
Katanin p80 Regulates Human Cortical Development by Limiting Centriole and Cilia Number
55
Neuron Article Katanin p80 Regulates Human Cortical Development by Limiting Centriole and Cilia Number Wen F. Hu, 1,2,3,5,6 Oz Pomp, 19 Tawfeg Ben-Omran, 12 Andrew Kodani, 18 Katrin Henke, 4,9 Ganeshwaran H. Mochida, 1,2,7,10 Timothy W. Yu, 1,7,11 Mollie B. Woodworth, 1,2,3,7 Carine Bonnard, 19 Grace Selva Raj, 19 Thong Teck Tan, 19 Hanan Hamamy, 21 Amira Masri, 23 Mohammad Shboul, 19 Muna Al Saffar, 1,2,13 Jennifer N. Partlow, 1,2,3 Mohammed Al-Dosari, 17 Anas Alazami, 14 Mohammed Alowain, 15,16 Fowzan S. Alkuraya, 14,16 Jeremy F. Reiter, 18 Matthew P. Harris, 4,9,24, * Bruno Reversade, 19,20,22,24, * and Christopher A. Walsh 1,2,3,5,6,7,8,24, * 1 Division of Genetics and Genomics, Department of Medicine 2 Manton Center for Orphan Disease Research 3 Howard Hughes Medical Institute 4 Department of Orthopedic Research Boston Children’s Hospital, Boston, MA 02115, USA 5 Program in Neuroscience 6 Harvard MD-PhD MSTP Program 7 Department of Pediatrics 8 Department of Neurology 9 Department of Genetics Harvard Medical School, Boston, MA 02115, USA 10 Pediatric Neurology Unit, Department of Neurology, Massachusetts General Hospital, Boston, MA 02114, USA 11 Program in Medical and Population Genetics, Broad Institute of MIT and Harvard University, Cambridge, MA 02142, USA 12 Clinical and Metabolic Genetics Division, Department of Pediatrics, Hamad Medical Corporation, Doha, 3050, Qatar 13 Department of Pediatrics, Faculty of Medicine and Health Sciences, United Arab Emirates University, Al-Ain, United Arab Emirates 14 Department of Genetics 15 Department of Medical Genetics King Faisal Specialist Hospital and Research Centre, Riyadh 11211, Saudi Arabia 16 Department of Anatomy and Cell Biology, College of Medicine, Alfaisal University, Riyadh 11533, Saudi Arabia 17 Department of Pharmacognosy, College of Pharmacy, King Saud University, Riyadh 11451, Saudi Arabia 18 Department of Biochemistry and Biophysics, Cardiovascular Research Institute, University of California, San Francisco, San Francisco, CA 94158, USA 19 Institute of Medical Biology, Human Genetics and Embryology Laboratory 20 Institute of Molecular and Cellular Biology A*STAR, Singapore 138648, Singapore 21 Department of Genetic Medicine and Development, Geneva University, Geneva 1211, Switzerland 22 Department of Paediatrics, National University of Singapore, Singapore 119260, Singapore 23 Department of Paediatrics, Faculty of Medicine, University of Jordan, Amman, 11942, Jordan 24 Co-senior author *Correspondence: [email protected] (M.P.H.), [email protected] (B.R.), [email protected] (C.A.W.) http://dx.doi.org/10.1016/j.neuron.2014.12.017 SUMMARY Katanin is a microtubule-severing complex whose catalytic activities are well characterized, but whose in vivo functions are incompletely understood. Human mutations in KATNB1, which encodes the noncatalytic regulatory p80 subunit of katanin, cause severe microlissencephaly. Loss of Katnb1 in mice confirms essential roles in neurogenesis and cell survival, while loss of zebrafish katnb1 reveals specific roles for katnin p80 in early and late developmental stages. Surprisingly, Katnb1 null mutant mouse embryos display hallmarks of aberrant Sonic hedgehog signaling, including holo- prosencephaly. KATNB1-deficient human cells show defective proliferation and spindle structure, while Katnb1 null fibroblasts also demonstrate a remarkable excess of centrioles, with supernumer- ary cilia but deficient Hedgehog signaling. Our results reveal unexpected functions for KATNB1 in regulating overall centriole, mother centriole, and cilia number, and as an essential gene for normal Hedgehog signaling during neocortical development. INTRODUCTION Although human genetics has identified essential roles for many centriole- and cilia-related proteins during human develop- ment, the functional relationships between these two crucial or- ganelles are less well understood. Mutations in genes encoding centrosomal proteins cause a wide range of syndromes, notably 1240 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Katanin p80 Regulates Human Cortical Development by Limiting Centriole and Cilia Number

Neuron
Article
Katanin p80 Regulates Human Cortical Developmentby Limiting Centriole and Cilia NumberWen F. Hu,1,2,3,5,6 Oz Pomp,19 Tawfeg Ben-Omran,12 Andrew Kodani,18 Katrin Henke,4,9 Ganeshwaran H. Mochida,1,2,7,10
Timothy W. Yu,1,7,11 Mollie B. Woodworth,1,2,3,7 Carine Bonnard,19 Grace Selva Raj,19 Thong Teck Tan,19
Hanan Hamamy,21 Amira Masri,23 Mohammad Shboul,19 Muna Al Saffar,1,2,13 Jennifer N. Partlow,1,2,3
Mohammed Al-Dosari,17 Anas Alazami,14 Mohammed Alowain,15,16 Fowzan S. Alkuraya,14,16 Jeremy F. Reiter,18
Matthew P. Harris,4,9,24,* Bruno Reversade,19,20,22,24,* and Christopher A. Walsh1,2,3,5,6,7,8,24,*1Division of Genetics and Genomics, Department of Medicine2Manton Center for Orphan Disease Research3Howard Hughes Medical Institute4Department of Orthopedic Research
Boston Children’s Hospital, Boston, MA 02115, USA5Program in Neuroscience6Harvard MD-PhD MSTP Program7Department of Pediatrics8Department of Neurology9Department of GeneticsHarvard Medical School, Boston, MA 02115, USA10Pediatric Neurology Unit, Department of Neurology, Massachusetts General Hospital, Boston, MA 02114, USA11Program in Medical and Population Genetics, Broad Institute of MIT and Harvard University, Cambridge, MA 02142, USA12Clinical and Metabolic Genetics Division, Department of Pediatrics, Hamad Medical Corporation, Doha, 3050, Qatar13Department of Pediatrics, Faculty of Medicine and Health Sciences, United Arab Emirates University, Al-Ain, United Arab Emirates14Department of Genetics15Department of Medical Genetics
King Faisal Specialist Hospital and Research Centre, Riyadh 11211, Saudi Arabia16Department of Anatomy and Cell Biology, College of Medicine, Alfaisal University, Riyadh 11533, Saudi Arabia17Department of Pharmacognosy, College of Pharmacy, King Saud University, Riyadh 11451, Saudi Arabia18Department of Biochemistry and Biophysics, Cardiovascular Research Institute, University of California, San Francisco, San Francisco,
CA 94158, USA19Institute of Medical Biology, Human Genetics and Embryology Laboratory20Institute of Molecular and Cellular Biology
A*STAR, Singapore 138648, Singapore21Department of Genetic Medicine and Development, Geneva University, Geneva 1211, Switzerland22Department of Paediatrics, National University of Singapore, Singapore 119260, Singapore23Department of Paediatrics, Faculty of Medicine, University of Jordan, Amman, 11942, Jordan24Co-senior author*Correspondence: [email protected] (M.P.H.), [email protected] (B.R.),
[email protected] (C.A.W.)
http://dx.doi.org/10.1016/j.neuron.2014.12.017
SUMMARY
Katanin is a microtubule-severing complex whosecatalytic activities are well characterized, but whosein vivo functions are incompletely understood.Human mutations in KATNB1, which encodes thenoncatalytic regulatory p80 subunit of katanin,cause severe microlissencephaly. Loss of Katnb1in mice confirms essential roles in neurogenesisand cell survival, while loss of zebrafish katnb1reveals specific roles for katnin p80 in early andlate developmental stages. Surprisingly, Katnb1null mutant mouse embryos display hallmarks ofaberrant Sonic hedgehog signaling, including holo-prosencephaly. KATNB1-deficient human cellsshow defective proliferation and spindle structure,
1240 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc
while Katnb1 null fibroblasts also demonstrate aremarkable excess of centrioles, with supernumer-ary cilia but deficient Hedgehog signaling. Ourresults reveal unexpected functions for KATNB1in regulating overall centriole, mother centriole,and cilia number, and as an essential gene fornormal Hedgehog signaling during neocorticaldevelopment.
INTRODUCTION
Although human genetics has identified essential roles for many
centriole- and cilia-related proteins during human develop-
ment, the functional relationships between these two crucial or-
ganelles are less well understood. Mutations in genes encoding
centrosomal proteins cause a wide range of syndromes, notably
.

Neuron
Katanin p80 Limits Centriole and Cilia Number
microcephaly, which is characterized by reduced brain size with
or without other features, such as reduced somatic size. Micro-
cephaly-associated mutations in genes encoding centrosomal
and pericentriolar proteins, including MCPH1, CDK5RAP2,
ASPM, CENPJ, WDR62, CEP152, CASC5, STIL, CEP135,
CEP63, NIN, PCNT, and POC1A, implicate a centrosomal
pathway important during human neurogenesis (Hu et al.,
2014; Mahmood et al., 2011; Thornton and Woods, 2009). How-
ever, cellular mechanisms by which centrioles control neurogen-
esis are still unclear.
A second class of diseases, broadly termed ‘‘ciliopathies,’’
manifest with a range of features, but common themes include
retinal disease, left-right asymmetry defects, polydactyly, hepa-
tobiliary disease, renal cysts, and obesity (Hildebrandt et al.,
2011). These diseases reflect functions of cilia in a wide range
of cellular processes, including mechanosensation, chemosen-
sation, and developmental responses to patterning signals and
growth factors (Nigg and Raff, 2009; Novarino et al., 2011; Oh
and Katsanis, 2012).
Although cilia and centrosomes both function centrally in
neocortical development, their interrelationships in the context
of neurogenesis are not well understood. The neocortex is
formed from a layer of neuroepithelial cells that divide
and expand to form radial glial progenitors, which subse-
quently give rise to intermediate progenitors and, ultimately,
neurons (Greig et al., 2013; Lui et al., 2011). Neuronal progen-
itors lining the lateral ventricle protrude apical primary cilia to
receive critical cues from the cerebrospinal fluid, such as
IGF2 and Sonic hedgehog (Shh), in order to maintain prolifera-
tive cell divisions (Lehtinen et al., 2011; Saade et al., 2013;
Tong et al., 2014). Mutations that impair the function of
the cilium disrupt progenitor polarity and neurogenesis (Higgin-
botham et al., 2013; Lee and Gleeson, 2011; Willaredt et al.,
2008). Recently, the ciliary membrane and a ciliary remnant
have been shown to remain tightly associated with the
mother, or older, more mature centriole in dividing apical neu-
roepithelial cells. The asymmetrical inheritance of both the
mother centriole and this apical ciliary membrane allows asym-
metrical cilium outgrowth and maintenance of apical stem cells
(Paridaen et al., 2013; Wang et al., 2009). These data suggest
that relationships between mother centrioles and cilia are
crucial to neurogenesis and the maintenance of stem cell
character.
Here, we describe recessive mutations in KATNB1, encoding
the p80 subunit of the microtubule-severing enzyme katanin,
that cause severe microcephaly with simplification of cortical
gyri and sulci, or microlissencephaly. By modeling the condition
in mice and zebrafish, we find that katanin p80 is a crucial regu-
lator of early embryonic development. Remarkably, KATNB1-
deficient cells show abnormalities of both the centriole and
primary cilium. In mouse embryonic fibroblasts (MEFs), loss
of katanin p80 leads to the formation of supernumerary centri-
oles capable of forming multipolar spindles and nucleating
ectopic cilia. Evidence of perturbed Shh signaling further sup-
ports disruption of ciliary function. Our data provide a new
mechanism of microcephaly that involves katanin p80-medi-
ated regulation of interactions between centriole and ciliary
signaling.
Ne
RESULTS
Mutations in KATNB1 Cause MicrolissencephalyThree unrelated Middle Eastern families presented with individ-
uals affected with severe microcephaly, global developmental
delay, and seizures. MRI of the affected individuals revealed
dramatically reduced brain size and cortical volume with simpli-
fied gyri, shallow sulci, and enlarged lateral ventricles posteriorly
(Figure 1A), with relative sparing of the midbrain, basal ganglia,
and cerebellum. Affected individuals also displayed mild facial
dysmorphisms and sloping foreheads, consistent with reduced
cranial volume (see Figure S1A available online).
Family 1 is a large Jordanian family with five affected individ-
uals from related, consanguineous nuclear families; siblings of
the affected individuals are all reported to be healthy (Figure 1B).
Family 2 originates fromSaudi Arabia, and the affectedmale pro-
band is the third child of healthy, first-cousin parents (Figure 1B0),with two healthy older siblings. Family 3 is of Palestinian origin,
and the affected individual is the fourth child of two healthy par-
ents with no reported consanguinity (Figure 1B0 0). A sibling of
Proband 3 died from a viral illness at age 2, while other siblings
are healthy. Several paternal first cousins were reported to
display a similar microcephaly and seizure phenotype, although
medical records and DNA samples were unavailable.
Medical genetic and neurological evaluation of the affected
individuals at birth and throughout life revealed dramatically
reduced head circumference, disproportionate to height and
weight (Figures S1B–S1D). Detailed clinical information on all
affected individuals is available in Table S1. The severely
reduced brain size, simplified gyri and enlarged ventricles,
especially posteriorly, and relative sparing of the brain stem
and cerebellum seen on MRI in affected individuals from
all three families bear a striking resemblance to the micro-
lissencephaly caused by mutations in NDE1 (Alkuraya et al.,
2011; Bakircio�glu et al., 2011), and so we use the same term
henceforth.
The consanguineous pedigrees implicated recessive inheri-
tance of rare, pathogenic variants. To identify the causative
mutations in these families, we undertook a combination of ho-
mozygosity mapping, whole-exome sequencing, and targeted
next-generation sequencing (see Experimental Procedures for
further details). In Family 1, mapping of shared regions that are
homozygous and identical-by-descent (IBD) in the affected indi-
viduals, and exclusion of common homozygous segments
shared by unaffected family members, identified a single shared
IBD candidate locus totaling 9 Mb on Chromosome 16 (Fig-
ure S1E). Subsequent whole-exome sequencing of Proband 1
revealed a single, unique homozygous variant present in the
region of IBD. Whole-exome sequencing in Proband 2 identified
3 homozygous, rare, protein-altering variants, and targeted
sequencing of coding exons within blocks of homozygosity
greater than 2 cM in Proband 3 identified seven homozygous,
rare, protein-altering variants. Crossreferencing all three families
identified homozygous deleterious mutations in a single, over-
lapping gene, KATNB1. Sanger sequencing confirmed precise
familial segregation of disease-associated variants (Figures
S1F–S1F0 0), and all three variants were absent from European
or ethnically matched control populations.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1241

