
Deciphering the regulatory network controlling flavonoid ...

www.landesbioscience.com Cell Cycle 2963
Cell Cycle 9:15, 2963-2972; August 1, 2010; © 2010 Landes Bioscience
extrA view extrA view
Key words: ChIP-seq, ChIP-chip, chromatin, DSB repair, γH2AX, AsiSI
Submitted: 05/14/10
Accepted: 05/20/10
Previously published online: www.landesbioscience.com/journals/cc/article/12412
DOI: 10.4161/cc.9.15.12412
*Correspondence to: Gaëlle Legube; Email: [email protected]
DNA double strand breaks (DSBs) are among the most deleterious forms
of lesions and deciphering the details of the chromatin landscape induced around DSBs represents a great challenge for molecular biologists.
Chromatin Immunoprecipitation, followed by microarray hybridisa-tion (ChIP-chip) or high-throughput sequencing (ChIP-seq), are powerful techniques that provide high-resolution maps of protein-genome interactions. However, applying these techniques to study chromatin changes induced around DSBs was previously hindered due to a lack of suitable DSB induction techniques.
We have recently developed an experimental system utilizing a restric-tion enzyme fused to a modified oes-trogen receptor ligand binding domain (AsiSI-ER), which generates multiple, sequence-specific and unambiguously positioned DSBs across the genome upon induction with 4-hydroxytamoxi-fen (4OHT).1 Cell lines expressing this construct represent a powerful tool to study specific chromatin changes dur-ing DSB repair, enabling high-resolu-tion profiling of DNA repair complexes and chromatin modifications induced around DSBs. Using this system, we have recently produced the first map of γH2AX, a DSB-induced chroma-tin modification, on two human chro-mosomes and have investigated its spreading properties.1 Here we provide additional data characterizing the cell lines, present a genome-wide profile of γH2AX obtained by ChIP-seq, and dis-cuss the potential of our system towards
Deciphering the chromatin landscape induced around DNA double strand breaks
Laurent Massip,1 Pierre Caron,1 Jason S. Iacovoni,2 Didier Trouche1 and Gaëlle Legube1,*1LBCMCP; CNRS and University of Toulouse; 2Bioinformatic Plateau IFR150; INSERM and University of Toulouse; Toulouse, France
investigations of previously uncharacter-ized aspects of DSB repair.
DSB-induced Chromatin Changes
Maintaining genome integrity is of crucial importance for multicellular organisms, as illustrated by the variety of human dis-eases associated with DNA repair defects. In eukaryotes, DNA associates with vari-ous proteins, mainly histones, to form a condensed structure known as chromatin, which tightly regulates its accessibility. Over the past few years, it has become evi-dent that chromatin, being the real sub-strate for all DNA related processes, plays a decisive role in DSB repair. The first hint for such a critical role came from the iden-tification of a DSB specific histone modi-fication: the phosphorylation of the H2A histone variant H2AX on S139, known as γH2AX,2,3 by PI3K kinase family mem-bers (ATM/ATR/DNAPK). γH2AX (or its yeast counterpart γH2A) is rapidly induced (within a few minutes) over large regions surrounding the break (reviewed in ref. 4), and is necessary for the accumu-lation and retention of DNA repair fac-tors into repair foci.5,6 As a consequence, H2AX null mice exhibit tumor suscepti-bility, sterility and immunodeficiency, a range of phenotypes consistent with DSB coping defects.7-9
