
Cell Cycle-Specific and Cell Type-Specific Expression of Rb in the Developing Human Retina
9
Cell Cycle–Specific and Cell Type–Specific Expression of Rb in the Developing Human Retina Thomas C. Lee, 1,2,3 Dena Almeida, 1,2 Nidia Claros, 1,2 David H. Abramson, 2,4 and David Cobrinik 1,2 PURPOSE. To define the pattern of Rb expression relative to cell cycle position and cell type in the developing human retina. METHODS. Cryosections of fetal week 11-18 retinas were immu- nostained for Rb and cell cycle– or cell type–specific markers. RESULTS. Rb was prominent in retinal progenitor cells (RPCs) expressing the cyclin D1, cyclin A, and cytoplasmic cyclin B markers of G 1 , S, and early to mid G 2 phases, but not in RPCs expressing the phosphohistone H3 marker of late G 2 and M. Rb was not detected in the earliest postmitotic ganglion, ama- crine, horizontal, and bipolar cell precursors migrating away from the ventricular layer, but was detected as such cells underwent further differentiation. Among photoreceptors, Rb was not detected in the earliest RXR() cone precursors or in the earliest Nrl() rod precursors, but subsequently rose to high levels in cones and to low levels in rods. Rb was promi- nent at the time when Mu ¨ller glia exit the cell cycle and was generally expressed in a pattern complementary to p27 Kip1 . CONCLUSIONS. Rb exhibits cell cycle–specific expression in RPCs, with loss in late G 2 -M and restoration in G 1 . Rb is re-expressed after postmitotic ganglion, amacrine, horizontal, and bipolar cell precursors migrate away from the ventricular layer; after the appearance of early cone and rod markers; but coinciding with Mu ¨ller glia cell cycle withdrawal. The results suggest that Rb does not mediate the initial proliferative arrest of retinal neurons, but may indirectly induce arrest in RPCs or maintain an arrest in postmitotic precursors. (Invest Ophthal- mol Vis Sci. 2006;47:5590 –5598) DOI:10.1167/iovs.06-0063 R etinoblastomas generally arise within the first 4 years of life and have been detected prenatally as early as fetal week (Fwk) 21. 1,2 The retinoblastoma gene (RB1) and protein (Rb) are crucial to the suppression of retinoblastoma, as RB1 muta- tion, deletion, or silencing occurs in all such tumors, and germline mutations predispose to numerous retinoblastoma foci. 3 Although these observations indicate that Rb suppresses tumorigenesis during early retinal development, the retinal cell type and developmental stage in which Rb functions have not been defined. 4,5 The human retina begins its development at Fwk 5 and initially consists of proliferating retinal progenitor cells (RPCs) within a neuroblastic layer (NBL). 6 RPC nuclei undergo a cell cycle– dependent migration within the NBL, in which mitotic nuclei are positioned at the ventricular (outermost) layer, S phase nuclei are located in the middle NBL, and G 1 and G 2 nuclei migrate between these positions. 7,8 Mitotic RPCs may give rise to additional RPCs or to postmitotic precursors of each of the mature retinal cell types, with the fate influenced by cell-intrinsic competence states and by extrinsic cues. 9 Retinal development proceeds in a central to peripheral direc- tion, such that proliferation ceases in the central retina by Fwk 12 and in the far periphery by Fwk 30. 10,11 Histopathological analyses have provided clues to the cell type in which Rb may suppress tumorigenesis. Retinoblasto- mas often contain differentiated cells that express markers of cones and, to a lesser extent, markers of rods and Mu ¨ller glia, but not markers of other retinal cells. 12,13 Similarly, retinoblas- toma cell lines express proteins that are specific to cones and rods but not other cell types. 14,15 In tumors, cells that express glial markers adjoin and may form concurrent with cells that express photoreceptor markers. 12,13 These findings suggest that retinoblastomas may derive from a cell that is committed to photoreceptor and glial differentiation or from an uncom- mitted cell that is restricted to such differentiation in the tumor environment. Among mammals, retinoblastoma is almost exclusively a human disease, as it has been diagnosed in only two individual animals. 16,17 Moreover, in mice, loss of Rb does not predispose to retinal tumors, and combined loss of Rb and either of the related proteins p107 or p130 results in tumors with amacrine but not photoreceptor differentiation. 18 –21 In this study, we sought to define the spatiotemporal pattern of Rb expression in the developing human retina, as a means of identifying cell types and developmental stages in which Rb may suppress tumorigenesis. METHODS Human fetal eyes were obtained under protocols approved by the Weill Medical College IRB and were studied in compliance with the tenets of the Declaration of Helsinki. Fetal age was determined by femur length. The cornea and lens were removed and the eyes fixed overnight at 4°C in 4% paraformaldehyde in PBS (PFA/PBS), incubated in 30% sucrose/PBS overnight at 4°C, embedded in one part 30% sucrose/PBS and two parts optimal cutting temperature compound (OCT; Miles Laboratories, Elkhart, IN), frozen, and sectioned at 10 to 15 m. For Rb staining, sections were postfixed in 4% PFA/PBS for 5 minutes, washed in 0.5 M NaCl/20 mM Tris (pH 8.0; TBS), treated with 1 mM EDTA/TBS for 5 minutes, washed with TBS, incubated in 0.1% H 2 O 2 for 15 minutes, washed in TBS, treated with ABC kit reagent A (Vector Laboratories, Burlingame, CA) in TBS for 15 minutes, washed in TBS, treated with ABC kit reagent B (Vector Laboratories) in TBS for 15 minutes, washed in TBS, blocked and permeabilized in 5% horse From the 1 Margaret M. Dyson Vision Research Institute, and 2 De- partment of Ophthalmology, Weill Medical College of Cornell Univer- sity, New York, New York; and the 4 Ophthalmic Oncology Service, Department of Surgery, Memorial Sloan-Kettering Cancer Center, New York, New York. 3 Current address: Division of Ophthalmology, Department of Sur- gery, Childrens Hospital Los Angeles, Los Angeles, California. Supported by The C.V. Starr Foundation, The Fred Gluck Founda- tion, The New York Community Trust, and The Fund for Ophthalmic Knowledge. Submitted for publication January 20, 2006; revised August 1, 2006; accepted October 11, 2006. Disclosure: T.C. Lee, None; D. Almeida, None; N. Claros, None; D.H. Abramson, None; D. Cobrinik, None The publication costs of this article were defrayed in part by page charge payment. This article must therefore be marked “advertise- ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact. Corresponding author: David Cobrinik, Margaret M. Dyson Vision Research Institute, Weill Medical College of Cornell University, 1300 York Ave, LC303, New York, NY 10021; [email protected]. Investigative Ophthalmology & Visual Science, December 2006, Vol. 47, No. 12 5590 Copyright © Association for Research in Vision and Ophthalmology
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cell Cycle-Specific and Cell Type-Specific Expression of Rb in the Developing Human Retina

Cell Cycle–Specific and Cell Type–Specific Expression ofRb in the Developing Human Retina
Thomas C. Lee,1,2,3 Dena Almeida,1,2 Nidia Claros,1,2 David H. Abramson,2,4 andDavid Cobrinik1,2
PURPOSE. To define the pattern of Rb expression relative to cellcycle position and cell type in the developing human retina.
