
BIBIT KRISAN (Dendranthema grandiflorum Kitam.) BEBAS ...
40
i LAPORAN PENELITIAN HIBAH BERSAING PERGURUAN TINGGI XVIII/2 TAHUN ANGGARAN 2011 PENYEDIAAN METODE SEROLOGI UNTUK SERTIFIKASI BIBIT KRISAN (Dendranthema grandiflorum Kitam.) BEBAS CHRYSANTHEMUM B CARLAVIRUS (CVB) Dr. Dr. Dr. Dr. Ir. I GEDE RAI MAY Ir. I GEDE RAI MAY Ir. I GEDE RAI MAY Ir. I GEDE RAI MAYA TEMAJA, MP TEMAJA, MP TEMAJA, MP TEMAJA, MP. Prof. Dr. Ir. MADE SUDANA, MS. Prof. Dr. Ir. MADE SUDANA, MS. Prof. Dr. Ir. MADE SUDANA, MS. Prof. Dr. Ir. MADE SUDANA, MS. Dr. Ir. GEDE SUASTIKA, MSc. Dr. Ir. GEDE SUASTIKA, MSc. Dr. Ir. GEDE SUASTIKA, MSc. Dr. Ir. GEDE SUASTIKA, MSc. Dibiayai dari Dana DIPA PNBP Universitas Udayana Tahun Anggaran 2011 dengan Surat Perjanjian Kontrak Nomor : 1696.A.10/UN14/KU.03.04/PERJANJIAN/2011, Tanggal 11 Mei 2011 FAKUL FAKUL FAKUL FAKULTAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGI TAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGI TAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGI TAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGI UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA DENPASAR DENPASAR DENPASAR DENPASAR
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of BIBIT KRISAN (Dendranthema grandiflorum Kitam.) BEBAS ...

i
LAPORAN PENELITIAN
HIBAH BERSAING PERGURUAN TINGGI XVIII/2
TAHUN ANGGARAN 2011
PENYEDIAAN METODE SEROLOGI UNTUK SERTIFIKASI BIBIT KRISAN (Dendranthema grandiflorum Kitam.) BEBAS CHRYSANTHEMUM B CARLAVIRUS (CVB)
Dr. Dr. Dr. Dr. Ir. I GEDE RAI MAYIr. I GEDE RAI MAYIr. I GEDE RAI MAYIr. I GEDE RAI MAYAAAA TEMAJA, MPTEMAJA, MPTEMAJA, MPTEMAJA, MP.... Prof. Dr. Ir. MADE SUDANA, MS.Prof. Dr. Ir. MADE SUDANA, MS.Prof. Dr. Ir. MADE SUDANA, MS.Prof. Dr. Ir. MADE SUDANA, MS. Dr. Ir. GEDE SUASTIKA, MSc.Dr. Ir. GEDE SUASTIKA, MSc.Dr. Ir. GEDE SUASTIKA, MSc.Dr. Ir. GEDE SUASTIKA, MSc.
Dibiayai dari Dana DIPA PNBP Universitas Udayana Tahun Anggaran 2011 dengan Surat Perjanjian Kontrak Nomor : 1696.A.10/UN14/KU.03.04/PERJANJIAN/2011,
Tanggal 11 Mei 2011
FAKULFAKULFAKULFAKULTAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGITAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGITAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGITAS PERTANIAN/PROGRAM STUDI AGROEKOTEKNOLOGI
UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA UNIVERSITAS UDAYANA DENPASARDENPASARDENPASARDENPASAR

ii
HALAMAN PENGESAHAN
A. Judul Penelitian: Penyediaan Metode Serologi Untuk Sertifikasi Bibit Krisan
(Dendranthema grandiflorum Kitam.) Bebas Chrysanthemum B
Carlavirus (CVB)
B. Ketua Peneliti
a) Nama lengkap : Dr. Ir. I Gede Rai Maya Temaja, MP.
b) Jenis kelamin : Laki-laki
c) Bidang keahlian : Fitopatologi
d) Pangkat/Golongan/NIP : Pembina Tk.I/IVb/19621009 198803 1 002
e) Unit kerja : Fakultas Pertanian Uniersitas Udayana
C. Tim peneliti
No Nama dan Gelar
Akademik
Bidang
Keahlian
Instansi Alokasi Waktu
(Jam/Minggu)
1 Dr. Ir. I Gede Rai
Maya Temaja, MP.
Fitopatologi Fak. Pertanian UNUD 18
2 Prof.Dr.Ir.Made
Sudana, MS.
Bioteknologi Fak. Pertanian UNUD 10
3 Dr. Ir. Gede Suastika,
M.Sc.
Virologi Fak. Pertanian IPB 18
D. Pendanaan dan jangka waktu penelitian
Jangka waktu penelitian yang diusulkan : 2 (dua) tahun
Biaya total yang diusulkan : Rp. 79.880.000,-
Biaya yang disetujui tahun II : Rp. 46.000.000,-
Denpasar, 27 Oktober 2011
Mengetahui Ketua Peneliti,
Dekan Fakultas Pertanian UNUD,
(Prof.Dr.Ir. I Nyoman Rai, MS) ( Dr. Ir. I Gede Rai Maya Temaja, MP)
NIP : 19630515 198803 1 001 NIP : 19621009 198803 1 002
Menyetujui
Ketua Lembaga Penelitian dan Pengabdian Masyarakat
Universitas Udayana
(Prof. Dr. Ir. I Ketut Satriawan, MT.)
NIP. 19640717 198903 1 001

iii
A. LAPORAN HASIL PENELITIAN
RINGKASAN
Chrysanthemum B Carlavirus (CVB) merupakan penyakit utama pada tanaman
krisan. Gejala yang ditimbulkan bervariasi dan sangat tergantung pada kultivar dan kondisi
lingkungan. Krisan terinfeksi CVB menunjukkan variasi gejala dari belang (mottle) ringan,
pemucatan tulang daun (vein-clearing) dan mosaik (mosaic) ringan pada daun; dan pecah
warna (color breaking) pada bunga. Penyakit ini terutama ditularkan melalui bahan
perbanyakan vegetatif tanaman, yaitu stek pucuk yang diambil dari tanaman induk
terinfeksi.
Dalam perdagangan internasional, sertifikasi bahan tanaman krisan bebas virus
adalah mutlak. Bahan tanaman krisan tersertifikasi untuk ekspor harus memenuhi
peraturan phytosanitary negara-negara pengimpor, terutama berkaitan dengan salah satu
organisme pengganggu tanaman karantina. Produksi tanaman krisan sehat membutuhkan
skrining bebas CVB untuk bahan tanaman krisan sebelum dipindahkan dari suatu negara
ke negara lainnya. Petunjuk sertiikasi krisan bebas CVB merekomendasikan infeksi CVB
dideterminasi dengan ELISA (serologi).
Keberhasilan dan ketelitian teknik serologi untuk mendeteksi dan mengidentifikasi
virus sangat tergantung pada ketersediaan pereaksi diagnostik seperti antiserum dengan
kualitas yang baik. Oleh sebab itu pada penelitian ini akan dilakukan kajian serologi CVB,
meliputi produksi antiserum dan pengujian metode serologi untuk deteksi sampel. Tujuan
penelitian ini adalah untuk mengetahui sifat antigenik siapan murni CVB isolat Indonesia
melalui immunisasi pada hewan mamalia dan menyediakan metode deteksi CVB berbasis
antiserum yang dapat digunakan untuk keperluan sertifikasi bibit krisan bebas virus.
Hasil penelitian menunjukkan bahwa imunisasi marmut menggunakan 150 µg
CVB murni dapat menghasilkan antiserum rata-rata 10,75 ml/ekor. Dengan metode
ELISA didapatkan bahwa titer antiserum tidak diserap, diserap dengan sap Nicotiana
benthamiana sehat dan antiserum murni adalah masing-masing 1/50.000, 1/50.000, dan
1/100.000.
Spesifisitas reaksi antiserum ditentukan dengan analisis Western blot dan
Immunosorbent electron microscopy (ISEM). Antiserum dengan pengenceran 1:1000
mampu mengkonfirmasi protein selubung CVB dengan ukuran pita sekitar 34 kDa, sesuai
dengan berat molekul protein selubung CVB yang digunakan untuk imunisasi pada

iv
hewan percobaan. Antiserum memberikan hasil dekorasi yang baik pada partikel CVB
yang diamati dibawah mikroskop elektron dengan metode ISEM.
Reaktivitas antiserum pada deteksi serologi diuji dengan I-ELISA dan TBIA.
Mendeteksi CVB dengan ELISA menunjukkan sensitifitas yang cukup tinggi dan virus
masih terdeteksi pada pengenceran cairan perasan tanaman terinfeksi 1/256. Pengenceran
sampel cairan perasan tanaman terinfeksi 1/16 menunjukkan reaksi yang kuat sampai pada
pengenceran antiserum 1000 kali dan masih menunjukkan reaksi positif sampai pada
pengenceran antiserum 100.000. Pada uji TBIA, reaksi positif masih nampak pada
pengenceran antiserum 10.000 kali. TBIA lebih menguntungkan dibandingkan dengan
teknik ELISA. Metode TBIA lebih mudah dan cepat, biayanya lebih murah karena hanya
diperlukan reagen yang lebih sedikit dan dapat menguji sampel lebih banyak. Metode ini
juga memungkinkan penyiapan (blotting) sampel dilakukan di lapangan, tanpa membawa
sampel ke laboratorium.