An exponentially growing number of chromatin modifications have since been shown to be involved in DSB repair (reviewed in refs. 10 and 11). Among these, the heterochromatin proteins HP1β and Kap1, are rapidly phosphorylated, leading to their transient eviction from chromatin, a mechanism proposed to promote DSB

2964 Cell Cycle volume 9 issue 15
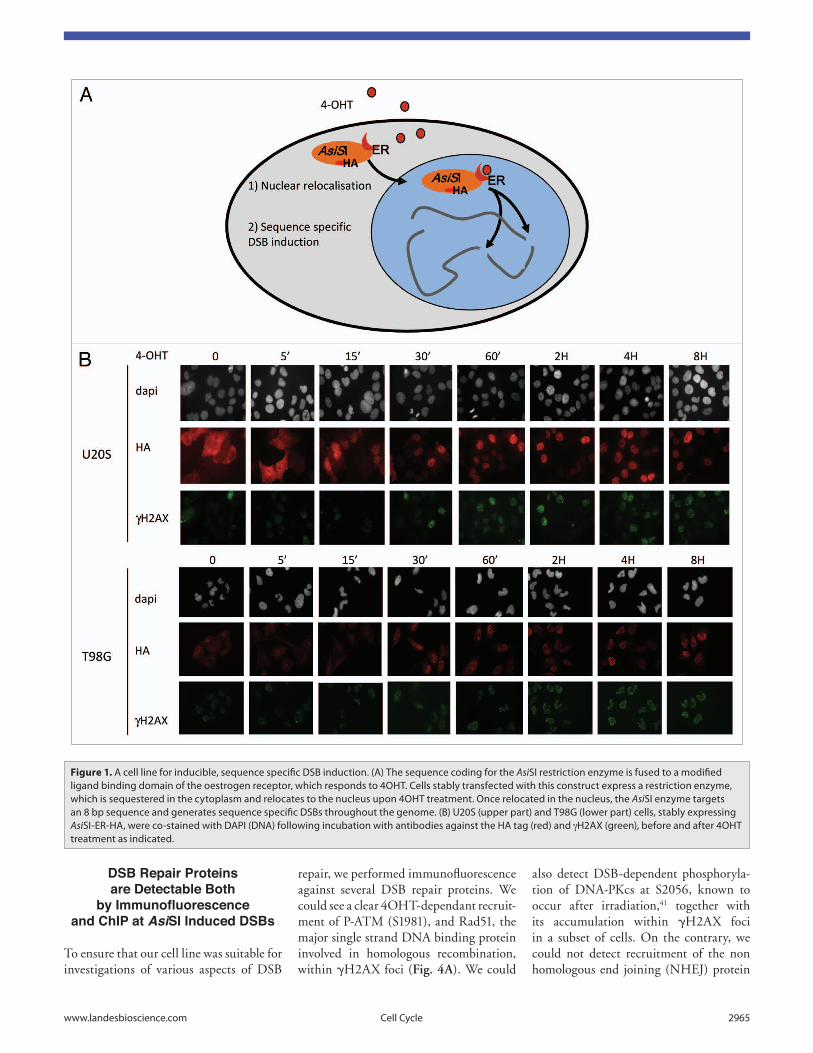
We raised both U20S and T98G sta-ble cell lines expressing AsiSI-ER, with the T98G line allowing cell cycle syn-chronization by serum starvation. In the absence of 4-hydroxytamoxifen (4OHT), the fusion protein is largely cytoplasmic and thus unable to induce DSBs, illus-trated by the lack or low background levels of γH2AX foci (Fig. 1B). 4OHT treatment rapidly triggered nuclear localisation of AsiSI-ER, being com-pleted within 15 to 30 min of treatment, depending upon the cell line (Fig. 1B). Nuclear localisation coincided with DSB induction, indicated by the apparition of γH2AX foci within 30 min of 4OHT treatment (Fig. 1B).
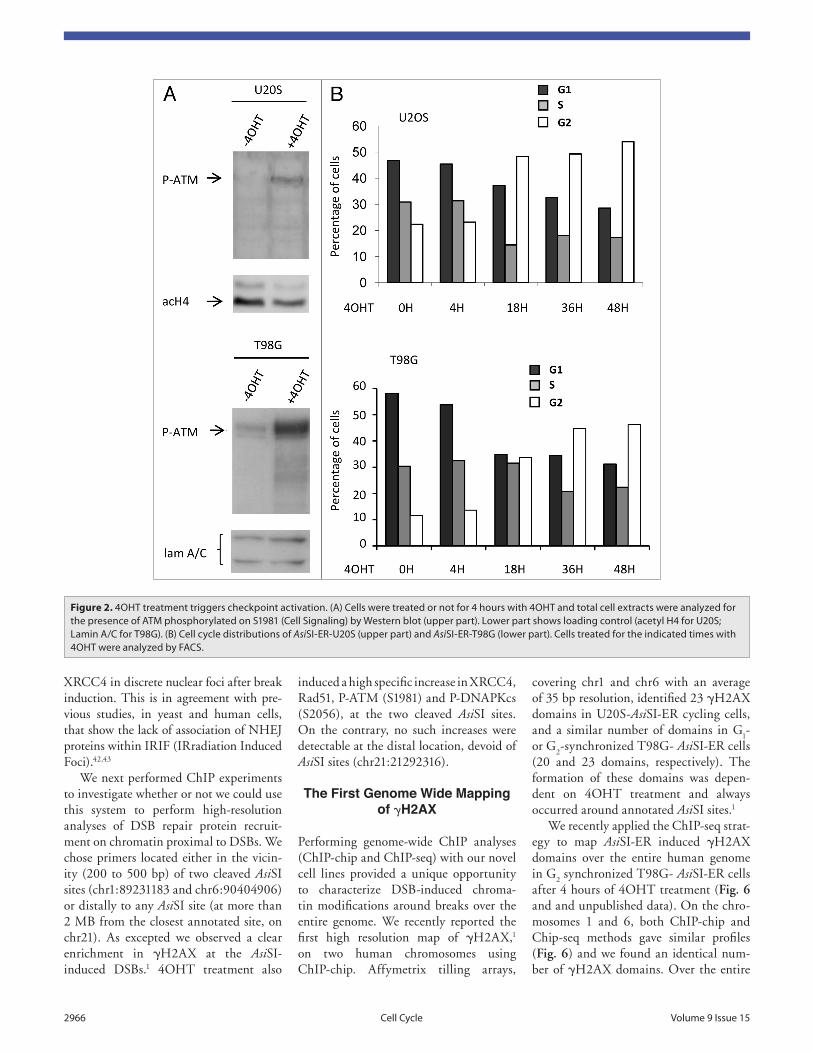
We first verified that the AsiSI-ER stable cell lines were able to properly acti-vate the DNA damage response (DDR) in the presence of 4OHT. In both cell lines, 4OHT treatment induced detectable phosphorylation of ATM on S1981 after 4H (Fig. 2A) and cell cycle arrest, primar-ily in G
2 phase, after 18H of treatment,
illustrating an activation of the G2/M
checkpoint (Fig. 2B). No cell death/apop-tosis was detected even 72H after DSB induction (data not shown). These data are in agreement with previous results obtained when transducing TAT-PvuII in U2OS cells. The continuous generation of PvuII-mediated DSBs led cells to accumu-late in G
2 but did not provoke apoptosis.40
In order to assess AsiSI-ER cutting efficiency in vivo at specific genomic locations and throughout the whole cell population, we developed a cleavage effi-ciency assay based on the use of biotinyl-ated double stranded oligonucleotides cohesive with the AsiSI site.1 Kinetic analyses performed around two AsiSI sites revealed that all sites did not behave in the same manner (Fig. 3): the site located at chr1:3092903 was not cleaved at all, independently of the length of 4OHT treatment. On the contrary the AsiSI site located at chr1:89231183 showed efficient cleavage after 15 min and reached peak levels after 4H of treatment. Our genome wide cleavage analyses revealed that some annotated AsiSI sites were inefficiently cut in vivo. We found this lack of cleavage to be likely due to the methylation status at specific sites, as AsiSI is inhibited by CpG methylation.1