METHODS. Cryosections of fetal week 11-18 retinas were immu-nostained for Rb and cell cycle– or cell type–specific markers.
RESULTS. Rb was prominent in retinal progenitor cells (RPCs)expressing the cyclin D1, cyclin A, and cytoplasmic cyclin Bmarkers of G1, S, and early to mid G2 phases, but not in RPCsexpressing the phosphohistone H3 marker of late G2 and M. Rbwas not detected in the earliest postmitotic ganglion, ama-crine, horizontal, and bipolar cell precursors migrating awayfrom the ventricular layer, but was detected as such cellsunderwent further differentiation. Among photoreceptors, Rbwas not detected in the earliest RXR�(�) cone precursors or inthe earliest Nrl(�) rod precursors, but subsequently rose tohigh levels in cones and to low levels in rods. Rb was promi-nent at the time when Muller glia exit the cell cycle and wasgenerally expressed in a pattern complementary to p27Kip1.
CONCLUSIONS. Rb exhibits cell cycle–specific expression inRPCs, with loss in late G2-M and restoration in G1. Rb isre-expressed after postmitotic ganglion, amacrine, horizontal,and bipolar cell precursors migrate away from the ventricularlayer; after the appearance of early cone and rod markers; butcoinciding with Muller glia cell cycle withdrawal. The resultssuggest that Rb does not mediate the initial proliferative arrestof retinal neurons, but may indirectly induce arrest in RPCs ormaintain an arrest in postmitotic precursors. (Invest Ophthal-mol Vis Sci. 2006;47:5590–5598) DOI:10.1167/iovs.06-0063
Retinoblastomas generally arise within the first 4 years of lifeand have been detected prenatally as early as fetal week
(Fwk) 21.1,2 The retinoblastoma gene (RB1) and protein (Rb)are crucial to the suppression of retinoblastoma, as RB1 muta-tion, deletion, or silencing occurs in all such tumors, andgermline mutations predispose to numerous retinoblastomafoci.3 Although these observations indicate that Rb suppressestumorigenesis during early retinal development, the retinal cell
type and developmental stage in which Rb functions have notbeen defined.4,5
The human retina begins its development at Fwk 5 andinitially consists of proliferating retinal progenitor cells (RPCs)within a neuroblastic layer (NBL).6 RPC nuclei undergo a cellcycle–dependent migration within the NBL, in which mitoticnuclei are positioned at the ventricular (outermost) layer, Sphase nuclei are located in the middle NBL, and G1 and G2
nuclei migrate between these positions.7,8 Mitotic RPCs maygive rise to additional RPCs or to postmitotic precursors ofeach of the mature retinal cell types, with the fate influencedby cell-intrinsic competence states and by extrinsic cues.9
Retinal development proceeds in a central to peripheral direc-tion, such that proliferation ceases in the central retina by Fwk12 and in the far periphery by Fwk 30.10,11
Histopathological analyses have provided clues to the celltype in which Rb may suppress tumorigenesis. Retinoblasto-mas often contain differentiated cells that express markers ofcones and, to a lesser extent, markers of rods and Muller glia,but not markers of other retinal cells.12,13 Similarly, retinoblas-toma cell lines express proteins that are specific to cones androds but not other cell types.14,15 In tumors, cells that expressglial markers adjoin and may form concurrent with cells thatexpress photoreceptor markers.12,13 These findings suggestthat retinoblastomas may derive from a cell that is committedto photoreceptor and glial differentiation or from an uncom-mitted cell that is restricted to such differentiation in the tumorenvironment.
Among mammals, retinoblastoma is almost exclusively ahuman disease, as it has been diagnosed in only two individualanimals.16,17 Moreover, in mice, loss of Rb does not predisposeto retinal tumors, and combined loss of Rb and either of therelated proteins p107 or p130 results in tumors with amacrinebut not photoreceptor differentiation.18–21 In this study, wesought to define the spatiotemporal pattern of Rb expressionin the developing human retina, as a means of identifying celltypes and developmental stages in which Rb may suppresstumorigenesis.
METHODS
Human fetal eyes were obtained under protocols approved by theWeill Medical College IRB and were studied in compliance with thetenets of the Declaration of Helsinki. Fetal age was determined byfemur length. The cornea and lens were removed and the eyes fixedovernight at 4°C in 4% paraformaldehyde in PBS (PFA/PBS), incubatedin 30% sucrose/PBS overnight at 4°C, embedded in one part 30%sucrose/PBS and two parts optimal cutting temperature compound(OCT; Miles Laboratories, Elkhart, IN), frozen, and sectioned at 10 to15 �m.
For Rb staining, sections were postfixed in 4% PFA/PBS for 5minutes, washed in 0.5 M NaCl/20 mM Tris (pH 8.0; TBS), treated with1 mM EDTA/TBS for 5 minutes, washed with TBS, incubated in 0.1%H2O2 for 15 minutes, washed in TBS, treated with ABC kit reagent A(Vector Laboratories, Burlingame, CA) in TBS for 15 minutes, washedin TBS, treated with ABC kit reagent B (Vector Laboratories) in TBS for15 minutes, washed in TBS, blocked and permeabilized in 5% horse
From the 1Margaret M. Dyson Vision Research Institute, and 2De-partment of Ophthalmology, Weill Medical College of Cornell Univer-sity, New York, New York; and the 4Ophthalmic Oncology Service,Department of Surgery, Memorial Sloan-Kettering Cancer Center, NewYork, New York.
3Current address: Division of Ophthalmology, Department of Sur-gery, Childrens Hospital Los Angeles, Los Angeles, California.
Supported by The C.V. Starr Foundation, The Fred Gluck Founda-tion, The New York Community Trust, and The Fund for OphthalmicKnowledge.
Submitted for publication January 20, 2006; revised August 1,2006; accepted October 11, 2006.
Disclosure: T.C. Lee, None; D. Almeida, None; N. Claros, None;D.H. Abramson, None; D. Cobrinik, None
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: David Cobrinik, Margaret M. Dyson VisionResearch Institute, Weill Medical College of Cornell University, 1300York Ave, LC303, New York, NY 10021; [email protected].
Investigative Ophthalmology & Visual Science, December 2006, Vol. 47, No. 125590 Copyright © Association for Research in Vision and Ophthalmology

serum�2% human serum in TBS (block 1), with 0.1% Triton-X-100 for20 minutes, incubated in Rb antibody G3-245 (1:200; BD-PharMingen,San Diego, CA) in block 1�0.05% Tween-20 overnight at 4°C, washedin TBS, incubated in biotinylated horse anti-mouse antibody (1:135;Vector Laboratories) in block 1 for 30 minutes, washed in TBS, washedin 0.1 M sodium bicarbonate and 0.15 M NaCl (balanced saline),incubated with FITC-conjugated streptavidin (1:100; Vector Laborato-ries) in balanced saline, and washed with TBS. For costaining, sectionswere incubated in 5% goat serum and 2% human serum in TBS (block2) for 20 minutes, incubated overnight with primary antibody (Sup-plementary Information, online at http://www.iovs.org/cgi/content/full/47/12/5590/DC1) in block 2, washed in TBS, incubated for 30minutes in block 2 with secondary antibody (Supplementary Informa-tion online), and washed in TBS. Sections were stained with (4�,6�-diamino-2-phenylindole (DAPI) and analyzed by indirect immunofluo-rescence or confocal microscopy.
