Characterization of primary cilia and Hedgehog signaling during development of the human pancreas...
14
SPECIAL ISSUE RESEARCH ARTICLE Characterization of Primary Cilia and Hedgehog Signaling During Development of the Human Pancreas and in Human Pancreatic Duct Cancer Cell Lines Sonja K. Nielsen, 1 Kjeld Møllgård, 2 Christian A. Clement, 1 Iben R. Veland, 1 Aashir Awan, 1 Bradley K. Yoder, 3 Ivana Novak, 1 and Søren Tvorup Christensen 1 * Hedgehog (Hh) signaling controls pancreatic development and homeostasis; aberrant Hh signaling is associated with several pancreatic diseases. Here we investigated the link between Hh signaling and primary cilia in the human developing pancreatic ducts and in cultures of human pancreatic duct adenocarcinoma cell lines, PANC-1 and CFPAC-1. We show that the onset of Hh signaling from human embryogenesis to fetal development is associated with accumulation of Hh signaling components Smo and Gli2 in duct primary cilia and a reduction of Gli3 in the duct epithelium. Smo, Ptc, and Gli2 localized to primary cilia of PANC-1 and CFPAC-1 cells, which may maintain high levels of nonstimulated Hh pathway activity. These findings indicate that primary cilia are involved in pancreatic development and postnatal tissue homeostasis. Developmental Dynamics 237:2039 –2052, 2008. © 2008 Wiley-Liss, Inc. Key words: primary cilia; pancreas; exocrine duct; development; cancer; Hedgehog signaling; Smoothened; Patched; Gli2; Gli3 Accepted 9 May 2008 INTRODUCTION Primary cilia are microtubule-based organelles that project into the extra- cellular environment from the mother centriole of most quiescent cells in the human body. Recent research has demonstrated that cilia function as sensory units that coordinate a vari- ety of signal transduction pathways essential for embryonic and postnatal development as well as tissue ho- meostasis in the adult body (Chris- tensen et al., 2007; Eggenschwiler and Anderson, 2007; Kiprilov et al., 2008). These pathways include Hedgehog (Hh), Wnt, and platelet-derived growth factor receptor (PDGFR) sig- naling as well as Ca 2 -signaling by means of TRP ion channels. Further- more, primary cilia may control be- havioral responses, such as occurs in the central nervous system where neuronal cilia function in a pathway that controls satiety responses (Dav- enport et al., 2007). Consequently, de- fects in ciliary assembly lead to a plethora of diseases and disorders, in- cluding kidney, liver and pancreatic cysts, blindness, obesity, diabetes, de- velopmental disorders, and other hu- man diseases now collectively referred to as ciliopathies (Badano et al., 2006; Satir and Christensen, 2007; Yoder, 2007; Fliegauf et al., 2007). Primary cilia on the epithelium of re- nal tubules control tissue homeostasis probably by acting as mechanosensors that monitor the composition and flow rate of urine in the nephron (Praetorius and Spring, 2003; Yoder, 2007). Ob- struction of this flow-sensing response in, for example, the IFT88 Tg737Rpw 1 Department of Biology, University of Copenhagen, Copenhagen, Denmark 2 Department of Cellular and Molecular Medicine, The Panum Institute, University of Copenhagen, Copenhagen, Denmark 3 Department of Cell Biology, University of Alabama at Birmingham, Birmingham, Alabama Grant sponsor: National Institutes of Health; Grant number: RO1 R01-HD056030. *Correspondence to: Søren Tvorup Christensen, Department of Biology, Section of Cell and Developmental Biology, The August Krogh Building, University of Copenhagen, Universitetsparken 13, DK-2100 Copenhagen OE, Denmark. E-mail: [email protected] DOI 10.1002/dvdy.21610 Published online 10 July 2008 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 237:2039 –2052, 2008 © 2008 Wiley-Liss, Inc.
Transcript of Characterization of primary cilia and Hedgehog signaling during development of the human pancreas...

SPECIAL ISSUE RESEARCH ARTICLE
Characterization of Primary Cilia andHedgehog Signaling During Development ofthe Human Pancreas and in Human PancreaticDuct Cancer Cell LinesSonja K. Nielsen,1 Kjeld Møllgård,2 Christian A. Clement,1 Iben R. Veland,1 Aashir Awan,1
Bradley K. Yoder,3 Ivana Novak,1 and Søren Tvorup Christensen1*
Hedgehog (Hh) signaling controls pancreatic development and homeostasis; aberrant Hh signaling isassociated with several pancreatic diseases. Here we investigated the link between Hh signaling andprimary cilia in the human developing pancreatic ducts and in cultures of human pancreatic ductadenocarcinoma cell lines, PANC-1 and CFPAC-1. We show that the onset of Hh signaling from humanembryogenesis to fetal development is associated with accumulation of Hh signaling components Smo andGli2 in duct primary cilia and a reduction of Gli3 in the duct epithelium. Smo, Ptc, and Gli2 localized toprimary cilia of PANC-1 and CFPAC-1 cells, which may maintain high levels of nonstimulated Hh pathwayactivity. These findings indicate that primary cilia are involved in pancreatic development and postnataltissue homeostasis. Developmental Dynamics 237:2039–2052, 2008. © 2008 Wiley-Liss, Inc.
Key words: primary cilia; pancreas; exocrine duct; development; cancer; Hedgehog signaling; Smoothened; Patched;Gli2; Gli3
Accepted 9 May 2008
INTRODUCTION
Primary cilia are microtubule-basedorganelles that project into the extra-cellular environment from the mothercentriole of most quiescent cells in thehuman body. Recent research hasdemonstrated that cilia function assensory units that coordinate a vari-ety of signal transduction pathwaysessential for embryonic and postnataldevelopment as well as tissue ho-meostasis in the adult body (Chris-tensen et al., 2007; Eggenschwiler andAnderson, 2007; Kiprilov et al., 2008).
These pathways include Hedgehog(Hh), Wnt, and platelet-derivedgrowth factor receptor (PDGFR) sig-naling as well as Ca2�-signaling bymeans of TRP ion channels. Further-more, primary cilia may control be-havioral responses, such as occurs inthe central nervous system whereneuronal cilia function in a pathwaythat controls satiety responses (Dav-enport et al., 2007). Consequently, de-fects in ciliary assembly lead to aplethora of diseases and disorders, in-cluding kidney, liver and pancreatic
cysts, blindness, obesity, diabetes, de-velopmental disorders, and other hu-man diseases now collectively referredto as ciliopathies (Badano et al., 2006;Satir and Christensen, 2007; Yoder,2007; Fliegauf et al., 2007).
Primary cilia on the epithelium of re-nal tubules control tissue homeostasisprobably by acting as mechanosensorsthat monitor the composition and flowrate of urine in the nephron (Praetoriusand Spring, 2003; Yoder, 2007). Ob-struction of this flow-sensing responsein, for example, the IFT88Tg737Rpw
1Department of Biology, University of Copenhagen, Copenhagen, Denmark2Department of Cellular and Molecular Medicine, The Panum Institute, University of Copenhagen, Copenhagen, Denmark3Department of Cell Biology, University of Alabama at Birmingham, Birmingham, AlabamaGrant sponsor: National Institutes of Health; Grant number: RO1 R01-HD056030.*Correspondence to: Søren Tvorup Christensen, Department of Biology, Section of Cell and Developmental Biology, TheAugust Krogh Building, University of Copenhagen, Universitetsparken 13, DK-2100 Copenhagen OE, Denmark.E-mail: [email protected]
DOI 10.1002/dvdy.21610Published online 10 July 2008 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 237:2039–2052, 2008
© 2008 Wiley-Liss, Inc.

mouse (hereafter referred to asTg737orpk), causes disruption of tissueorganization and cyst formation. TheTg737orpk mouse has a mutation inthe Tg737/IFT88 gene that encodes asubunit of the IFT particle complex Brequired for ciliary assembly (Pazouret al., 2000; Yoder et al., 2002a;Rosenbaum and Witman, 2002; Ped-ersen et al., 2008). Among the ciliarysignaling modules involved in renaltissue homeostasis are the TRP ionchannel polycystin 2 (PC-2) and poly-cystin-1 (PC-1), which are disruptedin human ADPKD (Yoder et al.,2002b; Pazour et al., 2002; Nauli etal., 2003). Bending of the ciliary axon-eme due to fluid movement has beenshown to induce a Ca2�-response(Praetorius and Spring, 2001), whichis dependent on PC-1 and PC-2. Thiscalcium response is thought to be im-portant in detecting fluid movementfor maintaining tissue function, be-cause mutations leading to its loss re-sult in cyst development (Delmas etal., 2004; Liu et al., 2005b; Kottgen,2007; Weimbs, 2007; Yoder, 2007). Inaddition, in the absence of flow PC-1may be proteolytically processed(Chauvet et al., 2004), leading to thenuclear translocation of the transcrip-tion factor STAT6 and co-activatorP100 (Low et al., 2006). Furthermore,defects in ciliary assembly may resultin up-regulation of the canonical Wntpathway and impair the ability of non-canonical Wnt signals to suppress thecanonical pathway (Gerdes et al.,2007; Corbit et al., 2008). Altered reg-ulation of the Wnt signaling pathwayin the kidney has been associated withuncontrolled cell proliferation and dif-ferentiation and was found to result incystic kidney disease (Cano et al.,2004; Merkel et al., 2007).
In addition to kidney defects, theloss of primary cilia in the Tg737orpk
mouse causes a series of abnormali-ties in the pancreas, including exten-sive cyst formation in ducts (Cano etal., 2004; Zhang et al., 2005). Thismay indicate a functional similaritybetween cilia in kidney and pancreaticduct systems. Cells of both exocrineand endocrine systems in the pan-creas possess primary cilia, includingislet cells and the ducts, but appar-ently not the acini (Kodama, 1983;Ashizawa et al., 1997; Cano et al.,2004, 2006; Zhang et al., 2005). Pan-
creas abnormalities in the Tg737orpk
mouse begin with dilations of the pan-creas ducts in late gestation, whichafter birth are accompanied by exten-sive formation of large, interconnectedcysts as well as apoptosis and vacuol-ization of acini. These changes arereminiscent of chronic pancreatitis,supporting the speculation that pri-mary cilia of ducts play an essentialrole in the development of the pan-creas (Cano et al., 2004; Zhang et al.,2005). In the dilated ducts and cysts,PC-2 is mislocalized to intracellularcompartments, the cytosolic localiza-tion of �-catenin is increased andthere is an increased expression ofTcf/Lef transcription factors. Thisfinding suggests that, as in the kid-ney, there is an alteration in the Wntsignaling pathway (Cano et al., 2004).
