
Cendawan Endofit - Universitas Muhammadiyah Makassar
200
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Cendawan Endofit - Universitas Muhammadiyah Makassar


Cendawan Endofit
i
CENDAWAN ENDOFIT
TEKNIK ISOLASI, IDENTIFIKASI
DAN POTENSI PEMANFAATAN CENDAWAN
ENDOFIT DALAM BUDIDAYA
TANAMAN

ii
Cendawan Endofit
Sanksi Pelanggaran Pasa 113UU No. 28 Tahun 2014 tentang Hak Cipta
Ketentuan PidanaPasal 113(1) Setiap Orang yang dengan tanpa hak melakukan pelanggaran hak ekonomi
sebagaimana dimaksud dalam Pasal 9 ayat (1) huruf i untuk Penggunaan Secara Komersial dipidana dengan pidana penjara paling lama 1 (satu) tahun dan/atau pidana denda paling banyak Rp lOO.OOO.OOO (seratus juta rupiah).
(2) Setiap Orang yang dengan tanpa hak dan/atau tanpa izin Pencipta atau pemegang Hak Cipta melakukan pelanggaran hak ekonomi Pencipta sebagaimana dimaksud dalam Pasal 9 ayat (1) huruf c, huruf d, huruf f, dan/atau huruf h untuk Penggunaan Secara Komersial dipidana dengan pidana penjara paling lama 3 (tiga) tahun dan/atau pidana denda paling banyak Rp 500.000.000,00 (lima ratus juta rupiah).
(3) Setiap Orang yang dengan tanpa hak dan/atau tanpa izin Pencipta atau pemegang Hak Cipta melakukan pelanggaran hak ekonomi Pencipta sebagaimana dimaksud dalam Pasal 9 ayat (1) huruf a, huruf b, huruf e, dan/atau huruf g untuk Penggunaan Secara Komersial dipidana dengan pidana penjara paling lama 4 (empat) tahun dan/atau pidana denda paling banyak Rp l.OOO.OOO.OOO,OO (satu miliar rupiah).
(4) Setiap Orang yang memenuhi unsur sebagaimana dimaksud pada ayat (3) yang dilakukan dalam bentuk pembajakan, dipidana dengan pidana penjara paling lama 10 (sepuluh) tahun dan/atau pidana denda paling banyak Rp 4.000.000.000,00 (empat miliar rupiah).

Cendawan Endofit
iii
Diterbitkan oleh:
CENDAWAN ENDOFIT
TEKNIK ISOLASI, IDENTIFIKASI
DAN POTENSI PEMANFAATAN CENDAWAN
ENDOFIT DALAM BUDIDAYA
TANAMAN
Dr. Syamsia, S.P., M. Si
Dr. Ir. Abubakar Idhan, M.P.
Dr. Amanda Patappari Firmansyah, S.P., M.P.
Noerfiryani, S.P., M. Si

iv
CENDAWAN ENDOFIT TEKNIK ISOLASI, IDENTIFIKASI DAN POTENSI PEMANFAATAN CENDAWAN ENDOFIT DALAM BUDIDAYA TANAMAN
Penulis: Dr. Syamsia,S.P.,M. Si Dr. lr. Abubakar ldhan,M.P. Dr. Amanda Patappari Firmansyah,S.P.,M.P. Noerfiryani,S.P.,M. Si
Cetakan Pertama: 2021 Ukuran: 15x23 cm, x + 188
ISBN: 978-623-7349-40-2
Editor:
Dr. lradhatullah Rahim,S.P.,M.P.
Penyuting:
Ma'ruf
Desain Sampuldan Tata Letak REESLITERA
Jl. Antang Raya No.99A, Makassar- Sulawesi Selatan Kontak!WA: 082191865019-085342101139
Penerbit: LPP UNISMUH MAKASSAR Anggota IKAPI
No. 021/Anggota Luar Biasa/SSL/2019
Hak cipta dilindungi Undang-Undang
Dilarang memperbanyak karya tulis ini dalam bentuk dan dengan
cara apapun tanpa ijin dari penerbit.

Cendawan Endofit
v
PRAKATA
Syukur Alhamdulillah, penulisan buku ajar ini dapat dirampungkan. Buku ajar ini merupakan hasil riset penulis yang terkait dengan cendawan endofit sejak tahun 2014 sampai sekarang. Buku ajar ini membahas tentang: pengertian dan manfaat cendawan endofit, peralatan dan bahan untuk isolasi cendawan, isolasi dan identifikasi cendawan, media pertumbuhan dan penyimpanan, serta manfaat cendawan endofit pada pertumbuhan tanaman.
Buku ajar ini diperuntukkan bagi mahasiswa Program Studi Agroteknologi khususnya yang memprogramkan mata kuliah Mikrobiologi, Pengantar Bioteknologi dan Bioteknologi Pertanian serta mata kuliah yang berkaitan dengan pemanfaatan mikroba dalam budidaya tanaman.
Ucapan terima kasih penulis sampaikan kepada Kementerian Riset dan Teknologi Badan Riset dan Inovsi Nasional (RISTEK-BRIN) telah mendanai kegiatan hibah penelitian skema Hibah Penelitian Disertasi (2014), Penelitian Produk Terapan (2015-2017), Penelitian Dasar Unggulan Perguruan Tinggi (2018-2020).
Penulis menyadari buku ajar ini masih banyak kekurangan. Oleh karena itu, penulis senantiasa menerima kritik dan saran yang sifatnya konstruktif demi penyempurnaan buku ini.
Makassar, 18 Februari 2021
Tim Penulis

vi
Cendawan Endofit
DAFTAR ISI
vvi
viiiix11244
88111414
1818192727
3131323738
PRAKATADAFTAR ISIDAFTAR TABELDAFTAR GAMBARBAB I PENDAHULUAN .....................................................
1.1 Pengertian Cendawan Endofit ..............................1.2 Manfaat Cendawan Endofit .................................Rangkuman ................................................................Tes Formatif ...............................................................
BAB II PERALATAN ISOLASI DAN PENGUJIAN CENDAWAN ENDOFIT ...........................................2.1 Peralatan ..............................................................2.2 Bahan ...................................................................Rangkuman ................................................................Tes Formatif ...............................................................
BAB III MEDIUM ISOLASI, PERTUMBUHAN DAN PENYIMPANAN CENDAWAN ENDOFIT ..............3.1 Komponen Medium .............................................3.2 Jenis Medium .......................................................Rangkuman ................................................................Tes Formatif ...............................................................
BAB IV ISOLASI DAN PEMURNIAN CENDAWAN ENDOFIT ...................................................................4.1 Persiapan Sampel .................................................4.2 Sterilisasi Permukaan ...........................................Rangkuman ................................................................Tes Formatif ...............................................................

Cendawan Endofit
vii
4242485657
606060656569698083848890108179182185
BAB V IDENTIFIKASI CENDAWAN ENDOFIT ..............5.1 Identifikasi Morfologi ............................................5.2 Identifikasi Molekuler ............................................Rangkuman ..................................................................Tes Formatif .................................................................
BAB VI METODE PENYIMPANAN CENDAWAN ENDOFIT ...................................................................
6.1 Penyimpanan Cendawan Endofit ...........................6.2 Metode Penyimpanan Cendawan Endofit ..............Rangkuman ..................................................................Tes Formatif .................................................................
BAB VII MANFAAT CENDAWAN ENDOFIT ...................7.1 Manfaat Langsung ..................................................7.2 Manfaat Tidak Langsung .......................................Rangkuman ..................................................................Tes Formatif .................................................................
BAB VIII PENUTUP ............................................................DAFTAR PUSTAKA .............................................................LAMPIRAN-LAMPIRAN ....................................................GLOSSARY ..........................................................................INDEKS ................................................................................BIODATA PENULIS .............................................................

viii
Cendawan Endofit
Daftar Primer yang umum digunakan untuk Barkoding CendawanVolume Reaksi untuk Amplifikasi PCRTingkat kemiripan Isolat Cendwan endofit asal tanaman Padi Lokal Sulawesi Selatan pada analisis BLASTN Tingkat Kemiripan isolat cendawan endofit asal tanaman cabai berdasarkan pada analisis BLASTN Absorbansi dan konsentrasi IAA dari isolat cendawan endofitKemampuan melarutkan fosfat dari isolat cendawan endofit
Tabel 1.
Tabel 2.Tabel 3.
Tabel 4.
Tabel 5.
Tabel 6.
DAFTAR TABEL

Cendawan Endofit
ix
Gambar 1.
Gambar 2.Gambar 3.Gambar 4.
Gambar 5.Gambar 6.Gambar 7.
Gambar 8.
Gambar 9.Gambar 10.
Gambar 11.Gambar 12.Gambar 13.Gambar 14.Gambar 15.Gambar 16.Gambar 17.
Timbangan digital kapasitas 200 g (a), 250 g (b), 5 kg (c)Autoklaf dengan sumber energi komporLaminar air flow buatan pabrik (a) dan Lab (b)Botol schott (a), gelas ukur (b), spatula (c) dan jarum Ose lurus (d)Bahan PDA sintetikBahan Ekstrak KentangProses pembuatan media PDA alami: kentang dipotong kecil (a), dimasak (b), ekstrak disaring ditambah dekstrose, agar-agar, aquades (c), dimasak (d), dipidah ke erlenmenyer (e), disterilisasi dengan autoklaf (f), dituang ke cawan petri (g)Sampel tanaman dari lokasi (kiri), bagian daun, batang dan akar sampel tanaman padi (kanan)Alur Sterilisasi PermukaanCendawan yang tumbuh pada permukaan batang, daun, dan akar (kiri), cendawan setelah dipindahkan ke medium PDA (kanan)Isolat cendawan endofit setelah permurnianIsolasi cendawan endofit dari tanaman obatCendawan endofit diisolasi dari tanaman cabaiCendawan berwarna putihWarna koloni abu-abuWarna kuning pada bagian tengahWarna Hitam dan hijau pinggir putih
DAFTAR GAMBAR

x
Cendawan Endofit
Gambar 18.Gambar 19.
Gambar 20.
Gambar 21.Gambar 22.Gambar 23.
Gambar 24.
Gambar 25.
Gambar 26.
Gambar 27.Gambar 28.
Gambar 29.
Gambar 30.
Gambar 31.
Tipe pertumbuhan konsentrisMakroskopis dan Mikroskopis Cendawan Endofit dari Padi Lokal EnrekangMorfologi isolat cendawan endofit dari Annona squamosaCendawan endofit dari tanaman karetMikroskopis kapang endofitik (kiri) Phoma (kanan)Hasil amplifikasi DNA cendawan endofit asal tanaman cabai menggunakan pasangan primer ITS1/ITS4Hasil amplifikasi PCR menggunakan pasangan primer NL1 dan NL4Hasil amplifikasi PCR menggunakan pasangan primer ITS1/ITS4Perubahan warna pink merupakan indikator kemampuan produksi IAAProduksi Giberelin Cendawan endofitSkema representasi dari asam organik yang dapat diproduksi oleh Mikroba Pelarut Fosfat dan digunakan untuk melarutkan bentuk fosfat anorganikZona bening yang terbentuk disekitar koloni cendawan merupakan indikator kemampuan melarutkan fosfatPerubahan warna supernata menjadi biru Merupakan indikator kemamuan melarutkan fosfatKemampuan produksi siderefor isolat cencawan endofit

Cendawan Endofit
1
BAB IPENDAHULUAN
1.1. Pengertian Cendawan EndofitSebelum membahas pengertian cendawan endofit, terlebih
dahulu harus dipahami pengertian dari endofit. Istilah endofit berasal dari Bahasa Yunani yaitu endo yang artinya dalam dan phyte yang artinya tanaman. Jadi pengertian endofit adalah semua organisme yang selama suatu periode siklus hidupnya berada dalam jaringan tanaman inang (Patil et al. 2015). Beberapa ahli memberikan definisi yang hampir sama tentang endofit. Azevedo et al., (2000), menjelaskan bahwa semua mikroorganisme yang berada dalam jaringan tanaman selama satu periode siklus hidupnya dianggap sebagai endofit. Demikian pula dengan Hilarino et al., (2011) dan Afandhi et al., (2018), mendefinisikan endofit sebagai mikroorganisme yang berada dalam jaringan tanaman selama periode tertentu dari siklus hidupnya, tanpa menimbulkan kerusakan pada tanaman.
Hal ini berarti, mikoroba endofit dapat didefinisikan sebagai mikroba dari kelompok bakteri atau cendawan yang menghabiskan seluruh siklus hidupnya dalam jaringan tanaman
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan pengertian cendawan endofit2. Membedakan mikroba endofit dengan cendawan endofit3. Menjelaskan manfaat cendawan endofit dalam bidang pertanian

2
Cendawan Endofit
tanpa menimbulkan gejala.Pengertian cendawan endofit telah didefinisikan oleh
beberapa ahli. Pengertian cendawan endofit menurut Durham, 2004; Wilia et al., 2011), cendawan endofit merupakan cendawan yang hidup dalam jaringan tanaman tanpa menunjukkan gejala. Selanjutnya menurut Clay 1988; Ariyanto, Abadi, and Djauhari (2013), yang dimaksud cendawan endofit adalah cendawan yang terdapat di dalam sistem jaringan tumbuhan, seperti daun, bunga, ranting, ataupun akar tumbuhan. Jadi berdasarkan pengertian tersebut, maka cendawan endofit dapat didefinisikan sebagai cendawan yang hidup dalam jaringan tanaman inang tanpa menimbulkan kerusakan pada tanaman inang.
1.2 Manfaat Cendawan EndofitCendawan endofit memiliki peranan penting dalam
kehidupan manusia seperti dalam bidang pertanian dan farmasi Menurut Gandjar, 1999; Hafsari and Asterina, (2013).
Peranan cendawan endofit dalam bidang pertanian diantaranya adalah sebagai pemacu pertumbuhan dan penghasil enzim. Menurut Sinaga, (2003); Ariyanto, Abadi and Djauhari, (2013), cendawan endofit ada yang berperan sebagai penghasil enzim seperti genus Aspergillus, Fusarium, dan Alternaria. Sedangkan cendawan endofit dari kelompok Trichoderma sp. dan Fusarium sp. dilaporkan oleh Amin et al., (1997) dapat meningkatkan pertumbuhan tanaman tomat.
Yadav and Yadav, (2017), membagi manfaat cendawan endofit kedalam dua kelompok yaitu manfaat langsung dan tidak langsung terhadap tanaman.a. Manfaat langsung, yaitu:
1. Produksi fitohormon (Fitohormon production)

Cendawan Endofit
3
2. Fiksasi Nitrogen (Nitrogen Fixation)3. Pelarutan Fosfat (Phosphate solubilization)4. Produksi siderophor (Siderophore production)
b. Manfaat tidak langsung, berupa:1. Bioremediasi (Bioremediation)
Bioremediasi merupakan metode untuk menghilangkan polutan dari lingkungan.
2. Fitoremediasi (Phytoremediation)Endofit membantu fitoremediasi dengan meningkatkan pertumbuhan tanaman, menurunkan fitotoksisitas logam, dan mempengaruhi translokasi dan akumulasi logam
3. Biokontrol (Biocontrol)Endofit memiliki berbagai mekanisme kontrol untuk menangkal pathogen dan hama tanaman.
4. Toleransi tanaman terhadap cekaman (Plant stress tolerance)Secara alami tumbuhan menghadapi berbagai tekanan dan bereaksi dengan memodifikasi metabolismenya sehingga menjadi toleran terhadap cekaman.Tanaman inang yang ditumbuhi mikroba endofit
mendapatkan banyak manfaat, seperti mempercepat pertumbuhan, meningkatkan daya tahan terhadap kekeringan dan serangan hama. Menurut Sukmadi, (2013), keberadaan mikroba endofit pada tanaman membantu tanaman berkompetisi di alam.
Pemanfaatan cendawan dalam bidang pertanian sudah banyak dikembangkan diantaranya pemanfaatan Tricoderma untuk meningkatkan ketahanan tanaman terhadap cekaman kekeringan. Demikian juga penggunaan cendawan dalam pengendalian hama dan penyakit. Manfaat endofit secara langsung dan tidak langsung akan dibahas lebih mendalam pada Bab VII.

4
Cendawan Endofit
Tes Formatif1. Endofit berasal dari Bahasa Yunani yaitu endo yang artinya
dalam dan pyte yang artinya…A. hidupB. ilmu
Istilah endofit berasal dari Bahasa Yunani yaitu endo yang artinya dalam, dan phyte yang artinya tanaman. Jadi pengertian endofit adalah semua organisme yang selama suatu periode siklus hidupnya berada dalam jaringan tanaman
Manfaat cendawan endofit pada tanaman diantaranya adalah:1. meningkatkan pertumbuhan vegetatif tanaman; 2. meningkatkan kemampuan tanaman untuk lebih toleran
terhadap kekeringan; 3. menghasilkan toksin yang melindungi tanaman dari
patogen
Manfaat cendawan endofit secara langsung pada tanaman adalah 1. Produksi fitohormon (Fitohormon production)2. Fiksasi Nitrogen (Nitrogen Fixation)3. Pelarutan Fosfat (Phosphate solubilization)4. Produksi siderophore (Siderophore production)
Manfaat cendawan endofit secara tidak langsung pada tanaman:1. Bioremediasi (Bioremediation)2. Fitoremediasi (Phytoremediation)3. Biokontrol (Biocontrol)4. Mekanisme Biokontrol (Biocontrol mechanism)5. Toleransi Tanaman terhadap cekaman (Plant stress
tolerance)
Rangkuman

Cendawan Endofit
5
C. tanamanD. makanan
2. Pengertian cendawan endofit adalah …A. Mikroba yang hidup dalam tanamanB. Cendawan yang hidup dalam jaringan tanamanC. Bakteri yang hidup dalam jaringan tanamanD. Mikroba yang hidup dalam akar tanaman
3. Manfaat cendawan endofit secara langsung pada tanaman adalah sebagai berikut, kecuali …A. Produksi fitohormonB. BiokontrolC. Pelarut fosfaD. Fiksasi Nitrogen
4. Manfaat cendawan endofit secara tidak langsung pada tanaman adalah
A. Fiksasi NitrogenB. Produksi SidereforeC. Pelarut fosfatD. Bioremediasi
5. Manfaat cendawan endofit bagi tanaman menurut Moore-Landecker adalah sebagai berikut, kecuali…A. Toleran terhadap kekeringanB. Meningkatkan pertumbuhan tanamanC. Memproduksi toksinD. Memproduksi siderefore
6. Bioremediasi adalah ….A. Menambah polutan B. Menghilangkan polutanC. Mengikat polutanD. Mengurai polutan

6
Cendawan Endofit
7. Endofit membantu fitoremediasi dengan cara berikut ini, kecuali…A. Mengurangi polutan B. Meningkatkan pertumbuhanC. Menurunkan fitotoksisitas logamD. Mempengaruhi translokasi dan akumulasi logam
Kunci Jawaban1. C2. B3. B4. D5. D6. B7. A
Tes Essay1. Jelaskan perbedaan mikroba endofit dan cendawan endofit.2. Jelaskan manfaat langsung dari cendawan endofit3. Moore-Landecker membedakan manfaat cendawan endofit
dalam 3 kelompok, tuliskan ketiga kelompok tersebut.
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban latihan
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 1.

Cendawan Endofit
7
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ---------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 90 – 100% = baik sekali 80 – 89% = baik 70 – 79% = cukup < 70% = kurang Bila Anda mencapai tingkat penguasaan 80% atau lebih,
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari kembali materi subunit 1 terutama bagian yang belum Anda kuasai.

8
Cendawan Endofit
2.1 Peralatan 1. Timbangan
Timbangan digunakan untuk mengetahui berat bahan yang akan digunakan dalam pembuatan medium seperti kentang, PDA, dekstrose, dan lain-lain. Jenis timbangan yang digunakan adalah timbangan digital. Kriteria timbangan yang dapat digunakan adalah: 1) timbangan analitik dengan kapasitas timbangan 200 g dan kemampuan membaca hingga 3 digit dibelakang koma atau 0,01 g (Gambar 1a); 2) timbangan dengan kapasitas 250 g dengan kemampuan membaca hingga 2 digit dibelakang koma atau 0,1 g (Gambar 1b); 3) timbangan dengan kapasitas 5 kg (Gambar 1c).
Gambar 1. Timbangan digital kapasitas 200 g (a), 250 g (b), 5 kg (c) (Dok.pribadi)
BAB IIPERALATAN DAN BAHAN ISOLASI
CENDAWAN ENDOFIT
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan peralatan dasar yang dibutuhkan dalam isolasi
cendawan endofit2. Menjelaskan bahan utama yang dibutuhkan dalam isolasi
cendawan endofit

Cendawan Endofit
9
2. AutoklafAutoklat digunakan untuk sterilisasi dengan metode uap
panas (Gambar 2). Autoklaf ada dua macam berdasarkan sumber energi yang digunakan yaitu: 1) autoklaf manual menggunakan sumber api kompor dan autoklaf otomatis menggunakan energi listrik. Kedua jenis autoklaf memiliki cara kerja yang sama dalam proses sterilisasi.
Gambar 2. Autoklaf dengan sumber energi kompor(Dokumen Pribadi)
Autoklaf dilengkapi dengan sarangan berupa panci yang terletak pada bagian dalam dari autoklaf sebagai tempat meletakkan bahan yang akan disterilisasi. Autoklaf diisi dengan air sampai batas tempat meletakkan sarangan. Sarangan diisi dengan bahan yang akan disterilkan. Kemudian tutup autoklaf diletakkan di bagian atas dan skrup diputar kencang sampai tidak bisa berputar lagi. Satu katup uap ditutup dan yang satu tetap dibiarkan terbuka. Autoklaf diletakkan diatas kompor yang sudah dinyalakan. Pada saat uap mulai keluar dari katup yang terbuka, katup tersebut ditutup agar suhu dan tekanan naik. Jika sudah mencapai tekanan 17, 5Psi atau suhu 121°C, api kompor

10
Cendawan Endofit
dikecilkan kemudian suhu dipertahankan selama 20-30 menit (Dwiyani 2015).
3. Laminar air flow Laminar air flow merupakan salah satu alat utama dalam
isolasi cendawan dan perbanyakan cendawan (Gambar 3). Semua kegiatan isolasi dan perbanyakan dilakukan di laminar air flow ini. Laminar air flow dilengkapi dengan lampu ultra violet (UV) yang dapat membunuh mikroorganisme. Selain itu terdapat lampu yang berfungsi sebagai penerang. Sebelum Laminar air flow digunakan, lampu UV dinyalakan selama 30 menit. Setelah itu, air flow dinyalakan untuk mendapatkan hembusan udara steril yang telah disaring oleh filter yang terdapat pada bagian atas dan bagian dalam laminar. Laminar siap digunakan.
Laminar yang telah selesai digunakan, segera dikosongkan dari bahan dan peralatan yang telah digunakan. Laminar kemudian disemprot dengan alkohol untuk mematikan sisa-sisa mikroba yang ada. Setelah disemprot kemudian dilap dengan tissue.
Gambar 3. Laminar air flow buatan pabrik (a) dan lab (b) (Dokumen pribadi)
4. Glassware dan Peralatan kecil lainnyaGlassware adalah peralatan kecil yang terbuat dari bahan

Cendawan Endofit
11
gelas seperti, botol schott, gelas ukur, cawan petri, objek glass, erlenmeyer dan lain-lain (Gambar 4).
Gambar 4. Botos schott (a), gelas ukur (b), spatula (c) dan jarum Ose lurus (d) (Dokumen Pribadi)
Peralatan kecil lainnya terdiri dari spatula, pinset, ose, dan lain lain yang terbuat dari bahan logam (stainless steel) (Gambar 4). Spatula merupakan pengaduk atau digunakan untuk mengambil bahan berupa serbuk. Pinset digunakan untuk menjepit sampel tanaman saat sterilisasi permukaan. Ose gunanya untuk mengambil misellum cendawan dan meletakkannya pada medium pertumbuhan.
2.2 Bahan 1. Potato Dextrose Agar
Potato Dextrose Agar (PDA) merupakan bahan sintetik untuk pembuatan medium PDA yang akan digunakan untuk isolasi dan perbanyakan cendawan. PDA dijual dalam bentuk serbuk (Gambar 5a). Harganya lumayan mahal, namun dapat diganti dengan membuat ekstrak dari kentang (Gambar. 5b)

12
Cendawan Endofit
Gambar 5. Bahan PDA sintetik(Dokumen pribadi)
Kelemahan penggunaan medium PDA sintetik adalah harganya cukup mahal, hal ini dapat diatasi dengan menggunakan bahan alami berupa ekstrak kentang yang diperoleh dari kentang segar.
Gambar 6. Bahan Ekstrak Kentang (Dokumen pribadi)

Cendawan Endofit
13
3. DekstroseDektrose atau glukosa adalah sejenis gula termasuk
monosakarida dengan rumurs molekul C6H12O6, banyak digunakan dalam industry permen, ice cream, sirup serta parbir-pabrik farmasi (Dwiyani 2015). Gula juga digunakan dalam pembuatan media untuk kultur jaringan sebagai sumber karbohidra untuk respirasi karena tanaman kultur bersifat heterotrof. Penambahan dekstrose pada pembuatan media untuk pertumbuhan mikroba teruma cendawan berfungsi sebagai sumber (Suryani, Santoso, and Juffrie 2010)energi bagi cendawan.
4. Agar-AgarAgar-agar terbuat dari ekstrak rumput laut dengan
karakteristik memiliki daya ikat terhadap air dan memadat membetuk gel(Suryani, Santoso, and Juffrie 2010). Fungsi agar-agar pada pembuatan medium padat adalah sebagai bahan pemadat medium. Oleh karena itu pada pembuatan medium cair tidak dilakukan penambahan agar-agar.
5. AquadesAguades adalah air hasil destilasi/penyulingan yang sama
dengan air murni atau H2O, karena H2O hampir tidak mengandung mineral (Suryani, Santoso, and Juffrie 2010).

14
Cendawan Endofit
Tes Formatif1. Salah satu alat yang dibutuhkan dalam isolasi cendawan adalah
autoklaf. Fungsi autoklaf adalah....a. menimbangb. sterilisasic. mengambil bahand. menjepit bahan
2. Laminar air flow memiliki dua jenis lampu. Salah satunya adalah lampu UV, yang berfungsi untuk….a. peneranganb. membunuh mikroorganismec. pemanasand. penguraian
3. Laminar air flow memiliki dua jenis lampu. Salah satunya adalah lampu neon yang berfungsi …
Rangkuman
Peralatan yang dibutuhkan untuk isolasi cendawan end-ofit diataranya adalah: 1) timbangan; 2) autoklaf; 3) laminar air flow; 4) Glassware.
Timbangan digunakan untuk mengukur bahan yang akan digunakan. Autoklaf untuk sterilisasi bahan dan pera-latan. Laminar air flow adalah alat untuk semua kegiatan mu-lai dari membuat medium, isolasi, dan perbanyakan cendawan endofit. Glassware adalah peralatan kecil yang terbuat dari ba-han gelas seperti botol schott, gelas ukur, erlenmeyer, cawan petri. Peralatan kecil seperti: spatula, pinset dan jarum ose.
Bahan yang dibutuhkan untuk medium isolasi cendawan adalah: 1) Potato Dextrose Agar; 2) kentang; 3) dekstrose; 4) agar-agar; 5) aquades.

Cendawan Endofit
15
a. peneranganb. sterilisasic. pemanasand. penguraian
4. Berikut ini adalah bahan untuk pembuatan media PDA alami, kecuali …a. Aquadesb. Alkoholc. Agar-agard. Kentang
5. Potato Dekstrose Agar dapat dibuat dari …a. Ekstrak tebub. Ekstrak kentangc. Ekstrak worteld. Eksrak kurma
6. Glassware adalah peralatan yang terbuat dari…a. stainless steelb. kertasc. gelasd. tanah liat
7. Berikut ini termasuk Glassware, kecuali…a. Erlenmeyerb. Spatulac. Gelas ukurd. Botos schott
8. Alat yang digunakan untuk menakar bahan yang akan digunakan adalah…a. Autoklafb. Spatulac. Gelas ukur

16
Cendawan Endofit
d. Timbangan9. Laminar air flow mempunyai dua buah saringan yang berada di
bagian atas dan dalam laminar yang berfungsi untuk …a. membunuh mikoroorganismeb. menerangi laminarc. mengalirkan udara steril ke dalam laminard. memanaskan laminar
10. Berikut ini peralatan yang terbuat dari bahan stainless steel, kecuali…
a. Pinset b. Spatulac. Gelas ukurd. Ose
Kunci Jawaban1. b2. b3. a4. b5. b6. c7. b8. d9. c10. c
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban latihan
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui

Cendawan Endofit
17
tingkat penguasaan Anda terhadap materi subunit 2.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ----------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 91 – 100% = baik sekali 81 – 89% = baik 71 – 79% = cukup < 70% = kurang Bila Anda mencapai tingkat penguasaan 80% atau lebih,
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari kembali materi subunit 1 terutama bagian yang belum Anda kuasai.