RESULTS
Distinct Rb Expression Patterns in Peripheral andCentral Retina
We initially evaluated Rb expression at Fwk 18. At this age,there is extensive proliferation in the peripheral retina, but noproliferation in the central retina.11 In the periphery, Rb wasprominent in nuclei throughout the middle NBL and in occa-sional nuclei in the outer NBL (Figs. 1A, 1B). A similar patternwas evident at Fwks 12 and 14, although the band of Rb(�)nuclei was positioned closer to the outer limiting membrane(OLM), due to the lack of rod precursors in the outer NBL atthese ages (Supplementary Fig. S1; all Supplementary Figuresare online at http://www.iovs.org/cgi/content/full/47/12/5590/DC1). In contrast, in the central retina, Rb was promi-nent in outer nuclear layer cells having the appearance ofcones and in inner nuclear layer (INL) positions typical ofMuller cell nuclei, but was detected at far lower levels else-where (Figs. 1C, 1D). Thus, the Rb expression pattern differedin the highly proliferating peripheral retina compared with thepostmitotic central retina and exhibited cell type–specific dif-ferences among postmitotic cells.
Rb Expression in RPCs
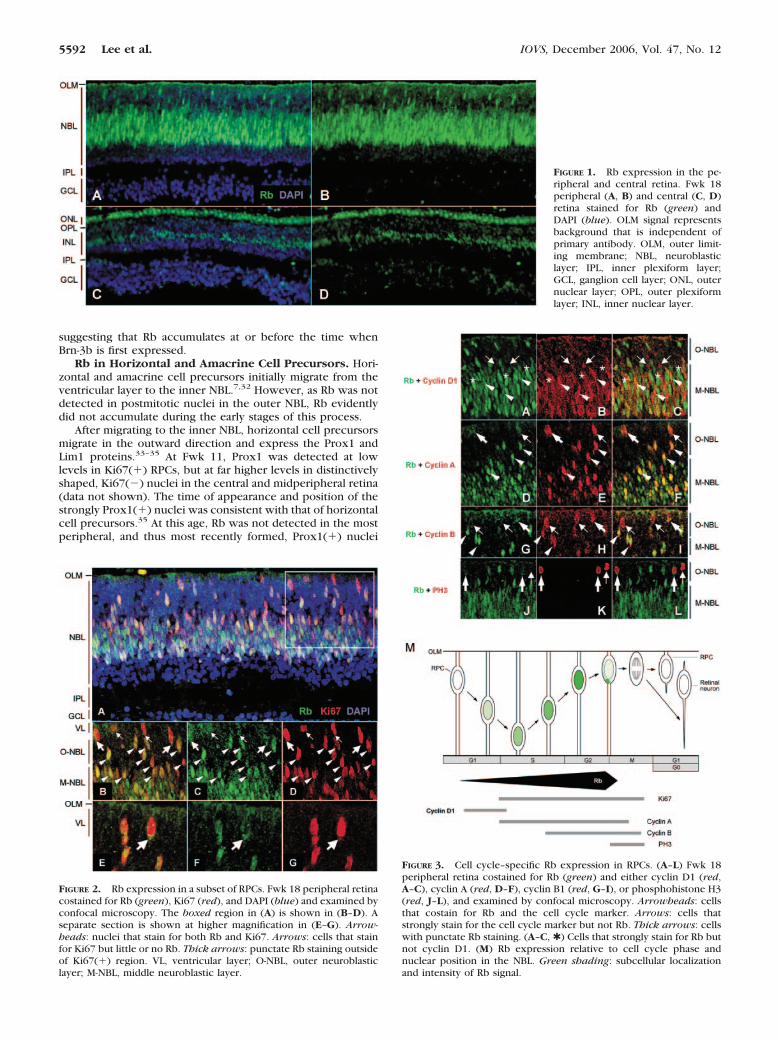
In the peripheral retina, Rb(�) nuclei were in positions thatare characteristic of RPCs (Figs. 1A, 1B). To determine whetherRb is expressed in such cells, we costained for Rb and theproliferation marker Ki67. Rb was prominent in all Ki67(�)nuclei in the middle and outer NBL (Figs. 2A–D, arrowheads),but was diminished or undetectable in Ki67(�) nuclei at theventricular layer (Fig. 2, arrows, and Supplementary Fig. S2online). Moreover, Rb had a punctate distribution outside ofthe Ki67(�) region in many ventricular layer cells (Figs. 2B–G;thick arrows).
The diminished Rb signal in ventricular layer RPCs suggeststhat Rb may be decreased in the G2- or M-phase nuclei that aretypically located in this position. To examine Rb in differentcell cycle phases, retinas were costained for Rb and a series ofcyclin proteins. Cyclin D1 was used as a marker of cells in G1,as it is expressed in G1 and degraded in S,22 and was notdetected in outer NBL positions typical of G2 and M.23,24 CyclinA was used as a marker of S, G2, and mitotic prophase,25 andcyclin B1 as a marker that accumulates in the cytoplasm in G2,enters the nucleus on entry into prophase, and is degraded inanaphase.26 Consistent with these assignments, cyclins D1 andA were detected in distinct cell populations (SupplementaryFig. S3 online).
As in other species, cyclin D1 was detected in the middleNBL (Fig. 3B). Rb was detected in nearly all cyclin D1(�)cells (Figs. 3A–C, arrowheads; Supplementary Fig. S2 on-
line), apart from rare cells at the interface of the outer andmiddle NBL (Figs. 3A–C, arrows). Notably, cyclin D1(�) cellsoften had weak Rb signals relative to the surrounding cells (Figs.3A–C, compare cells marked by arrowheads with those markedby asterisks).
Cells expressing cyclin A and cytoplasmic cyclin B weredetected in both the middle and outer NBL (Figs. 3E, 3H). Inthe middle NBL, nearly all cyclin A(�) and cytoplasmic cyclinB(�) cells had strong Rb signals (Figs. 3D–I, arrowheads; Sup-plementary Fig. S2 online). The higher Rb signal in cyclin A(�)and cytoplasmic cyclin B(�) cells, relative to cyclin D1(�)cells, suggests that Rb expression increases as RPCs progressfrom G1 into S and G2. In contrast, many cyclin A(�) and cyclinB(�) cells at the ventricular layer had little or no Rb (Figs.3D–I, arrows), although some cells had punctate Rb signals(Figs. 3D–I, thick arrows) similar to those in Ki67(�) cells insimilar positions (Fig. 2).
To define further the cell-cycle position in which Rbsignal declines, we costained for Rb and phosphorylatedhistone H3 (PH3), which is specific to late G2 and mitoticprophase, metaphase, and anaphase.27 PH3(�) cells hadeither a low, barely detectable nuclear Rb signal (notshown), a punctate Rb signal outside the PH-3(�) region(Figs. 3J–L, thick arrows), or no detectable Rb in the case ofanaphase cells (Figs. 3J–L, thin arrow). This observationsuggests that Rb levels decline at or before the time whenhistone H3 is phosphorylated in late G2.
