
Electrophoretic karyotype and gene assignment to resolved chromosomes of Trichoderma spp
Upload
independentCategory
view
8download
0
1600 Electrophoresis 2013, 34, 1600–1609
Xavier Subirats1∗,∗∗Victor U. Weiss2∗Irene Gosler1
Christoph Puls2
Andreas Limbeck2
Gunter Allmaier2
Ernst Kenndler3
1Max F. Perutz Laboratories,Department of MedicalBiochemistry, MedicalUniversity of Vienna, ViennaBiocenter (VBC), Vienna,Austria
2Institute of ChemicalTechnologies and Analytics,Vienna University ofTechnology, Vienna, Austria
3Institute for AnalyticalChemistry, University of Vienna,Vienna, Austria
Received December 19, 2012Revised January 29, 2013Accepted January 31, 2013
Research Article
Characterization of rhinovirus subviral Aparticles via capillary electrophoresis,electron microscopy and gas phaseelectrophoretic mobility molecular analysis:Part II
Human rhinoviruses (HRVs) are valuable tools in the investigation of early viral infec-tion steps due to their far reaching (although still incomplete) characterization. Duringendocytosis, native virions first loose one of the four capsid proteins (VP4); correspondingparticles sediment at 135S and were termed subviral A particles. Subsequently, the viralRNA genome leaves the viral shell giving rise to empty capsids. In continuation of ourprevious work with HRV serotype 2 (HRV2) intermediate subviral particles, in which wewere able to discriminate by CE even between two intermediates (AI and AII) of virusuncoating, we further concentrated on the characterization of AI particles with the elec-trophoretic mobility of around –17.2 × 10−9 m2/Vs at 20�C. In the course of our presentwork we related these particles to virions as previously described at the subviral A stage ofuncoating (and as such sedimenting at 135S) by determination of their protein and RNAcontent—in comparison to native virions AI particles did not include VP4, however, still93% of their initial RNA content. Binding of an mAb specific for subviral particles demon-strated antigenic rearrangements on the capsid surface at the AI stage. Furthermore, weinvestigated possible factors stabilizing intermediates of virus uncoating. We could ex-clude the influence of the previously suspected so-called contaminant of virus preparationon HRV2 subviral particle formation. Instead, we regarded other factors being part of thevirus preparation system and found a dependence of AI particle formation on the presenceof divalent cations.
Keywords:
HRV2 / inductively coupled plasma mass spectrometry / Rhinovirus / Subviralparticle DOI 10.1002/elps.201200686
1 Introduction
Human rhinoviruses (HRVs) from the Picornavirus family,nonenveloped viruses of about 30 nm diameter and com-posed of 60 copies each of four viral proteins (VPs) and anapproximately 7.1 kbases long viral RNA genome, are themain causative agents of common cold infections. Based onreceptor specificity upon viral cell infection, HRVs can bediscriminated into a major and a minor group: major groupvirions attach to intercellular adhesion molecule 1 (ICAM 1)for cell entry, minor group virions to members of the low-density lipoprotein receptor family. The receptor of the newly
Correspondence: Dr. Victor U. Weiss, Institute of Chemical Tech-nologies and Analytics, Vienna University of Technology, Getrei-demarkt 9/164, A-1060 Vienna, AustriaE-mail: [email protected]: +43-1-58801-15199
Abbreviations: HRV, human rhinovirus; ICAM 1, intercellu-lar adhesion molecule 1; PB, preparation buffer; SB, samplebuffer; VP, viral protein
discovered phylogenic species C is still unknown. In our workwe concentrated on one of the roughly 100 HRV serotypes,HRV2, a member of the minor virus group.
Virus/cell recognition is followed by endocytosis ofbound virions via clathrin-coated vesicles. During cell up-take, these vesicles bud from the cellular membrane and un-dergo maturation from early-to-late endosomes resulting inthe change of the originally relatively mild acidic pH (around6.0 to 6.5), at which the virus–receptor complex dissociates,to pH values below 5.6. At this pH, structural changes inthe virus capsid occur, allowing the virus to interact withthe endosomal membrane and to transfer its RNA genomethrough the lipid bilayer into the cytosol of the infected cell bystill largely unknown mechanisms (for a recent review referto [1]).
∗These authors contributed equally to this work.∗∗Additional corresponding author: Dr. Xavier Subirats, E-mail:[email protected]; Current address: Department of AnalyticalChemistry, University of Barcelona, Barcelona, Spain.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

Electrophoresis 2013, 34, 1600–1609 CE and CEC 1601
Figure 1. Scheme of HRV2 un-coating, that is, the forma-tion of subviral particles in thecourse of early viral cell infec-tion. The sequence of infectionevents is shown from left toright. At which point of infec-tion intermediate AII particles[17] are playing a role couldnot been answered so far.
According to the respective sedimentation of the virusand subviral particles, Lonberg-Holm and Yin [2] were ableto discriminate particles sedimenting at 150S, 135S and80S, respectively; 150S particles were shown to be nativevirions consisting of all four VPs and the viral RNA genome,whereas 135S particles (also named A particles) as intermedi-ates of viral cell infection have lost already VP4. Additionally,A particles presumably have undergone gross changes in thearrangement of the virus capsid; 80S particles (so-called Bparticles) are empty capsids; they are devoid of the viral RNAgenome as well and form endpoints of viral cell infection. Ascheme of virus uncoating is depicted in Fig. 1.
In comparison to intact virions, empty capsids have ex-panded by approximately 4% in diameter and exhibit a 1-nm-wide channel at the fivefold symmetry axis due to struc-tural rearrangements of VP1 on the capsid surface. Additionalstructural changes concern the area around the twofold axeson the proteinaceous virus capsid, where additional pores areformed [3,4]. More recently, Pickl-Herk, A. et al. (manuscriptin preparation) were able to identify these pores at the twofoldaxes as exit points of rhinoviral RNA.
Structural changes of the HRV2 capsid are also reflectedby changes in its antigenicity. B particles show reactivity to-ward the precipitating mAb 2G2 [5]. Subsequently, the 2G2binding site was determined at the star-shaped mesa on thesurface of the empty capsid after VP1 rearrangement in thecourse of virus uncoating; this structural epitope is not foundon intact virions but is only present on the surface of subviralparticles [6].
Intact virions as well as subviral B particles of HRV2 andtheir respective binding characteristics to receptor fragments,antibodies, and even receptor-decorated liposomes have al-ready been analytically assessed. In doing so, electrophoresiseither in the capillary (CE) or in its miniaturized chip formatallowed separation of particles in free solution (for a recentreview refer to [7]). Lately, also CE of related poliovirus wasof increasing interest [8, 9] as was the separation of bioparti-cles in general (for some exemplary reviews refer to [10, 11]).
