
Low-pH Adaptation and the Acid Tolerance Response of Bifidobacterium longum Biotype longum

Bi®dobacteriumspp. and
Lactobacillusacidophilus:biological,
biochemical,technological and
therapeuticalproperties relevant
for use asprobiotics
Ana M.P. Gomes andF. Xavier Malcata*
Escola Superior de Biotecnologia, UniversidadeCato lica Portuguesa Rua, Dr. Anto nio Bernardino deAlmeida, 4200 Porto, Portugal (fax:+351-2-5090351)
This review focuses on the biological properties and con-sequent technological roles of intestinal bacteria withpotential health-promoting capacities, and provides selectedexamples available in the literature that are pertinent to theaforementioned concepts. A comprehensive overview per-taining to the taxonomy and ecology, as well as nutritionaland health e�ects of Bi®dobacterium spp. and Lactobacillus
acidophilus, is provided; particular attention is paid to theirincorporation, and growth and acidi®cation features in fer-mented dairy products. The typical poor growth of thesespecies in milk is highlighted, and the use of bi®dogenic andgrowth factors, including their nature and function, is dis-cussed. E�orts to establish optimum environmental conditionsfor their growth are critically reviewed, in addition to thee�ects of the food and storage conditions on microbialsurvival. Criteria for selection of e�ective microbial strainsfor their probiotic e�ect are listed, and modi®cations toimprove fermentation e�ciency and shelf-life of ®nal dairyproducts are suggested; among these, the incorporation ofBi®dobacterium spp. and L. acidophilus into a solid matrix,such as cheese, is emphasized. # 1999 Elsevier Science Ltd.All rights reserved.
The use of Bi®dobacterium spp. and/or Lactobacillusacidophilus in fermented or culture-containing milksbecame popular by the end of the 1970s as a result ofthe tremendous increase in knowledge encompassing thetaxonomy and ecology of bi®dobacteria. Their popularityhas further increased owing to their reduced acidi®ca-tion during post-processing storage and their relativelyhigh yield of L(+)-lactic acid compared with D(ÿ)-lac-tic acid. In recent years, much work on bi®dobacteria,regarded as microorganisms targeted for technologicaland therapeutic applications, was performed in Japan, butother countries also became involved (e.g. Denmark,Germany, Poland, Russia, UK and USA) [1]. Suchconcerted work has led to many publications and patentapplications; the literature was reviewed by Rasic andKurmann [2] in a comprehensive paper that rapidlybecame a classic reference. The number of patents hasmeanwhile drastically increased, and pertain to anincreasing range of applications including new productdevelopment and starter culture production.
De®nitionThe word `probiotic', from the Greek `for life', has
over the past few years been used in several di�erentways. It was originally proposed to describe compoundsproduced by one protozoan that stimulated the growthof another [3], but in the early seventies, Sperti expan-ded the term to encompass tissue extracts that stimu-lated microbial growth. Parker [4] later applied the term
0924-2244/99/$ - see front matter Copyright # 1999 Elsevier Science Ltd. All rights reserved.PI I : S0924-2244 (99 )00033-3
Trends in Food Science & Technology 10 (1999) 139±157
* Corresponding author.
Review

to describe animal feed supplements that had a bene-®cial e�ect on the host by contributing to its intestinalmicrobial balance. Consequenlty, the word `probiotic'was applied to ``organisms and substances that con-tribute to intestinal microbial balance''. This generalde®nition was, however, unsatisfactory because animprecise word as `substances' might include a varietyof supplements, including antibiotics. Fuller [5] revisedthe de®nition to stress the importance of living cells asan essential component of an e�ective probiotic, andthus de®ned probiotic as ``a live microbial feed supple-ment which bene®cially a�ects the host animal byimproving its intestinal microbial balance''.The de®nition of probiotic was broadened in the cur-
rent decade [6]. Presently, probiotics are de®ned as``viable microorganisms (lactic acid and other bacteria,or yeasts applied as dried cells or in a fermented pro-duct) that exhibit a bene®cial e�ect on the health of thehost upon ingestion by improving the properties of itsindigenous micro¯ora''.
TaxonomyGenus Bi®dobacteriumBi®dobacteria were ®rst isolated and described in
1899±1900 by Tissier, who described rod-shaped, non-gas-producing, anaerobic microorganisms with bi®dmorphology, present in the faeces of breast-fed infants,which he termed Bacillus bi®dus. Bi®dobacteria aregenerally characterized as gram-positive, non-sporeforming, non-motile and catalase-negative anaerobes[7]. They have various shapes including short, curvedrods, club-shaped rods and bifurcated Y-shaped rods.Presently, 30 species are included in the genus Bi®do-bacterium, 10 of which are from human sources (dentalcaries, faeces and vagina), 17 from animal intestinaltracts or rumen, two from wastewater and one fromfermented milk (Table 1). Bi®dobacteria are phylogen-etically grouped in the actinomycete branch of gram-positive bacteria [7], that is characterized by a highguanine plus cytosine (G+C) content, which variesfrom 54 to 67 mol% (Table 2). In addition, there arenotable di�erences in physiological and biochemicalproperties, including cell-wall constituents (Table 2).They are saccharolytic organisms that produce aceticand lactic acids without generation of CO2, except dur-ing degradation of gluconate (Fig. 1). Heterofermenta-tion is initiated by splitting fructose-6-phosphate intoone C2 and one C4 moiety. The conversion of the C2
moiety to acetate is paralleled by the formation of hep-tose-7-phosphate from the C4 moiety concomitant withthe formation of a triose moiety derived from an addi-tional molecule of fructose-6-phosphate. The heptose-7-phosphate is subsequently split into two molecules ofacetate and one molecule of pyruvate. The second triosemoiety left from fructose-6-phosphate is converted intolactate. Therefore, the fermentation of two moles of
hexose results in three moles of acetate and two molesof lactate. The key enzyme in such glycolytic fermenta-tion, fructose-6-phosphate phosphoketolase, may beused as a taxonomic character in identi®cation of thegenus, but does not enable interspecies di�erentiation.Besides glucose, all bi®dobacteria from human origin
are also able to utilize galactose, lactose and, usually,fructose as carbon sources. The lactose transport systemfor B. bi®dum DSM 20082 was identi®ed recently as aproton symport, based on inhibition of lactose uptakeby inhibitors of ATP synthesis and by compounds thatinterfere with proton and metal ionophores [8]. Bi®do-bacterium spp. are, in some instances, also able to fer-ment complex carbohydrates; a recent study [9], inwhich 290 strains of 29 species of bi®dobacteria fromhuman and animal origin were surveyed for their abilityto ferment complex carbohydrates, has con®rmed thispotential. The substrates fermented by the largest num-ber of species were d-galactosamine, d-glucosamine,amylose and amylopectin. Porcine gastric mucin wasfermented only by B. bi®dum, whereas B. infantis wasthe only species that could ferment d-glucuronic acid.Strains of B. longum fermented arabinogalactan andarabic, ghatti and tragacanth gums.
Table 1. List of species (by alphabetical order) of the generaBi®dobacterium and Lactobacillus
Lactobacillus Bi®dobacterium
L. acetotolerans L. jenseniia B. adolescentisa
L. acidophilusa L. Johnsonii B. angulatuma
L. agilis L. kandleri B. animalisL. alimentarius L. ke®r B. asteroidesL. amylophilus L. ke®ranofaciens B. bi®duma
L. amylovorus L. malefermentans B. boumL. avarius L. mali B. brevaa
L. bifermentans L. minor B. catenulatuma
L. brevisa L. murinus B. choerinumL. buchneria L. orisa B. coryneformeL. casei subsp. caseia L. parabuchneria B. cuniculiL. collinoides L. paracaseia B. dentiuma
L. confusus L. pentosus B. gallicumL. coryniformis L. pontis B. gallinarumL. crispatusa L. plantaruma B. globosuma
L. curvatus L. reuteria B. indicumL. delbrueckii L. rhamnosusa B. infantisa
L. farciminis L. ruminis B. lactisL. fermentuma L. sake B. longuma
L. fructivorans L. salivariusa B. magnumL. fructosus L. sanfrancisco B. merycicumL. gallinarum L. sharpeae B. minimumL. gasseria L. suebicus B. pseudocatenulatuma
L. graminis L. vaccinostercus B. psudolongumL. halotolerans L. vaginalisa B. pullorumL. hamsteri L. viridescens B. ruminantiumL. helveticus B. saeculareL. hilgardii B. subtileL. homohiochii B. suisL. intestinalis B. thermophilum
a Species isolated from human sources
140 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

The optimum pH for growth is 6±7, with virtually nogrowth at pH 4.5±5.0 or below or at pH 8.0±8.5 orabove. Optimum growth temperature is 37±41�C, withmaximum growth at 43±45�C and virtually no growth at25±28�C or below. Comprehensive details of their biol-ogy are available in the extensive reviews by Rasic andKurmann [2] and Sgorbati et al. [7]. The latter haveprovided an updated report on the taxonomy status ofthe genus Bi®dobacterium, but their list of properlyidenti®ed bi®dobacteria does not include the moderatelyoxygen-tolerant species Bi®dobacterium lactis, whichwas identi®ed recently [10].
Genus LactobacillusIn 1990, Moro was the ®rst researcher to isolate faculta-
tive anaerobic straight rods from the faeces of breast-fedinfants, which he typi®ed as Bacillus acidophilus, a genericname for intestinal lactobacilli. Lactobacilli are in generalcharacterized as gram-positive, non-sporeforming andnon-¯agellated rods or coccobacilli [11]. The G+Ccontent of their DNA is usually between 32 and 51mol%. They are either aerotolerant or anaerobic andstrictly fermentative. Glucose is fermented pre-dominantly to lactic acid in the homofermentative case,or equimolar amounts of lactic acid, CO2 and ethanol(and/or acetic acid) in the heterofermentative counterpart.At present, 56 species of the genus Lactobacillus havebeen recognized (Table 1). Of these microorganisms, themost commonly suggested for dietary use areLactobacillusacidophilus strains, the de®nition of which has changedrecently.Comprehensive genetic studies have shown that the
original species actually consists of six DNA homologygroups, including L. crispatus, L. gallinarum, L. gasseri,L. amylovorus and L. johnsonii. Although these speciesare well de®ned, di�culties are often encountered inallocating newly isolated strains to each of these groups.Investigations based on agglutination tests, cell wallantigen and electrophoretic and antigenic characteristics
of d- and l-lactate dehydrogenases (LDH) also pointtoward heterogeneity of these species [12]. Lactobacillusacidophilus is a gram-positive rod with rounded endsthat occurs as single cells, as well as in pairs or in short
Table 2. Physiological and biochemical characteristics of Bi®dobacterium spp. and Lactobacillus acidophilusa
Character Bi®dobacterium spp. Lactobacillus acidophilus
Physiology Anaerobic Microaerophilic
Cell wall compositon:Peptidoglycan type Variable, basic amino acid in the tetrapeptide is either
ornithine or lysine, various types of cross-linkageLys-D Asp
Phospholipid composition/Teichoic acid Polyglycerolphospholipid and its lyso derivatives,alanylphosphatidylglycerol, lyso derivatives ofdiphosphatidylglycerol
Glycerol
DNA-base composition Mol% G+C(guanine+cytosine)
55±67 34±37
Lactic acid con®guration L DL
Sugar metabolism Heterofermentative Homofermentative
a Adapted from Kurmann and Rasic [17] and Mital and Garg [12]
Fig. 1. Formation of acetate and lactate from glucose by the bi®dumpathway. 1-hexokinase and fructose-6-phosphate isomerase, 2-fruc-tose-6-phosphate phosphoketolase, 3-transaldolase, 4-transketolase,5-ribose-5-phosphate isomerase, 6-ribulose-5-phosphate-3-epimerase,7-xylulose-5-phosphoketolase, 8-acetate kinase, 9-enzymes as inhomofermentative pathway (adapted from Rasic and Kurmann [2]).
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 141

