
The role of the ICOS-B7h T cell costimulatory pathway in transplantation immunity
Upload
independentCategory
view
1download
0
CD28/CTLA-4 and CD80/CD86 CostimulatoryMolecules are Mainly Involved in Acceptanceor Rejection of Human Liver TransplantAlfredo Minguela, Luis Marın, Alberto Torıo,Manuel Muro, Ana M. Garcıa-Alonso,Marıa R. Moya-Quiles, Francisco Sanchez-Bueno,Pascual Parrilla, and Marıa R. Alvarez-Lopez
ABSTRACT: CD28/CTLA-4 interactions with their spe-cific B7-ligands (CD80 and CD86) have decisive roles inantigenic and allogenic responses. Recently, experimentaltransplant studies demonstrated that donor-specific toler-ance is achieved by blocking these interactions. Thepresent study analyzes the expression of these co-stimu-latory molecules in peripheral blood cells from 74 liverrecipients and in 16 liver biopsies, which were classifiedinto acute-rejection (AR, n 5 27) and nonacute-rejection(NAR, n 5 47) groups, as well as their influence on thein vitro response of in vivo allosensitized cells. The resultsclearly indicate that in human liver transplant too, B7 andCD28/CTLA-4 expression on B and CD41 peripherallymphocytes respectively, contributes to graft acceptanceor rejection, and appears to be of crucial importance inmodulating the host alloresponse and specific-CTL gen-eration. In the NAR-group, costimulatory molecule ex-
pression remained at basal levels after transplant, whereasin the AR-group these molecules were significantly up-regulated on days of AR. CTLA-4 was observed in theinfiltrating lymphocytes in most of the biopsies, butCD80 or CD86 were not. Moreover, specific cytotoxicityfrom the in vivo primed cells was clearly suppressed in theNAR-patients with low co-stimulatory molecule expres-sion, whereas this activity was not modified but ratherstimulated in the AR-group. Together, these findingsindicate that intervention of CD28/CTLA-4/B7 signalingcould be therapeutically useful in clinical transplantation.Human Immunology 61, 658–669 (2000). © AmericanSociety for Histocompatibility and Immunogenetics,2000. Published by Elsevier Science Inc.
KEYWORDS: acute rejection; tolerance; citotoxicity;flow cytometry; T and B lymphocytes
ABBREVIATIONSAPC antigen presenting cellmAb monoclonal antibodiesAR acute rejectionMFI mean fluorescence intensityBD Becton DickinsonMLC mixed lymphocyte culturesCsA cyclosporine-ANAR non-acute-rejection
CTL cytotoxic T lymphocytesOLT orthotopic liver transplantDMSO dimethyl sulphoxidePBL peripheral blood lymphocytesFITC fluorescein isothiocyanatePBMC peripheral blood mononuclear cellsIL interleukinTCR T-cell receptor
INTRODUCTIONLiver transplant shows special tolerogenic behavior indifferent species [1], although in humans this topic has
not yet been fully explored. Leon et al. reported a lowexpression of co-stimulatory molecules in potential APCfrom liver grafts (epithelial and Kuppfer cells) [2], whichmight lead to the induction of a specific anergic state inrecipient Th lymphocytes due to a nonefficient antigenicpresentation [3–5]. In animal models, graft acceptancewas facilitated by blockade of CD28/CTLA-4 (CD152)interactions with their specific B7-ligands (CD80 andCD86) when using CTLA-4Ig or anti-B7 mAb [6–8].Moreover, a previous report demonstrated that CD28
From the Section of Immunology, (A.M., L.M., A.T., M.M., A.M.,G-A., M.R.M-Q., M.R.A-L.), and the Surgical Service (F.S-B., P.P.),University Hospital Virgen-Arrixaca, Murcia, Spain.
Address reprint requests to: M. R. Alvarez-Lopez, Seccion de Inmunolo-gıa, Hospital U. Virgen de la Arrixaca, El Palmar, 30120-Murcia, Spain;Fax: (134) 968-369678; E-Mail: [email protected].
Received June 21, 1999; revised January 21, 2000; accepted February11, 2000.
Human Immunology 61, 658–669 (2000)0198-8859/00/$–see front matter© American Society for Histocompatibility and Immunogenetics, 2000
Published by Elsevier Science Inc. PII S0198-8859(00)00113-0

might have a role in graft rejection or acceptance inhuman liver transplants [9].
Preliminary transplant studies indicated that directalloantigen recognition is the most efficient way to trig-ger an alloresponse [10], but it is currently accepted thatthe indirect presentation of alloantigens can also be in-volved. In heart [11] and liver transplants [12] thispathway constitutes an early event that precedes acuterejection episodes, suggesting that rejection is triggeredin the peripheral lymphoid tissues by autologous APC.
The existence of in vivo tolerized Th lymphocytes inassociation with self-antigens or as a result of experimen-tally-induced tolerance with heterologous antigens [13],suggests that a physiologic mechanism exists by whichsome population of normal APC can present antigens toTh lymphocytes in a tolerogenic manner [14]. The dif-ferent stimulatory ability of APC not only depends ontheir cognitive signals, but also on the presence of co-stimulatory molecules [15, 16]. Thus, dendritic cells,macrophages, and activated B lymphocytes with optimalexpression of CD80 and CD86 are efficient APC [17],whereas B7-negative resting B cells and T lymphocytesacting as nonefficient APC could induce anergy [18]. Infact, previous results by our group suggested that theexpansion of circulating B lymphocytes observed afterliver transplant can influence graft acceptance [19].
The present study investigated changes in the expres-sion of CD28/CTLA-4 and B7 costimulatory moleculesin PBMC from liver recipients, and their possible func-tional roles in the induction of allospecific effector cellsthat control liver graft rejection or acceptance.
MATERIALS AND METHODSPatientsA total of 88 orthotopic liver transplants (OLT) per-formed over a period of 2 years were studied. Afterexcluding patients who died or lost the graft within thefirst month after transplant due to causes other thanacute rejection (n 5 13), and those showing the firstacute rejection episode beyond the monitored period(n 5 1), the remaining 74 recipients, with differentdiagnoses (27 alcoholic cirrhosis, 19 hepatitis B or Cvirus cirrhosis, 10 carcinomas, 7 primary biliary cirrho-sis, 7 retransplants, and 4 amyloidosis), were divided intotwo groups: acute-rejection (AR-group, n 5 27) andnonacute rejection (NAR-group, n 5 47).
Immunosuppression consisted of standard triple-drugtherapy with methylprednisolone, azathioprine, andCsA. Methylprednisolone (1 g) was administered intra-venously during the intraoperative period and then dailyafter the transplant, with a gradual reduction from 200to 20 mg/day by postoperative day 7; Azathioprine at1–2 mg/kg per day was also administered intravenously
until it was tolerated orally, with a dose reduction whenleukocyte counts fell below 5000 cells/ml; two intrave-nous doses of conventional-CsA at 2 mg/kg per day ororal doses of 5–10 mg/kg per day were given to the first53 recipients (AR-group, n 5 16 and NAR-group, n 537), and Neoral-CsA was administered to the last 21recipients (AR-group, n 5 11 and NAR-group, n 5 10)who received two doses of 10–15 mg/kg per day througha jejunal tube. In both types of treatment, CsA bloodlevels were maintained between 250–350 ng/ml, andadditional individual regimens with a 1 g bolus of meth-ylprednisolone were used in case of acute rejection epi-sodes.
Histologic diagnosis of acute rejection was based onthe presence of at least two of the following features: (1)mononuclear, as well as mixed portal inflammation; (2)bile duct infiltration and damage; and (3) venous endo-thelitis of the portal tracts [9, 20]. Doppler echographywas performed to exclude hepatic ischemia due to hepaticartery or portal vein occlusion, and to establish an indi-cation for liver biopsy. Informed consent was obtainedfrom all recipients prior to experimental procedures.