After undergoing mitosis at the ventricular layer, early G1-phase RPC nuclei and postmitotic retinal precursor nucleimigrate through the outer NBL in the inward (vitread) direc-tion. However, all Rb(�) cells in the outer NBL expressedcyclin A (Figs. 3D–F and data not shown), which is specific toS, G2, and early M.25 The lack of Rb in cyclin A(�) cells impliesthat Rb was not appreciably expressed in early G0 or G1 nucleiduring their vitread migration.
In sum, Rb was present in cyclin D1(�) cells in the middleNBL, was detected at higher levels in cyclin A(�) and cytoplas-mic cyclin B(�) cells in the middle and outer NBL, and wasdiminished in cyclin A(�), cyclin B(�), and PH3(�) cells at theventricular layer (Fig. 3M). The results imply that Rb expres-sion increases during progression from G1 into S and G2,declines in late G2 and M coinciding with a punctate distribu-tion pattern, and does not reaccumulate until RPC nuclei re-turn to the middle NBL.
Rb Expression in Postmitotic Retinal Cells
After undergoing mitosis at the ventricular layer, RPCs give riseto additional RPCs or to postmitotic retinal precursors. Todefine the Rb expression pattern during the genesis of postmi-totic cells, we stained for Rb and early markers of the differentretinal cell types.
Rb in Ganglion Cell Precursors. Ganglion cells are pro-duced in the retinal far periphery at Fwk 12.11 Islet-1 is an earlymarker of such cells and is first expressed at the ventricularlayer soon after mitosis.28,29 Islet-1 was highly expressed inprecursors migrating through the NBL as well as in the incip-ient ganglion cell layer (GCL; Figs. 4A, 4B). Rb was not de-tected in migrating Islet-1(�) cells in the outer NBL (Figs.4A–C, arrows), but was prominent in Islet-1(�) cells at theinterface of the NBL and GCL (Figs. 4A–C, arrowheads). Rb wasalso detected at low levels in a subset of cells in the incipientGCL and in the more mature GCL at later ages (Figs. 1, 4),suggesting that Rb levels may decline during ganglion cellmaturation. To define further the timing of Rb expression, wecostained for Rb and Brn-3b, which is specific to postmitoticganglion cells.30,31 Rb was prominent in Brn-3b(�) cells at theinterface between the NBL and GCL (Figs. 4D–F, arrowheads),
IOVS, December 2006, Vol. 47, No. 12 Expression of Rb in the Developing Human Retina 5591
suggesting that Rb accumulates at or before the time whenBrn-3b is first expressed.
Rb in Horizontal and Amacrine Cell Precursors. Hori-zontal and amacrine cell precursors initially migrate from theventricular layer to the inner NBL.7,32 However, as Rb was notdetected in postmitotic nuclei in the outer NBL, Rb evidentlydid not accumulate during the early stages of this process.
After migrating to the inner NBL, horizontal cell precursorsmigrate in the outward direction and express the Prox1 andLim1 proteins.33–35 At Fwk 11, Prox1 was detected at lowlevels in Ki67(�) RPCs, but at far higher levels in distinctivelyshaped, Ki67(�) nuclei in the central and midperipheral retina(data not shown). The time of appearance and position of thestrongly Prox1(�) nuclei was consistent with that of horizontalcell precursors.35 At this age, Rb was not detected in the mostperipheral, and thus most recently formed, Prox1(�) nuclei
FIGURE 1. Rb expression in the pe-ripheral and central retina. Fwk 18peripheral (A, B) and central (C, D)retina stained for Rb (green) andDAPI (blue). OLM signal representsbackground that is independent ofprimary antibody. OLM, outer limit-ing membrane; NBL, neuroblasticlayer; IPL, inner plexiform layer;GCL, ganglion cell layer; ONL, outernuclear layer; OPL, outer plexiformlayer; INL, inner nuclear layer.
FIGURE 2. Rb expression in a subset of RPCs. Fwk 18 peripheral retinacostained for Rb (green), Ki67 (red), and DAPI (blue) and examined byconfocal microscopy. The boxed region in (A) is shown in (B–D). Aseparate section is shown at higher magnification in (E–G). Arrow-heads: nuclei that stain for both Rb and Ki67. Arrows: cells that stainfor Ki67 but little or no Rb. Thick arrows: punctate Rb staining outsideof Ki67(�) region. VL, ventricular layer; O-NBL, outer neuroblasticlayer; M-NBL, middle neuroblastic layer.
FIGURE 3. Cell cycle–specific Rb expression in RPCs. (A–L) Fwk 18peripheral retina costained for Rb (green) and either cyclin D1 (red,A–C), cyclin A (red, D–F), cyclin B1 (red, G–I), or phosphohistone H3(red, J–L), and examined by confocal microscopy. Arrowheads: cellsthat costain for Rb and the cell cycle marker. Arrows: cells thatstrongly stain for the cell cycle marker but not Rb. Thick arrows: cellswith punctate Rb staining. (A–C, ✱ ) Cells that strongly stain for Rb butnot cyclin D1. (M) Rb expression relative to cell cycle phase andnuclear position in the NBL. Green shading: subcellular localizationand intensity of Rb signal.
5592 Lee et al. IOVS, December 2006, Vol. 47, No. 12

(Figs. 5A–C), but was detected at low levels in occasionalProx1(�) cells in more central positions (Figs. 5D–F, arrow-heads). Similarly, at Fwk 18, Rb was not detected in the mostperipheral Prox1(�) horizontal cell precursors (Figs. 5G–I),but was highly expressed in the more mature horizontal cellsin the central retina (Figs. 5J–L). Rb expression in horizontalcell precursors was confirmed by costaining with Lim1 (Sup-plementary Fig. S4 online). The expression of Rb in the morecentral, but not the peripheral, Prox1(�) nuclei implies thatRb is expressed after the initial expression of Prox1 duringhorizontal cell maturation.
To date, we have not identified a marker that is specific topostmitotic amacrine precursors and compatible with Rb stain-ing (see Supplementary Information, http://www.iovs.org/cgi/content/full/47/12/5590/DC1). However, Rb was detected atlow levels in the inner INL where amacrine precursors accu-mulate (Fig. 1B), consistent with Rb’s having minimal expres-sion during amacrine cell maturation.
Rb in Cone Precursors. Rb was highly expressed in fovealcone precursors at Fwk 18 (Fig. 1C). To define the timing of Rbexpression in this lineage, we stained for Rb and several coneprecursor markers. One of the earliest cone markers in chicks
and mice is the retinoid X receptor � (RXR�).36–38 At Fwk 12,Rb was detected in only a subset of the RXR�(�) nuclei in thefovea (Figs. 6A–C, arrowheads), and in none of the youngerRXR�(�) nuclei at more peripheral positions (data not shown).At Fwk 15, Rb was detected in all the RXR�(�) nuclei in thecentral retina, but not in those in the periphery (Figs. 6D–I).Similarly, Rb was not detected in peripheral RXR�(�) nuclei atFwk 18 (data not shown). The increased prevalence of Rb inRXR�(�) cells in more developed retinal regions implies thatRb is expressed after RXR� appears.