Figure 1. Mutations in KATNB1 Cause Microlissencephaly
(A) MRI images of affected individuals show reduced cortical size (ctx), simplification of gyral folding pattern, enlarged lateral ventricles (lv) posteriorly and thinning
of the corpus callosum (cc), with relative sparing of the cerebellum (cb), basal ganglia (bg), thalamus (th), and brainstem (br). Scale bar, 50 mm.
(legend continued on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1242 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Neuron
Katanin p80 Limits Centriole and Cilia Number
KATNB1 encodes the p80 subunit of katanin, a microtubule-
severing enzyme comprised of a p60 catalytic subunit and a
p80 regulatory subunit (McNally and Vale, 1993). Family 1 carries
amutation that abolishes the initiator ATGcodon (Figure 1C), pre-
dicted to result either in complete loss of protein, or potential pro-
duction of an N-terminally truncated protein from a downstream
conserved in-frame methionine at position 57. Family 2 carries a
missense mutation that converts a highly conserved glycine to a
tryptophan (Figure 1C0). Family 3 carries a splice site mutation at
the exon-intron boundary of exon 6 that is predicted to abolish all
splice donor activity (Figure 1C0 0), as the first base pair of the
splice donor site is 100% conserved in eukaryotes (Mount,
1982; Senapathy et al., 1990). Katanin p80 contains sixWD40 re-
peats in the N terminus of the protein, and the mutations in
Families 2 and 3 are located within the first and third WD40
repeats, respectively (Figure 1D). The predicted N-terminal trun-
cated product in Family 1 also disrupts the WD40 domains.
KATNB1 DNA and protein sequences are highly evolutionarily
conserved. Using several different methods of model averaging
(Zhang et al., 2006), we calculated nonsynonymous to synony-
mous substitution ratios (Ka/Ks) between 0.03 and 0.08, indi-
cating extremely strong negative selection of nonsynonymous
variation. The specific proband nucleotide mutation sites are
also highly conserved, with GERP scores of 4.97, 4.28, and
5.01 and for the start codon, missense, and splice site mutation
sites, respectively (Cooper et al., 2005; Davydov et al., 2010).
Additionally, for the missense mutation, comparing the MultiZ
alignment across 100 vertebrate species, all but one species
contain glycine at that residue (Blanchette et al., 2004; Karolchik
et al., 2014). Together, these data indicate that theKATNB1 gene
and the proband mutation sites are extremely well conserved,
and that mutation of these sites is most likely pathogenic.
Mutations in KATNB1 Alter mRNA Splicing and ProteinQuantityThe high conservation of KATNB1 and the highly conserved pro-
tein structure of WD40 repeats (Neer et al., 1994) suggest that
the identified mutations would severely disrupt katanin p80 pro-
tein stability and function. Expression studies in primary fibro-
blasts and induced pluripotent stem cells (iPSCs) derived from
Proband 1 identified reductions in both katanin p80 and p60 in
human cells. The start codonmutation causes the loss of the first
methionine, and translation is predicted to initiate at the next in-
frame methionine 56 amino acids downstream (Figure S2B).
KATNB1 and KATNA1 mRNA are detectable at comparable
levels in Proband 1-derived iPSCs compared to control cell lines
by quantitative RT-PCR (Figure 2A). However, full-length katanin
p80 is not detectable in Proband 1-derived primary fibroblasts or
iPSCs, and in iPSCs we find a new, smaller katanin p80 protein
product consistent with the predicted N-terminal truncated
(B–B0 0) Pedigrees of families with microlissencephaly. Square, male; circle, fema
shading, reported affected individual, medical records unavailable; double lines
collected.
(C–C0 0) Mutation in Family 1 abolishes start ATG codon. Mutation in Family 3 is at a
tryptophan.
(D) Predicted protein structure of katanin p80. Mutations lie at first amino acid an
See also Figure S1, Table S1, and Movie S1.
Ne
form (Figure 2B). Remarkably, p60 protein was also undetect-
able in primary fibroblasts and greatly reduced in iPSCs despite
unchanged transcript levels (Figures 2A and 2B), most likely re-
sulting from a posttranscriptional event. A lymphoblastoid cell
line derived from Proband 2 showed an 84% reduction in katanin
p80 protein compared with a control line, with protein minimally
detectable after long exposure (Figure 2C), suggesting that the
missense allele is either null, due to defective function of the
mutant protein, or hypomorphic, with some residual protein
and residual activity. Interestingly, we also find a concomitant
64% decrease of the katanin p80 binding partner katanin p60
in Proband 2-derived lymphoblasts, compared with control (Fig-
ure 2C), suggesting that the loss of katanin p80 impairs katanin
p60 stability.
For Family 3, we analyzed mRNA splicing using a minigene
comprised of full-length cDNA, including the introns immediately
upstreamanddownstreamof the splicemutation site (Figure 2D).
We transfected tagged wild-type and mutated Proband 3 mini-
gene constructs into HEK293T cells, followed by RT-PCR, and
found that, compared with wild-type, the mutant minigene cre-
ates a smaller RNA product corresponding to skipping of exon
6 (dele6, Figure 2E). Sanger sequencing of the RT-PCR product
identified the presence of the exon 5–exon 7 boundary (Fig-
ure 2F). Expression studies showed that the splicingmutation re-
sults in stable, albeit functionally defective, protein. Transfection
of taggedwild-type and Proband 3minigenes, andwild-type and
exon 6-deleted (dele6) cDNAs, into HEK293T cells with quantita-
tive western blot revealed a 51%–77% reduction in katanin p80
from the mutant minigene and cDNAs compared with their wild-
type counterparts (Figure 2G). Because exon 6 is an in-frame
exon, and mutant protein is present at lower quantities than
wild-type, this mutant form may be hypomorphic. Whereas
tagged, wild-type katanin p80 localizes to the centrosome of
interphase cells (Figure 2H), most tagged dele6 protein accumu-
lates in the nucleus during interphase, with minimal localization
to the centrosome (Figure 2H), indicating that themisspliced pro-
tein is functionally perturbed. In summary, all three mutations
disrupt the N-terminal WD40 repeats of KATNB1, result in lower
levels of katanin p80 aswell as p60 protein, yet have the potential
to produce defective proteins with some partial function.
Katanin p80 Is Required for Embryonic SurvivalTo study functions of katanin p80 in vivo, we generated a Katnb1
gene-trap (gt) mouse line, with the gene-trap vector inserted
between coding exons 2 and 3 (Figure 3A). We confirmed by
RT-PCR that Katnb1 coding exon 2 splices fully to the
Engrailed-2 splice acceptor of the gene-trap vector, with no
detectable wild-type Katnb1 mRNA in homozygous gene-trap
mutants (Figures 3B and 3C). Katanin p80 protein is reduced
by 51% in heterozygous embryos and is undetectable in
le; red arrowhead, affected proband; black shading, affected individual; gray
, consanguineous marriages; diagonal line, deceased; asterisk, DNA sample
50 splice site. Missense mutation in Family 2 converts a conserved glycine to a
d in WD40 domains.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1243

Figure 2. Mutant KATNB1 Alleles Produce Less Protein Than Wild-Type Alleles, and dele6 Mutant Protein Is Mislocalized
(A) qRT-PCR of Proband 1-derived fibroblast and iPSC lines shows presence of KATNB1 and KATNA1mRNA at levels comparable to control cell lines. Unpaired
t test, p < 0.05; ns, not statistically significant.
(B) Western blot of Proband 1-derived fibroblast and iPSC lines shows absence of full-length katanin p80 and presence of smaller protein product compared to
control (top). Katanin p60 levels are also reduced (bottom).
(C) Western blot of Proband 2-derived lymphoblastoid cell lines shows near absence of katanin p80, detectable only at high exposure (top), and reduction of
katanin p60 (bottom). Quantification using LI-COR imaging system.
(legend continued on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1244 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Neuron
Katanin p80 Limits Centriole and Cilia Number
homozygous mutant embryos (Figure 3D), indicating that the
gene-trap allele is almost certainly null. Similar to our primary hu-
man cell line data (Figures 2B and 2C), katanin p60 protein levels
are reduced by 39% and 72% in the heterozygous and homozy-
gous Katnb1 gene-trap embryos, respectively (Figure 3E).
Homozygous null Katnb1 gene-trap embryos die embryoni-
cally, with dramatically reduced body size by embryonic day
15.5 (E15.5) (Figures 3F and 3G). Embryos are found along a
spectrum of phenotypic severity, and some very severely
affected embryos die as early as E12.5. All homozygous mutant
embryos are grossly morphologically abnormal, with features
including pallor of the liver, microophthalmia to anophthalmia,
and reduced limb bud outgrowth (Figure 3H). In addition, homo-
zygous mutant embryos show forebrain abnormalities ranging
frommicrocephaly to holoprosencephaly, in which the two hemi-
spheres of the brain fail to separate, resulting in a single ventricle
(Figures 3H–3J). Histopathological analysis of the embryos re-
vealed a lack of red blood cells in the liver, suggesting failed
definitive erythropoiesis, which is a potential cause of embryonic
death (data not shown) (Baron, 2013). Underdeveloped limb
buds, anophthalmia, and holoprosencephaly are commonly
seen in mouse mutants with defective Shh signaling (Chiang
et al., 1996; Hayhurst and McConnell, 2003; Litingtung et al.,
2002) and are associated with human mutations in SHH (Belloni
et al., 1996; Roessler et al., 1996), while Indian hedgehog,
another hedgehog family protein, has been implicated in defini-
tive erythropoiesis (Cridland et al., 2009). The overlap of pheno-
types between homozygous Katnb1 gene-trap mice and Shh
pathway mutants raises the unexpected possibility that katanin
p80 might be involved in modulating Shh signaling. These data
indicate that katanin p80 is required for embryonic development
and survival, and confirm the association of the human muta-
tions with the severe brain malformations.
Katanin p80 Is Required for Early Embryogenesis andFormation of Anterior Structures in ZebrafishAs a second, independent model of katanin loss of function dur-
ing development, we utilized the high conservation of the katanin
p80 protein sequence and gene structure in zebrafish (Figures
4A and S3A) and targeted the exon 6-intron 6 boundary near
the Proband 3 mutation site with TALEN endonucleases (Fig-
ure 4B; Table S2). We generated a series of mutant alleles
harboring different frameshift as well as in-frame deletions and
insertions at the target site (Figures 4B and S3B). Most of the
alleles are frameshift mutations and are predicted to lead to early
termination and, consequently, truncated proteins (Figure 4C
and S3B).
Mutant progeny from F1 carriers of truncated alleles are viable.
However, progeny of females deficient for katnb1, although able
(D) Schematic of the splice site minigene cDNA construct. Primer arrows show l
(E) RT-PCR results of minigene assay. Smaller product in splice mutant minigen
(F) Sanger sequencing of smaller spliced product (asterisk in E) confirms skippin
(G) Western blot of transfected minigenes and cDNAs shows reduced protein lev
imaging system.
(H) Wild-type EGFP-tagged katanin p80 localizes to the centrosome (left; Pericen
with minimal centrosomal localization (right). Scale bar, 10 um.
See also Figure S2.
Ne
to develop through 70% epiboly, show a wide spectrum of phe-
notypes stemming from defects in gastrulation and formation of
anterior structures, ranging from milder microcephaly to more
severe anencephaly and early embryonic death (Figures 4D,
4E, and S3C). The similar phenotypic results observed in mice
and zebrafish deficient for Katnb1 demonstrate that katanin
plays a major role during early embryogenesis and patterning
of anterior structures. Since the humans we identified with
KATNB1mutations survive beyond birth into childhood, the em-
bryonic lethality of Katnb1 deficiencies suggests that the three
identified human alleles may retain partial function. Interestingly,
the maternal rescue observed in the zebrafish (Figure 4E) indi-
cates that normal maternal katnb1 mRNA is sufficient to permit
generally normal growth and patterning during postembryonic
development in the absence of zygotic katnb1, suggesting that
variation in developmental timing of katanin p80 function also
affects phenotypic severity.
Katnb1 Is Ubiquitously Expressed in the Brain andCritical for Neocortical DevelopmentGiven the striking brain morphological phenotypes in the pro-
bands and homozygous gene-trap mouse embryos, we investi-
gated functions of katanin p80 during cortical development.
Katanin p80 is expressed in the mouse cerebral cortex
throughout development (Figure 5A). To evaluate cell-type-spe-
cific expression, we took advantage of the presence of a LacZ
cassette in the gene-trap vector and stained for LacZ protein
in the neocortex of heterozygous embryos. We found ubiquitous
staining throughout the proliferative ventricular zones and in the
cortical plate (Figure 5B), in agreement with available mouse
transcriptome data (Ayoub et al., 2011).
The dramatic reduction in brain size seen in homozygous
mutant mice reflects abnormal proliferation as well as cell death.
Following a pulse of bromodeoxyuridine (BrdU) to pregnant
dams at E12.5, a stage corresponding to early neurogenesis,
we find a 35% reduction in number of actively proliferating cells
in homozygous mutant embryos compared with wild-type,
confirming that fewer cycling progenitors are indeed present
(Figures 5C and 5D). Furthermore, we find that overall cortical
thickness is reduced by 54% at E13.5, a stage corresponding
to peak neurogenesis (Figures 5E–5H), with preserved cell polar-
ity (Figure S5A). To more specifically characterize cortical
defects, we performed immunohistochemistry for specific pro-
genitor subtypes and neurons. Sox2 immunoreactivity, which
identifies radial neuroepithelial progenitors, is reduced 32% in
Katnb1 null embryos. Tbr2 immunoreactivity, which identifies in-
termediate progenitors formed by asymmetric divisions of radial
neuroepithelial cells, is reduced more severely, with 72% deple-
tion. Neurons, labeled with DCX immunoreactivity and derived
ocation of PCR primers for (E).
e (asterisk) corresponds in size to deletion of exon 6 (dele6).
g of exon 6.
els in splice mutant allele relative to wild-type. Quantification using the LI-COR
trin), but EGFP-tagged dele6 mutant protein diffuses throughout the nucleus,
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1245