A series of papers of another group dealt with intermedi-ate A particles of poliovirus [12–15], confirming its identityas virus uncoating intermediate and showing its infectivity.Cryo-electron microscopy allowed reconstruction of virionsbinding to membrane surfaces as well as demonstrated thesite of RNA release near the twofold axis on the proteinaceousviral capsid. However, CE analysis of intermediate A particlesof HRV was previously only reported once in the context of thekinetics of thermal denaturation of the virus in the presenceof antiviral capsid binders [16] without going into more detailon particle characterization or formation conditions. There-fore, in the course of reconstruction of intermediate particlesof rhinovirus uncoating from cryo-electron microscopy im-ages (Pickl-Herk, A. et al., manuscript in preparation) wefurther concentrated on the characterization of particles de-tected upon CE.
In our previous work we found out that the uncoatingprocess to B via A particles could be directed as follows. Heatexposure (to 56�C, 10 min) of undiluted virus stock samplesled mainly to A particles, whereas the same procedure carriedout after dilution of the initial stock with sample buffer (SB)(100 mM sodium borate, pH 8.3) resulted mainly in B parti-cles. Acidification (to pH 5.8 or lower) led mainly to the forma-tion of A particles. A particles obtained from sample heatingand acidification were electrophoretically indistinguishable,however, seemingly both triggers of viral conversion acteddifferently on virus particles.
We were further able to differentiate two intermedi-ate A particles by CE, which we termed AI and AII [17]:AI particles exhibited an average electrophoretic mobility of−17.2 ± 0.4 × 10−9 m2/Vs at 20�C (n = 21); AII particles weremuch less stable and exhibited an electrophoretic mobility of−13.0 × 10−9 m2/Vs at 20�C. Furthermore, it is of note thatAII particles appear more heterogeneous (compared to AIparticles) as could be learned from their broader mobility dis-tribution (SD of the mobility was ±1.4, n = 11, 20�C). Wecould demonstrate that in the course of uncoating native viri-ons convert to AI particles; AII particles seemed to comprise
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

1602 X. Subirats et al. Electrophoresis 2013, 34, 1600–1609
another stage of virus uncoating. However, if AII particlesconstitute an intermediate of virus uncoating between theAI stage and 80S particles or if these particles can be foundalongside AI particles, is still under investigation. In all casesendpoints of virus conversion are subviral B particles de-void of VP4 as well as viral RNA. In the course of the presentmanuscript we focused on AI intermediates. From the sum ofthe experimental results concerning the particle compositionwe find AI particles of our research work indistinguishablefrom subviral A particles of rhinovirus uncoating as previ-ously described [2]. Additional experiments targeted possiblefactors influencing the formation of intermediate AI particlesas well as its antigenicity.
2 Materials and methods
2.1 Chemicals and reagents
Boric acid (99.99%), SDS (reagent plus grade), magne-sium chloride hexahydrate (bioreagent) for CE measure-ments, His-select R© Nickel Affinity Gel (suspension in 30%ethanol/water) and acetic acid were purchased from SigmaAldrich (Steinheim, Germany). Sodium hydroxide (pelletspure) as well as sodium acetate, benzoic acid, magnesiumchloride hexahydrate for ICP-MS measurements, sodiumchloride and calcium chloride dihydrate (all pro-analysis(p.a.)) as well as hydrochloric acid (37%, EMSURE ACS grade)were obtained from E. Merck (Darmstadt, Germany). EDTAwas purchased from Loba Chemie (Fischamend, Austria).The cell-permeant RNA selective fluorescent dye Syto RNA-Select (5 mM solution in DMSO, �ex/em = 490/530 nm)was from Molecular Probes (Eugene, OR, USA). Thesit R©
(polyethylene glycol dodecyl ether, membrane researchgrade), Atto 495 (free acid form of fluorophore, �ex/em =495/527 nm from Atto-Tec, Siegen, Germany), and DMSO(p.a.) were obtained from Fluka (Buchs, Switzerland). Stron-tium ICP standard (10 g/L in diluted nitric acid) was from theVWR Prolabo product range (Vienna, Austria). Water was ofMillipore grade (18.2 M� cm resistivity at 25�C).
2.2 Buffers
HRV2 stock solutions were in 50 mM sodium borate,pH 7.4 (preparation buffer, PB). BGE for CE separationswas 100 mM sodium borate, pH 8.3 additionally includingeither 10 mM Thesit R© or 10 mM SDS as surfactant. SBwas detergent-free BGE. BGE and SB were filtered (celluloseacetate membrane, 0.20 �m pore size syringe filters, Sarto-rius, Gottingen, Germany) and spinned (1 min at 800 rcfon a tabletop centrifuge, 5415D from Eppendorf, Hamburg,Germany) prior to CE. BGE employed for RNA release exper-iments additionally contained the fluorescent dye Syto RNA-Select at 0.5 �M concentration besides Thesit R© (BGE-LIF).Sample acidification was via sodium acetate buffer (50 mM,pH 5.0).
2.3 Biological material
RNase A from bovine pancreas (50 U/mg) was from Roche(Mannheim, Germany), the RNase inhibitor, recombinantRNasin (40 U/�L), from Promega (Madison, WI, USA).HRV2 was prepared and the concentration and purity of var-ious batches assessed by CE [18, 19]. In short, HRV2 wasgrown in HeLa cells (suspension culture), which were bro-ken up via repeated freezing/thawing cycles. After pelletingof cell debris, virions were found in the supernatant fromwhich they were pelleted prior to resuspension in 20 mMTris-HCl, pH 7.5 including 2 mM magnesium chloride. Fol-lowing steps included incubation with RNase, DNase (10 minat ambient temperature each), trypsin (5 min at 37�C), andN-laurylsarcosine (over night at 4�C). By slight changes in theHRV2 preparation protocol we were able to obtain a highlypure, contaminant-free virus batch (Weiss et al., manuscriptin preparation). Virions were pelleted and applied to sucrosedensity centrifugation (7.5–45% sucrose in 20 mM Tris-HCl,pH 7.5 including 2 mM magnesium chloride). HRV2 con-taining fractions were recovered; virions were again pelletedand resuspended in PB. A typical HRV2 preparation batchmeasured around 0.5 �M intact virions. The mAb 2G2 stockwas at 9.0 mg/mL concentration.
