
Calcium transport by cardiac sarcoplasmic reticulum and phosphorylation of phospholamban
23
Calcium transport by cardiac sarcoplasmic reticulum and phosphorylation of phospholamban M. Tada, M. Yamada, M. Kadoma, M. Inui and F. Ohmori 1 Division of Cardiology, First Department of Medicine, Osaka University School of Medicine, Osaka 553, and 1Department of Medicine, National Cardiovascular Center, Osaka 565, Japan Summary Active calcium transport by cardiac sarcoplasmic reticulum assumes a central role in the excitation-con- centration coupling of the myocardium, in that Ca2+-dependent ATPase (mol.wt. 100 000) of cardiac sarcoplasmic reticulum serves as an energy transducer and a translocator of Ca 2+ across the membrane. During the translocation of Ca 2+, the ATPase undergoes a complex series of reactions during which the phosphorylated intermediate EP is formed. We documented how the elementary steps of the ATPase reaction are coupled with calcium translocation, and provided evidences to indicate that two key steps of ATPase correspond to the conformational change of the enzyme, and appear to alter the affinity of the enzyme for Ca 2+. A line of evidence also indicated that Ca2+-dependent ATPase of cardiac sarcoplasmic reticulum is regulated by a specific protein named phospholamban (mol.wt. 22 000), which serves as a substrate for cyclic AMP-dependent protein kinase. Cyclic AMP-dependent phosphorylation of ph0spholamban resulted in a marked increase in the rate of turnover of the ATPase, by enhancing the rates of the key elementary steps, i.e. the steps at which the intermediate EP is formed and decomposed. Thus phospholamban is putatively thought to serve as a modulator of Ca2+-dependent ATPase of cardiac sarcoplasmic reticulum. A working model was proposed to interpret the mechanism. Also documented is a possibility that another protein kinase activatable by Ca 2+ and calmodulin is functional in regulating the phospholamban-ATPase system, thus suggesting the existence of a dual control system, in which both cyclic AMP- and calmodulin-dependent phosphorylation are in control of the Ca2+-dependent ATPase. Such a control mechanism may provide the interpretation, at the cellular level, that catecholamines exert actions on myocardial contractility. Thus, catecholamine-mediated increases in intracellular cyclic AMP could enhance calcium fluxes across the membrane of sarcoplasmic reticulum, thus resulting in the increased rates of relaxation and, at the same time, the increased rate and extent of contraction. Such a mechanism could also be operational in the tissues, other than the myocardium, in which catecholamines and other hormones serve as the 'first messenger', producing intracellular cyclic AMP as the 'second messenger'. Introduction Calcium fluxes across the membrane of sarco- plasmic reticulum assume central roles in the exci- tation-contraction coupling of striated muscles (I, 2). In skeletal muscle, release and subsequent ac- cumulation of Ca 2+ by sarcoplasmic reticulum trigger contraction and relaxation, respectively, of myofibrillar bundles. Calcium accumulation, often termed as calcium uptake, is carried out by the ATP-supported active transport of this cation across the membrane of sarcoplasmic reticulum (1). In the excitation-contraction coupling of the mam- malian myocardium(ventricular muscle), the mech- Molecular and Cellular Biochemistry 46, 73-95 (1982). 0300 8177/82/0462 0073/$ 04.60. © 1982, Martinus Nijhoff/Dr W. Junk Publishers, The Hague. Printed in The Netherlands.
-
Upload
yamaguchi-u -
Category
Documents
-
view
1 -
download
0
Transcript of Calcium transport by cardiac sarcoplasmic reticulum and phosphorylation of phospholamban

Calcium transport by cardiac sarcoplasmic reticulum and phosphorylation of phospholamban
M. Tada, M. Yamada, M. Kadoma, M. Inui and F. Ohmori 1 Division of Cardiology, First Department of Medicine, Osaka University School of Medicine, Osaka 553, and 1Department of Medicine, National Cardiovascular Center, Osaka 565, Japan
Summary
Active calcium transport by cardiac sarcoplasmic reticulum assumes a central role in the excitation-con- centration coupling of the myocardium, in that Ca2+-dependent ATPase (mol.wt. 100 000) of cardiac sarcoplasmic reticulum serves as an energy transducer and a translocator of Ca 2+ across the membrane. During the translocation of Ca 2+, the ATPase undergoes a complex series of reactions during which the phosphorylated intermediate EP is formed. We documented how the elementary steps of the ATPase reaction are coupled with calcium translocation, and provided evidences to indicate that two key steps of ATPase correspond to the conformational change of the enzyme, and appear to alter the affinity of the enzyme for Ca 2+.
A line of evidence also indicated that Ca2+-dependent ATPase of cardiac sarcoplasmic reticulum is regulated by a specific protein named phospholamban (mol.wt. 22 000), which serves as a substrate for cyclic AMP-dependent protein kinase. Cyclic AMP-dependent phosphorylation of ph0spholamban resulted in a marked increase in the rate of turnover of the ATPase, by enhancing the rates of the key elementary steps, i.e. the steps at which the intermediate EP is formed and decomposed. Thus phospholamban is putatively thought to serve as a modulator of Ca2+-dependent ATPase of cardiac sarcoplasmic reticulum. A working model was proposed to interpret the mechanism. Also documented is a possibility that another protein kinase activatable by Ca 2+ and calmodulin is functional in regulating the phospholamban-ATPase system, thus suggesting the existence of a dual control system, in which both cyclic AMP- and calmodulin-dependent phosphorylation are in control of the Ca2+-dependent ATPase.
Such a control mechanism may provide the interpretation, at the cellular level, that catecholamines exert actions on myocardial contractility. Thus, catecholamine-mediated increases in intracellular cyclic AMP could enhance calcium fluxes across the membrane of sarcoplasmic reticulum, thus resulting in the increased rates of relaxation and, at the same time, the increased rate and extent of contraction. Such a mechanism could also be operational in the tissues, other than the myocardium, in which catecholamines and other hormones serve as the 'first messenger', producing intracellular cyclic AMP as the 'second messenger'.
Introduction
Calcium fluxes across the membrane of sarco- plasmic reticulum assume central roles in the exci- tation-contraction coupling of striated muscles (I, 2). In skeletal muscle, release and subsequent ac- cumulation of Ca 2+ by sarcoplasmic reticulum
trigger contraction and relaxation, respectively, of myofibrillar bundles. Calcium accumulation, often termed as calcium uptake, is carried out by the ATP-supported active transport of this cation across the membrane of sarcoplasmic reticulum (1). In the excitation-contraction coupling of the mam- malian myocardium(ventricular muscle), the mech-
Molecular and Cellular Biochemistry 46, 73-95 (1982). 0300 8177/82/0462 0073/$ 04.60. © 1982, Martinus Nijhoff/Dr W. Junk Publishers, The Hague. Printed in The Netherlands.

74
anisms of calcium fluxes by sarcoplasmic reticulum are essentially similar, in that the Ca2+-dependent ATPase enzyme is responsible for calcium uptake during relaxation (1) and calcium release from sar- coplasmic reticulum could largely be responsible for initiating contraction (2, 3).
Adrenergic intervention is known to exhibit re- markable influence on the excitation-contraction coupling of the myocardium, notably the contrac- tility of ventricular and atrial muscles (inotropic effects). Cyclic AMP was found to serve as a second messenger mediating the fl-adrenergic actions of catecholamines on the myocardium (4, 5), although the underlying mechanisms to explain these effects have not entirely been clarified. In searching for possible subcellular mechanisms of cyclic AMP ef- fects, a protein kinase was found to be activated in a cyclic AMP-dependent manner (6). This enzyme catalyzes phosphorylation of phosphorylase kinase and glycogen synthetase, resulting in a marked in- crease in glycolysis (7, 8). i t was observed, however, that the positive inotropic responses evidently pre- ceded such metabolic changes when well-oxygen- ated myocardium was exposed to catecholamines (5), indicating that cyclic AMP-mediated increase in glycolysis may represent a response to meet the increased energy expenditure due to increased con- tractility and electrical activity of the myocardium (5).
In view of the pivotal role of calcium in the excitation-contraction coupling of the myocardi- urn, it is of importance to elucidate the processes through which cyclic AMP exerts actions on trans- and intracellular calcium movements. An increas- ing number of exPerimental data indicated that a cyclic AMP-dependent protein kinase catalyzes phosphorylation of cardiac membranes, notably sarcoplasmic reticulum and sarcolemma, with the concomitant alterations in membrane functions (9). In particular, active calcium transport by cardiac sarcoplasmic retieulum was found to be regulated by cyclic AMP-dependent phosphorylation of a specific membrane protein of 22 000 daltons (9), which was referred to by Tada et al. (10, 11) as phospholamban. Such an interaction between cal- cium and cyclic AMP within the myocardial cell may represent a key reaction step that mediate the inotropic actions of adrenergic agonists and antag- onists on the myocardium. Possible existence of a similar mechanism is also considered in other mus-
cular and nonmuscular tissues (9). Phospholamban is also phosphorylated by another protein kinase, that is activated by calcium and calmodulin.
This paper gives an outline of the present consen- sus on active calcium transport by cardiac sarco- plasmic reticulum and its relationship to phospho- rylation of phospholamban. Also documented will be a molecular mode by which phospholamban could interact with Ca2+-dependent ATPase en- zyme. Based on such molecular concepts that evolved quite recently, we shall discuss and inter- pret the physiological significance of the interplay between calcium and cyclic AMP in producing reg- ulation of motility in the myocardium and other tissues.
Structure and function of cardiac sarcoplasmic re- ticulum
Sarcoplasmic reticulum of the myocardium con- sists of a membrane-limited reticular structure of continuous vesicles and tubules that form a net- work surrounding the myofibrillar bundles (12, 12a) (Fig. 1). Another tubular structure running perpendicular to the long axis of the myocardial cell (the transverse tubule or Tsystem) was found not to belong to the structure of sarcoplasmic reticulum per se, but to be continuous with the surface mem- brane (sarcolernma), indicating that the lumen o f t system is a continuation of the extracellular space. In contrast, the lumen of sarcoplasmic reticulum forms a closed intracellular system, that is not con- tinuous with the extracellular space. The mem- brane ofT system, however, forms a junction with a part of sarcoplasmic reticulum (subsarcolemmal cisternae), that is thickened to form a foot-like structure (Fig. 1). The subsarcolemmal cisternae attached to the membrane of T system and surface membrane are believed to provide the basis for transmitting a signal from sarcolemma to sarco- plasmic reticulum, that triggers contraction and relaxation of myofibrils. Thus, during a contracti- on-relaxation cycle of the myocardium, the sarco- plasmic reticulum controls fluxes of calcium ions across its membrane, in that calcium is released from the sarcoplasmic reticulum to induce contrac- tion while relaxation is produced by the re-accumu- lation of calcium by sarcoplasmic reticulum (Fig. 2).
A rapid increase in intracellular Ca 2+ concentra-
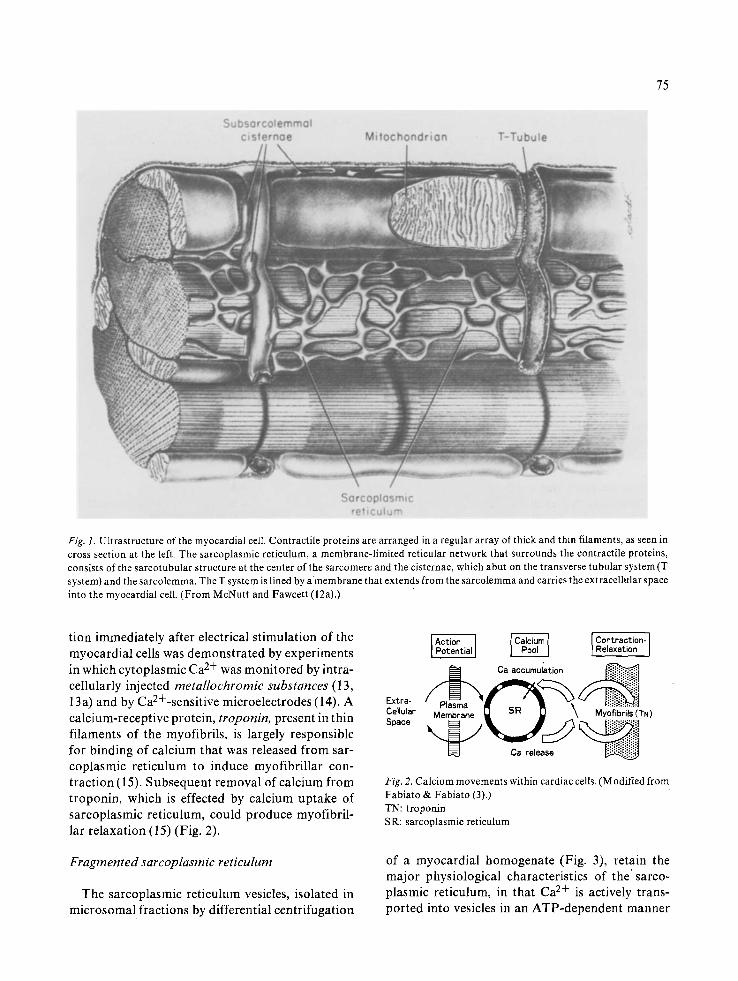
75
S JbSo~co~emmo~ c~sternae Mitocnond~ian T~Tubuie
Sorcoplosmic reticulurr,
Fig. 1. Ultrastructure of the myocardial cell. Contractile proteins are arranged in a regular array of thick and thin filaments, as seen in cross section at the left. The sarcoplasmic reticulum, a membrane-l imited reticular network that surrounds the contractile proteins, consists of the sarcotubular structure at the center of the sarcomere and the cisternae, which abut on the transverse tubular system (T system) and the sarcolemma. The T system is lined by a membrane that extends from the sarcolemma and carries the extracellular space into the myocardial cell. (From McNutt and Fawcett (12a).)
tion immediately after electrical stimulation of the myocardial cells was demonstrated by experiments in which cytoplasmic Ca 2+ was monitored by intra- cellularly injected metallochromic substances (13, 13 a) and by Ca2+-sensitive microelectrodes (14). A calcium-receptive protein, troponin, present in thin filaments of the myofibrils, is largely responsible for binding of calcium that was released from sar- coplasmic reticulum to induce myofibrillar con- traction (15). Subsequent removal of calcium from troponin, which is effected by calcium uptake of sarcoplasmic reticulum, could produce myofibril- lar relaxation (15) (Fig. 2).