(legend on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1246 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Figure 4. Maternal Contribution of katnb1 Is Essential for Zebrafish Gastrulation
(A) Katanin p80 protein and gene structure is highly conserved between humans and zebrafish.
(B) TALENs targeting the genomic region surrounding the zebrafish katnb1 exon 6-intron 6 boundary create a series of mutant alleles. Reference sequence (top).
Blue box, TALEN recognition sites; dash, deleted base pairs; red letters, inserted base pairs.
(C) Mutant alleles are predicted to alter protein structure by deletion of amino acids or early truncation of katanin p80 protein.
(D) Progeny of mutant females show a wide spectrum of phenotypes, ranging from early embryonic lethality and anencephaly to microcephaly.
(E) Maternal effect of katnb1 is observed in crosses of mutant females, compared with progeny arising from heterozygous carriers. Embryos developed normally
until 70% epiboly.
See also Table S2 and Figure S3.
Neuron
Katanin p80 Limits Centriole and Cilia Number
from asymmetrical divisions of radial neuroepithelial cells or from
the Tbr2-positive progenitors, are the most severely affected,
with an 83% reduction compared to wild-type (Figures 5E–5H).
These results reveal a striking reduction in cycling ventricular
zone progenitors in Katnb1 null cortex, but an even more pro-
found loss of cells that depend upon asymmetrical cell divisions,
Figure 3. Katnb1 Gene-Trap Mutants Die by E14.5, with Brain Phenoty
(A) Location of gene-trap insertion. Gene-trap insertion leads to early truncation
(B and C) RT-PCR confirms absence of wild-type allele in homozygous gene-trap
exon 2 into gene-trap cassette (C).
(D and E) Katanin p80 protein is absent in gt/gt mice (D), and less katanin p60 prot
the LI-COR imaging system.
(F) Homozygous gt/gt mice are present in expectedMendelian ratios before E14.5
embryonically lethal. Chi-square test, p < 0.0001.
(G) Compared with wild-type, homozygous gt/gt embryos are dramatically reduc
p < 0.001.
(H) Homozygous gt/gt embryos vary in size, with brain phenotypes ranging from
zygous gt/gt embryos are micropthalmic (embryo 2), while others have no eye dev
pallor of the liver. All pictured embryos are littermates. Scale bar, 5 mm (top) and
(I) Coronal sections through embryonic brains show reduced cortical size, thickne
of brain. Scale bar, 200 um.
(J) Distribution of homozygous gt/gt phenotypes recovered between E12.5 and E
Ne
including intermediate progenitors and neurons. These data
parallel the results of katanin p80 loss in the Drosophila brain
presented in an accompanying paper, in this issue of Neuron
(Mishra-Gorur et al., 2014). We hypothesized that cell death
might also be a factor in neuronal depletion, and indeed we
find apoptotic cells, marked by activated cleaved caspase 3, in
pes Ranging from Microcephaly to Holoprosencephaly
of Katnb1 mRNA, to mimic a null allele.
mice (B). Sanger sequencing of gene-trap RT-PCR product confirms splicing of
ein is produced in gt/gt mice, compared with wild-type (E). Quantification using
but are almost never found after E15.5, indicating thatKatnb1 loss of function is
ed in body size at E14.5. Error bars indicate mean ± SEM. One-way ANOVA,
microcephaly (embryos 2–4) to holoprosencephaly (embryo 5). Some homo-
elopment (embryos 3–5), and all have underdevelopment of the limb buds and
1 mm (bottom).
ss, and holoprosencephaly in homozygous gt/gt embryos. Dashed line, outline
15.5.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1247
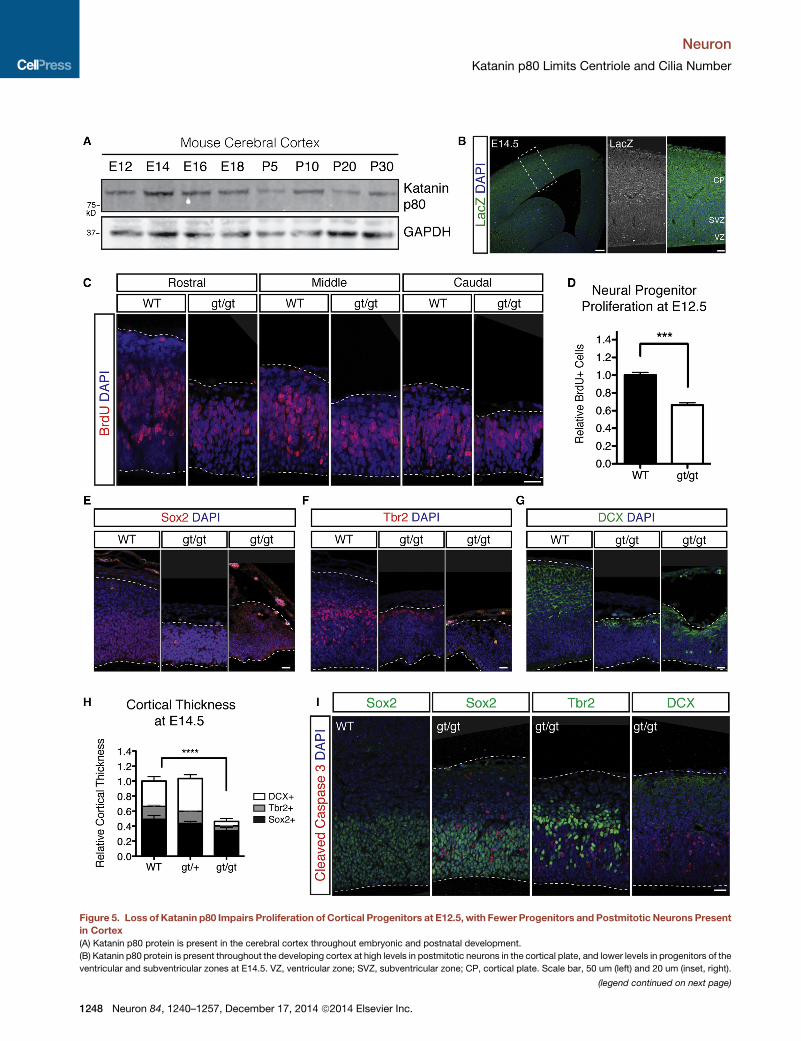
Figure 5. Loss of Katanin p80 Impairs Proliferation of Cortical Progenitors at E12.5, with Fewer Progenitors and Postmitotic Neurons Present
in Cortex
(A) Katanin p80 protein is present in the cerebral cortex throughout embryonic and postnatal development.
(B) Katanin p80 protein is present throughout the developing cortex at high levels in postmitotic neurons in the cortical plate, and lower levels in progenitors of the
ventricular and subventricular zones at E14.5. VZ, ventricular zone; SVZ, subventricular zone; CP, cortical plate. Scale bar, 50 um (left) and 20 um (inset, right).
(legend continued on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1248 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Neuron
Katanin p80 Limits Centriole and Cilia Number
homozygous mutant brains. Even in the most mildly affected
homozygous gene-trap embryos, apoptosis is abundant
throughout the brain, particularly in the proliferative ventricular
zones of the cortex, whereas apoptotic cells are absent from
wild-type brains (Figure 5I). We obtained similar results using
neural stem cells (NSCs) differentiated from human iPSCs.
Proband 1-derived neurospheres fail to grow and exhibit wide-
spread apoptosis (Figures S5B–S5D). Together, these data
indicate that katanin p80 is required for neuronal progenitor pro-
liferation and survival during cortical development.
Loss of Katanin p80 Disrupts Spindle Structure andMitosisTo better understand the cellular functions of katanin p80 during
human development, we studied proband-derived cell lines.
Katanin p80 and p60 localize to the spindle poles of mitotic cells
and to the pericentriolar matrix of interphase cells (Figures S5A–
S5C). In immortalized human lymphoblasts and human iPSCs
expressing mutant KATNB1 alleles, we find defects in centroso-
mal structure and function. Approximately 49% of S phase
Proband 2-derived lymphoblasts contain more than two centro-
somes, compared with 11% in control lymphoblasts (Figures 6A
and 6B). Instead of localizing perinuclearly, these centrosomes
cluster in the center of the cell, with the DNA arranged around
the centrosomes (Figure 6A). iPSCs derived from Proband 1 dis-
played mitotic abnormalities in 51% of cells, compared to only
4% of control iPSCs: 10% of cells displayed multipolar spindles,
with supernumerary chromosomes and excess kinetochores,
22% of cells displayed abnormal monoastral spindles, and an
additional 19% contain misaligned chromosomes (Figures 6C
and 6D). Flow cytometry analysis for DNA content on lympho-
blasts from Proband 2 revealed an increased proportion of cells
in S and G2 phase, compared with control lymphoblasts (Figures
6E and 6F), indicative of cell cycle perturbation and aneuploidy.
Katanin p80 Limits Centriole NumberWe further investigated centriole biogenesis in MEFs isolated
from gene-trap embryos. Katnb1 null MEFs are more severely
affected than proband-derived mutant cell lines, with 76% of
gene-trap MEFs containing multipolar mitotic spindles, com-
pared with 12% in wild-type MEFs (Figures 7A and 7D). We
also found binucleate homozygous gene-trapMEFs, likely result-
ing from aneuploid divisions (Shi and King, 2005), while we
almost never found binucleate wild-type MEFs (data not shown).
MEFs isolated from homozygous gene-trap embryos often failed
to proliferate (Figure S6C), and those homozygous gene-trap
MEFs that did grow well in vitro divided significantly more slowly
than wild-type MEFs (Figure 7F), consistent with the widespread
defects in proliferation observed in the mutant embryos.
(C) Fewer actively proliferating cortical progenitors are labeled by acute BrdU
sections in gt/gt cortex compared with wild-type. Scale bar, 20 um.
(D) Quantification of BrdU-positive cells per length of ventricular surface. Error b
(E–G) Homozygous gt/gt mutant cortex is reduced in thickness at E12.5 com
(F, Tbr2+) and neurons (G, DCX+) over radial glia (E, Sox2+). Scale bar, 20 um.
(H) Quantification of radial thickness of cortex, with percent composition of mark
(I) Apoptotic cells labeled by activated cleaved caspase 3 are abundant in homozy
See also Figure S4.
Ne
Defects in cytoskeletal structure in homozygous gene-trap
MEFs, with increased acetylated tubulin immunoreactivity
around the centrosome (Figure S6D), suggest increased overall
microtubule stability. Furthermore, we find more prominent
staining for CAMSAP2, a microtubule minus-end protein that
protects against depolymerization, as well as more linear,
streak-like staining for EB1, a microtubule plus-end protein (Fig-
ures S6E and S6F). Together, these data suggest that microtu-
bule dynamics are altered and that loss of katanin p80 results
in a more stabilized cytoskeleton.
Surprisingly, we found a high proportion of overduplicated
centrioles in homozygous gene-trap MEFs (Figure 7B), with
76% of cells displaying supernumerary centrioles (Figure 7E),
consistent with our multipolar spindle data. Transmission elec-
tron microscopy of centrosomes reveals that these ectopic cen-
trioles have normal 9-fold symmetry but are unpaired, unlike
wild-type, paired centrioles (Figures 7C, S6A, and S6B). These
supernumerary centrioles associate with multiple centrosomal
proteins, including g-tubulin, Stil, and Cep63 (Figure 7B), sug-
gesting they form functional centrosomes and may explain the
origin of the mitotic spindle defects we observe (Figures 7A
and 7D).
Loss of Katanin p80 Causes Supernumerary Cilia andAbrogates Sonic Hedgehog SignalingThe mother centriole is the older centriole in a cell, and normal
cells contain only one mother centriole. Examining the localiza-
tion of two mother centriolar proteins, Ninein and Cep164, re-
vealed the surprising presence of multiple mother centrioles in
Katnb1 homozygous gene-trap MEFs (Figure 8A), suggesting
defects in not just centriole number, but also centriolematuration
and identity.
Since the mother centriole nucleates the primary cilium, the
presence of multiple mother centrioles suggested that Katnb1
mutant cells may be capable of nucleating multiple cilia. Indeed,
after serum starvation to induce cilia growth, we identified many
cells with multiple Ift88- or Arl13b-positive cilia (Figure 8B). Over
35% of Katnb1 homozygous gene-trap MEFs possess supernu-
merary cilia (Figure 8D), nearly four times as many as wild-type
MEFs, confirming an excess of functional mother centrioles in
Katnb1 mutant cells. Some cells contain only a second, shorter
cilium, but other cells grow two or even three full-length cilia (Fig-
ures 8B and 8D). As with wild-type cells,Katnb1mutant MEFs do
not produce cilia without serum starvation (data not shown), sug-
gesting that the multiple cilia arise from supernumerary centri-
oles upon initiation of ciliogenesis, rather than from defective
ciliary resorption.
The overproduction of cilia in the absence of katanin p80
raised the question of whether homozygous gene-trap MEFs
injection at rostral (left), middle (middle), and caudal (right) matched coronal
ars indicate mean ± SEM. Unpaired t test, p < 0.001.
pared with wild-type, with preferential reduction in intermediate progenitors
ers in (E)–(G). Error bars indicate mean ± SEM. One-way ANOVA, p < 0.01.
gous gt/gt ventricular zones, even inmildly affected embryos. Scale bar, 20 um.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1249

Figure 6. Loss of Katanin p80 Causes Centrosome and Centriole Overduplication in Proband-Derived Cell Lines and MEFs
(A) Lymphoblasts derived from Proband 2 undergo centrosomal overduplication, with DNA surrounding centrally located centrosomes. Scale bar, 10 um.
(B) Quantification of proportion of lymphoblasts with overduplicated centrosomes. Unpaired t test, p < 0.01.
(C) Mitotic iPSCs derived from Proband 1 contain monoastral spindles, multipolar spindles, and lagging chromosomes (arrowhead). Scale bar, 10 um.
(D) Quantification of proportion of mitotic iPSCs with abnormal mitoses. Unpaired t test, p < 0.0001.
(E and F) A greater percentage of Proband 2-derived lymphoblasts are aneuploid and have skewed DNA content compared with control lymphoblasts by FACS
(E), indicating that the cell cycle is disrupted in proband-derived lymphoblasts (F).
See also Figure S5.
Neuron
Katanin p80 Limits Centriole and Cilia Number
also show alterations in ciliary signaling. The holoprosencephaly
seen in homozygous gene-trap embryos suggested a defect in
Shh signaling, which, in vertebrates, is dependent on cilia
(Huangfu and Anderson, 2005; Huangfu et al., 2003; Mahjoub
and Stearns, 2012). We triggered Shh signaling in MEFs using
Smoothened agonist (SAG) stimulation. SAG normally causes
Gli3 and Smoothened (Smo) to accumulate at the tips of primary
cilia. In contrast, Gli3 failed to accumulate in cilia in SAG-
stimulated MEFs lacking katanin p80 (Figure 8C), with a 77%
reduction in Gli3 localization to the ciliary tip in homozygous
1250 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc
gene-trap MEFs (Figure 8E). Smoothened accumulation in cilia
in SAG-stimulated MEFs is also decreased in homozygous
gene-trapMEFs, even in cells containing only a single cilium (Fig-
ure S7A). qRT-PCR analysis of downstream targets Gli1 and
Patched1, which are normally upregulated in response to Shh
signaling, showed a 5-fold increase in Gli1 induction and 11.8-
fold increase in Patched1 induction upon SAG stimulation in
wild-type MEFs, but no response in homozygous gene-trap
MEFs (Figures 8F and 8G). Together, these data demonstrate
that the loss of katanin p80 leads to overduplication of centrioles
.

Neuron
Katanin p80 Limits Centriole and Cilia Number
and excessive maternal centrioles, leading to supernumerary
cilia, and that the resulting multiple cilia fail to properly transduce
Shh signals.
DISCUSSION
Here, we identify katanin p80 as a critical regulator of human
brain development, and extend its functions beyond regulation
of microtubule severing and the mitotic spindle. Three indepen-
dent mutations in KATNB1, affecting protein levels and subcellu-
lar localization, result in severe microlissencephaly in humans,
and complete genetic ablation of Katnb1 leads to embryonic
lethality in mice and zebrafish. On a cellular level, loss of katanin
p80 causes the formation of supernumerary centrioles, including
extra mother centrioles. These centrioles result in multipolar
spindles and also nucleate supernumerary cilia, leading to
reduced Shh signaling. Together, these data implicate katanin
as a regulator of mother and daughter centriole number, and of
the ciliary number and function that follow, thus greatly expand-
ing our view of katanin function.
Despite the severe microlissencephaly seen in humans with
KATNB1 mutations, the human mutations we identified are
most likely hypomorphic. Katanin p80 is essential during early
embryogenesis in both mice and zebrafish. Humans do not
display phenotypes as extreme as those as seen in the loss-of-
functionmouse or zebrafishmutants, nor do they show evidence
of defects classic for ciliary or Shh dysfunction, such as polycy-
stic kidney disease, laterality defects, midline patterning defects
(including holoprosencephaly), or digit anomalies (Dubourg
et al., 2007; Hildebrandt et al., 2011; Novarino et al., 2011). Addi-
tionally, NSCs derived from human iPSCs do not develop excess
cilia or decreased Shh signaling seen in loss-of-function MEFs,
further supporting the hypomorphic nature of the human alleles.
This may reflect the specific mutations we have identified, all of
which disrupt the N-terminal WD40 repeats, and result in
reduced, but not completely abolished, protein levels; thus, the
mutated proteins may retain enough residual activity to rescue
the severe, complete loss-of-function phenotypes. Finally, a
previously published mouse carrying a missense mutation in
Katnb1 is viable, with defects observed only in spermatogenesis
(O’Donnell et al., 2012), suggesting that alleles of KATNB1 can
give rise to varied phenotypes of altered severity (see also
Mishra-Gorur et al., 2014).
Our results also indicate that there are distinct temporal roles
for katanin. Loss of maternal katnb1 results in severe defects of
gastrulation, whereas loss of zygotic zebrafish katnb1 is compat-
ible with survival and growth to viable, fertile adults, indicating
that katnb1 is necessary for tissue patterning during early devel-
opment, with additional roles during later development. There-
fore, the more severe phenotype seen in mice may represent
early loss of mammalian katanin p80 function, and the milder hu-
man disease phenotypic spectrummay arise from loss of katanin
p80 function later during development, possibly in a nervous-
system-specific manner.
The identification of mutations in KATNB1 suggests that
mutations in KATNA1, encoding the catalytic katanin p60 sub-
unit, might also cause microlissencephaly, given their strong
biochemical and functional interactions. In our cohort of micro-
Ne
cephalic and lissencephalic pedigrees, we have not yet identified
any individuals with KATNA1 mutations. Complete loss of
KATNA1 function may be incompatible with embryonic survival,
as is the case with Drosophila katanin p60 (Mao et al., 2014), or
mutations in KATNA1 may cause an entirely different human
phenotype. Further characterization and sequencing in individ-
uals will be necessary to more fully explore the human genetics
of katanin mutations.
The essential roles of KATNB1 in humans, mice, and zebrafish
are somewhat surprising, given our knowledge of the role of the
katanin complex from previous work, and illustrate the unique
insight hypomorphic humanmutations offers in uncovering novel
regulators of early embryonic development. Katanin is amicrotu-
bule-severing enzyme necessary for meiotic spindle assembly
(Mains et al., 1990; Srayko et al., 2006), determination of mitotic
spindle length (Loughlin et al., 2011; McNally et al., 2006),
severing at microtubule crossovers (Lindeboom et al., 2013;
Zhang et al., 2013), axon outgrowth (Ahmad et al., 1999; Karabay
et al., 2004; Yu et al., 2005), and cell motility (Zhang et al., 2011).
Furthermore, katanin has been shown to be required for
axonemal structure (Casanova et al., 2009; Dymek and Smith,
2012) and basal body release (Rasi et al., 2009). However, these
functions all relate to katanin’s microtubule-severing activity.
Our data point to roles for the katanin complex that have not pre-
viously been described or even suspected based on its microtu-
bule-severing function. The supernumerary cilia we observe are
not remnants from previous cell cycles that have not been disas-
sembled, but, rather, are formed de novo with each cell cycle.
Furthermore, our data implicate katanin p80 in functions inde-
pendent of targeting katanin p60 to the centrosome or regulating
katanin p60 catalytic activity (Hartman et al., 1998;McNally et al.,
2000).
Katanin p80, Centrioles, and CiliaWepresent evidence showing katanin’s involvement in control of
centriolar number and centriolar type, as well as in ciliogenesis
and Shh signaling, and in vivo evidence linking all three pro-
cesses. Katnb1-deficient cells show a striking excess of centri-
oles, including multiple mother centrioles. In asymmetrical
divisions of cortical progenitor cells, the mother centriole is pref-
erentially inherited by the stem cell that continues dividing,
whereas the daughter centriole is associated with the more
differentiated cell. Moreover, the mother centriole is associated
with a piece of ciliary membrane that serves to initiate a new
cilium in the cell inheriting the more mature, mother centriole,
allowing it to respond more quickly to extracellular growth sig-
nals, including Shh and IGF2 (Anderson and Stearns, 2009; Leh-
tinen et al., 2011; Paridaen et al., 2013; Piotrowska-Nitsche and
Caspary, 2012; Saade et al., 2013). Although overduplication of
centrioles has been produced by overexpression of genes such
as PLK4, SAS-6, STIL, CPAP, and CEP120 (Arquint and Nigg,
2014; Habedanck et al., 2005; Kleylein-Sohn et al., 2007; Lin
et al., 2013; Peel et al., 2007; Strnad et al., 2007; Tang et al.,
2011), or by loss of CCDC14, CEP76, Lin-23, and Sel-10 (Firat-
Karalar et al., 2014; Peel et al., 2012; Tsang et al., 2009), centriole
overduplication does not always cause excessmother centrioles
or cilia to form (Mahjoub and Stearns, 2012; Marthiens et al.,
2013), as overexpression of Plk4 in the mouse brain leads to
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1251