Flow CytometryCytometric analysis was performed on EDTA-anticoag-ulated peripheral blood samples that were routinely col-lected from overnight fasting patients (between 0700and 0800 h) during the preoperative (day 0) and post-operative periods: days 1 to 3, 4 to 6, 7 to 9, 10 to 13,14 to 17, 18 to 21, 22 to 25, and after day 25. Aftercollection, peripheral blood cells were immediatelystained by a standard immunofluorescence method withdifferent mAb, acquiring 83103 lymphocytes in aFACScan flow cytometer (Becton Dickinson [BD], SanJose, CA, USA), by setting up a light scatter gate usingan anti-CD45-PerCP/CD14-PE/HLA-I-FITC mAb com-bination. Mean fluorescence intensity (MFI), used as arelative molecule-density measurement of CD80-CD86on CD191 cells or monocytes and of CD28 on CD41
lymphocytes, was calculated by the PAINT-A-GATE-plus software (BD). The photoelectric components of theFACScan flow cytometer were adjusted daily, as de-scribed previously [9].
The absolute number of CD41CD281, CD41
CD281CTLA-41, CD191CD801, and CD191CD861
cells was calculated from the total number of lympho-cytes obtained by routine leukocyte count (CoulterT-540, Northwell Drive, UK), together with their esti-mated cytometric percentage values.
Fluorescent mAb, anti-CD45 (Hle-1, IgG1-PerCP;BD), anti-CD14 (Leu-M3, IgG2b-Phycoerythrin; BD),anti-HLA class-I (HLA-ABC, IgG2a-FICT; Serotec, Ox-ford, UK), anti-CD4 (Leu3a, IgG1-PerCP; BD), anti-CD19 (SJ25-C1, IgG1-TC; Caltag, San Francisco, CA,
659CD28/CTLA-4 and B7 Molecules in Human Liver Transplant

USA), anti-CD28 (CD28.2, IgG1-FITC; Immunotech,Marseille, France), anti-CD80 (BB1, IgM-FITC; Pharm-ingen, San Diego, CA, USA), anti-CD86 (IT2.2, IgG2b-Phycoerythrin; Pharmingen) and anti-CTLA-4 (BNI3.1,IgG2a-Phycoerythrin; Pharmingen) were used at saturat-ing concentration. Simultest IgG1-FITC/IgG2a-Phyco-erythrin (BD) and IgG1-TRICOLOR (Caltag) were alsoused to evaluate background fluorescence.
PBMC cultures were analyzed, after a 6-day incuba-tion, using flow cytometry by washing them in FACSbuffer [0.1% BSA and 0.01% sodium azide in PBS; BD],and staining them with fluorescent mAb for 30 min at4°C. After a final wash in FACS buffer, cells were im-mediately acquired. The MFI of each molecule was ex-pressed as the median value of triplicate assays.
Isolation of PBMC and SplenocytesHeparinized peripheral blood samples were obtainedfrom healthy controls and from conventional-CsA treatedrecipients pretransplant (day 0), and days 7 and 14post-transplant. Splenic samples were procured from thecorresponding liver donors. PBMC and splenocytes wereisolated by Ficoll-Hypaque (Nicomed Pharma, Oslo,Norway), washed twice, and resuspended in RPMI 1640medium (Life Technologies, Paisley, Scotland) supple-mented with 2 mM L-glutamine (Life Technologies), 50mg/ml streptomycin, 50 IU/ml penicillin (Flow Labora-tories, Irvine, UK), and 10% human AB-serum (Sigma,St. Louis, MO, USA; complete medium). PBMC andsplenocytes were cryopreserved until assay in 10%DMSO (Merck, Darmstadt, Germany) in liquid nitro-gen.
MLC and Generation of Cytotoxic Effector CellsPBMC from three patients of the AR-group with post-transplant increased levels of CD28, CD80, and CD86,and from three patients from the NAR-group with post-transplant values of these molecules similar or lower tothose observed before transplant, were used for cultures.In addition, the presence of similar HLA class-I (A, B, C)and HLA class-II (DR) compatibility was considered inall patients, as well as a percentage of CD41, CD81,CD191 cells, and monocytes similar to that observed inhealthy controls.
Proliferation assays of PBMC from healthy controlsand recipients (isolated on days 0, 7, and 14) weresimultaneously performed in a classical one-way MLC,using 5 3 104 responder cells and 5 3 104 25-Gy irra-diated stimulator splenic cells from the correspondingliver donor, distributed in 96-well V-bottom cultureplates at a final volume of 0.2 ml of complete mediumper well. After a 5-day incubation period at 37°C in ahumidified 5% CO2 atmosphere, cells were pulsed with1 mCi/well [3H]TdR (Nuclear Iberica; ICN Pharmaceu-
ticals, Irvine, CA, USA), and harvested onto glass filters18 h later (Skatron Instruments, Suffolk, UK). DNAincorporation of [3H]TdR was detected in a liquid scin-tillation beta-counter (Rackbeta; Wallac, Turka, Fin-land), and cell proliferation was expressed as the mediancounts per minute of the triplicate wells.
Generation of cytotoxic effector cells from patientsand controls was performed by culturing 1 3 106 PBMCresponder cells against 1 3 106 irradiated stimulatorcells in 2 ml of complete medium, in a round-bottomtube, at 37°C in a humidified 5% CO2 atmosphere for 6days. Autologous cultures were always used as controls.Blocking experiments of proliferation and cytotoxic T-cell generation were simultaneously performed by adding5 mg/ml of anti-CD80 and anti-CD86 (BB1 and IT2.2,Pharmingen), anti-CD28 (CD28.1-15E8; CLB, Amster-dam, The Netherlands), or anti-CTLA-4 (BNI3.1,Pharmingen) mAb, at the beginning of the culture.
Cytotoxicity AssaysSpecific cytotoxic activity was tested using donor PHA-splenic blasts as targets and cells from MLC as effectors,in a classical 4-h assay, as previously described [21].Briefly, 1 3 106 splenic cells per well in complete me-dium supplemented with 1 mg/ml PHA-P (Sigma) wereincubated for 3 days in 24-well flat-bottom tissue cultureplates at 37°C in a humidified 5% CO2 atmosphere.Blast cells were labeled afterwards for 3 h with 1 mCi/ml51Cr (Nuclear Iberica) and extensively washed to removeany unbound radioisotope. Effector cells were washedtwice in complete medium and viability was examinedwith 0.5% trypan-blue, then mixed with 2 3 103 51Cr-labeled blast cells to obtain effector/target cell ratios of40:1, 20:1, 10:1, and 5:1. After incubation at 37°C in5% CO2 atmosphere for 4 h and centrifugation at 400 g,51Cr concentration was measured in recovered superna-tants using a gamma-counter (Clinigamma; Wallac). Themedian value of the triplicates was used to calculate thepercentage of specific lysis using the following formula:[(experimental release 2 spontaneous release) / (maximalrelease 2 spontaneous release)] 3 100. The spontaneousand maximal 51Cr releases were determined in the ab-sence of effector cells or after lysis with 1 N HCl,respectively.