Fragmented sarcoplasmic reticulum
The sarcoplasmic reticulum vesicles, isolated in microsomal fractions by differential centrifugation
Extra- Cellular Space
~ Contraction- Relaxation I
Membrane ~ SR ~ \ Myofibrils (TN)
~ L rele2se[~ ~I Fig. 2. Calcium movements within cardiac cells. (Modified from Fabiato & Fabiato (3).) TN: troponin SR: sarcoplasmic retieulum
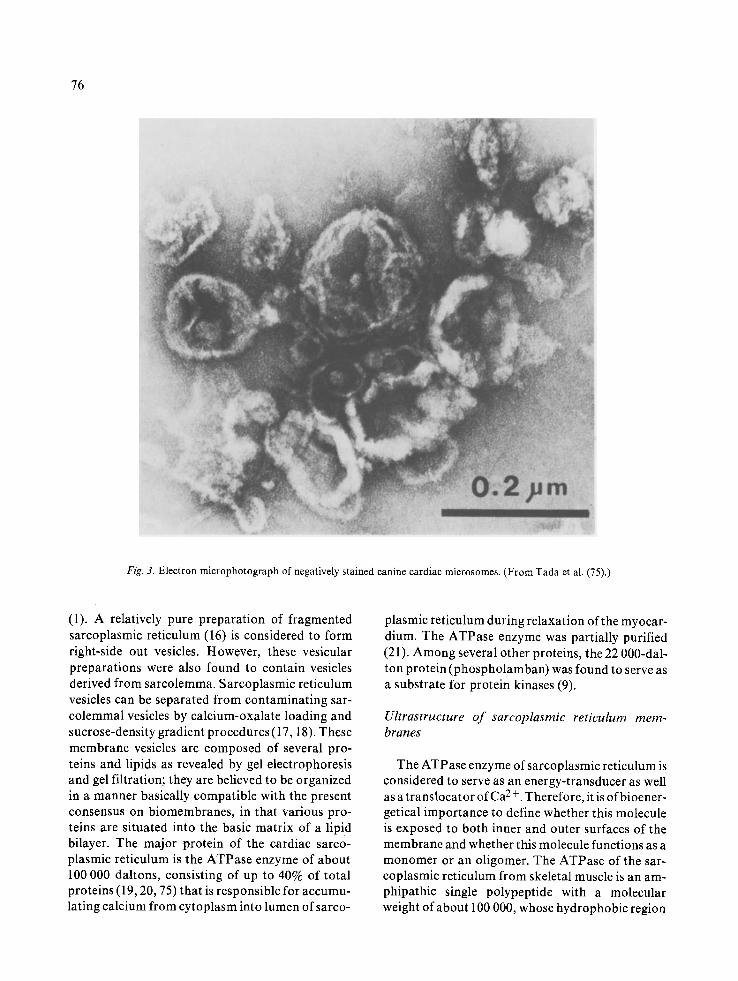
of a myocardial homogenate (Fig. 3), retain the major physiological characteristics of the' sarco- plasmic reticulum, in that Ca 2+ is actively trans- ported into vesicles in an ATP-dependent manner
76
Fig. 3. Electron microphotograph of negatively stained canine cardiac microsomes. (From Tada et al. (75).)
(1). A relatively pure preparation of fragmented sarcoplasmic reticulum (16) is considered to form right-side out vesicles. However, these vesicular preparations were also found to contain vesicles derived from sarcolemma. Sarcoplasmic reticulum vesicles can be separated from contaminating sar- colemmal vesicles by calcium-oxalate loading and sucrose-density gradient procedures (17, 18). These membrane vesicles are composed of several pro- teins and lipids as revealed by gel electrophoresis and gel filtration; they are believed to be organized in a manner basically compatible with the present consensus on biomembranes, in that various pro- teins are situated into the basic matrix of a lipid bilayer. The major protein of the cardiac sarco- plasmic reticulum is the ATPase enzyme of about 100 000 daltons, consisting of up to 40% of total proteins (19, 20, 75) that is responsible for accumu- lating calcium from cytoplasm into lumen of sarco-
plasmic reticulum during relaxation of the myocar- dium. The ATPase enzyme was partially purified (21). Among several other proteins, the 22 000-dal- ton protein (phospholamban) was found to serve as a substrate for protein kinases (9).
Ultrastructure o f sareoplasmic reticulum mem- branes
The ATPase enzyme of sarcoplasmic reticulum is considered to serve as an energy-transducer as well as a translocator of Ca 2+. Therefore, it is ofbioener- getical importance to define whether this molecule is exposed to both inner and outer surfaces of the membrane and whether this molecule functions as a monomer or an oligomer. The ATPase of the sar- coplasmic reticulum from skeletal muscle is an am- phipathic single polypeptide with a molecular weight of about 100 000, whose hydrophobie region

77
is in direct contact with the membrane lipid; part of its hydrophilic region is exposed to the outer sur- face of the membrane (1, 22).
X-ray diffraction and freeze-fracture studies of sarcoplasmic reticulum vesicles indicated that pro- teins are asymmetrically distributed across the membrane, the majority being contained in the cy- toplasmic leaflet (1, 23). The 9-nm particles found to be abundantly present in the cytoplasmic frac- ture faces were proposed, without direct evidence, to represent the structural feature of the transport ATPase. Although it is quite likely that the particles represent distinct intramembranous structures, rather than artifacts, it is not known whether each of these particles represents a mononer or an olig- omer.
These studies on the ultrastructural characteris- tics of the sarcoplasmic reticulum membranes sug- gest that the main body of the globular structure, which presumably represents part of the ATPase, is localized largely in the cytoplasmic leaflet of the lipid bilayer and that part of this structure is ex- posed to the external surface of the membrane. However, there is not enough evidence to support the view that the ATPase molecule spans the mem- brane and that a portion is exposed to the internal surface.
Proteolytic subfragmentation of the ATPase permitted to gain insights into intramolecular dis- tribution of the key functional features of the calci- um transport ATPase (1, 24). Two fragments with molecular weights of 52 000-60 000 and45 000-55 000 were obtained when sarcoplasmic reticulum vesi-
cles were briefly trypsinized under specific condi- tions (Table I). The larger subfragment (fragment I) contained the phosphorylation site (active site of the ATPase enzyme) and probably the calcium- binding sites. The smaller subfragment (fragment II), which did not contain these activities, was more hydrophobic and was assumed to largely represent the intramembranous structure seen in freeze-frac- ture studies. In contrast, fragment I is considered to largely represent the hydrophilic portion of the ATPase which exposes to the outer surface of the membrane. Further proteolysis of fragment I yielded subfragments of 30000-33000 daitons (fragment Ia) and 20 000-24 000 daltons (fragment Ib). Fragment Ia contained the phosphorylation site and fragment Ib contained calcium-binding sites. The use of antibodies against each fragment indicated that fragment Ia is externally located, whereas fragment Ib is only partially exposed. It is interesting to note that the transport state of the ATPase could determine the extent to which frag- ment Ib is exposed at the outer surface, suggesting that fragment Ib, possibly serving as calcium-bind- ing moiety, undergoes a profound shift of location from outer to inner portions of the membrane when the ATPase performs translocation of calcium (1, 77).
Active calcium transport by sarcoplasmic reticu- lum
During calcium transport the carrier protein (ATPase) must play at least three roles in a succes-
Table 1. Molecular weights of sarcoplasmic reticulum ATPase and its tryptic subfragments.
ATPase Primary degradation Secondary degradation
Fragment I Fragment II Fragment Ia Fragment Ib Reference
115 000 60 000 55 000 33 000 24 000 102 000 55 000 45 000 30 000 20 000
100 000 60 000 49 000 106 000 57 000 46 000 100 000 52 000 49 000 120 000" 65 000" 56 000" 115 000 or 56 000t 59 000t l19000t
32 000 21 000
Thorley-Lawson and Green (25, 26) Stewart, MacLennan, and co- workers (27-29) Yamamoto and Tonomura (30) Inesi and Scales (31) Louis et al. (32, 33) Yu et al. (34) Rizzolo et al. (35) and le Make et al. (36)
Values were determined by electrophoresis on SDS-polyacrylamide gels in all reports except those by Rizzolo et al. and le Maire et al. * ATPase protein labeled with diazotized diiodosulfanilic acid was subjected to subfragmentation. "~ Determined by sedimentation equilibrium measurements in SDS (35) or in deoxycholate (36). (From Tada et al. (1).)

78
sive manner: it has to recognize, translocate, and release calcium ions. In initiating transport, the carrier protein has to bind the particular ion by recognizing it from among many kinds of solutes that exist within the system (1). This role is presum- ably assumed by the high-affinity calcium binding (K D : 0.3-1.5 ~M) which was shown in the absence of substrate (ATP) to exist in sarcoplasmic reticu- lum vesicles as well as in purified ATPase prepara- tion (1). Such calcium binding should be followed by the translocation of calcium across the mem- brane, which requires an energy expenditure to in- duce conformational alteration of the carrier pro- tein. This property of ATPase will be detailed below. The affinity of the carrier for calcium thus changes and calcium is released from the carrier into internal lumen of the membrane vesicles. The system then returns to its original state.
A precise operational definition of the term translocation of cations (calcium) must be given in order to avoid confusion in assessing the experi- mental data. When calcium is incubated with mem- branes of sarcoplasmic reticulum, usually forming sealed vesicles, it is taken up by the membrane vesicles at the expenditure of energy (ATP), either by binding to the surface of the membrane or by actual uptake into the membrane vesicles. By oper- ational definition, calcium is considered to be actu- ally transported by the membrane when both of the following conditions are satisfied: a) calcium can- not be removed by the application, at the outer surface of vesicles, of the high concentration of a chelating agent ethylene glycol bis (/3-aminoethyl ether) N,N'-tetraacetic acid (EGTA), which by itself dose not penetrate the membrane; b) calcium can be readily removed by,the treatment of the membrane with calcium-specific ionophores (X-537A or A23187), detergents (Triton X-100) or alkali in the presence of EGTA. Such conditions would corre- spond either to an actual translocation of calcium from one side of the membrane to the other, or to an apparent translocation due to trapping of calcium inside the pocket formed by the carrier proteins.
Anions like oxalate were often included in the medium to enhance the amounts of calcium trans- ported into vesicles. The term oxalate-facilitated calcium uptake is often applied to describe these conditions. Oxalate is freely permeable to sarco- plamic reticulum membranes and could form a cal- cium-oxalate complex by binding with transported
calcium within the vesicular lumen, thus maintain- ing an intravesicular free Ca 2+ concentration and enabling more calcium to be transported. In the absence of oxalate most of the calcium is bound to the membrane, and only a small amount exists as the ionized form, thus leaving the calcium-binding capacity of the vesicles quite low. Phosphate could be substituted for oxalate, but the former is not always suitable for this purpose because it also enhances the reversal of calcium transport (see be- low).
ATP could serve to energize calcium uptake by sarcoplasmic reticulum, by inducing changes in conformation of ATPase enzyme. Thus, ATP is hydrolyzed when calcium is translocated across sarcoplasmic reticulum membrane. In intact vesi- cles, a tight coupling was found to exist between calcium and ATP, giving 2 mol of Ca 2+ transported for 1 mol of ATP hydrolyzed under a variety of conditions in the presence and absence of oxalate (1). In accord with these findings, 1 mol of ATPase enzyme was shown to contain approximately 2 mol of calcium-binding sites and I mol of ATP binding site. The stoichiometry was maintained unless the membrane vesicles became leaky, or unless the reaction conditions were thermodynamically unfa- vorable for the transport system.
Two classes of phosphoproteins in cardiac sarco- plasmic reticulum
Cardiac sarcoplasmic reticulum was found to form two classes of phosphoproteins. They differ in the molecular weights and their chemical character- istics are distinctly different (37, 38) (Table 2). The ATPase enzyme of cardiac sarcoplasmic reticulum, serving as an energy-transducer and a translocator of calcium across the membrane, forms a phospho- protein intermediate, in that the terminal phos- phate of ATP is incorporated into the ATPase en- zyme. The phosphoprotein formed is an acyl- phosphate which is labile in hydroxylamine (1, 38). In skeletal muscle, sarcoplasmic reticulum, such phosphorylation occurs at carboxyl group of aspar- tic acid residue(l). The amount of phosphorylation in cardiac sarcoplasmic reticulum varies from one preparation to the other, ranging from 0.6 to 1.3 nmoles P / m g microsomes in the presence of optim- al concentrations of ATP and Ca 2+ (20, 38-40).