2.4 Instrumentation
CE instrumentation was as described [17]. In short,CE with UV detection at indicated wavelengths was on3D CE instruments (Agilent, Waldbronn, Germany) em-ploying 50 �m id, 375 �m od fused silica capillaries(Polymicro, Phoenix, USA obtained from Optronis, Kehl,Germany) of Ltot/eff = 59.9 ± 0.1/51.4 ± 0.1 cm at 25 kV(approx. 42 kV/m field strength) positive polarity at indi-cated temperatures. Capillary preconditioning was by BGE for2 min, postconditioning by 1 M sodium hydroxide and wa-ter, 2 min, respectively. Sample application was by 50 mbarpressure for indicated times. For detection of fluorescent an-alytes, a capillary of Ltot/eff = 72.0/57.0 cm at 25 kV (approx.35 kV/m field strength) positive polarity at 25�C was em-ployed with a ZETALIF FL detection system (Picometrics,Ramonville St. Agne, France) equipped with an Argon laser(�ex = 488 nm). In case of UV detection, electropherogramswere aligned to the DMSO peak and the peak of the internalstandard according to [20]. Measurements were carried outat least in duplicate. When reasonable, a higher number ofmeasurements per sample was carried out.
ICP-MS analyses were carried out on an XSeries2 instru-ment from Thermo Fisher Scientific (Waltham, MA, USA)in duplicate. Standards and samples were introduced to thesystem at constant volumes (approx. 80 �L) via a sampleloop, a standard nebulizer, and a Peltier cooled spray cham-ber. Corresponding signals (24Mg, 25Mg, 26Mg, and 88Sr)were recorded over time (forward power: 1400 W; cool gas:13 Lpm, auxiliary gas: 0.80 Lpm, nebulizer gas: 0.93 LpmArgon (99.999% from Messer, Gumpoldskirchen, Austria),
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

Electrophoresis 2013, 34, 1600–1609 CE and CEC 1603
respectively). Areas of recorded peaks were corrected to theinternal standard (88Sr) and the resulting Mg/Sr intensity ra-tios related to given standard concentrations to obtain a linearcalibration in the range of 0.2–2.0 mM magnesium.
2.5 Sample preparation
Samples prepared for CE analysis typically valued in the rangeof several tens of nM HRV2 (with DMSO as neutral markerin the dilution range of around 1.3 × 103 and benzoic acid(in the range of 20 �g/mL) as internal standard). Additionof salts or EDTA was from higher concentrated stock solu-tions (to reduce virus stock dilution to a minimum) to indi-cated values prior to virus conversion. mAb addition for CEanalysis was at approximately 30-fold molar excess of 2G2over viral and subviral particles, respectively. Denaturationof virus was either by heating (10 min at 56�C in a waterbath) or by acidification via acetate buffer to pH 5.0. Undi-luted HRV2 stock solutions yielded highest amounts of AIparticles when heated as described. After dilution of the virusstock solution heat denaturation yielded mainly 80S parti-cles. Acidification resulted mainly in AI particle formation[17].
For analysis of the protein content, HRV2 was heat de-natured to yield AI particles. Storage solution of His-select R©
Nickel Affinity Gel was replaced by half concentrated SB.Spin size exclusion chromatography (SEC) was carried outas described [21] on Sephadex G50 (GE Healthcare, Uppsala,Sweden) after incubation of heat denatured HRV2 with sol-uble fragments of a recombinant receptor based on bindingdomain 3 of the very low-density lipoprotein receptor andgrafted with a His6-tag at its C-, and maltose binding proteinat its N-terminus (MBP-V33333) known to bind intact virions[22] as well as His-select R© Nickel Affinity Gel. Such, intactvirions were removed from the incubation solution (receptormediated complexation of intact virions to Ni2+ presentingagarose gel and retardation on filter) as well as protein mate-rial released from virus particles upon uncoating (1.5–30 kDafractionation range of sieving material for globular proteinsaccording to the manufacturing company). The spin fractionshowed highest amounts of AI particles after CE was sub-jected to further heat denaturation after additional dilutionin half concentrated SB including SDS (final SDS concentra-tion of sample at 8.5 mM). For comparison to intact virions,HRV2 stock solution was diluted 1: 20 v/v in BGE containingSDS and likewise heat denatured.
For experiments including RNase A digestion, HRV2was denatured by acetate buffer and incubated with RNaseinhibitor (final concentration 2 U/�L) and subsequently withRNase A (final concentration 0.008 U/�L).
For ICP-MS measurements magnesium chloride stan-dards were prepared in PB in the range of 0.2–2.0 mM (fivedata points including a matrix matched blank). Standards aswell as analyzed HRV2 preparations were consecutively di-luted 1:100 v/v in 5% HCl including strontium as internalstandard at a final strontium concentration of 50 mg/L.
3 Results and discussion
In our previous work we reported that sample heating allowedus to direct virus conversion to both, intermediate AI particlesor subviral B particles, respectively [17]. Sample acidification,on the other hand, mainly yielded AI particles, independentof applied incubation times (between 1 and 10 min) or pHvalues (between pH 5.8 and 5.0). Electrophoretic mobilitiesof intermediate AI particles were identical for both virus con-version techniques, heating or sample acidification.
Furthermore, we found the amount of AI particles thatcan be formed upon virus uncoating by sample heatingstrongly varying from one individual virus preparation to theother. Most surprisingly, the degree of sample dilution priorto heat denaturation (not the virus concentration!) directedthe outcome of viral conversion to empty capsids (subviralB particles). The more the initial virus preparation was di-luted, the lower was the amount of AI in relation to B parti-cles. Note that the virus concentration of the individual initialpreparations varied by nearly half an order of magnitude, al-though the preparation of virions from cell culture was iden-tical for all virus batches. We therefore assumed that anothersample component/other sample components might be re-sponsible for the stabilization of AI particles, which eitheroriginated from cell culture and were copurified with HRV2or originated from the buffers used in the virus preparationand purification procedures. Components of the latter weremainly sucrose (from gradient centrifugation), and some or-ganic (Tris) and inorganic (e.g., Mg2+) ions (see Section 2).Initial experiments allowed us to exclude sucrose to interferewith virus conversion (results not shown). Upon applicationof an orthogonal analysis method, gas phase electrophoreticmobility molecular analysis (GEMMA), which separates par-ticles according to their electrophoretical mobility diameterin the gas phase at ambient pressure, we could identify anadditional sample component, in the same overall size rangeas virions, however much more heterogeneous [17]. This ad-ditional sample component we could relate to the so-calledcontaminant from virus preparations that we previously de-tected by CE analysis [18,23] and which we found to be identi-cal to irregular shaped particles as found upon transmissionelectron microscopy (TEM) imaging [24, 25]. Further charac-terization of the contaminating material is at the momentstill ongoing. In the course of the present work we thereforeconcentrated on the contaminant (which we suspect as mate-rial co-purified with virions) and on ionic buffer constituentsas additional factors influencing AI particle formation.