chains. The typical size is 0.6±0.9 �m in width and 1.5±6.0 �m in length. It is non-¯agellated, non-motile andnon-sporeforming, and is intolerant to salt. This micro-organism does not contain cytochromes and, therefore, isbenzidine negative. In addition, it is microaerophilic, sosurface growth on solid media is generally enhanced byanaerobiosis or reduced oxygen pressure and 5±10%CO2. The physiological and biochemical characteristicsof this microorganism are given in Table 2. Most strainsof L. acidophilus can ferment amygdalin, cellobiose,fructose, galactose, glucose, lactose, maltose, mannose,salicin, sucrose, trehalose and aesculine [13]. Lactose isvirtually the only sugar present in milk, yet L. acid-ophilus has been reported to utilize sucrose more e�ec-tively than lactose [12]; such observations may beascribed to di�erences in �-galactosidase (EC 3.2.1.23)and �-fructofuranosidase (EC 3.2.1.26) activities. While�-fructofuranosidase is a constitutive enzyme, �-galac-tosidase may be induced in L. acidophilus [14]. More-over, both glucose and fructose moieties of sucrose areutilized by L. acidophilus, whereas the galactose moietyof lactose cannot be metabolized to an appreciabledegree. The glucose moiety is metabolized via the Emb-den±Meyerhof±Parnas pathway with lactic acid asessentially the sole end product. The yield of lactic acidis 1.8 mol/mol glucose, accompanied by minor amountsof other compounds. Acetaldehyde, a carbonyl ¯avour-ing molecule, may also result from metabolism of lac-tose, although in some instances it may be producedfrom metabolism of nitrogen-containing substances, e.g.threonine; a very high activity of threonine aldolase hasbeen found in L. acidophilus [15].Growth of L. acidophilus may occur at as high a tem-
perature as 45�C, but optimum growth occurs within35±40�C. Its acid tolerance varies from 0.3% to1.9% titratable acidity, with an optimum pH lyingat 5.5±6.0.
EcologyGenus Bi®dobacteriumBi®dobacteria are microorganisms of paramount
importance in the active and complex ecosystem of theintestinal tract of humans and other warm-bloodedanimals, and of honeybees [7]. They are distributed invarious ecological niches in the human gastrointestinaland genitourinary tracts, the exact ratio of which isdetermined mainly by age and diet. The indigenousmicro¯ora of infants is dominated by bi®dobacteria,which become established shortly after birth. Theirproliferation is stimulated by the glycoprotein compo-nents of �-casein in human colostrum and, to a lesserextent, human milk. The number of bi®dobacteriadecreases with increasing age of the individual andeventually becomes the third most abundant genus(accounting for ca. 25% of the total adult gut ¯ora)after the genera Bacteroides and Eubacterium [16]. The
onset of old age is characterized by signi®cant reduc-tions in the numbers of bi®dobacteria, whilst con-taminant bacteria such as clostridia and coliforms tendto increase, usually as a result of diminished secretion ofgastric juice in this age group [17]. Furthermore, thepro®le of constituent species changes; B. infantis and B.breve, typical of infants, are replaced by B. adolescentisin adults, whereas B. longum persists lifelong [18]. Thisage pro®le may obviously be in¯uenced by the dietaryintake of bi®dogenic factors [19] and by the hostphysiology [17].
Genus LactobacillusLactobacilli are distributed in various ecological
niches throughout the gastrointestinal and genital tractsand constitute an important part of the indigenousmicro¯ora of man and higher animals. Their distribu-tion is a�ected by several environmental factors, whichinclude pH, oxygen availability, level of speci®c sub-strates, presence of secretions and bacterial interactions.They are rarely associated with cases of gastrointestinaland extraintestinal infection, and strains employedtechnologically are regarded as non-pathogenic and safemicroorganisms. Furthermore, they have the reputationof health promoters, especially in the human gastro-intestinal and genitourinary tracts [20].
Growth performanceE�ect of substrate on growth
Genus Bi®dobacteriumThe fact that bi®dobacteria can grow in a semi-syn-
thetic medium containing only lactose, three free aminoacids (cysteine, glycine and tryptophan), several vitaminsand nucleotides, and some minerals contrasts with thegenerally recognized fastidiousness of lactobacilli withregard to nutritional requirements. Certain strains werefound to grow on a simpli®ed medium comprised oflactose (as fermentable carbohydrate), bu�ers, minerals,ammonium salts, cysteine, and the vitamins biotin andcalcium pantothenate [21]. Bi®dobacterium bi®dum var.pennsylvanicus, a mutant strain, requires pantothenateand N-acetylglucosamine-containing saccharides forgrowth and synthesis of its cell wall. The B. bi®dumstrain discovered by Tissier in 1899, as well as someother bi®dobacteria, require a nitrogen source (i.e. pep-tides), yet neither pantothenate nor N-acetylglucos-amine-containing saccharides are essential. A strikingdi�erence between lactobacilli and some strains of bi®do-bacteria is the ability of the latter to grow in a mediumcontaining nitrogen in ammonium form; the remainingbi®dobacteria strains require nitrogen from organicsources [22]. Evidence indicates that the nutritionalproperties of bi®dobacteria are species-dependent, soany new strain discovered or produced needs to beextensively studied in terms of its minimal nutritional
142 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

requirements; this is the case of the newly identi®edBi®dobacterium lactis strain [23].
Lactobacillus acidophilusLactobacilli have complex growth requirements. They
require low oxygen tension [13,24], fermentable carbo-hydrates, protein and its breakdown products [25], anumber of vitamins of the B-complex [26], nucleic acidderivatives [27], unsaturated free fatty acids [28], andminerals such as magnesium, manganese and iron [29]for their growth. Increased amount of thiol groups pre-sent in whey protein-enriched milks favours the growthof L. acidophilus, whereas peptone and trypsin stimulateits acid production [22]. Addition of tomato juice (as asource of simple sugars, minerals and vitamins of the B-complex) to skimmed milk has provided evidence forenhancement of both growth of (i.e. higher viablecounts and shorter generation times) and activity by (i.e.improved sugar utilization and lower pH) L. acidophilus[30]. These essential nutrients should, therefore, beavailable in the medium for establishment of a pre-dominant micro¯ora of lactobacilli; in their recent studyon optimization of cultivation of L. acidophilus, Tail-landier et al. [31] concluded that the optimum condi-tions are pH 6.0, 30�C, 40 g/L of glucose, 20 g/L ofpeptone, 20 g/L of yeast extract, 5 g/L of sodium acetateand 3 g/L of sodium citrate.
Milk as an unde®ned nutrient mediumBi®dobacteria are used in milk fermentations to a
limited extent because of their slow growth in thatmatrix which, in many instances, may be considered asan arti®cial medium. However, milk is an essentiallysatisfactory medium because it contains all essentialnutrients, except that amino acids and small peptidesare present at insu�cient concentrations (ca. 0.1 g/L) tosupport extended growth of bi®dobacteria [2,23].Nevertheless, cases of adaptation to milk upon multipletransfers have been reported [22]. Comparison of var-ious milk types (bovine, ovine and caprine milks) withrespect to their available nitrogen fraction has also beenexploited in an attempt to further probe the in¯uence ofthe raw material upon selected probiotic strains, but thehigher protein and vitamin contents of ovine milk werenot su�cient to sustain growth of B. lactis at the highrates required [32]. Moreover, the excess of fatty acidresidues present in caprine milk were indicated asresponsible for the poor growth of L. acidophilus.In general, bi®dobacteria grow better in rich synthetic
media, viz. TPY and MRS broths, than in milk; however,said media are complex and costly for large-scale propaga-tion of bi®dobacteria. Moreover, unless the cells harvestedfrom such media are extensively washed before incorpora-tion, they may confer o�-¯avours to the ®nished dairyproducts. To manufacture a quality product, both in termsof texture and viability of bi®dobacteria, a milk-based
medium is usually required because the casein contentof milk protein is higher than that of synthetic media(which are generally low in solids). Thus, improvementof growth conditions for the di�erent strains of bi®do-bacteria in milk, in particular via addition of variouseasily-available nitrogen sources or redox potential-lowering substances, has provided the impetus for manystudies [23,24,32±34], and less fastidious, mildly-acid-ifying strains can thus be claimed as key candidates forsuccessful bi®dobacteria-containing (bio)products.
Bi®dogenic and growth factors
De®nitionThere are clear di�erences between bi®dogenic factors
and growth factors for bi®dobacteria in terms of natureand function. Bi®dogenic factors are de®ned as com-pounds, usually of a carbohydrate nature, that survivedirect metabolism by the host and reach the large bowelor cecum, where they are preferentially metabolized bybi®dobacteria as source of energy. Bi®dogenic factorsmay fall under the new concept of prebiotics, which arede®ned as non-digestible food ingredients that bene®-cially a�ect the host by selectively stimulating thegrowth and/or activity of one, or a limited number of,bacteria in the colon, and which may thus improve thehealth of the host [35]. In contrast, growth factors arecompounds that promote the growth of bi®dobacteriain vitro but cannot be delivered to the large bowel orcecum to selectively promote proliferation of bi®do-bacteria [19]. Studies on the sources and applications foreither of these types of compounds have been welldocumented [19], yet comprehensive reviews of the stateof the art in this ®eld are still lacking. Table 3 sum-marizes established knowledge and more recent devel-opments in the isolation, identi®cation and applicationof both bi®dogenic and growth factors.
Bi®dogenic factorsFructooligosaccharides. The major carbohydrate
sources for bacterial growth in the colon are provided bydietary or endogenous means. In this respect, non-diges-tible oligosaccharides have been used in the diet speci®-cally to increase relative numbers of bi®dobacteria in thegut micro¯ora [36±39]. In particular, promising resultshave been achieved in vitro using inulin [37] and especiallyfructooligosaccharides (FOS), in both batch culture [38]and chemostat culture systems [39]. Among the di�erenttypes of (linear and branched) FOS studied, oligo-fructose, a linear chain molecule comprising glucose andfructose at a degree of polymerization of four, exhibitedthe highest bi®dogenic e�ect. Earlier in vivo clinical stu-dies reported a ten-fold increase of the bi®dobacterialpopulation in the large bowel upon addition of neosu-gars (i.e. short-chain FOS) to the human diet at a ratioof 15 g/d [36], which correlated well with the increased
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 143
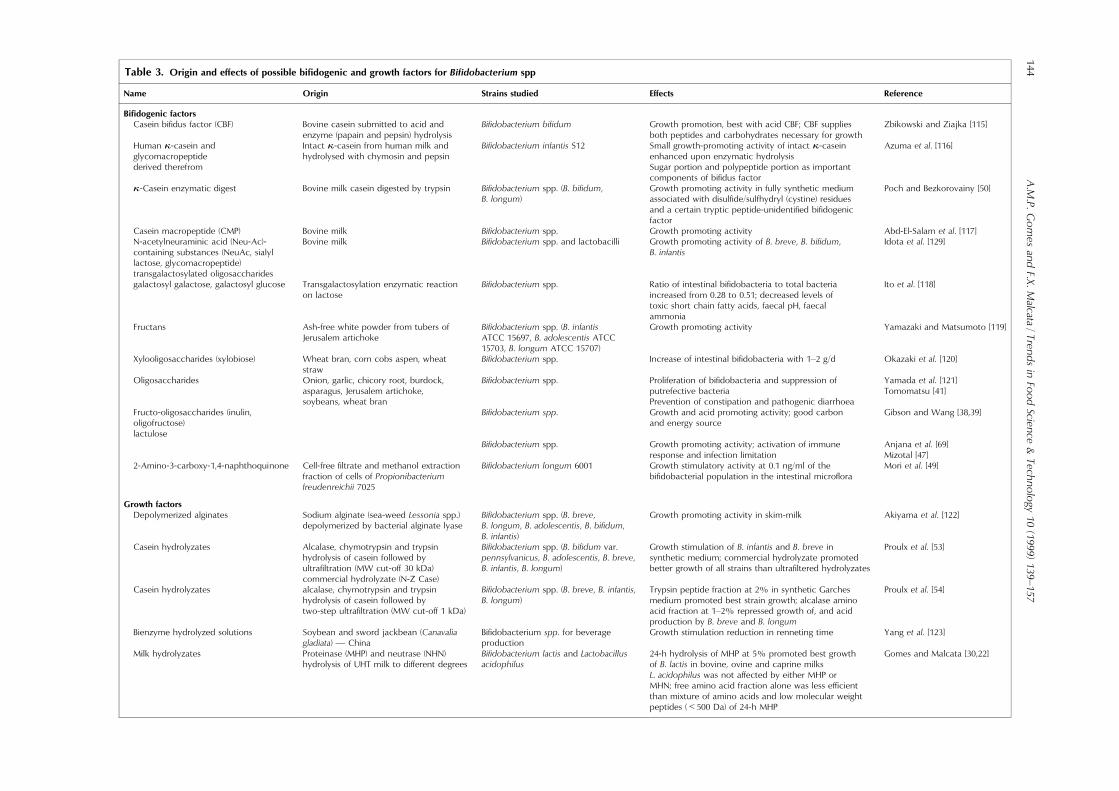
Table 3. Origin and e�ects of possible bi®dogenic and growth factors for Bi®dobacterium spp
Name Origin Strains studied E�ects Reference
Bi®dogenic factorsCasein bi®dus factor (CBF) Bovine casein submitted to acid and
enzyme (papain and pepsin) hydrolysisBi®dobacterium bi®dum Growth promotion, best with acid CBF; CBF supplies
both peptides and carbohydrates necessary for growthZbikowski and Ziajka [115]
Human �-casein andglycomacropeptidederived therefrom
Intact �-casein from human milk andhydrolysed with chymosin and pepsin
Bi®dobacterium infantis S12 Small growth-promoting activity of intact �-caseinenhanced upon enzymatic hydrolysisSugar portion and polypeptide portion as importantcomponents of bi®dus factor
Azuma et al. [116]
�-Casein enzymatic digest Bovine milk casein digested by trypsin Bi®dobacterium spp. (B. bi®dum,B. longum)
Growth promoting activity in fully synthetic mediumassociated with disul®de/sulfhydryl (cystine) residuesand a certain tryptic peptide-unidenti®ed bi®dogenicfactor
Poch and Bezkorovainy [50]
Casein macropeptide (CMP) Bovine milk Bi®dobacterium spp. Growth promoting activity Abd-El-Salam et al. [117]N-acetylneuraminic acid (Neu-Ac)-containing substances (NeuAc, sialyllactose, glycomacropeptide)
Bovine milk Bi®dobacterium spp. and lactobacilli Growth promoting activity of B. breve, B. bi®dum,B. infantis
Idota et al. [129]
transgalactosylated oligosaccharidesgalactosyl galactose, galactosyl glucose Transgalactosylation enzymatic reaction
on lactoseBi®dobacterium spp. Ratio of intestinal bi®dobacteria to total bacteria
increased from 0.28 to 0.51; decreased levels oftoxic short chain fatty acids, faecal pH, faecalammonia
Ito et al. [118]
Fructans Ash-free white powder from tubers ofJerusalem artichoke
Bi®dobacterium spp. (B. infantisATCC 15697, B. adolescentis ATCC15703, B. longum ATCC 15707)
Growth promoting activity Yamazaki and Matsumoto [119]
Xylooligosaccharides (xylobiose) Wheat bran, corn cobs aspen, wheatstraw
Bi®dobacterium spp. Increase of intestinal bi®dobacteria with 1±2 g/d Okazaki et al. [120]
Oligosaccharides Onion, garlic, chicory root, burdock,asparagus, Jerusalem artichoke,soybeans, wheat bran
Bi®dobacterium spp. Proliferation of bi®dobacteria and suppression ofputrefective bacteriaPrevention of constipation and pathogenic diarrhoea
Yamada et al. [121]Tomomatsu [41]
Fructo-oligosaccharides (inulin,oligofructose)lactulose
Bi®dobacterium spp. Growth and acid promoting activity; good carbonand energy source
Gibson and Wang [38,39]
Bi®dobacterium spp. Growth promoting activity; activation of immuneresponse and infection limitation
Anjana et al. [69]Mizotal [47]
2-Amino-3-carboxy-1,4-naphthoquinone Cell-free ®ltrate and methanol extractionfraction of cells of Propionibacteriumfreudenreichii 7025
Bi®dobacterium longum 6001 Growth stimulatory activity at 0.1 ng/ml of thebi®dobacterial population in the intestinal micro¯ora
Mori et al. [49]
Growth factorsDepolymerized alginates Sodium alginate (sea-weed Lessonia spp.)
depolymerized by bacterial alginate lyaseBi®dobacterium spp. (B. breve,B. longum, B. adolescentis, B. bi®dum,B. infantis)
Growth promoting activity in skim-milk Akiyama et al. [122]
Casein hydrolyzates Alcalase, chymotrypsin and trypsinhydrolysis of casein followed byultra®ltration (MW cut-o� 30 kDa)commercial hydrolyzate (N-Z Case)
Bi®dobacterium spp. (B. bi®dum var.pennsylvanicus, B. adolescentis, B. breve,B. infantis, B. longum)
Growth stimulation of B. infantis and B. breve insynthetic medium; commercial hydrolyzate promotedbetter growth of all strains than ultra®ltered hydrolyzates
Proulx et al. [53]
Casein hydrolyzates alcalase, chymotrypsin and trypsinhydrolysis of casein followed bytwo-step ultra®ltration (MW cut-o� 1 kDa)
Bi®dobacterium spp. (B. breve, B. infantis,B. longum)
Trypsin peptide fraction at 2% in synthetic Garchesmedium promoted best strain growth; alcalase aminoacid fraction at 1±2% repressed growth of, and acidproduction by B. breve and B. longum
Proulx et al. [54]
Bienzyme hydrolyzed solutions Soybean and sword jackbean (Canavaliagladiata) Ð China
Bi®dobacterium spp. for beverageproduction
Growth stimulation reduction in renneting time Yang et al. [123]
Milk hydrolyzates Proteinase (MHP) and neutrase (NHN)hydrolysis of UHT milk to di�erent degrees
Bi®dobacterium lactis and Lactobacillusacidophilus
24-h hydrolysis of MHP at 5% promoted best growthof B. lactis in bovine, ovine and caprine milksL. acidophilus was not a�ected by either MHP orMHN; free amino acid fraction alone was less e�cientthan mixture of amino acids and low molecular weightpeptides (<500 Da) of 24-h MHP
Gomes and Malcata [30,22]
144A.M.P.
Gomes
andF.X
.Malcata
/Tren
dsin
FoodScien
ce&Tech
nology
10(1999)
139±157

