740
adventitious root development
Carboxilic acid metabolic process (JA biosynthetic
process)
NO biosynthetic process
IAA biosynthetic
process
Glycosinolate biosynthetic process
Vitamin E biosynthetic process
Regulation of
telomerase
activity
Translation
Removal of
superoxide radicals
DNA repair
Cell cycle
Response to stress
Nucleosome assembly
DNA metabolic process
Response to biotic stimulus
‘Gamma effect’ ‘CK2mut + Gamma effect’(b)
(c)
30
172
72
DOWN
‘Gamma
59
203
53
UP
‘Gamma
‘CK2mut +Gamma
‘CK2mut +Gamma
Empty vector–Dex
CK2mut–Dex
CK2mut–Dex
+γ
Empty vector+Dex
CK2mut+Dex
CK2mut+Dex
+γ
1
2
3 4
(a)
1
2
4
3 ‘Gamma effect’
‘CK2mut + Gamma effect’
‘Empty vector effect’
‘CK2mut effect’
(e)
Correlation plot
‘Gamma effect’ fold change
‘CK
2m
ut
+G
am
ma
effect’
fold
change
5
10
15
20
0 5 10 15 20
TSO2
RAD51PARP1
BRCA1
59 genes
UP
correlated
30 genesDOWN
correlated
(d)
TSO2
1
2
3
RAD51
1
2
3
4
PARP1
4
8
12
16
−Dex +Dex
0 15 3 6 12 0 15 3 6 12
Expre
ssio
nle
vel(a
.u.)
2
4
6
8CYCB1;1
2
4
6
8BRCA1
–1.5 0 1.5
Log10 ratio
Mismatch repair
Homologous recombination
MSH6 At4g02070
RPA1
At4g19130
At5g45400
At5g08020
At5g61000
RPA2 At2g24490
GTF2H1 At1g55610
ERCC1 At3g05210
POLE1 At1g08260
PCNA At2g29570
At1g07370
Mfd - E. coli At3g02060
Phr – E. coli At1g04400
At2g47590
Nucleotide excission repair
+D
ex
–D
ex
+γ
+D
ex
+γ
Base excissionrepair
Photorepair
nucl. pool
rad6 pathway
at2g31320
at4g02390PARP
Poly(ADP-ribose) polymerase (PARP) enzymes
At3g46940DUT
RAD6A
MMS2
UBC13
At1g23260
At1g78870
At2g02760Time after γ(h)
467
144
3420
3611
262
202
112
102
3106
3508
. .
+D
ex
–D
ex
+
+D
ex
+
γ γ
effect’
effect’
effect’effect’
CK2 in DNA damage responses in Arabidopsis 631
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638
cycle-regulated genes in mutant seedlings (array data,
Figure S4b). Therefore, transcriptional under-expression of
the HR pathway in mutant plants may be due to G1/S cell-
cycle arrest. In spite of this, our array data also show that
radio-modulated genes involved in the HR pathway (such as
RAD51 or BRCA1) were IR-induced in mutant plants (Fig-
ure 2e), indicating that the pathway may be activated in a
cell cycle-independent manner under stress conditions.
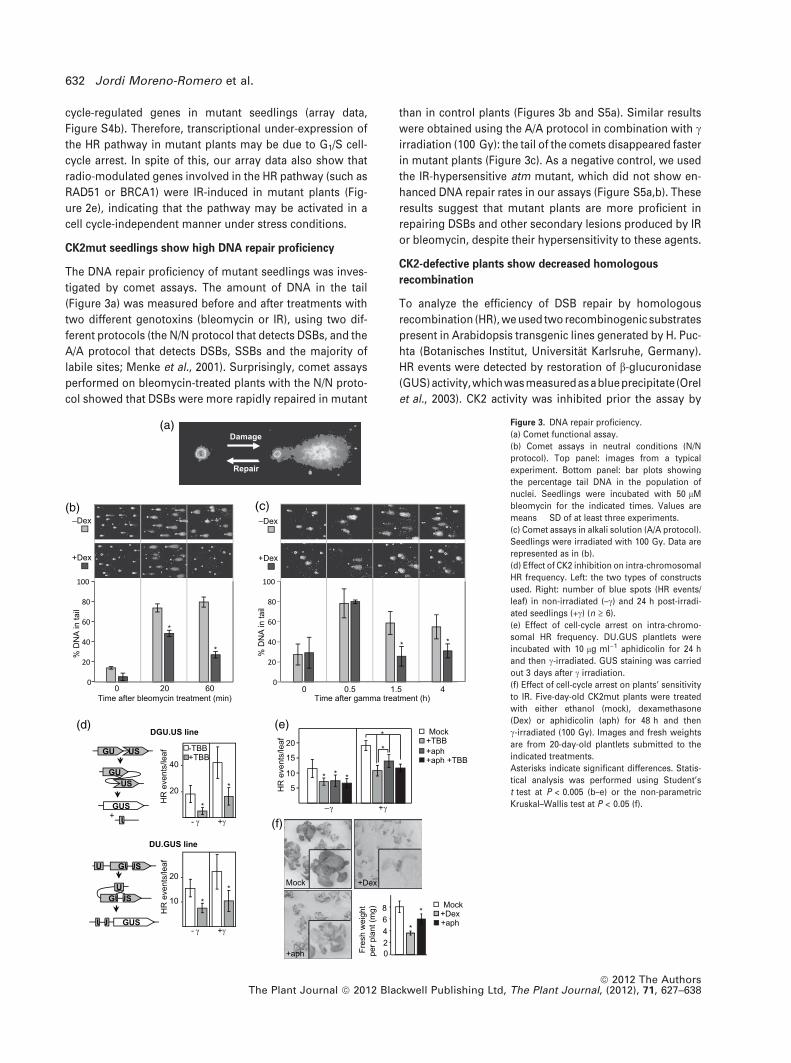
CK2mut seedlings show high DNA repair proficiency
The DNA repair proficiency of mutant seedlings was inves-
tigated by comet assays. The amount of DNA in the tail
(Figure 3a) was measured before and after treatments with
two different genotoxins (bleomycin or IR), using two dif-
ferent protocols (the N/N protocol that detects DSBs, and the
A/A protocol that detects DSBs, SSBs and the majority of
labile sites; Menke et al., 2001). Surprisingly, comet assays
performed on bleomycin-treated plants with the N/N proto-
col showed that DSBs were more rapidly repaired in mutant
than in control plants (Figures 3b and S5a). Similar results
were obtained using the A/A protocol in combination with cirradiation (100 Gy): the tail of the comets disappeared faster
in mutant plants (Figure 3c). As a negative control, we used
the IR-hypersensitive atm mutant, which did not show en-
hanced DNA repair rates in our assays (Figure S5a,b). These
results suggest that mutant plants are more proficient in
repairing DSBs and other secondary lesions produced by IR
or bleomycin, despite their hypersensitivity to these agents.