3.1 The role of the contaminant on stabilization
of AI particles
Changes in the HRV2 preparation protocol (Weiss V.U. et al.,manuscript in preparation) allowed us to generate a highlypure, contaminant-free virus preparation, which allowed usto target the effect of the contaminant on the in vitro for-mation of intermediate AI and subviral B particles. Figure 2
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

1604 X. Subirats et al. Electrophoresis 2013, 34, 1600–1609
Figure 2. AI particles are formed in vitro upon uncoating in theabsence of the “contaminant” of HRV2 preparations. This sam-ple does not contain the so-called contaminant, a constituent withhigh molecular mass, which is additionally present in virus prepa-rations prepared after standard protocols. (A) CE record of the“contaminant”-free pure virus preparation. (B) Upon heat denat-uration (10 min at 56�C) of such a diluted sample mainly emptycapsids (80S or subviral B particles) are obtained. The positionof the contaminant from standard virus preparations is indicatedby a highlighted background. (C) Heat denaturation of the HRV2stock dilution prior to dilution results in formation of AI instead ofB particles. (D) The formation of AI intermediates can be hinderedby addition of EDTA (e.g., 30 mM as shown) to the sample beforetriggering virus uncoating. Therefore the conversion of 150S toAI particles appears to be dependent on the presence of diva-lent metal cations. CE conditions: BGE, 100 mM sodium borate,pH 8.3, 10 mM Thesit; temperature 20�C, voltage 25 kV (approx.42 kV/m). Injection to CE was at 50 mbar pressure for 9 s. Detec-tion by UV absorption at 200 nm (obtained signals in mAU). Fortrace (A), initial virus stock solution was diluted 1: 20 in 100 mMsodium borate, pH 8.3. DMSO (1: 4 × 103) as neutral marker aswell as benzoic acid (in the range of 20 �g/mL) as internal stan-dard (i.s.) were added. To obtain AI particles, 0.6 �L HRV2 stockwas mixed with 0.3 �L SB (C) or 90 mM EDTA solution in SB(D), respectively, prior to sample heating (10 min, 56�C); samplewas thus diluted 1:1.5. For CE analysis, the sample was dilutedto HRV2 concentrations comparable to samples presented in(A) and (B). The spikes are indicative for particle aggregates.
depicts resulting electropherograms (aligned to the DMSOpeak and to the internal standard according to [20]); intactvirions (Fig. 2A) and B particles (Fig. 2B) migrate with sim-ilar, whereas distinguishable electrophoretic mobilities (seetable 1 in [17] corresponding to part I). Also the formationof intermediate AI particles was detectable (Fig. 2C). It is ofnote that sample heating (10 min to 56�C) was employed totrigger conversion from 150S to AI particles and that theseconditions did not lead to a complete conversion of all intactvirions. Nevertheless, since AI particles were recorded in thiscontaminant-free sample, we no longer suspect the contam-inant for playing a significant role in the formation of inter-mediates of virus uncoating. As a consequence we focusedon ionic buffer constituents and their impact on subviral AIparticles stabilization.
3.2 Mg2+ and other metal cations support formation
of intermediate AI particles
ICP-MS measurements of HRV2 stock solutions showed thatvirus preparations still contained magnesium ions in the mMrange (two preparations were tested yielding 2.06 ± 0.02 and1.76 ± 0.01 mM magnesium ions, respectively). However,since virions were pelleted after sucrose density centrifuga-tion (7.5–45% sucrose in 20 mM Tris-HCl, pH 7.5, including2 mM magnesium chloride) followed by resuspension in PBnot containing any additional magnesium ions, this findingindicated already some interaction between virions and cor-responding divalent cations. Furthermore, when repeatingthe formation of AI particles as depicted in Fig. 2C in thepresence of EDTA (Fig. 2D), no AI intermediates could bedetected. Therefore, the transition of intact virions to the in-termediate stage of virus uncoating seems to be dependenton the presence of divalent cations. Additional experimentsconfirmed this assumption: the formation of AI particles wassensitive to the presence of divalent cations in the SB as well asto increased ionic strengths as demonstrated for sample acid-ification in Fig. 3. Presence of magnesium chloride (Fig. 3C)or calcium chloride (Fig. 3B) led to a significant stabilizationof AI particles, and a reduced production of B particles. Thisfinding is in accordance with data available for coxsackievirus[26], rhinovirus serotype 2 [27], and poliovirus [28] showingeither (i) a decreasing amount of infective virions in plaqueassays upon dilution in a magnesium chloride containingbuffer (which was attributed to an increasing amount of in-termediate and not empty particles) or (ii) increasing amountsof 135S particles upon sucrose gradient centrifugation afterincubation with calcium chloride containing buffer or cellgrowth medium also containing divalent cations. However,application of CE analyses allows the exact determination ofconversion conditions for AI particles than by sample cen-trifugation on sucrose gradients previously described (e.g.,[13]).
Addition of sodium chloride (Fig. 3A) to the SB also ledto a significant reduction of the formation of B particles asfound already for 33 mM sodium chloride. Further additionof sodium chloride decreased the amount of these particleseven more. However, concomitantly, the peak shape for AIparticles as well as for the internal standard was distortedmost probably due to the strongly differing electric conduc-tance of the sample plug in relation to the BGE employed.Nevertheless, at addition of 333 mM sodium chloride to theSB, 80S particles could no longer be detected alongside inter-mediate AI particles.
On the exact mechanism of cationic stabilization of AIparticles we can only speculate at the moment. Possiblereasons for AI particle stabilization include interaction ofcations with the virus capsid, the RNA, or both, stabilizingtertiary and quaternary structures at the virus intermediateuncoating stage. However, we conclude from these find-ings that the presence of divalent metal cations and othersalts, and not of the contaminant or of nonionic componentslike sucrose found in HRV2 stock solutions from the virus
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

Electrophoresis 2013, 34, 1600–1609 CE and CEC 1605
Figure 3. CE depicting the influence of mono- and divalent cationaddition to the sample on the formation of AI particles. The influ-ence of the addition of magnesium chloride (C), calcium chloride(B), and sodium chloride (A) to the sample buffer (SB) duringvirus conversion via sample acidification (pH 5.0) was assessed.Increased addition of divalent or (to a far lesser extent) monova-lent cations decreased the amount of detected empty viral cap-sids (B particles, indicated by arrows) indicating stabilization ofAI particles during viral uncoating. Therefore, the conversion ofAI to B particles appears to be hindered by divalent cations. Highsalt concentrations led to peak distortions due to differences inthe electric conductance between sample plug and BGE. CE wasas in Fig. 2 with the exception of 8 s sample injection and elec-trophoresis at 25�C. Initial virus preparations were diluted 1:10 in100 mM sodium borate, pH 8.3, additionally adding chloride saltsas indicated prior to incubation at pH 5.0 at ambient temperaturefor 30 min. Samples were reneutralized prior to CE analysis andDMSO (1:4 × 103) as neutral marker and benzoic acid (around20 �g/mL) as i.s. were added.