Table 2. Comparison of the two phosphoproteins formed in the cardiac sarcoplasmic reticulum.
79
Parameter Phospholamban ATPase
1) Molecular Weight 2) P-amino acid 3) Nature of phosphoprotein
4) Amount of phosphorylation 5) Dependency of phospborylation on Ca 2+ 6) Sensitivity to trypsin 7) Physiological function
22 000 Serine Phosphoester (stable in hot acid and in hydroxylamine) 0.7-1.3 nmoles/mg Independent of Ca 2+* Labile to trypsin Substrate of cyclic AMP-dependent protein kinase Modulator of Ca2+-transport ATPase
100 000 Aspartic acid Acyl phosphate (labile in hydroxyl- amine 0.6 1.3 nmoles/nag Dependent on Ca 2+ Relatively resistant to trypsin Transports Ca 2+, which is directly coupled to the contraction-relax- ation cycle
* In the presence of calmodulin, the phosphorylation of phospholamban exhibits a marked dependency on Ca 2+ (See text for explanation). (From Yada et al. (37).)
Phosphorylat ion probably occurs at one of the tryptic subfragments of the ATPase, whose mo!ecu- lar weight is 30 000-33 000 daltons (fragment Ia).
Cardiac sarcoplasmic reticulum was found to form another phosphoprotein when reacted with a cyclic AMP-dependent protein kinase. The termi- nal phosphate of ATP is largely incorporated into a protein of 22 000 daltons (Fig. 4B). The phospho- protein formed exhibits stability characteristics of a phosphoester in which the phosphate is largely in- corporated into serine. This phosphorylatable pep- tide, presumably representing one of the major sites at which cyclic AMP-dependent protein kinase ex- erts its action, appears to function as a modulator of Ca2+-dependent ATPase and calcium pump of cardiac sarcoplasmic reticulum, thus referred to as phospholamban (see below).
These two phosphoproteins are interrelated, in that at phosphorylated state phospholamban could exert its effect on the ATPase enzyme by enhancing the turnover rate of the intermediary steps of the latter enzyme. In the following, characteristics of each of these two phosphorylation reactions are first documented before giving the detailed descrip- tion of their interrelationships in the subsequent sections.
Formation of A TPase intermediate
Active transport 0 fCa 2+ across the membrane of sarcoplasmic retieulum is an energy-requiring pro- cess mediated by a Ca 2+, Mg2+-dependent ATPase enzyme. Properties of this enzyme in the fragment-
ed sarcoplasmic reticulum have been extensively investigated, and the formation of the phosphory- lated intermediate EP was found to be critically important in this energy-transducing system. When the pre-steady state of Ca2+-dependent ATPase is followed by a rapid-mixing device in the presence of suboptimal concentration of ATP (5-50 #M) and Ca 2+ (1 14 #M), a reaction intermediate EP is rapidly formed, reaching maximal level within 50 msec at 20 °C (Fig. 5A). Essentially similar obser- vations were made in skeletal muscle sarcoplasmic reticulum (Fig. 5B). The liberation of Pi exhibits an initial lag phase, which coincides with the rapid increase in EP formation. Assuming that all of Pi is liberated from EP, subsequent increase in Pi libera- tion, which coincides with the completion of EP formation, is considered to represent the turnover of EP. These observations indicate that EP is the true intermediate of the ATPase reaction.
In understanding the mechanism by which the translocation of calcium across the membrane is coupled with the ATP hydrolysis, it is of a prime necessity to analyse kinetic properties of partial reactions constituting the overall ATPase reaction. It is also essential to consider the vectorial proper- ties of the actions of this enzyme; localization of the sites of action of substrates, products, and per- meates should always be distinguished during the active transport of cations. Based on the kinetic analyses of the formation of the phosphoenzyme and the liberation of products Pi and ADP under variety of conditions, the following reaction scheme was assumed to represent the mechanism of cou-
80
I I I I
2 A [
0
2o
o
nr
' 1 0 0 10 2 0 3 0
G E L SL ICE N U M B E R
2 6
2 0
15
10
5
0
i i
C
I I
I '
0 ' I() '
G E L SL ICE
Ill
i o '
N U M B E R
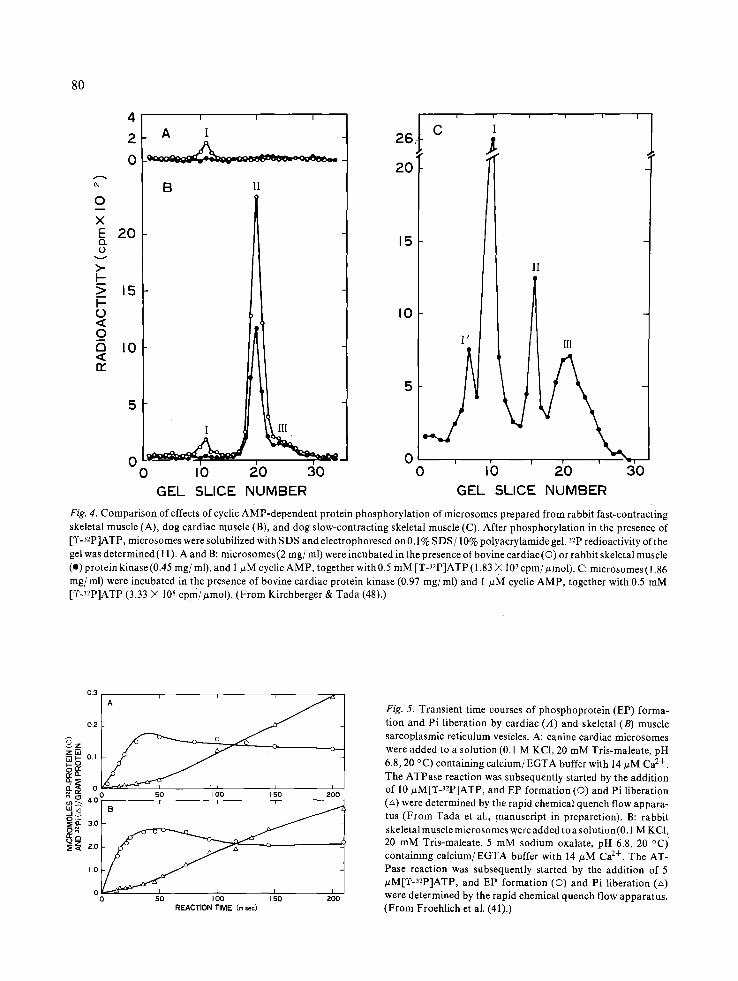
Fig. 4. Compariso n of effects of cyclic AMP-dependent protein phosphorylation of microsomes prepared from rabbit fast-contracting skeletal muscle (A), dog cardiac muscle (B), and dog slow-contracting skeletal muscle (C). After phosphorylation in the presence of [T-32p]ATP, microsomes were solubilized with SDS and electrophoresed on 0.1% SDS/10% polyacrylamide gel. 32p redioactivity of the gel was determined (11). A and B: microsomes (2 mg/ml) were incubated in the presence of bovine cardiac (©) or rabbit skeletal muscle (e) protein kinase (0.45 mg/ml), and 1 ~zM cyclic AMP, together with 0.5 mM [T-32p]ATP ( 1.83 >( 107 cpm/#mol). C: microsomes ( 1.86 mg/ml) were incubated in the presence of bovine cardiac protein kinase (0.97 rag/ml) and 1 #M cyclic AMP, together with 0.5 mM [TJ2P]ATP (3.33 × 108 cPm/#mol ). (From Kirchberger & Tada (48).)
0 .3
~_z o z w [] ~ O. i-o
5 0 I On 150 2 0 0 m ~ 4".0 B , , ,
o~h- 3.0
0 I t 5 0 I O0 150 2 0 0
R E A C T I O N T I M E (m see)
Fig. 5. Transient time courses of phosphoprotein (EP) forma- tion and Pi liberation by cardiac (A) and skeletal (B) muscle sarcoplasmic reticulum vesicles. A: canine cardiac microsomes were added to a solution (0.1 M KCI, 20 mM Tris-maleate, pH 6.8, 20 °C) containing calcium/EGTA buffer with 14 #M Ca 2+. The ATPase reaction was subsequently started by the addition of 10 #M[TJ2P]ATP, and EP formation (0) and Pi liberation (~) were determined by the rapid chemical quench flow appara- tus (From Tada et al., manuscript in preparetion). B: rabbit skeletal muscle microsomes were added to a solution(0.1 M KC1, 20 mM Tris-maleate, 5 mM sodium oxalate, pH 6.8, 20 °C) containing calcium/EGTA buffer with 14/zM Ca 2+. The AT- Pase reaction was subsequently started by the addition of 5 #M[q~J2p]ATP, and EP formation (O) and Pi liberation (z~) were determined by the rapid chemical quench flow apparatus. (From Froehlich et al. (41).)

81
piing between the ATPase and calcium transport (1):
E + ATP ° + 2Ca ° ~ ~Ca~ ~ EpCa~ + ADpO - ~ATP '7
E + ADP ° + Pi ° + 2Ca i (1)
where i and 0 indicate the inside and outside of the membrane vesicles, respectively. Two moles of Ca 2+ and 1 mol of ATP bind in a random sequence to 1 mol of the ATPase enzyme, E, at the outer surface of the membrane, forming the Michaelis
ECa~P ' , . 1 Calcium is subsequently translo- complex cared from outside to inside the membrane when the phosphorylated intermediate ECa2 ~ (simply de- signated as EP) is formed. Calcium is then released from the enzyme into the interior of the vesicles, with simultaneous decomposition of the phospho- rylated intermediate.
It should be noted that the ATPase enzyme was found to undergo distinct conformational changes during the transport cycle, in that both E and EP could form two different conformations, that ex- hibit different affinities for Ca 2+. The substrate- free enzyme was proposed to exhibit two different forms; a form of the enzyme having low affinity for Ca 2+ (E2) is transformed to another form of the enzyme having high affinity for Ca 2+ (E~), that allows Ca 2+ to bind (Equation 2). The step E2~E l is rate-determining. In analogy to the substrate-free enzyme, the phosphorylated intermediate EP was also shown to exhibit two forms, E~P and E2P, having high and low affinities for Ca 2+, respective- ly. Taking these findings into consideration, prob- able reaction mechanism of ATPase is summarized in the following equation (9):
The Michaelis complex Ca°E~ATP formed at the outside of membranes is immediately converted in- to E, type of phosphorylated intermediate (CaiE1--P), followed by the subsequent conversion into E 2 type of phosphorylated intermediate (MgE2--P). Calcium is translocated from outside to inside the membrane, when C a i E ~ P is formed. Calcium is released within the interior of vesicles when CaiE1--P is converted to MgE2--P. The in- termediate EP thus represents the sum of EIP and E2P. Among these intermediary steps, the step at which E 2 is converted to E 1 (Step i) and the step at which E~P is decomposed (Step iv) are the two major rate-determining steps, in the presence of saturating concentrations of Ca 2+, ATP and alkali metal salts. Upon addition o fADP, EIP could form E~ATP via reversal of Step iii, thus termed as ADP- sensitive intermediate, while E2P could not form E IATP upon addition of ADP, thus termed as ATP- insensitive intermediate. Shigekawa et al. (42, 43) found that EP decomposition was accelerated by alkali metal salts and free divalent cations (Mg 2+, Ca2+). In the presence of alkali metal salts the decomposition of E2P (Step v) is accelerated and conversion from E~P to E2P (Step iv) is rate-deter- mining, whereas in the absence of alkali metal salts the step at which E2P is decomposed (Step v) is rate- determining.
Most of the sequences in the ATPase reaction were found to be reversible. Thus the synthesis of ATP by the backward reaction is coupled with the reversal of calcium transport (1, 44). Under these conditions EP can be formed by the reaction of Pi with E both in the presence and absence of Ca 2+ concentration gradient across the membrane. Tran-
Ca°E~
Mg
MgE2
ATP o
(ii)
(vi)
pi o
Ca°EIATP
MgE2-Pi .
ADP o ;
) HOH
. " CaiEl~p (iii)
~ C a i
(iv) ~ Mg
(v) ", MgE2~ P
(2)
82
sient state kinetic studies of the reverse reaction under specified conditions allowed to determine the rate of EP formation when substrate-free enzymes are at two different states E 1 and E 2. The rate of EP formation from E 2 + Pi is much faster than that from E 1 + Pi since the step at which E 1 is converted to E 2 is rate-determining.