release of volatile fatty acids and concomitant decreasein pH (by 0.3 units), together with a reduction in thecounts of Enterobacteriaceae from 1�108 to 4�107 cfu/g.All FOS studied are oligosaccharides that occur in
nature, mainly of plant origin (Table 3), and are manu-factured industrially from sucrose syrup obtained fromsugar beet by an enzymatic process in which �-fructo-furanosidase (EC 3.2.1.26) derived from Aspergillusniger plays an essential role. These compounds are�(2!1) linked fructans, typically polydisperse, withdegrees of polymerization in the range 2±35; they areresistant to the strong hydrolytic capacities of thehuman intestinal enzymes and, therefore, reach thecolon essentially unaltered, where they behave likesoluble dietary ®bre and are digested by the colonic¯ora [40]. Other properties, applications and healthaspects closely related to their e�ect on bi®dobacteriagrowth in the colon have been discussed in severalreviews [41,42]. The increased intake of oligofructose-containing foods, or alternatively the increased incor-poration of these prebiotics into foods with a signi®cantcarbohydrate content, e.g. ice cream, cakes, infant for-mula feeds and milk-based products [43], o�er arational alternative to the more commonly employedincorporation of viable bi®dobacteria into fermentedmilks. In fact, the main advantage is that survival in theproduct becomes easier to assure.Xylooligosaccharides (e.g. xylobiose) and transga-
lactosylated oligosaccharides (e.g. lactosucrose) are alsoknown as e�ective promoters of proliferation of intest-inal bi®dobacteria (Table 3). The former compoundsoccur naturally, whereas the latter are available only viaindustrial synthesis. Their importance as bi®dogenicfactors is re¯ected in the large number of publicationsfocussed on the preparation of galactooligosaccharidesthrough bioconversion of lactose using various micro-bial enzymes [44±46]. Oligosaccharides that containmaltose, mannose and soya are also possible bi®dogenicfactors (Table 3).Several studies, reviewed by Modler [19] and Mizota
[47], reported that the lactose-derivative lactulose, adissaccharide composed of galactose and fructose (4-O-�-d-galactopyranosyl-d-fructose), can exert bi®dogenice�ects on bi®dobacteria resident in the lower intestineand hence can produce many of the desirable e�ectsascribed to FOS. These e�ects include improvement ofhyperammonaemia and suppression of production oftoxic short-chain fatty acids (butyric and valeric acids);in addition, this compound has many other health andmedical e�ects [47]. Lactulose has been used to increasethe numbers of viable lactobacilli in the intestine ofbottle-fed infants; other candidates, including suchsugar alcohols as lactitol and glucosyl-inositol (Table 3),may also be useful in increasing bi®dobacteria numbers.Neosugars, FOS and lactulose are commercially avail-
able and widely used in Japan and Europe as bi®dogenic
compounds. Although safe from a toxicological stand-point and responsible for many positive e�ects, they arenot necessarily ideal bi®dogenic factors: to be e�ective, thebi®dogenic factor must promote a selective fermentationin mixed culture so that the pro®le of the large intestinalmicro¯ora is altered towards a potentially healthiercommunity. Unfortunately, both types of compoundscan be metabolized by a number of gas-producing bac-teria and hence may cause intestinal discomfort.Quinone compounds. Roland et al. [48] reported
consistent data in vitro (mixed culture of B. bi®dum and48 h-old cultures of Propionibacterium freudenreichii inMRS broth) and in vivo (administration of 5�1010 cfuof propionibacteria to 19 males over 2 weeks) inattempts to check the validity of the hypothesis thatpropionibacterium (a microorganism used in the manu-facture of Swiss-type cheese) promotes the growth ofbi®dobacteria. A parallel study showed that, in a humanfaecal culture contained in a two-stage, continuous cul-ture system, addition of a quinone compound producedby P. freudenreichii cells stimulated the growth of bi®-dobacteria, thus suggesting a bi®dogenic e�ect regard-ing regulation of their numbers in the intestine [49](Table 3). However, more stringent studies are neededto clarify the speci®c action of this, and other, relatedquinone compounds upon bi®dobacterial growth.
Growth factorsWithin the range of compounds investigated, deriva-
tives of human and bovine milks (Table 3) have provengood candidates for the enhancement of growth of bi®-dobacteria in vitro. According to Poch and Bezkorovainy[50], growth-promoting activity of bovine milk �-caseinwas ascribed to its conjugated cystine residues, anobservation more recently con®rmed and re®ned in astudy aimed at comparing the growth-promoting e�ectsof various biological materials on B. longum [51] using asynthetic medium. All tentative growth-promoting fac-tors studied failed to exhibit their growth-promotingactivity when their dissul®de bonds were reduced oralkylated, thus suggesting that their composition in sulfur-containing peptides might be the key to said e�ect. Themajor whey proteins, �-lactalbumin and �-lactoglobulin,were also found to be excellent growth promoters, a traitvalidated after the studies by Petschow and Talbott [52]on the e�ectiveness of whey fractions of both humanmilk and bovine milk. Yeast extract, a commercial pro-duct, was found to be an e�ective growth promoter, andis most often used at concentrations that vary between0.1 and 0.5%(v/v). A previous study [33] has shown,however, that addition of yeast extract at 0.25%(v/v)did not stimulate growth of B. infantis, even though acidproduction was enhanced; it further demonstrated thegood growth-promoting activity of �-glycerophosphatein the presence of cysteine for B. bi®dum and B. infantis,but not for B. longum ATCC 15708.
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 145

Other biological compounds identi®ed as growth fac-tors for bi®dobacteria include threonine [25], cysteine[33], enzyme-treated chlorella, peptone and trypticase[19], as well as dextrin, maltose and extracts from car-rots (coenzyme A), potatoes and corn [2], and commer-cial casein hydrolysates and ultra®ltered hydrolysates[53,54] these hydrolysates are, in general, produced bybreaking down casein with proteases, peptidases orstrong acids to di�erent yields. Proulx et al. [53,54]claimed that peptides obtained from casein hydrolysatesmight be a preferable source of nitrogen than freeamino acids for dairy-related bi®dobacteria (Table 3).Apart from that, the development of hydrolysates withbetter quality at lower cost is essential. Gomes et al. [23]expanded on this work using milk (as economicalsource), hydrolysed to several extents by two alternativeproteinases, as nitrogen source to promote growth of,and acidi®cation by B. lactis and L. acidophilus; theseauthors established that both the nature of the enzymeand the degree of hydrolysis are important criteriatowards attainment of consistent growth of the afore-mentioned strains, an observation that con®rms thegrowth-limiting characteristics of available nitrogen inthe form of low molecular weight peptides (<500 Da)and free amino acids that are more easily assimilated byB. lactis (Table 3).
Nutritional and health values of fermented milkproductsHistorical backgroundNutritional and health aspects of functional foods
incorporating probiotic bacteria have received con-siderable attention, and eventually led to numerouspurported claims in the literature. These potentialadvantageous properties, initially summarized by Gurr[55] and more recently reviewed in detail by Gilliland
[56], Marteau and Rambaud [57] and Yaeshima [58], aresummarized in Table 4 and will be critically described inthe following subsections. Despite the many studies onthe bene®cial nutropharmaceutical aspects of probioticbacteria, the results obtained are highly variable andsometimes inconsistent with one another; hence, noclear, unequivocal evidence for the actual existence ofsome of these bene®ts is yet available, which then ren-ders health claims more di�cult to establish. To elim-inate these drawbacks, e�orts have been pursuedworldwide to rationally organize future research usingclearer and more reliable bases (via well-designed, ran-domized and placebo-controlled, double-blind studies)and exploiting statistically validated methods, with spe-cial emphasis on intestinal integrity and immune mod-ulation. Increasing numbers of colonization and dose-response studies de®ning the required doses have beenpublished [59].
Nutritional valueThe nutritional bene®ts of probiotics have been
mostly studied in milk-based products fermented withlactobacilli and bi®dobacteria. These products contain agreat many chemical compounds depending on the typeof milk used (usually cow's, ewe's or goat's), on the typeof microorganisms added (and their speci®c biochemicalactivities) and on the manufacturing processesemployed. They are characterized by a lower level ofresidual lactose and higher levels of free amino acidsand certain vitamins than non-fermented milks. Fur-thermore, they preferentially contain L(+)-lactic acid(that is more easily metabolized by human beings thanD(-)-lactic acid) produced by bi®dobacteria in additionto acetic acid [2], thereby preventing manifestation ofmetabolic acidosis in infants below one year of age.Moreover, the L(+)-lactic acid absorbed in the intestine
Table 4. Potential health and nutritional bene®ts of functional foods prepared with probiotic bacteria
Bene®cial e�ect Possible causes and mechanisms
Improved digestibility Partial breakdown of proteins, fats and carbohydratesImproved nutritional value Higher levels of B vitamins and certain free amino acids, viz. methionine, lysine and tryptophanImproved lactose utilization Reduced lactose in product and further availability of lactaseAntagonistic action towardsenteric pathogens
Disorders, such as functional diarrhoea, mucous colitis, ulcerated colitis, diverticulitis andantibiotic colitis controlled by acidity, microbial inhibitors and prevention of pathogen adhesionor pathogen activation
Colonisation in gut Survival in gastric acid, resistance to lysozyme and low surface tension of intestine, adherenceto intestinal mucosa, multiplication in the intestinal tract, immune modulation
Anticarcinogenic e�ect Conversion of potential pre-carcinogens into less harmful compoundsInhibitory action towards some types of cancer, in particular cancers of the gastrointestinaltract by degradation of pre-carcinogens, reduction of carcinogen-promoting enzymes andstimulation of the immune system
Hypocholesterolemic action Production of inhibitors of cholesterol synthesis. Use of cholesterol by assimilation andprecipitation with deconjugated bile salts
Immune modulation Enhancement of macrophage formation, stimulation of production of suppressor cells and -interferon
Vaccine vehicle Naturally occurring or rDNA vaccinal epitopes
146 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