Another signaling pathway essen-tial for pancreatic development andtissue homeostasis is the Hedgehog(Hh) pathway. Hh regulates cell pro-liferation and differentiation in nu-merous embryonic tissues and Shh isexpressed in many regions of the em-bryo where it functions as a key orga-nizer of tissue morphogenesis (Odentet al., 1999). One embryonic tissuewhere the initial absence of Shh sig-naling is required for development isin the pancreatic anlage, because ec-topic expression of Shh leads to trans-formation of pancreatic mesoderm intointestinal mesenchyme (Apelqvist etal., 1997). In addition to being an impor-tant inductive signal in embryonic de-velopment, Hh signaling is required inhomeostasis of mature tissues and isalso implicated in many human can-cers, including endodermal derived car-cinomas of the esophagus, stomach, andpancreas (Beachy et al., 2004) as well asneurodegenerative disorders (Bak etal., 2003). Although the mechanism isnot completely understood, it has beenshown that Hh signaling is coordinatedby the primary cilium (e.g., Huangfu etal., 2003; Corbit et al., 2005; Huangfuand Anderson, 2005; May et al., 2005;Haycraft et al., 2005; Liu et al., 2005a;Koyama et al., 2007; Vierkotten et al.,2007; Ruiz-Perez et al., 2007; Casparyet al., 2007; Rohatgi et al., 2007;Kiprilov et al., 2008). Regulation of theHh pathway is complex with Hh re-sponsive Gli transcription factors hav-ing either activator or repressor func-tions that depend on IFT proteins.
Binding of Hh to its receptor Patched(Ptc) occurs in the cilium and results intranslocation of Ptc out of the ciliumand into the cell body (Rohatgi et al.,2007; Kiprilov et al., 2008). In contrast,Smoothened (Smo), the transmem-brane effector of the pathway whose ac-tivity is suppressed by Ptc is targeted tothe cilium in the presence of Hh. Thisrelieves the inhibitory effects of Ptc onSmo and controls the activation or de-activation of Gli transcription factors.In the absence of IFT or cilia, the re-pressor and activator functions of theGli transcriptional regulators becomederegulated with the resulting pheno-type in a tissue being determined bywhether it is the activator or repressorfunction of the Gli transcription factorsthat is most critical. For example, theprocessing of full-length Gli3 (Gli3FL)to repressor form (Gli3R) is affected inIFT mutants, showing the importanceof functional IFT for regulating Glitranscription factors.
In the developing and mature pan-creas, deregulated Hh signaling re-sults in a series of diseases, includingannular pancreas, diabetes mellitus,chronic pancreatitis and pancreaticcancer (Lau et al., 2006). During theearly mouse embryonic stages, Hh sig-naling is kept at a low level in thepancreatic anlage to ensure the cor-rect establishment of organ bound-aries, as well as tissue architecture.At later developmental stages, Hh sig-naling is activated to promote prolif-eration and maturation of the tissue(Kawahira et al., 2005; Lau et al.,2006; Cano et al., 2007; van denBrink, 2007). In the mature pancreas,Hh signaling maintains endocrinefunctions, and aberrant Hh signalingcauses pancreatic ductal adenocarci-noma and chronic pancreatitis(Thayer et al., 2003; Berman et al.,2003; Kayed et al., 2003, 2006; Mortonet al., 2006; Cano et al., 2007).
The aim of our study was to exam-ine the expression of primary cilia inhuman pancreatic ducts and to inves-tigate the sensory competence of pri-mary cilia with regards to the Hh sig-naling pathway during pancreaticdevelopment and in pancreatic adeno-carcinoma cell lines, CFPAC-1 andPANC-1, which are isolated from met-astatic and primary tumors, respec-tively (Lieber et al., 1975; Schouma-cher et al., 1990). Initially, we show
2040 NIELSEN ET AL.

that localization of Hh signaling com-ponents is regulated during humanpancreatic development with Gli2 andSmo accumulating in the cilium atlater stages while the level of nuclearand cytosolic Gli3 expression is re-duced. These changes in localizationcorrelate with known activity of theHh pathway during pancreas develop-ment. We demonstrate that compo-nents of the Hh pathway also localizeto the cilium in CFPAC-1 and PANC-1cells and that these cells may maintainhigh levels of nonstimulated Hh path-way activity. These findings indicatethat Hh signaling coordinated by theprimary cilium in the pancreas is essen-tial for normal pancreatic developmentas well as postnatal tissue function.
RESULTS
Early Development of theHuman Pancreas
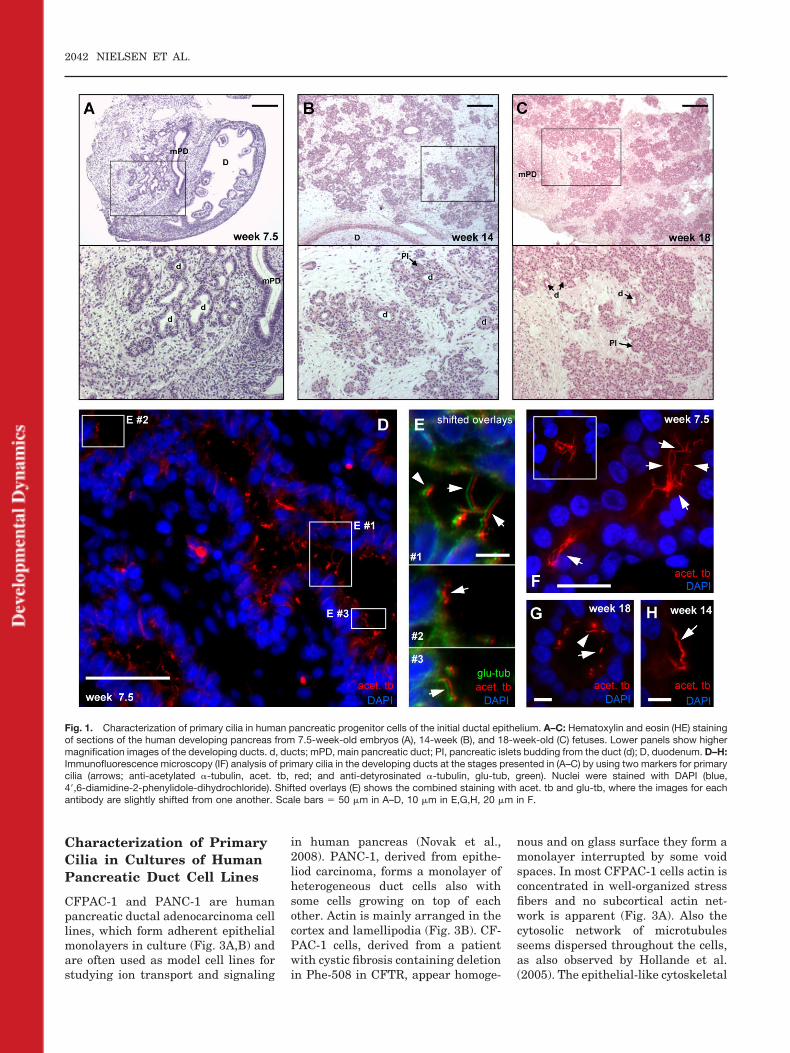
To investigate the presence of primarycilia in human pancreatic progenitorcells of the initial duct epithelium weanalyzed 3-�m-thick tissue sectionsfrom 7.5-week-old embryos and from14- and 18-week-old fetuses. Hema-toxylin and eosin (HE) staining of thepancreatic tissue and identification ofthe developing ducts at these stagesare presented in Figure 1A–C. In 5- to6-week-old embryos a dorsal and aventral out pocketing of the foregutduodenal endoderm result in the for-mation of separate dorsal and ventralpancreatic primordia. During the sev-enth embryonic week a fusion of thetwo epithelial pancreatic ducts takesplace followed by a rapid expansionand branching of the ductal epithe-lium (Fig. 1A). Pancreatic progenitorcells in this epithelium engage in twoseparate differentiation programs atdistinct developmental stages. At thefirst stage from week 8 to 10, the en-docrine progenitor cells appear as cellclusters budding from the central duc-tal epithelium. These buds form theislets of Langerhans in the centralpancreas at week 12, and at midges-tation, the endocrine compartment isestablished. At a much later stage, theexocrine progenitors are responsiblefor the differentiation and expansionof the exocrine acinar epithelium,which is mainly a late fetal or earlyperinatal event. Figure 1B shows a
section of the ductal system from a14-week-old fetus (Jackerott et al.,2006) with initial endocrine progeni-tors budded from the ductal epithelium,and Figure 1C shows a section from an18-week-old fetus with endocrine pro-genitors budded from the ductal epithe-lium as well as acinar progenitors sur-rounding the ductal epithelium.
Characterization of PrimaryCilia in Ducts of theDeveloping Human Pancreas
To identify primary cilia, we used twomarkers for cilia (anti-acetylated�-tubulin [acet. tb] and anti-detyrosi-nated �-tubulin [glu-tub]) in immuno-fluorescence microscopy (IF) analysisand found that epithelial cells in ductsat all stages form primary cilia thateither project into the duct lumen,form spiral-like structures or alignparallel to the lumen surface (Fig.1D–H). The cilia varied in lengthsfrom approximately 5 to 20 �m, and insome ducts long cilia seemed to formcontact or bridge with cilia fromneighboring epithelial cells or fromcells on the opposite side of the duct(Fig. 1E,F). At the late stage of embry-onic development (week 7.5), longacetylated-tubulin structures oftenprojected from epithelial cells in a“star”-like configuration by pointinginto the site of the developing duct(Fig. 1F and inset). Both short andlong primary cilia were observed dur-ing all three developmental stages.Long cilia were predominantly ob-served in week 7.5, possibly due to theprevalence of small intercalated andintralobular type ducts that do havelonger cilia (Kodama, 1983). The for-mation and orientation of primarycilia in the human developing ducts issimilar to that reported for the pan-creas in the rat and mouse (e.g., Ko-dama, 1983; Hidaka et al., 1995; Ash-izawa et al., 1997; Cano et al., 2004;Zhang et al., 2005).