18
Cendawan Endofit
3.1 Komponen MediumMedium merupakan suatu bahan yang terdiri atas campuran
zat makanan yang berfungsi sebagai tempat tumbuh mikroba (Octavia and Wantini 2017).
Persyaratan medium untuk pertumbuhan cendawan endofit pada dasarnya sama dengan persyaratan untuk pertumbuhan mikroba antara lain: pH media harus sesuai, steril dan tidak mengandung zat-zat penghambat serta mengandung semua nutrisi yang mudah digunakan mikroorganisme (Jutono, 1980;(Octavia and Wantini 2017). Medium Potato Dextrose Agar (PDA) cocok dan mendukung pertumbuhan cendawan dan memiliki pH 4.5 sampai 5.5 sehingga menghambat pertumbuhan bakteri yang membutuhkan pH netral (Cappucino & Sherman, 2014; (Jamilatun, Azzahra, and Aminah 2020).
Mikroorganisme termasuk cendawan membutuhkan nutrisi untuk pertumbuhan. Menurut Cappuino, 2014; (Octavia and Wantini 2017), nutrisi yang dibutuhkan untuk pertumbuhan mikroorganisme meliputi: karbon, nitrogen, unsur non logam
BAB IIIMEDIUM ISOLASI DAN PERTUMBUHAN
CENDAWAN ENDOFIT
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan pengertian medium dan komposisi medium 2. Menjelaskan jenis medium dan peruntukannya

Cendawan Endofit
19
seperti sulfur dan fosfor, unsur logam seperti Ca, Zn, Na, K, Cu, Mn, Mg dan Fe, vitamin, air dan energi.
3.2 Jenis MediumMedium berdasarkan bahan penyusunnya dibedakan atas:
1. Potato Dextrose Agar (PDA)Komposisi medium PDA instan yang dibuat oleh pabrik atau
perusahaan dalam bentuk bahan siap pakai mengandung ekstrak kentang, dextrose dan agar. Kentang mengandung karbohidrat, vitamin, dan mikronutrien lain yang dapat dimanfaatkan oleh cendawan, dextrose sebagai sumber gula dan energi dan agar sebagai bahan pemadat (Octavia and Wantini 2017).
Pembuatan Medium PDABahan dan cara pembuatan medium PDA menggunakan
bahan PDA sintetik dijelaskan oleh (Jamilatun, Azzahra, and Aminah 2020), sebagai berikut:a. Medium PDA instan
Bahan:• PDA : 39 GR• Aquades : 1000 mlCara membuat:1. Menimbang bahan PDA sintetik dan memasukkannya ke
dalam Erlenmeyer2. Menambahkan aquades dan mengaduk hingga bahan
tercampur3. Memanaskan bahan hingga mendidih dan homogen4. Mengukur pH (4.5-5.5), jika pH media kurang asam,
menambahkan asam tartat 10%5. Menutup erlenmeyer dengan kapas, kasa dan kerta kopi

20
Cendawan Endofit
6. Mensterilkan bahan dalam autoklaf pada suhu 121°C selama 15 dengan tekanan 2 atm
7. Menambahkan kloramfenikol 20 mL secara aseptis di dalam laminar air flow.
8. Menuang bahan ke dalam cawan petri dan biarkan memadat
9. Medium PDA siap digunakan
b. Medium PDA alamiBahan untuk pembuatan medium PDA alami adalah:1. Kentang : 200 g 2. Dextrose :20 g 3. Agar : 20 g4. Aquades : 1 LCara pembuatan medium PDA alami sebagaimana dijelaskan
oleh Gunawan et.al., 2006; Budiprakoso, 2010, sebagai berikut:1. Kentang dikupas sampai bersih dan dicuci kemudian
dipotong-potong dengan ukuran 2 cm x 2 cm ( Gambar 7a)
2. Potongan kentang dan air suling (aquades) dimasak (Gambar 7b), setelah setengah jam diangkat dan disaring untuk diambil ekstraknya (Gambar 7c)
3. Ekstrak kentang tersebut kemudian dimasak kembali dan ditambahkan agar-agar dan dekstrose (Gambar 7d)
4. Setelah mendidih, campuran tersebut kemudian diangkat dan dituangkan ke dalam erlemeyer/botol schott (Gambar 7e). Kemudian PDA tersebut dimasukkan ke dalam autoklaf untuk disterilisasi dengan menggunakan uap air jenuh bertekanan 15 lb/in 2 selama 15 menit pada suhu 121oC (Gambar 7f)

Cendawan Endofit
21
5. Setelah uap dalam autoklap hilang, agar-agar dituang ke cawan petri dan ditutup dengan plastik wrap dan dibiarkan hingga dingin dan mengeras dalam laminar air flow (Gambar 7g).
6. Media yang sudah dingin dan padat siap disimpan dalam tempat penyimpanan. Setiap hari diperiksa. Jika ada medium yang kontaminasi, segara dipisahkan. Media yang tidak kontaminasi dapat digunakan untuk isolasi cendawan endofit.
2. Carrot Sukrose Agar (CSA) Bahan dan cara membuat medium CSA sebagaimana
dijelaskan oleh (Jamilatun, Azzahra, and Aminah 2020), adalah sebagai berikut:
Gambar 7. Proses pembuatan media PDA alami: kentang dipotong kecil (a), dimasak (b), ekstrak disaring ditambah dekstrose, agar-agar, aquades (c), dimasak (d), dipidah ke
erlenmenyer (e), disterilisasi dengan autoklaf (f), dituang ke cawan petri (g). (Dokumen pribadi)

22
Cendawan Endofit
Bahan:• wortel : 200 g• Sukrose : 20 g• Agar : 15 g • Aquades : 1500 mLCara membuat:1. Mencuci wortel dengan dengan air bersih2. Memotong wortel menjadi bentuk dadu dengan ukuran
sisi lebih kurang 0.5 cm3. Mencuci potongan wortel dengan aquades4. Menghaluskan wortel dengan menggunakan mortar
tanpa diberi air5. Masukkan wotel yang telah halus ke dalam erlenmeyer6. Menambahkan aquades 500 mL7. Memanaskan wortel sampai mendidik8. Menyaring rebusan wortel 9. Menambahkan sukrose dan agar-agar10. Menambahkan aquades hingga volume 1000 mL11. Memanaskan hingga mendidih dan homogen12. Mengukur pH medium13. Mensterilkan medium dengan autoclave14. Menambahkan kloramfenikol 15. Menuang ke dalam cawang petri dalam kondisi aseptis
dalam laminar air flow16. Media siap digunakan setelah dingin dan memadat
3. Dichloran Rose Bengal Chloramphenicol Agar (DRBC) Dichloran Rose Bengal Chloramphenicol Agar (DRBC)
adalah media yang dikembangkan oleh King et al. (1979) (Idriati, Priyanto, and Triwibowo 2010) dan merupakan modifikasi dari

Cendawan Endofit
23
Rose bengal chloramphenicol (RBC) Agar dari Javis (1973). Perbedaan dengan RBC, media DRBC mengandung dichloran (0.002 g/L) dan rose Bengal konsentrasi 0.025 g/L (Idriati, Priyanto, and Triwibowo 2010).
Bahan dan cara pembuatan medium Carboxymethylcellulose agar (CMC), Glucose Ammonium Nitrate Agar (GAN), Malt Extract Agar (MEA, Media Czapex Agar (CZA), Oat meal Agar (OA), Miru atau low carbon agar (LCA), Miru atau low carbon agar (LCA) dijelaskan dalam Handbook of Industrial Mycology, Volume 22, Tahun 2005, sebagai berikut:
3. Carboxymethylcellulose agar (CMC)Bahan:• CMC : 2 g• MgSO4.7H2O : 0.05 g• Na2HP4 : 0,5 g• NaCl : 0,23 g• Yeast : 0,2 g• congo red : 0.5%• Aguadest : 100 mL
4. Glucose Ammonium Nitrate Agar (GAN) Bahan:• Glukose : 10 g• NH4NO3 : 1 g• KH2PO4 : 1 g• MgSO4.7H2O : 0.5 g• Rose Bengal : 0.03 g• Agar : 20 g• Aguades : 1000 mL

24
Cendawan Endofit
5. Malt Extract Agar (MEA)Bahan:• Malt extract : 20 g• Agar : 20 g• Air : 1000 mL
6. Czapex Agar (CZA) Bahan:• Sukrose : 30 g• NaNO3 : 2 g
7. Oat meal Agar (OA)Bahan:• Rolled oats : 30 G• Agar : 20 g• Air : 1000 mL• MgSO4.7H2O : 0.5 g• KCl : 0.5 g• FeSO4.7H2O : 0.01 g• FeSO4.7H2O : 0.001 g• Agar : 15 g• Aguades : 1000 mL
8. Miru atau low carbon agar (LCA) Bahan:• Glukose : 1 g• KH2PO4 : 1 g• MgSO4.7H2O : 0.2 g• KCl : 0.2 g• NaNO3 : 2 g

Cendawan Endofit
25
• Yeast extract : 0.2 g• Agar : 15 g• Aquades : 1000 mL
9. Cornmeal Agar (CMA) Bahan:• Yellow cornmeal : 40 g• Agar : 15-20 g• Aquades : 1000 mL Medium berdasarkan tujuan penggunaannya oleh (Ilyas.
2007) sebagai berikut: 1. medium untuk isolasi2. medium untuk identifikasi; 3. medium untuk penyimpananPenjelasan masing jenis medium adalah sebagai berikut:
1. Medium untuk Isolasi CendawanMedium untuk isolasi cendawan berfungsi meransang
pertumbuhan hifa dan perkecambahan spora. Medium tersebut hanya mengandung sedikit nutrient dan biasanya dijual bebas, contoh : Dichloran Rose Gengal Chloramphenicol Agar (DRBC), dan Glucose Ammonium Nitrate Agar (GAN), Water Agar (WA), miura atau low carbon agar ( LCA).
2. Medium untuk identifikasiMedium untuk identifikasi yang umum digunakan adalah
Potato Dextrose Agar (PDA), Malt Extract Agar (MEA), Miura Agar/Low Carbon Agar (LCA), Czapex Agar (CZA), dan Oat meal Agar (OA).

26
Cendawan Endofit
3. Medium untuk penyimpananMedium untuk penyimpanan digunakan menyimpan
isolat selama mungkin, dengan tujuan agar tingkat viabilitas cendawan tetap terjaga selama penyimpanan. Medium yang umum digunakan adalah Carboxymethyl cellulose agar (CMC), Cornmeal Agar (CMA) dan Potato Carrot Agar (PCA).
Medium berdasarkan bentuknya ada dua yaitu:
Medium PDA dan PDB terbuat dari bahan yang sama, hanya saja pada pembuatan medium PDB tidak digunakan agar-agar sehingga medium PDB berbentuk cair.
a.b.
Medium Padat, berbentuk padat dan menggunakan agar-agar sebagai bahan pemadat, contoh medium padat adalah PDAMedium Cair, berbentuk cait dan dalam pembuatannya tidak menggunakan agar-agar, contoh medium cair adalah PDB

Cendawan Endofit
27
Tes Formatif1. Suatu bahan yang teridri atas campuran zat makanan yang berfungsi sebagai tempat tumbuh mikroba merupakan pengertian dari …
a. menimbangb. sterilisasic. mengambil bahand. medium
Rangkuman
Medium merupakan bahan yang teridiri atas campuran zat makanan yang berfungsi sebagai tempat tumbuh mikroba sehingga harus memenuhi beberapa persyaratan diantaranya adalah pH media harus sesuai, steril dan tidak mengandung zat-zat penghambat serta mengandung nutrisi yang dibutuh-kan mikroba.
Nutrisi yang dibutuhkan untuk pertumbuhan mikoror-ganisme meliputi: karbon, nitrogen, unsur non logam dan log-am, vitamin, air dan energi.
Jenis medium berdasarkan bahan penyusunnya dibeda-kan atas: Potato Dextrose Agar (PDA), Carboxymethylcellu-lose agar (CMC), Glucose Ammonium Nitrate Agar (GAN), Malt Extract Agar (MEA), Czapex Agar (CZA), Oat Meal Agar (OA), Miru atau Low Carbon Agar (LCA) dan Cornmeal Agar (CMA).
Medium berdasarkan peruntukannya dibedakan atas: medium untuk isolasi, medium untuk identifikasi dan medium untuk penyimpanan.
Medium yang digunakan untuk isolasi adalah DRBC, GAN, WA, LCA, medium untuk identifikasi adalah PDA, MEA, LCA, CZA dan OA, sedangkan medium untuk penyim-panan adalah CMC, CMA dan PCA.

28
Cendawan Endofit
2. Medium untuk pertumbuhan mikroba harus memenuhi syarat sebagai berikut, kecuali …
a. sterilb. pH sesuaic. mengandung nutrisid. mengandung zat penghambat
3. Nutrisi yang dibutuhkan untuk pertumbuhan mikroorganisme adalah sebagai berikut, kecuali…
a. Vitaminb. Airc. Lemakd. Karbon
4. Berikut ini adalah bahan untuk pembuatan media PDA alami, kecuali …
a. Aquadesb. Alkoholc. Agar-agard. Kentang
5. Potato Dekstrose Agar dapat dibuat dari …a. Ekstrak tebub. Ekstrak kentangc. Ekstrak worteld. Eksrak kurma
6. Medium yang merupakan modifikasi dari RBC adalah…a. DRBCb. GANc. MEAd. LCA
7. Medium yang dapat digunakan untuk identifikasi cendawan endofit adalah

Cendawan Endofit
29
a. PCAb. MEAc. CMd. WA
8. Medium yang dapat digunakan untuk isolasi dan identifikasi adalah…
a. Potato Carrot Agarb. Low Carbon Agarc. Malt Extract Agard. Oat Meal Agar
9. Medium PDA digunakan untuk …a. isolasib. identifikasic. penyimpanand. isolasi dan identifikasi
10. Medium yang bahannya mengadung wortel adalah…a. PCAb. PDAc. MEAd. LCA
Kunci Jawaban1. d2. d3. c4. b5. b6. a7. b

30
Cendawan Endofit
8. b9. b10.a
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban latihan
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 3.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ----------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 92 – 100% = baik sekali 82 – 89% = baik 72 – 79% = cukup < 70% = kurang Bila Anda mencapai tingkat penguasaan 80% atau lebih,
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari kembali materi subunit 1 terutama bagian yang belum Anda kuasai.

Cendawan Endofit
31
4. 1 Persiapan SampelCendawan dapat diisolasi dari daerah disekitar perakaran
tanaman (rhizosfir), daun (filosfir) dan jaringan tanaman (endofit). Isolasi cendawan endofit dilakukan pada jaringan tanaman sehat dan diambil dari bagian daun, akar, dan batang.
Cendawan endofit paling banyak ditemukan pada akar, dibandingkan bagian batang dan cabang. Hal ini disebabkan karena jaringan akar tanaman secara morfologi, fisik, dan kimia menyediakan habitat bagi beragam komunitas mikroorganisme, termasuk bagi cendawan endofit (Ramdan et al. 2014).
Sebelum melakukan isolasi cendawan pada jaringan tanaman perlu dilakukan persipan sampel tanaman. Sampel tanaman yang diambil dari lokasi pertanaman disimpan dalam kantong platik dan diberi label nama tanaman, tanggal dan lokasi pengambilan sampel.
Sampel tanaman dibersihkan dari sisa-sisa tanah yang melekat dengan mencuci sampel tanaman di bawah air mengalir sehingga sisa tanah dan debu yang menempel pada tanaman ikut
BAB IVISOLASI DAN PEMURNIAN
CENDAWAN ENDOFIT
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan teknik persiapan sampel tanaman2. Menjelaskan teknik sterilisasi permukaan3. Menjelaskan teknik isolasi dan permurnian isolate cendawan
endofit

32
Cendawan Endofit
terbuang.Sampel tanaman yang telah bersih dikering anginkan untuk
menghilangkan sisa air yang melekat pada bagian tanaman. Selanjutnya sampel dipisahkan berdasarkan bagian-bagiannya yaitu akar, batang dan daun sebelum dilakukan sterilisasi permukaan.
Gambar 8. Sampel tanaman dari lokasi (kiri), bagian daun, batang dan akar sampel padi (kanan) (dokumen pribadi)
4. 2 Sterilisasi PermukaanSterilisasi permukaan merupakan tahap awal dalam kegiatan
isolasi cendawan dengan tujuan untuk membersikan permukaan tanaman dari mikroba dan memastikan cendawan yang diisolasi berasal bagian dalam jaringan tanaman.
Sterilisasi permukaan dapat menggunakan alkohol 70% dikombinasikan beberapa bahan kimia lain seperti natrium hipoklorit (NaOCl), H202 (3%) dan KMnO4 (2%) (Zang et al, 2006; Agusta, 2009). Ada beberapa metode yang dapat digunakan dalam sterilisasi permukaan sampel untuk isolasi cendawaan endofit. Metode sterilisasi permukaan dikemukan oleh beberapa ahli sebagai berikut:

Cendawan Endofit
33
1) Metode sterilisasi Permukaan menurut Pimentel et al. 2016; Russo et al., 2016)
Metode ini menggunakan alkohol 70%, Natrium hipoklorit 3%, dan aquades steril. Tahapan sterilisasi adalah sebagai berikut: 1) Sampel tanaman yang diambil dari bagian daun, batang, dan akar dicuci dua kali dalam air suling; 2) dilakukan perendaman selama 1 menit dalam 70% (v/v) etanol, 3) Bahan direndam lagi selama 4 menit dalam natrium hipoklorit (3%, v/v tersedia klorin) dan, 5) dicuci tiga kali dalam aquadest steril.
2) Metode sterilisasi permukaan menurut Fisher et al., 1994; Amirita et al., 2012)
Bahan yang digunakan dalam metode ini adalah alkohol 75%, dan Natrium hipoklorit 4%. Tahapan sterilisasi meliputi: 1) Sampel tanaman direndam pertama dalam 75% etanol selama 60 detik; 2) kemudian direndam lagi dalam Natrium hipoklorit 4% selama 180 detik; 3) direndam kembali dalam tanol 75% selama 30 detik.
3) Metode Sterilisasi permukaan menurut Dobranic et al., (1995; (Russo et al. 2016)
Metode ini menggunakan etanol 70%, Natrium hipoklorida 4%, dan air suling. Tahapan sterilisasi: 1) Sampel tanaman dicelupkan dalam etanol 70% selama 5 detik; 3) direndam dalam natrium hipoklorida 4% selama 90 detik; 4) akhirnya dibilas dalam air suling steril untuk 10 detik.
4) Metode sterilisasi permukaan menurut (Yunianto et al., (2012); Akmalasari et al., 2013).

34
Cendawan Endofit
Metode ini menggunakan alkohol 70%, NaOCl, dan aquades steril. Tahapan sterilisasi, yaitu: 1) Sampel tanaman dimasukkan ke dalam larutan alkohol 70% selama 1 menit; 2) direndam dalam NaOCl selama 5 menit; 3) dimasukkan dalam alkohol 70 % selama 30 detik; 4) dibilas dengan aquades steril 5 detik dengan tiga kali ulangan. Bahan yang disterilisasi kemudian dikeringkan dengan tissue steri l ± 1 menit.
5) Metode sterilisasi permukaan menurut Ezra et al., (2004) ;(Qadri et al. 2013)
Metode ini menggunakan etanol 90% dan Sodium hypochlorite 1%. Tahapan sterilisasi: 1) Sampel tanaman dimasukkan ke dalam 1% sodium hypochlorite; 2) dibilas dengan air suling; 3) dibilas dengan etanol 90% ethanol.
Setelah dilakukan sterilisasi permukaan selanjutnya bagian permukaan akar, batang, dan daun disayat. Bagian dalam permukaan daun, batang, dan akar diletakkan pada medium isolasi cendawan dan diinkubasi selama (2-14) x 24 jam. Cendawan endofit yang tumbuh dimurnikan pada medium Potato Dextrose Agar/PDA (Yunianto et al., 2012; Akmalasari et al., (2013).
Gambar 9. Alur Sterilisasi Permukaan

Cendawan Endofit
35
4.3 Isolasi Cendawan EndofitIsolat cendawan endofit yang tumbuh pada permukaan
jaringan tanaman dipindahkan ke medium PDA. Isolat cendawan yang tumbuh pada medium PDA umumnya terdiri atas beberapa jenis, ditandai dengan muncul beberapa warna miselia cendawan. Untuk mendapatkan isolat cendawan murni dari kegiatan isolasi yang telah dilakukan, maka perlu dilakukan pemurnian atau purifikasi.
Gambar 10. Cendawan yang tumbuh pada permukaan batang, daun, dan akar (kiri), cendawan setelah dipindahkan ke medium PDA (kanan) (dokumen pribadi)
4.4 Pemurnian Isolat Cendawan EndofitPemurnian atau purifikasi dilakukan pada setiap koloni
cendawan yang dianggap berbeda. Perbedaan koloni berdasarkan morfologi makroskopis, yaitu meliputi warna dan bentuk koloni. Hasil purifikasi akan didapatkan isolat murni atau spora tunggal.
Cendawan endofit yang telah berhasil diisolasi dari berbagai jenis tanaman seperti tanaman pangan, perkebunan, hortikultura (sayuran, buah-buahan, tanaman hias dan tanaman obat-obatan), dan tananaman kehutanan (sengon, mangrove, ebony). Contoh cendawan endofit yang telah diisolasi dari padi lokal Sulawesi Selatan (Gambar 11), cendawan endofit dari tanaman obat

36
Cendawan Endofit
(Gambar 12), cendawan endofit dari cabai (Gambar 13).
Gambar 11. Isolat cendawan endofit setelah permurnian(Dokumen pribadi)
Cendawan endofit telah diisolasi oleh (G, A, and Kannan 2015) pada jaringan daun dan batang tiga jenis tanaman obat yaitu Terminalia arjuna, Catharanthus roseus, and Azadirachta indica
Gambar 12. Isolasi cendawan endofit dari tanaman obat (G, A, and Kannan 2015).

Cendawan Endofit
37
Gambar 13. Cendawan endofit diisolasi dari tanaman cabai (Legiastuti and Aminingsih 2012).
Rangkuman
Isolasi cendawan endofit melalui tiga tahapan yaitu: 1) per-siapan sampel; 2) sterilisasi; 3) isolasi; 4) pemurnian/purifikasi
Persiapan sampel diawali dengan pembersihan sampel dari sisa tanah yang melekat pada tanaman dan pemisahan bagian-bagian sampel tanaman menjadi akar, batang dan daun. Masing-masing ba-gian dipotong kecil-kecil sebelum dilakukan sterilisasi permukaan.
Sterilisasi permukaan sampel tanaman untuk isolasi cendawan endofit menggunakan alkohol, natrium hipoklorit, dan aguades. Sterilisasi permukaan dilakukan secara bertahap.
Isolasi cendawan endofit dilakukan dengan meletakkan seti-ap potongan yang telah disterilisasi permukaan pada medium PDA dan diinkubasi pada suhu kamar hingga permukaan sampel tumbuh cendawan. Setiap jenis cendawan yang tumbuh pada permukaan sampel tanaman dipindahkan ke medium PDA yang baru.
Pemurnian dilakukan dengan memindahkan setiap isolat yang berbeda ke medium PDA yang baru sampai diperoleh isolat murni atau spora tunggal.

38
Cendawan Endofit
Tes Formatif1. Tahapan dalam isolasi cendawan ENDOFIT adalah sebagai
berikut, kecuali …A. Persiapan sampelB. Sterilisasi permukaanC. PurifikasiD. Pemanasan
2. Bahan yang biasa digunakan untuk sterilisasi permukaan adalah, kecuali… A. Air mineralB. AlkoholC. AquadesD. Natrium hipoklorit
3. Pembersihan sampel tanaman di bawah air mengalir merupa-kan tahapan …
A. Penyimpanan cendawanB. Perbanyakan cendawanC. Persiapan sampelD. Isolasi cendawan
4. Pemindahan setiap isolate ke medium PDA yang baru meru-pakan kegiatan pada tahap ….A. Persiapan sampelB. Pemurnian C. Sterilisasi permukaan D. Identifikasi
5. Pembilasan sampel dengan alkohol pada tahap sterilisasi permukaan dilakukan sebanyak…A. satu kali B. tiga kaliC. dua kali

Cendawan Endofit
39
D. empat kali6. Pembilasan sampel dengan air steril dilakukan tahap…
A. awalB. akhirC. awal dan akhirD. setiap tahap
7. Pembilasan dengan natrium hipokloris dilakukan setelahpembilasan dengan …A. AlkoholB. Air SterilC. Alkohol dan Air sterilD. Air steril dan Alkohol
8. Kegiatan pemisahan akar, batang dan daun sampel tanamandilakukan pada tahap…A. PurifikasiB. PemurnianC. Persiapan sampelD. Sterilisasi permukaan
9. Metode sterilisasi permukaan menurut Ezra menggunakan etanol dengan konsentrasi ….
A. 70%B. 80%C. 90%D. 95%
10. Cendawan yang diisolasi dari daerah disekitar perakarantanaman disebut cendawan…A. filosfirB. endofitC. rhizosfirD. patogen

40
Cendawan Endofit
Kunci Jawaban1. D 2. B3. C4. B5. C6. B7. A8. C9. C10. C
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban latihan
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 4.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ----------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 90 – 100% = baik sekali 80 – 89% = baik 70 – 79% = cukup < 70% = kurang
Bila Anda mencapai tingkat penguasaan 80% atau lebih,

Cendawan Endofit
41
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari kembali materi subunit 1 terutama bagian yang belum Anda kuasai.

42
Cendawan Endofit
5.1 Identifikasi MorfologiIdentifikasi cendawan penting dilakukan karena cendawan
di alam sangat melimpah dan beragaman jenisnya. Identifikasi bertujuan untuk mengetahui jenis cendawan dan membedakannyan dengan jenis yang lain. Identifikasi dapat dilakukan secara konvensional dan molekuler. Identifikasi secara konvensional berdasarkan pada karakter morfologi.
Identifikasi secara morfologi telah berhasil dilakukan oleh beberapa peneliti seperti: Gazis & Chaverri, (2010), menjelaskan bahwa identifikasi secara morfologi berdasarkan pada permukaan koloni, warna, struktur misellium, jenis anamorf, konidiomata, konidia, dan konidiofor (ukuran, warna, bentuk, ornamen, dll.), sedangkan Gandjar, (1999); Hafsari dan Asterina, (2013); (Syamsia 2016), mengidentifikasi cendawan secara morfologi dengan mengamati karakter makroskopi dan mikroskopi. Karakter makroskopi meliputi meliputi: warna dan permukaan koloni; garis-garis radial dari pusat koloni ke arah tepi koloni; dan lingkaran-lingkaran konsentris. Karakter mikroskopi meliputi:
BAB VIDENTIFIKASI CENDAWAN ENDOFIT
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan identifikasi cendawan endofit secara morfologi 2. Menjelaskan identifikasi cendawan endofit secara molekuler

Cendawan Endofit
43
bentuk hifa; ada atau tidaknya rhizoid; bentuk sel reproduksi seksualnya dan aseksualnya.
a. Karakter MakroskopisKarakter makroskopi yang umum diamati pada identifikasi
secara morfologi diantaranya adalah: 1. Warna dan permukaan koloni
Warna permukaan koloni cendawan endofit bervariasi, ada yang berwana putih (Gambar 14), ada juga yang berwana putih saat awal petumbuhan namun perlahan lahan berubah menjadi abu-abu (Gambar 15), kuning pinggir putih (Gambar 17), hitam (Gambar 17a), hijau pinggir putih (Gambar 17b).
Gambar 14. Cendawan berwarna putih (kiri) (Syamsia, 2016) dan Kanan (Syamsia Syamsia et al. 2020)
Gambar 15. Warna koloni abu-abu (Legiastuti and Aminingsih 2012)

44
Cendawan Endofit
Gambar 16. Warna kuning pada bagian tengah (Legiastuti and Aminingsih 2012) dan foto koleksi pribadi (kiri)
Gambar 17. Warna Hitam (Legiastuti and Aminingsih 2012) dan hijau pinggir putih (Syamsia, 2016).
2. Tipe pertumbuhanTipe pertumbuhan cendawan ada 2 yaitu tipe pertumbuhan
konsentris dan menyebar. Tipe konsentris pertumbuhannya secara terarur ke arah tepi, sedangkan tipe menyebar pertumbuhannya tidak beraturan. Contoh tipe pertumbuhan konsentris pada Gambar 18.
Gambar 18. Tipe pertumbuhan konsentris (Legiastuti and Aminingsih 2012)

Cendawan Endofit
45
b. Karakter MikroskopisPengamatan mikroskopis tidak dapat dilakukan dengan
kasat mata, perlu menggunakan preparat dan diamati di bawah mikroskop. Langkah pertama untuk pengamatan mikroskopis adalah pembuatan preparat. Teknik Pembuatan preparat melalui beberapa tahapan sebagaimana dijelaskan oleh Ariyanto, Abadi, and Djauhari (2013), sebagai berikut:
1. Menyiapkan object glass, cover glass, dan tissue, 2. Cendawan yang telah diisolasi pada medium PDA diambil
dengan jarum ose dan ditutup dengan menggunakan cover glass.
3. Preparat diletakkan pada wadah yang telah diberi alas tissue lembab dan inkubasi selama 2-3 hari.
4. Preparat kemudian diamati di bawah mikroskop untuk mengetahui karakter mikroskopis cendawan yang diamati
Berikut ini beberapa contoh hasil identifikasi secara morfologi dari cendawan endofit. Hasil identifikasi cendawan endofit yang diisolasi dari tanaman padi lokal Enrekang berdasarkan pengamatan karakter makroskopis dan mikroskopis morfologi teridentifikasi sebagai cendawan Aspergillus sp1 Aspergillus sp2, Aspergillus candidus, dan Acremonium sp (Syamsia, et al., 2016). Karakter morfologi masing-masing cendawan endofit dijelaskan sebagai berikut: 1. Aspergillus sp1
Warna koloni bagian atas putih, bagian bawah krem, permukaan berbulu halus, konidia berbentuk bulat oval, warna konidia hialin, permukaan konidiofor halus berwarna coklat muda, phialid berbentuk tegak, hifa bersepta.

46
Cendawan Endofit
2. Aspergillus sp2 Warna bagian atas koloni hijau pinggir putih, permukaan bawah krem, permukaan koloni berbulu halus dengan tipe pertumbuhan radial. Konidia berbentuk bulat dan hialin, Permukaan konidiofr halus dan berwarna coklat muda, phialid berbentuk tegak.
3. Aspergillus candidus Warna koloni bagian atas dan bawah putih, permukaan koloni halus dan bergelombang, dan tipe pertumbuhan radial. Konidia berbentuk elips berukuran Elips berukuran 2.2 – 3.4 µm dengan warna hialin. Permukaan konidiofor halus dan berwarna krem, phialid berbentuk tegak, berukuran panjang 5.6 -10 µm dan lebar 2.2 – 3.4 µm, dan hifa hialin.
4. Acremonium sp Warna permukaan atas koloni putih, permukaan bahwa krem/coklat muda, tekstur permukaan koloni kasar, konidia berbentuk bulat silinder dan berwarna hialin. Permukaan konidiofor halus dan warna transparan. Philaid berbentuk tegak dan berlendir di setiap puncak phialid. Serta hifa hialin.
Gambar 19. Makroskopis dan Mikroskopis Cendawan Endofit dari Padi Lokal Enrekang (Syamsia 2016)

Cendawan Endofit
47
Identifikasi morfologi cendawan endofit dari tanaman Annona squamosa (Yunianto et. al., 2012) pada Gambar 20.
Gambar 20 Morfologi isolat cendawan endofit dari Annona squamosa (Yunianto et al., 2012)
Cendawan endofit asal tanaman karet juga berhasil diisolasi dan dikarakterisasai (Gambar 21) (Amaria, Taufiq, and Harni 2013)
Gambar 21 Cendawan endofit dari tanaman karet (Amaria, Taufiq, and Harni 2013)

48
Cendawan Endofit
Gambar 22. Penampakan mikroskopis kapang endofitik (kiri) Phoma (kanan) Pestalotiopsis (Pembesaran 400X) (Ilyas 2007a)
Kelemahan Idenifikasi secara morfologiIdentifikasi secara morfologi sudah banyak mengungkap
jenis-jenis cendawan endofit yang ada pada berbagai jenis tanaman. Namun demikian identifikasi secara morfologi seringkali tidak dapat memberikan kepastian identitas isolat.
Beberapa penyebab ketidakpastian identitas isolate yang diisolasi secara morfologi dikemukakan oleh Hyde dan Soytong (2008); Legiastuti & Aminingsih, (2012) adalah: 1) morfologi cendawan endofit dapat berubah; 2) beberapa cendawan endofit memiliki pertumbuhan sangat lambat; 3) sering terjadi sporulasi. Sedangkan menurut (Diaz, Hennell, and Sucher 2013), beberapa spesies kelihatan sama, padahal sebenarnya tidak sama atau berbeda.
5.2 Identifikasi MolekulerIdentifikasi morfologi secara makroskopis dan mikroskopis
perlu diverifikasi dengan metode lainnya, yaitu metode identifikasi molekuler (Rahayu, Saryono, and Nugroho 2015). Menurut Legiastuti & Aminingsih (2012), identifikasi cendawan endofit menggunakan teknik PCR dan perunutan DNA memiliki kepekaan yang tinggi, cepat, dan akurat. Identitas cendawan

Cendawan Endofit
49
endofit dapat diketahui hingga tingkat spesies berdasarkan pada analisis BLASTN hasil perunutan DNA.
PCR (Polymerase Chain Reaction). Analisis genetik dengan menggunakan metode PCR memanfaatkan cara replikasi DNA dengan bantuan primer yang mengapit daerah tertentu dan optimasi suhu dilakukan untuk mendapatkan kondisi PCR yang optimal. Sehingga dihasilkan produk PCR spesifik yaitu terbentuk pita DNA tebal (Ludyasari 2005).
Bahan yang digunakan untuk setiap tahapan dalam identifikasi molekuler dijelaskan oleh (Diaz, Hennell, and Sucher 2013) adalah: a. Bahan untuk Fermentasi dan ekstrasi cendawan
- Czapek-Dox broth;- Aguades steril; - Miselia cendawan sekitar 500 mg per 250 ml;- Kertas saring
a. Bahan Elektroforesis Gel Agarose- Gel Agarose- 50x TAE buffer- Ethidium bromide (10 mg/mL)- 5x Nucleic acid- 1- kd ladder
b. Bahan Ektrasksi DNA- Kertas saring steril 0.22 µm- Nitrogen cair- mortal dan pestel- DNeasy plant mini kit (QIAGEN)
c. Bahan untuk Amplifikasi PCR- iProof High-Fidelity PCR kit (Bio- Rad)- DNA Engine® Peltier Termal Cycler (Bio-Rad)

50
Cendawan Endofit
d. Bahan PCR Reaction Clean-Up- PureLinkTMPCR Purification Kit (Invitrogen)
Tahapan identifikasi cendawan menurut (Diaz, Hennell, and Sucher 2013) adalah: 1. Fermentasi cendawan
Metode fermentasi cendawan adalah sebagai berikut:a. Media cair Czapek Dox dibuat dengan menambahkan 45,5
g Czapek Dox ke dalam botol Schott 1-L dan menambahkan aquades steril. Sterilisasi media dengan diautoklaf selama 15 menit
b. Media Czapek Dox cair yang telah disterilisasi dituangkan ke dalam erlenmeyer 250 mL.
c. Miselia cendawan dikeluarkan dari cawan petri menggunakan jarum steril sekali pakai dan memasukkan ke dalam Erlenmeyer yang telah telah diisi media Czapek Dox Broth
d. Erlenmeyer ditutup dengan kain tipis untuk memungkinkan aerasi.
e. Media berisi cendawan diinkubasikan pada temperatur 25°C dengan agitasi pada 120 rpm selama 7-14 hari.
2. Ekstraksi DNA Genomik menggunakan DNeasy Plant Mini Kit
a. Media cair yang mengandung miselia disaring menggunakan penyaring vakum steril 0,22 m.
b. Miselia sekitar 20 mg berat basah dipindahkan ke mortar dan ditambahkan nitrogen cair. Miselia digiling dengan mortal di bawah nitrogen cair sampai terbentuk bubuk. Nitrogen cair harus terus-menerus diisi ulang agar miselia tidak mencair.