is used as energy source, with an energy yield of 15 kJ/g,that compares well with 16 kJ/g for lactose [55].Lactobacillus acidophilus and bi®dobacteria have also
been reported to synthesize folic acid, niacin, thiamine,ribo¯avin, pyridoxine and vitamin K, which are slowlyabsorbed by the body [2,60]. The vitamins of the B-complex are frequently obtained as natural ingredientsin foods, so addition of bi®dobacteria to the diet willmore e�ectively help meet those requirements. Thebioavailability to the host of such minerals as calcium,zinc, iron, manganese, copper and phosphorusmay also beenhanced upon consumption of fermented dairy productscontaining bi®dobacteria via lowering the gastric pH(which facilitates ionization of minerals, a requirement forabsorption) and improved digestibility of the protein [2].
Therapeutic e�ectsE�ects on intestinal disturbances and intestinal infectionsThe intact intestinal epithelium, together with the
normal intestinal micro¯ora represents a barrier tomovement of pathogenic bacteria, antigens and othernoxious substances from the gut lumen to the blood. Inhealthy subjects this barrier is stable, thus protecting thehost and assuring normal intestinal function. Wheneither the normal micro¯ora or the epithelial cells aredisturbed, as triggered by dietary antigens, pathogens,chemicals or radiation, defects in the barrier mechan-isms become apparent; altered permeability facilitatesblood invasion by pathogens, foreign antigens and otherharmful substances. Several reviews [20,57] have docu-mented the use of probiotic bacteria to treat intestinaldisorders, e.g. acute rotavirus diarrhoea in children, aswell as food allergy and colonic disorders driven bypelvic radiotherapy and sometimes associated withdevelopment of colon cancer. These examples areamong those included in Table 5 that illustrate the stateof the art in what concerns application of several pro-biotic strains for therapeutic purposes. In all such dis-ease states, altered intestinal micro¯ora, impaired gutbarrier and di�erent types of intestinal in¯ammation areoften present [20], thereby o�ering a rationale for thee�ective use of probiotic bacteria, not only for treat-ment but especially for prevention of such changes.Among the possible mechanisms responsible forfavourable clinical responses, one may quote (i) pro-motion of the immunologic and nonimmunologicdefense barriers in the gut [61,62] (ii) capacity to preventpathogen adherence or pathogen activation via produc-tion of inhibitory metabolites, such as organic acids,hydrogen peroxide, bacteriocins and deconjugated bilesalts [63±65], or via ferrous iron uptake thus making itunavailable for pathogens [66], and (iii) modi®cation ofbacterial enzyme activity and subsequent in¯uence on gutmucosal permeability [20] (see Table 5). Probiotic bacteriaare able to survive gastric conditions (characterized by pH1.5±2.0 during fasting and 4.0±5.0 after a meal), colonize
the intestine (at least temporarily) by adhering to theintestinal epithelium in a process mediated by adhesins onthe surface of the bacteria (proteins, polysaccharides andcell wall-associated components), and o�er new dietaryalternatives for the stabilization of the intestinal micro-¯ora. In clinical nutrition, for instance, special diets areused for the treatment of intestinal in¯ammation,radiation enteritis and in¯ammatory bowel disease:probiotic bacteria do play a role in alleviating thesedisturbances, so the introduction of such strains intoclinical foods and special dietary foods is in order.
Potential antitumor activityThe colonic ¯ora has been shown to be involved in
colonic carcinogenesis [20] This e�ect is mediated bymicrobial enzymes such as �-glucuronidase, �-glucosi-dase, nitroreductase and urease, which convert pro-carcinogens into carcinogens. Experiments performed inanimal models showed that a few strains of L. acid-ophilus and Bi®dobacterium spp. are able to decrease thelevels of enzymes responsible for activation of someprocarcinogens, and consequently decrease the risk oftumor development [12,17]. Studies in the human bodyusing the same enzymes as end-points demonstrated ageneral reduction in microbial enzyme activities andconcomitant decrease in faecal mutagenicity [57,67] Thebene®cial e�ect could be accounted for by a favourablechange in the composition of the intestinal ¯ora followingintroduction of the aforementioned bacteria.Furthermore, it has been demonstrated, at least for
Lactobacillus casei (Shirota strain), that there is apotential opportunity in the area of `dietary prevention'of cancer. Morotomi [68] concluded that the parenteraladministration of this strain had antitumor and immu-nostimulating e�ects on experimentally implantedtumors; similar e�ects have been observed in the case oforal administration. The feeding of B. longum to ratswas recently shown to signi®cantly reduce formation ofsuch preneoplastic markers as azoxymethane-inducedcolonic aberrant crypt foci; furthermore, the productionof glutathione S-transferase, a Phase II enzyme marker,was also increased [69].The mechanisms that appear to be involved in anti-
tumor activity of systemically administered bi®dobacteriainclude activation of such non-speci®c cellular factors aspolymorphonuclear leucocytes [70], macrophages [71]and natural killer cells via regulation of -interferonproduction [72]; -interferon also exhibits antiviral andantiproliferative e�ects. Orally administered bi®do-bacteria play a role in increasing, to some extent, theproduction of IgA antibodies [73] and the functions ofPeyer's patch cells [74,75]. It should be noted that closelyrelated strains of Bi®dobacterium spp. and L. acidophilusdi�er between each other in immunostimulatory prop-erties, so some strains may not have all those propertiesnecessary for activity. Marin et al. [76] have recently
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 147
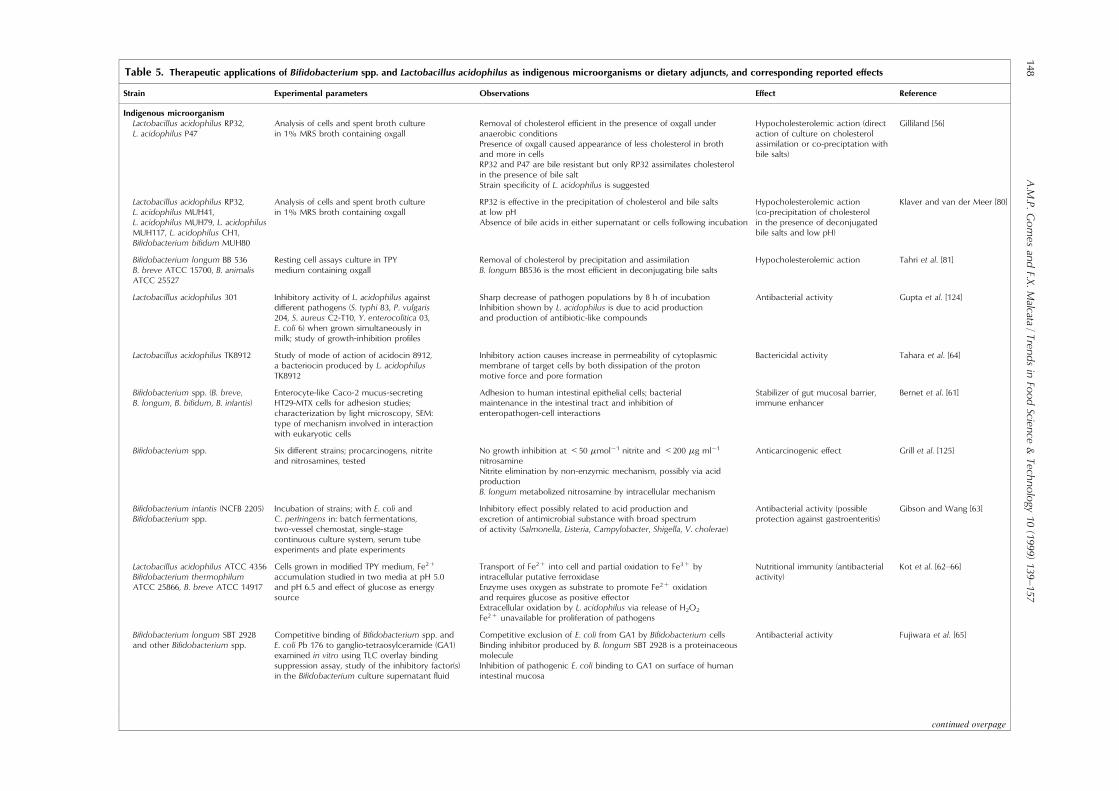
Table 5. Therapeutic applications of Bi®dobacterium spp. and Lactobacillus acidophilus as indigenous microorganisms or dietary adjuncts, and corresponding reported e�ects
Strain Experimental parameters Observations E�ect Reference
Indigenous microorganismLactobacillus acidophilus RP32,L. acidophilus P47
Analysis of cells and spent broth culturein 1% MRS broth containing oxgall
Removal of cholesterol e�cient in the presence of oxgall underanaerobic conditionsPresence of oxgall caused appearance of less cholesterol in brothand more in cellsRP32 and P47 are bile resistant but only RP32 assimilates cholesterolin the presence of bile saltStrain speci®city of L. acidophilus is suggested
Hypocholesterolemic action (directaction of culture on cholesterolassimilation or co-preciptation withbile salts)
Gilliland [56]
Lactobacillus acidophilus RP32,L. acidophilus MUH41,L. acidophilus MUH79, L. acidophilusMUH117, L. acidophilus CH1,Bi®dobacterium bi®dum MUH80
Analysis of cells and spent broth culturein 1% MRS broth containing oxgall
RP32 is e�ective in the precipitation of cholesterol and bile saltsat low pHAbsence of bile acids in either supernatant or cells following incubation
Hypocholesterolemic action(co-precipitation of cholesterolin the presence of deconjugatedbile salts and low pH)
Klaver and van der Meer [80]
Bi®dobacterium longum BB 536B. breve ATCC 15700, B. animalisATCC 25527
Resting cell assays culture in TPYmedium containing oxgall
Removal of cholesterol by precipitation and assimilationB. longum BB536 is the most e�cient in deconjugating bile salts
Hypocholesterolemic action Tahri et al. [81]
Lactobacillus acidophilus 301 Inhibitory activity of L. acidophilus againstdi�erent pathogens (S. typhi 83, P. vulgaris204, S. aureus C2-T10, Y. enterocolitica 03,E. coli 6) when grown simultaneously inmilk; study of growth-inhibition pro®les
Sharp decrease of pathogen populations by 8 h of incubationInhibition shown by L. acidophilus is due to acid productionand production of antibiotic-like compounds
Antibacterial activity Gupta et al. [124]
Lactobacillus acidophilus TK8912 Study of mode of action of acidocin 8912,a bacteriocin produced by L. acidophilusTK8912
Inhibitory action causes increase in permeability of cytoplasmicmembrane of target cells by both dissipation of the protonmotive force and pore formation
Bactericidal activity Tahara et al. [64]
Bi®dobacterium spp. (B. breve,B. longum, B. bi®dum, B. infantis)
Enterocyte-like Caco-2 mucus-secretingHT29-MTX cells for adhesion studies;characterization by light microscopy, SEM:type of mechanism involved in interactionwith eukaryotic cells
Adhesion to human intestinal epithelial cells; bacterialmaintenance in the intestinal tract and inhibition ofenteropathogen-cell interactions
Stabilizer of gut mucosal barrier,immune enhancer
Bernet et al. [61]
Bi®dobacterium spp. Six di�erent strains; procarcinogens, nitriteand nitrosamines, tested
No growth inhibition at <50 �molÿ1 nitrite and <200 �g mlÿ1
nitrosamineNitrite elimination by non-enzymic mechanism, possibly via acidproductionB. longum metabolized nitrosamine by intracellular mechanism
Anticarcinogenic e�ect Grill et al. [125]
Bi®dobacterium infantis (NCFB 2205)Bi®dobacterium spp.
Incubation of strains; with E. coli andC. perfringens in: batch fermentations,two-vessel chemostat, single-stagecontinuous culture system, serum tubeexperiments and plate experiments
Inhibitory e�ect possibly related to acid production andexcretion of antimicrobial substance with broad spectrumof activity (Salmonella, Listeria, Campylobacter, Shigella, V. cholerae)
Antibacterial activity (possibleprotection against gastroenteritis)
Gibson and Wang [63]
Lactobacillus acidophilus ATCC 4356Bi®dobacterium thermophilumATCC 25866, B. breve ATCC 14917
Cells grown in modi®ed TPY medium, Fe2+
accumulation studied in two media at pH 5.0and pH 6.5 and e�ect of glucose as energysource
Transport of Fe2+ into cell and partial oxidation to Fe3+ byintracellular putative ferroxidaseEnzyme uses oxygen as substrate to promote Fe2+ oxidationand requires glucose as positive e�ectorExtracellular oxidation by L. acidophilus via release of H2O2
Fe2+ unavailable for proliferation of pathogens
Nutritional immunity (antibacterialactivity)
Kot et al. [62±66]
Bi®dobacterium longum SBT 2928and other Bi®dobacterium spp.
Competitive binding of Bi®dobacterium spp. andE. coli Pb 176 to ganglio-tetraosylceramide (GA1)examined in vitro using TLC overlay bindingsuppression assay, study of the inhibitory factor(s)in the Bi®dobacterium culture supernatant ¯uid
Competitive exclusion of E. coli from GA1 by Bi®dobacterium cellsBinding inhibitor produced by B. longum SBT 2928 is a proteinaceousmoleculeInhibition of pathogenic E. coli binding to GA1 on surface of humanintestinal mucosa
Antibacterial activity Fujiwara et al. [65]
continued overpage
148A.M.P.
Gomes
andF.X
.Malcata
/Tren
dsin
FoodScien
ce&Tech
nology
10(1999)
139±157
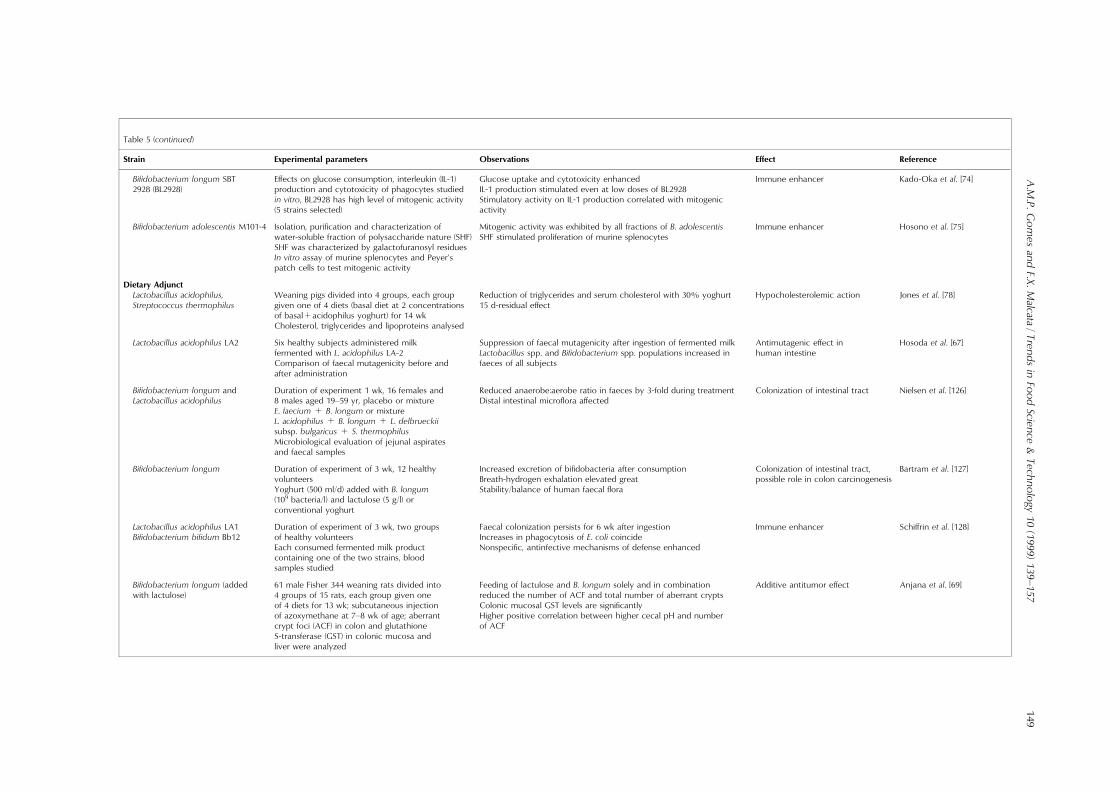
Table 5 (continued)
Strain Experimental parameters Observations E�ect Reference
Bi®dobacterium longum SBT2928 (BL2928)
E�ects on glucose consumption, interleukin (IL-1)production and cytotoxicity of phagocytes studiedin vitro, BL2928 has high level of mitogenic activity(5 strains selected)
Glucose uptake and cytotoxicity enhancedIL-1 production stimulated even at low doses of BL2928Stimulatory activity on IL-1 production correlated with mitogenicactivity
Immune enhancer Kado-Oka et al. [74]
Bi®dobacterium adolescentis M101-4 Isolation, puri®cation and characterization ofwater-soluble fraction of polysaccharide nature (SHF)SHF was characterized by galactofuranosyl residuesIn vitro assay of murine splenocytes and Peyer'spatch cells to test mitogenic activity
Mitogenic activity was exhibited by all fractions of B. adolescentisSHF stimulated proliferation of murine splenocytes
Immune enhancer Hosono et al. [75]
Dietary AdjunctLactobacillus acidophilus,Streptococcus thermophilus
Weaning pigs divided into 4 groups, each groupgiven one of 4 diets (basal diet at 2 concentrationsof basal+acidophilus yoghurt) for 14 wkCholesterol, triglycerides and lipoproteins analysed
Reduction of triglycerides and serum cholesterol with 30% yoghurt15 d-residual e�ect
Hypocholesterolemic action Jones et al. [78]
Lactobacillus acidophilus LA2 Six healthy subjects administered milkfermented with L. acidophilus LA-2Comparison of faecal mutagenicity before andafter administration
Suppression of faecal mutagenicity after ingestion of fermented milkLactobacillus spp. and Bi®dobacterium spp. populations increased infaeces of all subjects
Antimutagenic e�ect inhuman intestine
Hosoda et al. [67]
Bi®dobacterium longum andLactobacillus acidophilus
Duration of experiment 1 wk, 16 females and8 males aged 19±59 yr, placebo or mixtureE. faecium + B. longum or mixtureL. acidophilus + B. longum + L. delbrueckiisubsp. bulgaricus + S. thermophilusMicrobiological evaluation of jejunal aspiratesand faecal samples
Reduced anaerobe:aerobe ratio in faeces by 3-fold during treatmentDistal intestinal micro¯ora a�ected
Colonization of intestinal tract Nielsen et al. [126]
Bi®dobacterium longum Duration of experiment of 3 wk, 12 healthyvolunteersYoghurt (500 ml/d) added with B. longum(109 bacteria/l) and lactulose (5 g/l) orconventional yoghurt
Increased excretion of bi®dobacteria after consumptionBreath-hydrogen exhalation elevated greatStability/balance of human faecal ¯ora
Colonization of intestinal tract,possible role in colon carcinogenesis
Bartram et al. [127]
Lactobacillus acidophilus LA1Bi®dobacterium bi®dum Bb12
Duration of experiment of 3 wk, two groupsof healthy volunteersEach consumed fermented milk productcontaining one of the two strains, bloodsamples studied
Faecal colonization persists for 6 wk after ingestionIncreases in phagocytosis of E. coli coincideNonspeci®c, antinfective mechanisms of defense enhanced
Immune enhancer Schi�rin et al. [128]
Bi®dobacterium longum (addedwith lactulose)
61 male Fisher 344 weaning rats divided into4 groups of 15 rats, each group given oneof 4 diets for 13 wk; subcutaneous injectionof azoxymethane at 7±8 wk of age; aberrantcrypt foci (ACF) in colon and glutathioneS-transferase (GST) in colonic mucosa andliver were analyzed
Feeding of lactulose and B. longum solely and in combinationreduced the number of ACF and total number of aberrant cryptsColonic mucosal GST levels are signi®cantlyHigher positive correlation between higher cecal pH and numberof ACF
Additive antitumor e�ect Anjana et al. [69]
A.M.P.
Gomes
andF.X
.Malcata
/Tren
dsin
FoodScien
ce&Tech
nology
10(1999)
139±157149