Development of the HumanPancreas Correlates WithCiliary Localization of HhSignaling Components inDuct Epithelial Cells
It was previously shown that thegraded response to Hh-signaling con-
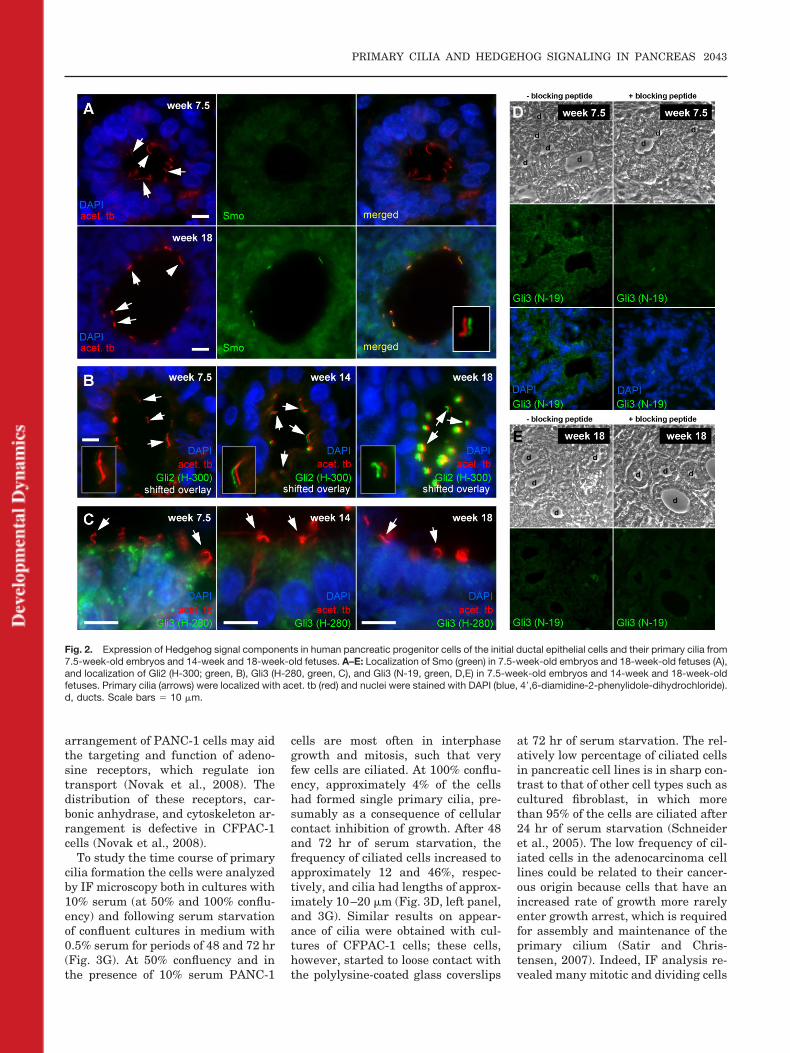
trols pancreatic organogenesis in themouse, where Hh signaling is at lowlevels during early embryonic stagesto ensure the correct establishment oforgan boundaries and tissue architec-ture. Hh signaling is then activated atlater developmental stages to promoteproliferation and maturation of thetissue (Kawahira et al., 2005; Lau etal., 2006; Cano et al., 2007; van denBrink, 2007). In Hh signaling, Gli2 invivo acts primarily as a transcrip-tional activator, whereas Gli3 mainlyworks as a transcriptional repressorto control development. To addressthe potential role of primary cilia inhuman pancreatic development, weinvestigated the expression and cellu-lar distribution of Smo, Gli2, and Gli3in developing pancreatic ducts by IFanalysis. In other cell types, such asfibroblasts (Rohatgi et al., 2007) andhuman embryonic stem cells, hESC(Kiprilov et al., 2008), Smo becomesconcentrated while Ptc levels decreasein primary cilia upon stimulation witha Hh ligand, which may result in ac-tivation or deactivation of Gli tran-scription factors, possibly in the ci-lium (Huangfu and Anderson, 2006;Christensen and Ott, 2007). We foundthat Smo (Fig. 2A) and Gli2 (Fig. 2B)are absent from primary cilia in pan-creatic ducts of embryos (week 7.5),but are concentrated in the cilia ofducts of early fetuses (week 14), andeven more so in 18-week-old fetuses.We then investigated the localizationof Gli3 in IF analysis using two differ-ent Gli3 antibodies that were raisedagainst amino acids 1-280 of Gli3(Gli3-H-280) and against a peptidemapping at the N-terminus of Gli3(Gli3-N-19). Both antibodies showed adecrease in staining intensity and cel-lular distribution of Gli3 during devel-opment. At week 7.5, Gli3 strongly lo-calized to duct epithelial cells and waspresent in the nuclei and in an intensepunctate pattern in the cytosol of ductepithelial cells. In contrast, in ducts of14- and 18-week-old fetuses, Gli3staining was reduced (Fig. 2C–E). Asa control, a blocking peptide againstanti-Gli3-N-19 decreased localizationof the antibody to the duct epithelium(Fig. 2D). These results could reflectactivation of the pathway because Hhis known to repress Gli3 expression(Marigo et al., 1996; Buscher andRuther, 1998; Schweitzer et al., 2000).
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2041
Characterization of PrimaryCilia in Cultures of HumanPancreatic Duct Cell Lines
CFPAC-1 and PANC-1 are humanpancreatic ductal adenocarcinoma celllines, which form adherent epithelialmonolayers in culture (Fig. 3A,B) andare often used as model cell lines forstudying ion transport and signaling
in human pancreas (Novak et al.,2008). PANC-1, derived from epithe-liod carcinoma, forms a monolayer ofheterogeneous duct cells also withsome cells growing on top of eachother. Actin is mainly arranged in thecortex and lamellipodia (Fig. 3B). CF-PAC-1 cells, derived from a patientwith cystic fibrosis containing deletionin Phe-508 in CFTR, appear homoge-
nous and on glass surface they form amonolayer interrupted by some voidspaces. In most CFPAC-1 cells actin isconcentrated in well-organized stressfibers and no subcortical actin net-work is apparent (Fig. 3A). Also thecytosolic network of microtubulesseems dispersed throughout the cells,as also observed by Hollande et al.(2005). The epithelial-like cytoskeletal
Fig. 1. Characterization of primary cilia in human pancreatic progenitor cells of the initial ductal epithelium. A–C: Hematoxylin and eosin (HE) stainingof sections of the human developing pancreas from 7.5-week-old embryos (A), 14-week (B), and 18-week-old (C) fetuses. Lower panels show highermagnification images of the developing ducts. d, ducts; mPD, main pancreatic duct; PI, pancreatic islets budding from the duct (d); D, duodenum. D–H:Immunofluorescence microscopy (IF) analysis of primary cilia in the developing ducts at the stages presented in (A–C) by using two markers for primarycilia (arrows; anti-acetylated �-tubulin, acet. tb, red; and anti-detyrosinated �-tubulin, glu-tub, green). Nuclei were stained with DAPI (blue,4�,6-diamidine-2-phenylidole-dihydrochloride). Shifted overlays (E) shows the combined staining with acet. tb and glu-tb, where the images for eachantibody are slightly shifted from one another. Scale bars � 50 �m in A–D, 10 �m in E,G,H, 20 �m in F.
2042 NIELSEN ET AL.
arrangement of PANC-1 cells may aidthe targeting and function of adeno-sine receptors, which regulate iontransport (Novak et al., 2008). Thedistribution of these receptors, car-bonic anhydrase, and cytoskeleton ar-rangement is defective in CFPAC-1cells (Novak et al., 2008).
To study the time course of primarycilia formation the cells were analyzedby IF microscopy both in cultures with10% serum (at 50% and 100% conflu-ency) and following serum starvationof confluent cultures in medium with0.5% serum for periods of 48 and 72 hr(Fig. 3G). At 50% confluency and inthe presence of 10% serum PANC-1
cells are most often in interphasegrowth and mitosis, such that veryfew cells are ciliated. At 100% conflu-ency, approximately 4% of the cellshad formed single primary cilia, pre-sumably as a consequence of cellularcontact inhibition of growth. After 48and 72 hr of serum starvation, thefrequency of ciliated cells increased toapproximately 12 and 46%, respec-tively, and cilia had lengths of approx-imately 10–20 �m (Fig. 3D, left panel,and 3G). Similar results on appear-ance of cilia were obtained with cul-tures of CFPAC-1 cells; these cells,however, started to loose contact withthe polylysine-coated glass coverslips
at 72 hr of serum starvation. The rel-atively low percentage of ciliated cellsin pancreatic cell lines is in sharp con-trast to that of other cell types such ascultured fibroblast, in which morethan 95% of the cells are ciliated after24 hr of serum starvation (Schneideret al., 2005). The low frequency of cil-iated cells in the adenocarcinoma celllines could be related to their cancer-ous origin because cells that have anincreased rate of growth more rarelyenter growth arrest, which is requiredfor assembly and maintenance of theprimary cilium (Satir and Chris-tensen, 2007). Indeed, IF analysis re-vealed many mitotic and dividing cells
Fig. 2. Expression of Hedgehog signal components in human pancreatic progenitor cells of the initial ductal epithelial cells and their primary cilia from7.5-week-old embryos and 14-week and 18-week-old fetuses. A–E: Localization of Smo (green) in 7.5-week-old embryos and 18-week-old fetuses (A),and localization of Gli2 (H-300; green, B), Gli3 (H-280, green, C), and Gli3 (N-19, green, D,E) in 7.5-week-old embryos and 14-week and 18-week-oldfetuses. Primary cilia (arrows) were localized with acet. tb (red) and nuclei were stained with DAPI (blue, 4�,6-diamidine-2-phenylidole-dihydrochloride).d, ducts. Scale bars � 10 �m.
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2043
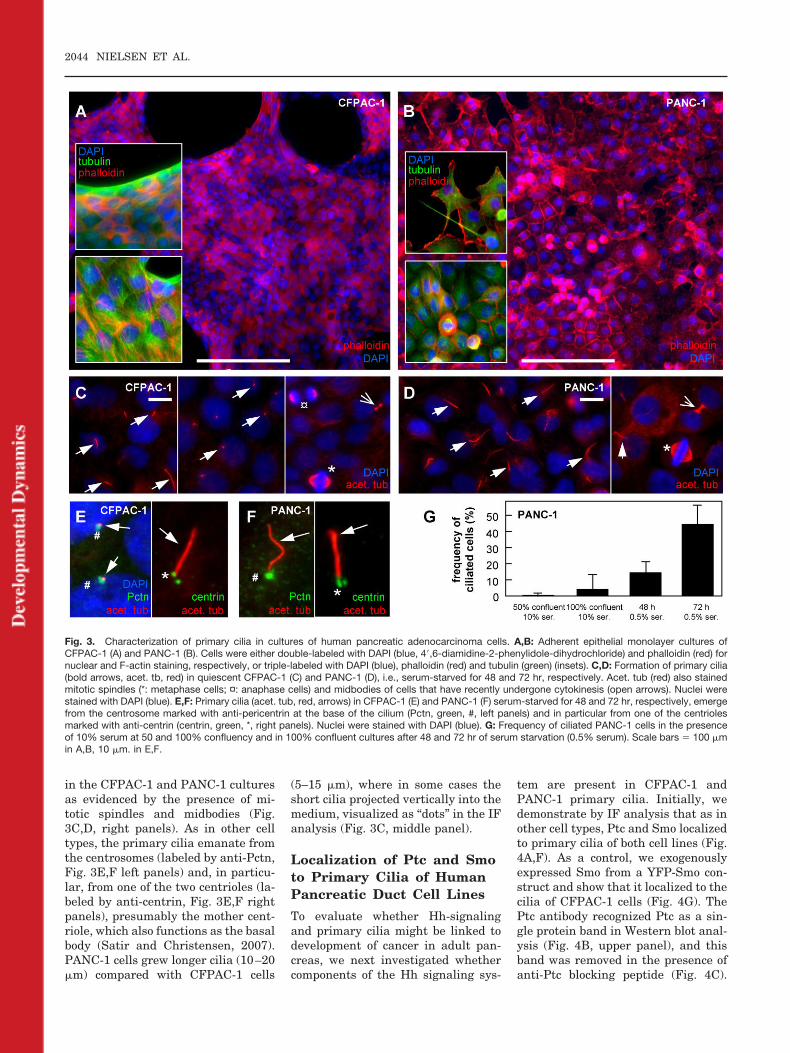
in the CFPAC-1 and PANC-1 culturesas evidenced by the presence of mi-totic spindles and midbodies (Fig.3C,D, right panels). As in other celltypes, the primary cilia emanate fromthe centrosomes (labeled by anti-Pctn,Fig. 3E,F left panels) and, in particu-lar, from one of the two centrioles (la-beled by anti-centrin, Fig. 3E,F rightpanels), presumably the mother cent-riole, which also functions as the basalbody (Satir and Christensen, 2007).PANC-1 cells grew longer cilia (10–20�m) compared with CFPAC-1 cells
(5–15 �m), where in some cases theshort cilia projected vertically into themedium, visualized as “dots” in the IFanalysis (Fig. 3C, middle panel).