preparation process are responsible for AI particle stabiliza-tion during virus uncoating. The recorded electrophoreticmobility (−17.1 ± 0.4 × 10−9 m2/Vs at 20�C (n = 21) af-ter sample heating and −17.3 ± 0.4 × 10−9 m2/Vs at 20�C(n = 21) after sample acidification, respectively) of AI particlesin a BGE including the nonionic detergent Thesit R© addition-ally supports this conclusion; As AI particles migrate with amuch higher electrophoretic net mobility than intact virionsor 80S particles [17], more negative charges are accessibleon the intermediate AI particle surface (mind that the parti-cle diameter does not change significantly for virus particlesduring uncoating) for interaction with cations. The decreaseof the concentration of such ions upon sample dilution mostprobably leads to the reduced recorded number of AI particlesas previously described [17].
3.3 AI particle composition
After heat denaturation in BGE containing SDS as deter-gent, CE analysis of the decomposition products of the virionsyields peaks for the four VPs and the viral RNA, respectively[29]. As stated, intermediates of viral uncoating sedimentingat 135S still include viral RNA but have already lost one of thefour VPs, VP4. In order to get insight into the compositionof AI particles, we prepared them by heating a HRV2 stocksolution, and removed still remaining intact virions and freeproteins. The intact virions were complexed to His6-taggedrecombinant receptor fragments followed by binding to Ni2+
presenting agarose beads. Upon application of spin SEC tothis mixture, bound native 150S virions as well as free smallerproteins like VP4 (with 7.35 kDa, according to [30]) were sep-arated from subviral assemblies (see Section 2). The resultingfractions were checked by CE and the fraction showing thehighest content of AI was subjected after addition of SDS tofurther heating. Under these conditions, the subviral particlesare decomposed into the capsid proteins and RNA. The ob-tained electropherogram is presented in Fig. 4 as dashed linein comparison to the decay products formed from 150S viri-ons after thermal denaturation in the same BGE (solid line).It can be seen that both, purified AI samples as well as 150Svirions, exhibit comparable peak patterns, with the importantexception of one VP peak, which is missing with AI (indicatedby the gray background). As this peak had previously been as-signed to VP4 [29] it is clear that this protein is not part ofAI particles. Additionally, the peak assigned to the viral RNAwas slightly diminished in the AI sample, probably causedby the partial RNA degradation. Such degradation can hardlybe avoided in the course of sample preparation due to om-nipresent RNases. Concomitantly, several smaller peaks werefound in the electropherogram (e.g., at 7.4 min migrationtime), which were suggested to correspond to smaller RNAfragments, products from the enzymanic RNA degradation.
3.4 RNA content of AI particles
We observed that in the presence of RNase inhibitor, incu-bation of HRV2 at the low pH of 5.0 led to the formation
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com
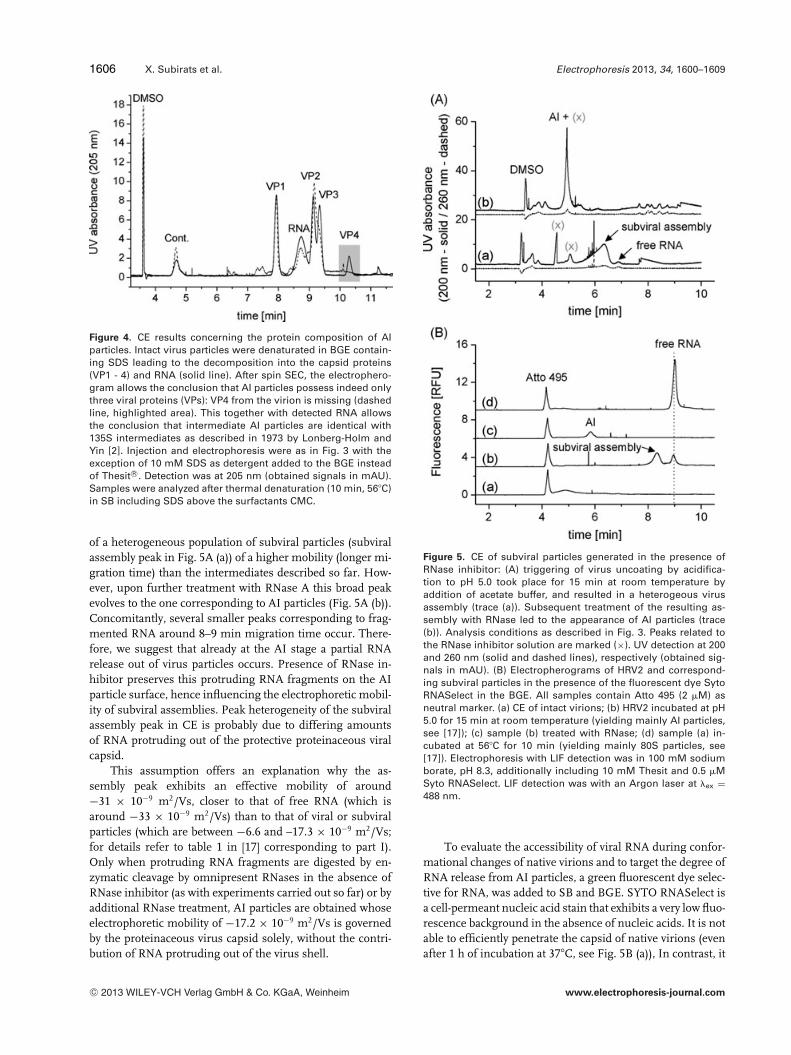
1606 X. Subirats et al. Electrophoresis 2013, 34, 1600–1609
Figure 4. CE results concerning the protein composition of AIparticles. Intact virus particles were denaturated in BGE contain-ing SDS leading to the decomposition into the capsid proteins(VP1 - 4) and RNA (solid line). After spin SEC, the electrophero-gram allows the conclusion that AI particles possess indeed onlythree viral proteins (VPs): VP4 from the virion is missing (dashedline, highlighted area). This together with detected RNA allowsthe conclusion that intermediate AI particles are identical with135S intermediates as described in 1973 by Lonberg-Holm andYin [2]. Injection and electrophoresis were as in Fig. 3 with theexception of 10 mM SDS as detergent added to the BGE insteadof Thesit R©. Detection was at 205 nm (obtained signals in mAU).Samples were analyzed after thermal denaturation (10 min, 56�C)in SB including SDS above the surfactants CMC.