indicated, following an in vitro study of 14 bi®dobacteriastrains, that four, used commercially for incorporationin dairy foods (i.e. Bi®dobacterium Bf-1, Bf-6, Bf-12 andBf-13) showed a great capacity for cytokine stimulationby macrophages, and could thus potentially modulatethe immune response either directly, or indirectly viaincorporation as food additives.
Potential hypocholesterolemic actionsSeveral studies indicated a statistically signi®cant
reduction in serum cholesterol during consumption oflarge doses (680±5000 mL/d) of certain fermented dairybioproducts [57] and the theory postulated for thehypocholesterolemic e�ect was the presence of suchorganic acids as uric, orotic and hydroxymethylglutaric,which actually inhibit cholesterol synthesis [77]. Unfor-tunately, these data have not permitted extrapolation tomore realistic consumption conditions (the doses usedare indeed excessively high), and were impoverished bythe lack of controls and the fact that in some cases thestrains used were ill-de®ned. Although a few experi-mental animal studies (Table 5) were successful in posi-tively attributing cholesterol-lowering properties to L.acidophilus contained in dairy products administered asacidophilus yoghurt to pigs [78] and mice [79], resultscannot be extrapolated to humans since there are dif-ferences in the regulation of cholesterol metabolismbetween animals and humans. Besides the hypocholes-terolemic action, Akalin et al. [79] also reported a highernumber of faecal lactobacilli and fewer coliforms inmice that received acidophilus yoghurt than in thosethat received plain yoghurt, thus indicating that L.acidophilus established itself more e�ectively in themurine intestinal tract than did L. bulgaricus in theyoghurt diet.The mechanisms by which fermented milk products
are able to diminish serum cholesterol have not to datebeen fully demonstrated. In vitro studies by Gilliland[56] showed that bi®dobacteria and L. acidophilus areable to utilize cholesterol in growth media, both byassimilation and precipitation with deconjugated bilesalts under acidic conditions. Their suggestion that thepresence of bile salts is a prerequisite for cholesterolassimilation by bi®dobacteria was con®rmed by theobservations of Klaver and van der Meer [80] (Table 5),and more recently by those of Tahri et al. [81,82], whowent one step further and investigated the nature of thebile salts; according to these authors [82], assimilationof cholesterol was higher in the presence of trihydrox-yconjugated bile salts than dihydroxyconjugated bilesalts. The aforementioned bacterial assimilation of cho-lesterol in the intestine may reduce its absorption fromthe digestive tract into the blood system.Despite the existing and ever increasing body of
knowledge, considerable research is still warranted to fullyelucidate causal relationships between the consumption
of probiotic bacteria and reduced serum cholesterollevels in humans.
Dairy products supplemented with Bi®dobacteriumspp. and/or Lactobacillus acidophilusGeneral characteristicsThe species most frequently employed in the produc-
tion of probiotic dairy products are of human intestinalorigin because it is generally accepted that they are bet-ter suited to the physiological needs of the host and canmore easily colonize his/her intestine than wild strains,or strains that exist in the colon of other animals. Thesehuman-borne strains include Bi®dobacterium ado-lescentis, B. bi®dum, B. breve, B. infantis, B. longum,Lactobacillus acidophilus, L. casei subsp. rhamnosus andEnterococcus faecium [22,83]. Results of recent studiessupport the use of selected strains of B. longum as dietaryadjuncts in dairy products, with B. adolescentis and B.infantis as adequate alternatives [84]. It is again worthnoting that each strain within these species exhibits uniqueproperties, viz. growth rate, metabolic rate, proteolyticactivity and ¯avour promotion. Consequently, carefulmanagement of such factors via tailored manufacturingtechnologies will enable these species to meet withvarying degrees of success in multiple industrial appli-cations. Cases of milk products fermented by bi®do-bacteria of animal origin have also been reported: thesestrains are much easier to cultivate and can withstandthe adverse conditions encountered during industrialproduction, viz. low pH and presence of detectable levelsof oxygen. In addition to a bene®cial role after consump-tion [85,86], the recently identi®ed B. lactis strain is also apromising candidate due to its good acid and oxygen tol-erance [23,87].The most frequent functional food products in the
market are of dairy origin (Table 6); Japan, a leadingcountry in their manufacture, produces and marketsmore than 50 di�erent dairy products containing viablecells. Similar trends are also observed in such developedEuropean countries as France, Germany and Sweden(where probiotic products account for �25% of all fer-mented milk products). It is estimated that there are�80 bi®do-containing products available in the worldmarket, and more than 45 dairy plants in Europe cur-rently produce bi®do-acidophilus products [83]. Besidesthe commercial products, research studies have beenmore and more focused on fermented milks [12,88],yoghurt [1], frozen yoghurt and desserts [89,90], icecream [91,92], cheese [93±95,106], fermented soya milk[96] and soya yoghurt [97]. The designation of new pro-ducts is normally obtained via adding the pre®x `Bio' toits traditional (sometimes partially simpli®ed) name (e.g.Biogurt, Biodrink, Biokys). The probiotic strains ofbi®dobacteria and L. acidophilus may be added, eitheralone or following combination with other lactic acidbacteria during fermentation, to the ®nal fermented
150 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

product, or to the fresh product before distribution.General accounts of the physicochemical and technolo-gical aspects of commercial fermented milk productscontaining Bi®dobacterium spp. and L. acidophilus havebeen provided by Kurmann and Rasic [17], Mital andGarg [12] and Tamine et al. [60].In addition to food products containing probiotic
bacteria, there are various health products as well aspharmaceutical preparations containing probiotics,available in the market. In general, these consist inencapsulated freeze-dried bacterial populations that areused in the treatment of gastrointestinal disturbances(diarrhoea, including side e�ects of antibiotic therapy),constipation and certain hepatic diseases.
Technological characteristicsProduction of high-quality fermented milk products
containing Bi®dobacterium spp. and Lactobacillus acid-ophilus is a major challenge to dairy plants owing to thesensitive character of the microorganisms in these bio-products, which adds to the usual di�culties encoun-tered in novel food products (i.e. poor palatability andconsequent limited consumer acceptability). In parti-cular, bi®dobacteria tend to exhibit weak growth andacid production in milk, which invariably requires longfermentation times and conditions of anaerobiosis, low
redox potential at least in the early phase of growth[24,98] and, often, addition of growth-promoting fac-tors to the milk [19,23,32,53,54], as previously discussed.In addition, bi®dobacteria produce, during fermenta-tion, acetic and lactic acids at the ratio 3:2, so excessivegrowth may yield products with vinegar-like taste andaroma, which are obviously not acceptable to con-sumers [83]. Careful selection of the strains employedand good monitoring throughout the manufacturingprocess are therefore compulsory in attempts to controle�ciently the metabolic products and hence the ®nalpH. The use of combined cultures of bi®dobacteria andL. acidophilus or other lactic acid bacteria, viz. Lacto-bacillus delbrueckii subsp. bulgaricus and Streptococcusthermophilus, S. thermophilus alone or mesophilic aro-matic cultures, has also been advocated as a solution formany such problems [23,83,94,99]; increased growthrates and reduction of fermentation time, absence ofcertain sensory and texture defects and furtherimprovement of nutritional value of `bi®dus' productsare advantages brought about by the latter possibility.Adverse e�ects with respect to viability have, however,been reported for some strains of Bi®dobacterium [99]and L. acidophilus [98].A large number of studies has been published per-
taining to characteristics of good probiotics [5,83] and
Table 6. Commercial products containing Bi®dobacterium spp. and Lactobacillus acidophilusa
Product Country of origin Microorganisms
A-38 Denmark Lactobacillus acidophilus, Bi®dobacterium bi®dum, Leuconostoc mesenteroidesspp. cremoris, mesophilic lactococci
Acidophilus buttermilk USA Lactobacillus acidophilus, Leuconostoc mesenteroides spp. cremoris, mesophiliclactococci
Progurt Bi®dobacterium bi®dum, Lactobacillus acidophilus, mesophilic lactococciAcidophilus milk Several countries Lactobacillus acidophilusAcidophilus yeast milk Former USSR Lactobacillus acidophilus, Saccharomyces fragilis, S. cerevisiaeA-B Yoghurt France Bi®dobacterium bi®dum, Lactobacillus acidophilusCultura Denmark IbidemMilky Italy IbidemNu-Trish A/B Milk USA IbidemBiomild Several countries Bi®dobacterium spp., Lactobacillus acidophilusAcidophilus yoghurt(ACO-yoghurt)
Several countries Lactobacillus acidophilus, L. delbrueckii subsp. bulgaricus, Streptococcusthermophilus
B-Active France Lactobacillus acidophilus, Bi®dobacterium bi®dum, L. delbrueckii subsp. bulgaricus,Streptococcus thermophilus
Fresh BA UK IbidemKyr Italy IbidemYoplus Australia IbidemBiogarde Germany Lactobacillus acidophilus, Bi®dobacterium bi®dum, Streptococcus thermophilusO®lus France IbidemPhilus Norway IbidemBi®dus milk Several countries Bi®dobacterium bi®dum, B. longumBi®ghurt Germany Bi®dobacterium bi®dum, Streptococcus thermophilusBiogurt Germany Lactobacillus acidophilus, Streptococcus thermophilusBiokys Czech Republic Bi®dobacterium bi®dum, Lactobacillus acidophilus, Pediococcus acidilacticiMil-Mil Japan Lactobacillus acidophilus, Bi®dobacterium bi®dum, B. breveAkult Japan Lactobacillus acidophilus, Bi®dobacterium bi®dum, B. breve, L. casei subsp. casei
a Adapted from Kurmann [22] and Hoier [83]
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 151