CK2-defective plants show decreased homologous
recombination
To analyze the efficiency of DSB repair by homologous
recombination (HR), we used two recombinogenic substrates
present in Arabidopsis transgenic lines generated by H. Puc-
hta (Botanisches Institut, Universitat Karlsruhe, Germany).
HR events were detected by restoration of b-glucuronidase
(GUS)activity,whichwasmeasuredasablueprecipitate (Orel
et al., 2003). CK2 activity was inhibited prior the assay by
(d)
GUS
GU
GU US
US20
40
− γ
HR
events
/leaf
+γ
−TBB+TBB
10
20
− γ +γ
HR
events
/leaf
GUUS
GUUSU
+ U
GUSUU
U
DGU.US line
DU.GUS line
Damage
Repair
(a)
(b) (c)
0
20
40
60
80
100
0 20 600
20
40
60
80
100
0 0.5 1.5 4
*
** *
+Dex
(e)
(f)
5
10
15
20
–γ +γ
Mock+TBB
+aph+aph +TBB
HR
events
/leaf
*
* **
0
2
4
6
8
Fre
sh
we
ight
per
pla
nt(m
g)
*
*
*
Mock+Dex+aph
–Dex
+Dex
–Dex
Time after bleomycin treatment (min) Time after gamma treatment (h)
%D
NA
inta
il
%D
NA
inta
il
*
*
*
*Mock +Dex
+aph
Figure 3. DNA repair proficiency.
(a) Comet functional assay.
(b) Comet assays in neutral conditions (N/N
protocol). Top panel: images from a typical
experiment. Bottom panel: bar plots showing
the percentage tail DNA in the population of
nuclei. Seedlings were incubated with 50 lM
bleomycin for the indicated times. Values are
means � SD of at least three experiments.
(c) Comet assays in alkali solution (A/A protocol).
Seedlings were irradiated with 100 Gy. Data are
represented as in (b).
(d) Effect of CK2 inhibition on intra-chromosomal
HR frequency. Left: the two types of constructs
used. Right: number of blue spots (HR events/
leaf) in non-irradiated (–c) and 24 h post-irradi-
ated seedlings (+c) (n ‡ 6).
(e) Effect of cell-cycle arrest on intra-chromo-
somal HR frequency. DU.GUS plantlets were
incubated with 10 lg ml)1 aphidicolin for 24 h
and then c-irradiated. GUS staining was carried
out 3 days after c irradiation.
(f) Effect of cell-cycle arrest on plants’ sensitivity
to IR. Five-day-old CK2mut plants were treated
with either ethanol (mock), dexamethasone
(Dex) or aphidicolin (aph) for 48 h and then
c-irradiated (100 Gy). Images and fresh weights
are from 20-day-old plantlets submitted to the
indicated treatments.
Asterisks indicate significant differences. Statis-
tical analysis was performed using Student’s
t test at P < 0.005 (b–e) or the non-parametric
Kruskal–Wallis test at P < 0.05 (f).
632 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

incubation of plants with tetrabromobenzotriazol (TBB)
(Shugar, 1994), a specific inhibitor that does not interfere with
the activity of other Arabidopsis kinases (Espunya et al.,
1999). TBB-treated seedlings exhibited fewer blue sectors
than control plants in all the conditions tested (Figure 3d),
indicating lower HR frequency. However, the number of blue
sectors significantly increased upon IR (blue spots were seen
24 h after IR), both in control and TBB-treated seedlings,
suggesting that CK2 is not required to activate the HR path-
way. In order to check whether the lower HR efficiency of TBB-
treated plants was due to an indirect effect of cell-cycle arrest
produced by CK2 inhibition, we assessed the effect of aphi-
dicolin, a drug that blocks the cell cycle at G1/S, on HR profi-
ciency. Figure 3(e) shows that, in non-irradiated seedlings,
the number of HR events was similarly decreased in the
presence of either aphidicolin or TBB (blue spots were seen
72 h after IR). Moreover, c irradiation induced significant
higherHR activation inaphidicolin-treatedplants than inTBB-
treated plants (1.85-fold versus 1.47-fold, respectively), and
the simultaneous presence of TBB and aphidicolin did not
have an additive effect on HR activation. To assess the long-
term effects of aphidicolin on plants’ sensitivity to IR, plants
were irradiated with 100 Gy and fresh weights were mea-
sured13 daysafter IR.Figure 3(f) showsthat IRaffectsgrowth
significantly more in CK2-defective plants than in aphidicolin-
treated plants. Moreover, aphidicolin-treated plants were
able to develop true leaves, whereas CK2-defective plants
were developmentally arrested. As previously demonstrated,
a 100 Gy dose is lethal for CK2mut plants, but not for aphi-
dicolin-treated plants. We conclude that, although cell-cycle
arrest may contribute to the weaker HR activity in non-irradi-
ated CK2-defectiveseedlings, additional mechanismsmay be
responsible for the impaired DDR in the same plants.
IR-induced histone H2AX phosphorylation and cyclin
CYCB1;1 expression are unaffected in CK2mut plants
One of the first molecular events induced by the DDR is
phosphorylation of histone H2AX. To determine whether
this mechanism was activated in CK2mut plants, protein
extracts from c-irradiated (100 Gy) seedlings were prepared
at 20 min after IR, and immunoblotted with an antiserum
specific to the phosphorylated form of H2AX (c-H2AX). We
detected a band of the expected molecular weight, the
intensity of which was similar in mutant and control plants
(Figure 4a). This finding was corroborated by performing a
time-course analysis of c-H2AX accumulation: both mutant
and control plants showed the maximum band intensity at
20 min after IR, rapidly decreasing afterwards (Figure 4b).
Another molecular marker of DDR activation is up-regu-
lation of mitotic cyclin CYCB1;1 (Culligan et al., 2006).
CYCB1;1 accumulation was monitored during 3 days post-
IR (100 Gy), using the translational fusion CYCB1;1::GFP
introduced into the CK2mut background. GFP fluorescence
was similar in c-irradiated mutant and control plants
(c)
4
8
12
0 4 24 48 72 0 4 24 48 72
–Dex +Dex
Root
tip
fluore
scence
(a.u
.)