(legend on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1252 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Neuron
Katanin p80 Limits Centriole and Cilia Number
centrosomal overduplication and microcephaly, but without de-
fects in cilia number or Shh signaling (Marthiens et al., 2013). Our
findings of multiple mother centrioles in Katnb1mutant cells, and
multiple cilia but with defective ciliary Shh signaling, suggest that
katanin p80 may serve an essential role in the link between
centriole and cilium to cell fate in the cortical ventricular zone.
The precise mechanism by which loss of katanin p80 causes
centriole overduplication remains unexplored. Duplication of
centrioles is normally limited to once per cell cycle, and presence
of a daughter centriole inhibits centriole duplication (La Terra
et al., 2005; Loncarek et al., 2008; Wong and Stearns, 2003).
However, certain cell lines are capable of excess duplication
when artificially arrested in S phase (Balczon et al., 1995), and
overexpression of various centriolar and pericentriolar matrix
proteins causes overduplication (Loncarek et al., 2008;Mahjoub,
2013).
Centriole duplication can occur from a template mother
centriole or by de novo synthesis, which require similar factors
(Khodjakov et al., 2002; La Terra et al., 2005). In the absence
of katanin p80, both excessive templating of themother centriole
and unbridled de novo centriole synthesis are possibilities.
Immunostaining and electron microscopy show centrioles
clustered together, rather than scattered throughout the cell,
suggesting that some templating may occur. However, the
centrioles we observe are not paired in the classic orthogonal
orientation of templated, duplicating centrioles and suggest
the de novo pathway may nonetheless be active. Furthermore,
centriole duplication has been proposed to be regulated by con-
trolling the localization of duplication factors (Firat-Karalar et al.,
2014). The increased microtubule stability with loss of katanin
may prevent trafficking of positive and negative regulators of
centriole duplication to and away from the centrosome, resulting
in unbridled duplication.
There is emerging evidence in neurodevelopment for genes
linking centrosome, spindle, and cilia biology. Ciliopathy syn-
dromes are associated with rare brain malformations, though
the mechanism by which they occur and their significance is still
unclear (Lee and Gleeson, 2011; Nigg and Raff, 2009; Oh and
Katsanis, 2012). Our results suggest that defects in cilia may
cause isolated nervous system disease, potentially related to
its roles in later development. Loss of two other centrosomal
microcephaly genes,CDK5RAP2 andNDE1, are known to cause
excess centrosomes and multipolar spindles and have recently
been shown to affect cilia length and number as well (Alkuraya
et al., 2011; Bakircio�glu et al., 2011; Barrera et al., 2010; Bond
et al., 2005; Kim et al., 2011; Lizarraga et al., 2010). Mutations
in NDE1 were the first identified cause of microlissencephaly
Figure 7. Loss of Katanin p80 Causes Centriole Overduplication
(A) Homozygous gt/gt MEFs have multipolar mitotic spindles and misaligned chr
(B) Centrioles, labeled by Centrin, are overduplicated in homozygous gt/gt ME
(middle), and Cep63 (bottom) associate with supernumerary centrioles with norm
(C) Serial transmission electron micrograph sections through centrosomes sho
contain only one pair of centrioles. Arrowhead, centriole. Scale bar, 200 nm.
(D) Quantification of mitotic spindle abnormalities in (A). Unpaired t test, p < 0.00
(E) Quantification of centriolar overduplication in (B). Unpaired t test, p < 0.0001.
(F) Homozygous gt/gt MEFs grow more slowly than wild-type or gt/+ MEFs. Two
See also Figure S6.
Ne
(Alkuraya et al., 2011; Bakircio�glu et al., 2011), with a phenotype
very similar to that of KATNB1mutations, and katanin p80 phys-
ically interacts with Ndel1, a close paralog of Nde1 (Toyo-Oka
et al., 2005).Nde1 andCdk5Rap2 also encode centrosomal pro-
teins essential for neurogenesis, though loss of neither Nde1 nor
Cdk5Rap2 is as severe as Katnb1 loss in mice (Barrera et al.,
2010; Feng and Walsh, 2004; Lizarraga et al., 2010). An
appealing model, therefore, is that CDK5RAP2, NDE1, and
KATNB1 may control the pathway limiting centriole duplication,
and that the presence of excessive centrioles, particularly
mother centrioles, in dividing neuroepithelial cells results in per-
turbed ciliary growth factor signaling, defective proliferation, and
excessive cell death.
EXPERIMENTAL PROCEDURES
Human Subjects
A combination of homozygosity mapping, whole-exome sequencing, and tar-
geted sequencing of coding exons within areas of homozygosity was per-
formedandfilteredasdescribed in theSupplementalExperimentalProcedures.
All studies were reviewed and approved by the Institutional Review Board of
Boston Children’s Hospital, Harvard Medical School, and local institutions.
Animals
Targeted gene-trap ESCs (EUCOMM) were injected into C57BL/6 blastocysts
to generate chimeric mice. Male chimeras were bred to WT C57BL/6 females
(Charles River) to transmit the gene-trap allele. The mice were maintained on a
C57/BL6 background. For pulse-labeling experiments, E12.5 timed pregnant
mice were injected with 75 mg/kg BrdU intraperitoneally and euthanized
30 min later.
TALENs targeting the exon 6-intron 6 boundary of zebrafish katnb1
(ENSDARG00000005456) were obtained from ZGENEBIO Biotech Co. Ltd
(Table S2). To generate katnb1 mutants, 150–200 pg of RNA was injected
into one-cell-stage eggs (albino/slc45a2).
Culture Systems
Epstein-Barr virus-immortalized lymphoblastoid cell lines established from
Proband 2 were maintained in RPMI-1640 (GIBCO) with 10% fetal bovine
serum (FBS, GIBCO), and 1 mM penicillin and streptomycin (P/S). Primary
fibroblasts were established from Proband 3 and were maintained in RPMI-
1640 (GIBCO) with 10% FBS, and 1 mM P/S. Human iPSCs (Figure S2A)
were generated using retroviral vectors encoding human KLF4, SOX2,
OCT4, and C-MYC cDNA (Addgene) and grown in knockout DMEM with
20% knockout serum replacement, 2 mM L-glutamine, 1% NEAA, 0.1 mM
b-mercaptoethanol, 0.1% P/S, and 4 ng/ml bFGF on irradiated fibroblast
feeders. iPSCs were dissociated to form embryoid bodies (EBs) and induced
in neural induction medium (DMEM/F12 supplemented with 20%KOSR, 2mM
L-glutamine, 0.2 mM NEAA, 0.1 mM 2-mercaptoethanol, and 1 mM sodium
pyruvate) for 7 days. EBs were then adhered onto laminin-coated plates in
neural precursor medium (neurobasal with 2 mM L-glutamine, 1% B27, 1%
N2, and 20 ng/ml bFGF) for 7–14 days to form rosettes, which were manually
isolated and expanded as NSCs in neural induction medium.
omosomes (arrowhead). PHH3, phosphorylated histone H3. Scale bar, 10 um.
Fs compared with wild-type MEFs. Centrosomal proteins g-tubulin (top), Stil
al stoichiometry. Scale bar, 10 um.
w multiple unpaired centrioles in gene-trap MEFs, whereas wild-type MEFs
01.
-way ANOVA, p < 0.001.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1253
(legend on next page)
Neuron
Katanin p80 Limits Centriole and Cilia Number
1254 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc.

Neuron
Katanin p80 Limits Centriole and Cilia Number
Trypsin-dissociated MEFs isolated from E13.5 mouse embryos were grown
in DMEM, High Glucose (HyClone) with 15% FBS, and 1 mM P/S and L-gluta-
mine. Cilia were induced by serum starvation for 48 hr in Optimem (GIBCO)
media, then treated with DMSO or 100 nM SAG for an additional 24 hr.
Additional details are available in Supplemental Experimental Procedures.
Quantitative RT-PCR
See Supplemental Experimental Procedures.
Western Blotting
See Supplemental Experimental Procedures.
Immunostaining
See Supplemental Experimental Procedures.
SUPPLEMENTAL INFORMATION
Supplemental Information includes seven figures, two tables, one movie, and
Supplemental Experimental Procedures and can be found with this article at
http://dx.doi.org/10.1016/j.neuron.2014.12.017.
AUTHOR CONTRIBUTIONS
W.F.H. designed, performed, and analyzed experiments to characterize the
mutations in Families 2 and 3; generated the gene-trap mouse line and zebra-
fish mutant lines; designed, performed, and analyzed all mouse and zebrafish
experiments; helped analyze the centriolar and ciliary phenotypes; designed,
performed, and analyzed EM experiments, and wrote the manuscript. O.P. de-
signed, performed, and analyzed experiments relating to Family 1. T.B.-O.
identified Family 3. A.K. and J.F.R. designed and performed immunostaining
experiments in mouse cell lines, and identified and analyzed centriolar and
ciliary phenotypes. K.H. generated the zebrafish mutant lines and designed,
performed, and analyzed all zebrafish experiments. G.H.M. designed and per-
formed initial genome-wide linkage studies, and G.H.M. and T.W.Y. designed
and performed targeted capture sequencing on Family 3. M.B.W. performed
and analyzed mouse BrdU labeling experiments and helped analyze centriolar
duplication phenotypes in Family 2 andmouse cell lines. C.B. analyzed the IBD
mapping and exome sequencing of Family 1. G.S.R. and T.T.T. reprogrammed
fibroblasts into iPSCs and performed immunostaining for Family 1. H.H. iden-
tified and A.M. and M.S. provided clinical information for Family 1. M.A.S. and
J.N.P. organized clinical information and subject samples for Families 2 and 3.
M.A-.D., A.A., M.A., and F.S.A. identified, provided clinical information for, and
performed genetic analyses for Family 2. M.P.H. directed the zebrafish
research. B.R. directed the research for Family 1. C.A.W. directed the overall
research and wrote the manuscript.
ACKNOWLEDGMENTS
We thank the individuals and their families for their participation. We thank
Boston Children’s Hospital (BCH) Transgenic Core Laboratory, Dana-Farber/
Harvard Cancer Center Rodent Histopathology Core, Dana-Farber Cancer
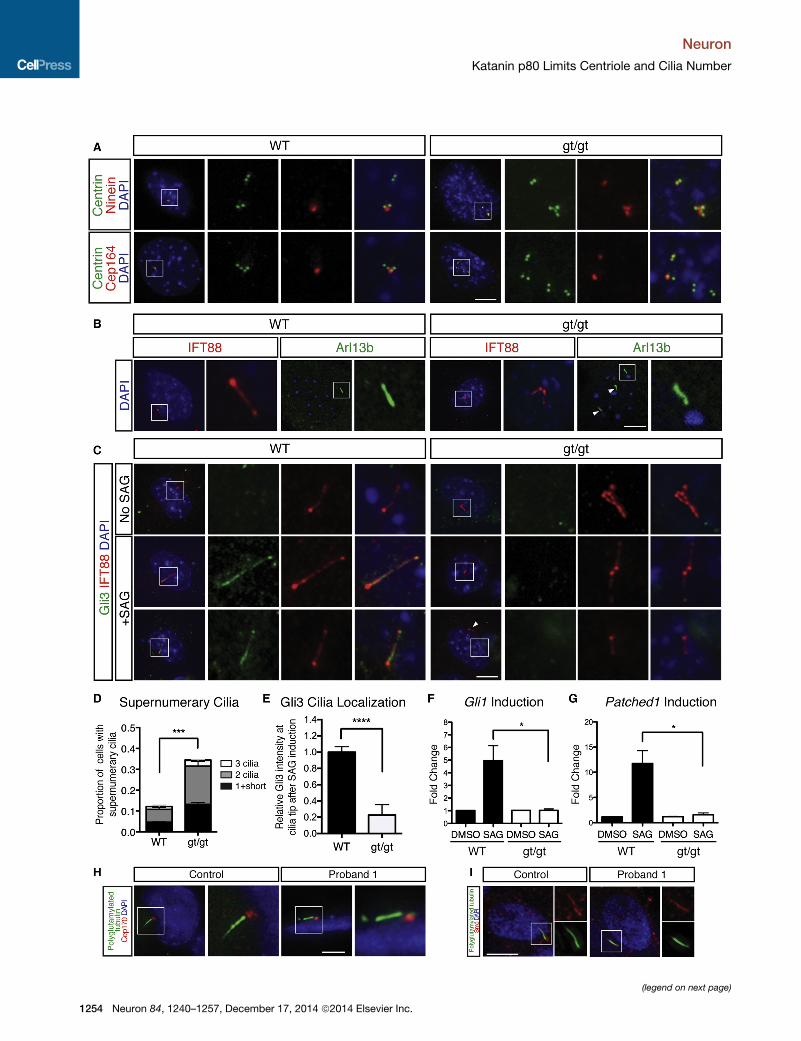
Figure 8. In the Absence of Katanin p80, Fibroblasts Produce Supernu
(A) Homozygous gt/gt MEFs contain multiple mother centrioles that label with N
(B) Homozygous gt/gt MEFs grow multiple cilia marked by IFT88 and Arl13b (rig
numerary cilia not shown in inset. Scale bar, 10 um.
(C) After stimulation by Smoothened agonist (SAG), gt/gt MEFs fail to relocate Gli3
MEFs robustly relocate Gli3 to the cilium after SAG stimulation (left). Scale bar, 1
(E) Quantification of proportion of ciliated MEFs with supernumerary cilia from (B
(E) Quantification of cilial tip Gli3 intensity in SAG-stimulated cells from (C). Unpa
(F and G) DownstreamSonic hedgehog targetsGli1 (F) and Patched1 (G) are induc
type MEFs. Error bars represent mean ± SEM. Unpaired t test, p < 0.05.
(H) NSCs derived from Proband 1 do not grow supernumerary cilia. Scale bar, 1
(I) After stimulation by Smoothened agonist (SAG), control and mutant NSCs sho
See also Figure S7.
Ne
Institute Neoplasia Flow Cytometry Core, Beth Israel Deaconess Medical
Center (BIDMC) Histology Core Laboratory, Harvard Medical School
Electron Microscopy Facility, and Genome Institute of Singapore for use of fa-
cilities. We thank Tamara Caspary, Erich Nigg, Suzie Scales, and James Silli-
bourne for kind gifts of reagents. This research was supported by NIH NINDS
F31NS083111 and NIGMS T32GM007753 (W.F.H.); NIH NHLBI T32HL007731
(A.K.); Manton Center for Orphan Disease Research and F. Hoffman-La Roche
Ltd. (G.H.M.); Clinical Investigator Training Program at Harvard-MIT Health
Sciences and Technology and BIDMC in collaboration with Pfizer, Inc. and
Merck & Company, Inc, and the Nancy Lurie Marks Junior Faculty MeRIT
Fellowship (T.W.Y.); Leonard and Isabelle Goldenson Research Fellowship
(M.B.W.); KACST grant 09-MED941-20 (F.S.A.); NIH NIAMS R01AR054396,
NIGMS R01GM095941, Burroughs Wellcome Fund, Packard Foundation,
and Sandler Family Supporting Foundation (J.F.R.); Strategic Positioning
Fund for Genetic Orphan Diseases, A*STAR Investigatorship from the Agency
for Science, Technology and Research in Singapore, Branco Weiss Founda-
tion fellow, and A*STAR and EMBO Young Investigator (B.R.); NIH NIDCR
1U01DE024434-01 and BCH Orthopedic Surgical Foundation (K.H. and
M.P.H.); and NIH NINDS R01NS035129 and NIMH RC2MH089952, the Qatar
National Research Fund National Priorities Research Program, and the Man-
ton Center for Orphan Disease Research (C.A.W.). C.A.W. is an Investigator
of the Howard Hughes Medical Institute.
Accepted: December 3, 2014
Published: December 17, 2014
REFERENCES
Ahmad, F.J., Yu, W., McNally, F.J., and Baas, P.W. (1999). An essential role for
katanin in severing microtubules in the neuron. J. Cell Biol. 145, 305–315.
Alkuraya, F.S., Cai, X., Emery, C., Mochida, G.H., Al-Dosari, M.S., Felie, J.M.,
Hill, R.S., Barry, B.J., Partlow, J.N., Gascon, G.G., et al. (2011). Human muta-
tions in NDE1 cause extrememicrocephaly with lissencephaly [corrected]. Am.
J. Hum. Genet. 88, 536–547.
Anderson, C.T., and Stearns, T. (2009). Centriole age underlies asynchronous
primary cilium growth in mammalian cells. Curr. Biol. 19, 1498–1502.
Arquint, C., and Nigg, E.A. (2014). STIL microcephaly mutations interfere with
APC/C-mediated degradation and cause centriole amplification. Curr. Biol. 24,
351–360.
Ayoub, A.E., Oh, S., Xie, Y., Leng, J., Cotney, J., Dominguez, M.H., Noonan,
J.P., and Rakic, P. (2011). Transcriptional programs in transient embryonic
zones of the cerebral cortex defined by high-resolution mRNA sequencing.
Proc. Natl. Acad. Sci. USA 108, 14950–14955.
Bakircio�glu, M., Carvalho, O.P., Khurshid, M., Cox, J.J., Tuysuz, B., Barak, T.,
Yilmaz, S., Caglayan, O., Dincer, A., Nicholas, A.K., et al. (2011). The essential
role of centrosomal NDE1 in human cerebral cortex neurogenesis. Am. J. Hum.
Genet. 88, 523–535.
Balczon, R., Bao, L., Zimmer, W.E., Brown, K., Zinkowski, R.P., and Brinkley,
B.R. (1995). Dissociation of centrosome replication events from cycles of DNA
merary Cilia with Abrogated Shh Signaling
inein (top) and Cep164 (bottom). Scale bar, 10 um.
ht), while wild-type MEFs grow only a single cilium (left). Arrowheads, super-
to the cilium, indicating a deficit in Sonic hedgehog signaling (right). Wild-type
0 um.
). Error bars represent mean ± SEM. Unpaired t test, p < 0.01.
ired t test, p < 0.0001.
ed at lower levels following SAG stimulation in gt/gt MEFs compared with wild-
0 um.
w similar Smoothened (Smo) localization to the cilium. Scale bar, 10 um.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1255