Phosphorylation of 22 O00-dalton protein (Phos- pholamban)
A phosphoester phosphoprotein was formed when cardiac sarcoplasmic reticulum was incubat- ed with cyclic AMP in the absence (45, 46) and presence (47) of cyclic AMP-dependent protein ki- nase. Formation of this phosphoprotein was inde- pendent of Ca 2+ up to 0.1 mM, but was markedly dependent on cyclic AMP between 0.1 and 10/~M with the half-maximal activation occurring at 0.2 #M (47). The phosphoprotein formed was stable in hydroxylamine and in hot acid, but was unstable in hot alkali (Table 3). Tada et al. (11) found that a single microsomal protein of approximately 22 000 daltons bound most of the 32p label when cardiac microsomes incubated with [T-32p]ATP and cyclic AMP-dependent protein kinase were solubilized in sodium dodecyl sulfate (SDS) and electrophoresed on SDS-polyacrylamide gel (Fig. 4B). In the pres- ence of NaF, inhibitor of phosphoprotein phospha- tase, phosphorylation of this protein increased sev-
Table3. Comparison of stability of two kinds of phosphoproteins formed in cardiac microsomes.
Treatment Amount of phospho- Amount of phoshoryl- protein intermediate ated phospholamban of ATPase (22 000-dalton protein)
nmolP/mg % nmolP/mg % protein protein
None; 0 °C (= control) 0.74 100 0.67 100 None; 90 °C 0.00 0 0.62 92.5 0.5 N NaOH; 25 °C 0.00 0 0.31 46.3 0.5 N NaOH; 90 °C 0.00 0 <0.01 <1.0 Control (0.8 M NaC1); 0.71 100 0.61 I00 30 °C
0.8 M hydroxylamine; <0.01 <1.0 0.60 98.4 30 °C
(From Tada et al. (38).)
eral fold, and the other two proteins of 55 000 and 11 000 daltons were also phosphorylated to a lesser extent. The 55 000-dalton phosphoprotein was at- tributed to autophosphorylation of the regulatory subunits of protein kinase. These findings were vir- tually in agreement with those by La Raia & Mor- kin (45) who also found phosphorylation of 20 000- dalton microsomal protein, catalyzed by an endogenous protein kinase. These reports were subsequently followed by a number of evidence (Table 4), supporting that the protein of 20 000- 24 000 daltons is the major site at which cyclic AMP-dependent protein kinase could react.
Table 4. Molecular weights of proteins phosphorylated by cyclic AMP-dependent protein kinase in cardiac sarcoplasmic reticulum.
Investigators Year Animals
Other P hospholamban phosphoprotein
Molecular weight Molecular weight (1000 daltons) (1000 daltons)
LaRaia & Morkin (45) 1974 Rabbit 20K Tada et al. (11) 1975 Dog 22K IlK Kirchberger & Tada (48) 1976 Dog, cat, guinea pig 22K 11K
rabbit Schwartz et al. (49) 1976 Dog 20K Wray & Gray (50) 1977 Dog 20K Will et al (51) 1978 Pigeon 22K 15K St. Louis & Sulakhe (52) 1979 Pig 22K 56K 16K Jones et al. (18) 1979 Dog 20K 7K Lamers & Stinis (53) 1980 Rat 24K 9K Bidlack & Shamoo (54) 1980 Dog 22K 6K Le Peuch et al. (55) 1980 Dog 22K IlK 7K Tada et al. (56) 1982 Dog 22K 11K
(From Tada & Katz (9).)

Since the 22 000-dalton protein was associated with functional alterations in sarcoplasmic reticu- lum (see below), it was termed as phospholamban, i.e. phosphate receptor (9, 11, 57), (,ka#j3auetu = receive).
Existence of phospholamban phosphorylation was confirmed in sarcoplasmic reticulum prepara- tions virtually freed from sarcolemmal vesicles by means of density gradient centrifugations (18, 51, 53) (Table 4). The reported molecular weight for phospholamban ranged between 20 000 and 24 000 daltons. Under the same conditions other proteins with lower molecular weights were also phosphory- lated. Within the range below 10 000 daltons, a phosphoprotein of 6 000 (54, 58) or 7 000 (18, 55) daltons was found. Chemical characteristics of this protein were distinct from phospholamban. Anoth- er phosphoprotein of larger molecular weight, 15 000 (51) or 16 000 (52) daltons, was noted in some reports. Phosphorylation of this protein was reported to occur at the threonine residue (51), while that of 22 000-dalton phospholamban at the serine residue (47, 51, 55). A phosphoprotein of 11 000 daltons, appearing under a specified condi- tions, is attributable to a putative monomer of phospholamban (see below). Phospholamban was also found to be phosphorylated by calmodulin- dependent protein kinase (see below).
Cyclic AMP regulation of calcium transport
A wide variety of proteins were found to be phosphorylated by a cyclic AMP-dependent pro- tein kinase. However, functional alterations due to such phosphorylation were documented in only a few proteins (59). In cardiac sarcoplasmic reticu- lure, virtually a single polypeptide (phospholam- ban) was shown to undergo such phosphorylation, and resultant alterations in membrane function, i.e. active calcium transport, were well correlated with cyclic AMP-dependent phosphorylation. A line of evidence indicate that phospholamban could serve as a regulator controlling the active calcium trans- port.
Active calcium transport
ATP-supported active calcium transport by membrane vesicles of sarcoplasmic reticulum was
83
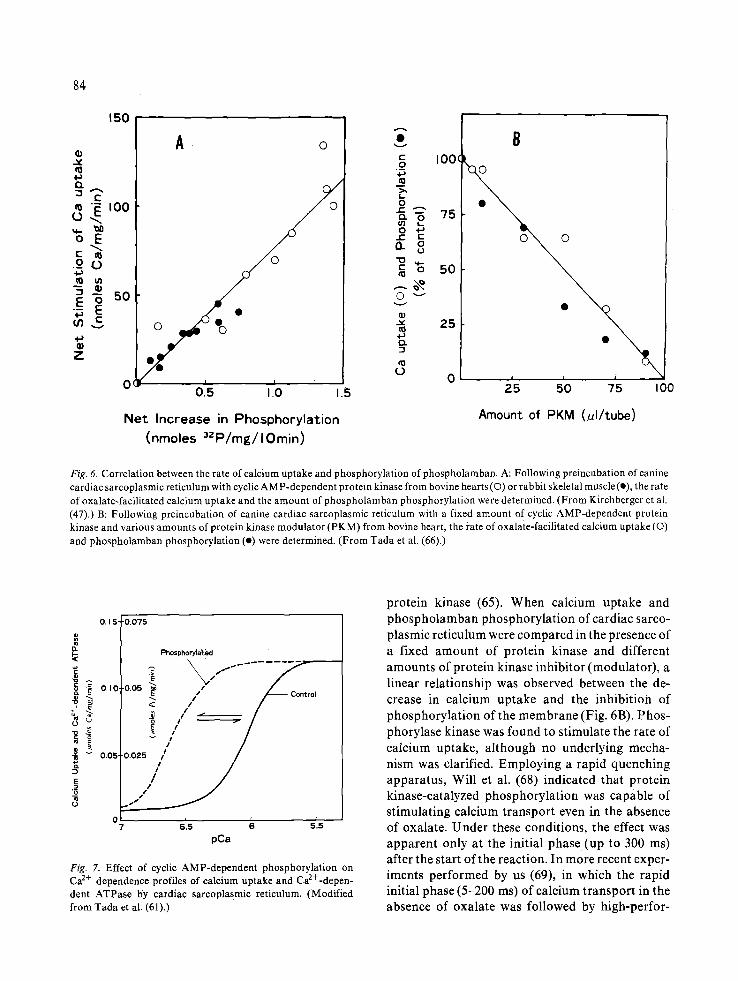
found to be markedly augmented when membrane vesicles were previously phospyorylated by incuba- tion with cyclic AMP-dependent protein kinase (47, 57, 60, 61). Under these conditions, where phospholamban within the membrane of cardiac sarcoplasmic reticulum was specifically phosphory- lated, the rate of oxalate-facilitated calcium uptake was more than doubled. Calcium uptake by sarco- plasmic reticulum from fast-contracting skeletal muscle did not exhibit such stimulation (48), nor was there significant phosphorylation of similar protein (Fig. 4A). The stimulation of calcium up- take by cardiac sarcoplasmic reticulum was well correlated with the increase in phospholamban phosphorylation (47, 62), but not with the 11 000- dalton phosphoprotein (62). Protein kinase ob- tained from cardiac muscle (type II kinase) effec- tively produced such stimulation, while that obtained from skeletal muscle (type I kinase) was also functional, though to a much lesser extent (Fig. 6A). Other reports also indicated the existence of such stimulation caused by endogenous and exo- genous protein kinases (45, 49, 63). When the rate of calcium uptake by cardiac sarcoplasmic reticu- lum was examined at different Ca 2+ concentrations in the presence and absence of cyclic AMP-depen- dent phosphorylation, the rate of calcium uptake by cardiac sarcoplasmic reticulum in the absence of cyclic AMP-dependent phosphorylation exhibited half-maximal activation at around 1 # M Ca 2+, whe- reas those in the presence of cyclic AMP-dependent phosphorylation of phospholamban exhibited half- maximal activation at lower Ca 2+ concentrations (--0.3 #M) (Fig. 7). These results suggest that cyclic AMP-dependent phosphorylation of phospholam- ban may increase an apparent affinity of calcium transport system(Ca2+-dependent ATPase) of car- diac sarcoplasmic reticulum for Ca 2+. These effects of protein kinase on calcium transport of cardiac sarcoplasmic reticulum were confirmed by treat- ment of cardiac sarcoplasmic reticulum with phos- phoprotein phosphatase (64, 65) or protein kinase modulator (66, 67), which can antagonize the effect of cyclic AMP-dependent protein kinase. When phosphorylated sarcoplasmic reticulum was incu- bated with phosphoprotein phosphatase obtained from bovine heart, most of the phosphorylated pro- tein (phospholamban) was dephosphorylated (64). Resultant dephosphorylation of the membrane led to complete reversal of the effects produced by
84
150
v
4~ n
cO £) ~ I00
o E
• £ 0
~J
Z
A ¸ o
I
0.5 1.10 1.5
Net Increase in Phosphorylation (nmoles 3ZP/mg/lOmin)
v
c o
o I- Q.
© v
n
0
c o o
,oO oo 25 e ~
i 5'0 0 25 75 I00
Amount of PKM (ul/tube)
Fig. 61 Correlation between the rate of calcium uptake and phosphorylation of phospholamban, A: Following preincubation of canine cardiac sarcoplasmic reticulum with cyclic AMP-dependent protein kinase from bovine hearts (©) or rabbit skeletal muscle (I), the rate of oxalate-facilitated calcium uptake and the amount of phospholamban phosphorylation were determined. (From Kirchberger et al. (47).) B: Following preincubation of canine cardiac sarcoplasmic reticulum with a fixed amount of cyclic AMP-dependent protein kinase and various amounts of protein kinase modulator (PKM) from bovine heart, the rate of oxalate-facilitated calcium uptake (©) and phospholamban phosphorylation (e) were determined. (From Tada et al. (66).)
0.15-
i ~ 0,,0.
+1 E
~ ~ 0.05-
I
-0.075
0 7 615
Phosphoryla~.~l
.o.o / " oo<,o.
/ / / /
0.025 i i
pCa
Fig. 7. Effect of cyclic AMP-dependent phosphorylation on Ca 2+ dependence profiles of calcium uptake and Ca2+-depen- dent ATPase by cardiac sarcoplasmic reticulum. (Modified from Tada et al. (61).)