0.2
0.6
1.0
0.3 hours post-IR
Band
inte
nsity
(a)
wt
132
78
45.7
18.4
32.5
7.6
H2A
X-P
0 0.3 0 0.3
– Dex + Dex
0
0.5
1.0
1.5
2.0
2.5
0 0.3 4 12
wt
(b)0 0.3 4 12 0 0.3 4 12
18.4
18.4
– Dex +Dex
CK2mut
wt
h post-IR
h post -IR
ratio
–D
ex/+
Dex
Band
inte
nsity
ratio
–D
ex/+
Dex
h post -IR
Figure 4. c-H2AX accumulation and CYCB1;1 expression in response to IR.
(a,b) Immunoblot detection of c-H2AX. Plants received 100 Gy of IR and then
were returned to growth chambers and harvested 20 min after removal from
the gamma source (a) or at the indicated times (b). Images shown are from a
typical experiment of three. A Coomassie-blue stained gel is shown in (a) (left
panel). The bottom panels in (a) and (b) show quantification of c-H2AX band
intensities, represented as )Dex/+Dex ratios, using data from three different
experiments (means � SD). The horizontal lines show the corresponding
ratios for wild-type plants.
(c) Time course of CYCB1;1-GFP expression in root tips. GFP fluorescence was
monitored and quantified (bar plots) at the indicated times. a.u., arbitrary
units.
CK2 in DNA damage responses in Arabidopsis 633
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

(Figure 4c), although the initial CYCB1;1 levels were lower in
the mutant, as reported previously (Moreno-Romero et al.,
2008).
These data indicate no significant changes in the initial
responses of CK2mut plants to IR-induced DNA damage,
confirming that their phenotype of hypersensitivity is not
due to defects in the activation of signalling pathways.
Chromatin structure in CK2mut plants
Due to the close relationship between chromatin structure
and the DNA damage response, we assessed the sensi-
tivity of the chromatin of CK2mut plants to micrococcal
nuclease (MNase). MNase is able to produce nicks in the
inter-nucleosomal DNA, and its efficiency depends on the
degree of chromatin compaction. Figure 5(a) shows that
the chromatin from CK2mut plants was more sensitive to
MNase digestion than the chromatin from control plants
(Figure 5a). This was confirmed by performing a time-
course analysis with 2.5 U ml)1 of nuclease at 4�C (Fig-
ure 5b) or 37�C (Figure S6a). In all the conditions tested,
nuclei from CK2mut plants were digested faster, leading to
accumulation of DNA bands of lower molecular weight
(Figure S6b). Several laboratories have demonstrated a
genetic link between chromatin structure, DNA repair and
transcriptional gene silencing (Takeda et al., 2004; Elmayan
et al., 2005). The array data from CK2mut seedlings
showed significant transcriptional changes in genes in-
volved in chromatin structure, such as histone modifica-
tion, heterochromatin formation or nucleosome assembly
(Figure S7a). Moreover, determination of transcript levels
of FAS1 and FAS2 (encoding two subunits of the CAF-1
complex) and of MIM by RT-PCR showed that they were
under-expressed in mutant seedlings, whereas those of
MSI1 (the third subunit of CAF-1) did not show significant
changes (Figure 5c). It has been reported that loss-of-
function fas1, fas2 and mim mutants are hypersensitive to
MMS, and that fas1 and fas2 mutations activate the
transcriptionally silent information (TSI) element (Mengiste
et al., 1999; Takeda et al., 2004). TSI repeats, concentrated
in pericentromeric chromosomal regions, are heavily
methylated and silenced in wild-type Arabidopsis, and
their expression is activated in some mutants affected in
epigenetic regulation (Steimer et al., 2000). In contrast,
CK2-defective plants show normal basal TSI transcript
levels. Upon IR, TSI levels increased 13.5-fold in CK2mut
plants and only 2.8-fold in control plants (Figure 5d). In
order to assess whether the IR-induced TSI over-expres-
sion in CK2mut plants was an indirect effect of the altered
auxin responses reported for this mutant (Marques-Bueno
et al., 2011), we measured TSI levels in post-IR plants
previously incubated with either the synthetic auxin ana-
logue 1-naphthaleneacetic acid (NAA) or the auxin trans-
port inhibitor NPA. Figure 5(e) shows that neither
compound affects TSI levels in wild-type plants, indicating
that impairment of auxin transport is likely not responsible
for the TSI release of silencing. Interestingly, incubation
with NPA increases TSI over-expression and IR sensitivity
in CK2mut plants (Figures 5e and S1, respectively).
(a) MNase (U mL–1)–Dex +Dex
0 1 5 10 20 40 0 1 5 10 20 40g
gc/g0 gt/g01.00
Conc.
0.89
0.21
0.12
0.04
0.03
1.00
0.70
0.08
0.01
0.00
0.00
(b) MNase (hours)
0 0.5 2 2.5 0 0.5 2 2.5–Dex +Dex
gTime
1.00
1.06
1.08
0.68
1.00
0.72
0.25
0.15
(d)(c)
0
2
4
6
8
Exp
ress
ion
leve
l (a.
u.)
FAS1MIM FAS2 MSI10
2
4
0 1 7Days post-IR
(e)
0
2
4
IAANPA
––
+–
–+
––
+–
–+
Exp
ress
ion
leve
l (a.
u.)
Exp
ress
ion
leve
l (a.
u.)
+Dex–Dex
+Dex–Dex
+Dex–Dex
Figure 5. Chromatin structure.
(a,b) Chromatin digestions with micrococcal nuclease (MNase). Gel electrophoresis of digested nuclei with different concentrations of MNase for 15 min at 37�C (a)
or with 2.5 U ml)1 of MNase at 4�C for the indicated times (b). The intensities of genomic bands at time 0 (g0), or after digestion with variable concentrations of
MNase (gc) or for different incubation times (gt) were determined by gel scanning.
(c) Expression levels of sequences related to chromatin-structure maintenance, measured by semi-quantitative RT-PCR.
(d,e) TSI expression. TSI transcript levels were measured by quantitative RT-PCR at the indicated times after IR (100 Gy IR) (d), or at 13 days after IR in plants
previously incubated with either 1 lM IAA or 10 lM NPA (e). Expression data were normalized to those of EF1A. Values are means � SD of three biological
replicates. a.u., arbitrary units.
634 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638
Administrator
Resaltado
Administrator
Resaltado
Administrator
Resaltado

In addition, microarray data showed that gene silencing in
either telomeric regions (data were analyzed as in Schonrock
et al., 2006, for telomeric genes present in the array),
pericentromeric regions of chromosomes 2 and 4 (Nakahig-
ashi et al., 2005), or transposable-related elements, was
maintained in CK2mut plants (Figure S7b,c,d, respectively).