Neuron
Katanin p80 Limits Centriole and Cilia Number
synthesis and mitotic division in hydroxyurea-arrested Chinese hamster ovary
cells. J. Cell Biol. 130, 105–115.
Baron, M.H. (2013). Concise Review: early embryonic erythropoiesis: not so
primitive after all. Stem Cells 31, 849–856.
Barrera, J.A., Kao, L.-R., Hammer, R.E., Seemann, J., Fuchs, J.L., and
Megraw, T.L. (2010). CDK5RAP2 regulates centriole engagement and cohe-
sion in mice. Dev. Cell 18, 913–926.
Belloni, E., Muenke, M., Roessler, E., Traverso, G., Siegel-Bartelt, J., Frumkin,
A., Mitchell, H.F., Donis-Keller, H., Helms, C., Hing, A.V., et al. (1996).
Identification of Sonic hedgehog as a candidate gene responsible for holopro-
sencephaly. Nat. Genet. 14, 353–356.
Blanchette, M., Kent, W.J., Riemer, C., Elnitski, L., Smit, A.F.A., Roskin, K.M.,
Baertsch, R., Rosenbloom, K., Clawson, H., Green, E.D., et al. (2004). Aligning
multiple genomic sequences with the threaded blockset aligner. Genome Res.
14, 708–715.
Bond, J., Roberts, E., Springell, K., Lizarraga, S.B., Scott, S., Higgins, J.,
Hampshire, D.J., Morrison, E.E., Leal, G.F., Silva, E.O., et al. (2005). A centro-
somal mechanism involving CDK5RAP2 and CENPJ controls brain size. Nat.
Genet. 37, 353–355.
Casanova, M., Crobu, L., Blaineau, C., Bourgeois, N., Bastien, P., and Pages,
M. (2009). Microtubule-severing proteins are involved in flagellar length control
and mitosis in Trypanosomatids. Mol. Microbiol. 71, 1353–1370.
Chiang, C., Litingtung, Y., Lee, E., Young, K.E., Corden, J.L., Westphal, H., and
Beachy, P.A. (1996). Cyclopia and defective axial patterning in mice lacking
Sonic hedgehog gene function. Nature 383, 407–413.
Cooper, G.M., Stone, E.A., Asimenos, G., Green, E.D., Batzoglou, S., and
Sidow, A.; NISC Comparative Sequencing Program (2005). Distribution and in-
tensity of constraint in mammalian genomic sequence. Genome Res. 15,
901–913.
Cridland, S.O., Keys, J.R., Papathanasiou, P., and Perkins, A.C. (2009). Indian
hedgehog supports definitive erythropoiesis. Blood Cells Mol. Dis. 43,
149–155.
Davydov, E.V., Goode, D.L., Sirota, M., Cooper, G.M., Sidow, A., and
Batzoglou, S. (2010). Identifying a high fraction of the human genome to be
under selective constraint using GERP++. PLoS Comput. Biol. 6, e1001025.
Dubourg, C., Bendavid, C., Pasquier, L., Henry, C., Odent, S., and David, V.
(2007). Holoprosencephaly. Orphanet J. Rare Dis. 2, 8.
Dymek, E.E., and Smith, E.F. (2012). PF19 encodes the catalytic subunit of
katanin, p60, and is required for assembly of the flagellar central apparatus
in Chlamydomonas. J. Cell Sci. 125, 3357–3366.
Feng, Y., and Walsh, C.A. (2004). Mitotic spindle regulation by Nde1 controls
cerebral cortical size. Neuron 44, 279–293.
Firat-Karalar, E.N., Rauniyar, N., Yates, J.R., 3rd, and Stearns, T. (2014).
Proximity interactions among centrosome components identify regulators of
centriole duplication. Curr. Biol. 24, 664–670.
Greig, L.C., Woodworth, M.B., Galazo, M.J., Padmanabhan, H., and Macklis,
J.D. (2013). Molecular logic of neocortical projection neuron specification,
development and diversity. Nat. Rev. Neurosci. 14, 755–769.
Habedanck, R., Stierhof, Y.-D., Wilkinson, C.J., and Nigg, E.A. (2005). The Polo
kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 7, 1140–1146.
Hartman, J.J., Mahr, J., McNally, K., Okawa, K., Iwamatsu, A., Thomas, S.,
Cheesman, S., Heuser, J., Vale, R.D., and McNally, F.J. (1998). Katanin, a
microtubule-severing protein, is a novel AAAATPase that targets to the centro-
some using a WD40-containing subunit. Cell 93, 277–287.
Hayhurst, M., and McConnell, S.K. (2003). Mouse models of holoprosence-
phaly. Curr. Opin. Neurol. 16, 135–141.
Higginbotham, H., Guo, J., Yokota, Y., Umberger, N.L., Su, C.-Y., Li, J., Verma,
N., Hirt, J., Ghukasyan, V., Caspary, T., and Anton, E.S. (2013). Arl13b-regu-
lated cilia activities are essential for polarized radial glial scaffold formation.
Nat. Neurosci. 16, 1000–1007.
Hildebrandt, F., Benzing, T., and Katsanis, N. (2011). Ciliopathies. N. Engl. J.
Med. 364, 1533–1543.
1256 Neuron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc
Hu, W.F., Chahrour, M.H., and Walsh, C.A. (2014). The diverse genetic land-
scape of neurodevelopmental disorders. Annu. Rev. Genomics Hum. Genet.
15, 195–213.
Huangfu, D., and Anderson, K.V. (2005). Cilia and Hedgehog responsiveness
in the mouse. Proc. Natl. Acad. Sci. USA 102, 11325–11330.
Huangfu, D., Liu, A., Rakeman, A.S., Murcia, N.S., Niswander, L., and
Anderson, K.V. (2003). Hedgehog signalling in themouse requires intraflagellar
transport proteins. Nature 426, 83–87.
Karabay, A., Yu, W., Solowska, J.M., Baird, D.H., and Baas, P.W. (2004).
Axonal growth is sensitive to the levels of katanin, a protein that severs micro-
tubules. J. Neurosci. 24, 5778–5788.
Karolchik, D., Barber, G.P., Casper, J., Clawson, H., Cline, M.S., Diekhans, M.,
Dreszer, T.R., Fujita, P.A., Guruvadoo, L., Haeussler, M., et al. (2014). The
UCSC Genome Browser database: 2014 update. Nucleic Acids Res. 42
(Database issue), D764–D770.
Khodjakov, A., Rieder, C.L., Sluder, G., Cassels, G., Sibon, O., and Wang,
C.-L. (2002). De novo formation of centrosomes in vertebrate cells arrested
during S phase. J. Cell Biol. 158, 1171–1181.
Kim, S., Zaghloul, N.A., Bubenshchikova, E., Oh, E.C., Rankin, S., Katsanis, N.,
Obara, T., and Tsiokas, L. (2011). Nde1-mediated inhibition of ciliogenesis
affects cell cycle re-entry. Nat. Cell Biol. 13, 351–360.
Kleylein-Sohn, J., Westendorf, J., Le Clech, M., Habedanck, R., Stierhof,
Y.-D., and Nigg, E.A. (2007). Plk4-induced centriole biogenesis in human cells.
Dev. Cell 13, 190–202.
La Terra, S., English, C.N., Hergert, P., McEwen, B.F., Sluder, G., and
Khodjakov, A. (2005). The de novo centriole assembly pathway in HeLa cells:
cell cycle progression and centriole assembly/maturation. J. Cell Biol. 168,
713–722.
Lee, J.E., and Gleeson, J.G. (2011). Cilia in the nervous system: linking cilia
function and neurodevelopmental disorders. Curr. Opin. Neurol. 24, 98–105.
Lehtinen, M.K., Zappaterra, M.W., Chen, X., Yang, Y.J., Hill, A.D., Lun, M.,
Maynard, T., Gonzalez, D., Kim, S., Ye, P., et al. (2011). The cerebrospinal fluid
provides a proliferative niche for neural progenitor cells. Neuron 69, 893–905.
Lin, Y.-N., Wu, C.-T., Lin, Y.-C., Hsu, W.-B., Tang, C.-J.C., Chang, C.-W., and
Tang, T.K. (2013). CEP120 interacts with CPAP and positively regulates
centriole elongation. J. Cell Biol. 202, 211–219.
Lindeboom, J.J., Nakamura, M., Hibbel, A., Shundyak, K., Gutierrez, R.,
Ketelaar, T., Emons, A.M.C., Mulder, B.M., Kirik, V., and Ehrhardt, D.W.
(2013). A mechanism for reorientation of cortical microtubule arrays driven
by microtubule severing. Science 342, 1245533.
Litingtung, Y., Dahn, R.D., Li, Y., Fallon, J.F., and Chiang, C. (2002). Shh and
Gli3 are dispensable for limb skeleton formation but regulate digit number
and identity. Nature 418, 979–983.
Lizarraga, S.B., Margossian, S.P., Harris, M.H., Campagna, D.R., Han, A.-P.,
Blevins, S., Mudbhary, R., Barker, J.E., Walsh, C.A., and Fleming, M.D.
(2010). Cdk5rap2 regulates centrosome function and chromosome segrega-
tion in neuronal progenitors. Development 137, 1907–1917.
Loncarek, J., Hergert, P., Magidson, V., and Khodjakov, A. (2008). Control of
daughter centriole formation by the pericentriolar material. Nat. Cell Biol. 10,
322–328.
Loughlin, R., Wilbur, J.D., McNally, F.J., Nedelec, F.J., and Heald, R. (2011).
Katanin contributes to interspecies spindle length scaling in Xenopus. Cell
147, 1397–1407.
Lui, J.H., Hansen, D.V., and Kriegstein, A.R. (2011). Development and evolu-
tion of the human neocortex. Cell 146, 18–36.
Mahjoub, M.R. (2013). The importance of a single primary cilium.
Organogenesis 9, 61–69.
Mahjoub, M.R., and Stearns, T. (2012). Supernumerary centrosomes nucleate
extra cilia and compromise primary cilium signaling. Curr. Biol. 22, 1628–1634.
Mahmood, S., Ahmad, W., and Hassan, M.J. (2011). Autosomal Recessive
Primary Microcephaly (MCPH): clinical manifestations, genetic heterogeneity
and mutation continuum. Orphanet J. Rare Dis. 6, 39.
.

Neuron
Katanin p80 Limits Centriole and Cilia Number
Mains, P.E., Kemphues, K.J., Sprunger, S.A., Sulston, I.A., and Wood, W.B.
(1990). Mutations affecting the meiotic and mitotic divisions of the early
Caenorhabditis elegans embryo. Genetics 126, 593–605.
Mao, C.-X., Xiong, Y., Xiong, Z., Wang, Q., Zhang, Y.Q., and Jin, S. (2014).
Microtubule-severing protein Katanin regulates neuromuscular junction devel-
opment and dendritic elaboration in Drosophila. Development 141, 1064–
1074.
Marthiens, V., Rujano, M.A., Pennetier, C., Tessier, S., Paul-Gilloteaux, P., and
Basto, R. (2013). Centrosome amplification causes microcephaly. Nat. Cell
Biol. 15, 731–740.
McNally, F.J., and Vale, R.D. (1993). Identification of katanin, an ATPase that
severs and disassembles stable microtubules. Cell 75, 419–429.
McNally, K.P., Bazirgan, O.A., and McNally, F.J. (2000). Two domains of p80
katanin regulate microtubule severing and spindle pole targeting by p60 kata-
nin. J. Cell Sci. 113, 1623–1633.
McNally, K., Audhya, A., Oegema, K., and McNally, F.J. (2006). Katanin con-
trols mitotic and meiotic spindle length. J. Cell Biol. 175, 881–891.
Mishra-Gorur, K., Ca�glayan, A.O., Schaffer, A.E., Chabu, C., Henegariu, O.,
Vonhoff, F., Akgumusx, G.T., Nishimura, S., Han, W., Tu, S., et al. (2014).
Mutations in KATNB1 cause complex cerebral malformations by disrupting
asymmetrically dividing neural progenitors. Neuron 84, this issue, 1226–1239.
Mount, S.M. (1982). A catalogue of splice junction sequences. Nucleic Acids
Res. 10, 459–472.
Neer, E.J., Schmidt, C.J., Nambudripad, R., and Smith, T.F. (1994). The
ancient regulatory-protein family of WD-repeat proteins. Nature 371, 297–300.
Nigg, E.A., and Raff, J.W. (2009). Centrioles, centrosomes, and cilia in health
and disease. Cell 139, 663–678.
Novarino, G., Akizu, N., and Gleeson, J.G. (2011). Modeling human disease in
humans: the ciliopathies. Cell 147, 70–79.
O’Donnell, L., Rhodes, D., Smith, S.J., Merriner, D.J., Clark, B.J., Borg, C.,
Whittle, B., O’Connor, A.E., Smith, L.B., McNally, F.J., et al. (2012). An essen-
tial role for katanin p80 and microtubule severing in male gamete production.
PLoS Genet. 8, e1002698.
Oh, E.C., and Katsanis, N. (2012). Cilia in vertebrate development and disease.
Development 139, 443–448.
Paridaen, J.T.M.L., Wilsch-Brauninger, M., and Huttner, W.B. (2013).
Asymmetric inheritance of centrosome-associated primary cilium membrane
directs ciliogenesis after cell division. Cell 155, 333–344.
Peel, N., Stevens, N.R., Basto, R., and Raff, J.W. (2007). Overexpressing
centriole-replication proteins in vivo induces centriole overduplication and
de novo formation. Curr. Biol. 17, 834–843.
Peel, N., Dougherty, M., Goeres, J., Liu, Y., and O’Connell, K.F. (2012). The C.
elegans F-box proteins LIN-23 and SEL-10 antagonize centrosome duplica-
tion by regulating ZYG-1 levels. J. Cell Sci. 125, 3535–3544.
Piotrowska-Nitsche, K., and Caspary, T. (2012). Live imaging of individual cell
divisions in mouse neuroepithelium shows asymmetry in cilium formation and
Sonic hedgehog response. Cilia 1, 6.
Rasi, M.Q., Parker, J.D.K., Feldman, J.L., Marshall, W.F., and Quarmby, L.M.
(2009). Katanin knockdown supports a role for microtubule severing in release
of basal bodies before mitosis in Chlamydomonas. Mol. Biol. Cell 20, 379–388.
Roessler, E., Belloni, E., Gaudenz, K., Jay, P., Berta, P., Scherer, S.W., Tsui,
L.C., and Muenke, M. (1996). Mutations in the human Sonic Hedgehog gene
cause holoprosencephaly. Nat. Genet. 14, 357–360.
Ne
Saade, M., Gutierrez-Vallejo, I., Le Dreau, G., Rabadan, M.A., Miguez, D.G.,
Buceta, J., and Martı, E. (2013). Sonic hedgehog signaling switches the
mode of division in the developing nervous system. Cell Rep. 4, 492–503.
Senapathy, P., Shapiro, M.B., and Harris, N.L. (1990). Splice junctions, branch
point sites, and exons: sequence statistics, identification, and applications to
genome project. Methods Enzymol. 183, 252–278.
Shi, Q., and King, R.W. (2005). Chromosome nondisjunction yields tetraploid
rather than aneuploid cells in human cell lines. Nature 437, 1038–1042.
Srayko, M., O’toole, E.T., Hyman, A.A., and Muller-Reichert, T. (2006). Katanin
disrupts the microtubule lattice and increases polymer number in C. elegans
meiosis. Curr. Biol. 16, 1944–1949.
Strnad, P., Leidel, S., Vinogradova, T., Euteneuer, U., Khodjakov, A., and
Gonczy, P. (2007). Regulated HsSAS-6 levels ensure formation of a single pro-
centriole per centriole during the centrosome duplication cycle. Dev. Cell 13,
203–213.
Tang, C.-J.C., Lin, S.-Y., Hsu, W.-B., Lin, Y.-N., Wu, C.-T., Lin, Y.-C., Chang,
C.-W., Wu, K.-S., and Tang, T.K. (2011). The humanmicrocephaly protein STIL
interacts with CPAP and is required for procentriole formation. EMBO J. 30,
4790–4804.
Thornton, G.K., and Woods, C.G. (2009). Primary microcephaly: do all roads
lead to Rome? Trends Genet. 25, 501–510.
Tong, C.K., Han, Y.-G., Shah, J.K., Obernier, K., Guinto, C.D., and Alvarez-
Buylla, A. (2014). Primary cilia are required in a unique subpopulation of neural
progenitors. Proc. Natl. Acad. Sci. USA 111, 12438–12443.
Toyo-Oka, K., Sasaki, S., Yano, Y., Mori, D., Kobayashi, T., Toyoshima, Y.Y.,
Tokuoka, S.M., Ishii, S., Shimizu, T., Muramatsu, M., et al. (2005).
Recruitment of katanin p60 by phosphorylated NDEL1, an LIS1 interacting
protein, is essential for mitotic cell division and neuronal migration. Hum.
Mol. Genet. 14, 3113–3128.
Tsang, W.Y., Spektor, A., Vijayakumar, S., Bista, B.R., Li, J., Sanchez, I.,
Duensing, S., and Dynlacht, B.D. (2009). Cep76, a centrosomal protein that
specifically restrains centriole reduplication. Dev. Cell 16, 649–660.
Wang, X., Tsai, J.-W., Imai, J.H., Lian, W.-N., Vallee, R.B., and Shi, S.-H.
(2009). Asymmetric centrosome inheritance maintains neural progenitors in
the neocortex. Nature 461, 947–955.
Willaredt, M.A., Hasenpusch-Theil, K., Gardner, H.A.R., Kitanovic, I.,
Hirschfeld-Warneken, V.C., Gojak, C.P., Gorgas, K., Bradford, C.L., Spatz,
J., Wolfl, S., et al. (2008). A crucial role for primary cilia in cortical morphogen-
esis. J. Neurosci. 28, 12887–12900.
Wong, C., and Stearns, T. (2003). Centrosome number is controlled by a
centrosome-intrinsic block to reduplication. Nat. Cell Biol. 5, 539–544.
Yu, W., Solowska, J.M., Qiang, L., Karabay, A., Baird, D., and Baas, P.W.
(2005). Regulation of microtubule severing by katanin subunits during neuronal
development. J. Neurosci. 25, 5573–5583.
Zhang, Z., Li, J., Zhao, X.-Q., Wang, J., Wong, G.K.-S., and Yu, J. (2006).
KaKs_Calculator: calculating Ka and Ks through model selection and model
averaging. Genomics Proteomics Bioinformatics 4, 259–263.
Zhang, D., Grode, K.D., Stewman, S.F., Diaz-Valencia, J.D., Liebling, E., Rath,
U., Riera, T., Currie, J.D., Buster, D.W., Asenjo, A.B., et al. (2011). Drosophila
katanin is a microtubule depolymerase that regulates cortical-microtubule
plus-end interactions and cell migration. Nat. Cell Biol. 13, 361–370.
Zhang, Q., Fishel, E., Bertroche, T., and Dixit, R. (2013). Microtubule severing
at crossover sites by katanin generates ordered cortical microtubule arrays in
Arabidopsis. Curr. Biol. 23, 2191–2195.
uron 84, 1240–1257, December 17, 2014 ª2014 Elsevier Inc. 1257