Localization of Ptc and Smoto Primary Cilia of HumanPancreatic Duct Cell Lines
To evaluate whether Hh-signalingand primary cilia might be linked todevelopment of cancer in adult pan-creas, we next investigated whethercomponents of the Hh signaling sys-
tem are present in CFPAC-1 andPANC-1 primary cilia. Initially, wedemonstrate by IF analysis that as inother cell types, Ptc and Smo localizedto primary cilia of both cell lines (Fig.4A,F). As a control, we exogenouslyexpressed Smo from a YFP-Smo con-struct and show that it localized to thecilia of CFPAC-1 cells (Fig. 4G). ThePtc antibody recognized Ptc as a sin-gle protein band in Western blot anal-ysis (Fig. 4B, upper panel), and thisband was removed in the presence ofanti-Ptc blocking peptide (Fig. 4C).
Fig. 3. Characterization of primary cilia in cultures of human pancreatic adenocarcinoma cells. A,B: Adherent epithelial monolayer cultures ofCFPAC-1 (A) and PANC-1 (B). Cells were either double-labeled with DAPI (blue, 4�,6-diamidine-2-phenylidole-dihydrochloride) and phalloidin (red) fornuclear and F-actin staining, respectively, or triple-labeled with DAPI (blue), phalloidin (red) and tubulin (green) (insets). C,D: Formation of primary cilia(bold arrows, acet. tb, red) in quiescent CFPAC-1 (C) and PANC-1 (D), i.e., serum-starved for 48 and 72 hr, respectively. Acet. tub (red) also stainedmitotic spindles (*: metaphase cells; ¤: anaphase cells) and midbodies of cells that have recently undergone cytokinesis (open arrows). Nuclei werestained with DAPI (blue). E,F: Primary cilia (acet. tub, red, arrows) in CFPAC-1 (E) and PANC-1 (F) serum-starved for 48 and 72 hr, respectively, emergefrom the centrosome marked with anti-pericentrin at the base of the cilium (Pctn, green, #, left panels) and in particular from one of the centriolesmarked with anti-centrin (centrin, green, *, right panels). Nuclei were stained with DAPI (blue). G: Frequency of ciliated PANC-1 cells in the presenceof 10% serum at 50 and 100% confluency and in 100% confluent cultures after 48 and 72 hr of serum starvation (0.5% serum). Scale bars � 100 �min A,B, 10 �m. in E,F.
2044 NIELSEN ET AL.

Expression of Ptc at the mRNA levelwas previously shown to be up-regu-lated as a consequence of the autono-mous operation of an active Hh signal-ing process in pancreatic cancer cellsin which Ptc mRNA in PANC-1 cells isup-regulated at a higher level thanthat of CFPAC-1 cells (Berman et al.,2003; Thayer et al., 2003). Consistentwith these earlier findings, we showthat the protein level of Ptc in PANC-1was higher than that in CFPAC-1cells (Fig. 4B, upper and lower pan-els). As further controls for antibodyspecificity, we show that Ptc localizedto primary cilia of wild-type mouseembryonic fibroblasts (wt MEFs) butnot to mutant Ptc�/� MEFs (Fig. 4D)as previously demonstrated by Ro-hatgi et al. (2007). Then we stimu-lated wt MEFs with the Smo agonist,SAG, that activates Hh-signaling(Chen et al., 2002) to analyze changesin protein levels of Ptc, which is up-regulated in response to activation ofthe Hh pathway. As shown in Figure4E, the Ptc is present at a low level inwt MEFs in the absence of SAG,whereas it increased after Hh path-way activation. This increase was de-pendent on the presence of the pri-mary cilium, because the level of Ptcwas kept at a low level in the presenceof SAG in MEFs derived from theTg737orpk mouse (Fig. 4E). These re-sults support the idea that the ciliumis required for activation of the Hhpathway, and that aberrant Hh sig-naling in pancreatic cancer may beassociated with increased Hh signal-ing by means of the primary cilium.
Another observation was that Ptclocalized only weakly to the primarycilium (Fig. 4A), while Smo localizedvery strongly and in a punctate pat-tern in the cilium of the pancreaticduct cancer cells (Fig. 4F). This is insharp contrast to that observed in cul-tures of normal cells such as MEFsand hESCs, where Ptc is highly con-centrated in the cilium and Smo ismostly absent from the cilium in non-stimulated cells (Rohatgi et al., 2007;Kiprilov et al., 2008). Stimulation ofthe pancreatic duct cells with SAG didnot further increase ciliary localiza-tion of Smo in CFPAC-1 cells (Fig.4H), supporting the conclusion thatthe Hh pathway is maintained at ahigh and autonomous level in thesecells.
Localization of GliTranscription Factors toPrimary Cilia of HumanPancreatic Duct Cell Lines
Gli2 and Gli3 are the primary tran-scription factors that are being regu-lated in Hh signaling. The full-lengthtranscription factors mainly functionas transcriptional activators, but inthe absence of the Hh signaling theymay undergo proteolytic processingand function as transcriptional re-pressors (Wang et al., 2000; Liting-tung et al., 2002; Pan et al., 2006). Toinvestigate the expression and local-ization of Gli2 in growth-arrestedPANC-1 and CF-PAC-1 cells we usedthree different antibodies raisedagainst amino acids 841-1140 map-ping near the C-terminus of Gli2(Gli2-H-300), a peptide mapping nearthe N-terminus of Gli2 (Gli2-N-20),and a peptide mapping within an in-ternal region of Gli2 (Gli2-G-20). Allthree antibodies localized uniquely tothe primary cilium and most often asan enrichment at the tip of the cilium(Fig. 5A,B, left panels), similar towhat has been observed in primarycilia of the murine limb bud and hESC(Haycraft et al., 2005; Kiprilov et al.,2008). Ciliary localization was abol-ished in the presence of blocking pep-tides to the two peptide antibodies(Fig. 5B, right panels). Also, localiza-tion of Gli2 to the ciliary tip was con-firmed by transfection and overex-pression of a green fluorescent protein(GFP) -Gli2 construct in NIH3T3 fi-broblasts (Fig. 5C). To confirm anti-body specificity, we performed West-ern blotting analysis with the peptideantibodies in the absence and in thepresence of blocking peptides. Asshown in Fig. 5D, Gli2-N-20 specifi-cally recognized both full-length (ca.170 kDa) and processed (ca. 70 kDa)forms of Gli2 that function as activa-tor (Gli2(FL)) and repressor (Gli2(R))forms in Hh signaling, respectively(Pan et al., 2006). In contrast, Gli2-G-20 recognized only Gli2(FL) (Fig.5E). This finding suggests that the ac-tivator forms of Gli2 are present in theprimary cilium and that ciliary Gli2may be part of the Hh signaling ma-chinery that is up-regulated in pan-creatic cancer cells.
We then analyzed lysates from CF-PAC-1 cells by Western blotting anal-
ysis using the Gli3 antibodies, Gli3-N-19 and Gli3-H-280. Gli3-N-19recognized both full-length (Gli3(FL)at ca. 180 kD) and processed forms(Gli3(R) at ca. 80 kDa) of Gli3 (Fig.5F,G). In contrast, Gli3-H-280 pre-dominantly recognized the processedversion of Gli3 (Fig. 5F), supportingthe conclusion that this antibodymainly identifies the repressor form ofGli3 under the experimental condi-tions carried out in this work. Thesedata further support the assumptionthat the observed decrease in stainingintensity and cellular distribution ofGli3 in the epithelium of the pancre-atic ducts during fetal development(Fig. 2) is due to the increased activa-tion of the Hh signaling that favorsthe processing of transcriptional acti-vators of the Hh pathway. Based onthese observations, it was thereforeinteresting to investigate the expres-sion and localization of Gli3(R) in pan-creatic cancer cells upon stimulationof the Hh pathway using Gli3-H-280in IF analysis. As seen in Figure 5H,Gli3-H-280 stained CFPAC-1 cellsvery weakly in both the absence andin the presence of SAG. In contrast,Gli3-H-280 strongly stained the nu-cleus as well as small cytoplasmicpuncta in NIH3T3 cells and this stain-ing was largely absent in the presenceof SAG (Fig. 5I). Furthermore, weused transfected NIH3T3 cells withGFP constructs of either the GFP-Gli3(R) and Gli3(FL) to evaluate thecellular localization of activator andrepressor forms of Gli3. Similar tothat seen in nonstimulated NIH3T3cells with Gli3-H-280 (Fig. 5I) andpreviously in mouse limb bud mesen-chyme cells, we found that GFP-Gli3(R) localized predominantly to thenucleus and to small cytosolic puncta,but not to the primary cilium (Fig. 5J,left panels). In contrast, GFP-Gli3(FL)seemed to localize to the primary ci-lium with minimal localization in thenucleus and without cytosolic puncta(Fig. 5J, right panels). These resultsare in line with our conclusion thatGli3-H-280 recognizes the repressorform of Gli3, which is concentrated inthe nucleus in the absence of Hh sig-naling, and that Gli3-repressor formsare expressed at a low level in CF-PAC-1 cells.