v
SUMMARY
Chrysanthemum B Carlavirus (CVB) is important disease of the florist’s
chrysanthemum. The symptoms of CVB disease was variable and are highly depend on
both of cultivar and environmental conditions. Infected chrysanthemum of CVB exhibited
a variety of symptoms ranging from mild mottle, vein-clearing and mild mosaic of leaves;
and color breaking of flowers. The disease is mainly transmitted by propagation, when
cuttings have been taken from infected mother plants.
In international trade, certification of virus-free chrysanthemum material is
absolutely essential. Certified chrysanthemum materials for export should in any case
satisfy the phytosanitary regulation of importing countries, especially respect to any of the
pathogens quarantine pests. Production of healthy plants of chrysanthemum demands the
screening of chrysanthemum materials for CVB before they can be transported from one
country to another. Guidelines for CVB in a certification scheme recomended that CVB
infection checked by ELISA (serology).
The success and accuracy of serological techniques to detect and identify viruses is
dependent on the availability of diagnostic reagents such as antibody with good quality.
Therefore in this study will be conducted serological studies of CVB, including antibody
production and examination of serological methods for sampel detection. The objective of
this study is to determine the antigenic characteristics of pure preparations of CVB
Indonesian isolates through immunization in mammals and provide an antibody-based
detection method that can used for certification of chrysanthemum virus-free.
The result showed that guinea pig immunization using 150 µg of purified virus was
able to produce 10.75 ml of antiserum. Using ELISA, the titre of crude antiserum,
antiserum absorbed by healthy Nicotiana benthamiana sap, and of purified antiserum was
1/50.000, 1/50.000, and 1/100.000 respectively.
Specificity of antiserum was determined by Western blot and immunosorbent
electron microscopy (ISEM) analysis. Antiserum dilution 1:1000 able to confirm the CVB
coat protein which molecular weight around 34 kDa, corresponding to the molecular
weight of CVB coat protein that used for immunization in animal experiments. Antiserum
gave good results decoration on CVB particles were observed under the electron
microscope by ISEM.
Antiserum reactivity in the serological detection tested using I-ELISA and TBIA
methods. Detecting by ELISA showed a fairly high sensitivity and the virus is still

vi
detectable at sap of infected plant dilutions 1/256. Sap of infected plant dilutions 1/16
showed a strong reaction to the antiserum dilution 1/1000 and still showed a positive
reaction to the antiserum dilution 1/100 000. In TBIA test, positive reaction was still
visible at antiserum dilution 1/10 000 times. TBIA more profitable than the ELISA
technique. TBIA prosedure is easier, faster, and less cost than ELISA. This method also
allows the samples blotting were conducted in the field, without bringing to the laboratory.

vii
PRAKATA
Krisan (Dendranthema grandiflorum Kitam.) merupakan salah satu jenis tanaman
hias bunga yang sangat diminati dan memiliki nilai ekonomi yang relatif tinggi di
Indonesia serta mempunyai prospek pemasaran cerah, baik itu pasar domestik maupun
ekspor. Kendala utama yang dihadapi petani dan pengusaha krisan adalah serangan
chrysanthemum B carlavirus (CVB) yang seringkali menjadi ganjalan ekspor bahan
tananam krisan, karena negara-negara pengimpor krisan mensyaratkan semua bahan
tanaman krisan yang diekspor harus sudah disertifikasi bebas CVB. Kendala ini semata-
mata muncul karena belum tersedianya metode deteksi yang handal untuk patogen ini.
Perlu dikaji metode deteksi CVB berbasis serologi yang memenuhi persyaratan sensitif,
massal dan ekonomis sehingga dapat diterapkan untuk pemenuhan kebutuhan sertifikasi.
Penulis mengucapkan terima kasih kepada Direktorat Jendral Pendidikan Tinggi,
Kementerian Pendidikan Nasional dan Rektor Universitas Udayana yang telah berkenan
memberikan dukungan dana penelitian untuk melakukan kajian penyediaan metode
serologi untuk sertifikasi bibit krisan bebas CVB. Penulis berharap keluaran penelitian ini
bermanfaat dalam program sertifikasi bibit krisan di Indonesia; sebagai perangkat
penunjang yang handal dalam menjalankan peraturan karantina internasional untuk
mencegah masuknya CVB isolat luar negeri yang mungkin terbawa dalam bahan tanaman
krisan yang diimpor ke dalam wilayah Indonesia; dan sebagai sumbangan pada
pengkayaan ilmu pengetahuan.
Denpasar, Oktober 2011
Ketua Pelaksana Kegiatan

viii
DAFTAR ISI
Halaman
HALAMAN PENGESAHAN ......................................................................... ii
A. LAPORAN HASIL PENELITIAN ............................................................. iii
RINGKASAN ............................................................................................... iii
SUMMARY .................................................................................................. v
PRAKATA ................................................................................................... vii
DAFTAR ISI ................................................................................................ viii
DAFTAR GAMBAR .................................................................................... ix
I. PENDAHULUAN ..................................................................................... 1
II. TUJUAN DAN MANFAAT PENELITIAN ........................................... 2
III. STUDI PUSTAKA ................................................................................. 3
IV. METODE PENELITIAN ....................................................................... 8
V. HASIL DAN PEMBAHASAN ............................................................... 15
VI. KESIMPULAN ...................................................................................... 28
DAFTAR PUSTAKA ................................................................................... 29

ix
DAFTAR GAMBAR
Halaman
1. Hasil pemurnian virus dengan pemisahan virus melalui ultrasenrifugasi
dengan gradien kepekatan sesium sulfat ....................................................
15
2. Profil spektrofotometri siapan virus murni ................................................... 16
3. Jumlah tanaman N. benthamiana, N. clevelandii, N. tabacum var. White
Burley, N. tabacum var. Havana, P. hybrida, C. amaranticolor dan C.
quinoa yang menunjukkan titer virus tinggi ................................................
17
4. Bentuk partikel CVB pada mikroskop elektron ........................................... 18
5. Hasil analisis SDS-PAGE (A) dan Western blot (B) protein selubung CVB
isolat Indonesia .............................................................................................
19
6. Nilai absorbansi antiserum pada panjang gelombang 280 ........................... 20
7. Titer antiserum tidak diserap (As K), antserum diserap (As S) dan
antiserum murni (As M) berdasarkan ELISA. I .........................................
22
8. Nilai absorbansi antiserum tidak diserap, diserap dan antiserum murni
pada uji ELISA terhadap isolat CVB dari berbagai lokasi (Cianjur,
Medan, Malang dan Bali) ............................................................................
23
9. Hasil analisis western blot protein selubung CVB menggunakan
antiserum yang diproduksi ..........................................................................
24
10. Partikel CVB pada pengamatan dengan mikroskop elektron ..................... 24
11. Reaktivitas antiserum pada pengenceran 1/100, 1/500, 1/1000, 1/5000
dan 1/10.000 terhadap isolat CVB dari berbagai lokasi pada uji serologi
dengan TBIA ................................................................................................
26
12. Deteksi serologis CVB pada tanaman krisan dengan teknik TBIA dengan
pengenceran antiserum 1/10.000 ................................................................
27

1
I. PENDAHULUAN
Krisan (Dendranthema grandiflorum Kitam.) adalah salah satu jenis bunga yang
banyak diminati oleh masyarakat dunia karena daya tarik warna, bentuk, dan ukurannya
yang beranekaragam. Di Indonesia, krisan banyak dibudidayakan oleh petani dalam skala
kecil maupun dalam skala besar oleh perusahaan agribisnis terutama di daerah sejuk seperti
Bogor, Bandung, Sumatera Utara dan daerah lain. Disamping untuk memenuhi kebutuhan
dalam negeri, krisan diproduksi terutama untuk memenuhi kebutuhan luar negeri seperti
negara-negara Eropa, Jepang, dan negara Asia lainnya. Ekspor bisa dalam bentuk bunga
potong, tetapi yang lebih banyak adalah ekspor dalam bentuk stek batang (PT. Saung
Mirwan, komunikasi pribadi).
Dalam budidaya tanaman krisan, banyak kendala yang dihadapi oleh para petani
dan pengusaha. Salah satu kendala tersebut adalah serangan chrysanthemum B Carlavirus
(CVB) (Verma et al. 2003). Infeksi CVB pada tanaman krisan dapat bermanifestasi dalam
berbagai gejala. Pertumbuhan tanaman menjadi terhambat (stunting) dengan perakaran
yang terbatas. Daun-daun berukuran kecil dan berwarna hijau pucat. Kandungan
anthocyanin pada batang sangat rendah. Pembungaan lebih cepat (beberapa hari)
dibandingkan dengan yang sehat. Bunga yang dihasilkan berukuran sangat kecil dengan
warna lebih pudar. Akibat infeksi virus yang sangat fatal ini mendorong negara-negara
pengimpor krisan menerapkan peraturan Karantina Internasional secara ketat.
Implikasinya adalah semua bahan tanaman krisan yang diekspor harus sudah disertifikasi
bebas CVB.
Indonesia yang mempunyai potensi luar biasa dalam mengembangkan produksi
krisan seringkali terganjal ekspornya akibat keberadaan CVB dalam jaringan tanaman (PT.
Saung Mirwan, komunikasi pribadi). Kendala ini semata-mata muncul karena belum
tersedianya metode deteksi CVB yang handal yang dapat digunakan dalam sertifikasi.
Metode berbasis serologi yang akan dikembangkan dalam penelitian ini diharapkan dapat
menjawab kebutuhan yang sangat urgen ini.