ImmunohistochemistryPercutaneous liver biopsies were immediately embeddedin OCT compound (Sakura Finetechnical, Tokyo, Japan)and snap-frozen in liquid nitrogen. Sections (6 mm thick)were prepared and air dried for 1 h, fixed in 95% ethanolat 220°C for 10 min, washed three times for 5 min in0.16 M PBS pH 7.1, and stained using a standardimmunophosphatase protocol (super sensitive detectionkit; Biogenex, San Ramon, CA, USA). Briefly, sections
660 A. Minguela et al.

were treated with avidin, biotin (Avidin-biotin blocking;Biogenex), and human IgG heat-aggregated for 20 mineach to block endogenous biotin and nonspecific bind-ing, and washed three times in PBS after each blockingstep. Samples were marked for 45 min in a humidifiedchamber with 5 mg/ml anti-CD80 (BB1; Pharmingen),anti-CD86 (IT2.2; Pharmingen), or anti-CTLA-4(BNI3.1; Pharmingen) mAb in antibody-diluting buffer(Biomeda, Foster City, CA, USA), followed by a 20-minincubation period with biotinylated anti-mouse immu-noglobulin antibody and alkaline phosphatase-conju-gated streptavidin. After each procedure step, three5-min washes in PBS were performed. Finally, sectionswere treated for 10 to 15 min with freshly preparedchromogen, Fast-red/0.1%-Levamisole (Sigma), counter-stained with hematoxylin (Sigma), and mounted withhistomed (Concepta; Biosystems, Barcelona, Spain). Theentire procedure was performed at room temperature andnonspecific staining was controlled by incubation withimmunoglobulin class-matched nonspecific antibody.
Statistical AnalysisStatistical analysis of data was performed using BMDPsoftware [22]. Briefly, variance analysis of repeated mea-sures (BMDP2V.6) and stepwise discriminant analysis(BMDP7M) were carried out to detect differences be-tween groups and between pre- and post-transplant pe-riods. Data were expressed as mean 6 SD, and only pvalues below 0.05 were considered significant.
RESULTSCostimulatory Molecules in ConventionalCsA-treated PatientsAll patients had significant decreases in the absolutenumber of CD41CD281 PBL after transplant, and in-creases of CD191 lymphocytes from days 4 to 17 post-transplant. Clear differences were observed in the num-ber of CD191CD801 and CD191CD861 cells betweenthe acute rejection (AR) and nonacute rejection (NAR)groups in the total post-transplant period. Thus, in theNAR-group CD191CD801 and CD191CD861 cellsremained at pretransplant levels, whereas in the AR-group they significantly increased from days 7 to 17 afterthe graft (p , 0.05; Figure 1A). In this period, acuterejection was detected in 13 of 16 AR patients. The threeremaining recipients that showed acute rejection beyondday 17 post-transplant had lower increases of the B7 andCD28 molecules in this early period.
In the NAR-group, the CD80 and CD86 mean fluo-rescence intensity (MFI) of CD191 lymphocytes re-mained at basal levels, and the CD28 expression onCD41 lymphocytes gradually decreased. In contrast, allthese molecules were significantly upregulated in pa-
tients from the AR group on days with the highestfrequency of acute rejection (days 7 to 17 post-trans-plant; p , 0.01; Figure 1B), peaking on the day ofrejection diagnosis (Figure 1D). After antirejection treat-ment, CD28 recovered to the pretransplant level, butCD80 and CD86 upregulation persisted 4 to 6 daysbeyond diagnosis. The CD86 MFI in monocytes declinedin the post-transplant period and the CD80 expressionwas undetectable, but in no case were there significantdifferences between the AR and NAR groups (Figure1B).
Figure 1C represents CD80 and CD86 expression onCD191 lymphocytes from a representative patient of theAR group, with the histologic rejection diagnosed onday 10 post-transplant. The upregulation of CD80 andCD86 on CD191 lymphocytes began as early as day 4and peaked on day 12, when it was observed that themajority of CD191 cells became B71.
Costimulatory Molecules inNeoral-CsA-treated Patients
AR patients treated with Neoral-CsA exhibited increasesin the absolute number of all B lymphocyte subsets, aswell as the CD80 and CD86 MFI on CD191 cells duringthe acute rejection process. These changes, particularly ofthe CD80 molecule, were less important than thosedetected in patients receiving conventional-CsA (Figure2A). Recipients from the NAR-group treated with Ne-oral-CsA also did not modify their pretransplant expres-sion of CD80 and CD86 on CD191 lymphocytes. Figure2B shows the post-transplant evolution of CD80 andCD86 molecules in a patient from the AR group treatedwith Neoral-CsA. At the time of the rejection diagnosis(day 10 post-transplant), CD86 was upregulated in mostof the CD191 lymphocytes, but only a few of these cellsexpressed CD80.
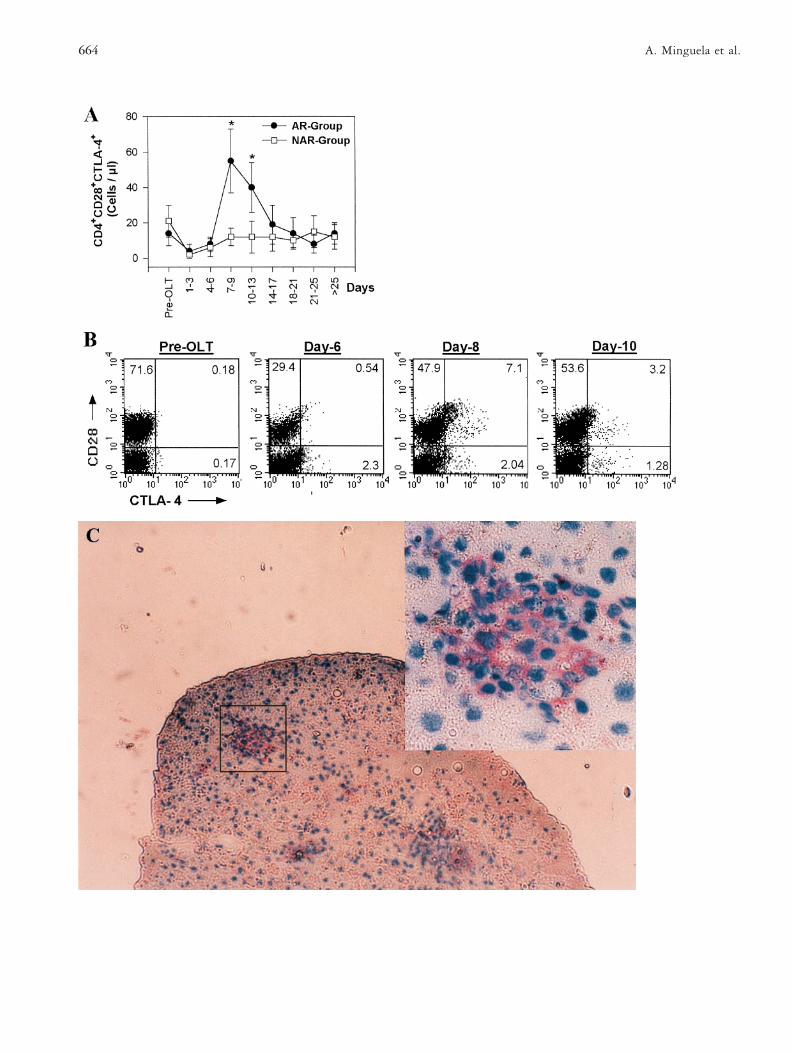
Under Neoral-CsA regimes, the CD28 expression onCD41 lymphocytes was similar to that observed in pa-tients treated with conventional-CsA, but additionally,in recipients treated with Neoral-CsA it was possible toevaluate the CTLA-4 evolution. The absolute number ofCD41CD281CTLA-41 cell subset remained at pre-transplant levels in the NAR patients, whereas this cellsubset was significantly augmented in the AR group ondays 7 to 13 post-transplant (p , 0.05; Figure 3A).During this period, 9 of 11 patients suffered from acuterejection. CD28 and CTLA-4 expression on PBL from arepresentative patient of the AR-group is shown in Fig-ure 3B, where it can be observed that a new populationof CTLA-41 lymphocytes (7.1% of total), which alsooverexpress the CD28 molecule, emerged 24 h before theday of rejection diagnosis (day 9).