ChIP.16,19,20,33,34 More recently, insertion of HO cut sites elsewhere on the genome was used to study γH2A distribution in a few different contexts.35 In mammalian cells, the lab of M. Jasin has developed an elegant system based on the expression of the rare-cutting enzyme I-SceI. This homing endonuclease targets an 18 bp palindromic sequence that is not present in mammalian genomes. Introduction of a transgene carrying this sequence per-mits induction of a single localised DSB upon expression of I-SceI.31 This system has been utilized to study DNA repair fidelity,36 checkpoint sensitivity37 and, more recently, histone modifications sur-rounding DSBs by ChIP.38,39 Finally, the lab of M. Kastan has used the I-PpoI homing endonuclease to generate a few endogenous, sequence-specific DSBs in human cells.30
However, none of these mammalian systems allows precise investigations of several breaks located in various chro-matin contexts. The I-SceI-based sys-tem31 leads to only one localized DSB, therefore limiting analyses to one single break. While the I-PpoI-based system30 generates several endogenous sequence-specific DSBs, they are mainly located in ribosomal genes, which limits the analy-sis of DSB repair to a very specific type of loci and hinders the use of genome-wide studies since ribosomal genes tend to fall into unassembled regions of the human genome due to their repetitive sequences.
A Cell System to Induce Multiple Sequence Specific DSBs Over the Entire Genome
Recently, we have developed stable human cell lines expressing a restriction enzyme, with an 8 bp recognition sequence, fused to a modified oestrogen receptor ligand binding domain (AsiSI-ER)1 (Fig. 1A). Since some epigenetic modifications induced around DSBs can spread over more than 1 Mbp, it was important to choose a restriction enzyme that rarely cuts on the genome to analyse chroma-tin modifications occurring around each break. For this reason, we chose the AsiSI enzyme, which according to the human genome sequence, should generate frag-ments longer than 1 Mbp.
accessibility.12,13 In addition, H2AX phos-phorylation is followed by the recruitment of Histone Acetyltransferases (HATs) catalysing H3/H4 acetylation,14-18 pro-posed to help chromatin relaxation. Both acetylation and phosphorylation con-tribute to the recruitment of nucleosome remodelling factors, such as INO80, and SWI/SNF, further enhancing DSB acces-sibility.16,17,19,20 Recently ubiquitination and sumoylation pathways have also been shown to participate, with several studies revealing complex recruitment patterns of E3 Ub and Sumo ligases, together with various Ub- and Sumo-ylated proteins, around DSBs,21-27 (reviewed in refs. 28 and 29). Combined, these chromatin modifications not only generate a chro-matin state permissive to DNA repair, but also contribute to the recruitment of DSB repair complexes and activation of DSB signalling and checkpoint path-ways. Thus, obtaining detailed informa-tion concerning changes in the chromatin landscape induced by DSBs represents an important prerequisite to understanding the damage repair process.