2
II. TUJUAN DAN MANFAAT PENELITIAN
2.1 Tujuan
Tujuan penelitian ini adalah mengetahui sifat antigenik siapan murni CVB isolat
Indonesia melalui immunisasi pada hewan mamalia dan menyediakan metode deteksi CVB
berbasis antiserum yang dapat digunakan untuk keperluan sertifikasi bibit krisan bebas
virus.
2.2 Manfaat
Hasil penelitian ini diharapkan dapat memberikan sumbangan pada pengkayaan
ilmu pengetahuan dan teknologi antara lain melalui penyediaan metode deteksi CVB
secara serologis yang handal dan dapat diterapkan dalam program sertifikasi bibit
krisan di Indonesia. Metode deteksi CVB yang akan dikembangkan dalam penelitian ini
diharapkan dapat digunakan sebagai perangkat penunjang yang handal dalam menjalankan
peraturan karantina internasional, yakni mencegah masuknya CVB isolat luar negeri yang
mungkin terbawa dalam bahan tanaman krisan yang diimpor ke dalam wilayah Indonesia.
Demikian juga sebaliknya dapat mencegah peristiwa ditolaknya ekspor bibit krisan
Indonesia karena sebelumnya telah dilakukan pemeriksaan/deteksi yang memadai melalui
metode serologi yang akan dihasilkan dalam penelitian ini. Sumbangan pada pengkayaan
ilmu pengetahuan yang lain adalah publikasi hasil penelitian yang diusulkan pada jurnal
ilmiah nasional maupun internasional.

3
III. STUDI PUSTAKA
3.1 Kisaran Inang dan Gejala Infeksi CVB
Chrysanthemum B carlavirus (CVB), atau nama lainnya adalah chrysanthemum
mild mosaic virus, chrysanthemum virus B, chrysanthemum Q virus, chrysanthemum dwarf
mottle, chrysanthemum necrotic mottle, gynura latent virus, dan chrysanthemum vein
mottle virus adalah virus dari genus carlavirus (Hollings & Stone, 1972; Hakkart & Maat,
1974).
CVB mempunyai kisaran inang yang sempit, menginfeksi tanaman krisan dan
sekitar 10 spesies pada 5 famili dikotiledon (Hollings, 1957). Pada penelitian Verma et al.
(2003), dari 22 spesies tanaman yang diinokulasi untuk uji kisaran inang, hanya 5 spesies
yang terinfeksi, yaitu Nicotiana clevelandii, N. glutinosa, N. rustika, Petunia hybrida dan
Vicia faba. Sedangkan penelitian Suastika et al. (1997) menemukan bahwa CVB mampu
menginfeksi tanaman G. savateri, N. clevelandii, N. benthamiana, N. occidentalis, Petunia
hybrida, Helichrysum bracteatum, Zinnia elegans, Chenopodium amaranticolor, C. quinoa,
Sesamum indicum dan Tetragonia expansa. Sedangkan 13 spesies tanaman lainnya dari 7
famili yang diuji, tidak terinfeksi.
Moran (1987) melaporkan bahwa beberapa spesies tanaman yang sesuai untuk
penyebaran CVB antara lain Aster amellus, Chrysanthemum carinatum, C. morifolium, N.
clevelandii, N. glutinosa, N. rustika, N. affinis. Sedangkan yang bukan inang CVB antara
laian Datura stramonium, C. amaranticolor, C. quinoa, Gompphera globosa, Capsicum
annuum, Cucumis sativus, Licopersicon esculentum.
Walaupun memiliki inang yang terbatas, tetapi dilaporkan CVB tersebar pada
pertanaman krisan di seluruh dunia. Infeksi virus ini pada tanaman krisan menyebabkan
perubahan fisiologi tanaman, yang berakibat gejala motling daun atau vein-clearing yang
sangat mild pada beberapa kultivar, dapat juga berkembang gejala brown necrotic streak.
Namun demikian tidak jarang kultivar yang terinfeksi virus ini tidak menunjukkan gejala
(symptomless) (Hollings & Stone, 1972). Beberapa varietas terinfeksi menunjukkan
penurunan kualitas bunga dibandingkan dengan tanaman yang bebas virus. Penurunan
kualitas bunga terutama karena pada tanaman terinfeksi warna mahkota bunga terputus-
putus (flower breaks), mengalami distorsi dan kerdil. Kadang-kadang pada krisan
terinfeksi CVB berkembang brown necrotic streaks pada floret (Hollings & Stones, 1972;
Moran, 1987).

4
Survey Verma et al. (2003) di Himachal Pradesh (India) menemukan bahwa
tanaman krisan diserang CVB dengan gejala vein banding, mosaic, mottling, dan vein
clearing yang lemah. Pada tanaman dengan daun menunjukkan gejala mosaic yang keras,
bunganya juga mengalami malformasi.
3.2 Penularan CVB
Semua anggota genus Carlavirus diketahui dapat ditularkan secara mekanis, dan
sebagian besar juga ditularkan secara non persisten melalui kutudaun (Foster, 1992),
walaupun ada yang ditularkan oleh kutu kebul (Bemisia tabaci) yaitu cowpea mild mottle
virus (CPMMV) (Badge et al.1996; Hull, 2002). CPMMV juga dapat ditularkan melalui
benih, seperti halnya pea strike virus (PeSV) dan red clover vein mosaic virus (RCVMV)
(Hull, 2002).
CVB bisa ditularkan secara non persisten oleh kutudaun, melalui penyambungan
dan secara mekanik dengan cairan perasan tanaman sakit, tetapi tidak dapat ditularkan
melalui kontak antar tanaman dan melalui benih. Di lapangan virus ini dapat ditularkan
oleh kutudaun Myzus persicae, Macrosiphum euphorbiae, Aulacorthum solani, Coloradoa
rufomaculata, dan Macrosiphoniella sanborni (Hollings & Stone, 1972; Moran, 1987).
Hollings & Stone (1972) melaporkan bahwa periode makan akuisisi kutudaun 30-
45 menit dan periode puasanya hingga 1 jam. Frekuensi penularan virus meningkat setelah
serangga dilaparkan 3 jam sebelum periode makan akuisisi.
3.3 Morfologi dan taksonomi CVB
CVB adalah virus yang mempunyai partikel berbentuk batang agak lurus dan
flexuous, dengan ukuran 680-685 nm x 12-13 nm. Partikel lebih lurus dan kelihatan kurang
flexuous dibandingkan Carnation latent virus, dan cenderung terputus-putus bila dilakukan
pengecatan dengan phosphotungstate (Hollings & Stone, 1972; Suastika et al. 1997; Foster,
1992; Verma et al. 2003).
Menurut Hull (2002), CVB termasuk ke dalam genus mengambang (floating
genera), karena genus-genus pada kelompok ini tidak bisa dimasukkan dalam famili.
Sampai saat ini ada 20 genus virus tanaman yang dimasukkan dalam floating genera ini
dengan karakterisasi mempunyai asam nukleat ribonukleat dalam bentuk utas tunggal
(single strand (ss) RNA) positive-sense dan tidak memenuhi syarat untuk dikelompokkan
pada familia tertentu.

5
Salah satu genus virus yang masuk dalam kelompok floating genera adalah
Carlavirus yang anggota jenisnya dicirikan dengan tipe Carnation latent virus (CLV),
mempunyai partikel bentuk batang agak flexuous dengan ukuran panjang 610-700 nm dan
diameter 12-15 nm. Virion mengandung molekul RNA tunggal linier dalam bentuk
positive-sense utas tunggal (positive-sense ssRNA), panjangnya 7,4-8,5 nm, dan memiliki
poly (A) pada ujung 3’. Asam nukleatnya diselubungi oleh coat protein tunggal dengan
berat 31-36 kDa (Hull, 2002; Lee et al. 2003; Zavriev et al. 1991; Foster, 1992; Lawrence
et al. 1995).
Lebih dari 35 spesies virus tergolong dalam genus Carlavirus, enam diantaranya
sudah dilaporkan sekuen lengkap nukleotidanya yaitu Actonitum latent virus (AcoLV),
Blueberry scorch virus (BlScV), Garlic latent virus (GarLV), Hop latent virus (HpLV),
Lily symptomless virus (LSV), dan Potato virus M (PVM) (Cohen et al. 2000; Chen et al.
2002; Fuji et al. 2002; Hataya et al. 2000; Zavriev et al. 1991). Beberapa spesies dari
carlavirus sudah disekuensing pada daerah terminal 3’nya, termasuk diantaranya PVS,
HeLVS, CLV, LSV, CVB, dan CPMMV (Badge et al. 1996; Foster & Mills, 1992).
Semuanya menunjukkan organisasi genom yang mirip, dengan kemiripan ukuran open
reading frame (ORF), dan tingkat homologi yang tinggi pada sekuen asam amino diantara
protein berkorespondensi.
RNA dengan poly A pada ujung 3’, memiliki 6 ORF yaitu ORF1 menyandi
polipeptida (223 kDa) yang merupakan replikase virus; ORF2 (25 kDa), ORF3 (12 kDa)
dan ORF4 (7 kDa) membentuk triple gene block (TGB) yang berperan dalam pergerakan
virus dari sel ke sel; ORF5 menyandi protein selubung (34 kDa) dan ORF6 (11-16 kDa)
menyandi protein cystein-rich yang belum diketahui fungsinya.
3.4 Identifikasi dan Deteksi CVB
Identifikasi virus secara biologis dapat dilakukan melalui uji kisaran inang dan
atau melalui uji hubungan virus dan serangga vektornya. Cara tersebut telah dilakukan
untuk mendeteksi dan mengidentifikasi CVB (Hollings & Stones, 1972; Moran, 1987;
Ram et al. 2005; Verma et al. 2003; Suastika et al. 1997).
Uji serologi dengan memanfaatkan reaksi antigen dan ector y merupakan cara
lain dalam mengidentifikasi virus penyebab penyakit tumbuhan. Kegunaan yang lain dari
uji serologi ini adalah mengukur konsentrasi virus dalam jaringan tumbuhan, mendeteksi
virus tumbuhan dalam tubuh serangga ector dan untuk mengetahui hubungan
kekerabatan antar virus (Agrios, 1997). Deteksi dan identifikasi secara serologi sudah