of a heterogeneous population of subviral particles (subviralassembly peak in Fig. 5A (a)) of a higher mobility (longer mi-gration time) than the intermediates described so far. How-ever, upon further treatment with RNase A this broad peakevolves to the one corresponding to AI particles (Fig. 5A (b)).Concomitantly, several smaller peaks corresponding to frag-mented RNA around 8–9 min migration time occur. There-fore, we suggest that already at the AI stage a partial RNArelease out of virus particles occurs. Presence of RNase in-hibitor preserves this protruding RNA fragments on the AIparticle surface, hence influencing the electrophoretic mobil-ity of subviral assemblies. Peak heterogeneity of the subviralassembly peak in CE is probably due to differing amountsof RNA protruding out of the protective proteinaceous viralcapsid.
This assumption offers an explanation why the as-sembly peak exhibits an effective mobility of around−31 × 10−9 m2/Vs, closer to that of free RNA (which isaround −33 × 10−9 m2/Vs) than to that of viral or subviralparticles (which are between −6.6 and –17.3 × 10−9 m2/Vs;for details refer to table 1 in [17] corresponding to part I).Only when protruding RNA fragments are digested by en-zymatic cleavage by omnipresent RNases in the absence ofRNase inhibitor (as with experiments carried out so far) or byadditional RNase treatment, AI particles are obtained whoseelectrophoretic mobility of −17.2 × 10−9 m2/Vs is governedby the proteinaceous virus capsid solely, without the contri-bution of RNA protruding out of the virus shell.
Figure 5. CE of subviral particles generated in the presence ofRNase inhibitor: (A) triggering of virus uncoating by acidifica-tion to pH 5.0 took place for 15 min at room temperature byaddition of acetate buffer, and resulted in a heterogeous virusassembly (trace (a)). Subsequent treatment of the resulting as-sembly with RNase led to the appearance of AI particles (trace(b)). Analysis conditions as described in Fig. 3. Peaks related tothe RNase inhibitor solution are marked (×). UV detection at 200and 260 nm (solid and dashed lines), respectively (obtained sig-nals in mAU). (B) Electropherograms of HRV2 and correspond-ing subviral particles in the presence of the fluorescent dye SytoRNASelect in the BGE. All samples contain Atto 495 (2 �M) asneutral marker. (a) CE of intact virions; (b) HRV2 incubated at pH5.0 for 15 min at room temperature (yielding mainly AI particles,see [17]); (c) sample (b) treated with RNase; (d) sample (a) in-cubated at 56�C for 10 min (yielding mainly 80S particles, see[17]). Electrophoresis with LIF detection was in 100 mM sodiumborate, pH 8.3, additionally including 10 mM Thesit and 0.5 �MSyto RNASelect. LIF detection was with an Argon laser at �ex =488 nm.
To evaluate the accessibility of viral RNA during confor-mational changes of native virions and to target the degree ofRNA release from AI particles, a green fluorescent dye selec-tive for RNA, was added to SB and BGE. SYTO RNASelect isa cell-permeant nucleic acid stain that exhibits a very low fluo-rescence background in the absence of nucleic acids. It is notable to efficiently penetrate the capsid of native virions (evenafter 1 h of incubation at 37�C, see Fig. 5B (a)), In contrast, it
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

Electrophoresis 2013, 34, 1600–1609 CE and CEC 1607
can be concluded from Fig. 5B (b), that RNA protruding fromintermediate particles as found in the subviral assembly aswell as free RNA in solution (Fig. 5B (d)) is readily labeled.Moreover, penetration into AI particles occurs (Fig. 5B (c)).These findings support the assumption of a partial RNA re-lease from AI particles. However, since AI particles still showsome residual fluorescence (Fig. 5B (c)), it can be followedthat a part of the RNA still remains within AI particles and isthus protected from enzymatic cleavage.
A most relevant question arising from the results pre-sented targets the amount of viral RNA still encapsulatedwithin AI particles (it is of note, that 135S particles obtainedby Lonberg-Holm and Yin [2] were described to contain thewhole viral RNA genome). We tried to answer this questionby comparing the ratios of peak areas obtained by UV ab-sorption at 205 and 260 nm for the following reasons. At205 nm, UV absorption is mainly caused by peptide bonds –most proteins exhibit a very similar absorbance coefficient atthis wavelength [31] – whereas at 260 nm absorption is dueto the nitrogenous bases in nucleotides (RNA in the presentcase). Based on the integrated peak areas of free and encap-sulated RNA from our own electropherograms recorded at260 nm, and in accordance to reference [32], we assumed thatthe absorbance of encapsulated RNA does not differ substan-tially from that of the free polynucleotide. For intact HRV2 theratio of peak areas measured at 205 and 260 nm, A205/A260,from 12 different preparations was 5.5 (±0.1), and the cal-culations based on peaks migrating between 7 and 9 minfrom RNA fragments released from the viral genome leadto a A205/A260 peak area ratio of 1.4 (±0.1). The correspond-ing value for AI particles from three different preparationswas 5.5 (±0.2), revealing nearly the same ratio as for nativeHRV2. If we subtract the contribution of VP4 (again basedon data from our own electropherograms) to the absorbancefrom that of the native virion, the estimated ratio for AI the-oretically is 5.2 (±0.1). Thus the measured ratio A205/A260
of 5.5 for AI was slightly, but significantly higher than theestimated value of 5.2, suggesting a partial release of RNA.Considering that the absorbance of AI particles at 205 nm(AAI
205) is the sum of capsid proteins (Acapsid205 ) and RNA (ARNA
205 )contributions, and that ARNA
205 /ARNA260 = 1.4, it follows:
AAI205 = Acapsid
205 + 1.4ARNA260 (1)
Provided that the absorbance of the capsid at 260 nm is neg-ligible, the measured and estimated AAI
205/AAI260 ratios are, re-
spectively:
(Acapsid205 )measured + 1.4(ARNA
260 )measured
(ARNA260 )measured
= 5.5 (2)
(Acapsid205 )estimated + 1.4(ARNA
260 )estimated
(ARNA260 )estimated
= 5.2 (3)
Then, assuming that the absorbance of the measured andestimated contribution of the viral capsid is the same,(Acapsid
205 )measured = (Acapsid205 )estimated, Eqs. (2) and (3) could be
combined and expressed as:
5.5(ARNA
260
)measured
− 1.4(ARNA
260
)measured
= 5.2(ARNA
260
)estimated
− 1.4(ARNA
260
)estimated
(4)
and after simplification:(ARNA
260
)measured
= 0.93(ARNA
260
)estimated
(5)
We can thus conclude that the contribution of the RNAto the UV absorbance for the AI particles is 7% lower than itwere if the particles would contain the complete genom. Withother words, we suggest that 93% of the initial RNA remainsin average encapsulated within AI particles.