protein kinase (65). When calcium uptake and phospholamban phosphorylation of cardiac sarco- plasmic reticulum were compared in the presence of a fixed amount of protein kinase and different amounts of protein kinase inhibitor (modulator), a linear relationship was observed between the de- crease in calcium uptake and the inhibitioia of phosphorylation of the membrane (Fig. 6B). Phos- phorylase kinase was found to stimulate the rate of calcium uptake, although no underlying mecha- nism was clarified. Employing a rapid quenching apparatus, Will et al. (68) indicated that protein kinase-catalyzed phosphorylation was capable of stimulating calcium transport even in the absence of oxalate. Under these conditions, the effect was apparent only at the initial phase(up to 300 ms) after the start of the reaction. In more recent exper- iments performed by us (69), in which the rapid initial phase (5-200 ms) of calcium transport in the absence of oxalate was followed by high-perfor-
Ca2+-dependent A TPase
85
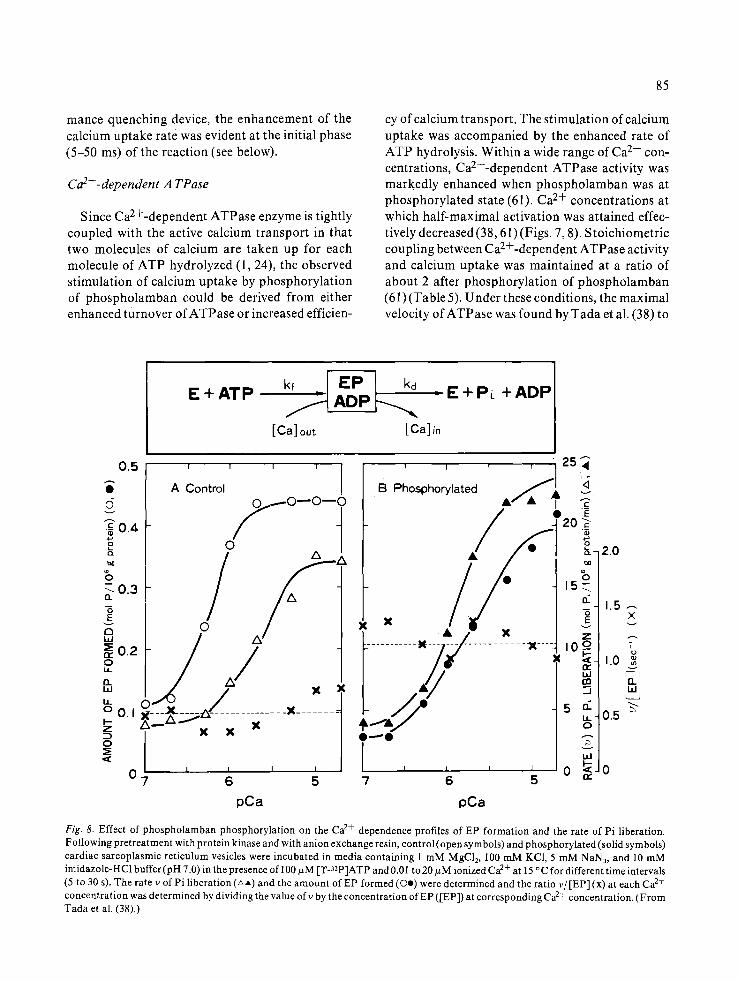
Since Ca2+-dependent ATPase enzyme is tightly coupled with the active calcium transport in that two molecules of calcium are taken up for each molecule of ATP hydrolyzed (1, 24), the observed stimulation of calcium uptake by phosphorylation of phospholamban could be derived from either enhanced turnover of ATPase or increased efficien-
E + A T P
cy of calcium transport. The stimulation of calcium uptake was accompanied by the enhanced rate of ATP hydrolysis. Within a wide range of Ca 2+ con- centrations, Ca2+-dependent ATPase activity was markedly enhanced when phospholamban was at phosphorylated state (61). Ca 2+ concentrations at which half-maximal activation was attained effec- tively decreased (38, 61) (Figs. 7, 8). Stoichiometric coupling between Ca2+-dependent ATPase activity and calcium uptake was maintained at a ratio of about 2 after phosphorylation of phospholamban (61) (Table 5). Under these conditions, the maximal velocity of ATPase was found by Tada et al. (38) to
(5
2 Q .
I1
E v
0 W
o LL n h i
h 0 I-- Z
0
kf kd - ! A D P ' - E + ADP ~ , _ A D P ~ , ~ , _ _ +P~
[Ca]out [Ca]in
0.1
0 .5 I I I I I
I A Control 0 / 0 ~ 0 - - 0
O.4 / 0
0.3
0 .2
0 "7 6 5
mance quenching device, the enhancement of the calcium uptake rate was evident at the initial phase (5-50 ms) of the reaction (see below).
, , , 25~"
B Phosphorylated • / A --~
t-----: ..... - / / - .......... .---:.. -l.o i. I ' "
7 6 5 re
pCa pCa
2.0
X
i
1.0 g 03 v
fl_ l.d
0.5
Fig. 8. Effect of phospholamban phosphorylat ion on the Ca 2+ dependence profiles of EP formation and the rate of Pi liberation. Following pretreatment with protein kinase and with anion exchange resin, control (open symbols) and phosphorylated (solid symbols) cardiac sarcoplasmic ret iculum vesicles were incubated in media containing 1 m M MgC12, 100 mM KC1, 5 mM NAN3, and 10 mM imidazole-HC1 buffer (pH 7.0) in the presence of 100 ~zM [T-32p]ATP and 0.0 ! to 20 ~M ionized Ca 2+ at 15 ° C for different time intervals (5 to 30 s). The rate v of Pi liberation (AA) and the amount of EP formed (Oe) were determined and the ratio v~ [EP] (x) at each Ca 2+ concentrat ion was determined by dividing the value ofv by the concentration o fEP ([EP]) at corresponding Ca 2+ concentration. (From Tada et al. (38).)

86
Table 5. Effects of protein kinase on stoichiometric relationship between calcium uptake and Ca2+-dependent ATPase activity of canine cardiac microsomes.
Experiment Pretreatment Calcium uptake Ca2+-dependent S toichiometric ATPase activity ratio (Ca:Pi)*
A nmoles Ca/ min/ mg nmoles Pi/ min/ mg
Control 61 33 1.9 Protein kinase 92 52 1.8 Protein kinase + cyclic AMP 143 68 2.1 Control 81 50 1.6 Protein kinase 126 71 1.8 Protein kinase + cyclic AMP 167 103 1.6
* Calculated by dividing the rate of calcium uptake by the rate of CaZ+-dependent ATPase. (From Tada et al. (61).)
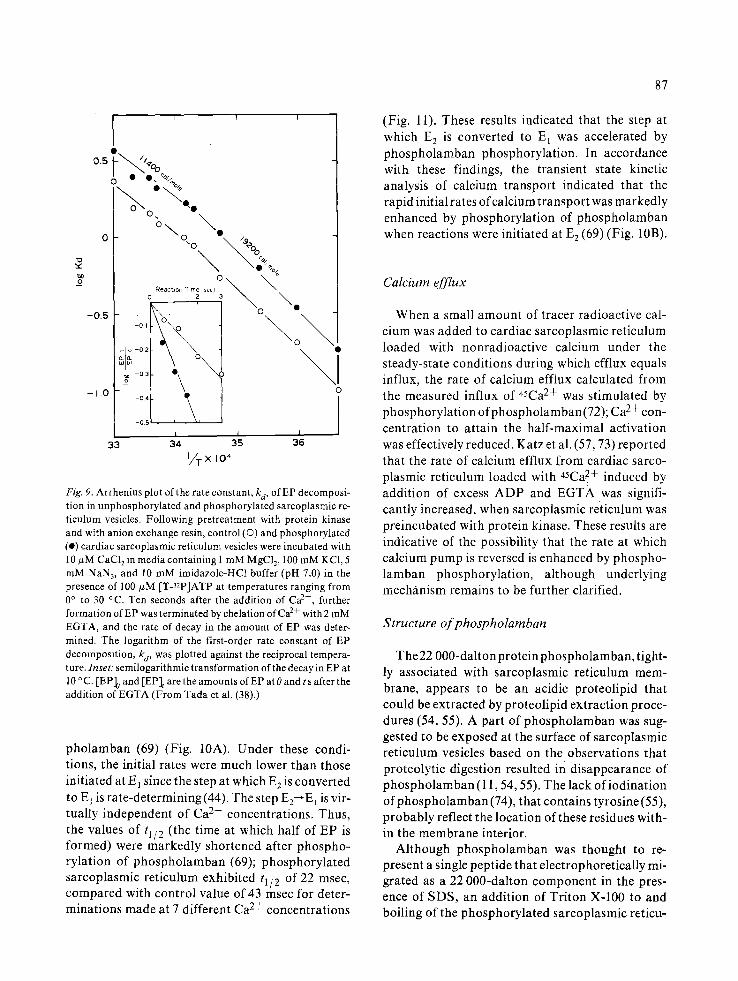
be markedly increased. Wray & Gray (50) also found augmentation of Ca2+-dependent ATP hy- drolysis by endogenous protein kinase. It is likely, therefore, that the turnover rate of a normally coupled transport is enhanced when phospholam- ban is at phosphorylated state. During the translo- cation of Ca 2+, the ATPase enzyme undergoes a complex series of reactions in which a number of intermediates are formed (see Equation 2). Essen- tially operational are the following two key reac- tions: the steps at which EP is formed and decom- posed (Equation I). Therefore, these steps were examined by special means under conditions where phospholamban is phosphorylated. Steady-state levels of EP were not altered by phospholamban phosphorylation at saturating concentrations of Ca 2+ and ATP (37, 38). However, lowering the concentrations of Ca 2+ (<10 #M) and ATP (<5 #M) resulted in decreased levels of EP when phos- pholamban phosphorylation was operational (38, 69). At Ca 2+ concentrations below 10/~M, phos- phorylation of phospholamban brought reduction in EP while the rate of Pi liberation(v) was elevated (Fig. 8). Thus, the ratio v/[EP] (the rate of Pi liberation per unit of EP concentration), which was independent of Ca 2+ (0.1-10 #M), was doubled by phospholamban phosphorylation (Fig. 8), suggest- ing that the rate of EP decomposition is markedly enhanced (38). These results are consistent with the findings that the rate constant, kd, of decay in EP was enhanced (38, 70) when phospholamban was at phosphorylated state. Under these conditions, the k d value was markedly elevated within a wide range of the temperature between 0 and 30 °C (38). The Arrhenius plots for k d of both control and phos-
phorylated sarcoplasmic reticulum indicated dif- ferent slopes below and above 18 o C. The activation energies remained constant after phosphorylation (11.4 and 19.2 kca l /mol at high and low tempera- tures, respectively), while free energy markedly in- creased by phosphorylation of phospholamban (Fig. 9). At ATP concentrations below 5 #M, EP levels were also lowered by phosphorylation of phospholamban (69), suggesting that phospholam- ban phosphorylat ion can enhance the rate of EP decomposit ion relative to that of EP formation when the latter is inhibited at low ATP concentra- tions.
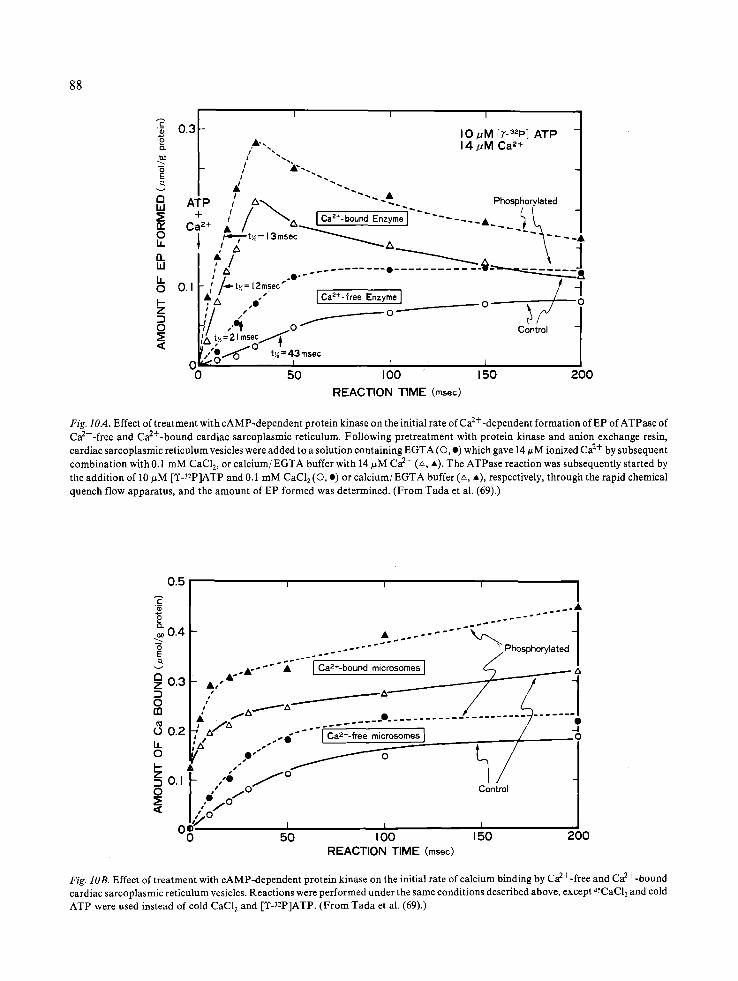
The steps at which EP is formed are extremely rapid processes, taking place within tens of millise- conds (69). Employing a high-performance rapid mixing device (71), the rate of EP formation was determined under conditions where the reactions were initiated at two different states of the ATPase enzyme. State E 1 represents the form of enzyme that binds calcium (Equation 2) when sarcoplasmic reticulum is incubated with C a / E G T A buffer con- taining micromolar concentration of ionized Ca 2+, while E 2 represents another form of enzyme that does not bind calcium (Equation 2) when sarco- plasmic reticulum is incubated with EGTA (69). Reactions were terminated at time intervals of 5 to 200 msec. In reactions initiated at E 1, the initial rates of EP formation were virtually unaltered, while EP levels were slightly augmented, by phos- pholamban phosphorylaion (69, 70) (Fig. 10A). In reactions initiated at E2, in which EP formation is accomplished via conversion of E 2 to Ej (see Equa- tion 2), the initial rates of EP formation were markedly enhanced by phosphorylation of phos-
- 0 . 5
- 1 . 0
E I r I
• / o.5
6 • e .°%
0~0. @'@ O\ \.
o o ° \% ~°%
0
t-°\:\! -01
o~ - 0 3
- 0 4
- 0 E I
I I I 36 3 3 3 4 3 5
I~T X 0 ~
! !