Cendawan Endofit
51
c. Miselia dalam nitrogen cair dituang ke dalam tabung centrifuge 2-mL dan biarkan nitrogen cair menguap.
d. Ekstraksi genom DNA (gDNA) menggunakan mini kit DNeasy ® tanaman, sesuai petunjuk perusahaan/pabrik (Qiagen DNeasy ® Plant Handbook, 07/2006. Gunakan air suling sebagai pelarut elusi akhir sebagai ganti buffer AE.
3. Penilaian Kuantitas dan Kemurnian DNA dengan UV Spektrofotometri
a. Spektrum UV dari solusi DNA diukur antara 220 dan 320 nm. Absorbansi harus antara 0,1 dan 1 AU.
b. Konsentrasi DNA diperkirakan dengan mengukur absorbansi larutan pada 260 nm. Absorbansi dilipatgandakan sebesar 50 mg/mL/AU untuk mendapatkan konsentrasi dsDNA yang diperkirakan.
c. Kemurnian DNA dari protein diperkirakan dengan mengukur rasio antara absorbansi pada 260 nm dan absorbansi pada 280 nm. DNA murni memiliki A 260/A 280 rasio 1,7-1,9.
d. Kemurnian DNA dari EDTA, karbohidrat, dan fenol diperkirakan dengan mengukur rasio antara absorbansi 260 nm dan absorbansi 230 nm. DNA murni memiliki A 260/ Rasio 230 dari 2,0–2,3.
4. Penilaian Kualitas gDNA dengan Elektroforesis Gel Agarosa
a. Gel agarosa 0,7% disiapkan/dibuat dengan melarutkan 0,35 g bubuk agarosa dalam 50 mL 1 × TAE.
b. gDNA (sekitar 20 mg) dicampur dengan buffer ke dalam gel. Digunakan 1-kb ladder untuk referensi ukuran.
c. Gel di running pada 100 V selama sekitar 60 menit.

52
Cendawan Endofit
d. Gel diwarnai dalam larutan etidium bromida (0,5 mg / mL).e. Gel divisiualisasikan di bawah sinar UV untuk menentukan
kualitas gDNA. Pita tunggal harus jelas di bagian atas gel. Jika gDNA telah rusak, maka goresan biasanya terjadi.
5. Amplifikasi PCR Cendawan wilayah ITS Mengatur reaksi PCR menggunakan kit DNA polimerase
iProof hi-fi delity sesuai dengan instruksi pabrik. Bisa menggunakan reaksi 50-uL per 1 unit iProof DNA polimerase untuk memastikan DNA yang cukup dihasilkan untuk pemurnian, kontrol kualitas, dan pengurutan. Urutan primer disajikan pada Tabel 1 dan volume reaksi disajikan pada Tabel 2.
Tabel 1. Daftar Primer yang umum digunakan untuk Barkoding Cendawan (Diaz, Hennell, and Sucher 2013)
Primer Sequence 5’-3’NSI1 (forward) GAT TGA ATG GCT TAG TGA GGNLB4 (reverse) GGA TTC TCA CCC TCT ATG ACITS1 (forward) TCC GTA GGT GAA CCT GCG GITS4 (reverse) TCC TCC GCT TAT TGA TAT GC
Tabel 2. Volume Reaksi untuk Amplifikasi PCR (Diaz, Hennell, and Sucher 2013)
Komponen Volume per reaksi5 x Proof buffer 10 µL
dNTP mix 1 µLForward primer (10 µM) 2,5 µLReverse primer (10 µM) 2,5 µL
DNA template 50 mgProof DNA polymerase 0,5 µL
H2O steril Sampai 50 µL

Cendawan Endofit
53
Amplifikasi DNA templat menggunakan program suhu yang diadaptasi dari White et al., (1990): (Diaz, Hennell, and Sucher 2013), sebagai berikut: denaturasi awal pada suhu 95°C selama 60 detik; denaturasi 95°C, 30 detik; dan annealing pada suhu 60°C, 40 detik; ekstensi 72°C, 90 detik; perpanjangan waktu terakhir 72°C, 300 detik. Denaturasi, annealing, dan ekstensi diulang 35 kali.
6. Penilaian Kualitas Produk PCR dengan Elektroforesis Gel Agarosa
a. Menyiapkan gel agarosa 1,0% dengan melarutkan 0,5 g bubuk agarose, dalam 50 mL TA 1 ×.
b. Sekitar 20 mg produk PCR yang dicampur dengan menambahkan buffer ke gel. G 1-kb ladder untuk referensi ukuran.
c. Gel dijalankan pada 100 V selama sekitar 60 menit.d. Gel diwarnai dengan larutan etidium bromida (0,5 mg /
mL).e. Gel divisualisasikan di bawah sinar UV untuk menentukan
kualitas gDNA.Satu pita harus jelas. Jika ada beberapa band atau tidak ada
band maka PCR di atas perlu dimodifikasi. Secara umum, suhu annealing dan konsentrasi Mg2+ adalah parameter yang dapat disesuaikan.
7. Pembersihan PCRBersihkan produk amplifikasi PCR menggunakan Kit
Purifikasi PCR PureLink ™, seperti yang dijelaskan oleh instruksi pabrik, kecuali untuk perubahan berikut:
(a) Buffer HC digunakan untuk pengikatan DNA untuk

54
Cendawan Endofit
menghilangkan produk samping PCR dengan berat molekul rendah.
(b) 30 mL air suling digunakan untuk mengelusi DNA yang dimurnikan.
Gambar 23. Hasil amplifikasi DNA cendawan endofit asal tanaman cabai menggunakan pasangan primer ITS1/
ITS1(Legiastuti and Aminingsih 2012)
Gambar 24. Hasil amplifikasi PCR menggunakan pasangan primer NL1 dan NL4 (Yunianto et al. 2012)

Cendawan Endofit
55
Gambar 25. Hasil amplifikasi PCR menggunakan pasangan primer ITS1/ITS4 (S. Syamsia et al. 2019).
Contoh hasil identifikasi molecular dari 2 isolat cendawan endofit memiliki kemiripan dengan Podoscypha bolleana dan Coprinopsis cinerea (Tabel 3).
Tabel 3. Tingkat kemiripan Isolat Cendwan endofit asal tanaman Padi Lokal Sulawesi Selatan pada analisis BLASTN (Syamsia Syamsia et al. 2019)

56
Cendawan Endofit
Tabel 4. Tingkat Kemiripan solate cendawan endofit asal tanaman cabai berdasarkan pada analisis BLASTN
Rangkuman
Cendawan endofit dapat diidentifikai secara morfologi dan molekuler. Identifikasi morfologi didasarkan pada pengamatan makrokopis dan mikroskopis. Makroskopis berdasarkan pada warna koloni, tipe permukaan koloni, dan tipe pertumbuhan. Mikroskopis berdasarkan pada bentuk hifa, struktur misellium, jenis anamorf, konidiomata, konidia, dan konidiofor (ukuran, warna, bentuk, ornamen, tepi, dan lain-lain).
Identifikasi secara molekuler menggunakan teknik PCR dan perunutan DNA. Identifikasi dapat diketahui hingga tingkat sepesis berdasarkan analisis BASTN hasil perunutan DNA. Tahapan Identifikasi cendawan secara molekuler meliputi:a. Fermentasi cendawanb. Ekstraksi DNA Genomikc. Penilaian Kuantitas dan Kemurnian DNA dengan spektrofotome tri UVd. Penilaian Kualitas gDNA dengan Elektroforesis Gel Agarosae. Amplifikasi PCR Cendawan wilayah ITSf. Penilaian kualitas Produk PCR dengan elektroforesis Gel Agaorsa
Primer yang umum digunakan untuk barcoding cendawan adalah: pasangan primer NSI1/NLB4 dan ITS2/ITS4.

Cendawan Endofit
57
Tes Formatif1. Berikut ini kriteria yang digunakan dalam identifikasi
cendawan secara makroskopis, kecuali …a. Warna kolonib. Permukaan kolonic. Hifad. Tipe pertumbuhan
2. Karakter mikroskopis yang digunakan dalam identifikasi secara morfologi adalah …a. Warna kolonib. Tipe Pertumbuhanc. Permukaan kolonid. Spora
3. Tahap pertama dalam identifikasi molekuler adalah…a. Ekstraksi DNA genomicb. Amplifikasi PCRc. Fermentasi cendawand. Penilaian kalitas gDNA dengan elektroforesis gel solate
4. Bahan untuk fermentasi cendawan adalah…a. Gel agaroseb. Ethidium bromidec. Czapek-Dox brothd. Nitrogen cair
5. Dneasy plant mini kit adalah salah satu bahan untuk…a. Fermentasi Cendawanb. Ektraksi DNAc. Amplifikasi PCRd. Elektroforesis gel agarose
6. Ethidium bromide merupakan bahan yang digunakan pada tahapan

58
Cendawan Endofit
a. Fermentasi Cendawanb. Ektraksi DNAc. Amplifikasi PCRd. Elektroforesis gel agarose
7. Pasangan primer yang umum digunakan untuk barcoding cendawan adalah….a. NS1/ITS1b. NS1/ITS4c. ITS1/ITS4d. NS1/ITS1
8. Nitrogen cair merupakan bahan yang digunakan pada tahapan…a. Fermentasi Cendawanb. Ektraksi DNAc. Amplifikasi PCRd. Elektroforesis gel agarose
9. Identifikasi secara morfologi memiliki kelemahan antara lain, kecuali…a. sering terjadi sporulasib. spesies sering kelihatan sama, padahal sebenarnya samac. pertumbuhan sangat cepatd. morfologi dapat berubah
10. Kelebihan identifikasi secara molekuler antara lain, keculai…a. Kepekaan tinggi, namun lambatb. Kepekaan rendah, dan akuratc. Kepekaan tinggi, lambat dan akuratd. Kepekaan tinggi, cepat dan akurat

Cendawan Endofit
59
Kunci Jawaban1. c2. d3. c4. c5. b
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban solate
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 5.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ------------------------------------------ x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 90 – 100% = baik sekali 80 – 89% = baik 70 – 79% = cukup < 70% = kurangBila Anda mencapai tingkat penguasaan 80% atau lebih,
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari solate materi subunit 1 terutama bagian yang belum Anda kuasai.
1. c2. d3. c4. c5. b6. d7. c8. b9. c10. d

60
Cendawan Endofit
6. 1 Penyimpanan Cendawan EndofitPrinsip penyimpanan mikroba adalah menghetikan atau
mengurangi laju penggunaan energi sel selama masa penyimpanan sehingga dapat diaktifkan Kembali karena masih memiliki energi sel yang cukup atau viable (Bjerketorp, et al. 2006 (Najmiyati and Dominikus H Akhadi 2013). Tujuan utama penyimpanan/preservasi mikroba adalah:
1. Mereduksi atau mengurangi laju metabolisme darimiroorganisme hingga sekecil mungkin dengan tetapmempertahankan viabilitasnya (daya hidupnya)
2. Memelihara sebaik mungkin biakan, sehingga diperolehangka perolehan (recovery) dan kehidupan (survival)yang tinggi dengan perubahan ciri-ciri minimum(Machmud, 2001) (Yulinery and M.Dewi 2012)
6. 2 Metode Penyimpanan Cendawan EndofitMenurut (Wulandari 2019), metode penyimpanan mikroba
termasuk cendawan dibedakan atas 3 yaitu:
BAB VIMETODE PENYIMPANAN
CENDAWAN ENDOFIT
Setelah mempelajari bab ini, mahasiswa diharapkan mampu:1. Menjelaskan tujuan penyimpanan cendawan endofit2. Menjelaskan metode metode penyimpanan cendawan endofit

Cendawan Endofit
61
1. Jangka pendekMetode penyimpanan ini memiliki jangka waktu yang solatesingkat, seperti dari isolasi sampai identifikasi.
2. Jangka menengahMetode penyimpanan jangka menengah sesuai dengan duraiproyek penelitian.
3. Jangka panjangMetode penyimpanan jangka panjang untuk pengumpulan,konservasi atau referensi penelitian.
Metode penyimpanan cendawan menurut (Najmiyati and Dominikus H Akhadi 2013) ada dua yaitu: a. Metode Penyimpanan jangka pendek
Penyimpanan jangka pendek terkait dengan kegiatanpenelitian di laboratorium.
b. Metode Penyimpanan jangka panjangPenyimpanan jangka panjang terkait dengan kegiatan koleksidan konservasi stok mikroba agar dapat ditumbuhkan solateseperti semula jika dibutuhkan.
Secara detail (Ilyas 2007b), menguraikan metode penyimpanan/preservasi cendawan ke dalam 4 metode yaitu:
a. Metode Penyimpanan transfer kultur berkala(Subculturing method)
b. Metode Penyimpanan Kering (Drying Method)c. Metode Penyimpanan Beku (Cryopreservation/Frezing
Method)d. Metode Penyimpanan Kering-Beku (Freze Drying/
Liophilization Method)

62
Cendawan Endofit
a. Metode Penyimpanan transfer kultur berkala (Subculturingmethod)
Metode kultur berkala hanya dilakukan terhadap solate yang daya simpannya pendek, yaitu hanya dapat bertahan hidup 6 bulan sehingga setelah periode tersebut perlu ditumbuhkan solate (subculture) pada media yang baru. Demikian juga solate yang disimpan pada parafin cair dan aquades steril disubkultur setelah 4-5 tahun.
Media penyimpanan yang digunakan pada metode penyimpanan transfer kultur berkala yaitu media agar, minyak mineral, dan aquades steril. Media agar dapat menggunakan media CMC, PDA, CMA, dan PCA, sedangkan media cair menggunakan miyak mineral seperti paraffin cair atau paraffin medis.
Metode preservasi dalam paraffin cair pertama kali digunakan oleh Sherf (1943) dan diaplkasikan berturut turut oleh (Norris, 1994; Wenham, 1964; Little dan Gordo, 1967; Smith et al, 1970; Onions, 1971; (Suciatmih and Rachmat 2005)
Metode Preservasi dalam paraffin cair dan dalam air dijelaskan (Suciatmih and Rachmat 2005) adalah sebagai berikut:
1) Preservasi dalam parafin caira) Menumbuhkan cendawan dalam tabung reaksi atau botl
berisi media PDA miring.b) Cendawan yang sudah menutupi permukaan agar ( 3-5
hari) direndam dengan paraffin cair yang telah distrilisasidengan autoclave pada suhu 121 °C selama 15 menitsebanyak 2 kali.
c) Banyaknya parafin cair yang diperlukan kira-kira 1 cm diatas permukaan cendawan.

Cendawan Endofit
63
d) Menyimpan tabung reasi atau botol berisi cendawan danparafin cair disimpan dalam lemari pada suhu ruang (±27° C).
2) Preservasi dalam aira. Sterilisasi botol atau tabung reaki yang berisi aquades
± 15 ml (sesuai ukurang botol/tabung reaksi) denganautoclave pada suhu 121 °C selama 15 menit sebanyak2 kali.
b. Menumbuhkan masing-masing cendawan pada cawanpetri yang berisi medium PDA sampai umur 3-5 hari.
c. Memotong medium PDA yag telah ditumbuhi cendawanberbentuk dadu dengan ukuran 0.5 cm x 0.5 cm (mediumdan cendawan).
d. Memasukkan potongan berbentuk dad uke dalam tabungreaksi yang berisi air yang telah distrilkan.
e. Menyimpan botol atau tabung reaksi pada suhu ruang (±27° C).
b. Metode Penyimpanan Kering (Drying Method)Metode penyimpanan kering dilakukan dengan cara
menurunkan kadar air di sekitar mikroba sehingga laju metabolisme turun bahkan terhenti. Penyimpanan kering dapat dilakukan dengan menggunakan bahan pengering berupa silica gel anhidrit dan tanah atau pasir kering steril.
c. Metode Penyimpanan Beku (Cryopreservation/FrezingMethod)
Metode penyimpanan beku dilakukan dengan cara menurunkan suhu di sekitar mikroba sehingga laju metabolisme

64
Cendawan Endofit
turun. Aktivitas metabolisme sel dapat ditekan pada suhu di bawah -70oC, bentuk kristas es yang lebih stabil, dan tidak membahayakan viabilitas akan dicapai pada suhu di bawah -80oC.
d. Metode Penyimpanan Kering-Beku (Freeze Drying/Liophilization Method)
Penyimpanan kering beku adalah metode penyimpanan material biologis yang dapat mempertahankan bentuk, struktur, dan aktifitas dari produk material biologis. Metode penyimpanan kering beku yaitu liophilization drying (L-drying) dan freeze-driying. Proses pengeringan pada L-drying melalui proses sublimasi, sedangkan pada freeze drying pengeringan dilakukan secara evaporasi.
Proses pengeringan pada metode L-drying dilakukan melalui proses evaporasi, sampel dibuat hampa udara dan dikeringakn darinfase cair tanapa melalui proses pembekuan terlebih dahulu.
Penyimpanan sampel mikroba dalam tabung ampul menggunakan teknik L-drying (T= - 48 ° C, vaccum gauge =0.0 pa) selama ± 3 jam sampai tercapai kondisi vakum. Ampul ditutup (sealing) dengan pemanas api yang melelhkan leher tabung kaca ampul yang sudah ditutup dilakukan dengan penembakan aliran listrik dengan menggunakan alat Spark Tester. Tabung ampul yang berisi sampel akan berwarna biru apabila kondisi di dalam tabung ampul tersebut vakum (Yulinery and M.Dewi 2016).

Cendawan Endofit
65
Tes Formatif1. Salah satu tujuan dari penyimpanan mikroba adalah...
a. meningkatkan laju metabolismeb. mereduksi laju metabolismec. mensurunkan nilai survivald. menurunkan nilai recovery
2. Berdasarkan jangka waktunya metode penyimpanan dibedakan atas:a. jangka panjang
Rangkuman
Tujuan utama penyimpanan mikroba adalah mereduksi laju metabolisme mikroba dengan tetap mempertahankan viabilitasnya dan memelihara biakan sehingga memiliki nilai recovery dan survival yang tinggi.
Metode penyimpanan berdasarkan jangka waktunya dibagi atas 3 yaitu jangka pendek, menengah dan panjang. Jangka pendek untuk kegiatan isolasi sampai identifikasi, jangka menengah sesuai dengan durasi proyek dan jangka panjang untuk pengumpulan, konservasi atau referensi penelitian.
Berdasarkan prosesnya metode penyimpanan dibedakan atas 4 yaitu: metode penyimpanan transfer kultur berkala (Subculturing method), metode penyimpanan transfer kultur berkala (Subculturing method), metode penyimpanan beku (Cryopreservation/Frezing Method), metode penyimpanan kering-beku (Freze Drying/ Liophilization Method).
Metode transfer kultur berkala menggunakan media agar, minyak mineral dan aquades, metode penyimpanan kering menggunakan bahan silika gel anhidrit dan tanah atau pasir kering steril, metode penyimpanan kering-beku menggunakan tabung ampul yang ditutup dengan metode pemanasan.

66
Cendawan Endofit
b. jangka pendekc. jangka menengahd. a, b dan c benar
3. Media pasir digunakan pada metode penyimpanan …a. keringb. bekuc. kultur berkalad. kering beku
4. Berikut ini bahan yang dapat digunakan untuk metodepenyimpanan kultur berkala, kecuali …a. PDAb. media agarc. minyak minerald. aquades
5. Bahan silika gel an hidrit digunakan pada metodepenyimpanan…a. kultur berkalab. keringc. bekud. kering beku
6. Metode penyimpanan yang menggunakan bahan ampuladalah…a. kultur berkalab. keringc. bekud. kering beku
7. Metode penyimpanan yang menggunakan paraffin cairadalah…a. kultur berkalab. kering

Cendawan Endofit
67
c. bekud. kering beku
8. Metode penyimpanan yang dilakukan dengan menurunkankadar air di sekitar mikorba adalah …a. kultur berkalab. keringc. bekud. kering beku
9. Metode penyimpanan dengan menurunkan suhu di sekitarmikroba adalah …a. kultur berkalab. keringc. bekud. kering beku
10. Metode penyimpanan yang menggunakan proses sublimasiadalah…
a. Freze Drying Methodb. Liophilization Methodc. Frezing Methodd. Dying Method
Kunci Jawaban1. B2. D3. A4. A5. B
1. B2. D3. A4. A5. B6. D7. A8. B9. C10. B

68
Cendawan Endofit
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban solate
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 6.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ---------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 90 – 100% = baik sekali 80 – 89% = baik 70 – 79% = cukup < 70% = kurang Bila Anda mencapai tingkat penguasaan 80% atau lebih,
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari solate materi subunit 1 terutama bagian yang belum Anda kuasai.

Cendawan Endofit
69
7.1 Manfaat Langsung 7.1.1 Penghasil Fitohormon
Istilah fitohormon atau phytohormone berasal dari solat Yunani yaitu “phytoes” yang artinya tanaman dan “hormoaein” yang artinya zat perangsang. Jadi fitohormon dapat didefinisikan sebagai zat-zat yang dapat merangang pertumbuhan dan proses fisiologis tanaman (Dewi 2008).
Fitohormon adalah senyawa-senyawa yang dalam konsentrasi yang rendah mempengaruhi proses-proses fisiologis dan proses lain pada tanaman. Proses tersebut seperti pertumbuhan, differensiasi dan perkembangan, pembukaan stoma, translokasi serta serapan hara (Dewi 2008).
Fitohormon dapat disintesis secara alami oleh tanaman dan dapat pula diperoleh dari mikroba yang bersimbiosis dengan tanaman. Mikroba penghasil fitohormon umumnya hidup di daerah rizosfer dan endofit tanaman. Salah satu mikroba penghasil fitohormon adalah cendawan endofit.
Fitohormon dikelompokkan oleh (Dewi 2008), menjadi 6
BAB VIIMANFAAT CENDAWAN ENDOFIT
Setelah mempelajari bab ini mahasiswa diharapkan mampu:1. Menjelaskan manfaat langsung cendawan endofit2. Menjelaskan manfaat tidak langsung cendawan endofit

70
Cendawan Endofit
kelompok yaitu: 1) Auksin; 2) Giberelin; 3) Sitokinin; 4) Etilen; 5) Brasinosteroid; 6) Asam Absisat.
1. AuksinAuksin merupakan fitohormon yang paling banyak di alam
dan paling aktif adalah Indole-3-Acetic Acid (IAA) (Tsavkelova et al., 2005;(Sukmadi 2013).
Mekanisme produksi fitohormon oleh cendawan endofit sebagai pemacu pertumbuhan tanaman (PGP) pada tanaman inang hampir sama dengan mekanisme produksi fitohormon pada cendawan rizosfer atau PGPR. Cendwan endofit meningkatkan pertumbuhan tanaman kelompok non legume dengan meningkatkan pertumbuhannya melalui produksi indole acetic acid, gibberellic acid, ethylene, dan auxins (Yadav and Yadav 2017).
Produksi fitohormon oleh cendawan dapat dideteksi dengan menggunakan reagen Salkowski (12 gL-1 FeCl3 dalam 429 ml L-1 H2SO4) dan precursor L-tryptophan (0.1 gL-1) (Glickman dan Dessaux, 1995). Perubahan warna pink pada kultur fitrat cendawan endofit setelah ditambahkan reagen Salkowski merupakan indikator adanya kemampuan isolate cendawan endofit dalam memproduksi isolate IAA (Gambar 27). Deteksi kemampuan produksi isolate IAA secara kuantitatif dengan mengukur absorbansi dari supernata menggunakan spektrofotometer UV pada panjang gelombang 535 nm dan konsentrasi IAA diperoleh dengan membandingkan dengan kurva standar. Kemampuan beberapa isolate cendawan endofit dalam memproduksi IAA dapat dilihat pada Tabel 5.

Cendawan Endofit
71
Gambar 26. Perubahan warna pink menunjukkan kemampuan produksi IAA (Dokumen Pribadi)
Tabel 5. Absorbansi dan konsentrasi IAA dari solate cendawan endofit (Syamsia et al. 2015).
2. GiberelinGiberelin (GA) pertama kali diisolasi dari Gibberella
fujikuroi (Hasan 2011). Menurut (Wiraatmaja 2017), jenis

72
Cendawan Endofit
gibereling yang ditemukan pada cendawan yaitu: GA1, GA2, GA3,GA4,GA7,GA9. Salah satu fungsi giberelin adalah mematahkan dormansi atau mempercepat perkecambahan. Giberelin berperan dalam pertumbuhan tanaman dan membatu dalam mengatasi stress abiotik (Khan et al., 2013); (Khan et al. 2017).
Cendawan penghasil giberelin seperti Fusarium, Aspergillus dan Penicilium (Leitão and Enguita 2016); (Alori, Glick, and Babalola 2017). Mikroba pelarut fosfat berpengaruh pada pertumbuhan dan perkembangan tanaman (Tallapragada and Gudimi, 2011); (Alori, Glick, and Babalola 2017).
Deteksi kemampuan cendawan dalam menghasilkan giberelin dengan cara kalorimetri dengan menggunakan pereaksi zinc acetate dan potassium ferrosianida. Metode ini telah digunakan oleh beberapa peneliti seperti: Glick 1995 dan Caron 1995); (Kafrawi, Kumalawati, and Mulyani 2015).
Hasil deteksi kemampuan isolate cendawan endofit dalam memproduksi giberelin bervariasi (Gambar 28). Hasil penelitian yang dilakukan (Gusmiaty et al. 2019), menunjukkan bahwa cendawan rizosfir jabon merah mampu memproduksi 2, 370 mgl-1. Demikian juga penelitian Iradhatullah, et al., (2019), juga menunjukkan bahwa cendawan pelapuk asal tanaman kakao mempu memproduksi giberelin, dan konsentrasi tertinggi dihasilkan oleh cendawan Tremella sp (4.10 µgl-1). Demikian juga dengan penelitian yang dilakukan oleh (Hasan 2011), Aspergillus flavus, A. niger, Fusarium oxysporum, Penicillium corylophilum, P. cyclopium, P. funiculosum and Rhizopus stolonifera mampumemproduksi giberelin.

Cendawan Endofit
73
Gambar 27. Produksi Giberelin Cendawan endofit dari padi lokal (Syamsia et al., 2020)
3. SitokininSitokinin berperan dalam pertumbuhan dan perkembangan
sel. Menurut Hartman (2002); (Kurniati, Sudartini, and Hidayat 2017), permulaan terbetnuknya akan tidak hanya dipengaruhi oleh auksin, tetapi juga oleh sitokinin dan giberelin dan sejumlah kofaktor pembentuk akar lainnya.
Sitokinin adalah turunan dari adenin basa nitrogen dan dibekdan atas atas: trans- zeatin (tZ), cis-zeatin (cZ), dihydrozeatin (DHZ) dan isopentenyladenine (iP), benzylaminopurine (BAP) dan toplins (T) (Stirk & Van Staden, 2010; Spichal, 2012); (Noble et al. 2014).
Salah satu cendawan yang dapat memproduksi sitokinin adalah Rhizopogon ochraceorubens (Crafts and Miller 1974). Kemampuan cendawan endofit dalam memproduksi sitokinin masih terbatas bila dibandingkan dengan mikorhiza.
7.1. 2 Pelarut FosfatFosfar merupakan unsur hara utama, kedua setelah nitrogen
yang dibutuhkan tanaman. Sebagian besar fosfor tanah, sekitar

74
Cendawan Endofit
95-99% berada dalam bentuk fosfat tidak larut dan tidak dapatdimanfaatkan oleh tanaman (Vassileva et al., 1998; Pradhan andSukla 2005).
Pemberian pupuk ke dalam tanah merupakah salah satu cara untuk meningkatkan ketersediaan fosfat dalam tanah, namun cara belum efektif dalam meningkatkan serapan pada tanaman. Menurut Omar, 1998; Pradhan and Sukla, 2005), sebagian besar fosfat setelah diaplikasikan mengalami transfomasi secara cepat ke dalam bentuk tidak larut.
Menurut Tallapragada, P., and Gudimi, M., 2011; Alori, Glick, and Babalola (2017), mikroba pelarut fosfat atau Phosfate Solubizing Microorganisms (PSM) memberikan pengaruh terhadap pertumbuhan dan perkembangan tanaman.
Menurut Yadav and Yadav (2017), beberapa endofit melepaskan asam organik ke dalam tanah yang melarutkan kompleks fosfat dan mengubahnya menjadi orto-fosfat yang dapat diserap oleh tanaman.
Gambar 28. Skema representasi dari asam solate yang dapat diproduksi oleh Mikroba Pelarut Fosfat dang digunakan untuk
melarutkan bentuk fosfat anorganik (Alori, Glick, and Babalola 2017)

Cendawan Endofit
75
Uji Kemampuan cendawan endofit dalam melarutkan fosfat secara kualitatif menggunakan medium Pikovskaya padat yang mengandung 0.5 % tricalsum fosfat (Ca3PO4) sebagai sumber fosfat (Sundara dan Shinha, 1962; Subba Rao, 1982; Syamsia et al. 2015). Kemampuan isolat cendawan melarutkan fosfat ditandai oleh adanya zona bening disekitar cendawan.
Gambar 29. Zona bening yang terbentuk disekitar koloni cendawan merupakan indikator kemampuan melarutkan fosfat
(Dokumen Pribadi)
Uji pelarutan fosfat secara kuantitatif menggunakan media Pikovskaya cair dan pengukuran tingkat pelarutan fosfat menggunakan spektrofotometer.
Gambar 30. Perubahan warna supernata menjadi biru menunjukkan kemamuan melarutkan fosfat (Dokumen pribadi)

76
Cendawan Endofit
Tabel 6. Kemampuan melarutkan fosfat dari solate cendawan endofit (Syamsia et al. 2015).
7.1.3 Produksi SidereforeIstilah siderefor berasal dari Bahasa Yunani yang berarti
pembawa besi (Neilands, 1995; Farida 2012). Menurut Schwyn and Neilands, 1987; Ahmed and Holmström 2014, Siderefore adalah agen pengkhelat besi dengan ukuran molekul kecil yang dihasilkan oleh mikroorganisme. Yadav and Yadav (2017), mendefinisikan Siderofor sebagai senyawa pengkhelat besi yang dapat memanfaatkan zat besi untuk tanaman dan menghilangkan keracunan besi. Berdasarkan pengertian tersebut maka keberadaan siderefore sangat penting bagi pertumbuhan tanaman untuk mengatasi kekurangan/kelebihan unsur hara besi bagi tanaman.
Mikroorganisme dapat menghasilkan siderefore pada kondisi kekurangan Fe (Schwyn and Neilands, 1987; Ahmed and Holmström 2014). Selain itu menurut Calvente et al., (1999); dan Halo, Al-Yahyai, and Al-Sadi (2018), Siderefor yang dihasilkan

Cendawan Endofit
77
oleh Rodhotorula glutinis dapat menekan penyakit blue rot pada apel.
Beberapa tanaman dilaporkan dapat menghasilkan siderefore seperti: barley dan gandum dapat memperoleh Fe dari sumber yang tidak larut (Kraemer et al., 2006;(Ahmed and Holmström 2014).
Jenis SidereforeSiderefore dibagi dalam tiga kelompok fungsional yang
khas, yaitu: hidroksamat, katekolat dan karboksilat (Ahmed and Holmström 2014).
Bakteri sebagian besar menghasilkan siderefore tipe katekolat (enterobaktin) dan sebagian menghasilkan karboksilat (rhizoabaktin) dan hidroksamat (ferrioksamin) (Matzanke, 1991; (Ahmed and Holmström 2014). Cendawan menghasilkan siderefore tipe katekolat dan hidroksamat (Fekete et al., 1989; Fekete; 1993; (Ahmed and Holmström 2014).
Kemampuan tanaman dan mikroorganisme dalam memproduksi siderefore menjadi sangat penting. Hal ini terkait dengan permasalahan yang sering dihadapi dalam pertumbuhan dan produksi tanaman terutama terkait kelebihan dan kekurangan unsur hara besi.
Cendawan menghasilkan siderefore dan mengkhelat besi, dengan membentuk senyawa kompleks dengan logam berat seperti Cd, Cu dan Zn (Johnstone TC, Nolan EM, 2015; Hussein and Joo 2019)
Menurut Sharma & Johri, 2003; (Prihatiningsih, Djatmiko, and Lestari 2017) keberadaan siderefore dapat menguntungkan tanaman karena dapat menghambat pertumbuhan pathogen. Pathogen akan mengalami kekurangan Fe 3+ akibat terikatnya

78
Cendawan Endofit
Fe 3+ oleh siderefor Hal ini menjadi peluang untuk pemanfaatan siderefore sebagai pemacu pertumbuhan dan pengendali pathogen. Deteksi kemampuan cendawan dalam memproduksi siderefor dapat dilakukan secara kualitatif dan kuantitaif.
Cendawan penghasil sideroforMikroorganisme yang mampu menghasilkan siderefore
adalah bakteri dan cendawan. Beberapa bakteri berhasil diisolasi dari tanaman memiliki kemampuan menghasilkan siderefore seperti Bacillus subtilis yang diisiolasi dari rizosfer kentang (Prihatiningsih, Djatmiko, and Lestari 2017), Pseudomonas fluoresens (Pratama, Advinda, and Fifendy 2018). Kelompok cendawan seperti Trichoderma (Anke et al. 1991), Cendawan pelapuk coklat Gloephyllum trabeum (Ahmed and Holmström 2014).
Deteksi kemampuan memproduksi siderefore menggunakan reagen Hathaway yang telah digunakan oleh Sivasakthivalen dan Stella Stella (2012) dan Wang et al. (1993); (Kafrawi, Kumalawati, and Mulyani 2015). Hasil skrining terhadap 8 isolat cendawan endofit dalam memproduksi siderefore dapat dilihat pada Gambar 32. Isolat cendawan endofit memiliki kemampuan memproduksisiderefor tipe katekol lebih tinggi dibanding Na. Salisilat. IsolatE4 merupakan isolate terbaik dalam memproduksi katekol danNa. Salisilat.
Gambar 31. Kemampuan produksi siderefore isolate cencawan endofit (Syamsia et al. 2020)

Cendawan Endofit
79
Menurut Neilands (1995); (Farida 2012), siderofor dapat digunakan dalam pengendalian penyakit tumbuhan dengan memanfaatkan peranannya untuk menyerap besi dari lingkungan dan menyediakan mineral yang penting bagi sel mikroba.
7.1. 4 Produksi Enzim Enzim merupakan golongan protein yang paling banyak
terdapat dalam sel hidup dan mempunyai fungsi penting sebagai biokatalisator pada reaksi-reaksi biokimia. Enzim dihasilkan oleh semua mahluk hidup untuk mengkatalisis reaksi biokimia dala tubuh mahluk hidup. Enzim dapat diekstraksi dari mikroorganisme karena mikroorganisme menghasilkan enzim dalam jumlah dan jenis yang sangat bervariasi. Mikroorganisme juga dapat dikulturkan untuk memperoleh enzim yang dihasilkannya
Salah satu peranan enzim dalam bidang pertanian adalah pemanfaatan dalam penanganan limbah pertanian secara biologi. Limbah pertanian memiliki komponen utama lignoselulosa. Lignoselulosa merupakan komponen utama dari biomassa yang terdiri dari tiga polimer yaitu: selulosa 35-50%, hemiselulosa 20-35%, dan lignin 10-25% (Saha, 2004; Setiawati et al. 2019).
Jenis EnzimBerdasarkan tempat kerjanya enzim dapat dibedakan
mejadi 2 macam, yaitu enzim intraseluler dan ekstraseluler. Enzim intraseluler merupakan enzim yang langsung digunakan di dalam sel sedangkan Enzim ekstraseluler merupakan enzim yang dilepas dari sel ke lingkungan untuk menghidrolisis molekul polimer di lingkungan, seperti selulosa, hemiselulosa, lignin (Maratun Sholihati, Baharuddin, and Santi 2015).