3.5 Comparison of the biological activity
Endpoints of viral conversion, that is, B particles sedimentingat 80S, were shown to bind mAb 2G2, leading to aggregationof the subviral particle: the peak of B particles in Fig. 6E ismissing in Fig. 6F as a result of binding to mAb 2G2 andsubsequent removal from CE samples (see also [5]). Intactvirions, on the other hand, were not susceptible to bindingto this antibody (compare Fig. 6A and B, indicated by a graybackground and [5]). Thus, mAb 2G2 binds to regions on thevirus capsid undergoing gross changes upon virus conver-sion. In fact, the antigenic determinant was found to be atthe star-shaped mesa on the empty viral capsid after VP1 re-arrangement during virus conversion [6]. We therefore tookinterest in binding of mAb 2G2 toward AI particles. It can beconcluded from comparison of Fig. 6C with D (peak positionof AI is indicated by a gray background) that AI particles wereprecipitated by mAb 2G2 as well. It follows that antigenic de-terminants for binding of mAb 2G2 are already expressed onthe viral capsid upon VP4 egress.
4 Concluding remarks
CE of intermediate particles of virus conversion from nativevirions (150S particles) to empty capsids (80S or subviral Bparticles) enabled us to differentiate two intermediate speciesexhibiting an average electrophoretic mobility of about−17.2 × 10−9 m2/Vs or −13.0 × 10−9 m2/Vs, respectively,at 20�C, which we termed AI and AII particles [17]. In thecourse of our present research we were able to demonstratethe influence of divalent (like Mg2+ or Ca2+) and to a farlesser extent of monovalent cations (like Na+) on the forma-tion of intermediate AI particles during rhinovirus uncoating.Upon complexation of divalent cations by EDTA the transi-tion of virions from the 150S to the AI stage was hindered.On the other hand, addition of divalent cations like Mg2+ orCa2+ or addition of sodium chloride led to stabilization ofAI particles. However, in those latter cases transition of AIparticles to empty viral capsids (80S particles) was limited.Therefore, it appears that the presence and the concentrationof especially divalent cations are crucial for the formation of
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com
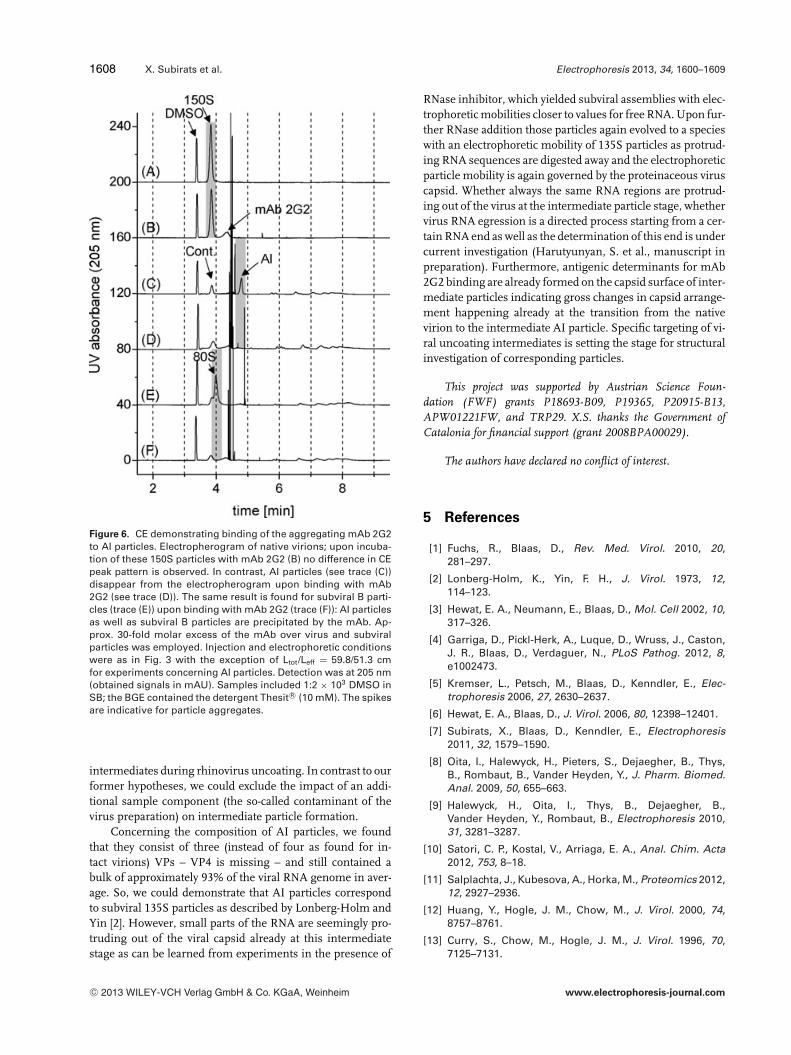
1608 X. Subirats et al. Electrophoresis 2013, 34, 1600–1609
Figure 6. CE demonstrating binding of the aggregating mAb 2G2to AI particles. Electropherogram of native virions; upon incuba-tion of these 150S particles with mAb 2G2 (B) no difference in CEpeak pattern is observed. In contrast, AI particles (see trace (C))disappear from the electropherogram upon binding with mAb2G2 (see trace (D)). The same result is found for subviral B parti-cles (trace (E)) upon binding with mAb 2G2 (trace (F)): AI particlesas well as subviral B particles are precipitated by the mAb. Ap-prox. 30-fold molar excess of the mAb over virus and subviralparticles was employed. Injection and electrophoretic conditionswere as in Fig. 3 with the exception of Ltot/Leff = 59.8/51.3 cmfor experiments concerning AI particles. Detection was at 205 nm(obtained signals in mAU). Samples included 1:2 × 103 DMSO inSB; the BGE contained the detergent Thesit R© (10 mM). The spikesare indicative for particle aggregates.
intermediates during rhinovirus uncoating. In contrast to ourformer hypotheses, we could exclude the impact of an addi-tional sample component (the so-called contaminant of thevirus preparation) on intermediate particle formation.