We could not analyze gene expression in heterochromatic
regions in c-irradiated CK2mut plants, because the array
data were obtained at 1.5 h post-IR, which is too short a time
to detect such changes. However, the above data enabled us
to conclude that CK2mut plants have an altered, less
compacted chromatin structure, and that these plants show
de-repression of gene silencing in the presence of stressors.
DISCUSSION
Studies in yeast and mammals suggest that the protein ki-
nase CK2 is involved in the DNA damage response. This was
supported by experimental data showing that CK2 mutants
or inhibition of CK2 activity caused hypersensitivity to DNA-
damaging agents. Moreover, key proteins of the DDR
machinery have been shown to be phosphorylated by CK2
(Krohn et al., 2003; Koch et al., 2004; Loizou et al., 2004;
Cheung et al., 2005; Yamane and Kinsella, 2005). The Ara-
bidopsis mutant used in this work (CK2mut plants) exhibits
hypersensitivity to a broad range of genotoxins, such as IR,
UV-C, bleomycin and MMS. Moreover, like the much-stud-
ied Arabidopsis atm mutant (Garcia et al., 2003; Culligan
et al., 2006), CK2mut root tips do not recover from exposure
to 100 Gy IR, and, instead, terminally differentiate into root
hair-producing organs. Interestingly, hypersensitivity to IR
was detected when CK2 activity was depleted either before
or after irradiation, suggesting that CK2 participates in long-
term DNA damage responses and/or survival.
Unexpectedly, comet assays showed that CK2mut plants
exhibited enhanced proficiency in repairing IR- and bleomy-
cin-induced DSB lesions. DSB repair involves two different
pathways: homologous recombination (HR) and non-homol-
ogous end joining (NHEJ). The efficiency of the HR pathway
was checked by using two recombinant substrates, DGU.US
and DU.GUS, which exploit restoration of GUS marker gene
by two alternative mechanisms: Rad51-independent (for
DGU.US) and Rad51-dependent (for DU.GUS). CK2-defective
plants showed a lower number of HR events with either
substrate in both irradiated and non-irradiated plants. This
supports the idea that HR is less active in CK2-defective
plants, and that the rapid DSB repair activity measured by
comet assays is due to enhanced NHEJ mechanisms. Our
experiments with aphidicolin demonstrated that cell-cycle
blockage at G1/S has a clear negative impact on HR activa-
tion, but this is not sufficient to explain the lower number of
HR events in post-irradiated CK2-defective seedlings.
According to the hierarchical action of DSB repair pathways
proposed by Charbonnel et al. (2011), HR is a late-onset
pathway that repairs DSBs that were not processed by NHEJ.
As CK2mut plants show very rapid DNA repair kinetics, a
possible explanation is that most breaks are repaired by
NHEJ pathways, which involves the use of non-conservative
mechanisms and can lead to loss of genetic material and/or
accumulation of errors that may be deleterious. In particular,
the still unidentified fourth pathway postulated by Charbon-
nel et al. (2011) that confers severe genomic instability might
be favoured in CK2mut plants.
ATM-dependent phosphorylation of histone H2AX, one of
the earlier responses to DSB induction, is conserved in
plants (Friesner et al., 2005). The kinetic of H2AX phosphor-
ylation/dephosphorylation in IR-treated CK2mut plants
strongly suggests that both sensing of the stimulus and
activation of the early steps in the signalling cascade are not
affected by loss of CK2 activity. This is concordant with the
observation that radio-modulated genes are induced at the
appropriate times in the mutant, although to a lesser extent
than in wild-type plants. According to some authors, inhi-
bition of CK2 activity in mammals decreases the levels of c-
H2AX and the number of repair foci (Ayoub et al., 2008).
However, other authors have recently reported that CK2
inhibition only delays c-H2AX foci removal, reducing sur-
vival of irradiated mammal cells (Zwicker et al., 2011). In
either case, CK2 in mammals appears to be involved in
modulating c-H2AX levels, whereas our results suggest that
this is not the case in Arabidopsis. In mammals, CK2-driven
phosphorylation of MDC1 is essential for accumulation and
retention of the MRN complex and contributes to the
propagation of c-H2AX (van Attikum and Gasser, 2009).
Although homologous genes for the MRN complex have
been cloned in Arabidopsis and shown to play an important
role in DNA repair and meiotic recombination (Waterworth
et al., 2007), no plant homologues for MDC1 have been
identified so far in plants. Therefore, animals and plants may
differ mechanistically with regard to this process.
Chromatin dynamics is linked to DNA repair: minutes after
damage, important changes in chromatin compaction occur,
which are facilitated by histone modifications and chromatin
remodelling (van Attikum and Gasser, 2005; Groth et al.,
2007). In plants, many of the Arabidopsis mutants exhibiting
hypersensitivity to genotoxic treatments are also affected in
chromatin structure, such as fas1 and fas2 (Kirik et al., 2006),
bru1 (Takeda et al., 2004), mre11 (Bundock and Hooykaas,
2002), mim (Mengiste et al., 1999), and npr1 and npr2 (Zhu
et al., 2006). Some of these mutants also show de-repres-
sion of heterochromatic genes (Elmayan et al., 2005). The
chromatin of CK2-defective plants was hypersensitive to
MNase digestion, indicating either less compaction or
structural disorganization. Moreover, the expression of
genes such as FAS1 or MIM (which are involved in chroma-
tin remodelling) was decreased, supporting the idea that
CK2 activity regulates components directly involved in
chromatin structure. However, transcriptional gene silenc-
ing was maintained in the heterochromatic structures ana-
CK2 in DNA damage responses in Arabidopsis 635
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

lyzed (TSI, transposons and pericentromeric/telomeric re-
gions), as late as 2 days after dexamethasone treatment,
but, interestingly, TSI transcript levels increased dramati-
cally when CK2mut seedlings were subjected to genotoxic
stress. In this sense, CK2mut plants behaved similarly to
npr1 and npr2 mutants, which show normal basal TSI levels
but over-expression of this locus after genotoxic treatments.
NPR1 and NPR2 encode histone chaperones that act in the
maintenance of dynamic chromatin (Zhu et al., 2006).