661CD28/CTLA-4 and B7 Molecules in Human Liver Transplant
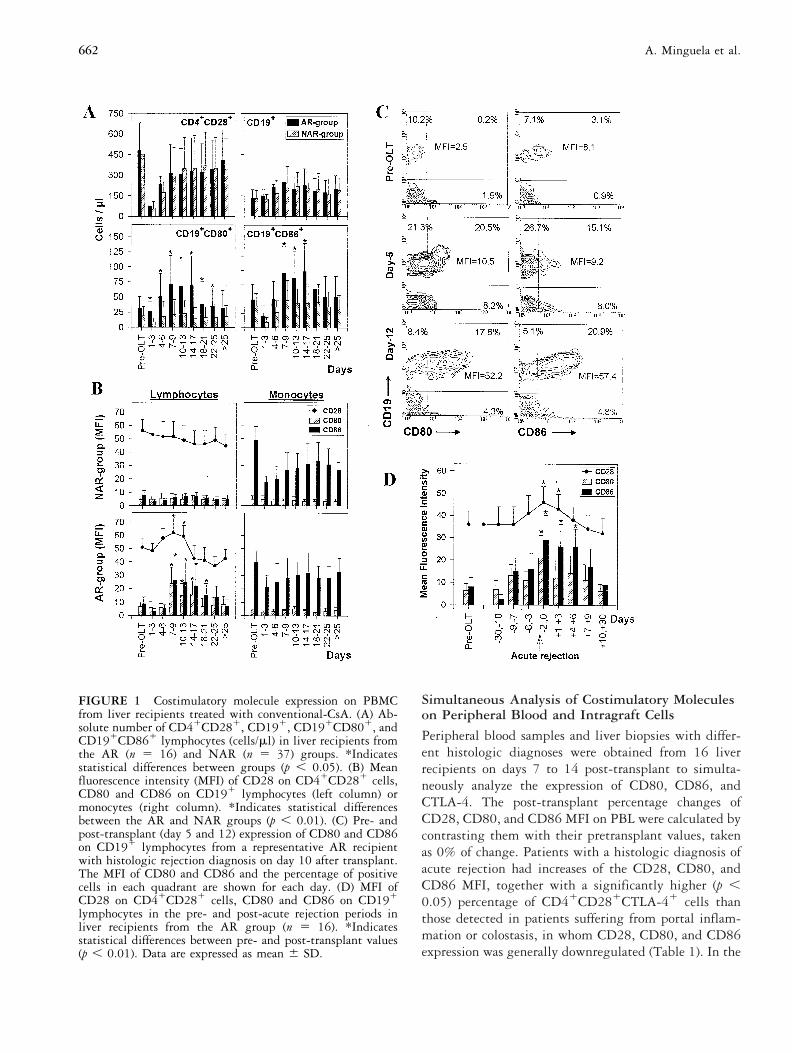
Simultaneous Analysis of Costimulatory Moleculeson Peripheral Blood and Intragraft Cells
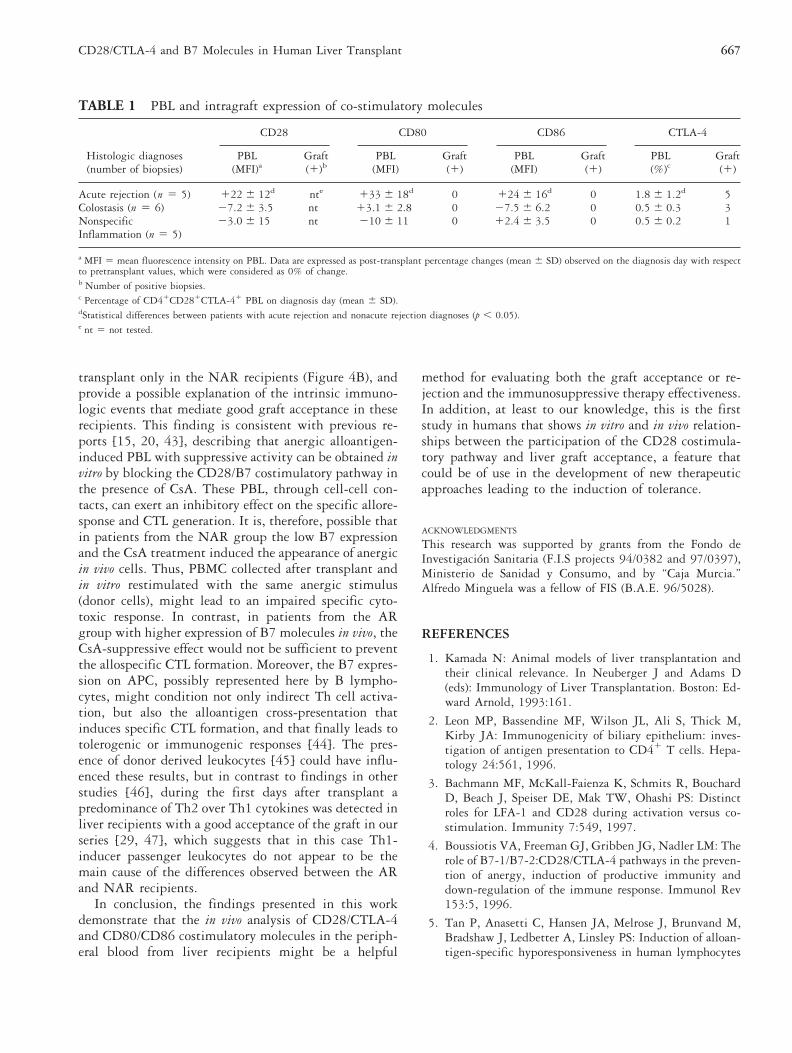
Peripheral blood samples and liver biopsies with differ-ent histologic diagnoses were obtained from 16 liverrecipients on days 7 to 14 post-transplant to simulta-neously analyze the expression of CD80, CD86, andCTLA-4. The post-transplant percentage changes ofCD28, CD80, and CD86 MFI on PBL were calculated bycontrasting them with their pretransplant values, takenas 0% of change. Patients with a histologic diagnosis ofacute rejection had increases of the CD28, CD80, andCD86 MFI, together with a significantly higher (p ,0.05) percentage of CD41CD281CTLA-41 cells thanthose detected in patients suffering from portal inflam-mation or colostasis, in whom CD28, CD80, and CD86expression was generally downregulated (Table 1). In the
FIGURE 1 Costimulatory molecule expression on PBMCfrom liver recipients treated with conventional-CsA. (A) Ab-solute number of CD41CD281, CD191, CD191CD801, andCD191CD861 lymphocytes (cells/ml) in liver recipients fromthe AR (n 5 16) and NAR (n 5 37) groups. *Indicatesstatistical differences between groups (p , 0.05). (B) Meanfluorescence intensity (MFI) of CD28 on CD41CD281 cells,CD80 and CD86 on CD191 lymphocytes (left column) ormonocytes (right column). *Indicates statistical differencesbetween the AR and NAR groups (p , 0.01). (C) Pre- andpost-transplant (day 5 and 12) expression of CD80 and CD86on CD191 lymphocytes from a representative AR recipientwith histologic rejection diagnosis on day 10 after transplant.The MFI of CD80 and CD86 and the percentage of positivecells in each quadrant are shown for each day. (D) MFI ofCD28 on CD41CD281 cells, CD80 and CD86 on CD191
lymphocytes in the pre- and post-acute rejection periods inliver recipients from the AR group (n 5 16). *Indicatesstatistical differences between pre- and post-transplant values(p , 0.01). Data are expressed as mean 6 SD.