80
Cendawan Endofit
a. SelulaseEnzim selulase yang dihasilkan oleh mikroba dapat
mendegradasi selulosa dengan cara mendegradasi molekul selulosa yang tidak larut menjadi mono atau disakarida sederhana larut sehingga dapat digunakan oleh mikroba sebagai sumber energi. Proses degradasi selulolase merupakan hasil kerja dari tiga komponen enzim secara sinergis, yaitu endoglukanase, eksoglukanase, dan β-glukosidase (Lymar et al., 1995);(Razie et al. 2019)
Munifah (2011); (Murtiyaningsih and Hazmi 2017), menambahkan bahwa enzim selulase adalah enzim yang dapat menghidrolisis selulosa dengan memutus ikatan glikosidik β-1,4 dalam selulosa, selodektrin, selobiosa, dan turunan selulosa lainnya menjadi gula sederhana atau glukosa.
Cendawan yang dapat mendegradasi selulose diantaranya: Aspergillus, Chaetomium, Fusarium, Pencillium, Rhizoctonia, Rhizopus dan Trichoderma (Razie et al. 2019)b. Kitinase
Kitinase merupakan enzim hidrolitik yang dapat menghidrolisis kitin pada ikatan β-1,4- glikosidiknya dengan menghasilkan derivat kitin seperti oligomer kitin yang mempunyai banyak manfaat (Purkan, et al. 2016). Enzim kitinase saat ini banyak digunakan sebagai agen degradasi karena dapat mendegradasi kitin menjadi produk yang ramah lingkungan dan dapat digunakan dalam bidang, pangan, dan lain-lain (Pratiwi et al. 2015; Elawati et al. 2018).
7.2 Manfaat Tidak Langsung7.2.1 Cekaman Abiotik
Cekaman abiotik seperti kekeringan, suhu tinggi atau

Cendawan Endofit
81
salinitas seringkali menyebabkan tanaman tidak dapat bertahan hidup. (Yulianti 2012).
a. Cekaman kekeringanKekeringan merupakan salah satu faktor perhambat
dalam pertumbuhan dan produksi tanaman. Menurut Nio dan Kandou, 2000); tingkat kerugian yang diakibatkan oleh cekaman kekeringan tergantung pada intensitas kekeringan dan lamanya tanaman mengalami kekurangan air.
Salah satu strategi yang dapat dilakukan dalam mengatasi cekaman kekeringan adalah dengan memanfaatkan cendawan endofit. Aly et al. (2011); (Khan et al. 2017), menjelaskan bahwa ada tiga teori yang terkait dengan symbiosis endofit dengan tanaman dalam memicu inangya mengaktifkan sistem pertahanannya, yaitu:1. Endofit menghasilkan senyawa oksigen reaktif untuk
mengoksidasi atau denaturasi sel inang untuk memacu tanaman meningkatkan ketahanannya terhadap cekaman yang menimpanya
2. Endofi menghasilkan berbagai macam antioksidan, asam fenol, dan derivatnya yang berperan dalam meningkatkan ketahanan tanaman terhadap tekanan dari luar
3. Endofit memicu petumuhan dan perkembangan akar lebih dalam untuk memperloeh hara dan air sehingga tanaman mampu bertahan dalam kondisi kering dan cepat pulih jika mengalami stress air.
Beberapa penelitian menunjukkan cendawan endofit berpotensi meningkatkan ketahanan tanaman terhadap cekaman kekeringan, seperti : 1) Beberapa spesies Trichoderma telah

82
Cendawan Endofit
dilaporkan mampu memperbaiki respon tanaman terhadap cekaman kekeringan, seperti adanya pengaruh pada panjang tajuk dan akar dari padi (Shukla, et al., 2012), 2) hipokotil yang lebih tinggi, berkurangnya kelayuan pada persemaian kakao setelah diberi perlakuan Trichoderma hamatum DIS 219b (Bae, et al., 2009); 3) peningkatan persentase perkecambahan dan pertumbuhan pada gandum dan tomat (Mastouri, et al., 2010; Hubbard, et al., 2012) pada kondisi cekaman kekeringan. Selain itu, penurunan laju fotosintesis, konduktansi stomata, dan peroksidase yang diakibatkan oleh cekaman kekeringan dapat ditekan dengan perlakuan Trichoderma (Bae, et al., 2009; Shukla, et al., 2012).
Penelitian (Yuspradana, Wiyono, and Widyastuti 2017), menggunakan cendawan cendawan endofit Acremonium sp, Curvularia sp, Penicilium sp, dan Nigrospora sp mampu meningkatkan ketahanan tanaman padi terhadap cekaman kekeringan. Penelitian yang dilakukan oleh Syamsia, (2015), cendawan endofit Penicillium sp, Aspergillus sp dan Aspergillus niger mampu meningkatkan pertambahan tinggi tanaman dan jumlah anakan padi pada kondisi cekaman kekeringan pada percobaan di green house.
b. Keracunan logam beratKemampuan tanaman bertahan hidup pada tanah-tanah
yang terkontaminasi logam berat adalah berkat adanya endofit yang memiliki kemampuan mendegradasi, mengeliminasi, atau menggunakan logam-logam tersebut dalam sistem metabolismenya (weyen et al., 2009; Aly et al, 2011;(Yulianti 2012).

Cendawan Endofit
83
7.1.2 Cekaman BiotikCekaman biotik adalah cekaman yang diakibatkan oleh
organisme hidup seperti hama, penyakit dan gulma. Mekanisme cendawan endofit dalam melindungi tanaman terhadap serangan pathogen ataupun serangga menurut Gao et al. 2010; (Yulianti 2012)
1. Penghambatan pertumbuhan pathogen secara langsungdengan senyawa antibiotic dan enzim litik yang dihasilkan
2. Penghambatang secara tidak langsung melalui perangsangan endofit terhadap tanaman menghasilkan metabolit sekunderseperti asam salisilat, asam jasmonat dan etilen yangberfungsi sebagai antimikroba seperti fitoaleksin
3. Perangsang pertumbuhan tanaman sehingga lebih kebaldan tahan terhadap serangan pathogen
4. Kolonisasi jaringan tanaman sehingga pathogen sulitpenetrasi
5. Hiperparasit
Rangkuman
Manfaat cendawan endofit terhadap pertumbuhan tanaman dibedakan atas dua yaitu: manfaat langsung dan tidak langsung.
Manfaat lansung meliputi: menghasilkan fitohormon seperti IAA dan Giberelin, melarutkan fosfat, memproduksi siderefor, memproduksi enszim.
Manfaat tidak langsung seperti: meningkatkan ketahanan tanaman terhadap cekaman abiotik (kekeringan dan keracunan logam berat), cekaman biotik (hama dan penyakit).
Produksi fitohormon terutama IAA dapat dideteksi dengan menggunakan reagen. Salkowski, Kemampuan melarutkan fosfat dengan menggunakan medium Pikovskaya, produksi siderefore dapat dideteksi dengan menggunakan reagen Hathaway.

84
Cendawan Endofit
Tes Formatif1. Salah satu manfaat dari cendawan endofit adalah menghasilkan
fitohormon. Fitohormon berasal dari Bahasa ….a. Inggrisb. Yunanic. Belandad. Indonesia
2. Fungsi utama siderefore adalah….a. mengikat nitrogenb. mengikat besic. mengikat fosfatd. mengikat enzim
3. Reagen yang digunakan untuk mendeteksi kemampuanproduksi IAA cendawan adalah….a. Pikovskayab. Salkowskic. L-Tryptophand. Tricalsium fosfat
4. Medium Pikovskaya mengandung ….a. L. tryptophanb. Enzimc. Tricalsium fosfatd. Besi
5. Terbentuknya zona bening disekitar koloni solate cendawanendofit merupakan indikator kemampuan ….a. melarutkan fosfatb. memproduksi IAAc. memproduksi sidereford. memproduksi giberelin
6. Isolat cendawan edondofit memiliki kemampuan produksi

Cendawan Endofit
85
yang tinggi terhada jenis siderefore….a. salisilatb. katekolc. kaboksilatd. hidrosamat
7. Berikut ini adalah cendawan penghasil giberelin, kecuali…a. Fusarium,b. Peniciliumc. Aspergillusd. Bacillus
8. Komponen utama limbah pertanian adalah ….a. giberelinb. lignoselulosac. sidereford. fosfat
9. Komponen terbesar dalam lignoselulosa adalah…a. pektinb. lininc. hemiselulosad. selulosa
10. Enzim yang dihasilkan oleh mikroba dan dapat mendegradasiselulase yang tidak larut menjadi monosakarida dan disakarida sederhana larut dan dapat digunakan sebagai sumber energiadalah…
a. amilaseb. pectinasec. selulased. kitinase

86
Cendawan Endofit
Kunci Jawaban1. b1. b2. b3. c4. a5. b6. d7. b8. d9. c
Umpan Balik dan Tindak Lanjut Cocokkanlah jawaban Anda dengan kunci jawaban solate
yang terdapat pada bagian akhir unit ini. Hitunglah jawaban Anda yang benar. Gunakanlah rumus di bawah ini untuk mengetahui tingkat penguasaan Anda terhadap materi subunit 1.
Rumus: Jumlah jawaban Anda yang benar
Tingkat penguasaan = ---------------------------------------- x 100% Jumlah soal
Arti tingkat penguasaan yang Anda capai: 91 – 100% = baik sekali 81 – 89% = baik 71 – 79% = cukup < 70% = kurang
Bila Anda mencapai tingkat penguasaan 80% atau lebih,

Cendawan Endofit
87
Anda dapat melanjutkan dengan unit selanjutnya. Selamat untuk Anda! Tetapi apabila tingkat penguasaan Anda masih di bawah 80%, Anda harus mempelajari solate materi subunit 1 terutama bagian yang belum Anda kuasai.

88
Cendawan Endofit
Cendawan endofit merupakan cendawan yang ada dalam jaringan tanaman dan berasosiasi dengan tanaman tanpa menimbulkan gejala pada tanaman inang. Keberadaan cendawan endofit dalam jaringan tanaman memberikan manfaat positif terhadap pertumbuhan tanaman dan sebaliknya cendawan endofit juga mendapatkan manfaat dari tanaman inang.
Teknik isolasi cendawan endofit menggunakan jaringan tanaman berupa batang, daun, akar, mahkota dan buah serta biji. Teknik isolasi cendawan endofit diawali dengan sterilisasi permukaan, kemudian dilanjutkan dengan inkubasi, dan pemindahan isolat cendawan yang tumbuh pada permukaan jaringan tanaman ke medium PDA. Isolasi diakhiri dengan kegiatan purifikasi atau pemurnian isolate cendawan endofit dengan memindahkan setiap isolate cendawan endofit ke medium PDA yang baru hingga diperoleh isolat murni atau spora tunggal.
Persayaratan medium untuk pertumbuhan cendawan endofit pada dasarnya sama dengan persyaratan untuk pertumbuhan mikroba pada umumnya yaitu: pH media harus sesuai, steril dan tidak mengandung zat-zat penghambat serta mengandung semua unsur nutrisi yang mudah digunakan mikroorganisme. Jenis medium yang umum digunakan adalah PDA, CSA, CMC, GAN, MEA, CZA, OA, LCA, CMA. Medium tersebut dapat digunakan untuk kegiatan isolasi, identifikasi dan penyimpanan isolate.
Identifikasi isolat cendawan endofit dapat dilakukan
BAB VIIIPENUTUP

Cendawan Endofit
89
secara morfologi dan molekuler. Identifikasi isolat cendawan endofit secara morfologi berdasarkan pada karakter makroskopi isolate cendawan endofit seperti warna koloni, tipe pertumbuhan, tekstur permukaan. Pengamatan mikroskopis meliputi bentuk hifa, ada tidaknya rhizoid, bentuk sel reproduksi seksualnya dan aseksualnya. Identifikasi secara molekuler dapat menggunakan teknik PCR dan perunutan DNA. Jenis primer yang umum digunakan untuk Barkoding cendawan adalah pasangan primer NSI1/NLB4 dan ITS1/ITS4.
Metode penyimpanan cendawan endofit bertujuan untuk mengurangi lagu metabilisme dan memelihara biakan agar tetap survival saat akan digunakan. Metode penyimpanan cendawan dapat dilakukan dengan metode penyimpanan kultur berkala menggunakan minyak parafin, media agar dan aquades steril dengan tujuan penyimpanan jangka pendek, metode penyimpanan transfer kultur berkala untuk tujuan jangka pendek, metode penyimpanan kering dengan menggunakan silica gel anhidrit dan tau pasir kering steril, metode penyimpanan beku pada suhu -70 oC dan metode penyimpanan kering beku dengan metode Ldrying dan Freeze driying.
Cendawan endofit memiliki manfaat langsung dan tidak langsung terhadap pertumbuhan tanaman. Manfaat langsung seperti: menghasilkan fitohormen (IAA dan Giberlin), memproduksi siderefor, melarutkan fosfat, memproduksi enzim. Sedangkan manfaat tidak langsung seperti: meningkatkan ketahanan tanaman terhadap cekaman abiotik dan biotik. Cekaman abiotik seperti cekaman kekeringan, keracunan logam berat sedangkan cekaman biotik seperti hama dan. penyakit.

90
Cendawan Endofit
DAFTAR PUSTAKA
Afandhi, Aminudin, Fery Abdul Choliq, Havinda Anggrilika W.S., and Hagus Tarno. 2018. “Distribution of the Endophytic Fungi in Apple Leaves.” AGRIVITA Journal of Agricultural Science 40 (1): 91–100. https://doi.org/10.17503/agrivita.v40i1.1563.
Agusta. A. 2009. Biologi dan Kimia Jamur Endofit. Bandung: Institut Teknologi Bandung
Aly A. H., A. Debbab, and P. Proksch. 2011. Fungal endophytes: unique plant inhabitants with great promises. Appl Microbiol Biotechnol. 90:1829–1845
Amin N, Malik W, Sarbini G. 1997. Penggunaan cendawan endofit sebagai biofertilizer dan biopestisida dalam sistem pertanaman tomat. Pros. Seminar Perhimpunan Bioteknologi Pertanian Indonesia. Hal 360 -366
Ahmed, E., and S. J.M. Holmström. 2014. “Siderophores in Environmental Research: Roles and Applications.” Microbial Biotechnology 7 (3): 196–208. https://doi.org/10.1111/1751-7915.12117.
Akmalasari, Iva, Endang Sri Purwati, and Stia Dewi. 2013. Isolasi dan Identifikasi Jamur Endofit Tanaman Manggis ( Garcinia mangostana L .), 30 Biosfera 82–89.
Alori, Elizabeth T., Bernard R. Glick, and Olubukola O. Babalola. 2017. “Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture.” Frontiers in Microbiology 8 (JUN): 1–8. https://doi.org/10.3389/fmicb.2017.00971.
Amaria, Widi, Efi Taufiq, and Rita Harni. 2013. “Seleksi Dan

Cendawan Endofit
91
Identifikasi Jamur Antagonis Sebagai Agens Hayati Jamur Akar Putih (Rigidoporus Microporus) Pada Tanaman Karet.” Buletn RISTRI 4 (1): 55–64. https://doi.org/10.21082/jtidp.v4n1.2013.p55-64.
Anke, Heidrun, Josiane Kinn, Karl-Erik Bergquist, and Olov Sterner. 1991. “Production of Siderophores by Strains of the GenusTrichoderma.” Biology of Metals 4 (3): 176–80. https://doi.org/10.1007/BF01141311.
Ariyanto, Eko Famuji, Abdul Latief Abadi, and Syamsuddin Djauhari. 2013. “Keanekaragaman Jamur Endofit Pada Daun Tanaman Padi (Oryza Sativa L.) Dengan Sistem Pengelolaan Hama Terpadu (PHT) Dan Konvensional DI Desa Bayem, Kecamatan Kasembon, Kabupaten Malang.” Jurnal HPT 1 (2): 37–51.
Azevedo, Joao Lucio, Walter Maccheroni Jr, Jose Odair Pereira, and Welington Luiz de Araujo. 2000. “Endophytic Microorganisms: A Review on Insect Control and Recent Advances on Tropical Plants.” Electronic Journal of Biotechnology 3 (1): 40–65. https://doi.org/10.2225/vol3-issue1-fulltext-4.
Bae, H., Sicher, R.C., Kim, M.S., Kim, SH., Strem. D.M. Melnick, R.L. and Bailey, B.A., 2009. The beneficial endophyteTrichoderma hamatum isolate DIS 219b promotes growthand delays the onset of the drought response in Theobromacacao. Journal of Experimental Botany,60(11): 3279–3295.
Brasilica, 25(4), 815– 821. http://doi.org/10.1590/S0102-33062011000 400008
Cappucino, J. G., & Sherman, N. (2014). Manual Laboratorium Mikrobiologi, Edisi 8. EGC, Jakarta.
Caron, M., C.L. Patten and S. Ghosh. 1995. Effects of Plant

92
Cendawan Endofit
Promoting Rhizobacteria Pseudomonas putida GR-122 on the Physiology of Canolla Roots. PlantGrowth Reg Soci Am, 22nd proceeding
Clay, K. 1988. Fungal Endophytes of Grasses: A Defensive Mutualism Between Plants and Fungi. Ecology. 69 (1): 10-16
Crafts, Carol B., and Carlos O. Miller. 1974. “Detection and Identification of Cytokinins Produced by Mycorrhizal Fungi.” Plant Physiology 54 (4): 586–88. https://doi.org/10.1104/pp.54.4.586.
Dewi, Intan Ratna. 2008. “Peranan Dan Fungsi Fitohormon Bagi Pertumbuhan Tanaman.” Makalah, 1–43.
Diaz, Patricia L, James R Hennell, and Nikolaus J Sucher. 2013. “Chapter 14 Genomic DNA Extraction and Barcoding of Endophytic Fungi,” no. February 2012. https://doi.org/10.1007/978-1-61779-609-8.
Durham, NC. 2004. Armies of fighting fungi protect chocolate trees. www.rpi.edu/ajayan/locker/publication s/natureajayanjan202004.
Dwiyani, Rindang. 2015. Kultur Jaringan Tanaman. Journal of Chemical Information and Modeling. Vol. 53. https://simdos.unud.ac.id/ uploads/file_penelitian_1_dir/127d33b953fcbc98107a8b381e77d5b9.
Elawati N, Pujyanto S, Kusdiyantin E. 2018. Karakteristik dan Sifat Kinetika Enzim Kitinase Asal Jamur Entomopatogen Beauveria bassiana . J.Bioteknologi dan Biosains Indonesia. Vol (5) 1
Ezra D, Hess WH, Strobel GA: New endophytic isolates of M. albus, a volatile antibiotic-producing fungus. Microbiology 2004, 150: 4023-4031. 10.1099/mic.0.27334-0

Cendawan Endofit
93
Farida, Ida. 2012. “Siderofor Sebagai Agens Antagonis Ralstonia Solanacearum Pada Tomat.”
Fisher, P.J., Petrini, O., Petrini, L.E., Sutton, B.C., 1994. Fungal endophytes from the leaves and twigs of Quercus ilex L. from England, Majorea and Switzerland. New Phytol., 127: 133-137.
Glick, B. R. 1995. The enhancement of plant growth by free-living bacteria. Canadian Journal Microbiology 41: 109-117
G, Arun Kumar, Robert Antony A, and V. Rajesh Kannan. 2015. “Exploration of Endophytic Microorganisms from Selected Medicinal Plants and Their Control Potential to Multi Drug Resistant Pathogens.” Journal of Medicinal Plant Studies 3 (2): 49–57. https://doi.org/10.1207/s15327655jchn 2303_4.
Gazis, Romina, and Priscila Chaverri. 2010. “Diversity of Fungal Endophytes in Leaves and Stems of Wild Rubber Trees (Hevea Brasiliensis) in Peru.” Fungal Ecology 3 (3): 240–54. https://doi.org/10.1016/j.funeco.2009. 12.001.
Gusmiaty, M Restu, B Bachtiar, and SH Larekeng. 2019. “Gibberellin And IAA Production by Rhizobacteria From Various Private Forest.” IOP Conference Series: Earth and Environmental Science 270 (May): 012018. https://doi.org/10.1088/1755-1315/270/1/012018.
Hafsari, Anggita Rahmi, and Isma Asterina. 2013. “Isolasi Dan Identifikasi Kapang Endofit Dari Tanaman Obat Surian (Toona Sinensis).” Jurnal Istek 7 (2): 175–91.
Halo, Boshra A., Rashid A. Al-Yahyai, and Abdullah M. Al-Sadi. 2018. “Aspergillus Terreus Inhibits Growth and Induces Morphological Abnormalities in Pythium Aphanidermatum and Suppresses Pythium-Induced Damping-off of

94
Cendawan Endofit
Cucumber.” Frontiers in Microbiology 9 (FEB): 1–12. https://doi.org/10.3389/fmicb.2018.00095.
Hasan, H.A.H. 2011. “Gibberellin and Auxin Production by Plant Root-Fungi and Their Biosynthesis under Salinity-Calcium Interaction.” Plant, Soil and Environment 48 (No. 3): 101–6. https://doi.org/10.17221/4207-PSE.
Hussein, Khalid Abdallah, and Jin Ho Joo. 2019. “Zinc Ions Affect Siderophore Production by Fungi Isolated from the Panax Ginseng Rhizosphere.” Journal of Microbiology and Biotechnology 29 (1): 105–13. https://doi.org/10.4014/jmb.1712.12026.
Hartman, H.T., Kester, D.E., dan Davies, F.T. 2002. Plant Propagation. Principles and Practices 7th ed. Pearson Education INC. New Jersey.
Hilarino, M. P. A., Silveira, F. A. de O. e, Oki, Y., Rodrigues, L., Santos, J. C., Corrêa Junior, A., … Rosa, C. A. (2011). Distribution of the endophytic fungi community in leaves of Bauhinia brevipes (Fabaceae). Acta Botanica
Hubbar, M., J. Germinda, V. Vujanovic. 2012. Fungal endphytes improve wheat seed germination under heat and drought stress. J. Botany. 90(2): 137-149
Hyde KD, Soytong K. 2008. The fungal endo- phyte dilemma. Fungal Divers. 33:163-173.
I.C. Pimentel, C. Glienke Blanco, J. Gabardo, M. R. Stuart, J. L. Azevedo. 2006 Identification and colonization of endophytic fungi from soybean (Glycine max (L.) Merril) under different environmental conditions Braz Arch Biol Technol, 49, pp. 705-711
Idriati, Ninoek, Nandang Priyanto, and Radestya Triwibowo. 2010. “Penggunaan Dichloran Rose Bengal Chloramphenicol

Cendawan Endofit
95
Agar (DRBC) Sebagai Media Tumbuh Kapang Pada Produk Perikanan.” Pasca Panen Dan Bioteknologi Kelautan Dan Perikanan 5 (2): 117–21.
Ilyas, Muhammad. 2007a. “Isolasi Dan Identifikasi Mikoflora Kapang Pada Sampel Serasah Daun Tumbuhan Di Kawasan Gunung Lawu , Surakarta , Jawa Tengah.” Biodiversitas 8 (April): 105–10.
———. 2007b. Seri Panduan Teknik Preservasi Fungi.Jamilatun, Makhabbah, Nisa Azzahra, and Aminah Aminah.
2020. “Perbandingan Pertumbuhan Aspergillus Fumigatus Pada Media Instan Modifikasi Carrot Sucrose Agar Dan Potato Dextrose Agar.” Jurnal Mikologi Indonesia 4 (1): 168–74. https://doi.org/10.46638/jmi.v4i1.69.
Javvis, B. 1973. Comparison of an improved rose Bengal-chlortetracycline agar with other media for selective isolation and enumeration of mold and yeasts in foods. J. Appl.Bacteriol. 36(4): 723-727
Jutono 1980. Pedoman Praktikumn Mikrobiologi Umum. Yogjakarta: Fakultas pertanian UGM
Johnstone TC, Nolan EM. 2015. Beyond iron: non-classical biological functions of bacterial siderophores. Dalton Trans. 14: 6320-6339.
Tsavkelova, E.A., T.A. Cherdyntseva, and A.I. Netrusov. 2005. Auxin Production by Bacteria Associated with Orchid Roots. Microbiology, Vol. 74, No. 1, pp. 46-53
Kafrawi, Zahraeni Kumalawati, and Sri Mulyani. 2015. “Skrining Isolat Plant Growth Promoting Rhizobacteri (PGPR) Dari Pertanaman Bawang Merah ( Allium Ascalonicum ) Di Gorontalo.” Seminar Nasional Mikrobiologi Kesehatan Dan Lingkungan, no. 29: 132–39.

96
Cendawan Endofit
Khan, Abdul Latif, Syed Abdullah Gilani, Muhammad Waqas, Khadija Al-Hosni, Salima Al-Khiziri, Yoon Ha Kim, Liaqat Ali, et al. 2017. “Endophytes from Medicinal Plants and Their Potential for Producing Indole Acetic Acid, Improving Seed Germination and Mitigating Oxidative Stress.” Journal of Zhejiang University: Science B 18 (2): 125–37. https://doi.org/10.1631/ jzus.B1500271.
Khan, A.L., Hussain, J., Al-Harrasi, A., et al., 2013. Endo- phytic fungi: a source of gibberellins and crop resistance to abiotic stress. Crit. Rev. Biotech., 35(1):1-13. http://dx.doi.org/10.3109/07388551.2013.800018
King, A. D., Hocking, A.D., and Pitt. J.I. 1979. Dichloran-rose Bengal medium for enumeration and isolation of molds from foods. Appl.Environ. Microbiolo.27(5): 595-964
Kraemer, S.M., Crowley, D.E., and Kretzschmar, R. (2006) Geochemical aspects of phytosiderophore promoted iron acquisition by plants. Adv Agron 91: 1–46.
Kurniati, Fitri, Tini Sudartini, and Dikdik Hidayat. 2017. “Aplikasi Berbagai Bahan ZPT Alami Untuk Meningkatkan Pertumbuhan Bibit Kemiri Sunan (Reutealis Trisperma (Blanco) Airy Shaw).” Jurnal Agro 4 (1): 40–49. https://doi.org/10.15575/1307.
Legiastuti, Tuti Susanti, and Tri Aminingsih. 2012a. “Identifikasi Cendawan Endofit Menggunakan Teknik Polymerase Chain Reaction.” Jurnal Fitopatologi Indonesia 8 (2): 31–36. https://doi.org/10.14692/jfi.8.2.31.
———. 2012b. “Identifikasi Cendawan Endofit Menggunakan Teknik Polymerase Chain Reaction.” Fitopatologi Indonesia 8 (April): 31–36.
Leitão, Ana Lúcia, and Francisco J. Enguita. 2016. “Gibberellins

Cendawan Endofit
97
in Penicillium Strains: Challenges for Endophyte-Plant Host Interactions under Salinity Stress.” Microbiological Research 183: 8–18. https://doi.org/ 10.1016/j.micres.2015.11.004.
Lymar, E.S., B. Li and V. Renganathan. 1995.Purification and characterization of a cellulose- binding β- glucosidase from cellulose degrading culture of Phanerochaete chrysosporium. Appl. Environ. Microbiol., 61:2976-2980.
Little GN and Gordon MA. 1967. Survival of fungus cultures maintained under mineral oil for twelve years. Mycologia 59, 733-736.
Ludyasari, Ayu. 2005. “Pengaruh Suhu Annealing Pada Program PCR Terhadap Keberhasilan Amplifikasi DNA Udang Jari (Metapenaeus Elgans De Man, 1907) Laguna Segara Anakan, Cilacap, Jawa Tengah.”
Machmud, M. 2001. Teknik penyimpanan dan pemeliharaan mikroba. Buletin Agrobio 4(1):24-32.
Mastouri. F, T. Bjorkman, G.E. Harman, 2010. Seed Treatment with Trichoderma harzianum alleviates biotic, abiotik, and physiological stresses in germinating seeds and seedlings. Phytophatology 100 (11): 1213-1221.
Matzanke, B.F. (1991) Structures, coordination chemistry and functions of microbial iron chelates. In CRC Handbook of Microbial Iron Chelates. Winkelmann, G. (ed.). Boca Raton, FL, USA: CRC Press, pp. 15–64
Munifah I, Chasanah E, Fawzya YN. 2011. Screening of cellulolytic bacteria from Indonesia’s marine environment. Di dalam: Prosiding Seminar ISISM (International Seminar of Indonesian Society for Microbiology); Bogor, 26 Juni 2011. Bogor: Perhimpunan Mikrobiologi Cabang Bogor.