Neuron, Volume 84
Supplemental Information
Katanin p80 Regulates Human Cortical Development by Limiting Centriole and Cilia Number Wen F. Hu, Oz Pomp, Tawfeg Ben-Omran, Andrew Kodani, Katrin Henke, Ganeshwaran H. Mochida, Timothy W. Yu, Mollie B. Woodworth, Carine Bonnard, Grace Selva Raj, Thong Teck Tan, Hanan Hamamy, Amira Masri, Mohammad Shboul, Muna Al Saffar, Jennifer N. Partlow, Mohammed Al-Dosari, Anas Alazami, Mohammed Alowain, Fowzan S. Alkuraya, Jeremy F. Reiter, Matthew P. Harris, Bruno Reversade, and Christopher A. Walsh

Supplemental Figures and Figure Legends
Supplemental Figure S1, related to Figure 1. KATNB1 affected individuals have
small head circumference relative to height and weight.
(A) Affected KATNB1 individuals show sloping foreheads indicative of reduced cranial
size, and facial dysmorphisms.
(B-D) KATNB1 affected individuals display head circumference well below 0.1st
percentile for ages between birth and five years, disproportionate to height and weight.
Proband data fitted with logarithmic regression line. Proband 1, purple; Proband 1
cousin (female), green; Proband 1 cousin (male), dark blue; Proband 2, red; Proband 3,
orange. Standard growth curves from World Health Organization data(WHO Multicentre
Growth Reference Study Group, 2006)
(E) Homozygosity mapping in Family 1 identifies a single shared identical-by-descent
(IBD) locus on Chromosome 16.
(F-F’’) Sanger sequencing traces showing homozygous mutations in probands and
heterozygous changes in the unaffected parents and siblings.


3
Supplemental Movie S1, related to Figure 1. MRI scans of Proband 2. Full sagittal,
axial and coronal inversion recovery MRI sequences of Proband 2 show reduced brain
size, simplification of gyral folding pattern, enlarged lateral ventricles posteriorly and
thinning of the corpus callosum, with sparing of the cerebellum, basal ganglia, thalamus
and brainstem.

5
Supplemental Figure S2, related to Figure 2. Characterization of iPSC colonies
and Family 1 mutation.
(A) Characterization of representative human iPSC colonies derived from Proband
1. Colonies with ES-like morphology were manually picked 4-5 weeks post-infection.
Several representative clones were characterized. These clones expressed pluripotency
markers, including surface antigens TRA1-60, TRA-1-81 and SSEA4, as well as the
nuclear transcription factors OCT4 and SOX2.
(B) Initiator codon mutation in Family 1 is predicted to result in N-terminal truncation of
56 amino acids. Asterisks, conserved residues.


7
Supplemental Figure S3, related to Figure 4. Sequences and characterization of
zebrafish katnb1 mutant alleles.
(A) Multiple sequence alignment of the human, mouse and zebrafish Katnb1, showing a
high level of conservation of the protein sequence between the species.
(B) Summary of identified TALEN-induced mutations in zebrafish katnb1. Genomic
region surrounding the exon 6 - intron 6 boundary in zebrafish is highlighted. TALEN
recognition sites are marked with blue boxes; exonic sequence (capital letters); intronic
sequence (lowercase). Wild-type sequence is listed above and the sequences of
identified alleles are detailed below; deleted base pairs (red dashes); inserted base
pairs (red letters). The effect on the genomic sequence is summarized to the right of
each sequence: - deletion; + insertion; presence (+) or absence (Δ) of the splice site;
allele names; frequency of the allele in 288 fish analyzed.
(C) Progeny from homozygous female carriers of katnb1 mutations develop normally
through epiboly. Summary of phenotypes appearing at 24 hpf ranging from mild
microcephaly, anencephaly and disrupted embryos.


9
Supplemental Figure S4, related to Figure 5. Katanin p80 in neural progenitor
cells.
(A) Apical complex marker, beta-catenin, localizes to the ventricular surface in both
wild-type and gene-trap developing mouse cortex, indicating intact cellular polarity.
Scale bar, 10 um.
(B) Neurospheres derived from Proband 1 iPSCs fail to grow. Scale bar, 100 um.
(C) Growth curves for neurospheres derived from Proband 1 and control. Two-way
ANOVA, p<0.0001.
(D) Neurospheres derived from Proband 1 show abundant apoptosis compared to
control cells. Scale bar, 100 um.


11
Supplemental Figure S5, related to Figure 6. Endogenous katanin complex
localizes to the centrosome and spindle pole.
(A) Endogenous katanin p80 and p60 localize to the spindle poles of mitotic HEK293T
cells. Scale bar, 10 um.
(B-C) Endogenous katanin p80 and p60 localize to the spindle poles of mitotic iPS cells
(B), and to the centrosome of interphase neural stem cells (NSCs) differentiated from
iPSCs (C). Localization is lost in mutant iPSCs and NSCs derived from Proband 1.
Scale bar, 10 um.


13
Supplemental Figure S6, related to Figure 7. Ultrastructural analysis of centrioles
and cytoskeletal immunohistochemistry.
(A) Gene-trap MEF centrosomes contain multiple unpaired centrioles by transmission
electron microscopy. Arrowhead, centriole. Scale bar, 200 nm.
(B) Cross-section of gene-trap MEF centriole shows normal 9-fold symmetry.
Arrowhead, centriole. Scale bar, 200 nm.
(C) Homozygous gt/gt MEFs fail to proliferate. Fisher’s exact test, p=0.03.
(D) Gene-trap MEF cytoskeleton is comprised of increased acetylated tubulin. Scale
bar, 10 um.
(E) Gene-trap MEFs exhibit increased staining for CAMSAP2, a microtubule minus-end
stabilizing protein. Scale bar, 10 um.
(F) Gene-trap MEFs show increased linear staining for EB1, a microtubule plus-end
protein, compared to the shorter, more puncta-like staining in wild-type MEFs. Scale
bar, 10 um.


15
Supplemental Figure S7, related to Figure 8. Cilia in neural progenitor cells.
(A) After stimulation by Smoothened agonist (SAG), gt/gt MEFs show reduced
Smoothened (Smo) localization to the cilium, indicating a deficit in Sonic hedgehog
signaling (right). Wild-type MEFs robustly relocate Smo to the cilium after SAG
stimulation (left). Scale bar, 10 um.


17
Supplemental Table S1, related to Figure 1. Clinical summary of affected
individuals.

Individual Proband 1 Proband 1 cousin Proband 1 cousin Proband 2 Proband 3Gender Male Female Male, deceased at 5 months Male Male
Parental consanguinity Yes Yes Yes Yes None reported, distant relation suspected based on homozygosity mapping
Prenatal and birth history Full term normal spontaneous vaginal delivery
Born by Caesarian section, no NICU admission
Unknown Unremarkable, full term normal spontaneous vaginal delivery
Microcephaly noted in 2nd trimester, full term normal spontaneous vaginal delivery
Head circumference at birth 30 cm (-‐3.5 SD) 27 cm (-‐5.9 SD) 27 cm (-‐5.9 SD) 29.5 cm (-‐3.9 SD) UnknownWeight at birth 3.7 kg (75th percentile) 3 kg (25th percentile) Unknown 3.3 kg (50th percentile) 3.2 kg (40th percentile)Length at birth Unknown Unknown Unknown 47 cm (-‐1.5 SD) UnknownAge at first evaluation 1 year old 9 months Unknown 3 months 8 monthsHead circumference at first evaluation 38 cm (-‐6.3 SD) 32 cm (-‐10.3 SD) Unknown 34.3 cm (-‐5.3 SD) 34 cm (-‐8.5 SD)Weight at first evaluation Unknown Unknown Unknown 7.2 kg (85th percentile) 6.8 kg (2nd percentile)Length at first evaluation Unknown Unknown Unknown 59.5 cm (-‐0.9 SD) 63 cm (-‐3.5 SD)Age at most recent evaluation 10 years 10 years Not applicable 3 years 4 yearsHead circumference at most recent evaluation
42.5 cm 41 cm Not applicable 39 cm (-‐7.4 SD) 39 cm (-‐7.7 SD)
Weight at most recent evaluation 22 kg 24 kg Not applicable 13 kg (20th percentile) 11.8 kg (0.5th percentile)Length at most recent evaluation 118.5 cm 120 cm Not applicable 90 cm (-‐1.6 SD) Unknown
Neurologic findings Spasticity in achilles tendon bilaterally
Increased tone in achilles tendon bilaterally with exaggerated deep tendon reflexes (+3) in knees and ankles, no clonus, babinski is downwards
Not applicable Spastic quadriplegia Mild hypertonia, increased deep tendon reflexes
Developmental history 10 years old: sitting alone, does not walk, no words (global delay )
10 years old: sitting alone, does not walk, no words
Not applicable Severe and global delay, no speech Head control at 1 year, rolled at 1.25 years, unable to sit unsupported at 4 years, no speech
Seizures 9 years old: seizures in the form of uprolling of eyes with decreased tone all over. Repeated after 6 months, treated with valproic acid
Developed seizures only once at the age of 7 years with cyanosis , uprolling of eyes and tonic contraction of upper limbs and lower limbs
Not applicable Yes, generalized tonic-‐clonic, onset 1 year, treated with Keppra
Yes, onset at 21 months, treated with phenobarbital
Hearing Grossly normal Grossly normal Not applicable Grossly normal Mild hearing loss at 45dB
Vision Grossly normal Grossly normal Not applicable Grossly normal (examined in a private clinic, report not available)
Bilateral vision loss with disc pallor and macular lesions
Dysmorphic features Microcephaly with decreased cranio-‐facial ratio and small cranial vault, large ears, synephrous
Microcephaly with decreased cranio-‐facial ratio and small cranial vault, large ears, synephrous, bulbous nose, almond eyes, big mouth
Not applicable Facial features consistent with microcephaly, single palmar creases, secondary craniosynostosis
Sloping forehead, round face, arched eyebrows, small nose with anteverted nares, thin upper lip, widely spaced teeth, increased internipplary distance, tapered fingers, small testes
Brain imaging MRI at 1 year and 11 years MRI at 10 years old Not applicable MRI at 17 months MRI at 11 months
cerebral cortex Simplification of the gyral pattern with thickened cortex (packygyria), broad flat gyri, smooth grey-‐white matter interface, brain atrophy
Simplification of the gyral pattern with thickened cortex (packygyria), broad flat gyri, smooth grey-‐white matter interface, shallow sylvian fissure
Moderately to severely simplified gyral pattern, normal cortical thickness
Severely simplified gyral pattern, normal cortical thickness
basal ganglia and thalami Grossly normal Grossly normal Grossly normal Grossly normal basal ganglia, small thalami
corpus callosum Partial agenesis of body and splenium (genu present)
Agenesis of the body and splenium (genu present), hypomyelination is noted
Partial agenesis (genu and body present)
Partial agenesis (only genu present)
ventricles Prominent ventricles enlarged ventricles (colpocephaly)
Enlarged ventricle (colpocephaly), the 4th ventricle is communicating with an enlarged cisterna magna, through a slit-‐like opening finding representing Dandy-‐Walker variant
Enlarged posteriorly Enlarged posteriorly, periventricular heterotopia
cerebellum and brain stem Atrophy involving the cerebellum and cerebellar vermis
Atrophy involving the cerebellum and cerebellar vermis
Grossly normal, enlarged cisterna magna
Grossly normal, enlarged cisterna magna
Other findings None None Not applicable None Oral pharyngeal dysphagia, elevated liver enzymes
Normal screening tests Blood Not available Not applicable Abdominal ultrasound Karyotype, toxoplasma, rubella, cytomegalovirus, herpes simplex, HIV, complete blood count, serum electrolytes, renal and thyroid function tests, creatine kinase, abdominal ultrasound

19
Supplemental Table S2, related to Figure 4. Plasmid and protein sequences of
TALENs used to generate zebrafish katnb1 mutants.

katnb1-L Plasmid sequence: gctgcttcgcgatgtacgggccagatatacgcgtggctccggtgcccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaaccggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttcccgagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccgccagaacacaatttaggtgacactatagaagctagcgtttaaacttaagctgatccactagtccagtgtggtggaattcgccatggactacaaagaccatgacggtgattataaagatcatgacatcgattacaaggatgacgatgacaagatggcccccaagaagaagaggaaggtgggcatccacggggtacccatggtagatttgagaactttgggatattcacagcagcagcaggaaaagatcaagcccaaagtgaggtcgacagtcgcgcagcatcacgaagcgctggtgggtcatgggtttacacatgcccacatcgtagccttgtcgcagcaccctgcagcccttggcacggtcgccgtcaagtaccaggacatgattgcggcgttgccggaagccacacatgaggcgatcgtcggtgtggggaaacagtggagcggagcccgagcgcttgaggccctgttgacggtcgcgggagagctgagagggcctccccttcagctggacacgggccagttgctgaagatcgcgaagcggggaggagtcacggcggtcgaggcggtgcacgcgtggcgcaatgcgctcacgggagcacccctcaacCTGACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTGTGCCAAGCGCACGGTTTAACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTGTGCCAAGCGCACGGCCTCACCCCAGAGCAGGTCGTGGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGC TGTGCCAAGCGCACGGATTAACCCCAGAGCAGGTCGTGGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTGTGCCAAGCGCACGGACTTACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTGTGCCAAGCGCACGGCCTTACCCCAGAGCAGGTCGTGGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTGTGCCAAGCGCACGGACTAACCCCAGAGCAGGTCGTGGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTGTGCCAAGCGCACGGGCTCACCCCAGAGCAGGTCGTGGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTGTGCCAAGCGCACGGGCTAACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTGTGCCAAGCGCACGGActcacgcctgagcaggtagtggctattgcatccaatggagggggcagacccgcactggagtcaatcgtggcccagctttcgaggccggaccccgcgctggccgcactcactaatgatcatcttgtagcgctggcctgcctcggcggacgacccgccttggatgcggtgaagaaggggctcccgcacgcgcctgcattgattaagcggaccaacagaaggattcccgagaggacatcacatcgagtggcaggatcccagctggtgaagagcgagctggaggagaagaagtccgagctgcggcacaagctgaagtacgtgccccacgagtacatcgagctgatcgagatcgccaggaacagcacccaggaccgcatcctggagatgaaggtgatggagttcttcatgaaggtgtacggctacaggggaaagcacctgggcggaagcagaaagcctgacggcgccatctatacagtgggcagccccatcgattacggcgtgatcgtggacacaaaggcctacagcggcggctacaatctgcctatcggccaggccgacgagatggagagatacgtggaggagaaccagacccggaataagcacctcaaccccaacgagtggtggaaggtgtaccctagcagcgtgaccgagttcaagttcctgttcgtgagcggccacttcaagggcaactacaaggcccagctgaccaggctgaaccacatcaccaactgcaatggcgccgtgctgagcgtggaggagctgctgatcggcggcgagatgatcaaagccggcaccctgacactggaggaggtgcggcgcaagttcaacaacggcgagatcaacttcagatcttgataactcgagtttaaacccgctgatcagcctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatgggcggccgccttctactgggcggttttatggacagcaagcgaaccggaattgccagctggggcgccctctggtaaggttgggaagccctgcaaagtaaactggatggctttcttgccgccaaggatctgatggcgcaggggatcaagctctgatcaagagacaggatgaggatcgtttcgcatgattgaacaagatggattgcacgcaggttctccggccgcttgggtggagaggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgccgtgttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgccctgaatgaactgcaagacgaggcagcgcggctatcgtggctggccacgacgggcgttccttgcgcagctgtgctcgacgttgtcactgaagcgggaagggactggctgctattgggcgaagtgccggggcaggatctcctgtcatctcaccttgctcctgccgagaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatccggctacctgcccattcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggatgatctggacgaagagcatcacgggctcgcgccagccgaactgttcgccaggctcaaggcgagcatgcccgacggcgaggatctcgtcgtgacccatggcgatgcctgcttgccgaatatcatggtggaaaatggccgcttttctggattcatcgactgtgccggctggtgtggcggaccgctatcaggacatagcgttggctaccgtgatattgctgaagagcttgcgcgaatggctgacgcttctcgtgcttacggtatcgccgcctcccgatgcatcagtgcacttttcggggaaatgtgcgcgaccctatttgtttatttttctaatacatcaaatatgtatccgctcatgagacaataacctgataaatgcttcaataatagcacgtgctaaaacttcatttttaaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcctttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgttcttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgt gcacacagcccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggag aaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatagt cctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcgg cctttttacggttcctggccttttgctggccttttgctcacatgttctt Protein: MDYKDHDGDYKDHDIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGFTHAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKIAKRGGVTAVEAVHAWRNALTGAPLNLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALDAVKKGLPHAPALIKRTNRRIPERTSHRVAGSQLVKSELEEKKSELRHKLKYVPHEYIELIEIARNSTQDRILEMKVMEFFMKVYGYRGKHLGGSRKPDGAIYTVGSPIDYGVIVDTKAYSGGYNLPIGQADEMERYVEENQTRNKHLNPNEWWKVYPSSVTEFKFLFVSGHFKGNYKAQLTRLNHITNCNGAVLSVEELLIGGEMIKAGTLTLEEVRRKFNNGEINFRS*