662 A. Minguela et al.
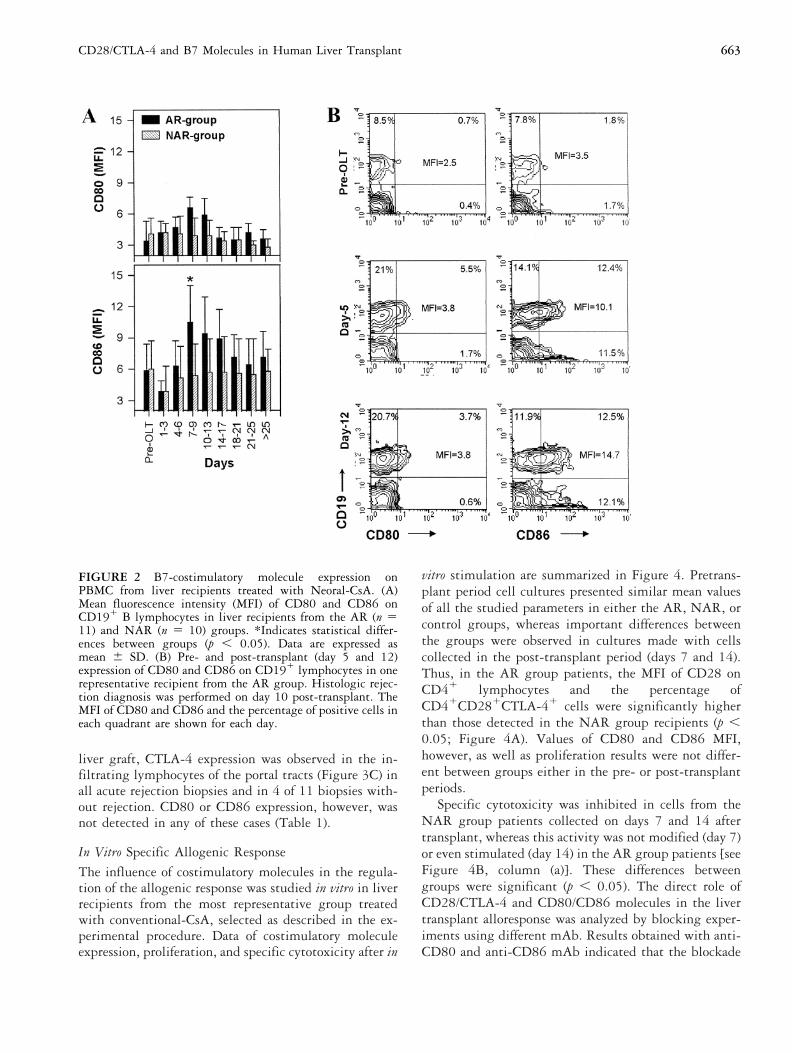
liver graft, CTLA-4 expression was observed in the in-filtrating lymphocytes of the portal tracts (Figure 3C) inall acute rejection biopsies and in 4 of 11 biopsies with-out rejection. CD80 or CD86 expression, however, wasnot detected in any of these cases (Table 1).
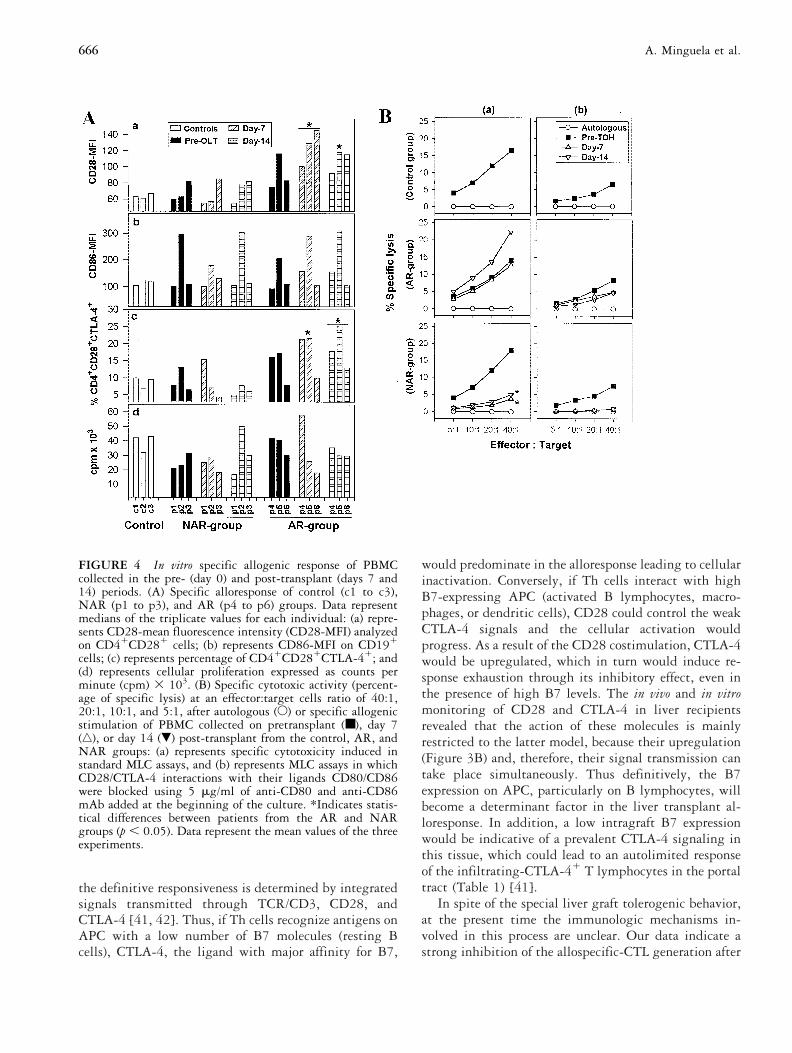
In Vitro Specific Allogenic ResponseThe influence of costimulatory molecules in the regula-tion of the allogenic response was studied in vitro in liverrecipients from the most representative group treatedwith conventional-CsA, selected as described in the ex-perimental procedure. Data of costimulatory moleculeexpression, proliferation, and specific cytotoxicity after in
vitro stimulation are summarized in Figure 4. Pretrans-plant period cell cultures presented similar mean valuesof all the studied parameters in either the AR, NAR, orcontrol groups, whereas important differences betweenthe groups were observed in cultures made with cellscollected in the post-transplant period (days 7 and 14).Thus, in the AR group patients, the MFI of CD28 onCD41 lymphocytes and the percentage ofCD41CD281CTLA-41 cells were significantly higherthan those detected in the NAR group recipients (p ,0.05; Figure 4A). Values of CD80 and CD86 MFI,however, as well as proliferation results were not differ-ent between groups either in the pre- or post-transplantperiods.
Specific cytotoxicity was inhibited in cells from theNAR group patients collected on days 7 and 14 aftertransplant, whereas this activity was not modified (day 7)or even stimulated (day 14) in the AR group patients [seeFigure 4B, column (a)]. These differences betweengroups were significant (p , 0.05). The direct role ofCD28/CTLA-4 and CD80/CD86 molecules in the livertransplant alloresponse was analyzed by blocking exper-iments using different mAb. Results obtained with anti-CD80 and anti-CD86 mAb indicated that the blockade
FIGURE 2 B7-costimulatory molecule expression onPBMC from liver recipients treated with Neoral-CsA. (A)Mean fluorescence intensity (MFI) of CD80 and CD86 onCD191 B lymphocytes in liver recipients from the AR (n 511) and NAR (n 5 10) groups. *Indicates statistical differ-ences between groups (p , 0.05). Data are expressed asmean 6 SD. (B) Pre- and post-transplant (day 5 and 12)expression of CD80 and CD86 on CD191 lymphocytes in onerepresentative recipient from the AR group. Histologic rejec-tion diagnosis was performed on day 10 post-transplant. TheMFI of CD80 and CD86 and the percentage of positive cells ineach quadrant are shown for each day.