criteria for selection thereof [17,20,84,100]; these aresummarized in Fig. 2. The most important propertiesinclude: (i) acid and bile tolerances, which are essentialto maintain high viable cell numbers during storage andduring passage through the digestive tract followingoral uptake [24,101±103]; (ii) adherence to the humanintestinal mucosa, which is needed for temporary colo-nization of the human gastrointestinal tract; and (iii)production of antimicrobial substances, with con-comitant inhibition of pathogens [20]. Furthermore, thestrains selected should produce a ®nal product posses-sing good taste and acceptable body and texture, aselection step that cannot be achieved unless the pro-duct is actually manufactured.It is also important that the strains selected can be
produced in large scale; in fact, owing to a slow propa-gation in milk, they will hardly be competitive in thepresence of other microorganisms and will, thus, beeasily outnumbered. Therefore, aseptic working condi-tions and special growth-promoting factors (as dis-cussed above) are pre-requisites to ensure high initialviable cell counts thereof. Voelki and Spillmann [104]optimized the process of preparation of starters forposterior manufacture of bi®dus milk and concluded thatactive starters can be prepared with two speci®c strains,Bi®dobacterium breve ATCC 15700 and B. longumATCC15707, under non-strictly anaerobic conditions, by usingmilk initially supplemented with readily available N-com-pounds (e.g. lactalbumin hydrolysate). A wide range ofstudies have made it clear that the production of startersof bi®dobacteria and L. acidophilus, in individual andhighly concentrated freeze-dried or deep frozen forms,
for direct inoculation of processed milk, o�ers great¯exibility in control of the desired sensory and micro-biological qualities (in terms of ratio of bi®dobacteria tolactobacilli) [83,105]. More recently, a ratio of 1:1 offrozen concentrates of B. lactis to L. acidophilus hasbeen considered adequate for optimum growth of, andacidi®cation by B. lactis when in coculture in plain milk,and the concomitant enhanced growth of L. acidophilushas also suggested a certain degree of symbiosis betweenthese strains [23]. Furthermore, it is important thatstrains selected for Direct-Vat-Set (DVS) starters canundergo concentration of up to 1010±1011 cfu/g in orderto permit the desired performance in commercial man-ufacture of fermented milk products. Apart from thisrequirement, the starter produced should also displayacceptable stability (usually guaranteed for 3±12 mo)throughout processing, and subsequent storage anddistribution. Direct inoculation of a dairy product witha frozen concentrated culture (ca. 5±10�1010 cfu/g) isrecommended at a rate of 0.05±0.1 mg per L of milk[23,83,94,106]Production of functional fermented milk products
also requires safety assessment of the probiotic strains; atentative strain must obviously be non-pathogenic fortechnological uses. There is a large consensus that lacticacid bacteria are benign microorganisms, with a well-established record of safety and no record of food poi-soning outbreaks. Several studies document the safetyof dairy strains [107], and an updated review on safetyof probiotic bacteria has been made available byDonohue and Salminen [108]. Doses of probiotic strainsas high as 1012 cfu/g have failed to exhibit toxicity, and
Fig. 2. Criteria used in selection of Bi®dobacterium spp. for application as dietary adjuncts.
152 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

among the di�erent species of the genus Bi®dobacteriumonly B. dentium was considered to actually be patho-genic to man in view of its role in dental caries [7].
Growth and survival characteristicsStrain viability in the carrier food by the time of con-
sumption is closely related to the aforementioned tech-nological requirements. Strain survival depends on pH(and its bu�ering capacity), presence of competingmicroorganisms, storage temperature and presence ofmicrobial inhibitors (e.g. NaCl and H2O2) in the foodmatrix [17]. Studies examining the viability of B. lactisand L. acidophilus, particularly in milk containing arange of experimental NaCl concentrations between 0and 6%(w/w) and stored at di�erent temperatureswithin the range of 5±15�C, indicated that pure culturesof L. acidophilus were more susceptible to higher NaClconcentrations than their B. lactis counterpart [32]. Inaddition, the mechanistic model made available by theseauthors considers the behaviour of the pure and mixedmicrobial populations to be described by speci®c deathrates that vary with temperature (following Arrheniusrelationships) and NaCl levels in the milk medium (fol-lowing simple inhibition kinetics), and is therefore use-ful in e�orts to rationalize the dynamics of cellpopulation survival under a variety of combinations ofeasily manipulated environmental conditions.Moreover, it is essential that fermented bioproducts
contain a satisfactory number of active cells at themoment of consumption, i.e. at least 106 cfu/mL,because the minimum therapeutic daily dose is usuallyconsidered as 108±109 viable cells, realizable through anintake of, say 100 g of fermented bioproduct containing106±107 viable cells/mL [2]. However, it is necessary thatsuch fermented milks are consumed regularly in orderto maintain the e�ect of these special microorganismson the composition of the intestinal micro¯ora. Cases ofmarked loss of viability in dairy products have beenreported for both Bi®dobacterium spp. and L. acid-ophilus, more often during refrigerated storage at lowpH [24,89,109], and re¯ect, as stressed previously, theneed for careful strain selection. These studies urgeprocessors to more deeply understand the implications ofadding acid-sensitive bacteria to highly acidic productslike yoghurt. In contrast, bi®dobacteria survive well inlow-acid products such as frozen yoghurt [89], ice-cream[92] or cheese when added exclusively as dietaryadjuncts [93]. New methods for enhancing the long-termsurvival of probiotic strains, and hence ensure that rea-sonable numbers of bacteria are delivered to the host,have been investigated either by replacing the carrierfood [92±94,106] or by improving the protection of acid-sensitive strains via microencapsulation with celluloseacetate phthalate [110], �-carrageenan [93] or Ca-alginate[111]; the ®rst method appears to be technologically andcommercially the most feasible. For example, a starter
composed solely of a mixture of L. acidophilus and B.lactis has been successfully used in the manufacture ofcheeses following modi®ed technologies of the well-knownGouda cheese [94] and of Cabra cheese, a cheese with alarge social and economic importance in the Mediterra-nean basin [106]. Growth behaviours of either strainwere dependent on the physicochemical characteristicsprevailing in both cheese-types, yet acid production wasnot a�ected. Survival of the probiotic strains was mon-itored during ripening, and the ®nal viable numberswere still above the accepted threshold. Both strainscontributed signi®cantly to ripening of both cheese-types, especially in terms of formation of low molecularmass peptides and free amino acids, but lipolysis wasnot greatly a�ected. The best compromise between sen-sorial, physicochemical and probiotic attributes of the®nal cheese was obtained for 3�107 and 7�106 cfu/mLof B. lactis and L. acidophilus, respectively, 3.5%(w/w)salt, addition of 0.3%(v/w) milk hydrolysate and ripen-ing for 70 d.The selection of oxygen-tolerant species may also be a
solution toward minimization of the adverse e�ect ofoxygen in fermented bioproducts containing Bi®do-bacterium spp. and Lactobacillus acidophilus. Themechanisms of susceptiblity of Bi®dobacterium to mole-cular oxygen have interested researchers at early stages[112], who estimated the oxygen sensitivity of twelvestrains by measuring the extent of growth inhibitionwhen they were grown in deep agar cultures and theextent of growth in aerated cultures; three categories ofBi®dobacterium species with regard to their oxygen sen-sitivity were accordingly suggested. These researcherswere not, however, able to establish a correlationbetween oxygen sensitivity and enzyme activities relat-ing to oxygen metabolism, and it was only in the presentdecade that studies provided a clearer insight into amore fundamental relationship. The initial study byShimamura et al. [113] has elucidated oxygen uptake byseveral bi®dobacterial species when carbohydrates aremetabolized, and reported that this mechanism is e�ec-tive in diminishing the impact of environmental oxygeneven when faced with an insu�cient energy source, e.g.glucose; involvement of NADH oxidase for oxygenuptake activity in Bi®dobacterium was thus demon-strated. In their follow-up study, Shimamura et al. [114]reported that all four Bi®dobacterium spp. strainsassayed expressed both NADH oxidase and NADHperoxidase activites, but to di�erent degrees; a stronginverse correlation between enzyme activites and degreeof oxygen sensitivity was consequently established. Bi®-dobacterium infantis, B. breve and B. longum were muchmore tolerant to oxygen (resulting from higher activitiesof the aforementioned enzymes) than B. adolescentiswas (arising from very low activities thereof); therefore,the former are better suited for industrial applications.These results permitted re®nement of their initial
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 153