Two other early cone markers are thyroid hormone recep-tor �2 (TR�2)39,40 and interphotoreceptor retinoid bindingprotein (IRBP), which is specific to cone precursors in theearly fovea.10 At Fwk 12, TR�2 and IRBP were detected only inthe fovea, and Rb was detected in all TR�2(�) and all IRBP(�)cells (Supplementary Fig. S5 online).
Rb in Rod Precursors. Rod precursors are initially posi-tioned below the outer limiting membrane. As additional rodnuclei accumulate, the earlier born rod nuclei are displaced toprogressively deeper layers.7,32 To evaluate Rb expression dur-ing rod development, we costained for Rb and Nrl, which isthe earliest known rod-specific protein.10,41 At Fwk 14, the
FIGURE 4. Rb expression in gan-glion cell precursors. Fwk 12 periph-eral retina costained for Rb (green)and either Islet-1 (red, A–C) orBrn-3b (red, D–F). Arrowheads: nu-clei that costain for Rb and eitherIslet-1 or Brn-3b. Arrows: nuclei thatstain for Islet-1 but not Rb. iGCL,incipient ganglion cell layer.
FIGURE 5. Rb expression in horizon-tal cell precursors. Fwk 11 peripheral(A–C) and central (D–F) retina orFwk 18 peripheral (G–I) and central(J–L) retina costained for Prox1 (red)and Rb (green). Arrows: nuclei thatstain for Prox1 but not Rb. Arrow-heads: nuclei that costain for Prox1and Rb. The positions of stronglyProx1(�) cells are consistent withthat of horizontal cell precursors atall ages and with that of a second,uncharacterized population in theFwk 18 INL.
IOVS, December 2006, Vol. 47, No. 12 Expression of Rb in the Developing Human Retina 5593

most peripheral (and hence, least mature) Nrl(�) nuclei werepositioned below the outer limiting membrane (Figs. 7A–C,arrows). Rb was not detected in these peripheral Nrl(�) nu-clei, but was detected in more mature Nrl(�) nuclei at morecentral and basal positions (Figs. 7A–C, arrowhead). This phe-nomenon was more clear in the Fwk 18 periphery, where Rbwas not evident in Nrl(�) nuclei near the outer limiting mem-brane, but was detected in more mature Nrl(�) nuclei at morebasal positions (Figs. 7D–G, arrowheads). Similarly, Rb wasdetected in Nrl(�) nuclei in the parafoveal region, albeit at farlower levels than in Nrl(�) cones (Supplementary Fig. S6online). Notably, Nrl was expressed at higher levels in older,basally positioned Rb(�) nuclei than in the younger, apicallypositioned nuclei that lacked Rb expression (Figs. 7D–G).
Rb in Bipolar Cell Precursors. Bipolar cells comprisemuch of the outer aspect of the INL and can be identified bytheir prominent nuclear expression of Chx10.42 Chx10 is alsoexpressed in RPCs, but at far lower levels and without thecentral-to-peripheral distribution of bipolar cell precursors.
At Fwk 15, the most peripheral, and thus youngest, stronglyChx10(�) bipolar cell nuclei were interspersed within the NBLand did not have detectable Rb (Figs. 8A–C). However, in thecentral retina, Chx10(�) nuclei formed a distinct layer, and inmost cases had moderate, above background Rb expression(Figs. 8D–F), indicating that Rb was first detected in bipolarcells after Chx10 accumulation.
Rb in Muller Glia. Rb was highly expressed in INL posi-tions that are characteristic of Muller glia nuclei at Fwk 18(Figs. 1C, 9F). This distribution was also evident at Fwk 15,although the pattern was less obvious due to higher Rb expres-sion in surrounding INL cells (Fig. 9C). Nonetheless, at Fwk 15each of the strongly Rb(�) INL nuclei was surrounded by theMuller cell–specific marker, cellular retinal–binding protein
(CRALBP)43 (Figs. 9A–C), confirming their Muller cell identity.Muller glia resemble RPCs in their morphology, gene ex-
pression profile, and ability to re-enter the cell cycle.44–46 Inkeeping with this relationship, Muller glia express cyclin D1for a limited time while they undergo cell cycle exit.46 Weconfirmed that cyclin D1 persisted in the Fwk 18 central retina,after proliferation had ceased (Fig. 9E and data not shown). Rbwas prominent in all cyclin D1(�) nuclei (Figs. 9D–F), indicat-ing that it is highly expressed as Muller glia exit the cell cycle.
Complementary Expression of Rb and p27Kip1
The lack of Rb in early postmitotic retinal neurons suggestedthat other cell cycle regulators mediate the proliferative arrestof such cells. Because p27Kip1 has an important antiprolifera-tive role in murine retina,23,24 we examined its expressionrelative to that of Rb.
In the Fwk 18 peripheral retina, p27 and Rb were generallyexpressed in a complementary pattern (Figs. 10A–C). A similarcomplementarity was evident at Fwk 14 (data not shown).RPCs comprising most of the the NBL prominently expressedRb but not p27, whereas rod precursors in the outer NBL andamacrine and ganglion cell precursors in the inner retina ex-pressed p27 but little or no Rb. Rare cells expressed both Rband p27 (Figs. 10A–C, arrowheads), particularly in outer NBLpositions where Rb and Nrl were coexpressed (see Figs. 7D–F),and in middle and inner NBL positions where Rb and Prox1were coexpressed (see Fig. 5). Thus, Rb may be superimposedover p27 expression during the differentiation of these cells.
Rb and p27 were also expressed in a complementary man-ner in the central retina. p27 was highly expressed and Rb hadminimal expression in positions typical of ganglion cells, am-acrine cells, horizontal cells, bipolar cells, and rods, whereas
FIGURE 6. Rb expression in coneprecursors. Fwk 12 (A–C) or Fwk 15(D–I) central (A–F) or peripheral(G–I) retina costained for Rb (green),RXR� (red), and DAPI (blue). Ar-rows: cells that stain for RXR� butnot Rb. Arrowheads: cells thatcostain for both RXR� and Rb.
FIGURE 7. Rb expression in rod pre-cursors. Fwk 14 midperipheral retina(A–C) and Fwk 18 peripheral retina(D–G) costained for Rb (green) andNrl (red). The boxed region in (D) isshown at higher magnification in (E–G). Arrows: cells that stain for Nrlbut not Rb. Arrowheads: cells thatcostain for Rb and Nrl. The direc-tions toward the retinal periphery orcenter are indicated for (A–C).
5594 Lee et al. IOVS, December 2006, Vol. 47, No. 12

little or no p27 was detected in the strongly Rb(�) Muller gliaand cones (Figs. 10G–I, arrows).
To assess the relationship between Rb and p27 expressionin earlier cones and Muller cells, we examined a transition zonehaving the most peripheral (and hence least mature) Rb(�)cone precursors and only rare Ki67(�) cells. In contrast to thecentral retina, Rb(�) cone precursors in this region had highlevels of p27 (Figs. 10D–F, arrowheads). This observation im-plies that cone precursors transition from a state in which theyexpress both Rb and p27 to a state in which they expresspredominantly Rb. However, we found no evidence of such atransition in Muller glia. Rather, the less mature glial cells in thetransition zone generally had no detectable p27 (Figs. 10D–F),whereas those in the central retina had low but detectable p27(Figs. 10G–I).