In conclusion, our results show that CK2-defective Arabid-
opsis plants are hypersensitive to a wide range of genotoxic
stress, suggesting that, as occurs in mammals, CK2 modu-
lates the DDR in plants. CK2-defective plants are able to
activate the DDR machinery, but preferentially use non-
conservative mechanisms for DNA repair, which may explain
plant lethality. The enhanced activity of NHEJ pathways may
be facilitated in these plants because, in contrast to the HR
pathway, expression of genes involved in NHEJ pathways is
not cell cycle-dependent, and thus is unaffected by depletion
of CK2 activity. Moreover, DNA damage sites may be more
accessible to repair factors in CK2mut plants due to their less
compacted chromatin, contributing to and amplifying the
rapid use of NHEJ pathways instead of HR. In this scenario,
CK2, by virtue of its dual activity as a regulator of chromatin
remodelling and cell-cycle progression, plays a role in
protecting cells from IR by avoiding extensive use of error-
prone DNA repair mechanisms that, although quickly repair-
ing the DNA, have deleterious effects.
EXPERIMENTAL PROCEDURES
Plant material
Growth of plants, in vitro germination and culture, and generation ofCK2mut and CYCB1;1::GFP x CK2mut transgenic plants have beendescribed by Moreno-Romero et al. (2008). Seeds containing therecombinogenic substrates DGU.US and DU.GUS were kindly pro-vided by Dr H. Putcha (Botanisches Institut, Universitat Karlsruhe,Germany). Expression of the CK2mut transgene was induced with1 lM dexamethasone dissolved in ethanol. Root lengths weremeasured on scanned plant images using a GS-700 imaging den-sitometer (Bio-Rad, Hercules, CA, USA) and IMAGEJ software (http://rsb.info.nih.gov/ij). Viability of root tip cells was tested by incubationwith Evans blue (0.25% w/v) (Sigma-Aldrich, St Louis, MO, USA) for1 h as described by Gaff and Okong’o-Ogola (1971). CYCB1;1::GFPexpression was monitored with a DMRB microscope (Leica Micro-systems GmbH, Wetzlar, Germany) equipped with a Leica DC200digital camera. Quantification of GFP fluorescence was performedusing LEICA Q500MC software, in non-saturating conditions. IAA,NAA and NPA (Sigma-Aldrich) were dissolved in ethanol at the gi-ven concentrations and added to Petri dishes. Aphidicolin (Sigma-Aldrich) was dissolved in dimethylsulfoxide and used at 10 lg ml)1
for the indicated times.
Genotoxic treatments
Unless otherwise indicated, genotoxic treatments were performedon 7-day-old seedlings grown on MS plates or liquid medium.Ionizing radiation (IR) was provided by a 137caesium source gener-ated by an IBL 437C irradiator (CIS Bio International, Bagnols/Ceze,
France) at a dose rate of 5.7–5.5 Gy min)1. UV-C radiation wasprovided by a Stratalinker UV crosslinker (Stratagene, AgilentTechnologies, Inc., Santa Clara, CA, USA). Bleomycin treatmentswere carried out for 1 h on 7-day-old seedlings grown on plates.MMS treatments were carried out for 7 days on 6-day-old seedlingsgrown in MS liquid medium and previously incubated with dexa-methasone or ethanol for 24 h.
RT-PCR assays
RNA extraction and quantitative RT-PCR assays were performed asdescribed by Moreno-Romero et al. (2008). For semi-quantitativeRT-PCR, the linear range of PCR product synthesis was establishedfor each primer pair, and the number of cycles was chosenaccordingly. Primer sequences and annealing temperature areshown in Table S2.
Hybridization and analysis of ATH1 Affymetrix arrays
Total RNA was isolated from frozen tissue samples using Trizol(Invitrogen, Carlsbad, CA, USA), and later purified using QiagenRNeasy Plant Mini Kit columns (Qiagen, Valencia, CA, USA). Threeindependent RNA preparations were made from pooled samples foreach of the conditions and lines. Microarray hybridizations (Gene-Chip� ATH1 from Affymetrix (Affymetrix, Inc., Santa Clara, CA,USA), with 22 810 sequences) were performed by the genomicfacilities of Progenika Biopharma (Derio, Spain) using the method-ology and equipment recommended by Affymetrix Inc. The com-plete array data for plants transformed with the empty vector (withand without dexamethasone) have been submitted to the Notting-ham Arabidopsis Stock Centre database (reference NASCARRAYS-642; http://affymetrix.arabidopsis.info/).
Bioinformatic analysis
Data analysis, including statistical analysis (ANOVA, P value <0.001),fold change values, plot representations, clustering and Venn dia-grams, was performed using ROSETTA RESOLVER software (RosettaBiosoftware, Seattle, WA, USA). Gene classification into functionalcategories was performed using BiNGO gene enrichment analysis(Maere et al., 2005). Microarray expression data of CK2 subunitswere obtained from the AtGenExpress project (http://www.arabid-opsis.org), data for the met1 mutant were obtained from Mathieuet al. (2007), and the list of transposable elements was obtainedfrom Slotkin et al. (2009).
CK2 activity assay, extraction of histones and
immunoblotting
CK2 kinase activity assays were performed as described by Moreno-Romero et al. (2008). The assays were done in triplicate, and back-ground signals (values obtained after adding 1 lM TBB to a reactiontest) were subtracted. Extraction of histones and immunoblottingwere performed as described by Friesner et al. (2005).
Comet assay
Comet assays were performed as described by Menke et al. (2001),with some modifications. Briefly, frozen material was chopped witha fresh razor blade on ice and under dim light in PBS containing10 mM EDTA. The suspensions of nuclei were filtered through80 lm mesh Sefar Nytal (Sefar AG, Heiden Switzerland) and mixedwith low-melting-point agarose (Bio-Rad) at 37�C, to a final con-centration of 0.38% w/v (alkali, A/A protocol) or 0.19% w/v (neutral,N/N protocol). Drops of 70 ll were loaded onto microscope slidespre-coated with 1% agarose, and covered with 20 · 20 mm coverslips, then incubated on ice for 5 min. In the A/A protocol, the slides
636 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

were incubated for 5 min in high-alkali buffer (0.3 M NaOH, 5 mM
EDTA, pH 13.5), prior to electrophoresis at 4�C in the same buffer(0.7 V cm)1), followed by a short neutralization step of 5 min in100 mM Tris/HCl, pH 7.5. In the N/N protocol, the slides were incu-bated for 1 h in high-salt buffer (2.5 M NaCl2, 10 mM Tris/HCl, pH 7.5,100 mM EDTA) on ice, followed by an equilibration step in 1· TBE(90 mM Tris/borate, 2 mM EDTA, pH 8.4) (3 · 5 min) prior to elec-trophoresis (1 V cm)1) at room temperature in TBE buffer. Theslides were then dehydrated in 75% v/v ethanol (5 min), followed by96% v/v ethanol (5 min), and air-dried. Dry agarose gels wererehydrated for 5 min in water and stained with 4¢,6-diamidino-2-phenylindole (DAPI) (1–5 lg ml)1). Images were captured using aLeica DRMB fluorescence microscope and quantified with COMET-
SCORE software, version 1.5 (Tritek Corp., Sumerduck, VA, USA). Atleast 100 nuclei were scored from two different comet gels for eachexperiment.