A Missing Tool for High-Resolution,
Genome-Wide Studies
Classical DSB induction methods involve either drugs or radiation, applied on the whole cell or on laser-localised track in the nuclei, which create heterogeneous DSBs at random positions in the genome. For this reason, most of the studies detailed above were performed by immunofluo-rescence, providing low-resolution, global and kinetic information. Chromatin immunoprecipitation (ChIP) allows high-resolution, molecular analyses of chro-matin but requires homogeneous DSB induction at known loci in all cells (as ChIP requires several million cells per assay). A few systems have been devel-oped that induce localized DSBs in yeast and mammalian cells.30-32 In S. cerevisae, expression of the HO endonuclease from a galactose inducible promoter triggers induction of a single DSB at a single anno-tated locus (MAT) in the yeast genome. This system has been widely used to study histone modifications and recruitment of DNA repair proteins around a DSB by
www.landesbioscience.com Cell Cycle 2965
also detect DSB-dependent phosphoryla-tion of DNA-PKcs at S2056, known to occur after irradiation,41 together with its accumulation within γH2AX foci in a subset of cells. On the contrary, we could not detect recruitment of the non homologous end joining (NHEJ) protein
repair, we performed immunofluorescence against several DSB repair proteins. We could see a clear 4OHT-dependant recruit-ment of P-ATM (S1981), and Rad51, the major single strand DNA binding protein involved in homologous recombination, within γH2AX foci (Fig. 4A). We could
DSB Repair Proteins are Detectable Both
by Immunofluorescence and ChIP at AsiSI Induced DSBs
To ensure that our cell line was suitable for investigations of various aspects of DSB
Figure 1. A cell line for inducible, sequence specific DSB induction. (A) the sequence coding for the AsiSi restriction enzyme is fused to a modified ligand binding domain of the oestrogen receptor, which responds to 4OHt. Cells stably transfected with this construct express a restriction enzyme, which is sequestered in the cytoplasm and relocates to the nucleus upon 4OHt treatment. Once relocated in the nucleus, the AsiSi enzyme targets an 8 bp sequence and generates sequence specific DSBs throughout the genome. (B) U20S (upper part) and t98G (lower part) cells, stably expressing AsiSi-er-HA, were co-stained with DAPi (DNA) following incubation with antibodies against the HA tag (red) and gH2Ax (green), before and after 4OHt treatment as indicated.
2966 Cell Cycle volume 9 issue 15
covering chr1 and chr6 with an average of 35 bp resolution, identified 23 γH2AX domains in U20S-AsiSI-ER cycling cells, and a similar number of domains in G
1-
or G2-synchronized T98G- AsiSI-ER cells
(20 and 23 domains, respectively). The formation of these domains was depen-dent on 4OHT treatment and always occurred around annotated AsiSI sites.1
We recently applied the ChIP-seq strat-egy to map AsiSI-ER induced γH2AX domains over the entire human genome in G
2 synchronized T98G- AsiSI-ER cells
after 4 hours of 4OHT treatment (Fig. 6 and and unpublished data). On the chro-mosomes 1 and 6, both ChIP-chip and Chip-seq methods gave similar profiles (Fig. 6) and we found an identical num-ber of γH2AX domains. Over the entire
induced a high specific increase in XRCC4, Rad51, P-ATM (S1981) and P-DNAPKcs (S2056), at the two cleaved AsiSI sites. On the contrary, no such increases were detectable at the distal location, devoid of AsiSI sites (chr21:21292316).
The First Genome Wide Mapping of γH2AX
Performing genome-wide ChIP analyses (ChIP-chip and ChIP-seq) with our novel cell lines provided a unique opportunity to characterize DSB-induced chroma-tin modifications around breaks over the entire genome. We recently reported the first high resolution map of γH2AX,1 on two human chromosomes using ChIP-chip. Affymetrix tilling arrays,
XRCC4 in discrete nuclear foci after break induction. This is in agreement with pre-vious studies, in yeast and human cells, that show the lack of association of NHEJ proteins within IRIF (IRradiation Induced Foci).42,43
We next performed ChIP experiments to investigate whether or not we could use this system to perform high-resolution analyses of DSB repair protein recruit-ment on chromatin proximal to DSBs. We chose primers located either in the vicin-ity (200 to 500 bp) of two cleaved AsiSI sites (chr1:89231183 and chr6:90404906) or distally to any AsiSI site (at more than 2 MB from the closest annotated site, on chr21). As excepted we observed a clear enrichment in γH2AX at the AsiSI-induced DSBs.1 4OHT treatment also
Figure 2. 4OHt treatment triggers checkpoint activation. (A) Cells were treated or not for 4 hours with 4OHt and total cell extracts were analyzed for the presence of AtM phosphorylated on S1981 (Cell Signaling) by western blot (upper part). Lower part shows loading control (acetyl H4 for U20S; Lamin A/C for t98G). (B) Cell cycle distributions of AsiSi-er-U20S (upper part) and AsiSi-er-t98G (lower part). Cells treated for the indicated times with 4OHt were analyzed by FACS.

www.landesbioscience.com Cell Cycle 2967
chromatin remodeling and transcription.1 Our results highlight the potential of the combination of AsiSI-ER stable cell lines and genome wide ChIP-chip and ChIP-seq techniques.
Exciting, future applications will be to investigate the three-dimensional distri-bution of DSBs. The sequence specific-ity and multiplicity of AsiSI-ER induced DSBs open new opportunities to use DNA FISH and high-resolution 3D tech-niques such as chromosome conforma-tion capture (3C) and HiC, to study how DSBs are organized within the nucleus. Combinatorial applications of such tech-niques will uncover the precise nature of the coordinate regulation of previously disparate research interests such as DNA damage repair, transcription and chroma-tin structure/function.