katnb1-R
Plasmid sequence: gctgcttcgcgatgtacgggccagatatacggctccggtgcccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttggggggaggggtcggcaattgaaccggtgcctagagaaggtggcgcggggtaaactgggaaagtgatgtcgtgtactggctccgcctttttcccgagggtgggggagaaccgtatataagtgcagtagtcgccgtgaacgttctttttcgcaacgggtttgccgccagaacacaatttaggtgacactatagaagctagcgtttaaacttaagctgatccactagtccagtgtggtggaattcgccatggactacaaagaccatgacggtgattataaagatcatgacatcgattacaaggatgacgatgacaagatggcccccaagaagaagaggaaggtgggcatccacggggtacccatggtagatttgagaactttgggatattcacagcagcagcaggaaaagatcaagcccaaagtgaggtcgacagtcgcgcagcatcacgaagcgctggtgggtcatgggtttacacatgcccacatcgtagccttgtcgcagcaccctgcagcccttggcacggtcgccgtcaagtaccaggacatgattgcggcgttgccggaagccacacatgaggcgatcgtcggtgtggggaaacagtggagcggagcccgagcgcttgaggccctgttgacggtcgcgggagagctgagagggcctccccttcagctggacacgggccagttgctgaagatcgcgaagcggggaggagtcacggcggtcgaggcggtgcacgcgtggcgcaatgcgctcacgggagcacccctcaacCTGACCCCAGAGCAGGTCGTGGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTGTGCCAAGCGCACGGTTTAACCCCAGAGCAGGTCGTGGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTGTGCCAAGCGCACGGCCTCACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTGTGCCAAGCGCACGGATTAACCCCAGAGCAGGTCGTGGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTGTGCCAAGCGCACGGACTTACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTGTGCCAAGCGCACGGCCTTACCCCAGAGCAGGTCGTGGCAATCGCGAGCAATAACGGCGGAAAACAGGCTTTGGAAACGGTGCAGAGGCTCCTTCCAGTGCTGTGCCAAGCGCACGGACTAACCCCAGAGCAGGTCGTGGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTGTGCCAAGCGCACGGGCTCACCCCAGAGCAGGTCGTGGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCCATTGCCTCGAATGGAGGGGGCAAACAGGCGTTGGAAACCGTACAACGATTGCTGCCGGTGCTGTGCCAAGCGCACGGGCTAACCCCAGAGCAGGTCGTGGCAATCGCCTCCAACATTGGCGGGAAACAGGCACTCGAGACTGTCCAGCGCCTGCTTCCCGTGCTTTGTCAGGCACACGGCCTCACTCCGGAACAAGTGGTCGCGATCGCAAGCCACGACGGAGGAAAGCAAGCCTTGGAAACAGTACAGAGGCTGTTGCCTGTGCTGTGCCAAGCGCACGGActcacgcctgagcaggtagtggctattgcatcccacgacgggggcagacccgcactggagtcaatcgtggcccagctttcgaggccggaccccgcgctggccgcactcactaatgatcatcttgtagcgctggcctgcctcggcggacgacccgccttggatgcggtgaagaaggggctcccgcacgcgcctgcattgattaagcggaccaacagaaggattcccgagaggacatcacatcgagtggcaggatcccagctggtgaagagcgagctggaggagaagaagtccgagctgcggcacaagctgaagtacgtgccccacgagtacatcgagctgatcgagatcgccaggaacagcacccaggaccgcatcctggagatgaaggtgatggagttcttcatgaaggtgtacggctacaggggaaagcacctgggcggaagcagaaagcctgacggcgccatctatacagtgggcagccccatcgattacggcgtgatcgtggacacaaaggcctacagcggcggctacaatctgcctatcggccaggccgacgagatgcagagatacgtgaaggagaaccagacccggaataagcacatcaaccccaacgagtggtggaaggtgtaccctagcagcgtgaccgagttcaagttcctgttcgtgagcggccacttcaagggcaactacaaggcccagctgaccaggctgaaccacaaaaccaactgcaatggcgccgtgctgagcgtggaggagctgctgatcggcggcgagatgatcaaagccggcaccctgacactggaggaggtgcggcgcaagttcaacaacggcgagatcaacttctgataactcgagtctagagggcccgtttaaacccgctgatcagcctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatgggcggccgccttctactgggcggttttatggacagcaagcgaaccggaattgccagctggggcgccctctggtaaggttgggaagccctgcaaagtaaactggatggctttcttgccgccaaggatctgatggcgcaggggatcaagctctgatcaagagacaggatgaggatcgtttcgcatgattgaacaagatggattgcacgcaggttctccggccgcttgggtggagaggctattcggctatgactgggcacaacagacaatcggctgctctgatgccgccgtgttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgccctgaatgaactgcaagacgaggcagcgcggctatcgtggctggccacgacgggcgttccttgcgcagctgtgctcgacgttgtcactgaagcgggaagggactggctgctattgggcgaagtgccggggcaggatctcctgtcatctcaccttgctcctgccgagaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatccggctacctgcccattcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggatgatctggacgaagagcatcacgggctcgcgccagccgaactgttcgccaggctcaaggcgagcatgcccgacggcgaggatctcgtcgtgacccatggcgatgcctgcttgccgaatatcatggtggaaaatggccgcttttctggattcatcgactgtgccggctggtgtggcggaccgctatcaggacatagcgttggctaccgtgatattgctgaagagcttgcgcgaatggctgacgcttctcgtgcttacggtatcgccgcctcccgatgcatcagtgcacttttcggggaaatgtgcgcgaccctatttgtttatttttctaatacatcaaatatgtatccgctcatgagacaataacctgataaatgcttcaataatagcacgtgctaaaacttcatttttaaatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcctttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgttcttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttctt Protein: MDYKDHDGDYKDHDIDYKDDDDKMAPKKKRKVGIHGVPMVDLRTLGYSQQQQEKIKPKVRSTVAQHHEALVGHGFTHAHIVALSQHPAALGTVAVKYQDMIAALPEATHEAIVGVGKQWSGARALEALLTVAGELRGPPLQLDTGQLLKIAKRGGVTAVEAVHAWRNALTGAPLNLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNNGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNGGGKQALETVQRLLPVLCQAHGLTPEQVVAIASNIGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGKQALETVQRLLPVLCQAHGLTPEQVVAIASHDGGRPALESIVAQLSRPDPALAALTNDHLVALACLGGRPALDAVKKGLPHAPALIKRTNRRIPERTSHRVAGSQLVKSELEEKKSELRHKLKYVPHEYIELIEIARNSTQDRILEMKVMEFFMKVYGYRGKHLGGSRKPDGAIYTVGSPIDYGVIVDTKAYSGGYNLPIGQADEMQRYVKENQTRNKHINPNEWWKVYPSSVTEFKFLFVSGHFKGNYKAQLTRLNHKTNCNGAVLSVEELLIGGEMIKAGTLTLEEVRRKFNNGEINF*

21
Extended Experimental Procedures
Human Subjects
We collected peripheral blood samples from the affected individuals and one or
both of their parents after obtaining written informed consent according to the protocols
approved by the participating institutions. All human studies were reviewed and
approved by the institutional review board of Boston Children’s Hospital, Harvard
Medical School and all local institutions.
Homozygosity Mapping, Exome Sequencing and Filtering in Family 1
Genomic DNA was extracted from saliva samples using the Oragen Purifier Kit.
Proband 1, the affected cousin, and their unaffected parents and siblings were
genotyped using Illumina Human OmniExpress 12v1 BeadChips following
manufacturer’s instructions. Call rates were above 99%. Gender and relationship were
verified using Illumina BeadStudio software. Mapping was performed by searching for
shared regions that are homozygous and identical-by-descent (IBD) in the 2 affected
individuals using custom programs written in the Mathematica (Wofram Research, Inc.)
data analysis software. Candidate regions were further refined by exclusion of common
homozygous segments with any unaffected family members. The confidence criteria to
identify IBD blocks were a minimum of 3 cM and a SNP density over the 5th percentile.
Centimorgan distances between SNPs were determined from the HapMap Phase II
recombination rate map. We identified a single shared identical-by-descent candidate
locus totaling 9 Mb on Chromosome 16 (hg19; chr16:55,216,971-64,204,977).
Whole exome sequencing of Proband 1 was performed using the Illumina
TruSeq Exome Enrichment Kit for exome capture using 1 ug of genomic DNA. Illumina

22
HiSeq2000 High-output mode was used for sequencing as 100 bp paired-end runs at
the UCLA Clinical Genomics Centre and at the UCLA Broad Stem Cell Research
Centre. 6 samples were pooled before the exome capture and sequenced across 2
flowcell lanes. Sequence reads were aligned to the human reference genome (Human
GRCh37 (hg19) build) using Novoalign (v2.07,
http://www.novocraft.com/main/index.php). PCR duplicates were identified by Picard
(v1.42, http://picard.sourceforge.net/) and Genome Analysis Toolkit (v1.1,
http://www.broadinstitute.org/gatk/)(McKenna et al., 2010) was used to re-align indels,
recalibrate the quality scores, call, filter, recalibrate and evaluate the variants. SNVs
and INDELs across the sequenced protein-coding regions and flanking junctions were
annotated using Variant Annotator X (VAX), a customized Ensembl Variant Effect
Predictor(Yourshaw et al., 2014). Each variant was annotated with information including
gene names and accession numbers, reference variant, variant consequences, protein
positions and amino-acid changes, conservation scores, population minor allele
frequencies, expression pattern, and PolyPhen2(Adzhubei et al., 2010), SIFT(Kumar et
al., 2009), Condel(González-Pérez and López-Bigas, 2011) predictions. An average
coverage of 62X was achieved across the exome with 86.2% of these bases covered at
≥10X. A total of 19,247 variants were identified across the RefSeq protein coding exons
and flanking introns (+/-2bp). Of these, 1515 variants (76 homozygous and 1439
heterozygous variants) were protein-changing variants with population minor allele
frequencies <1%.
A single, unique homozygous start codon mutation in KATNB1 (c.1A>G, hg19
chr16: 57,771,156) was present in the region of IBD. Sanger sequencing confirmed and

23
validated mutation segregation in all tested family members, using forward primer 5’-
TTCTGTCTGCCTGGATGTG and reverse primer 5’-GAAACAAAGCTGTGGGTTCAG.
Exome Sequencing and Filtering in Proband 2
Exome capture was performed using TruSeq Exome Enrichment kit (Illumina,
San Diego, CA, USA) following the manufacturer’s protocol. Samples were prepared as
an Illumina sequencing library, and in the second step, the sequencing libraries were
enriched for the desired target using the Illumina Exome Enrichment protocol. The
captured libraries were sequenced using the Illumina HiSeq 2000 Sequencer. The
reads were mapped against UCSC hg19 (http://genome.ucsc.edu/) by BWA (Burrows-
Wheeler Aligner) (http://bio-bwa.sourceforge.net/). SNPs and Indels were detected by
SAMTOOLS (http://samtools.sourceforge.net/).
Exome-derived data were filtered for variants as follows. A total of 82073 exome
variants were identified, of which 37,114 were homozygous, consistent with inheritance
from consanguineous parents. We excluded all variants that did not reside in a coding
or splice site region, or were present in dbSNP or the 1000Genomes database, leaving
175 variants. Subsequent steps involved removal of all variants which resided outside
the autozygome, or which were present in our Saudi population specific database (167
exomes) at an allele frequency greater than 0.005, or listed within the Exome Variant
Server, leaving only 4 variants. Finally the pathogenicity of all surviving missense
variants was established using Polyphen-2 and SIFT, and any variant predicted benign
by both algorithms was eliminated, with 3 variants remaining.

24
Gene Exonic Function Change Present in control
Saudi database
(167 exomes)
GLYATL3 frameshift deletion
NM_001010904:exon6:c.577_578del:
p.193_193del Yes
GJA10 nonsynonymous SNV NM_032602:exon1:c.669G>T: p.L223F Yes
KATNB1 nonsynonymous SNV NM_005886:exon3:c.97G>T: p.G33W No
KATNB1 was the only gene with variants that intersected with candidate variants in
Family 1 and Proband 3, and only the KATNB1 missense variant was completely absent
from our Saudi population specific database (c.97G>T, hg19 chr16:57,775,655). This
variant was confirmed by Sanger sequencing using the forward primer 5’-
GGTCACCCCTCCTTCACAAG and reverse primer 5’- TCACTCTTCAGCCTCCAACC.
Genome-wide SNP genotyping and homozygosity mapping in Proband 3
Proband 3 and his father were genotyped by the Illumina Human610-Quad
BeadChip array according to manufacturers’ instructions. Though the parents of the
affected individual were not reported to be consanguineous, there were 11 blocks of
homozygosity over 2.0 cM in Proband 3 that were not shared by his father. The largest
block was 13.4 cM in size, suggesting that the parents are likely to be distantly related.

25
Chromosome Interval Size (cM) Size (Mb)
16 rs8046668; rs12373064 13.4 14.1
14 rs11624171; rs4562969 9.5 5.8
16 rs2966097; rs7195057 6 1.7
5 rs1549221; rs2913861 5.5 2.9
5 rs3765045; rs13158449 2.9 1.9
20 rs242435; rs6129021 2.6 0.8
21 p terminus; rs2207843 2.6 4.3
1 rs7541950; rs11205321 2.6 2.4
9 rs10116618; rs28651528 2.4 4.8
9 rs28651528; rs10869153 2.2 26.8
14 rs2401756; rs9323838 2.1 0.9
Array capture and exon sequencing in Proband 3
We performed array capture followed by high-throughput Illumina sequencing of
all coding exons within the blocks of homozygosity over 2.0 cM in Proband 3 that were
not shared by his father. Array capture and sequencing were performed as previously
described(Yu et al., 2010) in Proband 3. Briefly, a custom microarray was designed with
oligonucleotide probes targeting all exons within the candidate interval. DNA bound to
the array was amplified and used to generate sequencing libraries. The average depth
of target coverage was 138X, with 86.6% of bases covered by ≥10 reads. After filtering
for standard quality and mapping confidence, a total of 1297 potential homozygous
sequence variants were identified.

26
Variant analysis and sequencing confirmation in Proband 3
Variants were annotated with respect to predicted molecular effect and presence
in known population datasets using ANNOVAR(Wang et al., 2010) as well as the
genomic mutation consequence calculator(Major, 2007). To identify candidate
pathogenic variants, variants were filtered to exclude those found in known population
datasets with a minor allele frequency (MAF) of 1% or greater in the 1000 Genomes
Project (http://www.1000genomes.org) or the NHLBI Exome Sequencing Project
(http://evs.gs.washington.edu/EVS), and to exclude those predicted to be molecularly
nonimpactful (intergenic, synonymous, or intronic and greater than 2bp from a splice
junction). Of the 1297 homozygous variants identified in the individual, 74 were protein-
altering, and only 7 variants were rare (MAF 0.01 as above) and protein-altering. Of the
7 variants, 3 are present in dbSNP132 and 2 others are present at very low frequency in
EVS; these 5 were all predicted to be tolerated by Polyphen-2 and SIFT, leaving two
remaining candidate variants.