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2045
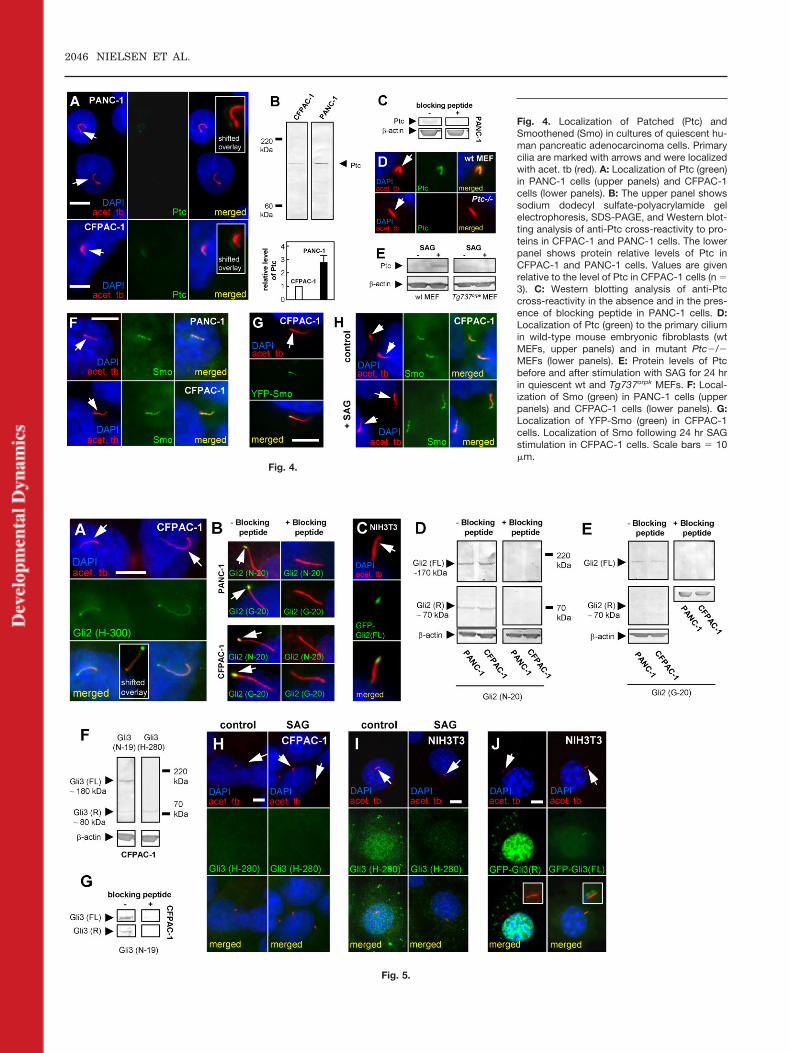
Fig. 4.
Fig. 5.
Fig. 4. Localization of Patched (Ptc) andSmoothened (Smo) in cultures of quiescent hu-man pancreatic adenocarcinoma cells. Primarycilia are marked with arrows and were localizedwith acet. tb (red). A: Localization of Ptc (green)in PANC-1 cells (upper panels) and CFPAC-1cells (lower panels). B: The upper panel showssodium dodecyl sulfate-polyacrylamide gelelectrophoresis, SDS-PAGE, and Western blot-ting analysis of anti-Ptc cross-reactivity to pro-teins in CFPAC-1 and PANC-1 cells. The lowerpanel shows protein relative levels of Ptc inCFPAC-1 and PANC-1 cells. Values are givenrelative to the level of Ptc in CFPAC-1 cells (n �3). C: Western blotting analysis of anti-Ptccross-reactivity in the absence and in the pres-ence of blocking peptide in PANC-1 cells. D:Localization of Ptc (green) to the primary ciliumin wild-type mouse embryonic fibroblasts (wtMEFs, upper panels) and in mutant Ptc�/�MEFs (lower panels). E: Protein levels of Ptcbefore and after stimulation with SAG for 24 hrin quiescent wt and Tg737orpk MEFs. F: Local-ization of Smo (green) in PANC-1 cells (upperpanels) and CFPAC-1 cells (lower panels). G:Localization of YFP-Smo (green) in CFPAC-1cells. Localization of Smo following 24 hr SAGstimulation in CFPAC-1 cells. Scale bars � 10�m.
2046 NIELSEN ET AL.

DISCUSSION
Recent research has shown that pri-mary cilia coordinate signal path-ways, which are essential in mamma-lian development, tissue homeostasisand behavioral responses. Accord-ingly, defective primary cilia assemblyand maintenance lead to a plethora ofdiseases and disorders, now collec-tively referred to as ciliopathies(Badano et al., 2006; Fliegauf et al.,2007; Satir and Christensen, 2007;Yoder, 2007). Previous reports haveshown that loss of primary cilia in theTg737orpk mouse causes a series of de-velopmental defects in exocrine andendocrine tissues of the pancreas, in-cluding extensive cyst formation inducts (Cano et al., 2004; Zhang et al.,2005). However, the function of pri-mary cilia in pancreatic organogenesisand adult tissue homeostasis islargely unknown. In this report, wedemonstrate that essential compo-nents of the Hedgehog (Hh) signalingpathway localize to primary cilia ofboth human pancreatic progenitorcells of the initial ductal epitheliumand in cultures of human pancreaticadenocarcinoma duct cell lines, andthat expression of ciliary Hh compo-nents may be linked to pancreatic or-ganogenesis and pancreatic cancer.
Hh Signaling and PrimaryCilia in the DevelopingHuman Pancreas
Previous studies have shown that thegraded response to activation of theHh signaling pathway is critical inpancreatic organogenesis in themouse, where Hh signaling is acti-vated only at late developmental
stages (Kawahira et al., 2005; Lau etal., 2006; Cano et al., 2007; van denBrink, 2007). It is likely that organo-genesis of the human pancreas alsodepends on the graded response to Hhsignaling, because the transcriptionalnetwork that controls pancreatic de-velopment is highly conserved inmammals (Wilson et al., 2003), al-though the time point of onset of de-velopment and maturation of the en-docrine system differs in humans androdents (Ostrer et al., 2006). Recently,the primary cilium was shown to actas a sophisticated switch by whichcells turn Hh signaling on and off bythe regulated movement of Smo intothe cilium and Ptc out of the ciliumupon stimulation of the Hh pathwayin fibroblasts (Rohatgi et al., 2007)and hESC (Kirprilov et al., 2008),where Smo probably functions to acti-vate Gli transcription factors (Chris-tensen and Ott, 2007). To analyze thesignificance of the primary cilium inHh signaling during organogenesis ofthe human pancreas, we studied sec-tions of the developing human pan-creas at different developmentalstages. These stages included embry-onic stage 7.5 weeks, which is beforedevelopment of the endocrine system,and stages 14 and 18 weeks, whereendocrine progenitors bud from theductal epithelium to form the endo-crine compartment (Fig. 1). Based onthe recent data from Rohatgi et al.(2007), our findings that Smo and Gli2are absent from pancreatic primarycilia at embryonic stage 7.5, buthighly concentrated in cilia in 14- and18-week-old fetuses (Fig. 2) suggestthat the Hh pathway has become ac-
tivated during developmental stages.This further supports the conclusionthat development of the human pan-creas is regulated by a graded Hh sig-naling response, and that this could becoordinated by the primary cilium. In-deed, the loss of Gli3 staining in theductal epithelium after week 7.5, thatis, before formation of the endocrinesystem, is consistent with the inter-pretation that Gli3 staining at week7.5 may represent the up-regulatedlevel of the repressor form of Gli3 (Fig.5F), consistent with low level of Hhsignaling at this developmental stage.Therefore, disruption of pancreatic de-velopment in mice with defects in pri-mary ciliary assembly (Cano et al.,2004, 2006; Zhang et al., 2005), maypartly be due to loss of coordinated Hhsignaling during genesis of the pan-creas.
Another interesting observation isthe presence of long cilia-like struc-tures that seem to bridge with corre-sponding structures emerging fromadjacent or opposite cells of the lumensurface (Fig. 1). This is particularlyprominent in the developing ducts atweek 7.5. Bridging of primary ciliawas also observed in collecting ductsof the mouse developing kidney (Liu etal., 2005b), but the function of physi-cal contact between individual pri-mary cilia is unknown. Althoughhighly speculative at this point, theintertwining of cilia could be part ofthe sensory signaling machinery thatcontrols the initial development ofducts and perhaps endocrine progeni-tors. In the developing kidney, the for-mation of tubular systems is con-trolled by inversin, which acts as a
Fig. 5. Expression and localization of Gli transcription factors in quiescent human pancreatic adenocarcinoma cells and NIH3T3 fibroblasts. Primarycilia are marked with arrows and were localized with acet. tb (red). A: Localization of Gli2 (H-300; green) to primary cilia of CFPAC-1 cells. The insetin the lower panel shows a shifted overlay of the cilium (red) and Gli3 (H-300, green). B: Localization of Gli2 (N-20) and Gli2 (G-20, both in green) inthe absence and in the presence of their corresponding blocking peptides in PANC-1 and CFPAC-1 cells. C: Localization of green fluorescent protein(GFP) -Gli2 (green) in NIH3T3 cells. D,E: SDS-PAGE and Western blotting analysis of anti-Gli2 (N-20) (D) and anti-Gli2 (G-20) (E) specificity to proteinsin CFPAC-1 and PANC-1 cells in the absence and in the presence of their corresponding blocking peptides. Anti-Gli2 (G-20) only recognized a bandat ca. 170 kDa. Abbreviations: Gli2(FL): Full length (activator) form of Gli2; Gli2(R): Processed (repressor) form of Gli2. F: SDS-PAGE and Westernblotting analysis of anti-Gli3 (N-19) and anti-Gli3 (H-280) specificity to proteins in CFPAC-1 cells. Gli3(FL): Full-length (mild activator) form of Gli3;Gli3(R): Processed (repressor) form of Gli3. Anti-Gli (N-19) recognized both Gli3 forms, while Gli3 (H-280) predominantly recognized the repressor formof Gli3. G: Western blotting analysis of anti-Gli3 (N-19) specificity in the absence and in the presence of its blocking peptide in CFPANC-1 cells.H,I: Changes in localization of Gli3 (H-280) in nonstimulated (left panels) vs. SAG-stimulated (right panels) in CFPAC-1 (H) and NIH3T3 (I) cells. Innonstimulated NIH3T3 cells, Gli3 localized similar to that of GFP-Gli3(R), but after SAG stimulation the nuclear and cytosolic levels of Gli3 weremarkedly decreased. In CF-PAC-1 cells the nuclear and cytosolic levels of Gli3 were low in both stimulated and nonstimulated cells. J: Expression oftwo different GFP-Gli3 (green) constructs in quiescent NIH3T3 cells, where the repressor form of Gli3 (GFP-Gli3(R)) strongly localizes to the nucleusand to cytosolic puncta (left panels) and the full-length form of Gli3 (GFP-Gli3(FL)) localizes to the primary cilium (right panels). Insets: Shifted overlayswith high resolution of GFP in the cilium. Nuclei were stained with DAPI (blue, 4�,6-diamidine-2-phenylidole-dihydrochloride). Scale bars � 5 �m in A,10 �m in J.
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2047

molecular switch between the canoni-cal and nonconical Wnt pathways (Si-mons et al., 2005), and inversin poten-tially signals from the primary ciliumto suppress �-catenin expression inthe canonical pathway and to up-reg-ulate the noncanonical pathway to or-ganize planar cell polarity and correcttubular formation (Christensen et al.,2007). Because loss of the primary ci-lium in the developing mouse pan-creas is associated with increased�-catenin expression (Cano et al.,2004; Zhang et al., 2005), it is possiblethat cilia in the developing pancreasregulate both Wnt and Hh signaling,although it is presently unknownwhether Hh or Wnt signaling is af-fected by physical contact betweenprimary cilia in the ductal epithelium.Further experiments are required toexplore this.