6
umum diaplikasikan untuk berbagai virus. Ada beberapa cara yang digunakan untuk
deteksi serologi antara lain ISEM, immunoflourescent staining (Hampton et al. 1990),
presipitasi dalam tabung, aglutinasi kloroplas, flokulasi lateks, gel double-diffusion test,
DIBA, immunoblotting atau western blotting, dan ELISA (Harlow & Lane 1999).
Metode serologi yang telah berhasil dikembangkan untuk mendeteksi virus
tumbuhan diantaranya yaitu metode DIBA digunakan untuk mendeteksi ZYMV
(Somowiyarjo et al. 1989). Abouzid et al. (2002) menggunakan ELISA dan western
blotting untuk mendeteksi beberapa protein selubung virus dari genus Begomovirus.
Bentuk partikel TYLCV telah berhasil dideteksi dan diidentifikasi oleh Attathom et al.
(1990) dengan menggunakan metode ISEM. Deteksi partikel virus menggunakan
antiserum dengan metode Immune Electron Microscopy (IEM) dilakukan oleh Sharma et
al. (2005) terhadap virus Cucumber mosaic virus (CMV), Lily mottle virus (LmoV) dan
Lily symptomless virus (LSV) pada tanaman lily. Deteksi dengan metode
immunoflourescent staining juga telah berhasil dilakukan oleh Sudarshana et al. (1997)
untuk mengetahui dinamika pergerakan Bean dwarf mosaic geminivirus (BDMV) dari sel
ke sel pada Phaseolus vulgaris. Pengujian presipitasi dengan memanfaatkan reaksi difusi
antara antigen dan antibodi telah berhasil dilakukan oleh Mahmood et al. (1997) untuk
mendeteksi Wheat streak mosaic tritimovirus (WSMV).
Deteksi CVB secara serologis menggunakan antiserum spesifik dilaporkan telah
dipakai untuk mendeteksi CVB pada tanaman krisan dengan metode double antibody
sandwich-enzyme linked immunosorbent assay (DAS-ELISA) (Ram et al. 2005; Verma et
al. 2003). Verma et al. (2003) juga menggunakan antiserum CVB untuk melihat partikel
virus pada kajian IEM. Pada tanaman Gymnaster savatieri antiserum CVB digunakan
untuk deteksi virus secara serologis dengan metode indirect-enzyme linked immunosorbent
assay (I-ELISA) dan untuk Immunosorbent electron microscopy (ISEM) (Suastika et al.
1997).
Sekarang ini, metode deteksi yang didasarkan pada analisis asam nukleat virus
mulai digunakan untuk mendeteksi dan mengidentifikasi virus. Sebagai contoh teknik
reverse transcriptase polymerase chain reaction (RT-PCR) dengan menggunakan primer
terbukti dapat digunakan untuk mendeteksi dan mengidentifikasi CMV, LmoV, LSV dan
Strawberry latent ringspot virus (SLRSV) pada tanaman lily (Sharma et al. 2005; Choi &
Ryu, 2003). Identifikasi molekuler dengan teknik RT-PCR juga telah digunakan untuk
mengkarakterisasi Daphne virus S (DVS) pada tanaman Daphne spp. (Lee et al. 2003);
Garlic Latent Virus (GarLV), Onion yellow dwarf virus (OYDV) dan Leek yellow stripe

7
virus (LYSV) pada tanaman bawang putih; Shallot yellow stripe virus (SYSV) pada
tanaman Allium fistulosum var. Caespitosum; dan LmoV pada tanaman lily (Chen et al.
2002).
Analisis asam nukleat CVB dengan teknik RT-PCR dilakukan oleh Levay dan
Zavriev pada tahun 1991. Ram et al. (2005) pada penelitiannya memproduksi tanaman
krisan bebas CVB menggunakan teknik RT-PCR untuk mendeteksi dan mengkarakterisasi
CVB dari tanaman yang berbeda dan tempat yang berbeda.
Metode ELISA dan RT-PCR adalah metode yang lebih dapat dipercaya dan lebih
sensitif sebagai metode pendeteksi virus atau indexing, dibandingkan dengan kajian
biologis. Sering terjadi bahwa virus tidak terdeteksi pada kajian biologis, tetapi
menunjukkan hasil posistif dengan metode ELISA dan RT-PCR yang dapat mendeteksi
virus pada konsentrasi rendah (Ram et al. 2005).

8
IV. METODE PENELITIAN
4.1 Perbanyakan Isolat CVB
Tanaman krisan terinfeksi CVB (diverifikasi melalui ELISA) yang telah dikoleksi
dari Kebun Percobaan Balai Penelitian Tanaman Hias di Segunung, Cianjur, Jawa Barat
digunakan sebagai bahan penelitian (sumber inokulum). Isolat CVB ini kemudian
diperbanyak pada tanaman tembakau (Nicotiana benthamiana). Bibit N. benthamiana
ditanam dari biji yang sehat. Tanaman umur tiga minggu diinokulasi secara mekanis
dengan cairan perasan tanaman N. benthamiana terinfeksi CVB. Sampel daun terinfeksi
digerus dan ditambahkan larutan bufer fosfat 0,01 M, pH 7,0 ditambahkan dengan
perbandingan 1 g daun per 5 ml larutan bufer fosfat (1:5 b/v). Cairan perasan inokulum ini
segera diinokulasikan ke bagian daun tanaman inang perbanyakan. Sebelum diinokulasi,
permukaan daun ditaburi dengan dioleskan karborundum 600 mesh, kemudian cairan
perasan inokulum dioleskan dengan cotton bud pada permukaan daun yang dilakukan
searah tulang daun tanpa digosok berlawanan arah. Setelah pengolesan inokulum,
dilakukan pembilasan sisa-sisa karborundum yang masih melekat pada permukaan daun
tanaman uji dengan akuades. Setelah 17 hari dari saat inokulasi, daun dan ranting
tanaman dipanen dan tunas-tunas yang tumbuh dibiarkan untuk dapat dipanen pada periode
panen berikutnya Daun dan ranting hasil panen digunakan sebagai bahan untuk pemurnian
virus.
4.2 Purifikasi CVB
Metode pemurnian CVB dilakukan sesuai dengan prosedur dari Foster (1998).
Daun tanaman N. benthamiana terinfeksi (200 g) digerus dan dibantu dengan penambahan
nitrogen cair. Serbuk hasil gerusan tersebut kemudian ditambah bufer borat 0,5 M, pH 7,8
(bufer borat 0,5M mengandung EDTA 0,005 M; driselase 0,5% w/v dan β-2
mercaptoethanol 1% v/v) dengan rasio 1:3 (w/v). Dipindahkan ke dalam gelas beaker dan
ditambahkan chloroform 15% (v/v), selanjutnya diaduk menggunakan mesin pengaduk
bermagnit (magnetic stirrer). Disentrifugasi pada 5000 g selama 20 menit (pada suhu 4oC).
Supernatan diambil ditambahkan polyethyleneglycol (PEG) 6% (w/v), diaduk pada 4oC
semalam. Ambil presipitat yang dihasilkan dengan sentrifugasi pada 10.000 g selama 15
menit dan diresuspensi pada bufer borat 0,5 M yang mengandung Triton X-100 0,5% (w/v).
Sentrifugasi pada kecepatan rendah (10.000 g selama 15 menit) dan pindahkan supernatan

9
ke tabung baru. Virus diendapkan dengan sentrifugasi pada kecepatan tinggi (30.000 g
selama 90 menit) dan pelet yang dihasilkan diresuspensi pada bufer borat 0,05 M yang
mengandung Triton-X 1% (w/v). Virus dimurnikan dengan sentrifugasi pada cesium
chloride (0,439 g/ml), sentrifugasi pada 110.000 g selama 20 jam pada suhu 5oC. Pita
virus diambil dari gradien cesium dan dilarutkan pada bufer borat 0,005 M (volume bufer
minimal dua kali). Virus dikumpulkan dengan sentrifugasi pada 40.000 g selama 90 menit
dan pelet diresuspensi dalam 0,5 ml akuades steril. Virus segera dapat digunakan, simpan
pada suhu 4oC selama 1-2 hari atau tempatkan pada suhu -20
oC untuk penyimpanan lama.
Tingkat kemurnian virus diukur menggunakan spektrofotometer pada panjang
gelombang 260 nm (A260) dan 280 nm (A280). Perbandingan antara nilai A260/A280 dapat
digunakan untuk mengetahui tingkat kemurnian virus (Dijkstra & de Jager, 1998).
Konsentrasi virus dihitung berdasarkan nilai A260 dan extinction coefficient (E%1.0
260.1 nmcm ) ,
dengan formulasi:
Keterangan:
C : konsentrasi virus (mg/ml)
A260 : nilai absorbansi pada panjang gelombang 260 nm
E %1.0
260.1 nmcm : nilai extinction coefficient
4.3 Produksi Antiserum CVB.
Antiserum diproduksi dengan melakukan imunisasi terhadap marmut jantan
berumur sekitar 6 bulan. Imunisasi dilakukan dengan menginjeksikan virus murni beserta
larutan adjuvant. Injeksi dilakukan di bawah kulit (sub cutaneous) 5 kali dengan interval
waktu 2 minggu. Injeksi pertama dengan dosis 50 µg virus murni yang ditambah
complete adjuvant dengan perbandingan 1:1. Injeksi selanjutnya dengan dosis 25 µg virus
murni yang ditambah incomplete adjuvant dengan perbandingan 1:1. Sepuluh hari setelah
penyuntikan terakhir, dilakukan panen darah dengan cara mengambil darah melalui jantung
dan disembelih. Butir darah merah dibiarkan mengendap dan disimpan dalam suhu 4oC
selama 12 jam. Cairan bening diambil dengan hati-hati kemudian disentrifugasi pada
kecepatan 8.000 g (rotor TMA-5, Tomy) selama 20 menit, selanjutnya antiserum yang
C = A260
E %1.0
260.1 nmcm