98
Cendawan Endofit
Matzanke, B.F. (1991) Structures, coordination chemistry and functions of microbial iron chelates. In CRC Handbook of Microbial Iron Chelates. Winkelmann, G. (ed.). Boca Raton, FL, USA: CRC Press, pp. 15–64
Munifah I, Chasanah E, Fawzya YN. 2011. Screening of cellulolytic bacteria from Indonesia’s marine environment. Di dalam: Prosiding Seminar ISISM (International Seminar of Indonesian Society for Microbiology); Bogor, 26 Juni 2011. Bogor: Perhimpunan Mikrobiologi Cabang Bogor.
Maratun Sholihati, Al, Maswati Baharuddin, and Santi. 2015. “Produksi Dan Uji Aktivitas Enzim Selulase Dari Bakteri Bacillus Subtilis.” Al Kimia, 78–90.
Murtiyaningsih, Hidayah, and Muhammad Hazmi. 2017. “Isolasi Dan Uji Aktivitas Enzim Selulase Pada Bakteri Selulolitik Asal Tanah Sampah.” Agritop 15 (2): 293–308. https://doi.org/10.3969/j.issn.1006-6896.2011.11.003.
Najmiyati, Erma, and Dominikus Dominikus H Akhadi. 2013. “Viabilitas Dan Kinerja Konsorsium Mikroba Pendegradasi Hidrokarbon Setelah Penyimpanan Dalam Pendingin Dan Penyimpanan Beku.” Jurnal Ecolab 6 (2): 81` – 89. https://doi.org/10.20886/jklh.2012.6.2.81-89.
Neilands JB. 1995. Siderophores: structure and fungtional of microbial iron transport compounds. The Journal of Biologycal Chemistry 270(45): 26723-26726.
Nio, S.A. dan F.E.F. Kandou. 2000. Respons pertumbuhan padi (Oryza sativa L.). sawah dan gogo pada fase vegetatif awal terhadap cekaman kekeringan. Eugenia 6:270-273.
Noble, Adam, Anna Kisiala, Amy Galer, Donna Clysdale, and R. J.Neil Emery. 2014. “Euglena Gracilis (Euglenophyceae) Produces Abscisic Acid and Cytokinins and Responds to

Cendawan Endofit
99
Their Exogenous Application Singly and in Combination with Other Growth Regulators.” European Journal of Phycology 49 (2): 244–54. https://doi.org/10.1080/09670262. 2014. 911353.
Norris D. 1944. Preservation of tube cultures of fungi and bacteria with liquid paraffin. Jour. Aust. Inst. Agri. Sci. 10, 77.
Octavia, Artha, and Sri Wantini. 2017. “Perbandingan Pertumbuhan Jamur Aspergillus Flavus Pada Media PDA (Potato Dextrose Agar ) Dan Media Alternatif Dari Singkong (Manihot Esculenta Crantz).” Jurnal Analis Kesehatan 6 (2): 625–31.
Omar SA (1998). The role of rock-phosphate-solubilizing fungi and vesicular-arbusular-mycorrhiza (VAM) in growth of wheat plants fertilized with rock phosphate. World J. Microbiol. Biotech. 14: 211– 218.
Onions AHS. 1971. Preservation of fungi. In: C Booth (ed): Methods in Microbiology 4 London and New York, Academic, 113-151.
Patil, Mohini G, Jyoti Pagare, Sucheta N Patil, Amanpreet K Sidhu, and K T H M College. 2015. “Extracellular Enzymatic Activities of Endophytic Fungi Isolated from Various Medicinal Plants” 4 (3): 1035–42.
Pradhan, N, and L B Sukla. 2005. “1 ’(0.573) Solubilization of Inorganic Phosphates by Fungi Isolated from Agriculture Soil.” African Journal of Biotechnology 5 (10): 850–54.
Pratama, Ilham, Linda Advinda, and Mades Fifendy. 2018. “Pengaruh Sumber Karbon Terhadap Produksi Siderefore Dari Bakteri Pseudomonad Fluoresen.” Bioscience 2 (2): 50. https://doi.org/10.24036 /020182210 406-0-00.
Pratiwi RS, Susanto TE, Wardani YAK, Sutrisno A (2015) Enzim

100
Cendawan Endofit
kitinase dan aplikasi di bidang industri: Kajian Pustaka. J Pangan Agroindustri 3: 878-887
Prihatiningsih, Nur, Heru Adi Djatmiko, and Puji Lestari. 2017. “Aktivitas Siderofor Bacillus Subtilis Sebagai Pemacu Pertumbuhan Dan Pengendali Patogen Tanaman Terung.” HPT Tropika 17 (2): 170–78.
Qadri, Masroor, Sarojini Johri, Bhahwal A. Shah, Anamika Khajuria, Tabasum Sidiq, Surrinder K. Lattoo, Malik Z. Abdin, and Syed Riyaz-Ul-Hassan. 2013. “Identification and Bioactive Potential of Endophytic Fungi Isolated from Selected Plants of the Western Himalayas.” SpringerPlus 2 (1): 1–14. https://doi.org/10.1186/2193-1801-2-8.
Rahayu, Fitri, Saryono, and Titania T Nugroho. 2015. “Isolasi DNA Dan Amplifikasi PCR Daerah ITS RDNA Fungi Endofit Umbi Tanaman Dahlia (Dahlia Variabilis) LBKURCC69.” Jom Fmipa 2 (1). https://doi.org/10.3724/SP.J.1042.2011.00117.
Ramdan, EP, Widodo Widodo, ET Tondok, S Wiyono, and SH Hidayat. 2014. “Cendawan Endofit Nonpatogen Asal Tanaman Cabai Dan Potensinya Sebagai Agens Pemacu Pertumbuhan.” Jurnal Fitopatologi Indonesia 9 (5): 139–44. https://doi.org/10.14692/jfi.9.5.139.
Razie, Fakhrur, Iswandi Anas, Atang Sutandi, Lukman Gunarto, and Sugiyanta Sugiyanta. 2019. “Aktivitas Enzim Selulase Mikroba Yang Diisolasi Dari Jerami Padi Di Persawahan Pasang Surut Di Kalimantan Selatan.” Jurnal Ilmu Tanah Dan Lingkungan 13 (2): 43. https://doi.org/10.29244 /jitl.13.2.43-48.
Russo, María L, Sebastián A Pelizza, Marta N Cabello, Sebastián A Stenglein, María F Vianna, and Ana C Scorsetti. 2016.

Cendawan Endofit
101
“Endophytic Fungi from Selected Varieties of Soybean ( Glycine Max L . Merr .) and Corn ( Zea Mays L .) Grown in an Agricultural Area of Argentina.” Revista Argentina de Microbiología 48 (2): 154–60. https://doi.org/10.1016/j.ram.2015. 11.006.
Saha, B.C., “Lignocellulose Biodegradation and Applications in Biotechnology. In: Lignocellulose Biodegradation”, Saha BC, Hayashi K (Ed.). American Chemical Society, Washington DC. p2-34, 2004
Setiawati, M.R, Ufah N, Hindersah R, Suryatmana P. 2019. Peran Mikroba Dekomposer Selulolitik dari Sarang Rayap dalam Menurunkan Kandungan Selulosa Limbah Pertanian Berselulosa Tingg. Jurnal Soilrens. Vol 17 (2)
Schwyn, B., and Neilands, J.B. (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160: 47–56.
Sharma A, & Johri BN. 2003. Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9, in maize (Zea maysL.) under iron limiting conditions. Microbiol. Res. 158(3): 243–248
Suciatmih, S, and R Rachmat. 2005. “Pengujian Survival Jamur Yang Dipreservasi Dalam Air Dan Parafin Cair [ Survival Testing of Preserved Fungi in Water and Liquid-Paraffin ].” Berita Biologi 7 (5): 241–48.
Sukmadi, R Bambang. 2013. “Aktivitas Fitohormon Indole-3-Acetic Acid (Iaa) Dari Beberapa Isolat Bakteri Rizosfer Dan Endofit.” Jurnal Sains Dan Teknologi Indonesia 14 (3): 221–27.https://doi.org/10.29122/ jsti.v14i3. 930.
Sinaga, M. S. 2003. Ilmu–Ilmu PenyakitTumbuhan. Bogor: Penebar Swadaya

102
Cendawan Endofit
Sivasakthivalen P dan D. Stella. 2012. Studies on Phytohormon Producing Potential of Agriculturally Beneficial Microbial (ABM) Isolats from Different Rhizospers Soils of Sunflower in Tamil Nadu. International Journal of Pharmaceutical and Biological Archives 2012; 3(5): 1150-1156.
Smith D H, Lewis FH and Fergus CL. 1970. Long term preservation of Botryosphaeri ribis and Dibotryon morbosum. Plant Disease Reporter 54,217-218.
Spichal, L. (2012). Cytokinins – recent news and views of evolu- tionally old molecules. Functional Plant Biology, 39: 267–284.
Stirk, W.A., Van Staden, J., Novak, O., Dolezal, K., Strnad, M., Dobrev, P.I., Sipos, G., Ordog, V. & Balint, P. (2011). Changes in endogenous cytokinin concentrations in Chlorella (Chlorophyceae) in relation to light and the cell cycle. Journal of Phycology, 47: 291–301.
Subba-Rao, S.N.S, 2010. Soil Microorganisms and Plant Growth (in bahasa Indonesia). Penerbit Universitas Indonesia. Jakarta
Shukla, N. R. P. Awasthi, L. Rawat, J. Kumar. 2012. Biochemical and Physiological Responses of Rice (Oryza savita L.) as Influenced by Trichoderma harzianum Under Drought Stress. Plant Physiology and Biochemistry 54: 78-88
Suryani, Isti, Agus Santoso, and M Juffrie. 2010. “Penambahan Agar-Agar Dan Pengaruhnya Terhadap Kestabilan Dan Daya Terima Susu Tempe Pada Mahasiswa Politeknik.” Jurnal Gizi Klinik Indonesia 7 (2): 85–91.
Syamsia. 2016. “Isolasi Dan Identifikasi Cendawan Endofit Tanaman Padi Lokal Enrekang.” Agrotan 2 (2): 61–67.
Syamsia, Tutik Kuswinanti, Elkawakib Syam’un, and Andi

Cendawan Endofit
103
Masniawati. 2015. “The Potency of Endophytic Fungal Isolates Collected from Local Aromatic Rice as Indole Acetic Acid (IAA) Producer.” Procedia Food Science 3: 96–103. https://doi.org/10.1016/j.profoo.2015.01.009.
Syamsia, S., A. Idham, A. Patappari, and N. Noerfitryani. 2019. “Isolation and Amplification DNA on Endophytic Fungi of Local Aromatic Rice Enrekang.” IOP Conference Series: Earth and Environmental Science 270 (1): 012054. https://doi.org/10.1088/1755-1315/270/1/012054.
Syamsia, S., Abubakar Idhan, Amanda Patapparai, and Noerfitryani Noerfitryani. 2019. “Molecular Identification of Endophytic Fungi from Local Rice and Growth Test on Several Types of Culture Media.” International Journal of Agriculture System 7 (2): 89–99. https://doi.org/10.20956/ijas.v7i2.2031.
Syamsia, S., Abubakar Idhan, Amanda Patappari, and Noerfiryani Noerfitriayani. 2020. “Potensi Cendawan Endofit Padi Lokal Sulawesi Selatan Sebagai Penghasil Siderefor. Jurnal Galung Tropika 9 (2): 187–94.
Syamsia, S., Abubakar Idhan, Amanda Patappari, 2020. Produksi Giberelin dan IAA Cendawan Endofit Asal Padi Lokal Sulawesi Selatan. Makalah yang disampaian pada seminar nasional Perhimpunan Agronomi Indonesia 2020 secara daring pada tanggal 17 Oktober 2020.
Vassileva M, Vassilev N, Fenice M, Federici F (2001). Immobilized cell technology applied in solubilization of insoluble inorganic (rock) phosphate and P plant acquisition. Bioresource Technol. 79: 263- 271
Wang Y, H.N. Brown, D.E. Crowley and P.J. Szanielo. 1993. Evidence for Direct Utilization of a Siderophore,

104
Cendawan Endofit
Ferrioxamine B, in Axenically Grown Cucumber. Plant Cell Environ., 16:579- 585.
Wilia, Weni, Alia, Yulia Novita, 2011. TriasEksplorasi Cendawan Endofit Dari Beberapa Varietas Kedelai Sebagai Agens Pemacu Pertumbuhan Tanaman. Jurnal Penelitian Universitas Jambi. Volume 13 (1): 33-38)
White T, Bruns T, Lee S, et al (1990) Amplifi cation and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis M, Gelfand D, Sninsky J, & White T (ed) PCR Protocols: A Guide to Methods and Applications. Academic Press Inc, New York
Wiraatmaja, I Wayan. 2017. Zat Pengatur Tumbuh Giberelin Dan Sitokinin. Bahan Ajar.
Wulandari, N. F. 2019. “Isolation and Preservation of Microfungi from Indonesia Islands.” IOP Conference Series: Earth and Environmental Science 308 (1): 1–6. https://doi.org/10.1088/1755-1315/308 /1/ 012079.
Yadav, Anurag, and Kusum Yadav. 2017. “Exploring the Potential of Endophytes in Agriculture : A Minireview” 6 (4): 102–6. https://doi.org/10.15406/apar.2017.06.00221.
Yulianti, Titiek. 2012. “Menggali Potensi Endofit Untuk Meningkatkan Kesehatan Tanaman Tebu Mendukung Peningkatan Produksi Gula.” Perspektif 11 (2): 111–22.
Yulinery, Titin, and Ratih M.Dewi. 2016. “Preparasi Deinococcus Radiodurans Dan Khamir Dalam Material Kecap L-Drying Sebagai Bahan Uji Profisiensi.” Jurnal Teknologi Lingkungan 13 (1): 93. https://doi.org /10.29122/jtl.v13i1.1409.
Yunianto, Prasetyawan, Syofi Rosmalawati, Indra Rachmawati, Wahyudi Priyono Suwarso, And Wahono Sumaryono.

Cendawan Endofit
105
2012. “Isolation and Identification of Endophytic Fungi from Srikaya Plants (Annona Squamosa) Having Potential Secondary Metabolites as Anti-Breast Cancer Activity.” Microbiology Indonesia 6 (1): 23–29. https://doi.org/10.5454 /mi.6.1.4.
Yuspradana, Rangga, Suryo Wiyono, and Rahayu Widyastuti. 2017. “Rice Resistance-Treated with Endophyte Fungi Against Drought Stress.” Internasional Journal of Sciences : Basic and Applied Research 36 (3): 256–70.
Zhang, H.W., Y.C. Song dan R.X. Tan, 2006. Biology and chemistry of encophytes. Natural Products Reports. 23: 753-771

106
Cendawan Endofit
LAMPIRAN-LAMPIRAN

Cendawan Endofit
107
Lampiran 1. Hasil riset, publikasi pada Procedia Food Science. https://doi.org/10.1016/j.profoo.2015.01.009, Judul The Potency of Endophytic Fungal Isolates Collected from Local Aromatic Rice as Indole Acetic Acid (IAA) Producer
The Potency of Endophytic Fungal Isolates Collected from Local Aromatic Rice as Indole Acetic Acid (IAA) Producer
Syamsiaa, Tutik Kuswinantib, Elkawakib Syam’unb and Andi Masniawatic
a. Agriculture Faculty of Muhammadiyah University, Makassarb. Agriculture Faculty of Hasanuddin University, Makassar
c. Dept. of Biology, Faculty of Natural Science, Hasanuddin University, Makassar
AbstractEndophytic fungi are fungi that live in plant tissues without causing disease symptoms and abnormalities in plants. This study aims to obtain endophytic fungal isolates from local aromatic rice and screening of their ability to produce the IAA hormone as a plant growth promoter. Totally 16 endophytic fungal isolates were isolated from aromatic rice tissue of Pulu Mandoti. Ten isolates obtained from stem, three isolates from the root and three isolates from leaf tissue. The ability of endophytic fungal isolates in producing hormones IAA varied from 0.635 to 2.651 mgl-1. Similarly, the ability to dissolve phosphate also varied from 0.005 – 3.719 mgl-1, and there is only 1 isolates that caused abnormal growth in rice seedlings.

108
Cendawan Endofit
Key words: endophytic fungi, local aromatic rice, IAA
INTRODUCTION South Sulawesi is one of the aromatic rice producing areas in
Indonesia. Pulu Mandoti has higher level of fragnant than other aromatic rice in this area, and cultivated only by people in the District Salukanang Enrekang.
Endophytic fungi are microorganisms that presence in plant tissues or organs such as seeds, leaves, flowers, twigs, stems and roots. Various functional compounds can be produced by endophytic fungi, and can act as anti cancer, antiviral, antibacterial, antifungal, plant growth hormones, insecticides and others [1.2] Indole acetic acid is essential compound for the growth and development of roots and shoots, many microbes including Plant Growth Promoting Rhizobacteria (PGPR) produce IAA [1]. Soil fungi and endophytes secrete plant growth-promoting substances such as indole acetic acid (IAA) and gibberellins [3.4.5]
Auxin was first isolated and characterized as plant growth, and indole-3-acetic acid (IAA) is a type of auxin [6.7]. Most of the genus Trichoderma produce auxin indole acetic acid (IAA), with or without precursor L-tryptophan [8]. Trichoderma isolated from rhizospher were more efficient in producing IAA compared T. asperellum T211 [9]. Deshwal et al. [10.11], observed that IAA produced by Pseudomonas aeruginosa MR-9 increases plant height, dry weight, number of nodules per plant, nodule fresh weight of Mucuna pruriens as many as 184, 124, 139, 180% compared with controls.
Essentially Phosphorus in the soil is presence in unavailable form for plants, and organisms associated with the plants can help in dissolving the mineral P to facilitate absorption by plants. The

Cendawan Endofit
109
fungus has been reported to dissolve P by producing of organic acids and is known to have a higher efficiency than bacteria in dissolving of phosphorus [12] The main objective of this research is to obtain endophytic isolates from local aromatic rice that has the ability to produce IAA hormones and dissolved phosphorus.
MATERIALS AND METHODSIsolation of Endophytic Fungus
Local aromatic rice plant samples used in this study is Pulu Mandoti which is aromatic rice that has the most fragrant aroma and the highest economic value among 8 types of aromatic rice that is cultivated by farmers in this regions. Samples taken from rice plantation located in District Salukanang. Isolation of endophytic fungi carried out on the roots, stems and leaves of the local aromatic rice plants. Sterilization plant parts done gradually by soaking them for 60 seconds in 70% ethanol, 3% NaOCl for 60 seconds, and 70% ethanol for 30 seconds. Then rinsed four times with sterile distilled water dried on sterile filter paper. Cut a small part of the plant to be grown in PDA medium.
Extract preparations Endophytic Fungus Endophytic fungi isolates that have been sucultured on PDA
(Potato Dextrose Agar) and incubated for 7 days. Five disks of fungal colony put on to liquid medium Potato Dextrose Broth (PDB) were incubated at 28o C in a shaker with 150 rpm/min for 7 days, and centrifuged at 5000 rpm for 25 min. The supernatan transferred in to new flask and the pellet was removed.
Detection and Quantification of IAA Capability of endophytic fungi in producing IAA hormones

110
Cendawan Endofit
was done using the method used by Bhagobathi and Joshi (2009) [13]. Supernatan of endophytic fungi mixed with Salkowsky reagent then incubated for 20 minutes. Observation of the color change before absorbance was measured using a spectrophotometer with a wavelength of 535 nm. IAA concentration of each isolate was compared to a standard curve.
Phosphate solubilizing ability The isolates were tested for their quantitative ability in
solubilizing phosphate by using Pikovskaya broth medium with Ca3(PO4)2 as the phosphate source [14]. Materials of Pikovskaya broth medium are glucose 10 g; Ca3(PO4)2 5 g; (NH4)2 SO4 0,5 g; MgSO.2H2O 0,1 g; MnSO4 25 mg; FeSO425 mg; KCl 0,2 g, yeast extract 0,5 g; and agar 15 g, dissolved in sterile water until volume of 1 l. Pipette 30 ml of the suspension and put in the Erlenmeyer, containt Pikovskaya broth medium, and incubated in rotary shaker at 150 rpm for 7 days. Filter the 20 ml culture with filter paper Whatman No. 42. Filtrate was centrifugated at 1000 rpm for 15 minutes, 5.0 ml of supernatan then poured into test tubes, added with 0.5 ml of concentrated reagents P (12 g ammonium molibdat, 0.277 g kaliumantimoltartat) and Reagent dye concentrated (0.53 g ascorbic acid), shaked for a few minutes, and let it standed for 30 minutes. The absorbance of solution was measured with the spectrophotometer at a wavelength of 693 nm. In the same way was done in the Erlenmeyer flask containing Pikovskaya broth medium uninoculated fungi as a control.
Fungal Isolate as Plant Growth Promoting Fungi (PGPF) The role of endophytic fungi as plant Growth Promoting
Fungi was tested against rice plants according to Hermawati

Cendawan Endofit
111
(2007) [15]. Rice seeds soaked in 1% NaOCl for 1 minute to remove pathogens and contaminants on the viability of the seed. Seeds soaked in a suspension of endophytic fungi for 1 day. Rice seed germinated in a petri dish that has been coated with 2 sheets of filter paper and moistened. Control were made by soaking the seeds in sterile water. The parameters measured were the percentage of germination and abnormal growth in rice seedlings for 7 days.
RESULTS AND DISCUSSION Isolation of endophytic fungi from local aromatic rice plants
obtained 16 isolates. Ten isolates from stem (KN1, KN2, KN3, KN 4, KN5, KN 6, KN7, KN11, KN 14, KN15), three isolates from leaf (KN9, KN10, KN13) and three isolates from roots KN8, KN12, KN16). This suggests that the presence of endophytic fungi in plant tissue is not the same and spread randomly. This is consistent with research from Stofwal that the existence of the type and number of endophytic fungi on each part of the plant is not the same [16.17]. Besides being very diverse in nature, these endophytes are a source of novel bioactive secondary metabolites [18.19]
Detection and Quantification of IAA Endophytic fungi isolates showed the highest IAA
production is isolate KN10 (2.651 mgl-1) and lowest KN6 isolates (0.635 mgl-1). Lower than the results of research Nenwani [12], F1 fungal isolates were able to produce IAA 11.45 μgml-1, also production IAA by bacteria in the presence of 5 mgml-1 tryptophan is 7.3 to 32.8 mgml-1 and 2.68 to 10.8 in the condition without tryptophan mgml-1 [20,21]. Similarly, bacteria from maize roots

112
Cendawan Endofit
in vitro resulted in the highest IAA on the KB3 isolates 1.1255 ppm [22]. The study of bacterial isolates in the rhizosphere of onion plants produce IAA Sulawesi varies from 0.76 to 2.33 ppm [23]. The highest IAA production produced by B. subtilis isolates ME 105, B. amyloliquefacaciens subsp. plantarum ME 3, P. polymyxa, and B. amyloliquefaciens subsp. plantarum ME8 (188, 151.9, 108.1 and 107 μgml-1) [24]
IAA produced by bacteria can promote plant growth by increasing the number of root hairs and lateral roots [25,26]. Endophytic fungi can increase the germination percentage of Vigna radiata and Cicer arietium by 95 and 87% compared to controls [13]. Gibberellins and auxin may play an important role in plant growth, reproduction, metabolism and response to various environmental cues [27].
Measurement of the IAA concentration produced by each isolate using spectrophotometer showed varying results, namely 0.635 – 2.651 mgl-1. (Table 1).

Cendawan Endofit
113
Figure 1. Abnormally growth of rice sprouts after inoculation with endophytic fungal isolates
Ability of Endophytic Fungal Isolate in Solubilize Phosfat Quantitative measurement using spectrofotometer at a
wavelength of 693 nm showed that sixteenth endophytic fungal isolates has the ability to dissolve phosphate that varies from 0.005 – 3.719 mgl-1 (Table 2). There are 7 isolates showed high phosphate solubility there are KN9, KN13, KN11, KN9, KN6, KN2 and KN2. This is accordance with the results of Pradhan and Sukla, [28,29] that found Aspergillus sp and Penicillium sp can sequentially dissolving 480 mg ml-1 of phosphate from 0.1 tricalsium phosphate (TCP) after 4 days.
The ability of isolates act as PGPF through applications on rice seed showed that of 16 isolates only 1 isolate (KN15) that caused abnormally growth on rice sprouts after 1-7 days. (Figure 1).

114
Cendawan Endofit
CONCLUSION There are 16 isolates of endophytic fungi from local aromatic rice that has the ability to produce the IAA hormone. The ability of tested isolates in producing IAA, varried from 0.635 to 2.651 mgl-1. Highest IAA production was shown by isolate KN10 (2.651 mgl-1). Similarly, the ability to dissolve phosphate also varied from 0.005 – 3.719 mgl-1. And there is only 1 isolates that showed abnormally growth in rice seedlings.
ACKNOWLEDGEMENTS Thanks to the Directorate of Higher Education for funding this research through a Doctoral Dissertation Research Grant through Kopertis DIPA IX fiscal year 2014.
REFERENCES [1] Strobel GA. Natural products from endophytic microorganism.
Journal of Natural Products. 2004; 67: 257-268. [2] Noverita, Dinah Fitria, Ernawati Sinaga. Isolation and
antibacterial activity assay of fungal endophyte of leaves and Rhizome Zingiber ottensii. (in Indonesia). Jurnal Farmasi Indonesia 2009; 4: 171 -176
[3] Khan, A. L., M. Hamayun, N. Ahmad, J. Hussain, S. M. Kang, Y.H. Kim. Salinity stress resistance offered by entophytic fungal interaction between Penicillium minioluteum LHL09 and Glycine max. L. J. Microbiol. Biotechnol. 2011; 21: 893-902.
[4] Chutima, R. and S. Lumyong. Production of indole-3-acetic acid by Thai native orchid- associated fungi. Symbiosis 2012;56: 35-44.
[5] Radhakrishnan, Ramalingam, kang-bo shim, byeong-won

Cendawan Endofit
115
lee, chung-dong Hwang, Suk-Bok Pae, Chang-Hwan Park, Sung-Up Kim, Choon-Ki Lee, and In-Youl Baek. IAA-producing Penicillium sp. NICS01 triggers plant growth and suppresses Fusarium sp.-induced oxidative stress in sesame (Sesamum indicum L.) J. Microbiol. Biotechnol. 2013;23: 856– 863
[6] Nakamura A, Umemura I, Gomi K. Production and characterization of auxin-insensitive rice by overexpression of a mutagenized rice IAA protein. Plant J. 2006; 46: 297-306
[7] Bilkay I.S, S Karakoc, N Aksöz, Indole-3-acetic acid and gibberellic acid production in Aspergillus niger. Turk J Biol 2010; 34: 313-318
[8] Oliveira AG, Junior AFC, Santos GR, Miller LO, Chagas LFB. Potencial de solubilizacão de fosfato e producão de AIA por Trichoderma spp. Rev. Verde. 2012; 7:149-155
[9] Resende. M. P, I. C. M C. Jakoby, L C R dos Santos, M. A Soares, F. D Pereira, E.L Souchie, and F G Silva, Phosphate solubilization and phytohormone production by endophytic and rhizosphere Trichoderma Isolates of Guanandi (Calophyllum brasiliense Cambess). Afr. J. Microbiol. 2014;27 :2616-2623.
[10] Deshwal, V.K., Devi, M.S., Bhajanka, N., Mistri, J., Bose, A. and Saini, N. Pseudomonas aeruginosa strains and their role in plant growth promotion in medicinal plant. global j. appl. agri.res.2011;1: 49-55.
[11] Deshwal V.K., and P Kumar. Plant growth promoting activity of Pseudomonads in rice crop. int.j.curr.microbiol.app.sci.2013;2(11): 152-157
[12] Nenwani.V, P Doshi, T. Saha and S. Rajkumar. Isolation

116
Cendawan Endofit
and Characterization of a fungal isolatae for Phosphate solubilization and plant growth promoting. Activity Journal of yeast and fungal research.2011; 1(1) pp. 009-014
[13] Bhagobaty R.K. and S.R. Joshi. Promotion of seed germination of Green gram and Chick pea by Penicillium verruculosum RS7PF, a root endophytic fungus of Potentilla fulgens L. Advanced Biotech. 2009.
[14] Subba-Rao, S.N.S, 2010. Soil Microorganisms and Plant Growth (in bahasa Indonesia). Penerbit Universitas Indonesia. Jakarta.
[15] Hermawati H. Effect of Endophytic Fungus of the Biology and Population Growth Aphis gossypii Glov (Homoptera: Aphididae) in pepper. (in bahasa Indonesia). Skripsi. IPB. 2007
[16] Stovall, M.E. An investigations of the fungus Balansia cyperi and its effect on purple nutsedge, Cyperus Rotundus. 1987.
[17] Sunariasih N P L. I Ketut Suada and Ni Wayan Suniti. Identification of Endophytic Fungi from Rice Grain and It’s Inhibiting Ability by In Vitro Against Pyricularia oryzae Cav. E- Jurnal Agroekoteknologi Tropika .2014; 3(2): 51-60
[18] Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology. 2007;88: 541–549.
[19] Sun, X.; Guo, L.D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 201; 47: 85–95.
[20] Ahmad, F., I. Ahmad, and M.S. Khan. 2005. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence

Cendawan Endofit
117
of tryptophan. Turk J. Biol.2005;29: 29-34. [21] Widiastuti H, Siswanto, dan Suharyanto. Characterization
and Selection of Azotobacter sp. In Enhancing Seed Germination and Growth of Plant. Buletin Plasma Nutfah. 2010;16 (2)
[22] Khairani.G. Isolation and Ability Test of Endofphytic Bacteria Producing IAA (indole Acetic Acid) Hormone from Crops Maize Root (Zea mays L) (in Indonesia). 2010.
[23] Kafrawi, Baharuddin, E. L. Sengin and A. Rosmana. Screening of Free-Living Indole Acetic Acid Producing Rhixobaceria From Shallot Rhizosheres In The Island of Sulawesi. International Journal of Scentific and Technoly Research. 2014; 3:118-121
[24] El-Meleigi, M. A, Ahmed A. Al-Rogaibah, Gmal H. Ibrahim and Khaled A. Al Gamhan, Role of Antibiosis and production of Indole-3-Acetic acid by bacilli strains in suppression of root pathogens and growth promotion of Alfalfa seedlings Int. J. Curr. Microbiol. App. Sci. 2014; 3 (6):685-696
[25] Okon Y and YKapulnik. Development and function of Azospirillium inoculated Roots. Plant and Soil 1986;90: 3-16
[26] Huen E, Screening of Soil Bacteria for Plant Growth Promotion Activities in vitro. Indonesian Journal of Agricultural Science. 2003; 4 (1): 27-31
[27] Waqas M, A L. Khan, M. Kamran, M. Hamayun, SM Kang, YH Kim and IJ Lee.Endophytic Fungi Produce Gibberellins and Indoleacetic Acid and Promotes Host-Plant Growth during Stress. Molecules. 2012;17: 10754-10773
[28] Pradhan N and Sukla LB. Solubization of inorganic phosphates by fungi isolated from agriculture soil. Arf. J.

118
Cendawan Endofit
Biotechnol. 2005;5 (10): 850 -854 [29] Handayani D. Potency of Aspergillus and Penicillium from
Dipterocarp Leaf Litter as Phosphate Solubilizer and Root Endosymbiont. (in Bahasa Indonesia) Thesis IPB.2011. p.71.