Material and Methods
Cell culture. Construction of the pBabe HA-AsiSI–ER vector, and generation of the stable cell lines were previously described.1 U20S-AsiSI-ER cells were cultured in DMEM GlutaMAX sup-plemented with antibiotics, 10% FCS (Invitrogen) and puromycin (1 µg/mL). T98G-AsiSI-ER were cultured in MEM GlutaMAX, supplemented with non-essential amino acid (NEAA), antibiotics, 10% FCS (Invitrogen) and puromycin (1 µg/mL). Cells were grown at 37°C under
15 minutes upon 4OHT (see our cleavage assay Fig. 3). Thus our system may also reveal suitable to study the kinetic of pro-tein recruitment on DSB.
In addition, stable AsiSI-ER cell lines offer the unique opportunity to investigate DSB-associated events with high-resolu-tion and at several distinct loci simultane-ously, providing unprecedented statistical analyses that will enable new biological dis-coveries. We were able to analyze γH2AX spreading over more than 20 MB, and thus uncovered previously undetermined features concerning γH2AX spreading. We found that γH2AX spread over 0.5 up to 2 MB of chromatin surrounding each break1 (Fig. 6) and observed an assy-metrical spreading around certain breaks, which revealed the existence of “boundar-ies” that limit γH2AX spreading within domains. Unfortunately, although we performed thorough comparisons with existing genomic and epigenomic data, we have not yet identified chromatin/DNA features that would account for these boundaries. However, we also found that γH2AX spreading was highly dis-continuous within domains and we iden-tified gene transcription as one of the determinants of this uneven distribution. Finally, we discovered that gene transcrip-tion remains unchanged within γH2AX domains after DSB induction, reveal-ing unexpected insights into the global orchestration of DSB repair-associated
genome, 4OHT treatment triggered the induction of 105 γH2AX domains. Our system thus generates approximately 200 DSBs in diploid G
1 cells (400 DSBs in
G2), which corresponds to an irradiation
of 5–10 Gy.
Characterizing the Chromatin Structure and Function
in DSB Repair
This new DSB-inducible system should be useful for various types of studies required by the DSB repair field. First, it permits investigation of “clean” DSB-triggered epigenetic events in the absence of any typical irradiation or genotoxic side effects. ROS production and/or com-plex chemical alterations of neighbour-ing nucleotides are indeed frequently associated with conventional clastogenic treatments and complicate result inter-pretation due to unrelated phenomenon such as kinase activations and base exci-sion repair (BER) engagement.44 Second, using ChIP followed by classical Q-PCR analysis allows investigation of DSB repair protein recruitment at a molecular level, which is sometimes critical in order to observe local events undetectable by conventional immunofluorescence. For example, we could see a clear recruitment of XRCC4 by ChIP, while no IRIF asso-ciation was detectable by IF. Importantly we can detect DSB induction already after
Figure 3. Kinetics of AsiSi-DSB induction. Genomic DNA, extracted at the indicated times relative to 4OHt treatment, was assayed for cleavage at two different AsiSi sites (described in Material and Methods). Briefly, a biotinylated oligonucleotide, cohesive with the AsiSi site, was ligated to genomic DNA prepared from 4OHt treated and untreated AsiSi-er-U20S cells and pulled down with streptavidin beads. An AsiSi-linearized plasmid, added to each sample before ligation, served as a normalization control. Pulled down DNA was analyzed by quantitative PCr amplification using two sets of primers flanking two AsiSi sites, and a primer pair located on the plasmid. the cleavage efficiency was calculated as a percentage relative to the signal obtained with the AsiSi-linearized plasmid.

2968 Cell Cycle volume 9 issue 15
anti-Phospho-ATM S1981 (10H11.E12, Cell Signaling Technology) with a rabbit monoclonal anti-γH2AX (Epitomics); a mouse monoclonal anti-γH2AX (JBW 301, Upstate Biotechnology) with either
Paraformaldehyde and immunofluores-cence was carried out following a stan-dard protocol. Combinations of the following antibodies were used: a mouse monoclonal anti-HA (HA11, Covance) or
a humidified atmosphere with 5% CO2.
Cells were treated with 300 nM 4OHT for the indicated periods.
Immunofluorescence. Cells grown on glass coverslips were fixed with 4%
Figure 4. recruitment of DSB repair proteins within AsiSi-induced γH2Ax foci. AsiSi-er-U20S cells were costained with DAPi (DNA) following incuba-tion with antibodies against gH2Ax and either AtM phosphorylated on S1981, DNAPKcs phosphorylated on S2056, xrCC4 or rad51, before and after 4OHt treatment.

www.landesbioscience.com Cell Cycle 2969
twice with ice-cold PBS and then lysed in the appropriate lysis buffer with sample reducing agent (Invitrogen). Liquid trans-fer of resolved proteins was performed onto PVDF membranes (Invitrogen). After 1 hour block in 5% non-fat dry milk/0.5% PBS-Tween, membranes were
system (Universal Imaging Corp., West Chester, PA).