10
dihasilkan dapat dimanfaatkan untuk pengujian serologi atau disimpan pada suhu – 20oC
dengan ditambah sodium azide 1%.
Antiserum yang dihasilkan bisa langsung digunakan untuk pengujian, dan untuk
meningkatkan kualitas dilakukan penyerapan menggunakan cairan perasan tanaman N.
benthamiana sehat serta pemurnian gamma-globulin menggunakan amonium sulfat.
Konsentrasi antiserum semua jenis antiserum (tidak diserap, diserap dan murni) diukur
berdasarkan nilai absorbansi 1,4 pada panjang gelombang 280 (A280) setara dengan
konsentrasi 1 mg/ml.
4.4 Penyerapan Antiserum Menggunakan Cairan Perasan Tanaman N. benthamiana
Sehat
Penyerapan antiserum dilakukan berdasarkan metode Dijkstra & de Jager (1998),
yaitu antiserum diserap dengan cairan perasan N. benthamiana sehat untuk menghindari
terjadinya reaksi silang. Daun N. benthamiana sehat digerus dalam larutan PBST dengan
perbandingan 1:20 (b/v), sehingga diperoleh cairan perasan tanaman. Antiserum dicampur
dengan cairan perasan tanaman tersebut degan perbandingan 1 : 1, digoyang dengan shaker
kemudian diinkubasikan pada suhu 37 oC selama 2 jam, dan pada suhu 4
oC selama 1 jam.
Campuran disentrifugasi pada 8.000 g (rotor TMA-5, Tomy) selama 15 menit, supernatan
diambil dan disimpan dalam suhu 4 oC selama 6 jam. Selanjutnya larutan disentrifugasi
pada 8000 g (rotor TMA-5, Tomy) selama 15 menit. Supernatan diambil dan siap
digunakan untuk uji serologi.
4.5 Isolasi Gamma-globulin Menggunakan Amonium Sulfat
Antiserum yang dihasilkan dimurnikan menggunakan metode presipitasi dengan
amonium sulfat jenuh menurut metode Clarck & Adams (1977). Satu ml antiserum
diencerkan dengan 9 ml akuades, kemudian ditambah 10 ml amonium sulfat jenuh secara
perlahan. Setelah diinkubasi selama 30 menit kemudian disentrifugasi pada 3.000 rpm
selama 15 menit. Hasil presipitat ditambah 2 ml PBS 0,01 M, pH 7,4 yang mengandung
sodium azide 0,01%. Campuran kemudian didialisis dalam 500 ml PBS 0,01M, pH 7,4
pada suhu 4 oC dengan 3 kali penggantian larutan penyangga setiap 8 jam.

11
4.6 Spesifisitas Antiserum
Spesifisitas antiserum yang diproduksi pada penelitian ini diamati dengan analisis
Western blot dan Immunosorbent electron microscopy (ISEM) .
Analisis Western Blot. Pengujian Western blot dilakukan berdasarkan metode
Towbin et al. (1979) menggunakan antiserum CVB. Siapan suspensi virus murni, protein
tanaman yang terinfeksi CVB dan tanaman sehat dielektroforesis pada 10% gel SDS-
PAGE, kemudian ditransfer ke membran nitroselulosa (Amersham Hybond-P PVDF
membrane, GE Health Care, UK). Membran diinkubasi dalam antiserum dengan
pengenceran 1:500 dalam tris buffer saline (TBS) (50 mM Tris-HCl, 50 mM NaCl, pH 7,5)
selama 1-2 jam. Membran dicuci dengan TBST (TBS + 0,5% Tween 20) selama 10 menit
sebanyak 3 kali. Setelah pencucian, membran diinkubasi dalam konjugat alkaline
phosphatase (Sigma Chemical Co., USA) pada pengenceran 1:1000 dalam TBS selama 2-3
jam. Kemudian dilakukan pencucian sebagaimana di atas, dan diwarnai dengan substrat
BCIP/NBT (Sigma Chemical Co.,USA). Semua tahapan pengujian ini dilakukan pada suhu
ruang. Reaksi positif ditunjukkan dengan perubahan warna ungu pada membran. Reaksi
pewarnaan dihentikan dengan mencuci membran dengan akuabides dan dikeringanginkan.
SDS-PAGE dilakukan menurut metode Leammli (Gall et al. 1980). Analisis
dilakukan terhadap siapan virus murni, protein tanaman yang terinfeksi CVB dan tanaman
sehat. Ekstraksi protein tanaman dilakukan dengan menggerus 0,2 g jaringan daun muda
tanaman uji dalam nitrogen cair hingga halus. Hasil gerusan dimasukkan ke dalam tabung
mikro ukuran 2 ml dan ditambahkan dua kali volume bufer ekstraksi (sucrose 30%, SDS
2%, 2-mercaptoethanol 1% dan Tris-HCl 60 mM, pH 6,8). Selanjutnya larutan tersebut
disentrifugasi pada kecepatan 13.000 g (rotor TMA-5, Tomy) selama 5 menit. Supernatan
dipisahkan dan digunakan sebagai sampel uji. Virus murni dan protein tanaman total
masing-masing ditambah loading dye (SDS 4%, 2-mercaptoethanol 2%, bromophenol blue
0,001%, glycerol 2% dan Tris-HCl 0,1 M, pH 8,8) dengan perbandingan 1:4 (v/v).
Campuran tersebut kemudian dipanaskan pada suhu 100 oC selama 10 menit dan masing-
masing 15 µl campuran dimasukkan ke dalam sumuran gel polyacrylamide 12%. Marker
protein yang digunakan adalah Low Molecular Weight (LMW) (Amersham Biosciences,
UK) yang mengandung phosphorilase b (97 kDa), albumin (66 kDa), ovalbumin (45 kDa),
carbonic anhydrase (30 kDa), trypsin inhibitor (20,1 kDa), α-lactabumin (14,4 kDa).

12
Analisis SDS-PAGE membutuhkan dua jenis gel yaitu separating gel dan stacking
gel. Separating gel dibuat dengan cara mencampur acrylamide 10% (1,8 ml) dengan bis-
acrylamide 0,25% (1,5 ml), Tris-HCl 0,375 M pH 8,8 (2,5 ml), SDS 0,1% (0,005 ml),
TEMED 0,025% (0,006 ml), APS (amonium persulphate) 0,025% (0,17 ml), dan 4 ml air.
Campuran dimasukkan ke dalam pelat gelas ukuran 10 x 7,2 cm yang telah dipasang pada
gel stand. Stacking gel dibuat dengan cara mencampur acrylamide 3% (0,06 ml) dengan
bisacrylamide 0,08% (0,04 ml), Tris-HCl 0,125 M (2,5 ml) pH 6,8, SDS 0,1% (0,1 ml),
TEMED 0,025% (0,006 ml), APS 0,025% (0,1 ml), dan 6,3 ml air. Campuran dituang di
atas separating gel, kemudian dipasang sisir dan setelah gel membeku sisir dicabut dan gel
siap digunakan. Marker protein, virus murni, dan protein tanaman total dimasukkan ke
sumuran yang telah dicetak oleh sisir pencetak, kemudian dielektroforesis pada 80 V
selama 3 jam.
Setelah dielektroforesis, protein divisualisasi dengan pewarnaan Coomassie blue
(Bio-Rad Laboratories, USA). Gel direndam dalam asam asetat glasial 12,5% selama 5
menit kemudian direndam dalam larutan Coomassie blue 0,25% dan diinkubasi selama 12
jam sambil digoyang. Gel dicuci menggunakan larutan penghilang warna yang terdiri atas
metanol 50% dan larutan asam asetat 10% sebanyak 3 kali masing-masing 10 menit.
Pengujian Western blot dilakukan berdasarkan metode Towbin et al. (1979)
menggunakan antiserum CVB. Siapan suspensi virus murni, protein tanaman yang
terinfeksi CVB dan tanaman sehat dielektroforesis pada 10% gel SDS-PAGE, kemudian
ditransfer ke membran nitroselulosa (Amersham Hybond-P PVDF membrane, GE Health
Care, UK). Membran diinkubasi dalam antiserum dengan pengenceran 1:500 dalam tris
buffer saline (TBS) (50 mM Tris-HCl, 50 mM NaCl, pH 7,5) selama 1-2 jam. Membran
dicuci dengan TBST (TBS + 0,5% Tween 20) selama 10 menit sebanyak 3 kali. Setelah
pencucian, membran diinkubasi dalam konjugat alkaline phosphatase (Sigma Chemical
Co., USA) pada pengenceran 1:1000 dalam TBS selama 2-3 jam. Kemudian dilakukan
pencucian sebagaimana di atas, dan diwarnai dengan substrat BCIP/NBT (Sigma Chemical
Co.,USA). Semua tahapan pengujian ini dilakukan pada suhu ruang. Reaksi positif
ditunjukkan dengan perubahan warna ungu pada membran. Reaksi pewarnaan dihentikan
dengan mencuci membran dengan akuabides dan dikeringanginkan.
Immunosorbent electron microscopy (ISEM). Teknik ISEM dilakukan
berdasarkan metode Dykstra (1992). Satu tetes suspensi antiserum diteteskan pada cawan
Petri yang sudah dilapisi membran parafilm. Grid yang sudah dilapisi colodion dan