Cendawan Endofit
119
Lamprian 2. Hasil riset, publikasi pada IOP Conf. Series: Earth and Environmental Science 270 (2019) 012054 doi:10.1088/1755-1315/270/1/012054. Judul: Isolation and Amplification DNA on Endophytic Fungi of Local Aromatic Rice Enrekang
Isolation and Amplification DNA on Endophytic Fungi of Local Aromatic Rice Enrekang
S Syamsia1, A Idham2, A Patappari3, N Noerfitryani4
1,2Agrotechnology, Faculty of Agriculture, Muhammadiyah University Makassar, Jalan Sultan Alauddin No. 259, Makassar, 90221, South Sulawesi, Indonesia. 3,4Agribusiness, Faculty of Agriculture, Muhammadiyah University Makassar, Jalan Sultan Alauddin No. 259, Makassar, 90221, South Sulawesi, Indonesia.
E-mail: [email protected]
Abstract. Local rice is one of the biodiversity that is currently not explored in detail. The exploration of endophytic fungi of local rice in South Sulawesi was carried out to find out the types of endophytic fungi in the local rice plant. Plant samples were taken from local rice plantations in the Tator, Enrekang, and Luwu regencies. DNA isolation and PCR amplification were the initial stages that need to be carried out in molecular identification. DNA fungi extraction was performed by using DNesay DNA extraction KIT. The result of DNA fungi isolation was electrophoresed with 0.8% agarose gel in TAE buffer (Tris Acetate EDTA). DNA isolation was successfully carried out by the formation of DNA bands. DNA amplification of two endophytic

120
Cendawan Endofit
fungi using the primary pairs of ITS1 and ITS4, with condition PCR of pre- denaturation at 95°C for 5 minute, denaturation at 94°C for 1 minute, annealing at 55,4°C for 1 minute, elongation at 72°C for 1 minute, final elongation at 72°C for 10 minute. The denaturation stage until the extension was repeated 35 times and produced DNA fragments with a molecular size of 700 bp and 600 bp.
1. Introduction Indonesia is the second most biodiverse mega-biodiversity
country in the world. This high amount of biodiversity is an invaluable asset that can be used for the welfare of the people. However, the utilization of this natural potential is still constrained by the lack of information and data regarding biodiversity potential.
Microorganisms are one of the biodiversity that needs to be explored in detail to explore its potential in the development of science and technology. Endophytic fungi are one of the microorganisms that have high diversity.
Endophytic fungi are fungi that live in plant tissues without showing symptoms. The highest endophytic concentration is found in crowns, stems, and leaves, while few live in at the roots — mycelia that grow between plant cells, mostly in the protective layer of leaves and reproductive structures. When the host is in the form of seeds, endophytes infect and spread from the plant part of the outer layer into the seed. This shows how endophytes move from plants in a seed production area. When seeds germinate and grow, endophytes infect and spread to the growth tissue of the host plant.
Endophytic fungi can be isolated from various types of

Cendawan Endofit
121
plants, one of which is rice. South Sulawesi is one of the local rice-producing regions in Indonesia. Local rice that is quite well known includes Pulu Mandoti from Enrekang, Pare Ambo from Toraja and Tarone from Luwu.
Some endophytic fungi have been isolated from rice plants including; 1) Nigrospora sp is an endophytic fungus which most often colonizes at rice stem midribs [1]; 2) Enrekang local rice endophytic fungi that were successfully isolated and morphologically identified are: Aspergillus sp1, Aspergillus sp2, Aspergillus candidus and Acremonium [2]; 3) Endophytic fungi have been isolated from rice plants and identified morphologically and have the potential as biocontrol agents from disease: Fusarium sp, Cladosporium sp, Phoma sp, Penicillium sp [3].
Identification of endophytic fungi from dahlia tubers has been successfully identified molecularly: Phialophorafa stigmata [4]. Based on the results of the analysis using ITS molecular markers, 4 isolates were identified as genera Lasiodiplodia, 4 isolates as Didymellaceae family, 10 isolates as the Phomopsis, 5 isolates as the genus Colletotrichum, 1 isolate as the genus Nemania. 1 isolate as genus Xylaria [5]. This study aimed to isolate the DNA of local Enrekang endophytic fungi and amplify the isolated DNA.
2. Materials and Methods 2.1. Isolation and purification of Endophytic Fungi
Samples of local rice plants were taken from local rice plants in Enrekang area. Methods of isolating endophytic fungi following the modified Rodriques method [6]. Plant sterilization was carried out at stages by soaking for 60 seconds in 70% ethanol, 3% NaOCl for 60 seconds and 70% ethanol for 30

122
Cendawan Endofit
seconds, then rinsed four times with sterile distilled water and dried on sterile filter paper. Plant parts were cut in small pieces and placed on potato dextrose agar (PDA) media and incubated at room temperature. Mycelium that growth on the roots, leaves and stems were grown on new PDA media until pure isolates was obtained. 2.2. Isolation of Fungi Endophytic DNA
DNA fungi extraction was carried out using DNeasy Plant Mini kit (QIAGEN) DNA extraction KIT. The extraction process was carried out following the manufacturer’s instructions. The fungi were mashed using a hitter. The solution is then incubated in a water bath. The solution was incubated in a water bath for 30 minutes at 65 °C. The solution was centrifuged to separate dirt and supernatant. 2.3. Fungi DNA Amplification Process
The DNA amplification process was started by making PCR mix consisting of hot start PCR mix (QIAGEN), it’s 1 and ITS 4 primers, DNA working fungi, and DDh2O. PCR mix solution was included in PCR machines for DNA amplification in vitro. The amplification process consists of initialization/initial denaturation at 95°C for 5 minutes, denaturation at 94 °C for 1 minute, attachment of primer to template DNA (annealing) at 55.4 °C for 1 minute, elongation of 72 °C for 1 minute, and extension of the final 72 °C for 10 minutes. The denaturation stage until the extension then repeated 35 times. DNA amplification process took place for ± 2 hours 16 minutes. 2.4. Separation of DNA Amplification Process
DNA amplification results were aligned to find out whether the amplification process was successful or not and to determine the size of the amplification product. Separation

Cendawan Endofit
123
of DNA amplification results was carried out using horizontal electrophoresis method. This method used 2% agarose and TAE buffer (Tris Acetate EDTA). The separation results were then placed in gel doc to see the results of separation using UV transilluminator and documented.
3. Results and Discussion 3.1. Isolated endophytic fungi
Endophytic fungi isolates which were successfully isolated from the local area of Enrekang rice plants as much as 8 isolates. Based on color and morphology observation items of the colonies, 2 isolates were selected for further identification. Selected isolates of endophytic fungi were re-grown on new PDA media, and DNA isolation was carried out at the 7th days. 3.2. Isolated of Fungi Endophytic DNA
The process of DNA isolation of endophytic fungi was successfully carried out through agarose gel electrophoresis. The formation of DNA bands on agarose gel observed under UV light showed that the DNA of both Enrekang local rice endophytic fungi had been isolated from mycelia was 7 days old. 3.3. PCR amplification
The use of a suitable primary pair and the right annealing temperature for each fungus is a determining factor in the success of PCR amplification. The primary pair used in this study was ITS1 as a specific fungus primer and ITS4 as a universal primer using an annealing temperature of 55.4°C. The results of the study [7] showed that the PCR process at 58°C for 1 minute gave optimum results. According to [8] the optimum annealing temperature greatly determines the success of the PCR. The high annealing temperature can inhibit the hybridization of the template so that

124
Cendawan Endofit
the resulting PCR product is less.
Figure 1. Results of DNA PCR amplification of two local Enrekang rice endophytic fungi isolates using ITS1 / ITS partner 4. 1 kb band path DND Ladder, Line 5 and 6 entophytic fungi DNA bands
The results of DNA amplification of endophytic fungi obtained were 600 bp and 700 bp. In Figure 1, it can be seen that the DNA band obtained is in the form of a single band and indicates that the DNA resulting from the transformation is pure enough.
DNA amplification of fungi using PCR technique often uses the primary pair of ITS1 /ITS4 which will amplify the area of ITS ribosomal DNA (rDNA). Ribosomal DNA (rDNA) is a genome coding region for ribosomal RNA components (RNA). This gene is widely used in phylogenetics, classification, and identification for fungi because of its universal nature, its conservative sequence structure and its presence in large numbers [8].
4. Conclusion DNA isolation was successfully performed using 0.8% agarose gel electrophoresis characterized by the formation of DNA bands. PCR amplification in the area of ITS-1 and ITS-2 rDNA can be carried out using the primary pairs of ITS1 and ITS4 with an annealing temperature of 55.4 0C and producing DNA fragments of 600 bp and 700 bp
Acknowledgement The author would like to thank Ristekdikti who has funded this

Cendawan Endofit
125
research through Higher Education Basic Research Scheme based on the Decree of the Director General of Strengthening Research and Development of the Ministry of Research, Technology and Higher Education of the Republic Indonesia Number 3 / E / KPT / 2018 on Acceptance of Research Funding in Higher Education in 2018
References [1] Irmawan D 2007 Kelimpahan dan kergaman cendawan
endofit pada beberapa varietas padi di Kuningan (Jawa Barat: Tasikmalaya dan Subang) p 7
[2] Syamsia 2016 Isolasi dan Identifikasi Cendawan Endofit Padi Aromatik Lokal Enrekang. J. Agrotan. 2 59-65
[3] Sucipto I 2016 Eksplorasi Bakteri dan Cendawan Endofit sebagai Agens Pengendali Penyakit Blas (Pyricularia oryzae) pada Padi Sawah (Bogor: Institut Pertanian Bogor) p 56
[4] Rahayu F and Nugroho T T 2015 Isolasi DNA dan Amplifikasi PCR Daerah ITS rDNA Fungi Endofit Umbi Tanaman Dahlia (Dahlia variabilis) LBKURCC69 Jom. Fmipa. 2 4-10
[5] Nuryadi W, Rakhmawati A and Prihatini I 2016 Isolasi dan Identifikasi Kapang Endofit dari Pohon Sengon Provenan Kepulauan Solomon Berdasarkan Morfologi dan Molekuler Analisis r DNA ITS Internal Transcribed Spacer J. Biologi 5 15-27
[6] Wilia W, Hayati I and Ristiyadi D 2012. Eksplorasi cendawan endofit dari tanaman padi sebagai aggens pemacu pertumbuhan tanaman Program Studi Agroekoteknologi, Fakultas Pertanian Universitas Jambi 1 73-79

126
Cendawan Endofit
[7] Ludyasari A 2005 Pengaruh Suhu Annealing pada Program PCR terhadap Keberhasilan Amplifikasi DNA Udang Jari (Metapenaeus elgans De Man, 1907) Laguna Segara Anakan, (Cilacap: Jawa Tengah) p 89
[8] Larekeng SH, Restu M, Arif A, Cahyaningsih Y F and Mukti J 2019. A Genetic Approach to Study Mating System on Jabon Merah (Anthocephalus macrophyllus Roxb.) from Three Different Provenances in South Sulawesi IOP Conf.Ser.: Earth Environ.Sci. 235 1-9
[9] Legiastuti T S and Aminingsih T 2012 Identifikasi Cendawan Endofit Menggunakan Teknik Polymerase Chain Reaction Fitopatologi Indonesia 8 31-36

Cendawan Endofit
127
Lamprian 3. Hasil riset, publikasi pada International Journal of Agriculture System Vol. 7 Issue 2, December 2019. Judul: Molecular Identification of Endophytic Fungi from Local Rice and Growth Test on Several Types of Culture Media. http://pasca.unhas.ac.id/ojs/index.php/ijas/article/view/2031/570
Molecular Identification of Endophytic Fungi from Local Rice and Growth Test on Several Types of Culture Media
Syamsia1, Abubakar Idhan1, Amanda Patapparai1, Noerfitryani Noerfitryani2, Rahmi Rahmi3, Iradhatullah Rahim4*
1 Department of Agrotechnology, Universitas Muhammadiyah Makassar, Indonesia
2 Department of Agribusiness, Universitas Muhammadiyah Makassar, Indonesia
3 Department of Aquaculture, Universitas Muhammadiyah Makassar, Indonesia
4 Department of Agrotechnology, Universitas Muhammadiyah Pare-Pare,
Indonesia * Corresponding author’s e-mail: [email protected]
ABSTRACTLocal rice is rice that has been cultivated for generations by the community and commonly cultivated without using chemical inputs. Endophytic fungi are fungi that live in the plant tissue and does not cause disease symptoms in the host plants. This study

128
Cendawan Endofit
aimed to molecular identifying isolates of MDTA and MDTB endophytic fungi which have been isolated from the local Pulu Mandoti rice plant tissue and growth test on the four types of culture media those were synthetic PDA, natural PDA, MPA, and MEA. The fungi DNA isolation using DNesay Kit. DNA sequencing analysis using the mega BLAST program showed that the MDTB fungus has similarities to Podoscypha bolleana strain 32034 no accession JQ675334 and Podoscypha bolleana strain 32032 no accession JQ675332, whereas the MDTA fungus has similarities to Coprinopsis cinerea A2S3-5 isolate and Coprinopsis cinerea strain CNRMA / F 07-32. The best culture media and sporulation of endophytic fungi is MPA media. This research is the first study to molecular identifying with endophytic fungi from local rice and viability test on the four types of culture media. The results of this study contribute to the diversity of local rice endophytic fungi in SulawesiKeywords: Coprinopsis cinere; culture media; Podoscypha bolleana; Sporulation
1. IntroductionEndophytic fungi are microorganisms that live in plant tissues without causing symptoms of damage to plants (Hilarino et al., 2011; Afandhi et al., 2018). Endophytic fungi have been isolated from several types of plants such as: 1) Apple (Malus domestica) (Afandhi et al., 2018); 2) Orchid (Cymbidium aloifolim L) (Shubha & Srinivas, 2017); 3) medicinal plant (Hedychium flavescens and Hedychium coronarium) (Uzma, Konappa, & Chowdappa, 2016); 4) medicinal plant (Asclepias sinaica) (Fouda, Hassan, Eid, & Ewais, 2015); 5) Medicinal Plant (Adhatoda vasica Nees, Coleus aromaticus Benth, Costus igneus N.E.Br and Lawsonia

Cendawan Endofit
129
inermis Linn) (Amirita et al., 2012); 6) Wild rubber trees (Hevea brasiliensis) (Gazis & Chaverri, 2010).
Endophytic fungi had an important role in agriculture such as stimulating or increasing plant growth, increasing plant resistance to pests, diseases, and nematodes, increasing plant resistance to drought stress. Some research results on the important role of endophytic fungi in agriculture are: 1) biocontrol agents for blast disease (Sucipto, Munif, Suryadi, & Tondok, 2015); 2) spur plant growth (Saylendra & Firnia, 2013); 3) biocontrol agent (Suciatmih, Yuliar, & Supriyati, 2011); 4) increase resistance to pathogenic pests (G, A, & Kannan, 2015); 5) increasing plant resistance to drought stress (Shukla, Awasthi, Rawat, & Kumar, 2012); 6) protect host plants from pest attacks (Faeth, 2002).
Exploration of endophytic fungi has been carried out on several types of plants to obtain a collection of endophytic fungi that can be utilized in agricultural fields. Local rice plants are one of the plants that had the potential of endophytic fungi that need to be explored in depth because they have some potential. According to Sitaresmi, Wening, Rakhmi, Yunani, & Susanto (2013), local rice naturally has resistance to pests and diseases, abiotic stress tolerant and has good rice quality and flavor favored by consumers in every location where the rice plant was cultivated.
Regional development centers of local rice in South Sulawesi are generally located in the mountainous area with an altitude 1000 m above sea level such us Tanah Toraja, Enrekang, and Luwu. Local rice varieties developed in Toraja are Pare Lalodo; Rogon; Pare Lea; Pare Kobo; Pare Ra’rari, Pare Ambo, Pare Tallang, Pare

130
Cendawan Endofit
Bau; Pare Birrang and Pare Bumbungan (Juhriah, A. Masniawati, Tabaru, & Astuti, 2013). While in Enrekang local rice was developed; Pare Salle; pare Pulu Lotong, pare Pinjan, pare Pulu Mandoti, pare Lambau, pare Pallan, Pare Solo, pare Mansur and pare Kamida (Maulana, Kuswinanti, Sennang, & Syaiful, 2014). Luwu has 5 local rice varieties, namely: Tarone, Dambo, Kamba, Mandi, and Remaja.
Exploration and identification of endophytic fungi are very important because endophytic fungi have an important role in the ecosystem. Besides that, it can be used as information and as a basis for developing the potential of endophytic fungi. Some endophytic fungi have been isolated from rice plants and morphologically identified, namely: 1) Fusarium sp, Cladosporium sp, Phoma sp, Penicillium sp (Sucipto, Munif, & Tondok, 2016); 2) Aspergillus sp1, Aspergillus sp2, Aspergillus candidus and Acremonium (Syamsia, 2016); 3) Nigrospora sp. 3, Penicillium, Trichoderma sp. 2, Nigrospora sp. 4, Verticillium, and brown sterile hyphae 2 (Asiah, Wiyono, & Triwidodo, 2011). Identification of fungi morphologically based on colonies and spores often less certainty of the identity of isolates. This is because the morphology endophytic fungi can be changed, other than that some endophytic fungus has very slow growth and frequent sporulation (Hyde dan Soytong 2008); (Legiastuti & Aminingsih, 2012)). According to (Diaz, Hennell, & Sucher, 2013), one solution to anticipate identification deficiencies morphologically is through molecular identification.
Molecular identification of endophytic fungi using comparative analysis of ribosome DNA sequences, especially the Internal

Cendawan Endofit
131
transcribed spacer (ITS) region using primary ITS2, ITS4 and ITS5 (Y.W. et al., 2009); primary ITS1 and ITS4 (Fernandes, Pereira, Silva, Bento, & Queiroz, 2015) and primary ITS5 and ITS4 (Rakhmana, Rahayu, Ardhi, & Wahyu, 2017); primary ITS1 and ITS2 (Alwakeel, 2013).
The types of culture media that can be used for fungi were Potato Dextrose Agar (PDA), Carrot Agar Potato (PCA), Sabouraud Dextrose Agar (SDA), Czapex Dox Agar (CDA), Corn Meal Agar (CMA), Nutrient Agar (NA), Malt Extract Agar (MEA) (Taurisia, Provorini, & Nurantoro, 2000).
This study aimed to identify endophytic fungi isolates from Enrekang local rice plants and test the ability of fungi to grow on several types of media. The molecular identification of endophytic fungi from Enrekang local rice has never been done before, so this is the first study. In addition, growth test of local rice in several types of media. The results of this study will provide the benefits of the diversity of endophytic fungi from local rice and further research for the development of potential endophytic fungi as PGPF in increasing agricultural production.
2. Materials and Method 2.1 Rejuvenation of Isolates and Endophytic Fungi DNA Extraction Two endophytic fungi isolate that were isolated from the roots and stems of Pulu Mandoti rice namely MDTB and MDTA were re-grown on PDA media. Mycelia of fungi isolates was harvested and extracted using DNesay DNA DNA kits from Qiagen. Miselia of fungi was taken and placed in mortal then crushed until smooth.

132
Cendawan Endofit
Mycelia was inserted into the microtube that has been filled with 400 µl buffer AP1, 40 µl PVP 26% and 4 µl RNase A stock (100 mg/ml) in the vortex then incubated in a water bath at 65 ° C for 30 minutes. The solution was then centrifuged.
2.2 Fungi DNA amplification Endophytic fungi DNA amplification using primers ITS1 and ITS4. The amplification started with the manufacture of PCR mix consisting of hotstar mix PCR (Qiagen), Primer ITS 1 and ITS 4, DNA working and DDh2O. The PCR mix solution was then inserted into the PCR machine for DNA amplification in vitro. The PCR solution was inserted into the PCR machine and the amplification process started with an initial denaturation at 95°C for 5 minutes, denaturation at 94°C for 1 minute, annealing at 55.4°C for 1 minute and the final extension of 72°C for 10 minutes. The denaturation stage until the extension was repeated 35 times. The DNA amplification process lasts for ± 2 hours 16 minutes.
2.3 Separation Process of DNA Amplification Results DNA amplification product separation was carried out using a horizontal electrophoresis method. This method was used 2% agarose and Tris Acetate EDTA (TAE) buffer 1 x. The results of separation then inserted in geldoc to see the results of separation used UV transuminator and documented. Separation DNA amplification product was repaired to determine whether the amplification process was successful or not and determine the size of the amplification product. 2.4 DNA Sequencing Analysis PCR products that were successfully amplified from MDTB and MDTA endophytic fungi isolates were sent to Genetic science, Jakarta for

Cendawan Endofit
133
sequencing. The sequencing results were then analyzed using the Basic Local Alignment Search Tool (BLAST) 2.5 Growth Test on Four Types of Culture Media Growth test of both endophytic fungi isolates on four types of culture media was carried out by growing each isolate in synthetic Potato Dextrose Agar (PDA) media, natural PDA, Malt Extract Agar (MEA) and Malt Peptone Agar (MPA). Endophytic fungus mycelium of two isolates were grown on media, Synthetic PDA (synthetic PDA), natural PDA (made from potato extract), MEA (15 g malt extract, 16 g agar / L sterile water), MPA (15 g malt extract, 20 g glucose, 5 g peptone and 16 agar / L sterile water). The isolates were incubated at room temperature. The growth of two isolates was measured based on the colony diameter until the 7th day (Rahim et al. 2015).
3. Results and Discussion 3.1 Molecular Identification The two of endophytic fungi isolates had different morphological characters, MDTB isolates were gray, the upper surface of the colony was compact and thick, the colony grows was very slow, small diameter (± 1.25 cm), the colony reverse was rather creamy. MDTB isolate was blackish white, smooth upper surface, the colony growth was very fast, large diameter (± 8.89 cm), colony reverse was white, the two isolates have concentric zones types.
Isolates DNA endophytic fungi were successfully amplified using primers pairs of ITS1 and ITS4 at an annealing temperature of 55.4 ° C. The formation of DNA bands on an agarose gel after electrophoresis showed the PCR amplification process was successful. The success of PCR amplification was determined by the primer pairs and the corresponding annealing temperature in

134
Cendawan Endofit
each fungus. This result was similar to the (Sibero et al., 2018), who succeeded in clarifying endophytic fungi R3 isolates isolated from coastal plants Hydnophytum formicarum from Sorong using primers pairs of ITS1 and ITS4. Likewise, the research of (Rahayu, Saryono, & Nugroho, 2015), succeeded in amplifying the DNA PCR of endophytic fungi from LBKURCC69 isolates that had been isolated from dahlia bulbs in the area ITS-1 and ITS-2 rDNA and primer pairs of ITS4 and ITS5. Research (Alwakeel 2013) using primer pairs of ITS1 and ITS 2. The results of electrophoresis analysis on the results of PCR produce a single band for each DNA amplification (Fig. 1). The size of the molecular weight of the two isolates was 700 bp. This result was different from the DNA band size of two endophytic fungi isolates isolated from dahlia plants, which were only 583 and 537 bp in size (Rakhmana et al., 2017), as well as the DNA size of endophytic fungi isolates from srikaya plants was 600 bp (Yunianto et al. 2012).

Cendawan Endofit
135
Fig. 1 The results of DNA amplification MDTB endophytic fungi isolates (1) amd MDTA (7) using primers pairs of ITS1 and ITS4, DNA ladder (1kb)
The results of identification using DNA barcoding proved that endophytic fungi from local Sulawesi Selatan rice with sample code 1 were homologs of Podoscypha bolleana strain 32034 no accession JQ675334 and Podoscypha bolleana strain 32032 no accession JQ675332 with 100% similarity level and sample code 8 was a homolog of Coprinopsis cinerea isolate A2S3-5 with 99% similarity level and Coprinopsis cinerea CNRMA / F 07-32 strain with 100% similarity level. Research (Sibero et al., 2018) proved that RS3 fungus was a homolog of Annulohypoxylon stygium DR47 strain with 99% similarity level, with accession number MG605083.1. Research by (Nuryadi et al., 2016), succeeded in identifying molecular fungi from dahlia tubers using ITS molecular markers, there were 4 isolates as Lasiodiplodia genus, 4 isolates as Didymellaceae family, 11 isolates were identified as Phomopsis genus, 5 isolates as Colletotrichum genus, 1 isolates as Nemania genus, and 1 isolate as Xylaria genus. Coprinopsis cinerea is a basidiomycete fungus that is used for many basic

136
Cendawan Endofit
studies, including research into the fungus development stage. C. cinerea fungus was easy to maintain, has a short life cycle and can be induced to develop fruit bodies in the laboratory and only takes 2 weeks to produce a ripe fruit body. The sequence results created in the FASTA format for phylogenetic tree construction using MEGA7. The tree was constructed using UPGMA models and Kimura-2- Parameter genetic distances. In the process of making the tree, Podoscypha bolleana Strain 32032, P. bolleana Strain 32034 having a close kinship to MDTA (Table 2.) and Coprinopsis cinereaStrain HN08 and C. cinerea A2S3-5 having a close kinship to MDTB (Table 3.). Dendogram based on the similarity value of the strain obtained from Gen-Bank. Each morphotype was grouped together and separated from each other. The phylogenetic tree produced (Fig 2). The tree showed that the position of the sample with its relative species. The position of MDTA with MDTB in the phylogenetic tree lies in one cluster so it showed that they were closely related. The genetic distances between MDTA and MDTB samples were calculated using the Kimura-2-Parameter method in MEGA7 according to (Kumar, Stecher, & Tamura, 2016). The results showed that the genetic distances between MDTA and MDTB with the highest boostrap value at 93%. These results indicate them likely that MDTA and MDTB endophytic fungi are closely related and even tend to be subspecies
Fig 2. Phylogenetic trees of MDTB and MDTA isolates
3.2 The Growth of endophytic fungi in four type of media The growth of the two endophytic fungi isolates in four types of media showed that the diameter of the colonies, the color of the

Cendawan Endofit
137
upper surface, the upper surface and zoning were basically the same for all types of media, except for texture and sporulation there were differences in MEA media, the texture of the two isolates very thin in the MEA medium and sporulate sporulation was very poor (Table 5 and Table 6).
The growth of MDTB endophytic fungi isolates in four types of media namely synthetic PDA, natural PDA, MPa and MEA (Fig. 3). Both endophytic fungi isolates have the ability to grow well in

138
Cendawan Endofit
all three types of media, namely PDA Synthetic, natural PDA and MPA. The media suitable for sporulation was MPA media. The result of this study was similar to research (Rahim, Kuswinanti, Asrul, & Rasyid, 2015) who was found MPA media as the best medium for foliage fungus growth. However, different research result (Devi, Misra, Saha, Devi, & Sinha, 2018) was getting the best media for fungi sporulation were MEA and OMA.
Fig. 3 The growth of MDTB endophytic fungi isolates in four types of media: Synthetic PDA (A), Natural PDA (B), MPA (C) and MEA (D)
In general, natural PDA media and synthetic PDA were still suitable for growth and sporulation of endophytic fungi. The result of the study (Noerfitryani & Hamzah, 2017), who use PDA media for the growth of Fusarium, Aspergillus and Trichoderma fungi. According to (Devi et al., 2018) PDA media was a medium commonly used for fungus growth because the formulation was simple and has the ability to support the growth of mycelia in almost all types of fungi. Likewise, research (Aini & Rahayu, 2015) showed that PDA media provide the best growth of fungi Candida albicans and Aspergillus niger than alternative media.
4. Conclusion The MDTB fungus has similarities to Podoscypha bolleana strain

Cendawan Endofit
139
32034 no accession JQ675334 and Podoscypha bolleana strain 32032 no accession JQ675332, whereas the MDTA fungus has similarities to Coprinopsis cinerea A2S3-5 isolate and Coprinopsis cinerea strain CNRMA/F 07-32. The best culture media and sporulation of endophytic fungi is MPA media.
Acknowledgements The author would like to thank Ristekdikti who has funded this research through Higher Education Basic Research Scheme based on the Decree of the Director General of Strengthening Research and Development of the Ministry of Research, Technology and Higher Education of the Republic Indonesia Number 3/E/KPT/2018 on Acceptance of Research Funding in Higher Education in 2018
References Afandhi, A., Choliq, F. A., Havinda Anggrilika, W. S., & Tarno,
H. (2018). Distribution of the endophytic fungi in apple leaves. Agrivita 40: 91–100
Aini, N., & Rahayu, T. (2015). Alternatif Media for Fungal Growth Using a Different Source of Carbohidrats Nurul. In Seminar Nasional XII Pendidikan Biologi FKIO: 861–866 [Indonesian]
Alwakeel, S. S. (2013). Molecular identification of isolated fungi from stored apples in Riyadh, Saudi Arabia. Saudi Journal of Biological Sciences 20: 311–317.
Amirita, A., Sindhu, P., Swetha, J., Vasanthi, N. S., & Kannan, K. P. (2012). Enumeration of endophytic fungi from medicinal plants and screening of extracellular enzymes. Word Journal of Science and Technolgy 2: 13–1.

140
Cendawan Endofit
Asiah, N., Wiyono, S., & Triwidodo, H. (2011). Keanekaragaman dan Kelimpahan Cendawan Endofit pada Batang Padi. [Skripsi]. Institut Pertanian Bogor [Indonesian]
Devi, K. S., Misra, D. K., Saha, J., Devi, P. S., & Sinha, B. (2018). Screening of Suitable Culture Media for Growth, Cultural and Morphological Characters of Pycnidia Forming Fungi. International Journal of Current Microbiology and Applied Sciences 7: 4207–4214.
Diaz, P. L., Hennell, J. R., & Sucher, N. J. (2013). Chapter 14 Genomic DNA Extraction and Barcoding of Endophytic Fungi. In Methods in Molecular Biology :171–179. Faeth, S. H. (2002). Are endophytic fungi defensive plant mutualists? Oikos 9: 25–36.
Fernandes, E. G., Pereira, O. L., Silva, C. C. da, Bento, C. B. P., & Queiroz, M. V. de. (2015). Diversity of endophytic fungi in Glycine max. Microbiological Research 181: 84–92.
Fouda, A. H., Hassan, S. E. D., Eid, A. M., & Ewais, E. E. D. (2015). Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Annals of Agricultural Sciences, 60: 95–104.
G, A. K., A, R. A., & Kannan, V. R. (2015). Exploration of endophytic microorganisms from selected medicinal plants and their control potential to multi drug resistant pathogens. Journal of Medicinal Plant Studies 3: 49–57.
Gazis, R., & Chaverri, P. (2010). Diversity of fungal endophytes in leaves and stems of wild rubber trees (Hevea brasiliensis) in Peru. Fungal Ecology 3: 240–254.
Hilarino, M. P. A., Silveira, F. A. de O. e, Oki, Y., Rodrigues, L., Santos, J. C., Corrêa Junior, A., … Rosa, C. A. (2011). Distribution of the endophytic fungi community in leaves

Cendawan Endofit
141
of Bauhinia brevipes (Fabaceae). Acta Botanica Brasilica, 25(4), 815–821.
Hyde KD, Soytong K. 2008. The fungal endophyte dilemma. Fungal Divers. 33:163- 173.
Juhriah, A. Masniawati, Tabaru, E., & Astuti, S. (2013). Morphological Characterization of Panicle Landrice’s from North Tana Toraja South Sulawesi. Jurnal Sainsmat II: 22–23. [Indonesian]
Kumar, S., Stecher, G., & Tamura, K. (2016). MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Molecular Biology and Evolution 33 :1870–1874.
Langer, E, (2017). Molecular phylogeny of Podoscypha (Basidiomycota, Meruliaceae) with description of a new genus, Crinipileoscypha gen. nov., inferred from nuclear and mitochondrial rDNA sequences; Unpublished
Legiastuti, T. S., & Aminingsih, T. (2012). Detection of Endophytic Fungi Using Polymerase Chain Reaction Technique. Fitopatologi Indonesia 8:31–36. Maulana, Z., Kuswinanti, T., Sennang, N. R., & Syaiful, S. A. (2014). Genetic Diversity of Locally Rice Germplasm from Tana Toraja and Enrekang Based on RAPD (Random Amplified Polymorphism DNA) Markers. International Journal of Scientific & Technology Research 3: 198–202.
Noerfitryani, N., & Hamzah, H. (2017). The Existence of Entomopatogenic Fungi on Rice Plants Rhizosphere. Internasional Journal of Bioscience and Biotechnology 5 :12–24.
Nuryadi, W., Rakhmawati, A., & Prihatini, I. (2016). Isolation and Identification of Endophytic Fungi of Sengon Trees Provenance Solomon Islands Based on Morphological and

142
Cendawan Endofit
Molecular (Analysis of rDNA ITS (Internal Transcribed Spacer). Jurnal Biologi 5 :15–27.
Rahim, I., Kuswinanti, T., Asrul, L., & Rasyid, B. (2015). Screening of Fungal Rot Isolates from Cocoa as Phosphate-Dissolving and their Growth Ability on Three Types of Media. Procedia Food Science 3: 104–111.
Rakhmana, S., Rahayu, F., Ardhi, A., & Wahyu, N. (2017). Molecular identification of endophytic fungi isolated from the tuber of Dahlia variabilis and exploration of their ability in producing ß- galactosidase. Biodiversitas 18 :145–152.
Saylendra, A., & Firnia, D. (2013). Potensi Cendawan Endofit Perakaran Jagung Sebagai Pemacu Pertumbuhan Tanaman. Jurnal Ilmu Pertanian dan Perikanan, 2: 135–140.
Shubha, J., & Srinivas, C. (2017). Diversity and extracellular enzymes of endophytic fungi associated with Cymbidium aloifolium L. African Journal of Biotechnology: 16: 2248–2258.
Shukla, N., Awasthi, R. P., Rawat, L., & Kumar, J. (2012). Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiology and Biochemistry 54 :78–88.
Sibero, M. T., Tarman, K., Radjasa, O. K., Sabdono, A., Trianto, A., & Bactiarini, T. U. (2018). Production of fungal pigment and identification of its producer through DNA barcoding approach. JPHPI 21: 99–108.
Sitaresmi, T., Wening, R. H., Rakhmi, A. T., Yunani, N., & Susanto, U. (2013). The Use of Local Variety Rice Germplasm in the Development of Improved Varieties. Iptek Tanaman Pangan 8: 22–30.