HR assay
Fourteen-day-old DGU.US or DU.GUS plantlets were treated witheither 10 lM TBB for 16 h or 10 lg ml)1 aphidicolin, or both, for 24 hin liquid medium, and then c-irradiated with 100 Gy and returned tothe growth chamber for the indicated times in each experiment.Aphidicolin was maintained in the growth medium for the wholetime, while TBB was removed after IR. The frequency of HR eventswas visualized as the number of blue tissue sectors after GUS his-tochemical staining. GUS activity was determined using a b-glucu-ronidase reporter gene staining kit (Sigma-Aldrich). Seedlings weremounted with 50% v/v glycerol and observed with a Leica DMRBmicroscope.
Micrococcal nuclease (MNase) assay
Nuclei from 7-day-old CK2mut plantlets (with or without dexa-methasone) were isolated by grinding plant material (2 g) in liquidnitrogen, and then resuspended in 0.25 M sucrose, 60 mM KCl,15 mM MgCl2, 1 mM CaCl2, 15 mM PIPES, pH 6.8, 0.8% Triton X-100and 1 mM PMSF. The homogenate was centrifuged for 20 min at10 000 g, and the pellet resuspended in 200–500 ll of 0.25 M
sucrose, 10 mM Tris/HCl pH 8.0, 10 mM MgCl2, 1% v/v Triton X-100,5 mM 2-mercaptoethanol and 1 mM PMSF. The suspension wasplaced on the top of an Eppendorf tube containing 1.7 M sucrose,10 mM Tris/HCl, pH 8.0, 10 mM MgCl2, 0.5% v/v Triton X-100, 5 mM
b-mercaptoethanol and 1 mM PMSF, and centrifuged at 4�C for 1 hat 12 000 g. The resulting pellet was resuspended in MNase buffer(0.3 M sucrose, 20 mM Tris/HCl, pH 7.5, 3 mM CaCl2), and the DNAconcentration was determined by measuring the absorbance at260 nm. Equal amounts of nuclei were digested, and the reactionwas stopped with 50 mM EDTA and 1% SDS. Samples were incu-bated with proteinase K overnight at 37�C, electrophoresed onagarose gels and visualized by gel staining with ethidium bromide.
ACKNOWLEDGEMENTS
We thank Dr H. Puchta (Botanisches Institut, Universitat Karlsruhe,Germany) for providing the DGU.US and DU.US reporter lines. Thiswork was supported by grants BFU2007-60569, BFU2010-15090 andConsolider Ingenio 2010 CSD2007-00036 from the Ministerio de Ed-ucacion y Ciencia, (Spain) and grants 2005SGR-00112 and 2009SGR-795 from the Generalitat de Catalunya, Catalunya, Spain. J.M.-R. andL.A. were recipients of fellowships from the Universitat Autonoma deBarcelona and the Ministerio de Educacion y Ciencia (Spain),respectively. We are indebted to the Arabidopsis InformationResource (TAIR) (http://arabidopsis.org) as an invaluable source ofdata, and to the Servei de Microscopia, Unitat Tecnica de Proteccio
Radioactiva and the Laboratori d’Analisi i Fotodocumentacio (Uni-versitat Autonoma de Barcelona, Spain) for technical support.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Influence of auxin transport on IR hypersensitivity.Figure S2. Expression and activity of CK2 after genotoxic treat-ments.Figure S3. Validation of array data.Figure S4. DNA damage response and the cell cycle.Figure S5. DSB repair proficiency.Figure S6. MNase sensitivity.Figure S7. Transcript profiling of chromatin-related GO categoriesand heterochromatic silenced regions.Table S1. Global changes of gene expression in response to IR.Table S2. Primers used for real-time RT-PCR.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Allende, J.E. and Allende, C.C. (1995) Protein kinases. 4. Protein kinase CK2: an
enzyme with multiple substrates and a puzzling regulation. FASEB J. 9, 313–323.
van Attikum, H. and Gasser, S.M. (2005) ATP-dependent chromatin remodeling
and DNA double-strand break repair. Cell Cycle, 4, 1011–1014.
van Attikum, H. and Gasser, S.M. (2009) Crosstalk between histone
modifications during the DNA damage response. Trends Cell Biol. 19, 207–
217.
Ayoub, N., Jeyasekharan, A.D., Bernal, J.A. and Venkitaraman, A.R. (2008) HP1-bmobilization promotes chromatin changes that initiate the DNA damage re-
sponse. Nature, 453, 682–686.
Ayoub, N., Jeyasekharan, A.D. and Venkitaraman, A.R. (2009) Mobilization and
recruitment of HP1b: a bimodal response to DNA breakage. Cell Cycle, 8, 2945–
2950.
Bray, C.M. and West, C.E. (2005) DNA repair mechanisms in plants: crucial sensors
and effectors for the maintenance of genome integrity. New Phytol. 168, 511–
528.
Britt, A.B. (1996) DNA damage and repair in plants. Annu. Rev. Plant Physiol. Plant
Mol. Biol. 47, 75–100.
Britt, A.B. (1999) Molecular genetics of DNA repair in higher plants. Trends Plant
Sci. 4, 20–25.
Bundock, P. and Hooykaas, P. (2002) Severe developmental defects, hypersensi-
tivity to DNA-damaging agents, and lengthened telomeres in Arabidopsis
MRE11 mutants. Plant Cell, 14, 2451–2462.
Chapman, J.R. and Jackson, S.P. (2008) Phospho-dependent interactions between
NBS1 and MDC1 mediate chromatin retention of the MRN complex at sites of
DNA damage. EMBO Rep. 9, 795–801.
Charbonnel, C., Gallego, M.E. and White, C.I. (2010) Xrcc1-dependent and ku-
dependent DNA double-strand break repair kinetics in Arabidopsis plants. Plant
J. 64, 280–290.
Charbonnel, C., Allain, E., Gallego, M.E. and White, C.I. (2011) Kinetic analysis of
DNA double-strand break repair pathways in Arabidopsis. DNA Repair (Amst)
10, 611–619.