Fig. 9. Arrhenius plot of the rate constant, kd, of EP decomposi- tion in unphosphoryla ted and phosphorylated sarcoplasmic re- t iculum vesicles. Following pretreatment with protein kinase and with anion exchange resin, control (O) and phosphorylated (e) cardiac sarcoplasmic reticulum vesicles were incubated with 10 #M CaC12 in media containing I mM MgC12, 100 mM KC1, 5 mM NAN3, and 10 mM imidazole-HC1 buffer (pH 7.0) in the presence of 100 #M [T-nP]ATP at temperatures ranging from 0 ° to 30 °C. Ten seconds after the addition of Ca 2+, further formation of EP was terminated by chelation of Ca 2+ with 2 mM EGTA, and the rate of decay in the amount of EP was deter- mined. The logarithm of the first-order rate constant of EP decomposition, ka, was plotted against the reciprocal tempera- ture. Inset: semilogarithmic t ransformation of the decay in EP at 10 ° C. [EP]o and [EP]t are the amounts of EP at 0 and t s after the addition of EGTA (From Tada et al. (38).)
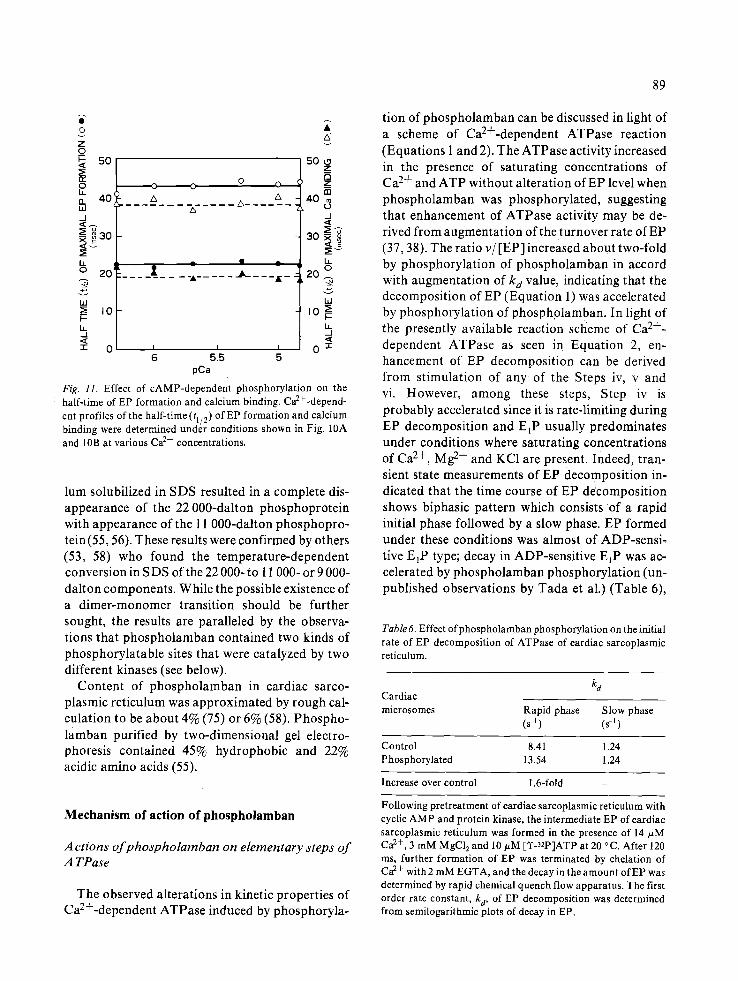
pholamban (69) (Fig. 10A). Under these condi- tions, the initial rates were much lower than those initiated at E 1 since the step at which E 2 is converted to E l is rate-determining (44). The step E2--E 1 is vir- tually independent of Ca 2+ concentrations. Thus, the values of tl/2 (the time at which half of EP is formed) were markedly shortened after phospho- rylation of phospholamban (69); phosphorylated sarcoplasmic reticulum exhibited t 1/2 of 22 msec, compared with control value of 43 msec for deter- minations made at 7 different Ca 2+ concentrations
87
(Fig. 11). These results indicated that the step at which E 2 is converted to E 1 was accelerated by phospholamban phosphorylation. In accordance with these findings, the transient state kinetic analysis of calcium transport indicated that the rapid initial rates of calcium transport was markedly enhanced by phosphorylation of phospholamban when reactions were initiated at E 2 (69) (Fig. 10B).
Calcium efflux
When a small amount of tracer radioactive cal- cium was added to cardiac sarcoplasmic reticulum loaded with nonradioactive calcium under the steady-state conditions during which efflux equals influx, the rate of calcium efflux calculated from the measured influx of 45Ca 2+ was stimulated by phosphorylation ofphospholamban(72); Ca 2+ con- centration to attain the half-maximal activation was effectively reduced. Katz et al. (57, 73) reported that the rate of calcium efflux from cardiac sarco- plasmic reticulum loaded with 45Ca2+ induced by addition of excess ADP and EGTA was signifi- cantly increased, when sarcoplasmic reticulum was preincubated with protein kinase. These results are indicative of the possibility that the rate at which calcium pump is reversed is enhanced by phospho- lamban phosphorylation, although underlying mechanism remains to be further clarified.
Structure of phospholamban
The22 000-dalton protein phospholamban, tight- ly associated with sarcoplasmic reticulum mem- brane, appears to be an acidic proteolipid that could be extracted by proteolipid extraction proce- dures (54, 55). A part of phospholamban was sug- gested to be exposed at the surface of sarcoplasmic reticulum vesicles based on the observations that proteolytic digestion resulted in disappearance of phospholamban (11,54, 55). The lack ofiodination of phospholamban (74), that contains tyrosine (55), probably reflect the location of these residues with- in the membrane interior.
Although phospholamban was thought to re- present a single peptide that electrophoretically mi- grated as a 22 000-dalton component in the pres- ence of SDS, an addition of Triton X-100 to and boiling of the phosphorylated sarcoplasmic reticu-
88
I I I I c
0.3 I0 ~M [T-32P] ATP o • 14pM Ca a+
a ATP / A ~ - ~. Phosphorylated ,., ,_/ - - , . _ , + / ~ Ca2+- bound Enzyme . . . . . . .
8 1 , , t~=13msec ~ ' ~ " '~ . . . . A ,' zx ~ a . . _ _ . . _ . _ \ 4
/ ; / ~ " ~ J ,' "~ .e . . . . . . . . . . . . . • . . . . . . . . . . . . ~ ~ A
O ~ 0.1 I- ~ /-~-t~:12rnsec" ~ / /
L'i " - ' " Ca2+-free Enzyme ] _ -'1 /,, , . , o _ _ _ _ _ _ _ _ _ _ _ _ o _ _ o
I:a ~,~ = 21 m s ~ " " Control
~/,.,'O/ t,=43msec 0 o..,"-" I I I 0 50 100 150 200
REACTION TIME (msec)
Fig. IOA. Effect of treatment with cAMP-dependent protein kinase on the initial rate of Ca2---dependent formation of EP of ATPase of Ca2+-free and Ca2+-bound cardiac sarcoplasmic reticulum. Following pretreatment with protein kinase and anion exchange resin, cardiac sarcoplasmic reticulum vesicles were added to a solution containing EGTA (O, e) which gave 14/~M ionized Ca 2+ by subsequent combination with 0.1 mM CaC12, or calcium/EGTA buffer with 14 #M Ca 2+ (~, *). The ATPase reaction was subsequently started by the addition of 10/~M [T-32p]ATP and 0.1 mM CaCI 2 (O, o) or calcium/EGTA buffer (L~, A), respectively, through the rapid chemical quench flow apparatus, and the amount of EP formed was determined. (From Tada et al. (69).)
0 , 5 I I I
® . . A
-- - . . . . . . Phosphorylated /
2 o.a , , , " ~ ~ -I :) ,, ~ z x ' - - ' - - - / / / o , ~ . - - - - - - - - - / / / m / f z ~ - " . / _ . . . . . . . . _ ~ . . . . . . . I
," ~ a . . . . . . . . . . . . . . . . . . . . . . . / • r / ' - ' ' ' " - I o 0.2 ~ 4 " " I Ca'+-fr,, microsomes ] / O
" ° I _~o,F ,,,,i / o / 1 -I 0 | , ~ 0 Control I
I ,.• .o" I
~ o! '~'°~ , , , I 0 50 I O0 150 200
REACTION TIME (msec)
Fig. lOB. Effect of treatment with cAMP-dependent protein kinase on the initial rate of calcium binding by Ca2+-free and Ca2+-bound cardiac sarcoplasmic reticulum vesicles. Reactions were performed under the same conditions described above, except 45CAC12 and cold ATP were used instead of cold CaC12 and [Y-3~P]ATP. (From Tada et al. (69).)
@
0
Z o ~- 50 < ~E (X O LL o. 4.0 ILl .-I
~Eo ~
b. o 20 ~
0 0 0 ,_'3
A A
- • -
: .............. ~ .A - . I K - - - - I -
I I I
6 55 5 pCa
A
50 o Z
Z
(D ._I <
h
2(3 o
Fig. 11. Effect of cAMP-dependent phosphorylation on the half-time of EP formation and calcium binding. Ca2+-depend - ent profiles of the half-time (tl/2) of EP formation and calcium binding were determined under conditions shown in Fig. 10A and 10B at various Ca 2+ concentrations.
lum solubilized in SDS resulted in a complete dis- appearance of the 22 000-dalton phosphoprotein with appearance of the 11 000-dalton phosphopro- tein(55, 56). These results were confirmed by others (53, 58) who found the temperature-dependent conversion in SDS of the 22 000- to 11 000- or 9 000- dalton components. While the possible existence of a dimer-monomer transition should be further sought, the results are paralleled by the observa- tions that phospholamban contained two kinds of phosphorylatable sites that were catalyzed by two different kinases (see below).
Content of phospholamban in cardiac sarco- plasmic reticulum was approximated by rough cal- culation to be about 4% (75) or 6% (58). Phospho- lamban purified by two-dimensional gel electro- phoresis contained 45% hydrophobic and 22% acidic amino acids (55).
Mechanism of action of phospholamban
Actions of phospholamban on elementary steps of A TPase
The observed alterations in kinetic properties of Ca2+-dependent ATPase induced by phosphoryla-
89
tion of phospholamban can be discussed in light of a scheme of Ca2+-dependent ATPase reaction (Equations 1 and 2). The ATPase activity increased in the presence of saturating concentrations of Ca 2+ and ATP without alteration of EP level when phospholamban was phosphorylated, suggesting that enhancement of ATPase activity may be de- rived from augmentation of the turnover rate of EP (37, 38). The ratio v/[EP] increased about two-fold by phosphorylation of phospholamban in accord with augmentation of k d value, indicating that the decomposition of EP (Equation 1) was accelerated by phosphorylation of phospholamban. In light of the presently available reaction scheme of Ca 2+- dependent ATPase as seen in Equation 2, en- hancement of EP decomposition can be derived from stimulation of any of the Steps iv, v and vi. However, among these stePS, Step iv is probably accelerated since it is rate-limiting during EP decomposition and E~P usually predominates under conditions where saturating concentrations of Ca 2+, Mg 2+ and KC1 are present. Indeed, tran- sient state measurements of EP decomposition in- dicated that the time course of EP decomposition shows biphasic pattern which consists of a rapid initial phase followed by a slow phase. EP formed under these conditions was almost of ADP-sensi- tive E~P type; decay in ADP-sensitive EIP was ac- celerated by phospholamban phosphorylation (un- published observations by Tada et al.) (Table 6),
Table 6. Effect ofphospholamban phosphorylation on the initial rate of EP decomposition of ATPase of cardiac sarcoplasmic reticulum.
Cardiac microsomes
¾
Rapid phase (s -~)
Slow phase (s -~)
Control 8.41 1,24 Phosphorylated 13.54 1.24
Increase over control 1.6-fold -
Following pretreatment of cardiac sarcoplasmic reticulum with cyclic AMP and protein kinase, the intermediate EP of cardiac sarcoplasmic reticulum was formed in the presence of 14 #M Ca 2+, 3 mM MgC12 and 10 ~zM [T-nP]ATP at 20 °C. After 120 ms, further formation of EP was terminated by chelation of Ca 2+ with 2 mM EGTA, and the decay in the amount of EP was determined by rapid chemical quench flow apparatus. The first order rate constant, ka, of EP decomposition was determined from semilogarithmic plots of decay in EP.

90
suggesting that phosphorylation of phospholam- ban may stimulate E~P--E2P (Step iv in Equation 2). The rate of EP formation was accelerated by phosphorylation of phospholamban when reac- tions were initiated at E 2 but not at E~ (Fig. 10), indicating that phosphorylation of phospholamban enhances a step of E2--E 1 (Step i in Equation 2), which is coupled with the reaction at which Ca 2+ affinity is greatly altered (69). Transient state mea- surements of reverse reaction of ATPase indicated that the rate of EP formation from Pi was augment- ed under phosphorylation of phospholamban when reactions were initiated at E t, but not at E2, suggest- ing that phospholamban may also enhance the rev- ersal rate of Step i (E~E2) (unpublished observa- tions by Tada et al.) (Table 7). It is of prime significance that both of the two key rate-determin- ing steps (Steps i and iv) during turnover of ATPase are enhanced when phospholamban is at phospho- rylated state. These are the two major steps at which the affinity of the enzyme for Ca 2+ is greatly al- tered. Taking it by and large, phospholamban would probably exert its action by regulating the cati0n-mediated conformational changes of the ATPase enzyme (see below).
It is also important to elucidate the relationship between phospholamban and ATPase in the mem- brane. Since a protein-protein interaction was sug- gested to exist between the ATPase and phospho- lamban, it is of interest to examine whether these rate-determining steps of ATPase are affected by phospholamban Ph0sphorylation when these pro- teins are solubilized by detergents.