Cendawan Endofit
143
Suciatmih, Yuliar, & Supriyati, D. (2011). Exploration of Entophytic Fungi from Lowland Rice as a Biocontrol Agen of Blast Disease in Lowland Rice. Jurnal Teknologi Lingkungan 12: 171–186.
Sucipto, I., Munif, A., Suryadi, Y., & Tondok, E. T. (2015). Eksploration of Entophytic Fungi from Lowland Rice. Jurnal Fitopatologi Indonesia 11: 211–218.
Sucipto, I., Munif, A., & Tondok, E. T. (2016). Exploration of Endophytic Bacteria and Fungi as Biocontrol Agent of Blast Disease (Pyricularia oryzae) on Rice. Institut Pertanian Bogor. Syamsia. (2016). Isolation and Identification of Endophytic Fungus on Local Aromatic Rice Plants of Enrekang. Agrotan 2: 61–67.
Taurisia, P. P., Provorini, M. W., & Nurantoro, I. (2000). The Effect of Media on The Growth and Biomass of Fungi Alternaria alternata (Fries) Keissler. Jurnal Biologi 19: 30 – 33. Teh Li Yee., & Latiffah Zakariah. (2014). The first report of Penicillium georgiense in Malaysia. Mycobiology 42: 274–278.
Uzma, F., Konappa, N. M., & Chowdappa, S. (2016). Diversity and extracellular enzyme activities of fungal endophytes isolated from medicinal plants of Western Ghats, Karnataka. Egyptian Journal of Basic and Applied Sciences 3: 335–342.
Y.W., H., Z.Y., C., S, S., K.D, H., H, C., & M, S. (2009). Molecular phylogenetic identification of endophytic fungi isolated from three Artemisia species. Fugal Diversity: 69 – 88. Retrieved from www.Plants Journal.com
Yunianto, P., Rosmalawati, S., Rachmawati, I., Priyono Suwarsono, W., & Sumaryono, W. (2012). Isolation and Identification of

144
Cendawan Endofit
Endophytic Fungi from Srikaya Plants (Annona squamosa) Having Potential Secondary Metabolites as Anti-Breast Cancer Activity. Microbiology Indonesia 6: 23–29.
Zhang, Y. and Li, D.C. 2012. Taxonomic studies of thermotolerant fungi. Department of Plant Pathology, Shandong Agricultural University

Cendawan Endofit
145
Lampiran 4. Hasil riset, publikasi pada jurnal Jurnal Galung Tropika, 9 (2) Agustus 2020. Judul: Potensi Cendawan Endofit Padi Lokal Sulawesi Selatan Sebagai Penghasil Siderefor https://jurnalpertanianumpar.com/index.php/jgt/article/view/659/pdf_93
Potensi Cendawan Endofit Padi Lokal Sulawesi Selatan Sebagai Penghasil Siderefor
Syamsia*Email: [email protected] Fakultas Pertanian,
Universitas Muhammadiyah Makassar Jl. Sultan Alauddin No. 259 Makassar, Sulawesi Selatan, 90221
Abubakar Idhan Email: [email protected] Fakultas Pertanian, Universitas
Muhammadiyah Makassar Jl. Sultan Alauddin No. 259 Makassar, Sulawesi Selatan, 90221
Amanda Patappari Email: [email protected] Fakultas Pertanian, Universitas Muhammadiyah Makassar Jl. Sultan Alauddin No.
259 Makassar, Sulawesi Selatan, 90221 Noerfitryani Email: [email protected] Fakultas
Pertanian, Universitas Muhammadiyah Makassar JL. Sultan Alauddin No. 259 Makassar, Sulawesi Selatan, 90221
ABSTRAKSiderefor adalah agen pengkelat (Chelating agent) besi yang dihasilkan oleh tanaman dan mikoorganisme pada kondisi kekurangan besi. Salah satu mikroorganisme yang memiliki

146
Cendawan Endofit
kemampuan memproduksi siderefor adalah cendawan. Penelitian ini bertujuan untuk mengetahui potensi cendawan endofit dalam memproduksi siderefor. Kemampuan cendawan dalam memproduksi siderefor terutama tipe Salisilat dan Katekol menggunakan medium Potato Dextrose Broth (PDB) dan Reagen Hathway. Sebanyak delapan isolat cendawan endofit asal padi lokal Sulawesi Selatan diuji kemampuan memproduksi siderefor. Delapan isolat cendawan endofit sebelum diuji diremajakan pada medium Potato Dextrose Agar (PDA) selama tujuh hari. Setiap isolat ditumbuhkan pada medium PDB sebelum diuji. Pengukuran absorbansi menggunakan spektrofotometer pada panjang gelombang 560 nm dengan Na. Salisilat 700 nm dengan 2.3 DHBA sebagai standar. Hasil penelitian menunjukkan isolat E4 dan E8 yang memiliki kemampuan memproduksi siderefor terbaik. Hasil penelitian memberikan informasi terbaru terkait potensi cendawan endofit asal padi lokal Sulawesi Selatan sebagai penghasil siderefor.
Kata kunci: siderefor; agen pengkelat; katekol; natrium salisilat.
ABSTRACTSiderophore is an iron-chelating agent produced by plants and microorganisms under iron deficiency conditions. One of the microorganisms that ability produce siderophore is a fungus. This study aimed to determine the potential of endophytic fungi in creating siderophore. The ability of fungi to produce siderophore, especially Salicylate and Catechol using Potato Dextrose Broth (PDB) liquid media and Hathway Reagent. Eight isolates of endophytic fungi from local rice South Sulawesi

Cendawan Endofit
147
were tested their ability to produce siderophore. Eight isolates of endophytic fungi before being tested were rejuvenated by growing on Potato Dextrose Agar (PDA) medium for seven days. Each isolate was grown on a PDB medium before being tested. The absorbance is measured using a spectrophotometer at a wavelength of 560 nm with Na. Salicylate 700 nm with 2.3 DHBA as standard. The results showed that the isolates of E4 and E8 had the capability to produce siderophores. The results provide the latest information regarding endophytic fungi’ potential from local rice in South Sulawesi as a siderophore producer.
Keywords: siderophore; chelating agent; catechol; sodium salicylate.
PENDAHULUANSiderefor adalah agen pengkelat besi dengan ukuran molekul
kecil yang dihasilkan oleh mikroorganisme dan tanaman dalam kondisi kekurangan besi (Schwyn & Neilands, 1987; Ahmed & Holmström, 2014). Siderefor merupakan senyawa dengan bobot molekul redah yang mampu mengkelat besi (Sharma & Johri, 2003; Prihatiningsih dkk., 2017). Dengan demikian keberadaan siderefor sangat penting bagi pertumbuhan tanaman untuk mengatasi kekurangan/kelebihan unsur hara besi bagi tanaman. Siderefor dibedakan dalam tiga kelompok, yaitu hidroksamat, katekolat dan karboksilat (Ahmed & Holmström, 2014).
Beberapa tanaman dilaporkan dapat menghasilkan siderefor, seperti barley dan gandum yang dapat mempe- roleh Fe dari sumber yang tidak larut (Kraemer, 2004; Ahmed & Holmström, 2014), dengan cara mengeluarkan pengkelat Fe (III) yang disebut

148
Cendawan Endofit
fitosiderofor yang membentuk kompleks kuat spesifik dengan Fe (III) (Ma, 2005; Ahmed & Holmström, 2014). Mikroor- ganisme juga mampu menghasilkan siderefor, seperti bakteri dan cendawan. Bakteri berhasil diisolasi dan memiliki kemampuan menghasilkan siderefor seperti Bacillus subtilis yang diisiolasi dari rizosfer kentang (Prihatiningsih dkk., 2017), Pseudomonad fluoresen (Pratama et al., 2018), dan isolat bakteri yang diisolasi dari rizosfer kentang varietas Hartapel (Kesaulya et al., 2015). Cenda- wan yang telah diisolasi dan mampu memproduksi siderefor, seperti Trichoderma (Anke et al., 1991), Cenda- wan pelapuk coklat Gloephyllum trabeum (Ahmed & Holmström, 2014), Aspergillus niger dan Penicillim oxalicum (Aziz et al., 2016).
Bakteri sebagian besar menghasil- kan siderefor tipe katekolat (enterobaktin) dan sebagian menghasilkan karboksilat (rhizoabaktin) dan hidroksamat (ferrioksamin) (Matzanke, 1991; Ahmed & Holmström, 2014). Sedangkan cenda- wan menghasilkan siderefor tipe katekolat dan hidroksamat (Fekete et al., 1989; Fekete; 1993; Ahmed & Holmström, 2014).
Kemampuan tanaman dan mikro-organisme dalam memproduksi siderefor menjadi sangat penting karena hal ini terkait dengan permasalahan yang sering dihadapi dalam pertumbuhan dan produksi tanaman terutama terkait kelebihan dan kekurangan unsur hara besi. Fungsi utama siderefor adalah untuk mengkelat besi, membentuk senyawa kompleks dengan logam berat seperti Cd, Cu dan Zn (Johnstone & Nolan, 2015; Hussein & Joo, 2019). Menurut Kobayashi dan Nsihizawa (2012), dan Ahmed dan Holmström (2014), besi merupakan unsur esensial untuk pertum- buhan tanaman. Sharma dan Johri (2003) yang didukung dengan Prihatiningsih dkk. (2017), bahwa

Cendawan Endofit
149
keberadaan siderefor dapat menguntungkan tanaman karena dapat menghambat pertumbuhan patogen. Patogen mengalami kekurangan Fe3+ akibat terikatnya Fe3+ oleh siderefor. Hal ini menjadi peluang pemanfaatan side- refor sebagai pemacu pertumbuhan dan pengendali patogen.
Beberapa cendawan asal rhizosfer tanaman telah diteliti menunjukkan kemampuan menghasilkan siderefor. Na- mun penelitian terkait kemampuan isolat cendawan endofit asal padi lokal Sulawesi Selatan dalam memproduksi siderefor belum pernah dilakukan. Berdasarkan uraian tersebut, penelitian ini dilakukan dengan tujuan untuk mengetahui potensi cendawan endofit asal padi lokal dalam memproduksi siderefor.
METODE PENELITIANPeremajaan Isolat Cendawan Isolat cendawan endofit koleksi hasil penelitian tahun 2018 merupakan cendawan yang diisolasi dari bagian akar, batang, daun padi lokal dari Kabupaten Enrekang dan Toraja Sulawesi Selatan. Delapan isolat cendawan diremajakan pada medium Potato Dextrose Agar (PDA) sebelum dilakukan pengujian Siderefor. Setiap isolat diambil satu lempeng dan diletakkan di atas medium PDA dan diinkubasi selama 7 hari. Isolat cendawan endofit yang telah tumbuh dan murni selanjutnya diperbanyak pada media PDA untuk dilakukan pengujian.
Persiapan Kultur Isolat Cendawan Endofit Setiap isolat cendawan endofit yang telah tumbuh pada medium PDA ditumbuhkan kembali pada medium Potato Dextrose Broth (PDB). Sebanyak 3 lempeng isolat cendawan endofit dimasukkan dalam erlenmeyer yang berisi medium PDB cair

150
Cendawan Endofit
dan diinkubasi pada suhu 27°C. Kemudian dishaker dengan 150 rpm/menit selama 7 hari. Medium PDB yang telah ditumbuhi isolat cendawan endofit disaring menggunakan kertas saring dan disentrifius 10.000 g selama 20 menit. Ini digunakan sebagai larutan uji (supernata) pada pengujian siderefor.
Uji Produksi Siderefor Secara KualitatifUji kemampuan Siderefor tipe Salisilat dan Katekol 189solate cendawan endofit menggunakan metode yang dijelaskan oleh Sivasakthivelan dan Stella (2012), dan Kesaulya et al. (2015) yang dimodifikasi. Sebanyak 20 ml kultur supernata diambil dan pH diatur hingga 2,0 dengan menambahkan larutan HCl. Kedalam 20 ml supernata ditambahkan 20 ml etil asetat dan dilakukan ektraksi sebanyak dua kali. Selanjutnya diambil 5 ml supernata dan dimasukkan dalam tabung reaksi kemudian ditambahkan 5 ml reagen Hathway (1 ml 0,1 M besi klorida dan 1 ml 0,1 N HCL ditambahkan ke dalam 100 ml air suling dan ditambahkan 1 ml 0,1 M kalium ferricyanide), selanjutnya diamati perubahan warna pada larutan supernata dan dibandingkan dengan kontrol.
Uji Produksi Siderofer Secara Kuantitatif Pengukuran siderefor secara kuantitatif dilakukan dengan mengukur absorbansi supernata dari setiap isolat cendawan endofit menggunakan spektrofometer pada panjang gelombang 560 nm, menggunakan Natrium Salisilat sebagai standar. Standar dibuat dari pengenceran dengan konsentrasi Natrium Salisilat berkisar dari 0 hingga 2 mg l-1, dengan menggunakan persamaan regresi Y = 0.179x + 0.027. dimana R2 =0.902 untuk pengujian Na. Salisisat sedangkan untuk Katekol menggunakan absorbansi ditentukan pada 700 nm dengan 2.3 DHBA sebagai standar.

Cendawan Endofit
151
Konsentrasi dalam filtrat kultur ditentukan dan dinyatakan sebagai mg l-1 dengan persamaan regresi Y = 0.209x +0.038 dimana R 2
= 0.955 (Kesauliya, 2015).
HASIL DAN PEMBAHASAN Peremajaan Isolat Cendawan Endofit Peremajaan isolat cendawan endofit dilakukan sebelum pengujian siderefor, karena isolat yang telah lama disimpan dengan media dimana isolat cendawannya sudah kering. Peremajaan isolat cendawaan bertujuan untuk mendapatkan isolat yang masih baru dan segar (Gambar 1).
Kultur Isolat Cendawan Endofit Penyiapan kultur isolat cendawan endofit diawali dengan penyiapan media PDB (Gambar 2a). Isolat cendawan endofit yang telah diremajakan ditum- buhkan pada media PDB untuk mendapatkan kultur isolat cendwan endofit yang akan digunakan pada pengujian Siderefor (Gambar 2b).
Deteksi Kemampuan Produksi Siderefor Secara Kualitatif Delapan isolat cendawan endofit yang diuji kemampuan
memproduksi siderefor secara kualitatif menunjukkan adanya kemampuan memproduksi sidrefor yang bervariasi berdasarkan peru- bahan warna supernata dan dibandingkan dengan kontrol (kuning). Perubahan warna supernata setiap isolat bervariasi sesuai dengan kemampuan isolat dalam menghasilkan siderefor (Gambar 3).
Berdasarkan perubahan warna, isolat E1, E4, E5 dan E8 secara kualitatif memperlihatkan kemampuan mempro- duksi siderefor terbaik. Ini ditunjukkan dengan indikator warna yang

152
Cendawan Endofit
lebih hijau dibandingkan isolat lain. Hasil penelitian Milagres et al. (1999), terhadap 11 cendawan berhasil mendeteksi 8 cen- dawan memiliki kemampuan produksi siderefor secara kualitatif dengan menggunakan media padat. Menurut Aziz et al. (2016), Aspergillus niger dan Penicillim oxalicum merupakan dua cendawan terbaik dalam memproduksi siderefor dari 16 cendawan yang diuji.
Deteksi Kemampuan Siderefor Secara Kuantitatif Produksi
Uji kemampuan produksi siderefor secara kuantitatif terhadap delapan isolat cendawan endofit dengan melakukan pengukuran absorbansi supernata. Setiap isolat cendawan endofit diukur absor- bansinya menggunakan spektrofotometer pada panjang gelombang 560 nm dan 700 nm. Hasil pengukuran secara kuantitatif menunjukkan bahwa produksi siderefor isolat cendawan endofit dalam bervariasi yaitu 0.18 – 2.53 mg l-1. Isolat cendawan endofit E4 menunjukkan kemampuan produksi siderefor terbaik untuk tipe Salisilat, yaitu 1.34 mgl-1 dan Katekol sebesar 2.53 mgl-1 (Gambar 4). Secara umum siderefor mikroba diklasifikasikan dalam tiga kelompok utama yaitu katekol, hidroksamat, dan karboksilat (Winkelmann, 2002; Ghosh et al., 2017).

Cendawan Endofit
153
Terdapat 3 isolat cendawan endofit yang memiliki kemampuan memproduksi siderefor tipe Katekol yang tinggi yaitu isolat E1, E4 dan E8, sedangkan untuk tipe Salisilat hanya isolate E4. Isolat E4 memiliki kemampuan produksi siderefor tipe Katekol dan Salisilat yang lebih tinggi dibandingkan dengan isolat lain. Kemam- puan isolat E4 dalam memproduksi siderefor tipe Katekol lebih tinggi dibandingkan tipe Salisilat. Hal ini sesuai dengan apa yang dikemukakan oleh Fekete et al. (1989), dan Fekete (1993). Ahmed dan Holmström (2014), bahwa siderefor yang dihasilkan oleh cendawan umumnya katekolat dan hidroksamat. Hal serupa dikemukakan oleh Plattner dan Diekmann (1994), serta Hussein dan Joo (2019), berpendapat bahawa struktur siderefor cendawan sebagian besar adalah hidroksamat yang mirip dengan siderefor bakteri.
KESIMPULAN DAN SARAN
Kedelapan isolat cendawan endofit memiliki kemampuan produksi siderefor tipe Salisilat dan Katekol yang berbeda, yaitu 0,17 – 1,34 mgl-1 dan 0, 53- 2,53 mgl-1 mgl-1. Isolat cendawan endofit E4 dan E8 merupakan isolat yang memiliki kemampuan produksi siderefor tipe Katekol terbaik. Perlu dilakukan penelitian lanjutan untuk menguji kemampuan konsorsium isolate cendawan endofit dalam memproduksi siderefor.
UCAPAN TERIMA KASIH Ucapan terima kasih ditujukan kepada Kementrian Riset
dan Teknologi/ Badan Riset dan Inovasi Nasional Republik Indonesia yang telah mendanai kegiatan Penelitian ini melalui Skim Penelitian Dasar Unggulan Perguruan Tinggi (PDUPT) selama tiga tahun (2018- 2020).

154
Cendawan Endofit
DAFTAR PUSTAKAAhmed, E., & Holmström, S.J.M. 2014. Siderophores in
Environmental Research: Roles and Applications. Microbial Biotechnology, 7(3), 196–208. https://doi.org/10.1111/1751- 7915.12117.
Anke, Heidrun, Kinn, J., Bergquist, KE., & Sterner, O. (1991). Production of Siderophores by Strains of the Genus Trichoderma. Biology of Metals, 4(3), 176–80. https://doi.org/10.1007/BF01141 311.
Aziz, Abdel, O.A., Helal, G.A., Galal, Y.G.M., Kader, A., & Rofaida, S. (2016). Fungal Siderophores Production in Vitro as Affected by Some Abiotic Factors.International Journal of Current Microbiology and Applied Sciences, 5(6), 210–22. https://doi.org/10.20546/ijcmas. 2016.506.025.
Fekete, F.A. (1993) Assays for microbial siderophores. In Iron Chelation in Plants and Soil Microorganisms. in Barton, L.L., & Hemming, B.C. (eds). New York: Academic Press, pp. 399–417. F
ekete, F.A., Chandhoke, V., & Jellison, J. (1989) Iron- binding compounds produced by wooddecaying basidiomycetes. Appl Environ Microbiol, 55 (pg. 2720– 2722).
Ghosh, S. K., Banerjee, S., & Sengupta, C. (2017). Bioassay, characterization and estimation of siderophores from some important antagonistic Fungi. Journal of Biopesticides, 10(2), 105-112.
Hussein, K. A., & Joo, J. H. (2019). Zinc Ions Affect Siderophore Production by Fungi Isolated from the Panax ginseng Rhizosphere. Journal of microbiology and biotechnology, 29(1), 105-113.

Cendawan Endofit
155
Johnstone, TC., & Nolan, EM. (2015). Beyond iron: non-classical biological functions of bacterial siderophores. Dalton Trans, 14 (pg. 6320-6339).
Kesaulya, H., Zakaria, B., & Syaiful, S. A. (2015). Isolation and physiological characterization of PGPR from potato plant rhizosphere in medium land of Buru Island. Procedia Food Science, 3, 190-199. https://doi.org/10.1016/j.profoo. 2015.01.021.
Kobayashi, T., & Nishizawa, N.K. (2012). Iron uptake, trans- location, and regulation in higher plants. Annu Rev Plant Biol, 63, 131–152.
Kraemer, S.M. (2004). Iron oxide dissolution and solubility in the presence of siderophores. Aquat Sci, 66, 3–18.
Ma, J.F. (2005). Plant root responses to three abundant soil minerals: silicon, aluminum and iron. Crit RevPlant Sci, 24, 267–281.
Matzanke, B.F. (1991). Structures, coordination chemistry and functions of microbial iron chelates. In CRC Handbook of Microbial Iron Chelates. Winkelmann, G. (ed.). Boca Raton, FL, USA: CRC Press, pp. 15–64.
Milagres, A. M., Machuca, A., & Napoleao, D. (1999). Detection of siderophore production from several fungi and bacteria by a modification of chrome azurol S (CAS) agar plate assay. Journal of Microbiological Methods, 37(1), 1-6. https://doi.org/10.14219/jada.arc hive.1964.0123.
Plattner, H., & Diekmann, H. (1994). Enzymology of siderophore biosynthesis. In Metal Ions in Fungi (G. Winkelmann & D. R. Winge, eds) pp. 99-116. Marcel Dekker, New York.
Pratama, Ilham, Advinda, L., & Fifendy, M. (2018). Pengaruh Sumber Karbon Terhadap Produksi Siderefor dari

156
Cendawan Endofit
Bakteri Pseudomonas Fluoresence. Bioscience, 2(2), 50. https://doi.org/10.24036/020182 210406-0-00.
Prihatiningsih, Nur, Djatmiko, HA., & Lestari, P. (2017). Aktivitas Siderofor Bacillus Subtilis Sebagai Pemacu Pertumbuhan Dan Pengendali Patogen Tanaman Terung. HPT Tropika, 17(2), 170–78.
Schwyn, B., & Neilands, J.B. (1987). Universal chemical assay for the detection and determination of siderophores. Anal Biochem, 160, 47–56.
Sharma, A., & Johri, BN. (2003). Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9, in maize (Zea mays L.) under iron limiting conditions. Microbiol. Res, 158(3), 243–248.
Sivasakthivelan, P., & Stella, D. (2012). Studies on the phytohormone producing potential of agriculturally beneficial microbial (ABM) isolates from different rhizosphere soils of sunflower in Tamil Nadu. International Journal of Pharmaceutical and Biological Archives, 3(5), 1150-1156.
Winkelmann, G. (2002). Microbial siderophore mediated transport. Biochemical Society Transactions, 30, 691-695.

Cendawan Endofit
157
Lampiran 5. Hasil riset, publikasi pada jurnal Agrotan. Judul: Isolasi Dan Identifikasi Cendawan Endofit Tanaman Padi Aromatik Lokal Enrekang. https://ejournals.umma.ac.id/index.php/agrotan/article/view/8/6
Isolasi Dan Identifikasi Cendawan Endofit Tanaman Padi Aromatik Lokal Enrekang
Isolation and Identification of Endophytic Fungus on Local Aromatic Rice Plants of Enrekang
Syamsia1)
E-mail: [email protected] 1)Fakultas Pertanian Universitas Muhammadiyah Makassar
ABSTRACTThis study aims to isolate and identify endophytic fungi in local aromatic rice plants of Enrekang. Isolation of endophytic fungi was carried on three local aromatic rice varieties from Enrekang namely Pulu Mandoti, Pulu Lotong and Pare Lambau. Isolation of the fungus was carried out at the roots, stems and leaves of the rice. Fungal isolates were purified and then identified macroscopically and microscopically. In this study 12 isolates of endophytic fungi were produced. The results of the identification of four isolates of endophytic fungus elected were Aspergillus sp1, Aspergillus sp2, Aspergillus candidus and acremonium. Keywords: Acremonium, Aspergillus Candidus, aromatic local
ABSTRAKPenelitian ini bertujuan untuk mengisolasi dan mengindetifikasi cendawan endofit tanaman padi aromatik lokal Enrekang. Isolasi

158
Cendawan Endofit
cendawan endofit dilakukan pada tigas jenis padi aromatik lokal Enrekang yaitu Pulu Mandoti, Pulu Lotong dan Pare Lambau. Isolasi cendawan dilakukan pada bagian akar, batang dan daun padi. Isolat cendawan yang telah dimurnikan kemudian didentifikasi secara makroskopis dan mikroskopis. Dalam penelitian ini dihasilkan 12 isolat cendawan endofit. Hasil identifikasi terhadap empat isolat cendawan endofit terpilih diperoleh Aspergillus sp1, Aspergillus sp2, Aspergillus candidus dan acremonium. Kata kunci: Acremonium, Aspergillus candidus, aromatik lokal
PENDAHULUANPadi aromatik lokal Enrekang merupakan padi yang
memiliki aroma pandan ditanam secara turun temurun oleh masyarakat di Kecamatan Salukanang Kabupaten Enrekang. Padi aromatik lokal termasuk padi umur dalam (sekitar 6 bulan) sehingga pertanaman hanya 1 kali dalam setahun. Jenis-jenis padi aromatik lokal di daerah ini adalah Pulu Mandoti, Pare Salle, Pare Pulu Lotong, Pare Pinjan, Pare Pallan, Pare Solo, Pare Mansur, Pare Kamida dan Pare Lambau.
Cendawan endofit biasanya hidup dalam jaringan tanaman tanpa menimbulkan gejala terhadap tanaman inang dan mendapat perhatian cukup besar beberapa dekade terakhir (Faeth, 2002). Hasil eksplorasi cendawan endofit pada tanaman padi di lahan PHT dan konvensional diperoleh cendawan Aspergillus sp., Penicillium sp., Nigrospora sp., Trichoderma sp., Curvularia sp, sedangkan jamur yang hanya terdapat di lahan PHT yaitu Mucor sp., Mastigosporium sp., Alternaria sp., Fusarium sp., dan Monosporium sp. Sedangkan jamur yang hanya ditemukan di lahan konvensional yaitu Verticillim sp dan Acremonium sp. (Ariyanto, et al. 2013). Cendawan endofit yang paling sering

Cendawan Endofit
159
mendominasi pelepah batang padi adalah Nigrospora sp. (Irmawan, 2007). Cendawan endofit yang terdapat pada tanaman manggis Phoma, sp., Aremonium sp., Penicillium sp., Geotrichum sp., Pestalotiopsis sp., Botryosphaeria sp., Colletotrichum sp. Chysosporium sp., Aspergillus sp., dan Blasomyces sp. (Akmalasari, et al. 2013)
Beberapa cendawan endofit telah diidentifikasi dan memiliki kemampuan sebagai biokontrol adalah: 1) cendawan A. niger memiliki kemampuan dalam menghambat pertumbuhan cendawan patogen karena memproduksi enzim hidrolitik seperti lipase, protease, selulase, pektinase (Schuster, et al. 2002). Cendawan endofit Nigrospora 1, Nigrospora 2 dan Nigrospora 3 mampu meningduksi ketahanan tanaman padi terhadap wereng batang padi coklat Nilaparvata lugens Stahl (Budiprakoso, 2010). Cendawan endofit Nigrospora sp. dapat memperlambat pertumbuhan populasi wereng batang coklat di laboratorium (Mawan, et al. 2015).
Beberapa hasil penelitian juga menunjukkan bahwa cendawan endofit sebagai biofertilizer yaitu; 1) inokulasi Aspergilus niger dapat meningkatkan pertumbuhan tanaman padi dan jagung dengan dosis pemupukan fosfor 50% (Sutoyo, 2009). Perlakuan filtrate cendawan Aspergillus sp dan Fusarium sp dapat mempengaruhi daya berkecambah benih, persentasi hidup, tinggi, diameter, NPA, IMB dan biomasa bibit sengon (Yuniarti, et al. 2013). Berdasarkan uraian tersebut, penelitian ini bertujuan untuk mengisolasi dan mengidentifikasi cendawan endofit dari tanaman padi aromatik lokal Enrekang.

160
Cendawan Endofit
METODOLOGIIsolasi Cendawan Endofit Padi Aromatik Lokal Enrekang
Isolasi cendawan endofit dari bagian akar, batang dan daun padi aromatik lokal Enrekang menggunakan metode metode Rodriques (Wilia et al., 2012) yang dimodifikasi. Tahapan awal sterilisasi dimulai dengan mencuci mencuci akar, batang dan daun padi dengan air mengalir. Kemudian sterilisasi permukaan dilakukan secara bertahap dengan merendam sampel tanaman (akar, batang dan daun) selama 60 detik dalam etanol 70%, NaOCl 3% selama 60 detik, dan etanol 70% selama 30 detik. Kemudian dibilas sebanyak empat kali dengan aquades steril dikeringkan di atas kertas saring steril. Bagian tanaman dipotong kecil untuk ditumbuhkan dalam media PDA.
Pemurnian isolat cendawan endofit Cendawan endofit yang telah tumbuh pada potongan
jaringan tanaman selama proses propagasi koloni yaitu memotong dan mentransfer secara aseptik bagian sebagian miselium cendawan ke dalam media kultur baru secara aseptik. Cendawan yang tumbuh diambil dengan menggunakan jarum ose yang sebelumnya dipijarkan di aatas api kemudian digoreskan ke media PDA yang baru dan diinkubasi pada suhu ruangan selama 3-5 hari.
Identifikasi cendawan endofit Identifikasi cendawan endofit dilakukan dengan mengamati
karakter morfologi makroskopi dan mikroskopi. Pengamatan makroskopi meliputi cendawan meliputi warna dan permukaan koloni, garis-garis radial dari pusat koloni ke arah tepi koloni dan lingkaran-lingkaran konsentris. Pengamatan mikroskopi preparat

Cendawan Endofit
161
meliputi bentuk hifa, ada atau tidaknya rhizoid, bentuk sel reproduksi seksualnya dan aseksualnya (Gandjar, 1999; Hafsari dan Asterina, 2013). Pengamatan morfologi mikroskopi dilakukan dengan membuat preparatat untuk pengamatan. Cara pembuatan preparat adalah gelas benda dibersihkan dengan alkohol kemudian dipanaskan sampai bebas lemak dan debu. Gelas benda ditetesi laktofenol pada bagian tengah. Biakan cendawan diambil secara aseptik menggunakan jarum ose, kemudian diletakkan di atas gelas benda yang telah ditetesi laktofenol, kemudian diberi sedikit alkohol. Preparat ditutup dengan kaca penutup dan dilewatkan di atas api lalu dilihat di bawah mikroskop untuk mendapatkan ciri mikroskopik- nya. Identifikasi dilakukan dengan mencocokkan karakteristik isolat cendawan dengan buku identifikasi. (Purwantisari dan Hastuti, 2009).
HASILHasil isolasi cendawan endofit pada bagian jaringan tanaman
padi aromatik lokal Enrekang didapatkan 12 isolat yaitu 6 isolat dari Pare Lambau dan masing-masing 3 isolat dari Pulu Lotong dan Pulu Mandoti. Karakteristik makroskopi dan mikroskopi terhadap empat isolat terpilih mempelihatkan hasil yang berbed-beda seperti yang tersaji pada Tabel 1.
Tabel 1. Karakteristik Makroskopi, Mikroskopi dan Identifikasi Cendawan Endofit Padi Aromatik Lokal Enrekag

162
Cendawan Endofit
PEMBAHASANIsolat PL- A
Isolat PL-A adalah isolat cendawan endofit yang diisolasi dari bagian akar padi Pulu Lotong Isolat PL-A mempunyai ciri-ciri: warna koloni bagian atas putih, bagian bawah krem, permukaan berbulu halus, Konidia berbentuk bulat oval, warna konidia hialin, permukaan konidiofor halus berwarna coklat muda, phialid berbentuk tegak, hifa bersepta. Hasil pengamatan makroskopi dan mikroskopi isolat ini sesuai dengan karakter Aspergillu sp. Secara mikroskopis cendwan Aspergillus mudah dikenali dan dibedakan dari cendawan marga laein, yaitu memiliki konidiofor yang tegak, tidak bersepta, tidak bercabang, dan ujun konidiofor membengkak membentuk vesikel. Pada permukaan vesikel ditutupi fialid yang menghasilkan konidia. Konidia tersusun satu sel (tidak bersepta), globus memiliki warna yang

Cendawan Endofit
163
beragam dan tersusun membentuk rantai basipetal (Barnett, 1995; Ilyas, 2006).
Gambar 1. Permukaan atas (a), permukaan bawah (b), mikroskopi (c) isolat PL-A
Isolat PL-D Isolat PL-D adalah isolat cendawan endofit dari bagian daun
padi Pulu Lotong memiliki ciri-ciri: warna bagian atas koloni hijau pinggir putih, permukaan bawah krem, permukaan koloni berbulu halus dengan tipe pertumbuhan radial. Konidia berbentuk bulat dan hialin, Permukaan konidiofr halus dan berwarna coklat muda, phialid berbentuk tegak, memiliki hifa bersepta dan bercabang. Hasil pengamatan makroskopi dan mikroskopi isolat ini sesuai dengan karakter Aspergillu sp. Menurut Domsch dan Gams (1980) dan Barnett dan Hunter (1972); Akmalasari, et al. (2013), menyatakan bahwa Aspergillus sp. Memiliki ciri warna koloni kuning, hijau, kekuning, hijau tua kebiruan, vesikel berentuk bulat atau elips, stipe berwarna kuning atau hyalin, mempunyai fialid, mempunyai konidia satu bersel bebentuk bulat sampai elips.
Gambar 2. Permukaan atas (a), permukaan bawah (b), mikroskopi (c) isolat PL-D
Isolat PM-A Isolat PM-A adalah isolat cendawan endofit yang diisolasi
dari padi Pulu Mandoti memiliki ciri-ciri: warna koloni bagian atas dan bawah putih, permukaan koloni halus dan bergelombang, tipe pertumbuhan radial. Konidia berbentuk elips berukuran

164
Cendawan Endofit
Elips berukuran 2.2 – 3.4 μm dengan warna hialin, Permukaan konidiofor halus dan berwarna krem, phialid berbentuk tegak Tegak, berukuran panjang 5.6 -10 μm dan lebar 2.2 – 3.4 μm, hifa hialin. Menurut Domsch dan Gams (1980) dan Barnett dan Hunter (1972); Akmalasari, et al. (2013), menyatakan bahwa Aspergillus sp. Memiliki ciri warna koloni kuning, hijau, kekuning, hijau tua kebiruan, vesikel berentuk bulat atau elips, stipe berwarna kuning atau hyalin, mempunyai fialid, mempunyai konidia satu bersel bebentuk bulat sampai elips. hialin. Menurut Domsch dan Gams, (1980) dan Gandjar et al. (1999); Akmalasari, et al. (2013), menyatakan bahwa Acremonium sp. Mempunyai ciri-ciri warna koloni putih sampai coklat, permukaan koloni dibagian tengah tampak seperti kapas, konidiofor bercabang umumnya dilapisi kromafil, mempunyai fialid. Konida bersel satu tampak agak bergerombol membentuk satu kepala, bentuk konidia menajang hingga bulat, hifa bersepta dan kadang-kadang terbentuk klamidospora.
Gambar 3. Permukaan atas (a), permukaan bawah (b), mikroskopi (c) Isolat PM-A.
Isolat PM-B Isolat PM-B adalah isolat cendwan endofit yang diisolasi
dari bagian batang padi Pulu Mandoti mempunyai ciri: warna permukaan atas koloni putih, permukaan bahwah krem/coklat muda., tekstur permukaan koloni kasar, Konidia berbentuk bulat silinder berwarna hialin. Permukaan konidiofor halus dan warna transparan. Philaid berbentuk tegak dan berlendir disetiap puncak phialid. Hifa

Cendawan Endofit
165
Gambar 4. Permukaan atas (a), permukaan bawah (b), mikroskopi, dan (c) Isolat PM-B.
KESIMPULAN Berdasarkan hasil isolasi dan pemurnian isolat diperoleh 12
isolat cendawan endofit dari padi aromatik lokal Enrekang. Hasil identifikasi terhadap empat isolat cendawan endofit yaitu isolat PL-A adalah Aspergillus sp 1, isolat PL-D adalah Aspergillus sp 2, isolat PM-A adalah Aspergillus candidus dan isolat PM-B adalah Acremonium.
DAFTAR PUSTAKAAkmalasari, I., Purwati, E. S., & Dewi, S. (2013). Isolasi dan
Identifikasi Jamur Endofit Tanaman Manggis (Garcinia mangostana L.). Biosfera, 30(2), 82–89.
Ariyanto, E. F., Abadi, A. L., & Djauhari, S. (2013). Keanekaragaman Jamur Endofit pada Daun Tanaman Padi (Oryza Sativa L.) dengan Sistem Pengelolaan Hama Terpadu (PHT) dan Konvensional DI Desa Bayem, Kecamatan Kasembon, Kabupaten Malang. Jurnal HPT, 1(2), 37–51.
Barnett, H.L. and B.B. Hunter. 1972. Illustrated Genera of Imperfect Fungi. Third Edition. Burgess Publishing. Company. Minnesote.
Domch, K.H and W. Gams. 1980. Compendium of soil fungi volume 1. Academic Press, London.
Gandjar, I., R.A. Samson, K. Twell- Vermeulen, A. Oetari dan I. Santoso. 1999. Pengenalan Kapang Tropik Umum. Yayasan Obor Indonesia, Jakarta.
Budiprakoso, B. (2010). Pemanfaatan Cendawan Endofit sebagai Penginduksi Ketahanan Tanaman Padi terhadap

166
Cendawan Endofit
Wereng Coklat Nilaparvata lugens (Stal). (Hemiptera: Delphacidae). Institut Pertanian Bogor.
Faeth, S. H. (2002). Are endophytic fungi defensive plant mutualists? Oikos, 98(1), 25–36. http://doi.org/10.1034/j.1600- 0706.2002.980103.x
Hafsari, A. R., & Asterina, I. (2013). Isolasi dan Identifikasi kapang Endofit dari Tanaman Obat Surian (Toona sinensis). Jurnal Istek, 7(2), 175–191.
Ilyas, M. (2006). Isolasi dan Identifikasi Kapang pada Relung Rizosfir Tanaman di Kawasan Cagar Alam Gunung Mutis, Nusa Tenggara Timur. Biodiversitas, 7(83), 216– 220. http://doi.org/10.13057/biodiv/d070 304
Irmawan, D. E. (2007). Kelimpahan dan Keragaman Cendawan ENdofit pada Beberapa Varietas Padi di Kuningan, Tasikmalaya dan Subagng Jawa Barat. Institut Pertanian Bogor.
Mawan, A., Buchori, D., & Triwidodo, H. (2015). Pengaruh cendawan endofit terhadap biologi dan statistik demografi wereng batang cokelat Nilaparvata lugens Stál (Hemiptera: Delphacidae). Jurnal Entomologi Indonesia, 12(1), 11– 19. http://doi.org/10.5994/jei.12.1.11
Purwantisari, S., & Hastuti, R. B. (2009). Isolasi dan Identifikasi Jamur Indigenous Rhizosfer Tanaman Kentang dari Lahan Pertanian Kentang Organik di Desa Pakis Magelang. Jurnal Bioma, 11(2), 45– 53.
Schuster, E., Dunn-Coleman, N., Frisvad, J., & Van Dijck, P. (2002). On the safety of Aspergillus niger - A review. Applied Microbiology and Biotechnology, 59(4–5), 426– 435. http://doi.org/10.1007/s00253- 002-1032-6
Sutoyo. (2009). Pengaruh Inokulasi Cendawan Endofit Akar.