Western blotting. Western blot analysis was performed using NuPAGE Tris-acetate 3–8% gels and reagents (Invitrogen) according to the manufac-turer indications. Briefly, cells were rinced
a rabbit anti-Phospho-DNA-PKcs S2056 (Abcam ab18192), anti-Rad51 (H92; SantaCruz Biotech sc-8349) or anti-XRCC4 (Abcam, ab14550). All second-ary antibodies were purchased from Molecular Probes. Image acquisition was performed with the MetaVue imaging
Figure 5. ChiP analyses of DSB repair proteins on AsiSi-induced DSBs. ChiP analysis was performed in AsiSi-er-U20S cells, before (-) and after (+) 4 hour 4OHt treatment, using no antibody (mock) and either anti P-AtM (S1981) (Cell Signaling 10H11.e12), anti P-DNAPK (S2056) (Abcam ab18193), anti rad51 (SantaCruz sc-8349), anti xrCC4 (Abcam ab145) or anti gH2Ax (Upstate 07–164) as indicated. enrichment was assessed by real time Q-PCr amplification using primers proximal to two AsiSi induced DSBs (chr1:89231183 and chr6:90404906) or distal (2 MB) to an AsiSi site located on chr21. the means and standard deviations of the mean (s.d.m) from three independent experiments are plotted.

2970 Cell Cycle volume 9 issue 15
Figure 6. γH2Ax mapping by ChiP-chip and ChiP-seq. (A) γH2Ax (epitomics 2212-1) ChiPs were performed on AsiSi-er-t98G cells synchronised in G2, before and after 4 hours of 4OHt treatment and were either hybridized to the Human tiling Array 2.0r A, which covers chromosomes 1 and 6 or subjected to deep sequencing (Solexa). the profiles obtained using both techniques are presented for chromosome 1 and 6. For ChiP-seq data, the y axis represents the number of sequenced tags, counted in 200 bp windows and smoothed by averaging over a 20 kb sliding window. For ChiP-chip data, the y axis represents the log2 (γH2Ax/input) smoothed over a sliding window of 500 probes. AsiSi site positions are indicated. Stars indicate non specific signal (repeat/pericentromeric regions). (B) Detailed views of ChiP-chip and ChiP-seq data around two AsiSi sites. Note the similarity between both profiles.

www.landesbioscience.com Cell Cycle 2971
8. Celeste A, Petersen S, Romanienko PJ, Fernandez-Capetillo O, Chen HT, Sedelnikova OA, et al. Genomic instability in mice lacking histone H2AX. Science 2002; 296:922-7.
9. Fernandez-Capetillo O, Mahadevaiah SK, Celeste A, Romanienko PJ, Camerini-Otero RD, Bonner WM, et al. H2AX is required for chromatin remodeling and inactivation of sex chromosomes in male mouse meiosis. Dev Cell 2003; 4:497-508.
10. Pandita TK, Richardson C. Chromatin remodel-ing finds its place in the DNA double-strand break response. Nucleic Acids Res 2009; 37:1363-77.
11. van Attikum H, Gasser SM. Crosstalk between histone modifications during the DNA damage response. Trends Cell Biol 2009; 19:207-17.
12. Ayoub N, Jeyasekharan AD, Bernal JA, Venkitaraman AR. HP1-beta mobilization promotes chromatin changes that initiate the DNA damage response. Nature 2008; 453:682-6.
13. Goodarzi AA, Noon AT, Deckbar D, Ziv Y, Shiloh Y, Lobrich M, et al. ATM signaling facilitates repair of DNA double-strand breaks associated with hetero-chromatin. Mol Cell 2008; 31:167-77.
14. Qin S, Parthun MR. Recruitment of the type B his-tone acetyltransferase Hat1p to chromatin is linked to DNA double-strand breaks. Mol Cell Biol 2006; 26:3649-58.
15. Tamburini BA, Tyler JK. Localized histone acetyla-tion and deacetylation triggered by the homolo-gous recombination pathway of double-strand DNA repair. Mol Cell Biol 2005; 25:4903-13.
16. Downs JA, Allard S, Jobin-Robitaille O, Javaheri A, Auger A, Bouchard N, et al. Binding of chromatin-modifying activities to phosphorylated histone H2A at DNA damage sites. Mol Cell 2004; 16:979-90.
17. Lee HS, Park JH, Kim SJ, Kwon SJ, Kwon J. A coop-erative activation loop among SWI/SNF, gamma-H2AX and H3 acetylation for DNA double-strand break repair. EMBO J 2010; 29:1434-45.
18. Sun Y, Jiang X, Xu Y, Ayrapetov MK, Moreau LA, Whetstine JR, et al. Histone H3 methylation links DNA damage detection to activation of the tumour suppressor Tip60. Nat Cell Biol 2009; 11:1376-82.
19. Tsukuda T, Fleming AB, Nickoloff JA, Osley MA. Chromatin remodelling at a DNA double-strand break site in Saccharomyces cerevisiae. Nature 2005; 438:379-83.
20. van Attikum H, Fritsch O, Hohn B, Gasser SM. Recruitment of the INO80 complex by H2A phos-phorylation links ATP-dependent chromatin remod-eling with DNA double-strand break repair. Cell 2004; 119:777-88.
21. Giannattasio M, Lazzaro F, Plevani P, Muzi-Falconi M. The DNA damage checkpoint response requires histone H2B ubiquitination by Rad6-Bre1 and H3 methylation by Dot1. J Biol Chem 2005; 280:9879-86.