Molecular model
Cyclic AMP-dependent phosphorylation of
Table 7. Effect of phospholamban phosphorylation on the re- verse ATPase reaction (EP formation from Pi).
Cardiac microsomes Initial velocity of EP formation
i~mol/ l Orns. g protein Control 0.029 Phosphorylated 0.076
Following pretreatment with cyclic A M P and protein kinase, cardiac sarcoplasmic reticulum was assayed for EP formation from P i in the presence of a trace amount of Ca 2+, 10 mM M g 2+ and 5 mM 32Pi at 20 ° C, employing a rapid chemical quench flow apparatus.
phospholamban profoundly altered the calcium transport by cardiac sarcoplasmic reticulum. Since such alterations were produced by modulating the kinetic properties of the Ca2+-dependent ATPase (11,38, 69), it was assumed that phospholamban is closely associated with the ATPase. Thus, as work- ing hypotheses, molecular models were presented in which phosphorylated phospholamban serves as an accelerator (37) or a desuppressor (63, 76) of the ATPase. Adding more recent informations, we now present a newer model (Fig. 12). The ATPase en- zyme is an amphipathic single polypeptide with a molecular weight of 100 000 which functions as a monomer or an oligomer during translocation of Ca 2+ (I, 77). A protein-protein interaction is sug- gested to exist between phospholamban and AT- Pase (38, 69), and the association appears to be tighter when phospholamban is at phosphorylated state (54, 75). In fact, the direct molecular interac- tion would be justified by the data, in which the ATPase enzyme and phospholamban remained as- sociated with each other after solubilization of sar- coplasmic reticulum with detergents (54, 55, 75). Although part of phospholamban is located at the cytoplasmic surface of sarcoplasmic reticulum
A. Phospholarnban at unphosphorylated state
Cao* ~" ~ ATPP j~ADP CYTOPLASM
ATPase Phospholarnban B. Phospholarnban at phosphorylated state
Ca z+ 0 E'~ AT L AjDP CYTOPLASM
ATP__ Phosp LUMEN OF SR
Fig. 12. Diagrammatic representation of the action of phospho- lamban on the calcium pump of cardiac sarcoplasmic reticulum. A S P represents an aspartic acid residue in the ATPase and S E R a serine residue in phospholamban. The phosphates attached to these amino acids are --P, the acyl phosphate ATPase interme- diate, and -P, the regulatory phosphoester. (From Tada and Katz (9).)

membrane(11,55), phospholamban is tightly bound to the membrane interior (54, 55, 74). Upon phos- phorylation, the phosphorylation site of phospho- lamban would be transposed within the membrane so that tryptic digestion becomes less effective (11, 54). A 1:1 stoichiometry is probably acceptable since protein contents of these two components and the amounts ofphosphoproteins formed within the membrane exhibited roughly a 1:1 molar ratio (38, 56, 58, 73, 75). During translocation of Ca 2+, the moiety of ATPase containing the active site (30 000 daltons) undergoes conformational change that causes the calcium-binding moiety (20 000 daltons) to translocate from outer to inner position (1, 77). Since both formation (69) and decomposition (38, 70) of EP were enhanced by phosphorylation of phospholamban, the conformational state of the 30 000-dalton moiety would probably be under di- rect control ofphospholamban. At phosphorylated state phospholamban would enhance the rate at which cation-mediated conformational changes of 30 000-dalton moiety take place, resulting in an increased rate of transposition of Ca2+-bound 20 000-dalton moiety.
Calmodulin-dependent phosphorylation of phos- pholamban
Le Peuch et al. (58) and Kranias et al. (78) report- ed that phosphate could be incorporated into 22 000-dalton protein, phospholamban, in the pres- ence of Ca 2+ and calmodulin. Formation of such phosphoprotein, whose stability characteristics were those of a phosphoester, was dependent on calmodulin but independent of cyclic AMP. They postulated that phospholamban could be phospho- rylated by an endogenous protein kinase associated with sarcoplasmic reticulum which was activated by calmodulin and Ca 2+. This membrane-bound protein kinase had characteristics very similar, but not identical, with glycogen phosphorylase b kinase (58). The maximal amount of phospholamban phosphorylation catalyzed by calmodulin-depen- dent protein kinase at the optimal Ca 2+ concentra- tion of 5-10 #M was about the same as that found in cyclic AMP-dependent protein kinase (56). While phosphorylation catalyzed by the two different ki- nases was found to occur independently, an addi- tive elevation of the amount of phosphorylation was seen when cardiac sarcoplasmic reticulum was
91
subjected to serial incubation under condition fa- vorable for two different kinases (56, 58).
S. Katz et al. (79) reported that the preincubation of cardiac sarcoplasmic reticulum with calmodulin resulted in an enhanced rate of calcium uptake. Such enhancement was found to be accompanied by calmodulin-dependent phosphorylation of phospholamban (56, 58, 80). However, it is not clear as to how such enhancement is related to cyclic AMP-mediated augmentation of the active calcium transport. Le P euch et al. (58) suggested that cyclic AMP-mediated control of active calcium transport was not seen unless calmodulin-depend- ent phosphorylation was operational, on the basis of the observations that in the absence of calmodu- lin-dependent phosphorylation of phospholamban calcium transport was not enhanced by cyclic AMP-dependent protein kinase. In contrast, Tada et al (56) found that cyclic AMP-mediated en- hancement of calcium transport can be seen when calmodulin-dependent phosphorylation was not operated. Such enhancement was observed inde- pendently when either of the systems was opera- tional and the effects were found to be additive, in accordance with the finding that phosphorylation by the two kinases occurred in independent and additive manners. It is possible that the Ca 2+ con- centrations employed by Le Peuch et al. (58) were far too high (0.1-0.5 mM) to see the physiological effect o f calcium control in cardiac sarcoplasmic reticulum. The stimulatory effects could be seen only when the Ca 2+ concentrations were within the range between 0.1 and 10 #M.
It is important to examine~whether calmodulin- dependent phosphorylation of phospholamban af- fects Ca2+-dependent ATPase activity. Le Peuch et al. (58) presented a hypothetical view that calmo- dulin did not affect ATPase activity. Again, the Ca 2+ concentration ranges employed in their exper- iment were too high to detect such a subtle change in Ca2+-dependent ATPase activity. Tada et al. (56) and Lopaschuk et al. (80) indicated that at physio- logical concentrations of Ca 2+ (0.1-50 #M), cal- modulin-dependent phosphorylation of phospho- lamban resulted in the stimulation of Ca 2+- dependent ATPase activity. It remains to be ex- tensively explored how the stimulatory effect produced by calmodulin is related to cyclic AMP- mediated enhancement of the calcium transport and ATPase activity.

92
Physiological relevance
Tada, Kirchberger & Katz first proposed that cyclic AMP-mediated acceleration of calcium up- take by cardiac sarcoplasmic reticulum may ex- plain the two principal mechanical effects of ca- techolamines on heart muscle; abbreviation of systole and increased contractility (11,57, 61) (Fig. 13). Some evidence (see 5, 9, 73, 81 for review) supports the view that the protein kinase-phospho- lamban system capable of accelerating the ATPase assumes a role in mediating catecholamine effects; this is summarized in Fig. 13. Cyclic AMP-mediat- ed increase in the rate of calcium accumulation by sarcoplasmic reticulum of intact heart could ex- plain abbreviation of systole because of the in- creased rate at which calcium would be removed from troponin. The increase in the rate of calcium accumulation following phospholamban Ph0spho- rylation could increase the amount of calcium stored in sarcoplasmic reticulum by retaining with- in the cell some of the calcium which would other- wise be lost during diastole. This could add to the
B-adranargic agent
i W
adenylate cyclase
ATP cAMP RmcAMP >_<' non-activated ~ RC C (activated protein kinase)
protein kinase)
phospholamban phosphorylated phospholamban
basal Ca-ATPase enanced Ca-ATPase and transDort and transport
increased Ca in SR
l abbreviation[ increased Ca release of systo e J in subsequent contractions
[ enhanced contractility I
Fig. 13. A possible mechanism for fl-adrenergic effects on con- tractility, mediated by enhanced Ca uptake by the sarcoplasmic reticulum (SR). RC represents protein kinase in soluble form or in association with SR membranes. R represents the regulatory subunit, C the catalytic subunit of the protein kinase. (Modified from Tsien (5).)
amount of calcium available for delivery to the contractile proteins in subsequent contractions, thus promoting augmentation of myocardial con- tractility (positive inotropic effect).
The proposed sequence of events, that is related to cyclic AMP-mediated changes in calcium tran- sients, may get support from experiments in intact heart, in that catecholamines produced relaxation at the very early phase, followed by augmentation of contraction (82). In fact, the onset of increased tension development after exposure of the heart to catecholamines was gradual, reaching the steady state after about 20 beats (83). Employing a skinned cardiac cell, which exhibited cycles of phasic con- tractions upon addition of an appropriate amount of Ca 2+ (84), Fabiato & Fabiato (85) demonstrated that a brief preincubation with cyclic AMP resulted in increased amplitude of contraction and faster rates of tension development and relaxation. The more direct evidence was presented by Allen & Blinks (13a) who measured intracellular Ca 2+ by aequorin, a calcium-sensitive bioluminescent pro- tein, and found that catecholamines augmented the initial rate of calcium release from sarcoplasmic reticulum during the early phase of contraction as well as the rate of reduction of Ca 2+ during relaxa- tion.
That cyclic AMP could mediate the inotropic effects of catecholamines was indicated by an in- crease in cyclic AMP prior to increased contractili- ty (5, 86). These results are at variance with findings by others (87, 88) who found no detectable increase in cyclic AMP when catecholamines covalently at- tached to glass beads augmented contraction. It should be further investigated whether a subtle in- crease in cyclic AMP, that might have been too low to detect, could have given rise to significant me- chanical changes. The dissociation of inotropic re- sponses from cyclic AMP formation may reflect some effect of catecholamines on sarcolemmal func- tions such as calcium channels. Alternatively, the cyclic AMP-protein kinase system could possibly be in a close contact with the membrane, forming a compartment. Immunocytochemical studies indi- cated that cyclic AMP (89) and possibly protein kinase (90) were predominantly localized at the area of sarcoplasmic reticulum and sarcolemma. That phospholamban is phosphorylated in viva was indicated by the findings (91,92, 92a) that the addition of isoproterenol to the heart perfused with

32p resulted in increased 32p incorporation into the 22 000-dalton microsomal protein (phospholam- ban) with simultaneous increase in the rate of ten- sion development and relaxation. Interestingly, the addition of acetylcholine was found to reduce the isoproterenol-induced increase in phospholamban phosphorylation (92). Calmodulin inhibitor (flu- phenazine) significantly reduced phosphorylation of phospholamban (91).
Kirchberger & Tada (48) were the first to report that cyclic AMP-phospholamban system could be operational in other muscular tissues. They found that microsomes isolated from slow-contracting skeletal muscle contained a phosphorylatable pep- tide similar to phospholamban (Fig. 4C), that ap- pears to exhibit stimulation of calcium uptake, while no phosphorylatable peptide was seen in fast- contracting skeletal muscle (Fig. 4A), nor was cal- cium uptake stimulated (48). Confirming the latter findings on cardiac and slow-contracting skeletal muscles, Schwartz et al (49) also found the in- creased rate of calcium uptake by sarcoplasmic re- ticulum from fast-contracting skeletal muscle with- out accompanying membrane phosphorylation. Possible existence of a similar mechanism was also considered in microsomes from other tissues such as platelets (93, 94) and vascular (95) and visceral (96) smooth muscles. In platelet microsomes a phospholamban-like protein was found to function as a regulator of calcium accumulation (97), while direct evidence was yet to be presented in smooth muscle. In hyperthyroid animals, the amounts of phospholamban in cardiac microsomes were markedly augmented (98), that accompanied an enhanced rate of calcium uptake (98, 99). The stim- ulation of calcium uptake produced by the cyclic AMP-phospholamban system in rat heart micro- somes was not altered by aging, while aged hearts exhibited a reduced rate of unstimulated micro- somal calcium uptake (100).
Acknowledgments
This work was supported by research grants from the Ministry of Education, Science and Culture and the Ministry of Health and Welfare of Japan and the Japan Heart Foundation, and by a Grant-in- Aid from the Muscular Dystrophy Association of America.
93
R eferenees
1. Tada, M., Yamamoto, T. & Tonomura, Y., 1978. Physiol. Rev. 58: 1-79.
2. Endo, M., 1977. Physiol. Rev. 57: 71-108. 3. Fabiato, A. & Fabiato, F., 1979. Ann. Rev. Physiol. 41:
473 484. 4. Sutherland, E. W. & Rail, T. W., 1960. Pharmacol. Rev.
12: 265-299. 5. Tsien, R. W., 1977. Adv. Cyclic Nuc. Res. 8: 363-420. 6. Krebs, E. G., 1972. Curr. Top. Cell. Regul. 5: 99-133. 7. Brostrom, M. A., Reimann, E. M., Walsh, D. A. & Krebs,
E. G., 1970. Adv. Enzyme Regul. 8:191 203. 8. Soderling, T. R. & Park, C. R., 1974. Adv. Cyclic Nuc. Res.
4: 283-333. 9. Tada, M. & Katz, A. M., 1982. Ann. Rev. Physiol. 44:
401-423. 10. Tada, M., Kirchberger, M, A., Iorio, J.-A. & Katz, A. M.,
1973. Circulation 48 (Suppl): IV-25. I 1. Tada, M., Kirchberger, M. A. & Katz, A, M., 1975. J. Biol.
Chem. 250: 2640-2647. 12. Katz, A. M., 1977. In: Physiology of the Heart, pp.