Hh Signaling and PrimaryCilia in Human PancreaticDuct Cell Lines
Hh is implicated as an important me-diator of human pancreatic carci-noma, as evidenced by, for example,the increased expression of positiveHh regulators and decreased expres-sion of negative Hh regulators in CF-PAC-1 or PANC-1 cells (Thayer et al.,2003; Berman et al., 2003). Here, weshow that essential components of theHh pathway, including Smo, Ptc, andGli2, are present in primary cilia ofhuman pancreatic ductal adenocarci-noma cell lines, CFPAC-1 andPANC-1, consistent with the idea thatthe primary cilium continues to coor-dinate Hh signaling in cells derivedfrom the mature pancreas. We alsosuggest that aberrant Hh signaling inthese cancer cell lines may be associ-ated with the autonomous activationof the signaling pathway in the ciliumas judged by high levels of Smo andlow levels of Ptc in the cilium, which isaccompanied by the formation ofGli2(FL) forms in the cilium and lowlevels of Gli3(R) in the nucleus.
In support of the hypothesis thatthe primary cilium may comprise thesite for aberrant Hh signaling in hu-man pancreatic cancer tumorigenesis,we show that in contrast to normalcells, which require Hh pathway stim-ulation in the cultures for Smo to en-
ter the cilium and activate Hh targetgenes (Rohatgi et al., 2007; Kiprilov etal., 2008), Smo is strongly localized tothe primary cilium in both CFPAC-1and PANC-1 cells, even in the absenceof external stimulation of the Hhpathway by SAG. Further and in con-trast to primary cultures of mouse em-bryonic fibroblasts, Ptc is expressed ata higher level than that of MEFs,which require the primary cilium andstimulation of the Hh pathway to in-crease their expression of Ptc. The ac-tivator form of Gli2, Gli2(FL), local-izes to the tip of primary cilium, whichmay comprise the site for activation ofGli transcription factors in Hh signal-ing. We then examined the expressionand localization of Gli3 before and af-ter SAG stimulation by comparing thelocalization of endogenous Gli3 by IFassessed using Gli3-H-280, which rec-ognizes the processed form of Gli3with localization of GFP-Gli3(FL) andGFP-Gli3(R) expressed in transfectgrowth-arrested NIH3T3 fibroblasts.As shown in Figure 5J, only GFP-Gli3(R) localized to the nucleus and tocytoplasmic puncta. These data areconsistent with the conclusion thatthe repressor form is absent from theprimary cilium and is repressing Hhtarget genes in the absence of Hh sig-naling. We then performed IF analysisin NIH3T3 cells in the absence and inthe presence of SAG and compared thestaining with Gli3-H-280 to localiza-tion of the protein expressed from theGFP constructs. As shown in Figure5I (left panels) anti-Gli3 stains non-stimulated cells similar to that of theGFP-Gli3(R) construct. Upon activa-tion of the Hh pathway nuclear Gli3staining is strongly reduced, support-ing the conclusion that the antibody inIF analysis is detecting the repressorform of Gli3. Finally, we examined thelocalization of Gli3 in CFPAC-1 cells,which showed very weak cytosolic andnuclear staining of Gli3 in both stim-ulated and nonstimulated cells. Thisresult would be expected if Hh signal-ing activity is elevated in these cellsbecause Hh signaling represses Gli3expression (Marigo et al., 1996; Bus-cher and Ruther, 1998; Schweitzer etal., 2000), and is further evidenced bythe accumulation of Smo in the ciliumof these cells.
CONCLUSIONS
Our data indicate a functional role ofthe primary cilium in coordinating Hhsignaling during development of thehuman pancreas and potentially in tu-morigenesis of the adult human pan-creas. During development the pri-mary cilium may sense and relay thegraded response to activation of theHh signaling pathway that controlshuman pancreatic organogenesis atthe onset of fetal development. In theadult, the autonomous activation ofthe Hh pathway in pancreatic tumor-igenesis may be linked to aberrant Hhsignaling in the primary cilium of thepancreatic cells. Further analysis willbe required to understand the poten-tial significance of Gli processing inthe cilium and how the switch be-tween activator and repressor forms ofGli2 and Gli3 is regulated by the ci-lium during development and tumori-genesis of the pancreas and poten-tially of other human tissues andorgans. Furthermore, it will be impor-tant to identify the molecular mecha-nisms that coordinate translocation ofSmo and Ptc in and out of the cilium,which ultimately may impinge on theactivation and deactivation of the Hhpathway, which is of relevance in hu-man health and development.
EXPERIMENTALPROCEDURES
Cell Cultures
The human pancreatic exocrine ductcell lines PANC-1 (ATCC, #CRL-1469)and CFPAC-1 (ATCC, #CRL-1918;passages 8–36) were from the Ameri-can Type Culture Collection and werecultured in Dulbecco�s modified Ea-gles medium with glutamax (DMEM;Invitrogen) and Iscove’s modified Dul-becco’s medium (IMDM; Invitrogen),respectively. Both growth media weresupplemented with 10% heat inacti-vated fetal calf serum (FCS; Invitro-gen) and 1% penicillin-streptomycin(Penicilin G sodium: 10,000 U/ml andstreptomycin G sodium: 10,000 �g/ml)(P/S; Invitrogen) and kept in a humid-ified air chamber at 37°C and 5% CO2.Passing of cells was performed bytrypsination. NIH3T3 Swiss mouseembryonic fibroblasts and wt, Ptc�/�and Tg737orpk mouse embryonic fibro-blasts (MEFs) were cultured as de-
2048 NIELSEN ET AL.

scribed previously for MEFs (Schnei-der et al., 2005).
Tissues
Human embryos and fetuses were ob-tained from extra uterine pregnan-cies, Caesarean sections, or prosta-glandin induced legal abortionsdonated to the Developmental BiologyUnit, ICMM, at the Panum Institute,University of Copenhagen, Denmark.The embryos and fetuses ranged from8 to 180 mm crown-rump length(CRL), corresponding to 6th–19th ovu-lation weeks. Informed consent wasobtained according to the guidelines ofthe Helsinki Declaration II. Addi-tional samples from legal first trimes-ter abortions from the Laboratory ofReproductive Biology, Rigshospitalet,and Frederiksberg Hospital (bothUniversity of Copenhagen) were alsoincluded in this study. Informed con-sent was obtained according to theHelsinki declaration II and approvedby the ethical committee of Copenha-gen and Frederiksberg Communities(KF 258206). Calculation of embry-onic and fetal age was based on infor-mation about the last menstrual pe-riod, and measurements of CRL andfoot lengths.
Antibodies, BlockingPeptides, and StainingReagents
Primary antibodies (dilutions in pa-renthesis for IF analysis): mousemonoclonal antibodies from Sig-maAldrich: anti-acetylated �-tubulin(acet. tb, cat. no. T6793; 1:5,000), anti-�-actin (cat. no. A5441; 1:5,000), and�-tubulin (cat. no. T5168; 1:2,000).Rabbit polyclonal anti-detyrosinated�-tubulin (glu-tub; ab48389; 1:500)was from Abcam. Polyclonal antibod-ies from Santa Cruz Biotechnology:goat anti-centrin (cat. no. sc-8719;1:500), goat anti-pericentrin (Pctn;cat.no. sc-28145; 1:500), rabbit anti-Gli2 transcription factor [Gli2(H-300),cat. no. sc-28674; Gli2(N-20), cat. no.sc-20290; Gli2(G-20), cat no. sc-20291;all diluted 1:200], rabbit anti-Gli3transcription factor [Gli3(H-280), cat.no. sc-20688; Gli3(N-19), cat. no. sc-6155; all diluted at 1:100], goat anti-Patched (Ptc; cat. no. sc-6149; 1:200).Two different rabbit anti-Smoothened
were used: (1) Smo (cat. no. LS-A2668;1:100) from MBL, and (2) Smo (cat. no.38686) from Abcam. Secondary anti-bodies for immunofluorescence mi-croscopy analysis (all from MolecularProbes and diluted at 1:600): AlexaFluor 568-conjugated goat anti-mouseIgG (cat. no. A11019), Alexa Fluor568-conjugated rabbit anti-mouse IgG(cat. no. A11061), Alexa Fluor 488-conjugated goat anti-rabbit IgG (cat.no. A11008), and Alexa Fluor 488-con-jugated donkey anti-goat IgG (cat. no.A11055). Secondary antibodies forWestern blot analysis (all from Jack-son ImmunoResearch and diluted1:5,000): alkaline phosphatase conju-gated goat anti-rabbit, rabbit anti-goat,and goat anti-mouse. Nuclei werestained with 4�,6-diamidine-2-pheny-lidole-dihydrochloride (DAPI, 1:600)from Molecular Probes, and F-actin wasstained with rhodamine phalloidin (1:100; Molecular Probes, Invitrogen).Blocking peptides sc-6155P, sc20290P,sc-20291P, and sc- sc-6149P were fromSanta Cruz Biotechnology.
Immunohistochemistry ofthe Developing HumanPancreas
Tissue specimens were dissected intoappropriate tissue blocks and fixed for12–24 hr at 4°C in one of the followingfixatives: 10% buffered formalin, 4%Formol-Calcium, Lillie’s AAF, orBouin’s fixatives. The specimens weredehydrated with graded alcohols,cleared in xylene, and embedded inparaffin wax (Merck, melting point52°C). Serial sections, 3–5 �m thick,were cut in transverse, sagittal, orhorizontal planes and placed on si-lanized slides. Representative sec-tions of each series were stained withhematoxylin and eosin, or with tolu-idine blue. For immunohistochemistry(IHC) sections were dewaxed, rehy-drated and washed in phosphate buff-ered saline (PBS; 137 mM NaCl, 2.6mM KCl, 6.5 mM Na2HPO4, and 1.5mM KH2PO4) as previously described(Teilmann and Christensen, 2005),followed by rinsing with blockingbuffer (5% bovine serum albumin inPBS) for 15 min before incubationwith primary antibodies for at least1.5 hr at room temperature or over-night at 4°C. The sections were thenwashed three times in blocking buffer,
incubated for 45 min in dark with flu-orochrome-conjugated secondary anti-bodies and DAPI in blocking buffer.After washing, sections were mountedin PBS with 70% glycerol and 2% N-propylgallate and sealed with nail pol-ish. Samples were observed on anEclipse E600 microscopes (Nikon, To-kyo, Japan) with EPI-FL3 filters andMagnaFire cooled CCD camera(Optronics, Goleta, CA), and digitalimages were processed using AdobePhotoshop 6.0.
Immunocytochemistry
Cells were grown on 25-mm-diametersterile polylysine-coated glass cover-slips for 3–5 days to �80% confluency,followed by serum starvation inDMEM/IMDM with 0.5% (PANC-1and CFPAC-1) and 0% (MEF) FCSand 1% P/S for 24, 48, and 72 hr, toinduce growth arrest and formation ofprimary cilia. The cells were brieflywashed in PBS, fixed for 15 min in 4%paraformaldehyde followed by washin blocking buffer and then permeabil-ized with 1% Triton X-100 in PBS for10 min. The coverslips were rinsedwith blocking buffer for 30 min beforeincubation with antibodies as de-scribed above for IHC, and mountedon microscope slides in anti-fademounting solution and sealed withnail polish. Samples were observed onan Eclipse E600 microscopes (Nikon,Tokyo, Japan) with EPI-FL3 filtersand MagnaFire cooled CCD camera(Optronics, Goleta, CA), and digitalimages were processed using AdobePhotoshop 6.0.