Concerning the composition of AI particles, we foundthat they consist of three (instead of four as found for in-tact virions) VPs – VP4 is missing – and still contained abulk of approximately 93% of the viral RNA genome in aver-age. So, we could demonstrate that AI particles correspondto subviral 135S particles as described by Lonberg-Holm andYin [2]. However, small parts of the RNA are seemingly pro-truding out of the viral capsid already at this intermediatestage as can be learned from experiments in the presence of
RNase inhibitor, which yielded subviral assemblies with elec-trophoretic mobilities closer to values for free RNA. Upon fur-ther RNase addition those particles again evolved to a specieswith an electrophoretic mobility of 135S particles as protrud-ing RNA sequences are digested away and the electrophoreticparticle mobility is again governed by the proteinaceous viruscapsid. Whether always the same RNA regions are protrud-ing out of the virus at the intermediate particle stage, whethervirus RNA egression is a directed process starting from a cer-tain RNA end as well as the determination of this end is undercurrent investigation (Harutyunyan, S. et al., manuscript inpreparation). Furthermore, antigenic determinants for mAb2G2 binding are already formed on the capsid surface of inter-mediate particles indicating gross changes in capsid arrange-ment happening already at the transition from the nativevirion to the intermediate AI particle. Specific targeting of vi-ral uncoating intermediates is setting the stage for structuralinvestigation of corresponding particles.
This project was supported by Austrian Science Foun-dation (FWF) grants P18693-B09, P19365, P20915-B13,APW01221FW, and TRP29. X.S. thanks the Government ofCatalonia for financial support (grant 2008BPA00029).
The authors have declared no conflict of interest.
5 References
[1] Fuchs, R., Blaas, D., Rev. Med. Virol. 2010, 20,281–297.
[2] Lonberg-Holm, K., Yin, F. H., J. Virol. 1973, 12,114–123.
[3] Hewat, E. A., Neumann, E., Blaas, D., Mol. Cell 2002, 10,317–326.
[4] Garriga, D., Pickl-Herk, A., Luque, D., Wruss, J., Caston,J. R., Blaas, D., Verdaguer, N., PLoS Pathog. 2012, 8,e1002473.
[5] Kremser, L., Petsch, M., Blaas, D., Kenndler, E., Elec-trophoresis 2006, 27, 2630–2637.
[6] Hewat, E. A., Blaas, D., J. Virol. 2006, 80, 12398–12401.
[7] Subirats, X., Blaas, D., Kenndler, E., Electrophoresis2011, 32, 1579–1590.
[8] Oita, I., Halewyck, H., Pieters, S., Dejaegher, B., Thys,B., Rombaut, B., Vander Heyden, Y., J. Pharm. Biomed.Anal. 2009, 50, 655–663.
[9] Halewyck, H., Oita, I., Thys, B., Dejaegher, B.,Vander Heyden, Y., Rombaut, B., Electrophoresis 2010,31, 3281–3287.
[10] Satori, C. P., Kostal, V., Arriaga, E. A., Anal. Chim. Acta2012, 753, 8–18.
[11] Salplachta, J., Kubesova, A., Horka, M., Proteomics 2012,12, 2927–2936.
[12] Huang, Y., Hogle, J. M., Chow, M., J. Virol. 2000, 74,8757–8761.
[13] Curry, S., Chow, M., Hogle, J. M., J. Virol. 1996, 70,7125–7131.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com

Electrophoresis 2013, 34, 1600–1609 CE and CEC 1609
[14] Bubeck, D., Filman, D. J., Cheng, N., Steven, A. C., Hogle,J. M., Belnap, D. M., J. Virol. 2005, 79, 7745–7755.
[15] Bostina, M., Levy, H., Filman, D. J., Hogle, J. M., J. Virol.2011, 85, 776–783.
[16] Okun, V. M., Nizet, S., Blaas, D., Kenndler, E., Elec-trophoresis 2002, 23, 896–902.
[17] Weiss, V. U., Subirats, X., Pickl-Herk, A., Bilek, G.,Winkler, W., Kumar, M., Allmaier, G., Blaas, D., Kenndler,E., Electrophoresis 2012, 33, 1833–1841.
[18] Okun, V. M., Ronacher, B., Blaas, D., Kenndler, E., Anal.Chem. 1999, 71, 2028–2032.
[19] Hewat, E. A., Neumann, E., Conway, J. F., Moser, R.,Ronacher, B., Marlovits, T. C., Blaas, D., EMBO J. 2000,19, 6317–6325.
[20] Reijenga, J. C., Martens, J. H., Giuliani, A., Chiari, M.,J. Chromatogr. B Analyt. Technol. Biomed Life Sci 2002,770, 45–51.
[21] Weiss, V. U., Bilek, G., Pickl-Herk, A., Blaas, D., Kenndler,E., Electrophoresis 2009, 30, 2123–2128.
[22] Moser, R., Snyers, L., Wruss, J., Angulo, J., Peters, H.,Peters, T., Blaas, D., Virology 2005, 338, 259–269.
[23] Kremser, L., Konecsni, T., Blaas, D., Kenndler, E., Anal.Chem. 2004, 76, 4175–4181.
[24] Bilek, G., Weiss, V. U., Pickl-Herk, A., Blaas, D., Kenndler,E., Electrophoresis 2009, 30, 4292–4299.
[25] Bilek, G., Matscheko, N. M., Pickl-Herk, A., Weiss, V. U.,Subirats, X., Kenndler, E., Blaas, D., J. Virol. 2011, 85,8368–8375.
[26] Cords, C. E., James, C. G., McLaren, L. C., J. Virol. 1975,15, 244–252.
[27] Lonberg-Holm, K., Gosser, L. B., Shimshick, E. J., J. Virol.1976, 19, 746–749.
[28] Wetz, K., Kucinski, T., J. Gen. Virol. 1991, 72 (Pt 10),2541–2544.
[29] Kremser, L., Petsch, M., Blaas, D., Kenndler, E., Elec-trophoresis 2006, 27, 1112–1121.
[30] Kremser, L., Bilek, G., Kenndler, E., Electrophoresis 2007,28, 3684–3690.
[31] Kreusch, S., Schwedler, S., Tautkus, B., Cumme, G. A.,Horn, A., Anal. Biochem. 2003, 313, 208–215.
[32] Porterfield, J. Z., Zlotnick, A., Virology 2010, 407,281–288.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.electrophoresis-journal.com
Copyright © 2022 FDOKUMEN



