137-159, Raven Press, New York. 12a. McNutt, N. S. & Fawcett, D. W., 1969. J. Cell Biol. 42:
1-45. 13. Ashley, C. C. & Ridgway, E. B., 1968. Nature 219:
1168 1169. 13a. Allen, D. G. & Blinks, J. R., 1978. Nature273:509 513. 14. Marban, E.,Rink, T.J.,Tsien, R.W.&Tsien, R.Y., 1980.
Nature 286: 845-850. 15. Ebashi, S., 1976. Ann. Rev. Physiol. 38: 293-313. 16. Harigaya, S. & Schwartz, A., 1969. Circ. Res. 25:781 794. 17. Levitsky, D. O., Aliev, M. K., Kuzmin, A. V., Levchenko,
T. S., Smirnov, V. N. & Chazov, E. I., 1976. Biochim. Biophys. Acta 443:468 484.
18. Jones, L. R., Besch, H. R., Jr., Fleming, J. W., McCon- naughey, M. M. & Watanabe, A. M., 1979. J. Biol. Chem. 254: 530-539. Affolter, H., Chiesi, M., Dabrowska, R. & Carafori, E., 1976. Eur. J. Biochem. 67:389 396. Suko, J. & Hasselbach, W., 1976. Eur. J. Biochem. 64: 123-130. Van Winkle, W. B., Pitts, B. J. R. & Entman, M. L., 1978. J. Biol. Chem. 253: 8671-8673. MacLennan, D. H. & Holland, P. C., 1976. In: The En- zymes of Biological Membranes (Martonosi, A., ed.), pp. 221-259, Plenum Press, New York.
23. Fleischer, S. Wang, C.-T., Hymel, L., Seeling, J., Brown, M. F., Herbette, L., Scarpa, A., McLaughlin, A. C. & Blasie, J. K., 1979. In: Function and Molecular Aspects of Biomembrane Transport (Quagliariello, E., Palmieri, F., Papa, S., Klingenberg, M., eds.), pp. 465 485, Elsevier/ North-Holland, Amsterdam.
24. MacLennan, D. H. & Holland, P. C., 1975. Ann. Rev. Biophys. Bioeng. 4: 377-404.
25. Thorley-Lawson, D. A. & Green, N. M., 1973. Eur. J. Biochem. 40: 403-413.
26. Thorley-Lawson, D. A. & Green, N. M., 1975. Eur. J. Bioehem. 59: 193-200.
19.
20.
21.
22.

94
271 MacLennan, D. H., Seeman, P., lles, G. H. & Yip, C. C., 1971. J. Biol. Chem. 246: 2702-2710.
28. Stewart, P. S. & MacLennan, D. H., 1974. J. Biol. Chem. 249: 985-993.
29. Stewart, P. S., MacLennan, D. H. & Shamoo, A. E., 1976. J. Biol. Chem. 251: 712-719.
30. Yamamoto, T. & Tonomura, Y., 1977. J. Biochem. Tokyo 82: 653-660.
31. Inesi, G. & Scales, D., 1974. Biochemistry 13:3298 3306. 32. Louis, C. F., Buonaffina, R. & Binks, B., 1974. Arch.
Biochem. Biophys. 161: 83-92. 33. Louis, C. & Shooter, E. M., 1972. Arch. Biochem. Biophys.
153:641 655. 34. Yu, B. P., Masoro, E. J. & Morley, T. F., 1976. J. Biol.
Chem. 251: 2037-2043. 35. Rizzolo, L. J., Le Maire, M., Reynolds, J. A. & Tanford,
C., 1976. Biochemistry 15: 3433-3437. 36 Le Maire, M., Jorgensen, K. E., R¢igaard, H. & Moller,~l.
V., 1976. Biochemistry 15: 5805-5812. 37. Tada, M., Ohmori, F., Kinoshita, N. & Abe, H., 1978. Adv.
Cyclic Nuc. Res. 9:355 369. 38. Tada, M., Ohmori, F., Yamada, M. & Abe, H., 1979. J.
Biol. Chem. 254: 319-326. 39. Pang, D. C. & Briggs, F. N., 1973. Biochemistry 12:
4905-4911. 40. Shigekawa, M., Finegan, J.-M. & Katz, A. M., 1976. J.
Biol. Chem. 251: 6894-6900. 41. Froehlich, J. P. & Taylor, E. W., 1976. J. Biol. Chem. 251:
2307-2315. 42. Shigekawa, M. & Dougherty, J. P., 1978. J. Biol. Chem.
253: 1451-1457. 43. Shigekawa, M., Dougherty, J. P. & Katz, A. M., 1978. J.
Biol. Chem. 253:1442 1450. 44. De Meis, L. & Vianna, A. L., 1979. Ann. Rev. Biochem. 48:
275-292. 45. LaRaia, P. J. & Morkin, E., 1974. Circ. Res. 35: 298-306. 46. Wray, H. L., Gray, R. R. & Olsson, R. A., 1973. J. Biol.
Chem. 248: 1496-1498. 47. Kirchberger, M. A., Tada, M. & Katz, A. M., 1974. J. Biol.
Chem. 249: 6166-6173~ 48. Kirchberger, M. A. & Tada, M., 1976. J. Biol. Chem. 251:
725-729. 49. Schwartz, A., Entman, M: L., Kaniike, K., Lane, L. K.,
Van Winkle, W. B ~, & Bornet, E. P., 1976. Biochim. Bio- phys. Acta 426: 57-72.
50. Wray, H. L. & Gray, R. R., 1977, Biochim. Biophys. Acta 461: 441-459.
51. Will, H., Levchenko, T. S., Levitsky, D. O., Smirnov, V. N. & Wollenberger, A., 1978. Biochim. Biophys. Acta 543: 175-193.
52. St. Louis, P. J. & Sulakhe, P. V., 1979. Arch. Biochem. Biophys. 198: 227-240.
53. Lamers, J. M. J. & Stinis, J. T., 1980. Biochim. Biophys. Acta 624: 443-459.
54. Bidlack, J. M. & Shamoo, A. E., 1980. Biochim. Biophys. Acta 632: 310-325.
55. Le Peuch, C. J., Le Peuch, D. A. M. & Demaille, J. G., 1980. Biochemistry 19: 3368-3373.
56. Tada, M., Inui, M., Yamada, M., Kadoma, M., Kuzuya, T., Abe, H. & Kakiuchi, S., 1982. J. Mol. Cell. Cardiol. (in press).
57. Katz, A. M., Tada, M. & Kirchberger, M. A., 1975. Adv. Cyclic Nuc. Res. 5: 453-472.
58. Le Peuch, C. J., Haiech, J. & Demaille, 2. G., 1979. Bio- chemistry 18: 5150-5157.
59. Nimmo, H. G. & Cohen, P., 1977. Adv. Cyclic Nuc. Res. 8: 145 266.
60. Kirchberger, M. A., Tada, M., Repke, D. I. & Katz, A. M., 1972. J. Mol. Cell. Cardiol. 4: 673-680.
61. Tada, M., Kirchberger, M. A., Repke, D. I. & Katz, A. M., 1974. J. Biol. Chem. 249: 6174-6180.
62. Kirchberger, M. A. & Chu, G., 1976. Biochim. Biophys. Acta 419: 559-562.
63. Hicks, M. J., Shigekawa, M. & Katz, A. M., 1979. Circ. Res. 44: 384-391.
64. Tada, M., Kirchberger, M. A. & Li, H.-C., 1975. J. Cyclic Nuc. Res. 1: 329-338.
65. Kirchberger, M. A. & Raffo, A., 1977.2. Cyclic Nuc. Res. 3: 45-53.
66. Tada, M., Ohmori, F., Nimura, Y. & Abe, H., 1977. J. Biochem. Tokyo 82: 885-892.
67. Ohmori, F., Tada, M., Kinoshita, N., Matsuo, H., Saka- kibara, H., Nimura, Y. & Abe, H., 1978. In: Recent Advan- ces in Studies on Cardiac Structure and Metabolism, Vol. 11 (Kobayashi, T., Sano, T., Dhalla, N. S., eds.) pp. 279 284, University Park Press, Baltimore.
68. Will, H., Blanck, J., Smettan, G. & Wollenberger, A., 1976. Bioehim. Biophys. Acta 449: 295-303.
69. Tada, M., Yamada, M., Ohmori, F., Kuzuya, T., Inui, M. & Abe, H., 1980. J. Biol. Chem. 255: 1985-1992.
70. Kranias, E. G., Mandel, F., Wang, T. & Schwartz, A., 1980. Biochemistry 19: 5434-5439.
71. Froehlich, J. P., Sullivan, J. V. & Berger, R. L., 1976. Anal. Biochem. 73: 331-341.
72. Kirchberger, M. A. & Wong, D., 1978. J. Biol. Chem. 253: 6941-6945.
73. Katz, A. M., 1979. Adv. Cyclic Nuc. Res. 11: 303-343. 74. Louis, C. F. & Katz, A. M., 1977. Biochim. Biophys. Acta
494: 255-265. 75. Tada, M., Yamada, M., Ohmori, F., Kuzuya, T. & Abe,
H., 1979. In: Cation Flux Across Biomembranes (Muko- hata, Y. & Packer, L., eds.) pp. 179 190, Academic Press, New York.
76. Katz, A. M., 1980. Trends Phrmacol. Sci. 1:434 436. 77. Yamamoto, T., Takisawa, H., Tonomura, Y., 1979. Curt.
Top. Bioenerg. 9: 179-236. 78. Kranias, E. G., Bilezikjian, L. M., Potter, J. D., Piascik, M.
T. & Schwartz, A., 1980. Ann. NY. Acad, Sci. 356: 279 29O.
79. Katz, S. & Remtulia, M. A., 1978. Biochem. Biophys. Res. Commun. 83: 1373-1379.
80. Lopaschuk, G. Richter, B. & Katz, S., 1980. Biochemistry 19: 5603-5607.
81. Stull, J. T. & Mayer, S. E., 1979. In: Handbook of Physiol- ogy. II. The Cardiovascular System. Vol. 1 (Berne, R. M., Sperelakis, N., eds.), pp. 741-774, American Physiological Society, Bethesda.
82. Morad, M. & Rolett, E. L., 1972. J. Physiol. (London) 224: 537-558.
83. Reuter, H., 1974. J. Physiol. London 242: 429-451. 84. Fabiato, A. & Fabiato, F., 1977. Circ. Res. 40: 119-129. 85. Fabiato, A. & Fabiato, F., 1975. Nature 253:556 558.

86. Schumann, H. J., Endoh, M. & Brodde, O. E., 1975. Nau- nyn-Schmiedebergs Arch. Pbarmacol. 289: 291-302.
87. Ingebretsen, W. R. Jr., Bercker, E., Friedman, W. F. & Mayer, S. E., 1977. Circ. Res. 40: 474-484.
88. Venter, J. C., Ross, J. Jr. & Kaplan, N. O., 1975. Proc. Natl. Acad. Sci. U.S.A. 72:824 828.
89. Ong., S. M. & Steiner, A. L., 1977. Science 195: 183-185. 90. Steiner, A, L., Koide, Y., Earp, H. S., Bechtel, P. J. &
Beavo, J. A., 1978. Adv. Cyclic Nuc. Res. 9:691 705. 91. Le Peuch, C. J., Guilleux, J.-C. & Demaille, J. G., 1980.
FEBS Lett. 114:165 168. 92. Lindemann, J. P., Jones, L. R. & Watanabe, A. M., 1980.
Clin. Res. 28: 471A (Abstract). 92a. Lindemann, J. P., Jones, L. R. & Watanabe, A. M., 1981.
Circulation 64 (Soppl) IV-22. 93. Haslam, R.J., Davidson, M. M. L., Davies, T., Lynham, J.
A. & McClenaghan, M. D., 1978. Adv. Cyclic Nuc. Res. 9: 533-552.
95
94. Kaser-Glanzmann, R., Jak/tbovfi, M., George, J. N. & Ltischer, E. F., 1977. Biochim. Biophys. Acta466: 429-440.
95. Bhalla, R. C., Webb, R. C., Singh, D. & Brock, T., 1978. Am. J. Physiol. 234:H508 H514.
96. Kimura, M., Kimura, I. & Kobayashi, S., 1977. Biochem. Pharmacol. 26: 994-996.
97. K~.ser-Glanzmann, R., Gerber, E. & LUscher, E. F., 1979. Biochim. Biopbys. Acta 558: 344-347.
98. Limas, C. J., 1978. Am. J. Physiol. 234:H426 H431. 99. Limas, C. J., 1978. Am. J. Physiol. 235:H745 H751.
100. Kadoma, M., Sacktor, B. & Froehlich, J. P., 1980. Fed. Proc. 39:2040 (Abstract).
Received 28 December 1981.




