22. Mailand N, Bekker-Jensen S, Faustrup H, Melander F, Bartek J, Lukas C, et al. RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins. Cell 2007; 131:887-900.
23. Stewart GS, Panier S, Townsend K, Al-Hakim AK, Kolas NK, Miller ES, et al. The RIDDLE syndrome protein mediates a ubiquitin-dependent signaling cas-cade at sites of DNA damage. Cell 2009; 136:420-34.
24. Galanty Y, Belotserkovskaya R, Coates J, Polo S, Miller KM, Jackson SP. Mammalian SUMO E3-ligases PIAS1 and PIAS4 promote responses to DNA double-strand breaks. Nature 2009; 462:935-9.
25. Huen MS, Grant R, Manke I, Minn K, Yu X, Yaffe MB, et al. RNF8 transduces the DNA-damage signal via histone ubiquitylation and checkpoint protein assembly. Cell 2007; 131:901-14.
26. Kolas NK, Chapman JR, Nakada S, Ylanko J, Chahwan R, Sweeney FD, et al. Orchestration of the DNA-damage response by the RNF8 ubiquitin ligase. Science 2007; 318:1637-40.
CCT TGC AAA CCA GTC CT; and chr6_90404906: FW: TGC CGG TCT CCT AGA AGT TG, REV: GCG CTT GAT TTC CCT GAG T) or distal (2.8 MB) to the AsiSI site chr21_21292316 (FW: TGG CTG GAA CTG CTT TCT TT, REV: GGT GAG TGA ATG AGC TGC AA).
The procedure for ChIP-chip experi-ment and analyses are described in ref. 1. For ChIP-seq, sequencing librar-ies were prepared using 10 ng of puri-fied DNA (averaged size 250–300 bp), by the Genomic Core facility (EMBL, Heidelberg) and subjected to high throughput sequencing (Illumina Genome Analyzer II, single read). After quality fil-tering, the number of uniquely mapped sequencing reads (on genome hg18) was counted over 200 bp windows. The signal presented here was further smoothed over a 20 kb sliding window.
Acknowledgements
We thank NEB Biolabs for providing AsiSI genomic DNA, the EMBL Genomic Core facility for high throughput sequencing and Dr. C. Normand for critical reading of the manuscript. Funding was provided by grants from the CNRS (PEPS), PRES_Toulouse University, ARC and ANR Jeunes Chercheurs.
References1. Iacovoni JS, Caron P, Lassadi I, Nicolas E, Massip L,
Trouche D, et al. High-resolution profiling of gam-maH2AX around DNA double strand breaks in the mammalian genome. EMBO J 2010; 29:1446-57.
2. Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol 1999; 146:905-16.
3. Rogakou EP, Pilch DR, Orr AH, Ivanova VS, Bonner WM. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J Biol Chem 1998; 273:5858-68.
4. Stucki M, Jackson SP. gammaH2AX and MDC1: anchoring the DNA-damage-response machinery to broken chromosomes. DNA Repair (Amst) 2006; 5:534-43.
5. Celeste A, Fernandez-Capetillo O, Kruhlak MJ, Pilch DR, Staudt DW, Lee A, et al. Histone H2AX phosphorylation is dispensable for the initial recogni-tion of DNA breaks. Nat Cell Biol 2003; 5:675-9.
6. Ward IM, Minn K, Jorda KG, Chen J. Accumulation of checkpoint protein 53BP1 at DNA breaks involves its binding to phosphorylated histone H2AX. J Biol Chem 2003; 278:19579-82.
7. Celeste A, Difilippantonio S, Difilippantonio MJ, Fernandez-Capetillo O, Pilch DR, Sedelnikova OA, et al. H2AX haploinsufficiency modifies genom-ic stability and tumor susceptibility. Cell 2003; 114:371-83.
incubated overnight with the following primary antibodies: anti-phospho-ATM S1981 (10H11.E12, 1/500); anti-AcH4 (Upstate 06–866,1/1,000) and anti-Lamin A/C (SantaCruz H-110, 1/500). Horseradish peroxidase-coupled sec-ondary antibodies were purchased from Amersham Biosciences and chemilumi-nescence Lumilight reagent was from Roche Diagnostic.
Cleavage efficiency assay. A detailed protocol for the cleavage efficiency assay was described previously in reference 1. To measure the percentage of broken loci, 0.5 ng of an AsiSI-linearized pREP plas-mid was added in the ligation mix. Pull down DNA was analyzed by Real-time Q-PCR with either control primers located on the pREP plasmid (FW: ACA TTG TTG GAG CCG AAA TC, REV: CGC AAG GAA TCG GTC AAT AC) or with primers located proximally to genomic AsiSI sites (chr1_89231183: FW: GAT TGG CTA TGG GTG TGG AC, REV: CAT CCT TGC AAA CCA GTC CT and chr1_3092903: FW: CGA GAT CCA AGG AAG TCG TG; REV: CCC CGG ACA CTT TAA AAG GA). Cleavage efficiency (as a percentage) was calculated related to the signal obtained with primers located on the AsiSI linear-ized plasmid.