663CD28/CTLA-4 and B7 Molecules in Human Liver Transplant
664 A. Minguela et al.

of these molecules induced a strong inhibition, not onlyof the cytotoxic response [Figure 4B, column (b)] butalso of the proliferative capacity, as well as the costimu-latory molecule overexpression (data not shown) in all theAR, NAR, and control groups. The results of the assaysperformed with anti-CD28 or anti-CTLA-4 mAb wereirrelevant.
DISCUSSIONThese results indicate that the acute rejection of humanliver transplants is clearly associated with an upregula-tion of CD28/CTLA-4-B7 molecules on T and B periph-eral blood lymphocytes. This fact suggests that cellularinteractions involving all these costimulatory moleculesmay have a decisive role in human liver graft acceptanceor rejection, probably by triggering CsA-resistant signalsthat control Th lymphocyte activation and allospecific-CTL generation [23, 24]. Studies performed in experi-mental transplant models demonstrate that the block-ade of CD28-B7 binding prolongs graft survival andinduces specific tolerance [6 – 8, 25]. In human livertransplants as well, previous reports indicate that theCD28 pathway appears to be important [9]. This isnow further supported by new in vivo and in vitrostudies of CD28-B7 modulation, which demonstratetheir relevance in the specific alloresponse and re-stricted CTL generation.
Because of the reduced number of mature dendriticcells in the liver [26] and the practical absence of co-stimulatory molecules on sinusoidal and biliar-epithelialcells [2, 15], direct allorecognition could be less relevantin this type of graft. The results of the present studydemonstrate a reduced expression of B7 molecules onliver graft samples obtained during acute rejection epi-sodes, together with the concomitant presence of acti-vated CD41 and CD191 lymphocytes overexpressingCD28/CTLA-4 or CD80/CD86 in peripheral blood (Fig-ure 1). These findings suggest that Th allospecific clones
are activated in peripheral tissues by alloantigen recog-nition on autologous APC (possibly B7-positive B lym-phocytes). Subsequently, these specifically activated Tcells can migrate to the graft where, by inflammatorycytokine secretion, they induce HLA class I and class IIupregulation on biliar duct and endothelial cells [27],which in turn become targets for the direct recognitionof T CD41 and CD81 lymphocytes.
In liver transplants, immunosuppressive agents andthe high post-transplant IL-10 serum concentration de-scribed by Le Moine et al. [28], and confirmed in ourown series [29], might hinder the ability of macrophagesand dendritic cells to stimulate Th1 but not Th2 lym-phocytes, without affecting B lymphocyte accessoryfunctions [30–33]. Figure 1B shows that after trans-plant, B7 molecules remain at basal levels in monocytesor are even downregulated during the acute rejectionprocess, unlike in B lymphocytes. Therefore, it is tempt-ing to speculate that, depending on their costimulatorymolecule levels, B lymphocytes are the APC that mod-ulate the host response that leads to graft rejection oracceptance [16]. Thus, resting B lymphocytes (B7 neg-atives) from NAR recipients would present alloantigensto Th cells in a tolerogenic manner favoring graft accep-tance, whereas activated B lymphocytes (B7 positives)from the AR patients using the CD28 pathway might beefficient APC [14, 18, 34]. Furthermore, the highercapacity of AR recipients to upregulate CD28 on Tlymphocytes, observed after specific in vitro stimulationof PBMC collected in the post-transplant period, sup-ports that the in vivo CD28 upregulation could be re-sponsible for the strong alloresponse triggered during theacute rejection process in these patients. In contrast, thelow CD28 expression displayed by T lymphocytes in theNAR patients could be indicative of nonefficient ortolerogenic TCR engagements [3, 35].
Despite the different CD28-B7 expression patterns,CsA blood concentrations were not different in patientsof the AR and NAR groups treated either with conven-tional or Neoral CsA (data not shown). Nevertheless,patients under Neoral-CsA regimes exhibited slightlyhigher and less variable CsA blood levels, which couldhave induced their lower expression of B7 molecules(Figure 2), particularly of those such as CD80, requiringa long lasting cellular stimulation to be upregulated[36]. In contrast, these changes did not affect CD28expression or the acute rejection rate.
Accumulating evidence supports that TCR and CD28transduce early stimulatory signals in the immunore-sponse, whereas in a later step, CTLA-4 appears to in-hibit it [37, 38] by apoptosis induction [39] or, moreprobably, by reducing IL-2 secretion [40]. More recentreports establish, however, the participation of CTLA-4in the early cellular activation events in such a way that
4™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™FIGURE 3 PBMC and liver graft expression of CTLA-4 inrecipients treated with Neoral-CsA. A) Absolute number ofCD41CD281CTLA-41 lymphocytes (cells/ml, mean 6 SD) inliver recipients from the AR (n 5 11) and NAR (n 5 10)groups. *Indicates statistical differences between groups (p ,0.05). (B) Pre- and post-transplant expression of CD28 andCTLA-4 on peripheral blood lymphocytes in one representa-tive recipient from the AR group with acute rejection diag-nosis on day 9 after graft. The percentage of positive cells ineach quadrant is shown for each day. (C) CTLA-4 expression onportal tract infiltrating cells in liver tissue obtained on day 7post-transplant from an AR-diagnosed recipient (originalmagnification 3 100). Superior right corner shows a detail ofthe infiltrating CTLA-41 lymphocytes in the portal tract(original magnification 3 400).
665CD28/CTLA-4 and B7 Molecules in Human Liver Transplant
the definitive responsiveness is determined by integratedsignals transmitted through TCR/CD3, CD28, andCTLA-4 [41, 42]. Thus, if Th cells recognize antigens onAPC with a low number of B7 molecules (resting Bcells), CTLA-4, the ligand with major affinity for B7,
would predominate in the alloresponse leading to cellularinactivation. Conversely, if Th cells interact with highB7-expressing APC (activated B lymphocytes, macro-phages, or dendritic cells), CD28 could control the weakCTLA-4 signals and the cellular activation wouldprogress. As a result of the CD28 costimulation, CTLA-4would be upregulated, which in turn would induce re-sponse exhaustion through its inhibitory effect, even inthe presence of high B7 levels. The in vivo and in vitromonitoring of CD28 and CTLA-4 in liver recipientsrevealed that the action of these molecules is mainlyrestricted to the latter model, because their upregulation(Figure 3B) and, therefore, their signal transmission cantake place simultaneously. Thus definitively, the B7expression on APC, particularly on B lymphocytes, willbecome a determinant factor in the liver transplant al-loresponse. In addition, a low intragraft B7 expressionwould be indicative of a prevalent CTLA-4 signaling inthis tissue, which could lead to an autolimited responseof the infiltrating-CTLA-41 T lymphocytes in the portaltract (Table 1) [41].
In spite of the special liver graft tolerogenic behavior,at the present time the immunologic mechanisms in-volved in this process are unclear. Our data indicate astrong inhibition of the allospecific-CTL generation after
FIGURE 4 In vitro specific allogenic response of PBMCcollected in the pre- (day 0) and post-transplant (days 7 and14) periods. (A) Specific alloresponse of control (c1 to c3),NAR (p1 to p3), and AR (p4 to p6) groups. Data representmedians of the triplicate values for each individual: (a) repre-sents CD28-mean fluorescence intensity (CD28-MFI) analyzedon CD41CD281 cells; (b) represents CD86-MFI on CD191
cells; (c) represents percentage of CD41CD281CTLA-41; and(d) represents cellular proliferation expressed as counts perminute (cpm) 3 103. (B) Specific cytotoxic activity (percent-age of specific lysis) at an effector:target cells ratio of 40:1,20:1, 10:1, and 5:1, after autologous (E) or specific allogenicstimulation of PBMC collected on pretransplant (■), day 7(‚), or day 14 (�) post-transplant from the control, AR, andNAR groups: (a) represents specific cytotoxicity induced instandard MLC assays, and (b) represents MLC assays in whichCD28/CTLA-4 interactions with their ligands CD80/CD86were blocked using 5 mg/ml of anti-CD80 and anti-CD86mAb added at the beginning of the culture. *Indicates statis-tical differences between patients from the AR and NARgroups (p , 0.05). Data represent the mean values of the threeexperiments.