DISCUSSION
This study aimed to define the pattern of Rb expression in thehuman fetal retina, from gestational weeks 11 to 18. Thisinterval is characterized by RPC proliferation, production ofeach of the retinal cell types, and transition from a proliferativeto a postmitotic state. The data showed that Rb is expressed ina cell cycle–specific manner in RPCs and accumulates at vari-ous times and levels in the different types of postmitotic retinalprecursors.
Cell Cycle–Specific Expression of Rb in RPCs
Others had previously shown that Rb is expressed in RPCs inthe developing mouse and human retina.47–49 By using cyclinsand phosphorylated histone H3 (PH3) as markers, we deducedthat Rb is expressed in G1 nuclei in the middle NBL, increasesexpression in S and early G2 nuclei traversing the NBL, declinesto undetectable levels from late G2 to anaphase at the ventric-ular layer, and does not reaccumulate until G1 nuclei return tothe middle NBL (Fig. 3M). This cell cycle-specific pattern inhuman RPCs is consistent with the general decline in Rbexpression in M- and G1-phase RPCs in mice,47 and furtherindicates that Rb is both dramatically and consistently dimin-ished during RPC mitosis.
The cyclical expression of Rb in RPCs contrasts with Rb’sconstitutive expression throughout the cell cycle in diversecell types that were previously analyzed with the same anti-body.50 In those other cell types, Rb associated with condens-ing chromatin in prophase, redistributed to the cytoplasm onnuclear envelope breakdown in metaphase, and returned todaughter nuclei on nuclear envelope restoration in telo-phase.50 However, in RPCs, Rb declined before prophase(when PH3 becomes prominent27) and was not restored totelophase, or G1, nuclei until such nuclei had migrated to themiddle NBL.
The mechanism by which Rb expression declines in late G2
and M is unknown. However, it was notable that some cellsnear the ventricular layer had a punctate Rb signal immediatelyoutside of their Ki67(�) or PH3(�) regions (Figs. 2B–G). As Rbwas not detected in metaphase or later M-phase nuclei, the
FIGURE 8. Rb expression in bipolar cell precursors. Fwk 15 peripheral(A–C) or central (D–F) retina costained for Chx10 (red) and Rb(green). Arrows: nuclei that stain for Chx10 but not Rb. Arrowheads:nuclei that costain for Rb and Chx10.
FIGURE 9. Rb expression in Mullerglia. Fwk 15 (A–C) or Fwk 18 (D–F)fovea costained for Rb (green), andeither CRALBP (red, A–C) or cyclinD1 (red, D–F). Arrowheads: cellsthat costain for Rb and eitherCRALBP or cyclin D1.
IOVS, December 2006, Vol. 47, No. 12 Expression of Rb in the Developing Human Retina 5595

punctate Rb signal may designate sites to which Rb translo-cates before its degradation. Of interest, an earlier study ofosteosarcoma cells showed that high Cdk activity can inducephosphorylation of Rb residue 567 and may thereby induce Rbcytoplasmic translocation and degradation.51 Thus, the dramat-ically increased Cdk activity at the G2–M transition may elicitcytoplasmic translocation and degradation of most of the Rb inRPCs, even though it apparently does not do so in other celltypes.
It seems plausible that Rb’s cell cycle–specific expressionserves a developmental role. However, the cyclical loss of Rb inthe M phase seems unlikely to regulate RPC fate, as Rb waslacking in M-phase cells both in the periphery (where RPCsmainly produce more RPCs) and in more central positions(where RPCs mainly produce postmitotic precursors). Alterna-tively, the decline in Rb expression before mitosis may pre-clude Rb from inhibiting proliferation early in the subsequentcell cycle. Our detection of occasional cyclin D1(�), Rb(�)cells in the outer NBL (Figs. 3A–C) suggests that Rb may bere-expressed only after RPCs acquire cyclin D1-dependent ki-nase activity that can suppress Rb function.
Delayed Expression of Rb in PostmitoticRetinal Precursors
Our analyses show that Rb is expressed in all cell types in thedeveloping human retina. This result is unsurprising in light ofRb’s widespread expression and diverse functions in manydevelopmental contexts.52 Moreover, Rb has been detected incell types other than amacrine cells in the early mouse retina.48
However, the present study also defined the timing of Rbexpression relative to that of cell type–specific markers and
p27 and thereby provides novel insights into the potential roleof Rb in retinal development.
Unexpectedly, Rb was not detected at the time when post-mitotic retinal neurons were born, but accumulated well afterthe terminal mitosis. For example, Rb was not detected inpostmitotic ganglion, horizontal, amacrine, and bipolar cellprecursors migrating away from the ventricular layer, but wasdetected at the time when ganglion cells first expressed Brn-3band after horizontal cells first expressed Prox1. Similarly, Rbwas not detected in nascent Chx10(�) bipolar precursors, butwas detected in more mature Chx10(�) cells. This evidencesuggests that Rb does not directly mediate cell cycle with-drawal or early differentiation events in postmitotic retinalprecursors.
The timing of Rb expression in developing photoreceptorswas of particular interest, given that retinoblastoma may derivefrom a photoreceptor-directed cell. Surprisingly, Rb was notdetected at the time when rod precursors first expressed Nrl inthe outer retinal layers, but was detected in more mature rodnuclei at more basal positions. This suggests that in rod pre-cursors, Rb mediates neither cell cycle withdrawal nor theinitial expression of Nrl. That Rb does not mediate a rodprecursor arrest is consistent with the lack of rod precursorproliferation in Rb-deficient mouse retinas.49 Nevertheless, thelack of Rb in early Nrl(�) precursors was unexpected, giventhat Rb-deficient mouse retinas had impaired rod differentia-tion and decreased Nrl mRNA.49 Our finding that Nrl was morehighly expressed in Rb(�) versus Rb(�) cells (Figs. 7E–G)suggests that Rb may promote Nrl expression during late butnot early rod differentiation.
As in rods, Rb was not detected in the earliest cones.Whereas cone precursors are evident at Fwk 9,10 Rb was
FIGURE 10. Complementary expression of Rb and p27Kip1. Fwk 18 retina costained for Rb (green) and p27Kip1 (red). Images of peripheral (A–C),transition zone (D–F), and central (G–I) retina are from the same section and were imaged in identical fashion. Arrowheads: nuclei that costainfor Rb and p27. Arrows: nuclei that stain strongly for Rb but not p27.
5596 Lee et al. IOVS, December 2006, Vol. 47, No. 12

detected in only a subset of such precursors in the centralretina at Fwk 12 and was detected in central but not midpe-ripheral cones at Fwks 15 and 18. Although we cannot deter-mine the precise length of the delay, the evidence suggests thatfoveal cones may arrest for 1 to 3 weeks before having detect-able Rb expression. In contrast to rods, Rb rose to high levelsin early cones. Spencer et al.48 similarly detected high Rbexpression in adult cones. We extend their finding by showingthat cone precursors prominently express Rb in the fetal pe-riod when retinoblastoma tumorigenesis begins.