Cendawan Endofit
167
Institut Pertanian Bogor. Wilia, W., Hayati, I., & Ristiyadi, D. (2012). Eksplorasi cendawan
endofit dari tanaman padi sebagai aggens pemacu pertumbuhan tanaman. Program Studi Agroekoteknologi, Fakultas Pertanian Universitas Jambi, 1(4), 73–79. Yuniarti, N., Suharti, T., & Bramasto, Y. (2013). Pengaruh Filtrat Cendawan Aspergillus sp dan Fusarium sp terhadap Viabilitas Benih dan Pertumbuhan Bibit Sengon (Paraserianthes falcataria). Jurnal Penelitian Kehutanan Wallace, 2(2), 93–103.

168
Cendawan Endofit
Lampiran 6
PRODUKSI GIBERELIN DAN IAA CENDAWAN ENDOFIT
ASAL PADI LOKAL SULAWESI SELATAN
Syamsia Syamsia1, Abubakar Idhan2, Amanda Patappari Firmansyah3
1,2,3 Program Studi Agroteknologi Fakultas Pertanian Universitas Muhammadiyah Makassar
Jl. Sultan Alauddin No. 259 Makassar Email: [email protected]
AbstrakGiberelin (GA3) dan Indole Acetat Acid (IAA) berperan dalan proses perkembangan dan fisiologis tanaman seperti perkecambahan biji, pertumbuhan, inisiasi bunga serta pembentukan bunga dan buah. Cendawan memiliki kemampuan menghasilkan fitohormon seperti Giberelin dan IAA. Penelitian ini bertujuan untuk mengetahui potensi isolat cendawan endofit yang telah diisolasi padi lokal dalam memproduksi Giberelin dan IAA. Pengujian IAA menggunakan media Potato Dextrose Broth (PDB) dengan penambahan L- Triptophan dan menggunakan reagen Salkowski. Pengujian Giberelin mengikuti metode Borrow. Pengukuran absorbansi panjang gelombang 535 nm untuk IAA dan Giberelin pada 254 nm menggunakan UV-VIS spectrophotometer. Penentuan konsentrasi IAA dan Giberelin berdasarkan kurva standar. Hasil penelitian menunjukkan

Cendawan Endofit
169
bahwa kemampuan isolat cendawan endofit dalam memproduksi giberelin bervariasi 0.81- 2.35 ppm dan IAA 0.23-2.36 ppm. Isolat E1 terbaik dalam memproduksi Giberelin dan E3 terbaik dalam produksi IAA.
Kata Kunci: fitohormon, L-Triptophan, Potato Dextrose Broth
1. PENDAHULUANGiberelin dan IAA merupakan hormon yang berperan
penting dalam pertumbuhan tanaman, reproduksi, metabolisme dan merespon berbagai perubahan lingkungan (Waqas et al. 2012).
Giberelin berperan dalam proses pertumbuhan tanaman seperti perkecambahan biji, pemanjangan batang, pembungaan dan perkembangan buah (Salazar-Cerezo et al. 2018).
IAA berperan dalam perkembangan akar, pembentukan kuncup dan bunga serta proses lain dalam tanaman, dan sangat diperlukan untuk pertumbuhan dan perkembangan tanaman (Reinhard et al. 2000; Lubna et al. 2018).
Giberelin ditemukan pada tumbuhan, alga, cendawan dan bakteri, namun konsentrasi tertinggi ditemukan pada jamur (Waqas et al. 2012). IAA banyak ditemukan pada tanaman dan berperan dalam pemanjangan sel, pembelahan sel dan diferensiasi sel serta inisiasi akar (Quint, 2006; Zao, 2010; Numponsak et al. 2018).
Cendawan endofit dapat memproduksi beberapa fitohormon seperti IAA, Giberelin dan Sitokinin (Wulandari and Suryantini 2019). Cendawan pelapuk yang diisiolasi dari tanaman kakao mampu memproduksi Giberelin (Rahim, et al. 2019). Cendawan yang diisolasi dari dari perakaran kelapa

170
Cendawan Endofit
sawit dapat memproduksi IAA (Sapareng et al. 2017).Beberapa cendawan endofit dapat menghasilkan
indole acetic acid (IAA) (Ansari et al. 2013; Waqas et al. 2014;(Lubna et al. 2018). Cendawan endofit Phoma glomerata dan Penicillium sp. LWL3 mampu memproduksi Giberelin dan IAA (Waqas et al. 2012).
Penelitian ini bertujuan untuk mengetahui kemampuan isolate cendawan endofit asal padi lokal Sulawesi Selatan dalam memproduksi IAA dan Giberelin.
2. BAHAN DAN METODE2.1 Bahan
Bahan yang digunakan adalah: isolate cendawan endofit, PDA, PDB, Zinc acetat, Potassium ferrocyanide, asam klorida, L-tryptophan, FeCl3
, dan H2SO4, aquades.
2.1 Peremajaan isolate Cendawan Endofite Koleksi isolate cendawan endofit yang telah diperoleh dari
penelitian sebelumnya. Sebanyak 8 isolat ditumbuhkan kembali pada media Potato Dextrose Agar (PDA) dan diinkubasi pada suhu kamar hingga umur 7 hari. Isolat yang telah tumbuh selanjutnya ditumbuhkan pada media Potato Dextrose Broth (PDB).
2.2 Produksi GiberelinProduksi hormon GA3 isolat cendawan pelapuk diukur
menggunakan metode Borrow et al. 1955; Kesaulya, 2015). Sebanyak 5 lempeng isolate cendawan endofit diambil dari media PDA dengan menggunakan cork bohrer, ditumbuhkan pada media PDB, dan diinkubasi pada suhu kamar selama 7

Cendawan Endofit
171
hari. Setelah itu, kultur isolat cendawan disentrifuse pada 8000 rpm selama 10 menit, dan 15 ml kultur isolate cendawan endofit diambil dan dimasukkan ke dalam tabung reaksi dan ditambahkan 2 ml larutan Zinc acetat. Setelah 2 menit, ditambahkan 2 ml larutan Potassium ferrocyanide dan disentrifuse pada 8000 rpm selama 10 menit. Supernata sebanyak 5 ml dimasukkan dalam tabung reaksi dan ditambahkan 5 ml asam klorida 30% dan dinkubasi pada suhu kamar selama 75 menit. Blanko dipersiapkan dengan asam klorida 5%. Absorbansi diukur pada panjang gelombang 254 nm menggunakan spektrofotometer UV-VIS. Konsentrasi GA3 dibandingkan dengan kurva standar GA3 dengan persamaan regresi Y = 0,888 X + 0,441 dimana R2 = 0, 921 (Kesaulya, 2015).
2.3 Produksi Indole Acetic Acid (IAA)Produksi IAA oleh cendawan endofit diuji menggunakan
media PDB dan reagen Salkowski. Isolat cendawan endofit yang telah diremajakan ditumbuhkan pada media PDB yang telah ditambahkan L-tryptophan (0,1 gl -1 ) pada suhu 28 C dalam kondisi gelap selama 5 hari. Supernata disentrifugasi selama 10 menit dengan 8000 rpm. Sebanyak 1 ml supernata dimasukkan dalam tabung reaksi dan ditambahkan 4 ml reagen Salkowski (12 gl1 FeCl3
dalam 429 l-1 ml H2SO4) (Glickman dan Dessaus, 1995; Kesaulya, 2015) dan disimpan dalam suhu 28 C dalam kondisi gelap selama 24 jam.

172
Cendawan Endofit
Absorbansi diukur pada panjang gelombang 535 nm pada spektrofotometer UV-VIS. Konsentrasi auksin diukur dengan menggunakan kurva standar IAA menggunakan persamaan regresi Y= 0.064X + 0.09 dimana R2 =0.995 (Kesaulya, 2015).
3. HASIL DAN PEMBAHASAN3.1 Peremejaan dan Persiapan Kultur Isolat Cendawan
Endofit Sebanyak 8 isolat cendawan endofit diremajakan dengan
cara ditumbuhkan pada media PDA hingga diperoleh isolat murni (Gambar 1a), selanjutnya isolat ditumbuhkan pada media PDB (Gambar 1b) yang akan digunakan untuk pengujian Giberelin dan IAA. Media PDB untuk pengujian IAA diberi tambahan L-tryptophan. Menurut Dewi et al. 2016); (Gusmiaty et al. 2018), L-tryptophan merupakan asam amino yang berperan sebagai prekursor dalam pembentukan IAA pada tanaman dan mikroorganisme
Gambar 1. Isolat Cendawan Endofit pada media PDA (a), media PDB (b)
3.2 Uji Produksi hormon GiberelinHasil uji kemampuan isolate cendawan endofit dalam
memproduksi Giberelin menunjukkan bahwa kedelapan isolat
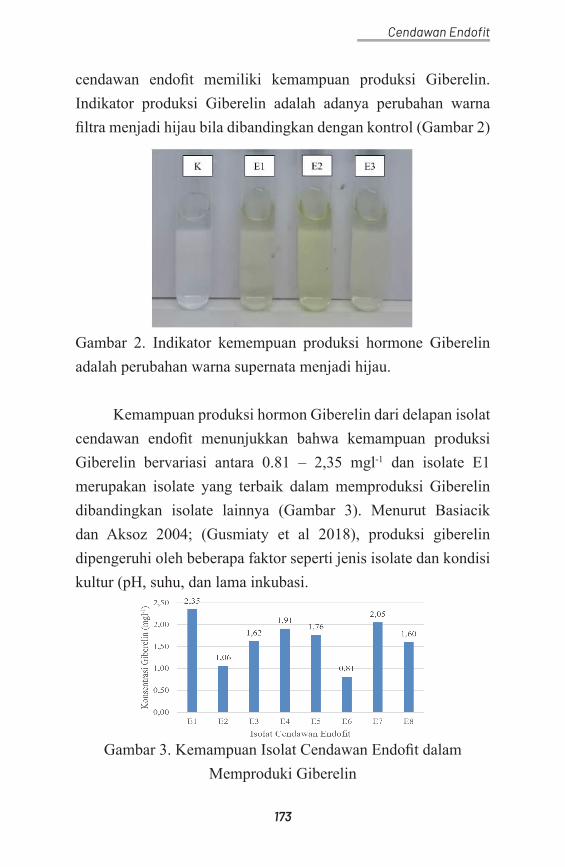
Cendawan Endofit
173
cendawan endofit memiliki kemampuan produksi Giberelin. Indikator produksi Giberelin adalah adanya perubahan warna filtra menjadi hijau bila dibandingkan dengan kontrol (Gambar 2)
Gambar 2. Indikator kemempuan produksi hormone Giberelin adalah perubahan warna supernata menjadi hijau.
Kemampuan produksi hormon Giberelin dari delapan isolat cendawan endofit menunjukkan bahwa kemampuan produksi Giberelin bervariasi antara 0.81 – 2,35 mgl-1 dan isolate E1 merupakan isolate yang terbaik dalam memproduksi Giberelin dibandingkan isolate lainnya (Gambar 3). Menurut Basiacik dan Aksoz 2004; (Gusmiaty et al 2018), produksi giberelin dipengeruhi oleh beberapa faktor seperti jenis isolate dan kondisi kultur (pH, suhu, dan lama inkubasi.
Gambar 3. Kemampuan Isolat Cendawan Endofit dalam Memproduki Giberelin

174
Cendawan Endofit
3.3 Produksi hormoan IAAHasil uji kemampuan isolate cendawan endofit dalam
memproduksi hormon IAA menunjukkan bahwa kedelapan isolate memiliki kemampuan produksi IAA bervarias. Indikator kemampuan memproduksi hormon IAA adalah perubahan warna supernata menjadi pink (Gambar 4), semakin cerah warna semakin tinggi kemampuan memproduksi IAA.
Gambar 4. Indikator kemampuan produksi IAA adalah perubahan warna kultur menjadi pink
Konsentrasi IAA yang dihasilkan oleh kedelapan isolate cendawan endofit menunjukkan bahwa kemampuan isolate cendawan endofit memproduksi IAA berkisar antara 0,23- 2,36 mgl-1. Isolat E1 memperlihatkan kemampuan terbaik dalam memproduksi IAA dibandingkan isolate lainnya (Gambar 5). Hasil tersebut hampir sama dengan kemampuan rhizobacteria dalam memproduksi IAA yaitu 2,370 mgl-1 (Gusmiaty et al 2018), namun penelitian (Sapareng et al. 2017) menunjukkan hasil yang lebih tinggi yaitu 8,429 mgl-1. Hal ini menunjukkan

Cendawan Endofit
175
bahwa kemampuan produksi IAA dipengaruhi oleh beberapa factor seperti, jenis mikorba, pH media dan nutrisi/unsur hara (Wulandari and Suryantini 2019).
Gambar 5. Kemampuan Isolat cendawan Endofit dalam Memproduksi IAA
4. KESIMPULAN DAN SARAN Isolat cendawan endofit memiliki kemampuan produksi
Giberelin bervariasi antara 0.81 – 2,35 mgl-1 dan IAA antara 0,23- 2,36 mgl-1. Isolat E1 terbaik dalam memproduksi Giberelin E1 dan E3 terbaik dalam memproduksi IAA.
Perlu dilakukan penelitian lanjutan untuk mengetahui konsorsium isolate terbaik dalam memproduksi Giberelin dan IAA
5. UCAPAN TERIMA KASIHTerima Kasih Kepada Kementrian Riset dan Teknologi/
Badan Riset dan Inovasi Nasional yang telah mendanai kegiatan ini melalui Hibah Penelitian Dasar Unggulan Perguruan Tinggi (PDUPT) selama tiga tahun (2018-2020)

176
Cendawan Endofit
6. DAFTAR PUSTAKAAnsari MW, Trivedi DK, Sahoo RK, Gill SS, Tuteja N. 2013.
A critical review on fungi mediated plant responses with special emphasis to Piriformospora indica on improved production and protection of crops. Plant Physiol Biochem. 70:403–410
Basiacik and Aksoz 2004 Optimation of carbon-nitrogen ratio for production of gibberellic acid by Pseudomonas sp. J Microbiol. 53 117–20
Dewi KT, Jodi S, Dwi A 2016. Isolasi dan Uji Aktivitas Bakteri Penghasil Hormon Tumbuh IAA (Indole3-Acetic Acid) dan Bakteri Perombak Protein dari Tanah Pertanian Tual, Maluku Tenggara in Prosiding Seminar Nasional Masyarakat Biodiversitas Indonesia (International Conference on Biodiversity: 271–276
Gilickmann E, Dessaux YA. 1995. Critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl Environ Microb 61: 793-796.
Gusmiaty, M Restu, B Bachtiar, and SH Larekeng. 2018. “Gibberellin And IAA Production by Rhizobacteria From Various Private Forest.” IOP Conference Series: Earth and Environmental Science 270 (May): 012018.
Kesaulya, Baharuddin, B.Zakariah, Syatrianty A. Syaiful. 2015. Isolation and Physiological Characterization of PGPR from Potato Plant Rhizosphere in Medium Land of Buru Island. Procedia Food Science 3 : 190-199.
Lubna, Sajjad Asaf, Muhammad Hamayun, Humaira Gul, In-Jung Lee, and Anwar Hussain. 2018. “Aspergillus Niger CSR3 Regulates Plant Endogenous Hormones and Secondary

Cendawan Endofit
177
Metabolites by Producing Gibberellins and Indoleacetic Acid.” Journal of Plant Interactions 13 (1): 100–111.
Numponsak, Tosapon, Jaturong Kumla, Nakarin Suwannarach, Kenji Matsui, and Saisamorn Lumyong. 2018. Biosynthetic Pathway and Optimal Conditions for the Production of Indole-3-Acetic Acid by an Endophytic Fungus, Colletotrichum Fructicola CMU-A109.Edited by Sabrina Sarrocco. PLOS ONE 13.
Quint M, Gray WM 2006. Auxin signaling. Curr Opin Plant Biol 9: 448–453.
Rahim, Iradhatullah, Suherman Seherman, and Hakzah Hakzah. 2019. “Produksi Hormon Giberelin Dari Cendawan Pelapuk Asal Tanaman Kakao.” In Prosiding Seminar Nasional SMIPT 2:26–27.
Borrow A, Brian PW, Chester VE, Curtis PJ, Hemming HG, Henehan C, Jeffreys EG, Lloyd PB, Nixon IS, Norris GL, Radley M. 1955. Gibberellic acid, a metabolic product of the fungus Gibberella fujikuroi: some observations on its production and isolation. J Sci Food Agric 6 (6): 340-348.
Reinhardt D, Mandel T, Kuhlemeier C. 2000. Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell. 12 (4):507–518.
Salazar-Cerezo, Sonia, Nancy Martinez-Montiel, Maria del Carmen Cruz-Lopez, and Rebeca D. Martinez-Contreras. 2018. “Fungal Diversity and Community Composition of Culturable Fungi in Stanhopea Trigrina Cast Gibberellin Producers.” Frontiers in Microbiology 9 (APR): 1–15.
Sapareng, Sukriming, Ambo Ala, Tutik Kuswinanti, and Burhanuddin Rasyid. 2017. “Capability of Rot Fungus Isolates from Oil Palm Empty Bunches in the Production of

178
Cendawan Endofit
Indole Acetic Acid (IAA).” International Journal of Current Microbiology and Applied Sciences 6 (11):2174–80.
Waqas, Muhammad, Abdul Latif Khan, Muhammad Kamran, Muhammad Hamayun, Sang Mo Kang, Yoon Ha Kim, and In Jung Lee. 2012a. “Endophytic Fungi Produce Gibberellins and Indoleacetic Acid and Promotes Host-Plant Growth during Stress.” Molecules 17 (9): 10754–73.
Wulandari, Reine Suci, and Rosa Suryantini. 2019. “Endophytic Fungi in Paraserianthes Falcataria: Production of Indole Acetic Acid.” Microbiology Indonesia 13 (1): 16–24.
Zhao Y. 2010. Auxin biosynthesis and its role in plant development. Annu Rev Plant Biol 61: 49–64.

Cendawan Endofit
179
GLOSARIUM
Annealing
Aquades
AuksinAutoklaf
Biokontrol
Bioremediasi
Cendawan endofit
Denaturasi
DekstroseDrying Method
Endofit
Ekstensi
:
:
::
:
:
:
:
::
:
:
merupakan tahap penempelan primer pada rantai tunggal DNAair hasil destilasi/penyulingan yang sama dengan air murni atau H2OHormon pemacu pertumbuhan tanamanAlat untuk sterilisasi dengan metode uap panasPemberantasan hama dan penyakit tanaman dengan menggunakan parasite atau musuh alamipenggunaan mikoorganisem untuk mengu-rangi polutan di lingkungan.Cendawan yang hidup dalam jaringan tanaman tanpa menimbulkan gejala pada tanaman inangProses pemisahan rantai DNA templat pada proses PCRgula sederhanaMetode penyimpanan kering dengan menurunkan kadar air di sekitar mikrobaDari Bahasa Yunani, endo artinya dalam dan phyte artinya tanaman. Jadi endofit adalah semua organisme yang selama periode siklus hidupnya berad dalam jaringan tanamanProses pemanjangan untai baru DNA

180
Cendawan Endofit
:
:
:
::::
:
:
::
:
Fitoremediasi
Filosfir
Frezing Method
FilosfirFilosfirFitohormonFreez-driying
Glasswar
Hifa
KitinaseLiophilization drying
L-tryptophan
dari sampel, dimulai dari posisi primer yang telah menempel di urutan basa nukleotida DNA target akan bergerak dari ujung 5’ menuju ujung 3’ dari untai tunggal DNA.Mekanisme menghilangkan polutan dari tanah atau perairan yang terkontaminasi dengan menggunakan tumbuhan Istilah yang digunakan untuk bagian tumbuhan yang berada di atas permukaan tanah sebagai habit mikroorganismeMetode penyimpanan beku dengan menurunkan suhu di sekitar mikroba sehingga laju metabolism turundaerah disekitar perakaran tanamanbagian daun tanamanHormone tumbuhMetode penyimpanan beku dengan proses evaporasiperalatan kecil yang terbuat dari gelas seperti botol shott, gelas ukur, Erlenmeyer, cawan petri dllstruktur cendawan berbentuk seperti tabung yang terbentuk dari pertumbuhan spora atau konidiaEnzim yang dapat menghidrolisis kitin Metode penyimpanan kering beku dengan proses sublimasimerupakan senyawa precursor pada

Cendawan Endofit
181
Makroskopis
Mikroskopis
Patogen
PhytohormonePurifikasi
RhizosfirRhizoid
Selulase
Sterilisasi permukaan
Subculturing Method
:
:
:
::
::
:
:
:
pengujian IAAbentuk atau struktur cendawan yang dapat dilihat secara langsung seperti warna, tipe pertumbuhan dan bentuk permukaanbentuk atau struktur cendawan yang hanya dapat diamati dengan menggunakan mikroskop, seperti: hifa, rhizoid, dan bentuk sporamikroorganisme parasit yang dapat menyebabkan penyakitzat pemacu pertumbuhan tanaman kegiatan pemurnian isolate cendawan endofit hingga diperoleh isolat murni atau spora tunggalDaerah sekitar perakaran tanamanstruktur menyerupai rambut atau benang-benangEnzim yang dapat mendegradasi selulosaMembersihkan permukaan sampel tanaman dari mikrobaMetode penyimpanan bersifat sementara dan perlu ditumbuhkan secara berkala dengan menumbuhkan pada medium tumbuh yang baru

182
Cendawan Endofit
INDEKS
AAnnealingAnnelaing 49Aquades:
Aquades 13, 14, 18, 19, 20, 22, 23, 26, 61Auksin
Auksin 63Autoklaf Autoklaf 9, 13, 14BBiokontrol:
Biokontrol 1,Bioremediasi
Bioremediasi 2 DDenaturasi Denaturasi 49,Dekstrose Dekstrose 12, 13, 17, ``18, 19Drying Method Drying Method 58FFitoremediasi
Fitoremediasi 2, Filosfir Filosfir 29, 37

Cendawan Endofit
183
Fitohormon Fitohormon 65, 66, 78, 79Freez-driying Freez driying 60, 61Frezing Method Frezing Method 60, 61GGlassware Glassware 11, 13HHifa
Hifa 39, 42KKitinase
Kitinase 75, 80LLaminar air flow
Laminar air flow 10, 13, 15Liophilization drying
Liophilizationg drying 60, 61, 63L-tryptophan L-tryptophan 66, 79MMakroskopis Makroskopis 39, 43, 53Mikroskopis Mikroskopis 41,44, 53PPurifikasi
Purifikasi 33, 35

184
Cendawan Endofit
RRhizosfir Rhizosfir 29, 37SSelulase Selulase 75Siderefor Siderefor 72, 74, 79, 80Subculturing Method
Subculturing Method 58, 61

Cendawan Endofit
185
Dr. Syamsia, SP., M. Si. Lahir di Pangkajene-Sidrap pada tanggal 15 Juni 1972, anak pertama dari pasangan H. Tayibe dan Hj. Alimang. Pendidikan SD dan SMP diselesaikan di Pangkajene-Sidrap. Tamat SMA Tahun 1989 di SMA Negeri 1 Makassar. Gelar Meraih Gelar Sarjana Pertanian pada Fakultas Pertanian Unhas tahun 1996. Lulus
Pendidikan Magister tahun 1999. Gelar Doktor diraih tahun 2015 di Program Pasca Sarjana Unhas. Terangkat sebagai CPNS Departemen Kehutanan sebagai Penyuluh Kehutanan pada tahun 2000. Terdaftar sebagai dosen tetap Fakultas Pertanian Universitas Muhammadiyah sejak tahun 2008 sampai sekarang dengan status dosen DPK LLDIKTI Wil. IX Sulawesi dan Gorontalo. Menjabat Ketua Program Studi Agribisnis periode (2010-2014). Aktif mengikuti kegiatan seminar nasional dan internasional. Menjadi anggota Perhimpunan Agronomi Indonesia (PERAGI) sejak tahun 2016 sampai sekarang. Tergabung dalam Forum Dosen Indonesia Komda Sulawesi Selatan. Meraih dana hibah penelitian skim Hibah Bersaing (2011-2014), Hibah Disertasi (2014), Hibah Produk Terapan (2015-2017), dan Hibah Penelitian Dasar Unggulan Perguruan Tinggi (2018-2020). Mendapatkan dana hibah pengabdian masyarakat IbM (tahun 2011 dan 2015), Kewirausahaan Kampus (2015-2017), PPDM (2018) dan PPUPIK (2018-2020). Mendapat penghargaan sebagai Poster Terbaik untuk penelitian Produk Terapan dari Kemenristek Dikti pada Tahun 2017.
BIODATA PENULIS

186
Cendawan Endofit
Dr. Ir. Abubakar Idhan, M. P. Lahir di Palampang pada tanggal 2 Juni 1958. Menyelesaikan Pendidikan SD dan SMP di Palampang Bulukumba. Menyelesaikan Pendidikan Sekolah Menengah Atas di SMA Negeri 3 Makassar. Gelas Insinyur diraih pada tahun 1986 di Jurusan Budidaya Tanaman Fakultas Pertanian Unhas. Gelas Magister Pertanian diraih penulis
pada Program Studi Sistem-Sistem Pertanian Pasca Sarjana. Penulis terdaftar sebagai mahasiswa program doktor pada tahun 2011 dan meraih gelar Doktor dalam Bidang Ilmu Pertanian pada tahun 2016. Menjabat sebagai Sekretaris LP3M Unismuh Makassar periode 2009-2016 dan saat ini sebagai Ketua LP3M Unismuh Makassar. Penulis telah beberapa kali mendapatkan dana penelitian Ristekdikti melalui skim penelitian Hibah Bersaing, Hibah Disertasi, dan Hibah Penelitian Terapan Unggulan Perguruan Tinggi. Demikian juga dengan skim pengabdian masyarakat diantaranya IbM, Kewirausahaan Kampus, IbDM dan PPUPIK. Berbagai artikel telah dipublikasin di beberapa jurnal internasional dan nasional terakreditasi.

Cendawan Endofit
187
Dr. Amanda Patappari Firmansyah, S.P., M. P. Lahir di Ujung Pandang, 9 Juli 1986 dari pasangan Alm. Drs Firmansyah Asape dan Ir. siLely Mardawati, M.P. Penulis meyelesaikan Sekolah Dasar di SD Ktolik Santo Yakobus Makassar, lalu melanjutkan ke Sekolah Menengah Pertama di SMP Negeri 5 Makassar. Pendidikan Sekolah Menengah Atas diselesaikan di SMU
Islam Athira Makassar, Gelar Sarjana Pertanian diraih tahun 2008 pada Prodi Ilmu Hama dan Penyakit Tumbuhan. Mendapat gelar Magister Pertanian pada Program Studi Sistem-Sistem Pertanian Pasca Sarjana Unhas tahun 2012. Gelar Doktor dalam Bidang Ilmu Pertania diraih pada tahun 2020 di Program Pasca Sarjana Unhas. Terdaftar sebagai dosen tetap Fakultas Pertanian Universitas Muhammadiyah Makassar sejak tahun 2014 sampai sekarang dengan status dosen tetap yayasan. Aktif mengikuti kegiatan seminar nasional dan internasional. Menjadi Anggota Perhimpunan Agronomi Indonesia (PERAGI). Penulis banyak melakukan penelitian terkait tanaman kakao, yakni pengelolaan hama dan penyakit tanaman kakao serta penelitian di bidang bahan alami tanaman sebagai pestisida nabati.

188
Cendawan Endofit
Noerfitryani, S. P., M. Si. Lulus S-1 di Program Studi Ilmu Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Hasanuddin Makassar tahun 2012, lulus S-2 di Program Studi Magister Ilmu Hama dan Penyakit Tumbuhan Universitas Hasanuddin Makassar tahun 2014. Saat ini menempuh pendidikan Doktor di China Agricultural University, Beijing-China.
Mengajar di Fakultas Pertanian Universitas Muhammadiyah Makassar sejak tahun 2015. Telah mengikuti International Training Program di India tahun 2017 dan Visiting Scholar Program di Beijing University Chemical Technology (BUCT), Beijing – China tahun 2019, serta aktif mengikuti kegiatan seminar nasional dan Internasional. Penerima Hibah skema Penelitian Dosen Pemula (PDP) dari Kementrian Riset, Tekhnologi dan Pendidikan Tinggi di tahun 2017 dan 2018, meneliti mengenai eksplorasi cendawan pada rhizosfer tanaman padi sebagai agens hayati.





