Flow cytometry analysis. Cells were seeded in 100 mm culture dishes, and col-lected after the indicated time of 4OHT treatment. Cells were fixed in 70% ice-cold Ethanol, spun down and incubated for 30 min with 500 µL propidium iodide staining solution (BD PharmingenTM) and analyzed by flow cytometry (FACScalibur) to determine their cell cycle distribution.
Chromatin immunoprecipitation and high throughput sequencing. ChIP assays were carried out according to the protocol described in ref. 1. 200 µg of chromatin was immunoprecipitated using 2 µg of an anti-Phospho-ATM S1981 (10H11.E12, Cell Signaling Technology), an anti-Phos-pho-DNA-PKcs S2056 (Abcam ab18192), an anti-Rad51 (H92; SantaCruz Biotech sc-8349), an anti-XRCC4 (Abcam, ab14550) or no antibody (mock). ChIP were analyzed by real-time QPCR using primers proximal to two AsiSI sites (chr1_89231183: FW: GAT TGG CTA TGG GTG TGG AC, REV: CAT

2972 Cell Cycle volume 9 issue 15
40. Kuhne C, Tjornhammar ML, Pongor S, Banks L, Simoncsits A. Repair of a minimal DNA double-strand break by NHEJ requires DNA-PKcs and is controlled by the ATM/ATR checkpoint. Nucleic Acids Res 2003; 31:7227-37.
41. Chen BP, Chan DW, Kobayashi J, Burma S, Asaithamby A, Morotomi-Yano K, et al. Cell cycle dependence of DNA-dependent protein kinase phos-phorylation in response to DNA double strand breaks. J Biol Chem 2005; 280:14709-15.
42. Jakob B, Scholz M, Taucher-Scholz G. Characterization of CDKN1A (p21) binding to sites of heavy-ion-induced damage: colocalization with proteins involved in DNA repair. Int J Radiat Biol 2002; 78:75-88.
43. Lisby M, Barlow JH, Burgess RC, Rothstein R. Choreography of the DNA damage response: spatio-temporal relationships among checkpoint and repair proteins. Cell 2004; 118:699-713.
44. Chen SK, Tsai MH, Hwang JJ, Chang WP. Determination of 8-oxoguanine in individual cell nucleus of gamma-irradiated mammalian cells. Radiat Res 2001; 155:832-6.
34. Martin SG, Laroche T, Suka N, Grunstein M, Gasser SM. Relocalization of telomeric Ku and SIR proteins in response to DNA strand breaks in yeast. Cell 1999; 97:621-33.
35. Kim JA, Kruhlak M, Dotiwala F, Nussenzweig A, Haber JE. Heterochromatin is refractory to gamma-H2AX modification in yeast and mammals. J Cell Biol 2007; 178:209-18.
36. Nickoloff JA, Brenneman MA. Analysis of recom-binational repair of DNA double-strand breaks in mammalian cells with I-SceI nuclease. Methods Mol Biol 2004; 262:35-52.
37. Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 2003; 421:499-506.
38. Murr R, Loizou JI, Yang YG, Cuenin C, Li H, Wang ZQ, et al. Histone acetylation by Trrap-Tip60 modu-lates loading of repair proteins and repair of DNA double-strand breaks. Nat Cell Biol 2006; 8:91-9.
39. Rodrigue A, Lafrance M, Gauthier MC, McDonald D, Hendzel M, West SC, et al. Interplay between human DNA repair proteins at a unique double-strand break in vivo. EMBO J 2006; 25:222-31.
27. Bekker-Jensen S, Rendtlew Danielsen J, Fugger K, Gromova I, Nerstedt A, Lukas C, et al. HERC2 coordinates ubiquitin-dependent assembly of DNA repair factors on damaged chromosomes. Nat Cell Biol 2010; 12:80-6.
28. Messick TE, Greenberg RA. The ubiquitin landscape at DNA double-strand breaks. J Cell Biol 2009; 187:319-26.
29. Panier S, Durocher D. Regulatory ubiquitylation in response to DNA double-strand breaks. DNA Repair (Amst) 2009; 8:436-43.
30. Berkovich E, Monnat RJ Jr, Kastan MB. Roles of ATM and NBS1 in chromatin structure modulation and DNA double-strand break repair. Nat Cell Biol 2007; 9:683-90.
31. Rouet P, Smih F, Jasin M. Expression of a site-specific endonuclease stimulates homologous recombination in mammalian cells. Proc Natl Acad Sci USA 1994; 91:6064-8.
32. Kramer KM, Brock JA, Bloom K, Moore JK, Haber JE. Two different types of double-strand breaks in Saccharomyces cerevisiae are repaired by similar RAD52-independent, nonhomologous recombina-tion events. Mol Cell Biol 1994; 14:1293-301.
33. Wolner B, van Komen S, Sung P, Peterson CL. Recruitment of the recombinational repair machinery to a DNA double-strand break in yeast. Mol Cell 2003; 12:221-32.
Copyright © 2022 FDOKUMEN




