In contrast to the delayed Rb expression in postmitoticretinal neurons, Rb was highly expressed in Muller glia at thetime of cell cycle exit. As Muller glia resemble quiescentRPCs,44–46 Rb’s expression may be governed by the sameprocess in the two cell types. Notably, p27 was lacking at thetime when Muller glia exit the cell cycle. This is consistentwith the lack of p27 in RPCs, but contrasts with the prominentexpression of p27 in postmitotic retinal neurons.
Relationship of Rb Expressionto Tumor Suppression
In mice, Rb was found to be highly expressed in each of thetissues in which it has crucial developmental roles.53,54 Thus,although a protein’s expression level does not necessarily re-flect its function, it is of interest to consider whether Rb mightsuppress retinoblastoma in the retinal cell types in which it isexpressed at high levels.
Rb was most highly expressed in RPCs. However, Rb seemsunlikely to directly induce a proliferative arrest in such cells, asRPCs are not known to exit the cell cycle without differenti-ating into one of the postmitotic retinal cell types. Moreover,our data argue that Rb does not accumulate in RPCs in prepa-ration for a role after such cells divide, because Rb expressiondeclined in late G2/M and was not rapidly restored to postmi-totic cells. Thus, if Rb were to suppress tumorigenesis throughits expression in RPCs, it would appear to do so by indirectlyeliciting cell cycle exit and differentiation after a subsequentmitosis. Notably, in cortical neurogenesis, progenitor cells un-dergoing neurogenic divisions have longer cell cycles thanthose undergoing proliferative divisions, and artificially length-ening G1 suffices to induce neurogenic differentiation after anintervening mitosis.55,56 Thus, Rb might indirectly promoteneurogenic differentiation by lengthening G1 or through othereffects that precede Rb’s degradation in late G2 and M.
Rb was also highly expressed at the time when Muller gliaexit the cell cycle. Because p27 was not concurrently ex-pressed, it is tempting to infer that Rb may be needed tosuppress Muller cell proliferation. However, arguing againstthis possibility are the lack of Muller cell markers in prolifer-ating retinoblastoma cells12,13 and evidence that adult Mullerglia express high levels of the Rb-related p107 and p130.48
In cone precursors, Rb rose to high levels concurrent witha decline in p27. This reciprocal increase in Rb and loss of p27was unique to cone precursors at the ages that we examined.Notably, Rb is prominent in cone precursors and mature conesin the human retina, but in neither developing nor adult conesin mice (this study and Refs. 21,48). Moreover, Rb’s expressionin human cone precursors correlates with its role as a retino-blastoma suppressor in humans and with the predominantcone phenotype of differentiated retinoblastoma cells.12,13
These observations suggest that Rb may have a human-specificfunction in cone precursors, that coincides with the decline inp27. Nevertheless, additional studies are needed to addresswhether Rb suppresses retinoblastoma in cone precursors, inother cells that have high Rb levels such as RPCs or Muller glia,or in still other cell types that had less prominent Rb expres-sion.
Acknowledgments
The authors thank John Saari, Thomas Jessel, Barbara Wiggert, HemantKhanna, Anand Swaroop, Lily Ng, and Douglas Forrest for gifts ofantibodies used in this study.
References
1. Abramson DH, Schefler AC. Update on retinoblastoma. Retina.2004;24:828–848.
2. Maat-Kievit JA, Oepkes D, Hartwig NG, Vermeij-Keers C, van KampIL, van de Kamp JJ. A large retinoblastoma detected in a fetus at 21weeks of gestation. Prenat Diagn. 1993;13:377–384.
3. Lohmann DR, Gallie BL. Retinoblastoma: revisiting the model pro-totype of inherited cancer. Am J Med Genet C Semin Med Genet.2004;129:23–28.
4. Dyer MA, Bremner R. The search for the retinoblastoma cell oforigin. Nat Rev Cancer. 2005;5:91–101.
5. Cobrinik D. Pocket proteins and cell cycle control. Oncogene.2005;24:2796–2809.
6. Barishak Y. Embryology of the Eye and Its Adnexa. New York:Karger; 2001.
7. Sidman RL. Histogenesis of mouse retina studied with thymidine-H3. In: Smelser G, ed. The Structure of the Eye. New York:Academic Press;1961:487–505.
8. Donovan SL, Dyer MA. Regulation of proliferation during centralnervous system development. Semin Cell Dev Biol. 2005;16:407–421.
9. Livesey FJ, Cepko CL. Vertebrate neural cell-fate determination:lessons from the retina. Nat Rev Neurosci. 2001;2:109–118.
10. O’Brien KM, Schulte D, Hendrickson AE. Expression of photore-ceptor-associated molecules during human fetal eye development.Mol Vis. 2003;9:401–409.
11. Provis JM, van Driel D, Billson FA, Russell P. Development of thehuman retina: patterns of cell distribution and redistribution in theganglion cell layer. J Comp Neurol. 1985;233:429–451.
12. Gonzalez-Fernandez F, Lopes MB, Garcia-Fernandez JM, et al. Ex-pression of developmentally defined retinal phenotypes in thehistogenesis of retinoblastoma. Am J Pathol. 1992;141:363–375.
13. Nork TM, Schwartz TL, Doshi HM, Millecchia LL. Retinoblastoma:cell of origin. Arch Ophthalmol. 1995;113:791–802.
14. Bogenmann E, Lochrie MA, Simon MI. Cone cell-specific genesexpressed in retinoblastoma. Science. 1988;240:76–78.
15. Di Polo A, Farber DB. Rod photoreceptor-specific gene expressionin human retinoblastoma cells. Proc Natl Acad Sci USA. 1995;92:4016–4020.
16. Fugaro MN, Kiupel M, Montiani-Ferreira F, Hawkins JF, JanovitzEB. Retinoblastoma in the eye of a llama (Llama glama). Vet Oph-thalmol. 2005;8:287–290.
17. Syed NA, Nork TM, Poulsen GL, Riis RC, George C, Albert DM.Retinoblastoma in a dog. Arch Ophthalmol. 1997;115:758–763.
18. Robanus-Maandag E, Dekker M, van der Valk M, et al. p107 is asuppressor of retinoblastoma development in pRb-deficient mice.Genes Dev. 1998;12:1599–1609.
19. MacPherson D, Sage J, Kim T, Ho D, McLaughlin ME, Jacks T. Celltype-specific effects of Rb deletion in the murine retina. GenesDev. 2004;18:1681–1694.
20. Zhang J, Schweers B, Dyer MA. The First Knockout Mouse Modelof Retinoblastoma. Cell Cycle. 2004;3:952–959.
21. Chen D, Livne-bar I, Vanderluit JL, Slack RS, Agochiya M, BremnerR. Cell-specific effects of RB or RB/p107 loss on retinal develop-ment implicate an intrinsically death-resistant cell-of-origin in ret-inoblastoma. Cancer Cell. 2004;5:539–551.
22. Guo Y, Yang K, Harwalkar J, et al. Phosphorylation of cyclin D1 atThr 286 during S phase leads to its proteasomal degradation andallows efficient DNA synthesis. Oncogene. 2005;24:2599–2612.