Chen, I.P., Haehnel, U., Altschmied, L., Schubert, I. and Puchta, H. (2003) The
transcriptional response of Arabidopsis to genotoxic stress – a high-density
colony array study (HDCA). Plant J. 35, 771–786.
Cheung, W.L., Turner, F.B., Krishnamoorthy, T., Wolner, B., Ahn, S.H., Foley, M.,
Dorsey, J.A., Peterson, C.L., Berger, S.L. and Allis, C.D. (2005) Phosphorylation
of histone H4 serine 1 during DNA damage requires casein kinase II in S. ce-
revisiae. Curr. Biol. 15, 656–660.
Culligan, K., Tissier, A. and Britt, A. (2004) ATR regulates a G2-phase cell-cycle
checkpoint in Arabidopsis thaliana. Plant Cell, 16, 1091–1104.
Culligan, K.M., Robertson, C.E., Foreman, J., Doerner, P. and Britt, A.B. (2006) ATR
and ATM play both distinct and additive roles in response to ionizing radiation.
Plant J. 48, 947–961.
CK2 in DNA damage responses in Arabidopsis 637
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638

Dohmann, E.M., Levesque, M.P., De Veylder, L., Reichardt, I., Jurgens, G., Schmid,
M. and Schwechheimer, C. (2008) The Arabidopsis COP9 signalosome is
essential for G2 phase progression and genomic stability. Development, 135,
2013–2022.
Elmayan, T., Proux, F. and Vaucheret, H. (2005) Arabidopsis RPA2: a genetic link
among transcriptional gene silencing, DNA repair, and DNA replication. Curr.
Biol. 15, 1919–1925.
Espunya, M.C., Combettes, B., Dot, J., Chaubet-Gigot, N. and Martinez, M.C.
(1999) Cell-cycle modulation of CK2 activity in tobacco BY-2 cells. Plant J. 19,
655–666.
Filhol, O. and Cochet, C. (2009) Protein kinase CK2 in health and disease. Cellular
functions of protein kinase CK2: a dynamic affair. Cell. Mol. Life Sci. 66, 1830–
1839.
Friesner, J.D., Liu, B., Culligan, K. and Britt, A.B. (2005) Ionizing radiation-dependent c-
H2AX focus formation requires ataxia telangiectasia mutated and ataxia telangi-
ectasia mutated and Rad3-related. Mol. Biol. Cell 16, 2566–2576.
Furukawa, T., Curtis, M.J., Tominey, C.M., Duong, Y.H., Wilcox, B.W., Aggoune, D.,
Hays, J.B. and Britt, A.B. (2010) A shared DNA-damage-response pathway for
induction of stem-cell death by UVB and by gamma irradiation. DNA Repair
(Amst) 9, 940–948.
Gaff, D.F. and Okong’o-Ogola, O. (1971) The use of non-permeating pigments for
testing survival of cells. J. Exp. Bot. 22, 756–758.
Garcia, V., Bruchet, H., Camescasse, D., Granier, F., Bouchez, D. and Tissier, A.
(2003) AtATM is essential for meiosis and the somatic response to DNA damage
in plants. Plant Cell, 15, 119–132.
Groth, A., Rocha, W., Verreault, A. and Almouzni, G. (2007) Chromatin challenges
during DNA replication and repair. Cell, 128, 721–733.
Issinger, O.G. (1993) Casein kinases: pleiotropic mediators of cellular regulation.
Pharmacol. Ther. 59, 1–30.
Kilian, J., Whitehead, D., Horak, J., Wanke, D., Weinl, S., Batistic, O., D’Angelo, C.,
Bornberg-Bauer, E., Kudla, J. and Harter, K. (2007) The AtGenExpressglobal stress
expression data set: protocols, evaluation and model data analysis of UV-B light,
drought and cold stress responses. Plant J. 50, 347–363.
Kim, J.H., Moon, Y.R., Kim, J.S., Oh, M.H., Lee, J.W. and Chung, B.Y. (2007)
Transcriptomic profile of Arabidopsis rosette leaves during the reproductive
stage after exposure to ionizing radiation. Radiat. Res. 168, 267–280.
Kimura, S. and Sakaguchi, K. (2006) DNA repair in plants. Chem. Rev. 106, 753–766.
Kinner, A., Wu, W., Staudt, C. and Iliakis, G. (2008) c-H2AX in recognition and
signaling of DNA double-strand breaks in the context of chromatin. Nucleic
Acids Res. 36, 5678–5694.
Kirik, A., Pecinka, A., Wendeler, E. and Reiss, B. (2006) The chromatin assembly
factor subunit FASCIATA1 is involved in homologous recombination in plants.
Plant Cell, 18, 2431–2442.
Koch, C.A., Agyei, R., Galicia, S., Metalnikov, P., Donnell, P., Starostine, A.,
Weinfeld, M. and Dronachar, D. (2004) Xrcc4 physically links DNA end pro-
cessing by polynucleotide kinase to DNA ligation by DNA ligase IV. EMBO J. 23,
3874–3885.
Krohn, N.M., Stemmer, C., Fojan, P., Grimm, R. and Grasser, K.D. (2003) Protein
kinase CK2 phosphorylates the high mobility group domain protein SSRP1,
inducing the recognition of UV-damaged DNA. J. Biol. Chem. 278, 12710–12715.
Lieber, M.R. (2010) The mechanism of double-strand DNA break repair by the non-
homologous DNA end-joining pathway. Annu. Rev. Biochem. 79, 181–211.
Loizou, J.I., El-Khamisy, S.F., Zlatanou, A., Moore, D.J., Chan, D.W., Qin, J., Sarno,
S., Meggio, F., Pinna, L.A. and Caldecott, K.W. (2004) The protein kinase CK2
facilitates repair of chromosomal DNA single-strand breaks. Cell, 117, 17–28.
Maere, S., Heymans, K. and Kuiper, M. (2005) BiNGO: a Cytoscape plugin to assess
overrepresentation of gene ontology categories in biological networks. Bioin-
formatics, 21, 3448–3449.
Mah, L.J., El-Osta, A. and Karagiannis, T.C. (2010) cH2AX: a sensitive molecular
marker of DNA damage and repair. Leukemia, 24, 679–686.
Marques-Bueno, M.M., Moreno-Romero, J., Abas, L., De Michele, R. and Martinez,
M.C. (2011) A dominant negative mutant of protein kinase CK2 exhibits altered
auxin responses in Arabidopsis. Plant J. 67, 169–180.
Mathieu, O., Reinders, J., Caikovski, M., Smathajitt, C. and Paszkowski, J. (2007)
Transgenerational stability of the Arabidopsis epigenome is coordinated by CG
methylation. Cell, 130, 851–862.