hypothesis [113] in that the pathway involving the twoNADH oxidative enzymes apparently operates as adefense mechanism to reduce oxygen toxicity in Bi®do-bacterium spp.Finally, a probiotic strain must show good surviva-
bility, both in the product and after digestion. It is clearfrom the above lines that certain strains are able to sur-vive well when exposed to gastric acids, or certainsecretions of the small intestine.
References
1 Reuter, G. (1990) `Bi®dobacteria Cultures as Components ofYoghurt-like Products' in Bi®dobacteria Micro¯ora 9, 107±118
2 Rasic, J.L. and Kurmann, J.A. (1983) `Bi®dobacteria and TheirRole', BirkhauÈ ser, Basel, Switzerland
3 Lilly, D.M. and Stillwell, R.H. (1965) `Probiotics: Growth PromotingFactors Produced by Microorganisms' in Science 147, 747±748
4 Parker, R.B. (1974) `Probiotics, The Other Half of the Anti-biotics Story' in Animal Nutrition and Health 29, 4±8
5 Fuller, R. (1989) `A Review: Probiotics in Man and Animals' inJournal of Applied Bacteriology 66, 365±378
6 Havenaar, R. and Huis in't Veld, J.H.J. (1992) `Probiotics; AGeneral Review' in The Lactic Acid Bacteria in Health and Dis-ease, (Wood, B., ed), pp. 151±170, Elsevier, Barking
7 Sgorbati, B., Biavati, B. and Palenzona, D. (1995) `The GenusBi®dobacterium' in The Lactic Acid Bacteria Vol. 2. The Generaof Lactic Acid Bacteria, (chapter 8), (Wood, B.J.B, Holzapfel,W.H., eds), pp. 279±306, Blackie Academic, London
8 Krzewinski, F., Brassart, C., Gavini, F. and Bouquelet, S. (1996)`Characterization of the Lactose Transport System in theStrain Bi®dobacterium bi®dum DSM 20082' in Current Micro-biology 32, 301±307
9 Crociani, F., Alessandrini, A., Mucci, M.M. and Biavati, B.(1994) `Degradation of Complex Carbohydrates by Bi®do-bacterium spp' in International Journal of Food Microbiology24, 199±210
10 Meile, L., Ludwig, W., Rueger, U., Gut, C., Kaufmann, P., Dasen,G., Wenger, S. and Teuber, M. (1997) `Bi®dobacterium lactis sp.nov., a Moderately Oxygen Tolerant Species Isolated from Fer-mented Milk' in Systematic and Applied Microbiology 20, 57±64
11 Hammes, W.P. and Vogel, R.F. (1995) `The Genus Lactobacillus'in The Lactic Acid Bacteria Vol. 2. The Genera of Lactic AcidBacteria, (Wood, B.J.B., Holzapfel, W.H., eds), pp. 19±54,Blackie Academic, London
12 Mital, B.K. and Garg, S.K. (1992) `Acidophilus Milk Products:Manufacture and Therapeutics' in Food Reviews International8, 347±389
13 Nahaisi, M.H. (1986) `Lactobacillus acidophilus: TherapeuticProperties, Products and Enumeration. (Chapter 6)' in Devel-opments in Food Microbiology, (Robinson, R.K., ed), pp. 153±178, Elsevier Applied Science Publishers, London, UK
14 Nielsen, J.W. and Gilliland, S.E. (1992) `The Lactose Hydrolyz-ing Enzyme from Lactobacillus acidophilus' in Cultured DairyProducts Journal 27, 20±24
15 Marshall, V.M. and Cole, W.M. (1983) `Threonine Aldolase andAlcohol Dehydrogenase Activities in Lactobacillus bulgaricusand Lactobacillus acidophilus and their Contribution to Fla-vour Production in Fermented Milks' in Journal of DairyResearch 50, 375±379
16 Finegold, S.M., Sutter, V.L. and Mathisen, G.E. (1983) `NormalIndigenous Intestinal Flora' in Human Intestinal Micro¯ora inHealth and Disease, (Hentges, D.J., ed), pp. 3±31, AcademicPress, New York, NY
17 Kurmann, J.A. and Rasic, J.L. (1991) `The Health Potential of
Products Containing Bi®dobacteria' in Therapeutic Propertiesof Fermented Milks, (Robinson, R.K., ed), pp. 117±157, ElsevierApplied Science Publishers, London, UK
18 Mitsuoka, T. (1990) `Bi®dobacteria and their Role in HumanHealth' in Journal of Industrial Microbiology 6, 263±268
19 Modler, H.W. (1994) `Bi®dogenic FactorsÐSources, Metabo-lism and Applications' in International Dairy Journal 4, 383±407
20 Salminen, S., Isolauri, E. and Salminen, E. (1996) `Clinical Usesof Probiotics for Stabilizing the Gut Mucosal Barrier: Success-ful Strains and Future Challenges' in Antonie van Leeu-wenhoek 70, 347±358
21 Hassinen, J.B., Durbin, G.T., Tomarelli, R.M. and Bernhart, F.W.(1951) `The Minimal Nutritional Requirements of Lactobacillusbi®dus' in Journal of Bacteriology 62, 771±777
22 Kurmann, J.A. (1998) `Starters for Fermented Milks: Starterswith Selected Intestinal Bacteria' in Bulletin of the InternationalDairy Federation 227, 41±55
23 Gomes, A.M.P., Malcata, F.X. and Klaver, F.A.M. (1998)`Growth Enhancement of Bi®dobacterium lactis Bo and Lacto-bacillus acidophilus Ki by Milk Hydrolyzates' in Journal ofDairy Science 81, 2817±2825
24 Klaver, F.A.M., Kingma, F. and Weerkamp, A.H. (1993) `Growthand Survival of Bi®dobacteria in Milk' in Netherlands Milk andDairy Journal 47, 151±164
25 Marshall, V.M., Cole, W.M. and Mabbitt, L.A. (1982) `Fermen-tation of Specially Formulated Milk with Single Strains of Bi®-dobacteria' in Journal of the Society of Dairy Technology 35,143±144
26 Rogosa, M. (1974) `Lactobacillus: Bergey's Manual of Determi-native Bacteriology', Williams and Wilkins, Baltimore, MA, USA
27 Desmazeaud, M. (1983) `L'e tat des Connaissances en MatieÁ rede Nutrition des Bacte ries Lactiques' in Le Lait 63, 267±316
28 Gyllenberg, H., Rossander, M. and Roine, P. (1956) `E�ect ofthe Fatty Acids of Human Milk and Cow's Milk on IntestinalLactobacilli' in Acta Paediatrica Stockholm 45, 147±160
29 Ledesma, O.V., de Ruiz Holgado, A.P., Olivier, G., de Giori,G.S., Raibaud, P. and Galpin, J.V. (1977) `A Synthetic Mediumfor Comparative Nutritional Studies of Lactobacilli' in Journalof Applied Bacteriology 42, 123±133
30 Babu, V., Mital, B.K. and Garg, S.K. (1992) `E�ect of Tomato JuiceAddition on the Growth and Activity of Lactobacillus acid-ophilus' in International Journal of Food Microbiology 17, 67±70
31 Taillandier, P., Gilis, F., Portugal, F.R., Laforce, P. and Stre-haiano, P. (1996) `In¯uence of Medium Composition, pH andTemperature on the Growth and Viability of Lactobacillusacidophilus' in Biotechnology Letters 18, 775±780
32 Gomes, A.M.P. and Malcata, F.X. (1998) `Use of Small Rumi-nants' Milks Supplemented with Available Nitrogen asGrowth Media for Bi®dobacterium lactis and Lactobacillusacidophilus' in Journal of Applied Microbiology 85, 839±848
33 Roy, D., Dussault, F. and Ward, P. (1990) `Growth Require-ments of Bi®dobacterium Strains in Milk' in Milchwissenschaft45, 500±502
34 Ventling, B.L. and Mistry, V.V. (1993) `Growth Characteristicsof Bi®dobacteria in Ultra®ltered Milk' in Journal of Dairy Sci-ence 76, 962±971
35 Gibson, G.R. and Roberfroid, M.B. (1995) `Dietary Modulationof the Human Colonic Microbiota: Introducing the Conceptof Prebiotics' in Journal of Nutrition 125, 1401±1412
36 Mitsuoka, T., Hidaka, H. and Eida, T. (1987) `E�ect of Fructo-oligosaccharides on Intestinal Micro¯ora' in Die Nahrung 31,427±436
37 Wang, X. and Gibson, G.R. (1993) `E�ects of the In Vitro Fer-mentation of Oligofructose and Inulin by Bacteria Growing inthe Human Large Intestine' in Journal of Applied Bacteriology75, 373±380
154 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

38 Gibson, G.R. and Wang, X. (1994) `Bi®dogenic Properties ofDi�erent types of Fructo-oligosaccharides' in Food Micro-biology 11, 491±498
39 Gibson, G.R. and Wang, X. (1994) `Enrichment of Bi®dobacteriafrom Human Gut Contents by Oligofructose using ContinuousCulture' in FEMS Microbiology Letters 118, 121±128
40 Roberfroid, M., Gibson, G.R. and Delzenne, N. (1993) `TheBiochemistry of Oligofructose, a Non-digestible Fiber: AnApproach to Calculate its Caloric Value' in Nutrition Reviews51, 137±146
41 Tomomatsu, H. (1994) `Health E�ects of Oligosaccharides' inFood Technology 48, 61±65
42 Mizota, T. (1996) `Functional and Nutritional Foods ContainingBi®dogenic Factors' in Bulletin of the International Dairy Fed-eration 313, 31±35
43 Go�, H.D. and Modler, H.W. (1996) `Bi®dobacteria, Fructooli-gosaccharides and Ice Cream' in Canadian Dairy 75, 10
44 Berger, J.L., Lee, B.H. and Lacroix, C. (1995) `OligosaccharidesSynthesis by Free and Immobilized �-Galactosidases fromThermus aquaticus YT-1' in Biotechnology Letters 17, 1077±1080
45 Yanahira, S., Kobayashi, T., Suguri, T., Nakakoshi, M., Miura,S., Ishikawa, H. and Nakajima, I. (1995) `Formation of Oligo-saccharides from Lactose by Bacillus circulans �-Galactosi-dase' in Bioscience, Biotechnology and Biochemistry 59, 1021±1026
46 Onishi, N. and Yokozeki, K. (1996) `Gluco-oligosaccharide andGalacto-oligosaccharide Production by Rhodotorula minutaIFO879' in Journal of Fermentation and Bioengineering 82,124±127
47 Mizota, T. (1996) `Lactulose as a Growth Promoting Factor forBi®dobacterium and its Physiological Aspects' in Bulletin of theInternational Dairy Federation 313, 43±48
48 Roland, N., Bougle, D., Lebeurrier, F., Arhan, P. and Maubois,J.-L. 1997. Propionibacterium freudenreichii Stimulate theGrowth of Bi®dobacterium bi®dum In Vitro and Increase FecalBi®dobacteria in Healthy Human Volunteers. In Proceedingsof Lactic 97 Symposium, Adria Normandie, pp. 103±104
49 Mori, H., Sato, Y., Taketomo, N., Kamiyama, T., Yoshiyama, Y.,Meguro, S., Sato, H. and Kaneko, T. (1997) `Isolation andStructural Identi®cation of Bi®dogenic Growth StimulatorProduced by Propionibacterium freudenreichii' in Journal ofDairy Science 80, 1959±1964
50 Poch, M. and Bezkorovainy, A. (1991) `Bovine Milk �-CaseinTrypsin Digest is a Growth Enhancer for the Genus Bi®do-bacterium' in Journal of Agriculture and Food Chemistry 39,73±77
51 Ibrahim, S.A. and Bezkorovainy, A. (1994) `Growth-promotingFactors for Bi®dobacterium longum' in Journal of Food Science59, 189±191
52 Petschow, B.W. and Talbott, R.D. (1990) `Growth Promotion ofBi®dobacterium Species by Whey and Casein Fractions fromHuman and Bovine Milk' in Journal of Clinical Microbiology28, 287±292
53 Proulx, M., Gauthier, S.F. and Roy, D. (1992) `Utilisation d'Hy-drolysats Enzymatiques de Case ine pour la Croissance desBi®dobacte ries' in Le Lait 72, 393±404
54 Proulx, M., Ward, P., Gauthier, S.F. and Roy, D. (1994) `Com-parison of Bi®dobacterial Growth-promoting Activity of Ultra-®ltered Casein Hydrolyzate Fractions' in Le Lait 74, 139±152
55 Gurr, M.I. (1987) `Nutritional Aspects of Fermented Milk Pro-ducts' in FEMS Microbiology Reviews 46, 337±342
56 Gilliland, S.E. (1990) `Health and Nutritional Bene®ts from LacticAcid Bacteria' in FEMS Microbiology Reviews 87, 175±188
57 Marteau, P. and Rambaud, J.C. (1993) `Potential of Using Lac-tic Acid Bacteria for Therapy and Immunomodulation in Man'in FEMS Microbiology Reviews 12, 207±220
58 Yaeshima, T. (1996) `Bene®ts of Bi®dobacteria to HumanHealth' in Bulletin of the International Dairy Federation 313,36±42
59 Saxelin, M., Pessi, T. and Salminen, S. (1995) `Fecal RecoveryFollowing Oral Administration of Lactobacillus Strain GG(ATCC 53103) in Gelatine Capsules to Healthy Volunteers' inInternational Journal of Food Microbiology 25, 199±203
60 Tamine, A.Y., Marshall, V.M. and Robinson, R.K. (1995) `Micro-biological and Technological Aspects of Milks Fermented byBi®dobacteria' in Journal of Dairy Science 62, 151±187
61 Bernet, M.F., Brassart, D., Neeser, J.R. and Servin, A. (1993)`Adhesion of Human Bi®dobacterial Strains to CulturedHuman Intestinal Epithelial Cells and Inhibition of Entero-pathogen-cell Interactions' in Applied and EnvironmentalMicrobiology 59, 4121±4128
62 Kot, E., Furmanov, S. and Bezkorovainy, A. (1995) `Ferrous IronOxidation by Lactobacillus acidophilus and its Metabolic Products'in Journal of Agricultural and Food Chemistry 43, 1276±1282
63 Gibson, G.R. and Wang, X. (1994) `Regulatory E�ects of Bi®-dobacteria on the Growth of Other Colonic Bacteria' inJournal of Applied Bacteriology 77, 412±420
64 Tahara, T., Oshimura, M. and Kanatani, K. (1996) `Mode ofAction of Acidocin 8912, A Bacteriocin Produced by Lacto-bacillus acidophilus TK8912' in Letters in Applied Microbiology23, 192±194
65 Fujiwara, S., Hashiba, H., Hirota, T. and Forstner, J.F. (1997)`Proteinaceous Factor(s) in Culture Supernatant Fluids of Bi®-dobacteria which Prevents the Binding of EnterotoxigenicEscherichia coli to Gangliotetraosylceramide' in Applied andEnvironmental Microbiology 63, 506±512
66 Kot, E., Haloftis, G. and Bezkorovainy, A. (1994) `Iron Accu-mulation by Bi®dobacteria at Low pO2 and in Air: Action ofPutative Ferroxidase' in Journal of Agricultural and FoodChemistry 42, 685±688
67 Hosoda, M., Hashimoto, H., He, F., Morita, H. and Hosono, A.(1996) `E�ect of Administration of Milk Fermented with Lacto-bacillus acidophilus LA-2 on Fecal Mutagenicity and Micro-¯ora in the Human Intestine' in Journal of Dairy Science 79,745±749
68 Morotomi, M. (1996) `Properties of Lactobacillus casei ShirotaStrain as Probiotics' in Asia Paci®c Journal of Clinical Nutrition5, 29±30
69 Anjana, C., Ramkishan-Rao, D., Chawan, C.B. and Shack-elford, L. (1997) `Bi®dobacterium longum and Lactulose Sup-press Azoxymethane-induced Colonic Aberrant Crypt Foci inRats' in Carcinogenesis 18, 517±521
70 Hatcher, G.E. and Lambrecht, R.S. (1993) `Augmentation ofMacrophage Phagocytic Activity by Cell-free Extracts ofSelected Lactic Acid-producing Bacteria' in Journal of DairyScience 76, 2485±2493
71 Fernandes, C.F. and Shahani, K.M. (1990) `Anticarcinogenicand Immunological Properties of Dietary Lactobacilli' inJournal of Food Protection 53, 704±710
72 De Simone, C., Ferrazzi, M., Di Seri, M., Momgio, F., Baldinelli,L. and Di Fabio, S. (1987) `The Immunoregulation of theIntestinal Flora Bi®dobacteria and Lactobacilli Modulate theProduction of -IFN Induced by Pathogenic Bacteria' inInternational Journal of Immunotherapy 3, 151±158
73 Lee, J., Ametani, A., Enomoto, A., Sato, Y., Motoshima, H., Ike,F. and Kaminogawa, S. (1993) `Screening for the Immunopo-tentiating Activity of Food Microorganisms and Enhancementof the Immune Response by Bi®dobacterium adolescentisM101-4' in Bioscience Biotechnology and Biochemistry 57, 2127
74 Kado-Oka, Y., Fujiwara, S. and Hirota, T. (1991) `E�ects ofBi®dobacteria Cells on Mitogenic Response of Splenocytesand Several Functions of Phagocytes' in Milchwissenschaft 46,626±628, 630
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 155