Sodium Dodecyl Sulfate-Polyacrylamide GelElectrophoresis and WesternBlot Analysis
Cell lysates were prepared by using1% sodium dodecyl sulfate (SDS) lysisbuffer. Lysates were sonicated andcentrifuged to separate off cell debris.Protein concentrations were mea-sured using a DC Protein Assay fromBio-Rad. Proteins were resolved bySDS-polyacrylamide gel electrophore-sis (SDS-PAGE) as described byChristensen et al. (2001). Briefly, pro-teins were separated under reducingconditions with NuPAGE 10% Bis-Tris precast gels (Invitrogen), fol-
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2049

lowed by electrophoretic transfer tonitrocellulose membranes (Invitro-gen). Before incubation with antibod-ies for 2 hr/over night the membraneswere blocked with 5% milk/TBST (10mM Tris/HCl [pH 7.5], 120 mM NaCl,0.1% Tween 20) and 0.5% Na-azide.Antibodies were diluted in 5% milk/TBST as indicated: anti-Ptc (1:200),anti-Gli2 (N-20) and (G-20) (1:200),anti-Gli3 (H-280) and (N-19) (1:100),anti-�-actin (1:5,000), and anti-�-tu-bulin (1:2,000). Membranes werewashed several times in TBST fol-lowed by incubation with alkalinephosphatase-conjugated secondaryantibodies. Blots were developed withBCIP/TNBT from KPL. The developedblots were scanned and band intensitywas estimated from arbitrary densito-metric values obtained using UN-SCAN-IT software. The blots werefurther processed for publication inAdobe Photoshop version 6.0.
SAG Stimulation
Cultures of PANC-1, CF-PAC1 cellsand NIH3T3 fibroblasts (80% conflu-ency) were serum starved for 48–72hr and incubated in the presence andin the absence of 1 �M SAG (AlexisBiochemicals, San Diego, CA) for 0and 24 hr, followed by IF analysiswith anti-Smo and anti-Gli3 and byWB analysis with anti-Ptc and anti-Gli2. Primary cilia were visualizedwith anti-acet. tb and nuclei withDAPI. All images were taken withequivalent time exposures.
Transfection of Cells WithFluorescent-Tagged Plasmids
CFPAC-1 cells were cultured on cov-erslips placed in six-well test plates(NUNC A/S, Denmark) and grown to70% confluency. The medium waschanged to DMEM/IMDM for 1 hr, fol-lowed by incubation with 1 �g/ml plas-mid (YFP-Smo, provided by P. Beachy)and 5 �l of lipofectamine 2000 (Invitro-gen) per well. After 4 hr of transfectionthe medium was changed to DMEM/IMDM with 10% FCS, and after thecells had reached 90% confluency theywere serum-starved for 48 hr to inducegrowth arrest. Cells were then fixed andpermeabilized and subjected to immu-nocytochemistry analysis as describedabove. NIH3T3 cells were cultured
and transfected under the same con-ditions as above, except that serum-free DMEM was used during the 4-hrtransfection period. The following plas-mids were used: full length Gli2::GFP,GFP-Gli2(FL), full length Gli3::GFP,GFP-Gli3(FL), Gli3repressor::GFP, GFP-Gli(R) (Buttitta et al., 2003). All the vec-tors were constructed in the pShuttlebackbone (Clontech).
ACKNOWLEDGMENTSWe thank Drs. Melissa Lutterodt andAnne Grete Byskov (Laboratory of Re-productive Biology, Rigshospitalet,Denmark) for expert help with collect-ing of human embryonic tissues. Wealso thank Dr. Kathryn Anderson(Memorial Sloan-Kettering CancerCenter, NY, USA) for the gift of thePtc�/� MEFs, and Simon Krabbeand Mette R. Hansen (Department ofBiology, University of Copenhagen,Denmark) for help with cultures of hu-man pancreatic ductal adenocarci-noma cell lines. This work was sup-ported by the Lundbeck Foundationand the Danish Science ResearchCouncil (S.T.C.), the Novo Nordisk/Novozymes Foundation (S.K.N.), theDanish Science Research Council(I.N.), funds from the Department ofBiology, University of Copenhagen(C.A.C.), and by a National Instituteof Health Grant (B.K.Y.).
REFERENCES
Apelqvist A, Ahlgren U, Edlund H. 1997.Sonic hedgehog directs specialised meso-derm differentiation in the intestine andpancreas. Curr Biol 7:801–804.
Ashizawa N, Endoh H, Hidaka K, Wa-tanabe M, Fukumoto S. 1997. Three-di-mensional structure of the rat pancreaticduct in normal and inflammated pan-creas. Microsc Res Tech 37:543–556.
Badano JL, Mitsuma N, Beales PL, Katsa-nis N. 2006. The ciliopathies: an emerg-ing class of human genetic disorders.Annu Rev Genomics Hum Genet 7:125–148.
Bak M, Hansen C, Tommerup N, LarsenLA. 2003. The Hedgehog signaling path-way—implications for drug targets incancer and neurodegenerative disorders.Pharmacogenomics 4:411–429.
Beachy PA, Karhadkar SS, Berman DM.2004. Tissue repair and stem cell renewalin carcinogenesis. Nature 432:324–331.
Berman DM, Karhadkar SS, Maitra A,Montes De Oca R, Gerstenblith MR,Briggs K, Parker AR, Shimada Y, Eshle-man JR, Watkins DN, Beachy PA. 2003.Widespread requirement for Hedgehog
ligand stimulation in growth of digestivetract tumours. Nature 425:846–851.
Buscher D, Ruther U. 1998. Expressionprofile of Gli family members and Shh innormal and mutant mouse limb develop-ment. Dev Dyn 211:88–96.
Buttitta L, Mo R, Hui CC, Fan CM. 2003.Interplays of Gli2 and Gli3 and their re-quirement in mediating Shh-dependentsclerotome induction. Development 130:6233–6243.
Cano DA, Murcia NS, Pazour GJ, HebrokM. 2004. Orpk mouse model of polycystickidney disease reveals essential role ofprimary cilia in pancreatic tissue organi-zation. Development 131:3457–3467.
Cano DA, Sekine S, Hebrok M. 2006. Pri-mary cilia deletion in pancreatic epithelialcells results in cyst formation and pancre-atitis. Gastroenterology 131:1856–1869.
Cano DA, Hebrok M, Zenker M. 2007. Pan-creatic development and disease. Gastro-enterology 132:745–762.
Caspary T, Larkins CE, Anderson KV.2007. The graded response to SonicHedgehog depends on cilia architecture.Dev Cell 12:767–778.
Chauvet V, Tian X, Husson H, Grimm DH,Wang T, Hieseberger T, Igarashi P, Ben-nett AM, Ibraghimov-Beskrovnaya O,Somlo S, Caplan MJ. 2004. Mechanicalstimuli induce cleavage and nucleartranslocation of the polycystin-1 C termi-nus. J Clin Invest 114:1433–1443.
Chen JK, Taipale J, Young KE, Maiti T,Beachy PA. 2002. Small molecule modu-lation of Smoothened activity. Proc NatlAcad Sci U S A 99:14071–14076.
Christensen ST, Ott CM. 2007. Cell signal-ing. A ciliary signaling switch. Science20;317:330–331.
Christensen ST, Guerra C, Wada Y, Valen-tin T, Angeletti RH, Satir P, HamasakiT. 2001. A regulatory light chain of cili-ary outer arm dynein in Tetrahymenathermophila. J Biol Chem 276:20048–20054.
Christensen ST, Pedersen LB, SchneiderL, Satir P. 2007. Sensory cilia and inte-gration of signal transduction in humanhealth and disease. Traffic 8:97–109.
Corbit KC, Aanstad P, Singla V, NormanAR, Stainier DY, Reiter JF. 2005. Verte-brate Smoothened functions at the pri-mary cilium. Nature 437:1018–1021.
Corbit KC, Shyer AE, Dowdle WE, Gaul-den J, Singla V, Reiter JF. 2008. Kif3aconstrains beta-catenin-dependent Wntsignalling through dual ciliary and non-ciliary mechanisms. Nat Cell Biol 10:70–76.
Davenport JR, Watts AJ, Roper VC, CroyleMJ, van Groen T, Wyss JM, Nagy TR,Kesterson RA, Yoder BK. 2007. Disrup-tion of intraflagellar transport in adultmice leads to obesity and slow-onset cys-tic kidney disease. Curr Biol 17:1586–1594.
Delmas P, Padilla F, Osorio N, Coste B,Raoux M, Crest M. 2004. Polycystins,calcium signaling, and human dis-eases. Biochem Biophys Res Commun322:1374 –1383.
2050 NIELSEN ET AL.

Eggenschwiler JT, Anderson KV. 2007.Cilia and developmental signaling. AnnuRev Cell Dev Biol 23:345–373.
Fliegauf M, Benzing T, Omran H. 2007.When cilia go bad: cilia defects and cil-iopathies. Nat Rev Mol Cell Biol 8:880–893.
Gerdes JM, Liu Y, Zaghloul NA, Leitch CC,Lawson SS, Kato M, Beachy PA, BealesPL, DeMartino GN, Fisher S, BadanoJL, Katsanis N. 2007. Disruption of thebasal body compromises proteasomalfunction and perturbs intracellular Wntresponse. Nat Genet 39:1350–1360.
Haycraft CJ, Banizs B, Aydin-Son Y,Zhang Q, Michaud EJ, Yoder BK. 2005.Gli2 and Gli3 localize to cilia and requirethe intraflagellar transport protein po-laris for processing and function. PLoSGenet 1:e53.
Hidaka K, Ashizawa N, Endoh H, Wa-tanabe M, Fukumoto S. 1995. Fine struc-ture of the cilia in the pancreatic duct ofWBN/Kob rat. Int J Pancreatol 18:207–213.
Hollande E, Salvador-Cartier C, Alvarez L,Fanjul M. 2005. Expression of a wild-type CFTR maintains the integrity of thebiosynthetic/secretory pathway in hu-man cystic fibrosis pancreatic duct cells.J Histochem Cytochem 53:1539–1552.
Huangfu D, Anderson KV. 2005. Cilia andHedgehog responsiveness in the mouse.Proc Natl Acad Sci U S A 102:11325–11330.
Huangfu D, Anderson KV. 2006. Signalingfrom Smo to Ci/Gli: conservation and di-vergence of Hedgehog pathways fromDrosophila to vertebrates. Development133:3–14.
Huangfu D, Liu A, Rakeman AS, MurciaNS, Niswander L, Anderson KV. 2003.Hedgehog signalling in the mouse re-quires intraflagellar transport proteins.Nature 426:83–87.