13
dikarbonisasi diletakkan secara perlahan pada tetesan suspensi antiserum. Bagian grid
yang terlapisi dibuat menempel pada suspensi dan diinkubasi selama 30 menit pada suhu
25oC. Grid dicuci dengan sepuluh tetes PBS. Siapan virus murni diteteskan pada bagian
lain di atas parafilm dan grid diletakan pada tetesan siapan virus murni tersebut, kemudian
diinkubasikan selama 15 menit. Grid dicuci sebagaimana pencucian sebelumnya. Grid
dikeringkan, dan segera setelah itu bagian grid yang terlapisi diletakkan pada cairan
pewarna. Grid dikeringkan dengan kertas saring dan diamati dengan mikroskop elektron
transmisi model JEOL 1010 yang dioperasikan pada 80 kV. Siapan virus murni tanpa
didekorasi dengan antiserum dipakai sebagai kontrol.
4.7 Reaktivitas Antiserum
Reaktivitas antiserum yang dihasilkan dikaji dengan uji serologi yaitu Enzyme-
Linked Immunosorbent Assay (ELISA) dan Tissue Immunobinding Assay (TIBA).
Enzyme-Linked Immunosorbent Assay (ELISA). Metode ELISA dilakukan
berdasarkan metode Stack & Macmillan (2005). Sampel tanaman terinfeksi CVB digerus
dalam sample extraction buffer dengan perbandingan 1:5 (b/v). Sampel tersebut
dimasukkan ke dalam masing-masing sumuran pada plat mikrotiter ELISA sebanyak 100
µl dan diinkubasi pada suhu 4 oC selama semalam. Selanjutnya masing-masing sumuran
plat mikrotiter dicuci sebanyak 6 kali dengan PBST (8 g sodium chloride, 0,2 g monobasic
potassium phosphate, 1,15 g dibasic sodium phosphate, 0,2 g potassium chloride, 0,2 g
sodium azide dan 0,5 g Tween-20 yang dilarutkan dalam 1000 ml H2O, pH 7,4). Sumuran
plat mikrotiter diisi dengan 100 µl antiserum yang telah dilarutkan dalam bufer ECI (0,2 g
bovine serum albumin, 2 g polyvinylpyrrolidone, 0,02 g sodium azide dan dilarutkan dalam
100 ml PBST, pH 7,4) dan kemudian diinkubasi pada suhu 37 oC selama 2 jam. Masing-
masing sumuran dicuci kembali dengan PBST dan diisi 100 µl konjugat (anti rabbit-IgG,
Sigma, USA) yang dilarutkan dalam bufer ECI (1:1.000) dan diinkubasi pada suhu ruang
selama 2 jam. Sumuran plat mikrotiter dicuci dengan PBST, kemudian diisi dengan 100
µl substrat PNP yang dilarutkan dalam bufer PNP (97 ml diethanolamine, 600 ml H2O, 0,2
g sodium azide dilarutkan dalam 1000 ml H2O, pH 9,8). Setelah diinkubasi pada suhu
ruang selama 30 menit dilakukan pengamatan secara kuantitatif dengan menggunakan
ELISA reader pada panjang gelombang 405 nm. Reaksi dihentikan dengan cara
menambahkan larutan NaOH 3 M sebanyak 50 µl ke dalam masing-masing sumuran.
Kontrol negatif yang digunakan adalah tanaman sehat dan bufer.

14
Tissue Immunobinding Assay (TIBA). Pengujian TBIA dilakukan berdasarkan
metode Lin et al. (1990). Sampel daun dipotong melintang pada bagian pangkal daun yang
masih putih dan dibuat tepi potongan yang rata. Potongan ini kemudian ditempelkan pada
membran nitroselulosa (Amersham Hybond-P PVDF membrane, GE Health Care, UK)
selama 1 menit. Sebelumnya membran diperlakukan dengan dicelup pada metanol absolut
selama 10 detik, kemudian dicuci masing-masing 5 menit di dalam akuabides dan
phosphate buffer saline (PBS) (137 mM NaCl, 1,5 mM KH2PO4, 8,0 mM Na2HPO4, 2,7
mM KCl, 3,0 mM NaN3, pH 7,4) dan dikeringanginkan di atas kertas saring yang telah
dicelup ke dalam PBS. Membran yang telah diblot ditempatkan dalam wadah plastik dan
diblocking dengan susu skim 2% (w/v) yang dilarutkan dalam PBS selama 60 menit pada
suhu ruang. Membran diinkubasikan dengan antiserum pada suhu ruang selama 1-2 jam,
atau pada suhu 4°C semalam. Antiserum diencerkan dalam PBS dengan seri pengenceran
sesuai perlakuan. Membran dicuci dengan PBST selama 10 menit sebanyak 3 kali pada
suhu ruang. Membran kemudian diinkubasikan dengan konjugat anti guinea pig-IgG
(Sigma Chemical Co. St. Louise, USA) pada pengenceran 1:1000 dalam PBS selama 2-3
jam pada suhu ruang. Membran dicuci sebagaimana di atas, dan diwarnai dengan substrat
5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium (BCIP/NBT) (Sigma
Chemical Co., USA) dalam bufer alkaline phosphate (AP) (0,1 M Tris Base, 0,1 M NaCl,
5,0 mM MgCl2.6H2O, pH 9,5). Reaksi positif ditunjukkan dengan perubahan warna ungu
pada tissue blot. Reaksi pewarnaan dihentikan dengan mencuci membran dengan
akuabides dan dikeringanginkan.

15
V. HASIL DAN PEMBAHASAN
5.1 Karakteristik Virus Murni
Pemurnian CVB menggunakan metode pemisahan virus dengan cara
ultrasentrifugasi dengan gradien kepekatan sesium sulfat berhasil memperoleh zona virus
yang membentuk suatu pita yang berbatas tegas yang mencerminkan lokasi virus berada
(Gambar 1).
Spektrofotometri terhadap suspensi virus murni pada panjang gelombang 260 nm
(A260) dan 280 nm (A280) menghasilkan nilai absorbansi berturut-turut 1,449 dan 1,183
(Gambar 2). Tingkat kemurnian virus dapat dihitung dari nisbah A260/A280 yaitu 1,22. Nilai
nisbah ini menunjukkan suspensi virus murni hasil purifikasi pada penelitan ini
mempunyai tingkat kemurnian yang cukup tinggi. Dijkstra & de Jager (1998) menyatakan
bahwa virus dengan bentuk partikel memanjang memiliki nilai A260/A280 sekitar 1,2;
sedangkan virus dengan partikel berbentuk isometrik memiliki nilai A260/A280 sekitar 1,7.
Gambar 1. Hasil pemurnian virus dengan pemisahan virus melalui ultrasenrifugasi dengan
gradien kepekatan sesium sulfat. Zona virus murni membentuk pita yang
berbatas tegas.
Berdasarkan asumsi bahwa extinction coefficient (E%1.0
260.1 nmcm ) carlavirus adalah 2,3
(Koenig, 1982), maka konsentrasi virus pada suspensi virus murni tersebut adalah 0,630
mg/ml. Dengan demikian total virus yang dihasilkan dari 200 g daun segar N.
benthamiana adalah 6,250 mg.
zona virus murni