27
Gene Exonic
Function
Change Exome
Variant
Server
6500
allele
frequency
dbSNP132 Polyphen
-2
SIFT
TPTE
nonsynonymous
SNV NM_199260:c.A1042G: p.K348E rs212146 NA Tolerated
LOC100288
966,POTED
nonsynonymous
SNV NM_001257362:c.G337A: p.G113S rs6517869 Benign Tolerated
LOC100288
966,POTED
nonsynonymous
SNV NM_001257362:c.A1603G: p.M535V rs78643169 Benign Tolerated
CDC42BPB
nonsynonymous
SNV NM_006035:c.C2948T: p.A983V 0.000923 Benign Tolerated
PROP1
nonsynonymous
SNV NM_006261:c.C425T: p.A142V 0.00246
Benign Tolerated
NOP16
frameshift
insertion
NM_001256539:c.586_587insAC:
p.R196fs
KATNB1 splicing NM_005886:exon6:c.432+1G>A
Of these 2 variants, whole-exome sequencing of Probands 1 and 2 led to the
elimination the NOP16 variant, leaving a single remaining candidate. This was a
homozygous single base pair change in KATNB1 in the first base of the intron
(c.432+1G>A, hg19 chr16:57,785,215). This variant was verified by Sanger sequencing
with forward primer 5’-TTGTCTCCGTGGGGAGTAAC and reverse primer 5’-
CTAGGGGATTCATGCAGAGC. The primers for Sanger confirmation of this variant
were designed with Primer3 (http://frodo.wi.mit.edu/primer3/)(Rozen and Skaletsky,
2000).

28
Sequencing of KATNB1 in Controls
The missense mutation was screened against 178 ethnically matched Saudi
Arabian controls and 82 tribe-specific controls. The splice site mutation is absent from
>800 exomes sequenced in the Walsh lab (many of whom are from individuals from the
Middle East with brain malformations or autism) and from the Exome Variant Server
(EVS, http://evs.gs.washington.edu/EVS/). No allele carrying the mutation was found in
these controls.
Animals
All animal experimentation was carried out under protocols approved by the
IACUCs of Harvard Medical School and Boston Children’s Hospital. Targeted gene-trap
ES cells (EUCOMM) were injected into C57BL/6 blastocysts to generate chimeric mice.
Male chimeras were be bred to WT C57BL/6 background females (Charles River
laboratories) to transmit the gene-trap allele. The mice are maintained on a C57/Bl6
background. For BrdU pulse-labeling experiments, E12.5 timed pregnant mice were
injected with 75 mg/kg BrdU intraperitoneally and euthanized 30 minutes later. For
histological analysis, brains were perfused with PBS and then followed by 4% PFA
before paraffin embedding and sectioning, or cryoprotected in 30% sucrose before
embedding in OCT (Tissue-Tek) and cryosectioning. Genotyping primers are available
upon request.
Transcription activator-like effector nucleases (TALENs) targeting the exon 6-
intron 6 boundary of zebrafish katnb1 (ENSDARG00000005456) were designed by and
ordered from ZGENEBIO Biotech Co. LTD (Supplementary Table 2). Plasmids were
linearized with NotI and RNA was subsequently synthesized using the mMESSAGE

29
mMachine SP6 Transcription kit (Life Technologies). To generate katnb1 mutants, 150-
200pg of RNA was injected into eggs (albino/slc45a2) at the 1-cell stage. Fish in the F1
generation, stemming from brother-sister matings of injected fish, were prescreened for
induced mutations by heteroduplex mobility assay(Ota et al., 2013). Genotyping was
subsequently confirmed by colony PCR and Sanger sequencing of individual fish to
identify the specific alleles. Genotyping primers are available upon request.
Culture Systems
HEK293T cells were grown in Dulbecco’s Modified Eagle Medium, High Glucose,
High Glucose (HyClone) with 10% fetal bovine serum (FBS, Gibco) and 1mM penicillin,
streptomycin and L-glutamine. Patient point mutations were generated using
QuikChange kit (Stratagene). For tagged KATNB1 mini-gene and cDNA
overexpression, we transfected 293T cells with specified plasmids using Fugene6
(Roche). Constructs were generated using cDNA for Refseq gene ID 10300.
Primary fibroblasts were established from Proband 1 and were maintained in
RPMI-1640 (Gibco) with 10% FBS, and 1mM Penicillin and Streptomycin. Human
iPSCs were generated according to published protocols(Takahashi et al., 2007) using
retroviral vectors encoding the human cDNAs of KLF4, SOX2, OCT4, and C-MYC
(Addgene) and grown in Knockout DMEM containing 20% knockout serum replacement,
2 mM L-glutamine, 1% NEAA, 0.1 mM b-mercaptoethanol, 1% NEAA, 0.1% penicillin
and streptomycin, and 4 ng/ml bFGF on irradiated fibroblast feeders. iPSC colonies
were manually picked after 3–4 weeks and cultured on irradiated MEFs. Control cell
lines are from an unrelated, unaffected ethnically-matched individual.

30
For neural stem cell (NSC) generation, iPSCs were dissociated into single cells
and 10,000 cells/well were seeded into a low attachment U-bottom 96-well plate to form
cell aggregates of embryoid bodies (EB) in neural induction medium (DMEM/F12
supplemented with 20% KOSR, 2mM L-glutamine, 0.2mM NEAA, 0.1mM 2-
mercaptoethanol and 1mM Sodium Pyruvate) for 7 days. EBs were then adhered onto
laminin-coated plates in neural precursor medium (Neurobasal medium supplemented
with 2mM L-glutamine, 1% B27, 1% N2 and 20ng/ml bFGF) for the formation of rosette-
like structures for additional 7-14 days. The neural rosettes were manually isolated and
further expanded as NSCs in neural induction medium. For formation of neurospheres,
8000 cells/well of NSCs were seeded into a low attachment U-bottom 96-well plate in
neural precursor medium to form neurospheres of equal sizes. Sphere size was
measured every day for 6 days.
Lymphoblastoid cell lines were established from Proband 2 following Epstein-
Barr virus (EBV) immortalization and were maintained in RPMI-1640 (Gibco) with 10%
FBS, and 1mM Penicillin and Streptomycin. For imaging, they were grown on Poly-L-
Ornithine (Sigma) coated coverslips. Control cell lines are from an unrelated, unaffected
individual.
Gene-trap MEFs were isolated from E13.5 embryos and dissociated by
trypsinization. Cells were grown in Dulbecco’s Modified Eagle Medium, High Glucose
(HyClone) with 15% fetal bovine serum (FBS, Gibco) and 1mM penicillin, streptomycin
and L-glutamine. Cilia were induced by serum starvation for 48 hrs in Optimem (Gibco)
media, then treated with DMSO or 100nM SAG for an additional 24 hours.

31
RT-PCR and Quantitative RT-PCR
For RT-PCR, RNA was harvested using RNAeasy Plus kit (Qiagen). 1 ug of RNA
was used to synthesize cDNA using Superscript First Strand Synthesis kit (Invitrogen).
For qRT-PCR, total RNA was extracted from Proband 1-derived fibroblasts and
iPSCs using Nucleospine RNAII Kit (Macherey-Naga) with DNAse digestion. RNA was
quantified using ND-1000 spectrophotometer (Biofrontier Technology) and first strand
cDNA was produced with M-MulV reverse transcriptase (Biolabs) using 1 µg of total
RNA input. Quantitative RT-PCR, normalized to GAPDH expression levels, was
performed with the SYBER Green Mix (Roche). Primers were as follows: KATNB1
forward 5’-ACTGGTGCTGGGCAAAG; KATNB1 reverse 5’-
TGCAGTTGGGCTTGTTGA; KATNA1 forward 5’-GGCGGGTTTGTCTTGAAATTC;
KATNA1 reverse 5’-GTACTGCCCAGTTCAATCGT.
For Shh qRT-PCR, MEFs were plated in 6-well plates, starved overnight in Opti-
MEM, and then treated with DMSO or 100nM SAG for an additional 24 hours. RNA was
harvested using an RNAeasy Plus kit (Qiagen). 1 ug of RNA was used to synthesize
cDNA using a First Strand cDNA synthesis kit (Maxima). The resulting cDNA was
probed with primers to Gli1, Patched and beta-Actin and amplified using Sybr GreenER
(Invitrogen). Primer and probe sequences as previously described(Garcia-Gonzalo et
al., 2011).
Western Blot
Lysate was prepared using RIPA buffer (50 mM Tris-HCl 7.4, 150 mM NaCl, 1%
NP-40, 0.5% sodium deoxycholate, and 0.1% SDS) containing a protease inhibitor
cocktail mix (Roche). Protein concentrations were normalized using the bicinchoninic

32
acid assay (Thermo Scientific). Extracted proteins were separated by electrophoresis on
Bis-Tris polyacrylamide gels (Life Technologies) or SDS polyacrylamide gels with DTT,
followed by transfer on polyvinylidene difluoride membrane (Millipore).
For quantitative Western blotting, membranes were blocked for 1 hr in Odyssey
Blocking Buffer (LICOR Biosciences) at room temperature, incubated them with primary
antibodies according to the antibody manufacturer’s instructions, and then incubated
them with fluorescent-dye-conjugated secondary antibodies (LICOR Biosciences).
Bands were detected, normalized and quantified with the Odyssey Infrared Imaging
System (Li-Cor Biosciences).
Antibodies: Flag (Cell Signaling, 1:1000); GAPDH (Abcam, 1:2000); Katanin p60
(Proteintech, 1:200; Atlas, 1:200); Katanin p80 (Proteintech, 1:500; Santa Cruz, 1:200;
LSBio, 1:2000; Sigma, 1:100)
Immunostaining
Cells were grown on glass coverslips and fixed in either 4% PFA for 20 minutes
at room temperature, or 100% Methanol for 3 minutes at -20C. Cells were blocked (5%
serum in PBS-0.1% Triton-X), incubated with antibodies and mounted using Gelvatol or
Fluoromount-G (Southern Biotech) mounting media.
Embryos were drop fixed in 4% PFA overnight, followed by PBS wash. Paraffin
sections (5 um) of brains were rehydrated and subjected to antigen retrieval with
Retrievagen A (BD Pharmingen), followed by blocking (5% serum in PBS),
permeabilization (0.1% Triton X-100), and antibody incubation. Alternatively,
cryosections (50 um) were permeabilized (0.1% Triton X-100), blocked, and incubated
with antibodies (24h-48h).

33
All samples were counterstained with DAPI or Hoechst 33258 (Sigma). Images were
acquired with a Zeiss LSM 700 on an upright Axio Imager.M2 equipped with 405, 488,
555 and 639 nm solid state lasers, motorized stage and a CCD AxioCam MR; an
inverted Zeiss Axio Observer D1; or a DeltaVision RT GE microscope.
For Gli3 immunocytochemistry quantification, Gli3 intensity was measured at the
ciliary tip and background intensity was subtracted.
Antibodies: Acetylated tubulin (Abcam, 1:500; MeOH); Arl13b (kind gift of Tamara
Caspary, 1:500; PFA); Beta catenin (BD Biosciences, 1:100; PFA); Beta tubulin
(Abcam, 1:400; PFA); BrdU (AbD Serotec, 1:500; PFA); CAMSAP2 (Proteintech,
1:1000; MeOH); Centrin (Millipore, 1:1000; MeOH); Centromere (Antibodies Inc, 1:100;
PFA); Cep164 (kind gift of Erich Nigg, 1:1000; MeOH); Cep170 (Abcam, 1:500; PFA);
Cep63 (Millipore, 1:500; MeOH); Cleaved Caspase 3 (Abcam, 1:500; PFA); Cyclin A
(Santa Cruz, 1:200; MeOH); DCX (Santa Cruz, 1:250; PFA); EB1 (BD Biosciences,
1:250; MeOH); Gamma tubulin (Sigma, 1:200, MeOH; Abcam, 1:300, PFA); GFP
(Abcam, 1:1000; PFA); Gli3 (kind gift of Suzie Scales, 1:1000; MeOH); GT335 poly
glutamylated tubulin (Enzo Life Sciences, 1:500; PFA); IFT88 (Proteintech, 1:500;
MeOH); Katanin p60 (Proteintech, 1:500; Atlas, 1:250; PFA); Katanin p80 (Proteintech,
1:500; Santa Cruz, 1:500; Sigma, 1:2000; PFA); LacZ (MP biomedicals, 1:250; PFA);
Ninein (kind gift of James Sillibourne, 1:2500; MeOH); OCT4 (Santa Cruz, 1:100; PFA);
Pericentrin (Abcam, 1:1000; PFA); Phosphorylated Histone H3 (Cell Signaling, 1:1000,
MeOH); Smoothened (Abcam, 1:500; PFA); Sox2 (Santa Cruz, 1:500; R&D, 1:200;
PFA); SSEA4 (Millipore, 1:100; PFA); STIL (Bethyl Labs, 1:200; MeOH); Tbr2 (Abcam,

34
1:500; PFA); TRA-1-60 (Millipore, 1:100; PFA); TRA-1-81 (Millipore, 1:100; PFA); Yo
Pro (Invitrogen, 1:500; PFA)
Electron Microscopy
Cells were grown on aclar coverslips and fixed for 20 minutes in a 1:1 dilution of
culture media to 5% Glutaraldehyde, 2.5% Paraformaldehyde, 0.06 % picric acid in 0.2
M sodium cacodylate buffer (pH 7.4), washed 3x in 0.1M Cacodylate buffer, then post-
fixed in 1% Osmium tetroxide (OsO4)/1.5% Potassiumferrocyanide (KFeCN6) for 30
min, washed in water 3x and incubated in 1% aqueous uranyl acetate for 30min
followed by 2 washes in water and subsequent dehydration in grades of alcohol (5min
each; 50%, 70%, 95%, 2x 100%). Cells were embedded in TAAB Epon (Marivac
Canada Inc. St. Laurent, Canada) and polymerized at 60 degrees C for 48 hrs. After
polymerization the aclar coverslip was peeled off and 1mm squares of the embedded
monolayer was glued onto an empty Epon block for sectioning.
Ultrathin serial sections (80nm) were cut on a Reichert Ultracut-S microtome,
picked up on to formvar/carbon coated copper grids, stained with lead citrate and
examined in a JEOL 1200EX Transmission electron microscope. Images were recorded
with an AMT 2k CCD camera.
Statistical Analyses
All data are represented as Mean ± SEM. Analyses were performed using at least 3
samples each from 3 different animals of each genotype, or using at least 2 replicates
for Proband-derived cell lines. ANOVAs were analyzed using Bonferroni test correction
for multiple comparisons. No F-tests were significant for unequal variances; therefore,
unpaired t-tests were analyzed assuming equal variances

35
References
Adzhubei, I.A., Schmidt, S., Peshkin, L., Ramensky, V.E., Gerasimova, A., Bork, P., Kondrashov, A.S., and Sunyaev, S.R. (2010). A method and server for predicting damaging missense mutations. Nat Meth 7, 248–249.
Garcia-Gonzalo, F.R., Corbit, K.C., Sirerol-Piquer, M.S., Ramaswami, G., Otto, E.A., Noriega, T.R., Seol, A.D., Robinson, J.F., Bennett, C.L., Josifova, D.J., et al. (2011). A transition zone complex regulates mammalian ciliogenesis and ciliary membrane composition. Nat Genet 43, 776–784.
González-Pérez, A., and López-Bigas, N. (2011). Improving the assessment of the outcome of nonsynonymous SNVs with a consensus deleteriousness score, Condel. Am J Hum Genet 88, 440–449.
Kumar, P., Henikoff, S., and Ng, P.C. (2009). Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat Protoc 4, 1073–1081.
Major, J.E. (2007). Genomic mutation consequence calculator. Bioinformatics 23, 3091–3092.
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., Garimella, K., Altshuler, D., Gabriel, S., Daly, M., et al. (2010). The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res 20, 1297–1303.
Ota, S., Hisano, Y., Muraki, M., Hoshijima, K., Dahlem, T.J., Grunwald, D.J., Okada, Y., and Kawahara, A. (2013). Efficient identification of TALEN-mediated genome modifications using heteroduplex mobility assays. Genes Cells 18, 450–458.
Rozen, S., and Skaletsky, H. (2000). Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol 132, 365–386.
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., and Yamanaka, S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861–872.
Wang, K., Li, M., and Hakonarson, H. (2010). ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res 38, e164.
WHO Multicentre Growth Reference Study Group (2006). WHO Child Growth Standards based on length/height, weight and age. Acta Paediatr Suppl 450, 76–85.
Yourshaw, M., Taylor, S.P., Rao, A.R., Martín, M.G., and Nelson, S.F. (2014). Rich annotation of DNA sequencing variants by leveraging the Ensembl Variant Effect Predictor with plugins. Brief. Bioinformatics.

36
Yu, T.W., Mochida, G.H., Tischfield, D.J., Sgaier, S.K., Flores-Sarnat, L., Sergi, C.M., Topçu, M., McDonald, M.T., Barry, B.J., Felie, J.M., et al. (2010). Mutations in WDR62, encoding a centrosome-associated protein, cause microcephaly with simplified gyri and abnormal cortical architecture. Nat Genet 42, 1015–1020.




