Meggio, F. and Pinna, L.A. (2003) One-thousand-and-one substrates of protein
kinase CK2? FASEB J. 17, 349–368.
Melander, F., Bekker-Jensen, S., Falck, J., Bartek, J., Mailand, N. and Lukas, J.
(2008) Phosphorylation of SDT repeats in the MDC1 N-terminus triggers
retention of NBS1 at the DNA damage-modified chromatin. J. Cell Biol. 181,
213–226.
Menges, M., de Jager, S.M., Gruissem, W. and Murray, J.A. (2005) Global
analysis of the core cell cycle regulators of Arabidopsis identifies novel
genes, reveals multiple and highly specific profiles of expression and pro-
vides a coherent model for plant cell cycle control. Plant J. 41, 546–566.
Mengiste, T., Revenkova, E., Bechtold, N. and Paszkowski, J. (1999) An SMC-like
protein is required for efficient homologous recombination in Arabidopsis.
EMBO J. 18, 4505–4512.
Menke, M., Chen, I., Angelis, K.J. and Schubert, I. (2001) DNA damage and repair in
Arabidopsis thaliana as measured by the comet assay after treatment with
different classes of genotoxins. Mutat. Res. 493, 87–93.
Moreno-Romero, J., Espunya, M.C., Platara, M., Arino, J. and Martinez, M.C.
(2008) A role for protein kinase CK2 in plant development: evidence obtained
using a dominant-negative mutant. Plant J. 55, 118–130.
Nagata, T., Yamada, H., Du, Z., Todoriki, S. and Kikuchi, S. (2005) Microarray
analysis of genes that respond to c-irradiation in Arabidopsis. J. Agric. Food
Chem. 53, 1022–1030.
Nakahigashi, K., Jasencakova, Z., Schubert, I. and Goto, K. (2005) The Arabidopsis
heterochromatin protein1 homolog (TERMINAL FLOWER2) silences genes
within the euchromatic region but not genes positioned in heterochromatin.
Plant Cell Physiol. 46, 1747–1756.
Orel, N., Kyryk, A. and Puchta, H. (2003) Different pathways of homologous
recombination are used for the repair of double-strand breaks within tandemly
arranged sequences in the plant genome. Plant J. 35, 604–612.
Parsons, J.L., Dianova, I.I., Finch, D., Tait, P.S., Strom, C.E., Helleday, T. and Dia-
nov, G.L. (2010) XRCC1 phosphorylation by CK2 is required for its stability and
efficient DNA repair. DNA Repair (Amst) 9, 835–841.
Ricaud, L., Proux, C., Renou, J.P., Pichon, O., Fochesato, S., Ortet, P. and Montane,
M.H. (2007) ATM-mediated transcriptional and developmental responses to c-
rays in Arabidopsis. PLoS One, 2, e430.
Riches, L.C., Lynch, A.M. and Gooderham, N.J. (2008) Early events in the mammalian
response to DNA double-strand breaks. Mutagenesis, 23, 331–339.
Salinas, P., Fuentes, D., Vidal, E., Jordana, X.,Echeverria,M.and Holuigue, L. (2006) An
extensive survey of CK2 a and b subunits in Arabidopsis: multiple isoforms exhibit
differential subcellular localization. Plant Cell Physiol. 47, 1295–1308.
Schonrock, N., Exner, V., Probst, A., Gruissem, W. and Hennig, L. (2006) Functional
genomic analysis of CAF-1 mutants in Arabidopsis thaliana. J. Biol. Chem. 281,
9560–9568.
Shugar, D. (1994) Development of inhibitors of protein kinases CKI and CKII and
some related aspects, including donor and acceptor specificities and viral
protein kinases. Cell. Mol. Biol. Res. 40, 411–419.
Slotkin, R.K., Vaughn, M., Borges, F., Tanurdzic, M., Becker, J.D., Feijo, J.A. and
Martienssen, R.A. (2009) Epigenetic reprogramming and small RNA silencing of
transposable elements in pollen. Cell, 136, 461–472.
Spycher, C., Miller, E.S., Townsend, K., Pavic, L., Morrice, N.A., Janscak, P.,
Stewart, G.S. and Stucki, M. (2008) Constitutive phosphorylation of MDC1
physically links the MRE11–RAD50–NBS1 complex to damaged chromatin. J.
Cell Biol. 181, 227–240.
Steimer, A., Amedeo, P., Afsar, K., Fransz, P., Mittelsten Scheid, O. and Pasz-
kowski, J. (2000) Endogenous targets of transcriptional gene silencing in Ara-
bidopsis. Plant Cell, 12, 1165–1178.
Su, T.T. (2006) Cellular responses to DNA damage: one signal, multiple choices.
Annu. Rev. Genet. 40, 187–208.
Takeda, S., Tadele, Z., Hofmann, I. et al. (2004) BRU1, a novel link between re-
sponses to DNA damage and epigenetic gene silencing in Arabidopsis. Genes
Dev. 18, 782–793.
Waterworth, W.M., Altun, C., Armstrong, S.J., Roberts, N., Dean, P.J., Young, K.,
Weil, C.F., Bray, C.M. and West, C.E. (2007) NBS1 is involved in DNA repair and
plays a synergistic role with ATM in mediating meiotic homologous recombi-
nation in plants. Plant J. 52, 41–52.
Yamane, K. and Kinsella, T.J. (2005) CK2 inhibits apoptosis and changes its cellular
localization following ionizing radiation. Cancer Res. 65, 4362–4367.
Yoshiyama, K., Conklin, P.A., Huefner, N.D. and Britt, A.B. (2009) Suppressor of
gamma response 1 (SOG1) encodes a putative transcription factor governing
multiple responses to DNA damage. Proc. Natl Acad. Sci. USA, 106, 12843–
12848.
Zhu, Y., Dong, A., Meyer, D., Pichon, O., Renou, J.P., Cao, K. and Shen, W.H. (2006)
Arabidopsis NRP1 and NRP2 encode histone chaperones and are required for
maintaining postembryonic root growth. Plant Cell, 18, 2879–2892.
Zwicker, F., Ebert, M., Huber, P.E., Debus, J. and Weber, K.J. (2011) A specific
inhibitor of protein kinase CK2 delays c-H2Ax foci removal and reduces clo-
nogenic survival of irradiated mammalian cells. Radiat. Oncol. 6, 15.
638 Jordi Moreno-Romero et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 627–638




