Jackerott M, Lee YC, Mollgard K, Kofod H,Jensen J, Rohleder S, Neubauer N,Gaarn LW, Lykke J, Dodge R, DalgaardLT, Sostrup B, Jensen DB, Thim L, NexoE, Thams P, Bisgaard HC, Nielsen JH.2006. Trefoil factors are expressed in hu-man and rat endocrine pancreas: differ-ential regulation by growth hormone.Endocrinology 147:5752–5759.
Kawahira H, Scheel DW, Smith SB, Ger-man MS, Hebrok M. 2005. Hedgehog sig-naling regulates expansion of pancreaticepithelial cells. Dev Biol 280:111–121.
Kayed H, Kleeff J, Esposito I, Giese T, Ke-leg S, Giese N, Buchler MW. 2003. Dis-tribution of Indian hedgehog and its re-ceptors patched and smoothened inhuman chronic pancreatitis. J Endorinol178:467–478.
Kayed H, Kleeff J, Osman T, Keleg S,Buchler MW, Friess H. 2006. Hedgehogsignaling in the normal and diseasedpancreas. Pancreas 32:119–129.
Kiprilov EK, Awan A, Desprat R, Velho M,Clement CA, Byskov AG, Andersen CY,Satir P, Bouhassira EE, Christensen ST,Hirsch RE. 2008. Human embryonicstem cells in culture possess primary
cilia with hedgehog signaling machinery.J Cell Biol 180:897–904.
Kodama T. 1983. A light and electron mi-croscopic study on the pancreatic ductalsystem. Acta Pathol Jpn 33:297–321.
Kottgen M. 2007. TRPP2 and autosomaldominant polycystic kidney disease. Bio-chim Biophys Acta 1772:836–850.
Koyama E, Young B, Nagayama M, Shi-bukawa Y, Enomoto-Iwamoto M,Iwamoto M, Maeda Y, Lanske B, Song B,Serra R, Pacifici M. 2007. ConditionalKif3a ablation causes abnormal hedge-hog signaling topography, growth platedysfunction, and excessive bone and car-tilage formation during mouse skeleto-genesis. Development 134:2159–2169.
Lau J, Kawahira H, Hebrok M. 2006.Hedgehog signaling in pancreas develop-ment and disease. Cell Mol Life Sci 63:642–652.
Lieber M, Mazzetta J, Nelson-Rees W,Kaplan M, Todaro G. 1975. Establish-ment of a continuous tumor-cell line(panc-1) from a human carcinoma of theexocrine pancreas. Int J Cancer 15:741–747.
Litingtung Y, Dahn RD, Li Y, Fallon JF,Chiang C. 2002. Shh and Gli3 are dis-pensable for limb skeleton formation butregulate digit number and identity. Na-ture 418:979–983.
Liu A, Wang B, Niswander LA. 2005a.Mouse intraflagellar transport proteinsregulate both the activator and repressorfunctions of Gli transcription factors. De-velopment. 132:3103–3111.
Liu W, Murcia NS, Duan Y, Weinbaum S,Yoder BK, Schwiebert E, Satlin LM.2005b. Mechanoregulation of intracellu-lar Ca2� concentration is attenuated incollecting duct of monocilium-impairedorpk mice. Am J Physiol Renal Physiol289:F978–F988.
Low SH, Vasanth S, Larson CH, Mukher-jee S, Sharma N, Kinter MT, Kane ME,Obara T, Weimbs T. 2006. Polycystin-1,STAT6, and P100 function in a pathwaythat transduces ciliary mechanosensa-tion and is activated in polycystic kidneydisease. Dev Cell 10:57–69.
Marigo V, Johnson RL, Vortkamp A, TabinCJ. 1996. Sonic hedgehog differentiallyregulates expression of GLI and GLI3during limb development. Dev Biol 180:273–283.
May SR, Ashique AM, Karlen M, Wang B,Shen Y, Zarbalis K, Reiter J, Ericson J,Peterson AS. 2005. Loss of the retro-grade motor for IFT disrupts localizationof Smo to cilia and prevents the expres-sion of both activator and repressor func-tions of Gli. Dev Biol 287:378–389.
Merkel CE, Karner CM, Carroll TJ. 2007.Molecular regulation of kidney develop-ment: is the answer blowing in the Wnt?.Pediatr Nephrol 22:1825–1838.
Morton JP, Mongeau ME, Klimstra DS,Morris JP, Lee YC, Kawaguchi Y, WrightVE, Hebrok M, Lewis BC. 2006. Sonichedgehog acts at multiple stages duringpancreatic tumorigenesis. Proc NatlAcad Sci U S A 104:5103–5108.
Nauli SM, Alenghat FJ, Luo Y, Williams E,Vassilev P, Li X, Elia AE, Lu W, BrownEM, Quinn SJ, Ingber DE, Zhou J. 2003.Polycystins 1 and 2 mediate mech-anosensation in the primary cilium ofkidney cells. Nat Genet 33:129–137.
Novak I, Hede SE, Hansen MR. 2008.Adenosine receptors in rat and humanpancreatic ducts stimulate chloridetransport. Pflugers Arch 456:437–447.
Odent S, Atti-Bitach T, Blayau M, MathieuM, Aug J, Delezo de AL, Gall JY, LeMarec B, Munnich A, David V, Veke-mans M. 1999. Expression of the Sonichedgehog (SHH) gene during early hu-man development and phenotypic ex-pression of new mutations causing holo-prosencephaly. Hum Mol Genet 8:1683–1689.
Ostrer H, Wilson DI, Hanley NA. 2006.Human embryo and early fetus research.Clin Genet 70:98–107.
Pan Y, Bai CB, Joyner AL, Wang B. 2006.Sonic hedgehog signaling regulates Gli2transcriptional activity by suppressingits processing and degradation. Mol CellBiol 26:3365–3377.
Pazour GJ, Dickert BL, Vucica Y, SeeleyES, Rosenbaum JL, Witman GB, ColeDG. 2000. Chlamydomonas IFT88 andits mouse homologue, polycystic kidneydisease gene tg737, are required for as-sembly of cilia and flagella. J Cell Biol151:709–718.
Pazour GJ, San Agustin JT, Follit JA,Rosenbaum JL, Witman GB. 2002. Poly-cystin-2 localizes to kidney cilia and theciliary level is elevated in orpk mice withpolycystic kidney disease. Curr Biol 12:R378–R380.
Pedersen LB, Schroder JM, Veland IR,Christensen ST. 2008. Assembly of pri-mary cilia. Dev Dyn 2008 [Epub ahead ofprint].
Praetorius HA, Spring KR. 2001. Bendingthe MDCK cell primary cilium increasesintracellular calcium. J Membr Biol 184:71–79.
Praetorius HA, Spring KR. 2003. The renalcell primary cilium functions as a flowsensor. Curr Opin Nephrol Hypertens12:517–520.
Rohatgi R, Milenkovic L, Scott MP. 2007.Patched1 regulates hedgehog signalingat the primary cilium. Science 317:372–376.
Rosenbaum JL, Witman GB. 2002. In-traflagellar transport. Nat Rev Mol CellBiol 3:813–825.
Ruiz-Perez VL, Blair HJ, Rodriguez-An-dres ME, Blanco MJ, Wilson A, Liu YN,Miles C, Peters H, Goodship JA. 2007.Evc is a positive mediator of Ihh-regu-lated bone growth that localises at thebase of chondrocyte cilia. Development134:2903–2912.
Satir P, Christensen ST. 2007. Overview ofstructure and function of mammaliancilia. Annu Rev Physiol 69:377–400.
Schneider L, Clement CA, Teilmann SC,Pazour GJ, Hoffmann EK, Satir P,Christensen ST. 2005. PDGFRalphaal-pha signaling is regulated through the
PRIMARY CILIA AND HEDGEHOG SIGNALING IN PANCREAS 2051

primary cilium in fibroblasts. Curr Biol15:1861–1866.
Schoumacher RA, Ram J, Iannuzzi MC,Bradbury NA, Wallace RW, Tom Hon C,Kelly DR, Schmid SM, Gelder FB, RadoTA, Frizzell RA. 1990. A cystic fibrosispancreatic adenocarcinoma cell line.Proc Natl Acad Sci U S A 87:4012–4016.
Schweitzer R, Vogan KJ, Tabin CJ. 2000.Similar expression and regulation ofGli2 and Gli3 in the chick limb bud.Mech Dev 98:171–174.
Simons M, Gloy J, Ganner A, BullerkotteA, Bashkurov M, Kronig C, Schermer B,Benzing T, Cabello OA, Jenny A,Mlodzik M, Polok B, Driever W, Obara T,Walz G. 2005. Inversin, the gene productmutated in nephronophthisis type II,functions as a molecular switch betweenWnt signaling pathways. Nat Genet 37:537–543.
Teilmann SC, Christensen ST. 2005. Local-ization of the angiopoietin receptorsTie-1 and Tie-2 on the primary cilia inthe female reproductive organs. Cell BiolInt 29:340–346.
Thayer SP, di Magliano MP, Heiser PW,Nielsen CM, Roberts DJ, Lauwers GY,Qi YP, Gysin S, Fernandez-del CastilloC, Yajnik V, Antoniu B, McMahon M,Warshaw AL, Hebrok M. 2003. Hedge-hog is an early and late mediator of pan-creatic cancer tumorigenesis. Nature425:851–856.
van den Brink GR. 2007. Hedgehog signal-ing in development and homeostasis ofthe gastrointestinal tract. Physiol Rev87:1343–1375.
Vierkotten J, Dildrop R, Peters T, Wang B,Ruther U. 2007. Ftm is a novel basalbody protein of cilia involved in Shh sig-nalling. Development 134:2569–2577.
Wang B, Fallon JF, Beachy PA. 2000.Hedgehog-regulated processing of Gli3produces an anterior/posterior repressorgradient in the developing vertebratelimb. Cell 100:423–434.
Weimbs T. 2007. Polycystic kidney diseaseand renal injury repair: common path-ways, fluid flow, and the function of poly-cystin-1. Am J Physiol Renal Physiol293:F1423–F1432.
Wilson ME, Scheel D, German MS. 2003.Gene expression cascades in pancreaticdevelopment. Mech Dev 120:65–80.
Yoder BK. 2007. Role of primary cilia inthe pathogenesis of polycystic kidneydisease. J Am Soc Nephrol 18:1381–1388.
Yoder BK, Tousson A, Millican L, Wu JH,Bugg CE Jr, Schafer JA, Balkovetz DF.2002a. Polaris, a protein disrupted inorpk mutant mice, is required for assem-bly of renal cilium. Am J Physiol RenalPhysiol 282:F541–F552.
Yoder BK, Hou X, Guay-Woodford LM.2002b. The polycystic kidney diseaseproteins, polycystin-1, polycystin-2, po-laris, and cystin, are co-localized in re-nal cilia. J Am Soc Nephrol 13:2508 –2516.
Zhang Q, Davenport JR, Croyle MJ, Hay-craft CJ, Yoder BK. 2005. Disruption ofIFT results in both exocrine and endo-crine abnormalities in the pancreas ofTg737(orpk) mutant mice. Lab Invest 85:45–64.
2052 NIELSEN ET AL.