666 A. Minguela et al.
transplant only in the NAR recipients (Figure 4B), andprovide a possible explanation of the intrinsic immuno-logic events that mediate good graft acceptance in theserecipients. This finding is consistent with previous re-ports [15, 20, 43], describing that anergic alloantigen-induced PBL with suppressive activity can be obtained invitro by blocking the CD28/B7 costimulatory pathway inthe presence of CsA. These PBL, through cell-cell con-tacts, can exert an inhibitory effect on the specific allore-sponse and CTL generation. It is, therefore, possible thatin patients from the NAR group the low B7 expressionand the CsA treatment induced the appearance of anergicin vivo cells. Thus, PBMC collected after transplant andin vitro restimulated with the same anergic stimulus(donor cells), might lead to an impaired specific cyto-toxic response. In contrast, in patients from the ARgroup with higher expression of B7 molecules in vivo, theCsA-suppressive effect would not be sufficient to preventthe allospecific CTL formation. Moreover, the B7 expres-sion on APC, possibly represented here by B lympho-cytes, might condition not only indirect Th cell activa-tion, but also the alloantigen cross-presentation thatinduces specific CTL formation, and that finally leads totolerogenic or immunogenic responses [44]. The pres-ence of donor derived leukocytes [45] could have influ-enced these results, but in contrast to findings in otherstudies [46], during the first days after transplant apredominance of Th2 over Th1 cytokines was detected inliver recipients with a good acceptance of the graft in ourseries [29, 47], which suggests that in this case Th1-inducer passenger leukocytes do not appear to be themain cause of the differences observed between the ARand NAR recipients.
In conclusion, the findings presented in this workdemonstrate that the in vivo analysis of CD28/CTLA-4and CD80/CD86 costimulatory molecules in the periph-eral blood from liver recipients might be a helpful
method for evaluating both the graft acceptance or re-jection and the immunosuppressive therapy effectiveness.In addition, at least to our knowledge, this is the firststudy in humans that shows in vitro and in vivo relation-ships between the participation of the CD28 costimula-tory pathway and liver graft acceptance, a feature thatcould be of use in the development of new therapeuticapproaches leading to the induction of tolerance.
ACKNOWLEDGMENTS
This research was supported by grants from the Fondo deInvestigacion Sanitaria (F.I.S projects 94/0382 and 97/0397),Ministerio de Sanidad y Consumo, and by “Caja Murcia.”Alfredo Minguela was a fellow of FIS (B.A.E. 96/5028).
REFERENCES
1. Kamada N: Animal models of liver transplantation andtheir clinical relevance. In Neuberger J and Adams D(eds): Immunology of Liver Transplantation. Boston: Ed-ward Arnold, 1993:161.
2. Leon MP, Bassendine MF, Wilson JL, Ali S, Thick M,Kirby JA: Immunogenicity of biliary epithelium: inves-tigation of antigen presentation to CD41 T cells. Hepa-tology 24:561, 1996.
3. Bachmann MF, McKall-Faienza K, Schmits R, BouchardD, Beach J, Speiser DE, Mak TW, Ohashi PS: Distinctroles for LFA-1 and CD28 during activation versus co-stimulation. Immunity 7:549, 1997.
4. Boussiotis VA, Freeman GJ, Gribben JG, Nadler LM: Therole of B7-1/B7-2:CD28/CTLA-4 pathways in the preven-tion of anergy, induction of productive immunity anddown-regulation of the immune response. Immunol Rev153:5, 1996.
5. Tan P, Anasetti C, Hansen JA, Melrose J, Brunvand M,Bradshaw J, Ledbetter A, Linsley PS: Induction of alloan-tigen-specific hyporesponsiveness in human lymphocytes
TABLE 1 PBL and intragraft expression of co-stimulatory molecules
Histologic diagnoses
CD28 CD80 CD86 CTLA-4
PBL Graft PBL Graft PBL Graft PBL Graft(number of biopsies) (MFI)a (1)b (MFI) (1) (MFI) (1) (%)c (1)
Acute rejection (n 5 5) 122 6 12d nte 133 6 18d 0 124 6 16d 0 1.8 6 1.2d 5Colostasis (n 5 6) 27.2 6 3.5 nt 13.1 6 2.8 0 27.5 6 6.2 0 0.5 6 0.3 3Nonspecific 23.0 6 15 nt 210 6 11 0 12.4 6 3.5 0 0.5 6 0.2 1Inflammation (n 5 5)
a MFI 5 mean fluorescence intensity on PBL. Data are expressed as post-transplant percentage changes (mean 6 SD) observed on the diagnosis day with respectto pretransplant values, which were considered as 0% of change.b Number of positive biopsies.c Percentage of CD41CD281CTLA-41 PBL on diagnosis day (mean 6 SD).dStatistical differences between patients with acute rejection and nonacute rejection diagnoses (p , 0.05).e nt 5 not tested.
667CD28/CTLA-4 and B7 Molecules in Human Liver Transplant

by blocking interaction of CD28 with its natural ligandB7/BB1. J Exp Med 177:165, 1993.
6. Lenschow DJ, Zeng Y, Hathcock KS, Zuckerman LA,Freeman G, Thistlethwaite JR, Gray GS, Nadler LM,Goodnow CC, Bluestone JA: Inhibition of transplant re-jection following treatment with anti-B-2 and anti-B-1antibodies. Transplantation 60:1171, 1995.
7. Akalin E, Chandraker A, Russell ME, Turka LA, HancockWW, Sayegh MH: CD28-B7 T cell costimulatory block-ade by CTLA4Ig in the rat renal allograft model: inhibi-tion of cell-mediated and humoral immune responses invivo. Transplantation 62:1942, 1996.
8. Lakkis FG, Konieczny BT, Saleem S, Baddoura FK, Lins-ley PS, Alexander DZ, Lowry RP, Pearson TC, Larsen CP:Blocking the CD28-B7 T cell costimulation pathwayinduces long term cardiac allograft acceptance in theabsence of IL-4. J Immunol 158:2443, 1997.
9. Garcıa-Alonso AM, Minguela A, Muro M, Ontanon J,Torıo A, Marın L, Lopez-Segura P, Miras M, Alvarez-Lopez M-R: CD28 expression on peripheral blood T lym-phocytes after orthotopic liver transplant: up-regulationin acute rejection. Hum Immunol 53:64, 1997.
10. Lechler RY, Batchelor JR: Restoration of immunogenicityto passenger cell-depletion kidney allografts by the addi-tion of donor strein dendritic cells. J Exp Med 155:31,1982.
11. Liu Z, Colovai AI, Tugulea S, Reed EF, Fisher PE, Man-cini D, Rose EA, Cortesini R, Michler RE, Suciu-Foca N:Indirect recognition of donor HLA-DR peptides in organallograft rejection. J Clin Invest 98:1150, 1996.
12. Malajoni ER, Cinti P, Orlandini A, Malajoni J, Tugulea S,Ho E, Liu Z, Suciu-Foca N, Cortesini R: Mechanism ofliver allograft rejection: the indirect recognition pathway.Hum Immunol 53:57, 1997.
13. Quill H: Anergy as a mechanism of peripheral T celltolerance. J Immunol 156:1325, 1996.
14. Gilbert KM, Weigle WO: B cell presentation of a tolero-genic signal to Th clones. Cell Immunol 139:58, 1992.
15. Van Gool SW, Vanderberghe P, de Boer M, Ceuppens JL:CD80, CD86 and CD40 provide accessory signals in amultiple-step T-cell activation model. Immunol Rev 153:47, 1996.