16
Gambar 2. Profil spektrofotometri siapan virus murni
Hasil pemurnian CVB telah dilaporkan oleh beberapa peneliti sebelumnya. Suastika
et al. (1997) memperoleh nilai A260/A280 sebesar 1,3 dan 12 mg virus murni per 100 g daun
segar ketika melakukan pemurnian CVB menggunakan N. clevelandii sebagai sumber
virus dan metode sentrifugasi gradien sukrose. Sedangkan Hollings & Stone (1972)
melaporkan bahwa nilai A260/A280 CVB adalah 1,55.
Pemurnian virus dari kelompok carlavirus yang lain dilaporkan dari penelitian Van
Lent et al. (1980) yang memurnikan Elderberry carlavirus (ECV). Purifikasi dilakukan
dengan metode klarifikasi melalui presipitasi dengan garam, konsentrasi melalui presipitasi
dengan PEG dan disentrifugasi dengan gradien sukrose didapatkan bahwa virus murni
memiliki nilai A260/A280 = 1,17 dan dari 100 g daun Sambucus segar diperoleh 2-3 mg
virus. Sharma et al. (2005) yang melakukan purifikasi terhadap LSV menggunakan gradien
sesium sulfat memperloleh virus murni dengan nilai A260/A280 = 1,4.
Tingkat kemurnian virus yang dihasilkan dari penelitian ini relatif tinggi yaitu di atas
rata-rata untuk virus yang partikelnya berbentuk memanjang yang memiliki rata-rata nilai
A260/A280 sekitar 1,2 (Dijkstra & de Jager, 1998). Keberhasilan pemurnian tersebut
mungkin disebabkan oleh 2 faktor, yaitu metode pemurnian dan tanaman inang sumber
virus yang digunakan. Metode pemurnian yang digunakan sesuai dengan prosedur dari
Foster (1998), yaitu meliputi ekstraksi daun (memisahkan molekul makro tanaman)
dengan kloroform yang ditambahkan enzim untuk maserasi jaringan (driselase), pencegah
oksidasi (β-2 mercaptoehtanol), agen kelasi (EDTA) dan detergen pemecah membran sel
(Triton X-100); klarifikasi ekstrak daun dengan sentrifugasi 5.000 g selama 20 menit;
mengkonsentrasikan virus dan memisahkan dari material tumbuhan yang bermolekul kecil
dengan PEG dan sentrifugasi pada 10.000 g selama 15 menit; sedimentasi dengan
1,449
1,183
17
sentrifugasi kecepatan tinggi (30.000 g selama 90 menit); dan pemurnian dengan
sentrifugasi pada cesium chloride, sentrifugasi pada 110.000 g selama 20 jam. Metode
tersebut relatif lebih sederhana dan mudah dilakukan sehingga memperkecil terjadinya
proses oksidasi. Terjadinya proses oksidasi sering menjadi kendala dalam setiap proses
pemurnian virus tumbuhan, karena proses oksidasi mengakibatkan degradasi virus.
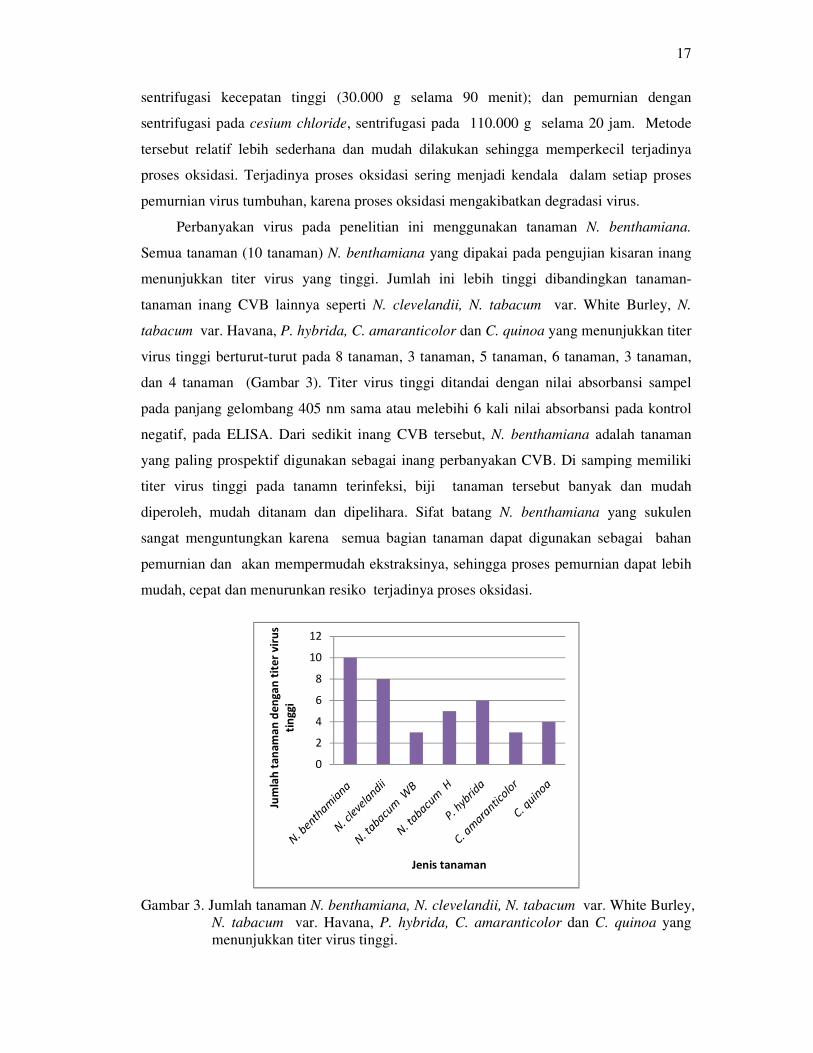
Perbanyakan virus pada penelitian ini menggunakan tanaman N. benthamiana.
Semua tanaman (10 tanaman) N. benthamiana yang dipakai pada pengujian kisaran inang
menunjukkan titer virus yang tinggi. Jumlah ini lebih tinggi dibandingkan tanaman-
tanaman inang CVB lainnya seperti N. clevelandii, N. tabacum var. White Burley, N.
tabacum var. Havana, P. hybrida, C. amaranticolor dan C. quinoa yang menunjukkan titer
virus tinggi berturut-turut pada 8 tanaman, 3 tanaman, 5 tanaman, 6 tanaman, 3 tanaman,
dan 4 tanaman (Gambar 3). Titer virus tinggi ditandai dengan nilai absorbansi sampel
pada panjang gelombang 405 nm sama atau melebihi 6 kali nilai absorbansi pada kontrol
negatif, pada ELISA. Dari sedikit inang CVB tersebut, N. benthamiana adalah tanaman
yang paling prospektif digunakan sebagai inang perbanyakan CVB. Di samping memiliki
titer virus tinggi pada tanamn terinfeksi, biji tanaman tersebut banyak dan mudah
diperoleh, mudah ditanam dan dipelihara. Sifat batang N. benthamiana yang sukulen
sangat menguntungkan karena semua bagian tanaman dapat digunakan sebagai bahan
pemurnian dan akan mempermudah ekstraksinya, sehingga proses pemurnian dapat lebih
mudah, cepat dan menurunkan resiko terjadinya proses oksidasi.
Gambar 3. Jumlah tanaman N. benthamiana, N. clevelandii, N. tabacum var. White Burley,
N. tabacum var. Havana, P. hybrida, C. amaranticolor dan C. quinoa yang
menunjukkan titer virus tinggi.
0
2
4
6
8
10
12
Jum
lah
ta
na
ma
n d
en
ga
n t
ite
r v
iru
s
tin
gg
i
Jenis tanaman

18
5.2 Morfologi dan Ukuran Partikel Virus
Hasil pengamatan dengan mikroskop elektron terhadap siapan virus murni
menunjukkan adanya partikel virus yang berbentuk batang agak lurus dan lentur dengan
ukuran panjang 685 nm dan lebar 12 nm (Gambar 4).
Gambar 4. Bentuk partikel CVB pada mikroskop elektron (pembesaran 40.000 kali).
A : siapan perasan tanaman terifeksi CVB. B : siapan virus murni.
Bentuk dan ukuran partikel virus ini sesuai dengan yang dilaporkan oleh peneliti
terdahulu. Hollings & Stone (1972) mendapatkan bentuk dan ukuran partikel CVB yang
sama dengan hasil pengamatan pada penelitian ini. Suastika et al. (1997) pada pengamatan
siapan murni CVB menemukan partikel virus berbentuk panjang agak lentur dengan
ukuran panjang 650-700 nm dan lebar 13 nm; dan pengamatan Verma et al. (2003) juga
mendapatkan bentuk partikel lentur dengan ukuran 680x12 nm.
5.3 Ukuran Protein Selubung Virus
Ukuran protein selubung ditentukan dengan SDS – PAGE dan Western blot.
Analisis SDS-PAGE menunjukkan adanya pita yang sama pada sampel daun N.
benthamiana terinfeksi CVB dan virus murni yaitu pita protein berukuran sekitar 34 kDa
(Gambar 5A), dan pita tersebut tidak nampak pada sampel daun N. benthamiana sehat. Hal
ini menunjukkan bahwa pita tersebut kemungkinan merupakan protein selubung dari virus.
Konfirmasi dengan analisis Western blot terhadap protein selubung virus menggunakan
antiserum CVB menunjukkan bahwa pita protein berukuran 34 kDa bereaksi positif
dengan antiserum CVB (Gambar 5B). Ini berarti bahwa memang benar pita dengan berat
molekul 34 kDa tersebut adalah protein selubung CVB. Hasil pengamatan morfologi virus
A B 200 nm 200 nm

19
dan ukuran protein selubung mendukung peneliti sebelumnya yang menemukan bentuk
dan ukuran partikel serta berat molekul protein selubung CVB yang sama (Suastika et al.
1997; Hollings & Stone,1972). Hull (2002) dan Lee et al. (2003) juga menyatakan bahwa
virus anggota kelompok carlavirus memiliki protein selubung tunggal dengan berat
molekul 31-36 kDa.
Gambar 5. Hasil analisis SDS-PAGE (A) dan Western blot (B) protein selubung CVB
isolat Indonesia. M : Marker. H : daun N. benthamiana sehat. I : daun N.
benthamiana terinfeksi CVB. C : virus murni.
5.4 Produksi Antiserum CVB
Konsentrasi antiserum. Antiserum diproduksi dengan melakukan imunisasi
terhadap marmut yaitu dengan menginjeksikan siapan virus murni CVB ke dalam tubuh
marmut. Masing-masing marmut menghasilkan antiserum rata-rata 10,75 ml. Nilai
absorbansi dari antiserum tidak diserap, antiserum yang sudah diserap menggunakan cairan
perasan N. benthamiana sehat serta antiserum murni (gamma globulin) yang dihasilkan
disajikan pada Gambar 6.
Absorbansi pada panjang gelombang 280 dari ketiga jenis antiserum diukur
menggunakan spektrofotometer untuk penentuan konsentrasi proteinnya. Setelah masing-
masing diencerkan sepuluh kali, untuk antiserum kasar, antiserum yang diserap dan
antiserum murni mempunyai nilai A 280 berturut-turut 1,569, 0,938 dan 0,667.
Berdasarkan asumsi bahwa nilai absorbansi 1,4 pada A 280 setara dengan konsentrasi 1
mg/ml, maka dapat dihitung konsentrasi antiserum kasar, antiserum yang diserap dan
antiserum murni berturut-turut 1,121 mg/ml, 0,670 mg/ml dan 0,476 mg/ml.
A B
34 kDa
34 kDa