75 Hosono, A., Lee, J., Ametani, A., Natsume, M., Hirayama, M.,Adachi, T. and Kaminogawa, S. (1997) `Characterization of aWater-soluble Polysaccharide Fraction with Immunopo-tentiating Activity from Bi®dobacterium Adolescentis M101-4'in Bioscience, Biotechnology and Biochemistry 61, 312±316
76 Marin, M.L., Lee, J.H., Murtha, J., Ustunol, Z. and Pestka, J.J.(1997) `Di�erential Cytokine Production in Clonal Macro-phage and T-cell Lines Cultured with Bi®dobacteria' in Journalof Dairy Science 80, 2713±2720
77 Fernandes, C.F., Shahani, K.M. and Amer, M.A. (1987) `Ther-apeutic Role of Dietary Lactobacilli and Lactobacilli FermentedDairy Products' in FEMS Microbiology Reviews 46, 343±356
78 Jones, G., Shahani, K.M. and Amer, M.A. (1985) `The E�ect ofAcidophilus Yoghurt on Serum Cholesterol, Triglyceride andLipoprotein Levels of Weanling Pigs' in Journal of Dairy Sci-ence 68, 84±91
79 Akalin, A.S., GoÈ ncË , S. and DuÈ zel, S. (1997) `In¯uence of Yogurtand Acidophilus Yogurt on Serum Cholesterol Levels in Mice'in Journal of Dairy Science 80, 2721±2725
80 Klaver, F. and van der Meer, R. (1993) `The Assumed Assim-ilation of Cholesterol by Lactobacillus and Bi®dobacteriumbi®dum is Due to their Bile Salt-deconjugation Activity' inApplied and Environmental Microbiology 59, 1120±1124
81 Tahri, K., Crociani, J., Ballongue, J. and Schneider, F. (1995)`E�ect of Three Strains of Bi®dobacteria on Cholesterol' inLetters in Applied Microbiology 21, 149±151
82 Tahri, K., Grill, J.P. and Schneider, F. (1997) `Involvement ofTrihydroxyconjugated Bile Salts in Cholesterol Assimilation byBi®dobacteria' in Current Microbiology 34, 79±84
83 Hoier, E. (1992) `Use of Probiotic Starter Cultures in DairyProducts' in Food Australia 44, 418±420
84 Martin, J.H. (1996) `Technical Considerations for IncorporatingBi®dobacteria and Bi®dogenic Factors into Dairy Products' inBulletin of the International Dairy Federation 313, 49±51
85 Weerkamp, A.H., Bovee-Oudenhoven, I. and van der Meer, R.(1996) `E�ect of Probiotics on Infections with Pathogenic Bac-teria in Human Gut Flora-associated Rats.' Poster J36, pre-sented at Fifth Symposium on Lactic Acid Bacteria-Genetics,Metabolism and Applications, Veldhoven, The Netherlands,8±12 September
86 Weerkamp, A.H., Klaver, F.A.M. and van der Meer, R. (1993)``Assimilation'' of Cholesterol by Lactobacilli is due to theirBile Salt Deconjugating Activity' in FEMS MicrobiologyReviews 12, P185
87 Gomes, A.M.P., Teixeira, M.G. and Malcata, F.X. (1998) `Viabi-lity of Bi®dobacterium lactis and Lactobacillus acidophilus inMilk: Sodium Chloride Concentration and Storage Tempera-ture' in Journal of Food Processing and Preservation 22, 221±240
88 Richardson, D. (1996) `Probiotics and Product Innovation' inNutrition and Food Science 4, 27±33
89 Laroia, S. and Martin, J.H. (1991) `E�ect of pH on Survival ofBi®dobacterium bi®dum and Lactobacillus acidophilus in Fro-zen Fermented Dairy Desserts' in Cultured Dairy ProductsJournal 26, 13±21
90 Modler, H.W. and Villa-Garcia, L. (1993) `The Growth of Bi®do-bacterium longum in a Whey-based Medium and Viability ofthis Organism in Frozen Yoghurt with Low and High Levels ofDeveloped Acidity' in Cultured Dairy Products Journal 28, 4±8
91 Modler, H.W., McKellar, R.C., Go�, H.D. and Mackie, D.A.(1990) `Using Ice Cream as a Mechanism to Incorporate Bi®-dobacteria and Fructooligosaccharides into the Human Diet'in Cultured Dairy Products Journal 25, 4±9
92 Hekmat, S. and McMahon, D.J. (1992) `Survival of Lactobacillusacidophilus and Bi®dobacterium bi®dum in Ice Cream for Use asa Probiotic Food' in Journal of Dairy Science 75, 1415±1422
93 Dinakar, P. and Mistry, V.V. (1994) `Growth and Viability of
Bi®dobacterium bi®dum in Cheddar Cheese' in Journal ofDairy Science 77, 2854±2864
94 Gomes, A.M.P., Malcata, F.X., Klaver, F.A.M. and Grande, H.J.(1995) `Incorporation of Bi®dobacterium sp. Strain Bo andLactobacillus acidophilus Strain Ki in a Cheese Product' inNetherlands Milk and Dairy Journal 49, 71±95
95 Blanchette, L., Roy, D., Belanger, G. and Gauthier, S.F. (1996)`Production of Cottage Cheese Using Dressing Fermented byBi®dobacteria' in Journal of Dairy Science 79, 8±15
96 Valdez, G.F. and de Giori, G.S. (1993) `E�ectiveness of Soy Milkas Food Carrier for Lactobacillus acidophilus' in Journal ofFood Protection 56, 320±322
97 Murti, T.W., Bouillanne, C., Landon, M. and Desmazeaud, M.J.(1992) `Bacterial Growth and Volatile Compounds in Yoghurt-type Products from Soymilk Containing Bi®dobacterium spp'in Journal of Food Science 58, 153±157
98 Dave, R.I. and Shah, N.P. (1997) `Viability of Yoghurt andProbiotic Bacteria in Yoghurts Made from Commercial StarterCultures' in International Dairy Journal 7, 31±41
99 Samona, A., Robinson, R.K. and Marakis, S. (1996) `Acid Produc-tion by Bi®dobacteria and Yoghurt Bacteria during Fermenta-tion and Storage of Milk' in Food Microbiology 13, 275±280
100 Johnson, M.C., Ray, B. and Bhowmik, T. (1987) `Selection ofLactobacillus acidophilus Strains for use in ``Acidophilus Pro-ducts''' in Antonie van Leeuwenhoek 53, 215±231
101 Clark, P.A., Cotton, L.N. and Martin, J.H. (1993) `Selection ofBi®dobacteria for use as Dietary Adjuncts in Cultured DairyFoods: IIÐTolerance to Simulated pH of Human Stomachs' inCultured Dairy Products Journal 28, 11±14
102 Ibrahim, S.A. and Bezkorovainy, A. (1993) `Survival of Bi®do-bacteria in the Presence of Bile Salt' in Journal of the Scienceof Food and Agriculture 62, 351±354
103 Gupta, P.K., Mital, B.K. and Garg, S.K. (1996) `Characterizationof Lactobacillus acidophilus Strains for Use as Dietary Adjunct'in International Journal of Food Microbiology 29, 105±109
104 Voelki, D. and Spillmann, H. (1996) `Optimization of the Pre-paration of Starters for Manufacture of Bi®dus Milk UnderPractical Conditions' in DMZ-Lebensmittelindustrie undMilchwirtschaft 117, 56±64
105 Blanchette, L., Roy, D. and Gauthier, S.F. (1995) `ProtectiveE�ect of Ultra®ltered Retentate Powder on Stability of Freeze-dried Cultures of Bi®dobacteria During Storage' in Milchwis-senschaft 50, 363±367
106 Gomes, A.M.P. and Malcata, F.X. (1998) `Development of aProbiotic Cheese Manufactured from Goat Milk: ResponseSurface Analysis via Technological Manipulation' in Journal ofDairy Science 81, 1492±1507
107 Adams, M.R. and Marteau, P. (1995) `On the Safety of LacticAcid Bacteria from Food' in International Journal of FoodMicrobiology 27, 263±264
108 Donohue, D. and Salminen, S. (1996) `Safety of ProbioticBacteria' in Asia Paci®c Journal of Clinical Nutrition 5, 25±28
109 Lankaputhra, W.E.V., Shah, N.P. and Britz, M.L. (1996) `Survivalof Bi®dobacteria During Refrigerated Storage in the Presenceof Acid and Hydrogen Peroxide' in Milchwissenschaft 51, 65±70
110 Rao, A.V., Shiwnarain, N. and Maharaj, I. (1989) `Survival ofMicroencapsulated Bi®dobacterium pseudolongum in Simu-lated Gastric and Intestinal Juices' in Canadian Institute ofFood Science and Technology Journal 22, 345±349
111 Kim, K.I., Baek, Y.J. and Yoon, Y.H. (1996) `E�ects of Rehydra-tion Media and Immobilization in Ca-alginate on the Survivalof Lactobacillus casei and Bi®dobacterium bi®dum' in KoreanJournal of Dairy Science 18, 193±198
112 de Vries, W. and Stouthamer, A.H. (1969) `Factors Determin-ing the Degree of Anaerobiosis of Bi®dobacterium Strains' inArchives Mikrobiologie 65, 275±287
156 A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157

113 Shimamura, S., Abe, F., Ishibashi, N., Miyakawa, H., Yaeshima,T. and Tomita, M. (1990) `Endogenous Oxygen Uptake andPolysaccharide Accumulation in Bi®dobacterium' in Agri-culture and Biological Chemistry 54, 2869±2874
114 Shimamura, S., Abe, F., Ishibashi, N., Miyakawa, H., Yaeshima,T., Araya, T. and Tomita, M. (1992) `Relationship betweenoxygen sensitivity and oxygen metabolism of Bi®dobacteriumspecies' in Journal of Dairy Science 75, 3296±3306
115 Zbikowski, Z. and Ziajka, S. (1986) `Hydrolyzed Casein as aSource of Bi®dus Factor' in Die Nahrung 30, 415±416
116 Azuma, N., Yamauchi, K. and Mitsuoka, T. (1984) `Bi®dusGrowth-promoting Activity of a Glycomacropeptide Derivedfrom Human �-Casein' in Agricultural and Biological Chem-istry 48, 2159±2162
117 Abd-El-Salam, M.H., El-Shibiny, S. and Buchheim, W. (1996)`Characteristics and Potential Uses of the Casein Macropep-tide' in International Dairy Journal 6, 327±341
118 Ito, M., Kimura, M., Deguchi, Y., Miyamori-Watabe, A.,Yajima, T. and Kan, T. (1993) `E�ects of TransgalactosylatedDisaccharides on the Human Intestinal Micro¯ora and theirMetabolism' in Journal of Nutritional Science and Vitaminology39, 279±288
119 Yamazaki, H. and Matsumoto, K. (1994) `Puri®cation of Jer-usalem Artichoke Fructans and their Utilisation by Bi®do-bacteria' in Journal of the Science of Food and Agriculture 64,461±465
120 Okazaki, M., Fujiwaka, S. and Matsumoto, N. (1990) `E�ect ofXylooligosaccharide on the Growth of Bi®dobacteria' in Bi®-dobacteria and Micro¯ora 9, 77±86
121 Yamada, H., Itoh, K., Morishita, Y. and Taniguchi, H. (1993)`Structure and Properties of Oligosaccharides from WheatBran' in Cereal Foods World 38, 490±492
122 Akiyama, H., Endo, T., Nakakita, R., Murata, K., Yonemoto, Y.and Okayama, K. (1992) `E�ect of Depolymerized Alginateson the Growth of Bi®dobacteria' in Bioscience Biotechnologyand Biochemistry 56, 355±356
123 Yang, R.D., Zhang, L.H., Chen, H.Y. and Lin, X.S. (1996)`Development of New E�cient Bi®dus Growth from Farm By-products' in China Dairy Industry 24, 11
124 Gupta, P.K., Mital, B.K. and Garg, S.K. (1996) `Inhibitory Activ-ity of Lactobacillus acidophilus Against Di�erent Pathogens inMilk' in Journal of Food Science and Technology of India 33,147±149
125 Grill, J.P., Crociani, J. and Ballongue, J. (1995) `E�ect of Bi®do-bacteria on Nitrites and Nitrosamines' in Letters in AppliedMicrobiology 20, 328±330
126 Nielsen, O.H., Jorgensen, S., Pedersen, K. and Justesen, T.(1994) `Microbiological Evaluation of Jejunal Aspirates andFaecal Samples after Oral Administration of Bi®dobacteriaand Lactic Acid Bacteria' in Journal of Applied Bacteriology 76,469±474
127 Bartram, H.P., Scheppach, W., Gerlach, S., Ruckdeschel, G.,Kelber, E. and Kasper, H. (1994) `Does Yogurt Enriched withBi®dobacterium longum A�ect Colonic Microbiology andFecal Metabolites in Healthy Subjects' in American Journal ofClinical Nutrition 59, 428±432
128 Schi�rin, E.J., Rochat, F., Link-Amster, H., Aeschlimann, J.M.and Donnet-Hughes, A. (1995) `Immunomodulation ofHuman Blood Cells Following the Ingestion of Lactic AcidBacteria' in Journal of Dairy Science 78, 491±497
129 Idota, T., Kawakami, H. and Nakajima, I. (1994) `Growth-pro-moting E�ects of N-acetylneuraminic Acid-containing Sub-stances on Bi®dobacteria' in Bioscience, Biotechnology andBiochemistry 58, 1720±1722
A.M.P. Gomes and F.X. Malcata / Trends in Food Science & Technology 10 (1999) 139±157 157
Copyright © 2022 FDOKUMEN




