16. Gilbert KM, Weigle WO: Tolerogenicity of resting andactivated B cells. J Exp Med 179:249, 1994.
17. Wilson JL, Cunningham AC, Kirby JA: Alloantigen pre-sentation by B cells: analysis of the requirement for B-cellactivation. Immunology 86:325, 1995.
18. Croft M, Joseph SB, Miner KT: Partial activation of naiveCD4 T cells and tolerance induction in response to pep-tide presented by resting B cells. J Immunol 159:3257,1997.
19. Minguela A, Alvarez-Lopez MR, Sanchez-Bueno F, TorıoA, Muro M, Robles R, Pons JA, Parrilla P, Garcıa-AlonsoAM: Presence of different T and B peripheral blood lym-phocytes subsets in liver transplantation after Cyclospor-
ine or OKT3 immunosuppressive treatment. TransplantProc 27:2317, 1995.
20. Mor E, Solomon H, Gibbs JF, Holman MJ, GoldsteinRM, Husberg BS, Gonwa TA, Klintmalm GB: Acutecellular rejection following liver transplantation: clinicalpathologic features and effect on outcome. Semin LiverDis 12:28, 1992.
21. Kristensen T: Human histocompatibility testing by T-cell-mediated lympholysis: a European standard CMLtechnique. Tissue Antigens 16:335, 1980.
22. Dixon WJ (ed): BMDP Statistical Software. Berkeley, CA:University Press of California Ltd., 1992.
23. Van Gool SW, de Boer M, Ceuppens JL: CD28 ligation bymonoclonal antibodies or B7/BB1 provides an accessorysignal for the Cyclosporin A-resistant generation of cyto-toxic T cell activity. J Immunol 150:3254, 1993.
24. June CH, Ledbetter JA, Gillespie MM, Lindsten T,Thompson CB: T-cell proliferation involving the CD28pathway is associated with Cyclosporin-resistant interleu-kin 2 gene expression. Mol Cell Biol 7:4472, 1987.
25. Sayegh MH, Turka LA: The role of T-cell costimulatoryactivation pathways in transplant rejection. N Engl J Med338:1813, 1998.
26. Thomson AW, Lu L: Are dendritic cells the key to livertransplant tolerance? Immunol Today 20:27, 1999.
27. Koskimies S, Lautenschlager I: Transplantation antigens.In Neuberger J, Adams D (eds): Immunology of LiverTransplantation. Boston: Edward Arnold, 1993:127.
28. Le Moine O, Marchant A, Durand F, Ickx B, Pradier O,Belghiti J, Abramovicz D, Gelin M, Goldman M, DeviereJ: Systemic release of Interleukin-10 during orthotopicliver transplantation. Hepatology 20:889, 1994.
29. Minguela A, Torıo A, Marın L, Muro M, Villar LM, DıazJ, Ramırez P, Parrilla P, Garcıa-Alonso AM, Alvarez-Lopez MR: Implication of soluble and membrane HLAclass-I and serum IL-10 in liver graft acceptance. HumImmunol 60:500, 1999.
30. Fiorentino DF, Zlotnick R, Mosmann TR, Howard M,O9Garra A: IL-10 inhibits cytokine production by acti-vated macrophages. J Immunol 147:3815, 1991.
31. Peguet-Navarro J, Moulon C, Caux C, Dalbiez-GauthierC, Banchereau J, Schmitt D: Interleukin-10 inhibits theprimary allogenic T cell response to human epidermalLanherhans cells. Eur J Immunol 24:884, 1994.
32. Ding L, Linsley PS, Huang LY, Germain RN, ShevachEM: IL-10 inhibits macrophage costimulatory activity byselectively inhibiting the up-regulation of B-7 expression.J Immunol 151:1224, 1993.
33. Willems F, Marchant A, Delville JP, Gerard C, DelvauxA, Velu T, de Boer M, Goldman M: IL-10 inhibits B7 andICAM-1 expression on human monocytes. Eur J Immunol24:1007, 1994.
34. Eynon EE, Parker DC: Small B cells as antigen-presenting
668 A. Minguela et al.

cells in the induction of tolerance to soluble proteinantigens. J Exp Med 175:131, 1992.
35. Viola A, Lanzavecchia A: T cell activation determined byT cell receptor number and tunable thresholds. Science273:104, 1996.
36. Hathcock KS, Laszlo G, Pucillo C, Linsley PS, Hodes RJ:Comparative analysis of B7-1 and B7-2 costimulatoryligands: expression and function. J Exp Med 180:631,1994.
37. Kearney ER, Walunas TL, Karr RW, Morton PA, LohDY, Bluestone JA, Jenkins MK: Antigen-dependentclonal expansion of a trace population of antigen-specificCD41 T cells in vivo is dependent on CD28 costimula-tion and inhibited by CTLA-4. J Immunol 155:1032,1995.
38. Tivol EA, Borriello F, Schweitzer AN, Lynch WP, Blue-stone JA, Sharpe AH: Loss of CTLA-4 leads to massivelymphoproliferation and fatal multiorgan tissue destruc-tion, revealing a critical negative regulatory role ofCTLA-4. Immunity 3:541, 1995.
39. Gribben GJ, Freeman GJ, Boussiotis VA, Rennert P,Jellis CL, Greenfield E, Barber M, Restivo VA, Ke XY,Gray GS, Nadler LM: CTLA-4 mediates antigen-specificapoptosis in human T cells. Proc Natl Acad Sci USA92:811, 1995.
40. Walunas T, Bakker CY, Bluestone JA: CTLA-4 ligationblocks CD28-dependent T cell activation. J Exp Med183:2541, 1996.
41. Chambers CA, Kummel MF, Boitel B, Hurwitz A, Sul-livan TJ, Fournier S, Cassell D, Brunner M, Allison JP:The role of CTLA-4 in the regulation and inhibition ofT-cell responses. Immunol Rev 153:27, 1996.
42. Thompson CB, Allison JP: The emerging role of CTLA-4as an immune attenuator. Immunity 7:445, 1997.
43. Comoli P, Montagna D, Moretta A, Zecca M, Locatelli F,Maccario R: Alloantigen-induced human lymphocytesrendered nonresponsives by combination of anti-CD80monoclonal antibodies and cyclosporin-A suppress mixedlymphocyte reaction in vitro. J Immunol 155:5506, 1995.
44. Carbone FR, Kurts C, Bennett SRM, Miller JFAP, HeathWR: Cross-presentation: a general mechanism for CTLimmunity and tolerance. Immunol Today 19:368, 1998.
45. Jonson JR, Hogan PG, Thomas R, Steadman C, CloustonAD, Balderson GA, Lynch SV, Strong RW, Powell EE:Peripheral blood chimerism following human liver trans-plantation. Hepatology 25:1233, 1997.
46. Bishop GA, Sun J, DeCruz DJ, Rokahr KL, Sedgwick JD,Sheil AGR, Gallagher ND, McCaughan GW: Toleranceto rat liver allografts. Donor cell migration and tolerance-associated cytokine production in peripheral lymphoidtissues. J Immunol 156:4925, 1996.
47. Minguela A, Torıo A, Marın L, Sanchez-Bueno F, Garcıa-Alonso AM, Ontanon J, Parrilla P, Alvarez-Lopez MR:Implication of Th1, Th2 and Th3 cytokines in liver graftacceptance. Transplant Proc 31:516, 1999.
669CD28/CTLA-4 and B7 Molecules in Human Liver Transplant
Copyright © 2022 FDOKUMEN




