23. Dyer MA, Cepko CL. p27Kip1 and p57Kip2 regulate proliferationin distinct retinal progenitor cell populations. J Neurosci. 2001;21:4259–4271.
24. Levine EM, Close J, Fero M, Ostrovsky A, Reh TA. p27(Kip1)regulates cell cycle withdrawal of late multipotent progenitor cellsin the mammalian retina. Dev Biol. 2000;219:299–314.
IOVS, December 2006, Vol. 47, No. 12 Expression of Rb in the Developing Human Retina 5597

25. Fung TK, Poon RY. A roller coaster ride with the mitotic cyclins.Semin Cell Dev Biol. 2005;16:335–342.
26. Pines J, Hunter T. Human cyclins A and B1 are differentiallylocated in the cell and undergo cell cycle-dependent nuclear trans-port. J Cell Biol. 1991;115:1–17.
27. Hendzel MJ, Wei Y, Mancini MA, et al. Mitosis-specific phosphor-ylation of histone H3 initiates primarily within pericentromericheterochromatin during G2 and spreads in an ordered fashioncoincident with mitotic chromosome condensation. Chromo-soma. 1997;106:348–360.
28. Rachel RA, Dolen G, Hayes NL, et al. Spatiotemporal features ofearly neuronogenesis differ in wild-type and albino mouse retina.J Neurosci. 2002;22:4249–4263.
29. Galli-Resta L, Resta G, Tan SS, Reese BE. Mosaics of islet-1-express-ing amacrine cells assembled by short-range cellular interactions.J Neurosci. 1997;17:7831–7838.
30. Xiang M. Requirement for Brn-3b in early differentiation of post-mitotic retinal ganglion cell precursors. Dev Biol. 1998;197:155–169.
31. Xiang M, Zhou L, Peng YW, Eddy RL, Shows TB, Nathans J. Brn-3b:a POU domain gene expressed in a subset of retinal ganglion cells.Neuron. 1993;11:689–701.
32. Hinds JW, Hinds PL. Differentiation of photoreceptors and hori-zontal cells in the embryonic mouse retina: an electron micro-scopic, serial section analysis. J Comp Neurol. 1979;187:495–511.
33. Liu W, Wang JH, Xiang M. Specific expression of the LIM/home-odomain protein Lim-1 in horizontal cells during retinogenesis.Dev Dyn. 2000;217:320–325.
34. Edqvist PH, Hallbook F. Newborn horizontal cells migrate bi-directionally across the neuroepithelium during retinal develop-ment. Development. 2004;131:1343–1351.
35. Dyer MA, Livesey FJ, Cepko CL, Oliver G. Prox1 function controlsprogenitor cell proliferation and horizontal cell genesis in themammalian retina. Nat Genet. 2003;34:53–58.
36. Hoover F, Seleiro EA, Kielland A, Brickell PM, Glover JC. RetinoidX receptor gamma gene transcripts are expressed by a subset ofearly generated retinal cells and eventually restricted to photore-ceptors. J Comp Neurol. 1998;391:204–213.
37. Mori M, Ghyselinck NB, Chambon P, Mark M. Systematic immu-nolocalization of retinoid receptors in developing and adult mouseeyes. Invest Ophthalmol Vis Sci. 2001;42:1312–1318.
38. Roberts MR, Hendrickson A, McGuire CR, Reh TA. Retinoid Xreceptor gamma is necessary to establish the S-opsin gradient incone photoreceptors of the developing mouse retina. Invest Oph-thalmol Vis Sci. 2005;46:2897–2904.
39. Ng L, Hurley JB, Dierks B, et al. A thyroid hormone receptor thatis required for the development of green cone photoreceptors.Nat Genet. 2001;27:94–98.
40. Sjoberg M, Vennstrom B, Forrest D. Thyroid hormone receptors inchick retinal development: differential expression of mRNAs for
alpha and N-terminal variant beta receptors. Development. 1992;114:39–47.
41. Swain PK, Hicks D, Mears AJ, et al. Multiple phosphorylatedisoforms of NRL are expressed in rod photoreceptors. J Biol Chem.2001;276:36824–36830.
42. Liu IS, Chen JD, Ploder L, et al. Developmental expression of anovel murine homeobox gene (Chx10): evidence for roles indetermination of the neuroretina and inner nuclear layer. Neuron.1994;13:377–393.
43. Bunt-Milam AH, Saari JC. Immunocytochemical localization of tworetinoid-binding proteins in vertebrate retina. J Cell Biol. 1983;97:703–712.
44. Close JL, Gumuscu B, Reh TA. Retinal neurons regulate prolifera-tion of postnatal progenitors and Muller glia in the rat retina viaTGF beta signaling. Development. 2005;132:3015–3026.
45. Walcott JC, Provis JM. Muller cells express the neuronal progenitorcell marker nestin in both differentiated and undifferentiated hu-man foetal retina. Clin Exp Ophthalmol. 2003;31:246–249.
46. Blackshaw S, Harpavat S, Trimarchi J, et al. Genomic analysis ofmouse retinal development. PLoS Biol. 2004;2:E247.
47. Donovan S, Schweers B, Martins R, Johnson D, Dyer MA. Compen-sation by tumor suppressor genes during retinal development inmice and humans. BMC Biol. 2006;4:14.
48. Spencer C, Pajovic S, Devlin H, Dinh QD, Corson TW, Gallie BL.Distinct patterns of expression of the RB gene family in mouse andhuman retina. Gene Expr Patterns. 2005;5:687–694.
49. Zhang J, Gray J, Wu L, et al. Rb regulates proliferation and rodphotoreceptor development in the mouse retina. Nat Genet. 2004;36:351–360.
50. Szekely L, Uzvolgyi E, Jiang WQ, et al. Subcellular localization ofthe retinoblastoma protein. Cell Growth Differ. 1991;2:287–295.
51. Ma D, Zhou P, Harbour JW. Distinct mechanisms for regulating thetumor suppressor and antiapoptotic functions of Rb. J Biol Chem.2003;278:19358–19366.
52. Liu H, Dibling B, Spike B, Dirlam A, Macleod K. New roles for theRB tumor suppressor protein. Curr Opin Genet Dev. 2004;14:55–64.
53. Jiang Z, Zacksenhaus E, Gallie BL, Phillips RA. The retinoblastomagene family is differentially expressed during embryogenesis. On-cogene. 1997;14:1789–1797.
54. Classon M, Harlow E. The retinoblastoma tumour suppressor indevelopment and cancer. Nat Rev Cancer. 2002;2:910–917.
55. Calegari F, Huttner WB. An inhibition of cyclin-dependent kinasesthat lengthens, but does not arrest, neuroepithelial cell cycleinduces premature neurogenesis. J Cell Sci. 2003;116:4947–4955.
56. Calegari F, Haubensak W, Haffner C, Huttner WB. Selective length-ening of the cell cycle in the neurogenic subpopulation of neuralprogenitor cells during mouse brain development. J Neurosci.2005;25:6533–6538.
5598 Lee et al. IOVS, December 2006, Vol. 47, No. 12




