20
Gambar 6. Nilai absorbansi antiserum pada panjang gelombang 280.
A : antiserum tidak diserap; B : antiserum yang diserap menggunakan cairan
perasan N. benthamiana sehat ; C : antiserum murni (gamma globulin)
A
B
C
1,569
0,938
0,667

21
Konsentrasi protein antiserum murni yang dipresipitasi menggunakan amonium
sulfat lebih kecil dibandingkan dengan antiserum yang tidak diserap maupun yang diserap.
Namun demikian, reaktivitasnya terhadap antigen paling tinggi dibandingkan dengan
antiserum lainnya. Menggunakan antiserum murni yang sudah dipisahkan dari protein lain
yang terdapat dalam suatu antiserum dapat meningkatkan akurasi dan spesifitasnya dalam
uji serologi. Gamma globulin murni juga dapat dilabel (konjugasi) dengan enzim tertentu
untuk pengujian secara DAS-ELISA, atau dilabel dengan unsur radioaktif untuk pengujian
dengan radio immuno assay.
Titer antiserum. Titer antiserum ditentukan dengan metode ELISA yaitu melalui
pengujian beberapa perlakuan pengenceran antiserum (100, 500, 1000, 5000, 10.000,
50.000, 100.000, 500.000 dan 1.000.000 kali). Hasil yang diperoleh menunjukkan bahwa
antiserum yang tidak diserap maupun yang diserap mempunyai titer 1/50.000, sedangkan
antiserum murni mempunyai titer 1/100.000 (Gambar 7).
Titer antiserum untuk serum tidak diserap dan diserap yang diproduksi pada
penelitian ini sedikit lebih tinggi dibandingkan dengan kelompok virus lain. Penelitian
Sulandari et al. (2004) pada produksi antiserum geminivirus mendapatkan bahwa
antiserum yang tidak diserap maupun yang diserap mempunyai titer 1/10.000, dan gamma
globulin mempunyai titer 1/100.000; sedangkan produksi antiserum geminivirus yang
dilakukan Haryadi (2006) menemukan titer antiserum yaitu 1/16.384. Titer antiserum
antara lain dipengaruhi oleh jenis virus dan hewan percobaan yang dipakai dalam produksi
antiserum. Makin besar berat molekul antigen, makin efektif berperan sebagai zat
imunogen untuk menstimulasi antiserum (Hull, 2002). Kemungkinan CVB yang memiliki
berat molekul protein selubung 34 kDa lebih imunogenik dibandingkan dengan
geminivirus pada penelitian Sulandari et al. (2004) yang memiliki berat molekul protein
selubung 29 kDa, sehingga CVB menstimulasi antiserum dengan titer yang relatif lebih
tinggi. Di samping itu, marmut yang dipakai sebagai hewan percobaan dalam produksi
antiserum ini menghasilkan antiserum berkualitas baik. Menurut Rollin & Kesel (1995),
marmut adalah sumber komplemen serum yang sangat baik; dan sering menjadi hewan
pilihan dalam produksi antiserum karena hanya memerlukan sedikit antigen untuk
imunisasi, volume serumnya relatif banyak, mudah pemeliharaannya, dan menghasilkan
antiserum yang baik kualitasnya.

22
Gambar 7. Titer antiserum tidak diserap (As K), antserum diserap (As S) dan antiserum
murni (As M) berdasarkan ELISA. I : sampel tanaman terinfeksi CVB.
S : sampel tanaman sehat
Reaktivitas ketiga siapan antiserum terhadap antigen sangat kuat dan dapat
dibedakan dengan jelas antara sampel tanaman sakit dengan tanaman yang sehat. Sampel
tanaman sakit yang berasal dari Cianjur, Medan, Malang dan Bali dapat dideteksi dengan
baik menggunakan tiga macam antiserum (Gambar 8). Antiserum yang sudah dimurnikan
(gamma-globulin) ternyata reaktivitasnya paling baik dibandingkan dengan antiserum tidak
diserap ataupun yang sudah diserap. Nisbah nilai absorbansi pada sampel tanaman krisan
sakit dibandingkan dengan sampel sehat lebih tinggi pada sampel menggunakan antiserum
murni dibandingkan dengan antiserum kasar dan yang diserap.
Spesifitas dan akurasi uji serologi sangat dipengaruhi oleh tingkat kemurnian
antiserum yang digunakan. Gamma-globulin merupakan protein spesifik yang sudah
dipisahkan dari protein lain yang terdapat dalam suatu antiserum. Protein tersebut akan
bereaksi dengan antigen tertentu sehingga spesifitas dan akurasinya lebih tinggi
dibandingkan antiserum kasar ataupun yang sudah diserap yang keduanya masih tercampur
dengan protein lain.
0
0.2
0.4
0.6
0.8
1
1.2
100
500
1000
5000
1000
0
5000
0
1000
00
5000
00
1000
000
Pengenceran antiserum
Nilai ab
so
rban
si p
ad
a A
405 n
m
I-As K
I-As S
I-As M
S-As K
S-As-S
S-As M

23
Gambar 8. Nilai absorbansi antiserum tidak diserap, diserap dan antiserum murni pada uji
ELISA terhadap isolat CVB dari berbagai lokasi (Cianjur, Medan, Malang dan
Bali)
5.5 Spesifisitas Antiserum
Antiserum yang diproduksi pada penelitian ini juga diuji untuk analisis Western
blot dan Immunosorbent electron microscopy (ISEM), untuk melihat spesifisitas reaksi
antiserum dengan protein selubung CVB. Menggunakan antiserum yang sudah dimurnikan
dapat mendeteksi adanya pita dengan ukuran sekitar 34 kDa (Gambar 9) sesuai dengan
ukuran protein selubung yang didapatkan pada uji SDS-PAGE. Antiserum dengan
pengenceran 1:1000 mampu mengkonfirmasi protein selubung CVB pada uji ini.
Protein 34 kDa yang terdeteksi pada analisis Western blot sesuai dengan berat
molekul protein selubung CVB yang digunakan untuk imunisasi pada hewan percobaan.
Hal ini membuktikan bahwa antiserum yang dihasilkan berasal dari virus murni yang
digunakan sebagai imunogen dan bukan dari antigen lain. Analisis Western blot juga
membuktikan tidak adanya background atau reaksi positif antiserum terhadap komponen
tanaman. Ini dapat dilihat dari bersihnya lajur sampel daun tanaman N. benthamiana sehat.
0
0.2
0.4
0.6
0.8
1
1.2
Tidak diserap Diserap Murni
Jenis antiserum
Nil
ai
ab
so
rban
si
pad
a A
405 n
m
Cianjur
Medan
Malang
Bali
Sehat
Bufer

24
Gambar 9. Hasil analisis western blot protein selubung CVB menggunakan antiserum
yang diproduksi. M : Marker. H : daun N. benthamiana sehat. I : daun N.
benthamiana terinfeksi CVB. C : virus murni.
Pengamatan partikel CVB pada mikroskop elektron dengan metode ISEM
menggunakan antiserum yang diproduksi pada penelitian ini memberikan hasil dekorasi
yang baik. Partikel virus berbentuk panjang agak lentur nampak lebih jelas pada grid yang
sebelumnya dicelupkan pada suspensi antiserum dibandingkan dengan siapan murni tanpa
antiserum (Gambar 10). Hal ini menunjukkan bahwa antiserum yang dihasilkan
memberikan reaksi yang spesifik terhadap protein selubung virus.
Gambar 10. Partikel CVB pada pengamatan dengan mikroskop elektron (pembesaran
40.000 kali). Partikel virus berbentuk panjang agak lentur nampak lebih jelas
pada siapan virus murni didekorasi dengan antiserum (A), dibandingkan
dengan siapan virus murni tanpa antiserum (B)
A B 200 nm 200 nm
34 kDa

25
5.6 Pengujian Metode Deteksi Serologi
Melihat reaktivitas antiserum pada deteksi serologi, dilakukan pengujian dua
metode deteksi yaitu I-ELISA dan TBIA. Pada kedua metode ini dibuat pengenceran
antiserum 100, 500, 1000, 5000, 10.000, 50.000, 100.000, 500.000 dan 1.000.000 kali dan
pada ELISA juga dibuat pengenceran cairan perasan tanaman sakit : bufer (b/v) yaitu ½, ¼,
1/8, 1/16, 1/32, 1/64, 1/128, 1/256 dan 1/512.
Uji serologi dengan ELISA menunjukkan bahwa semakin tinggi pengenceran
antiserum semakin lemah reaksi yang terjadi, atau semakin kecil nilai nisbah absorbansi
pada sampel dengan nilai absorbansi pada kontrol negatif. Demikian pula semakin tinggi
pengenceran sampel menunjukkan reaksi yang semakin lemah. Sensitifitas ELISA dalam
mendeteksi CVB cukup tinggi dan virus masih terdeteksi pada pengenceran cairan perasan
tanaman terinfeksi 1/256. Pengenceran sampel cairan perasan tanaman terinfeksi 1/16
menunjukkan reaksi yang kuat sampai pada pengenceran antiserum 1000 kali dan masih
menunjukkan reaksi positif sampai pada pengenceran antiserum 100.000 (Tabel 1).
Tabel 1. Reaksi antiserum terhadap cairan perasan tanaman terinfeksi CVB pada berbagai
seri pengenceran melalui metode I-ELISA
Pengenceran antiserum
Reaksi antiserum terhadap sampel
Bufer Tanaman Sehat*
Pengenceran cairan perasan tanaman terinfeksi CVB
1/2 1/4 1/8 1/16 1/32 1/64 1/128 1/256 1/512
100 - - +++ +++ +++ +++ +++ +++ ++ + -
500 - - +++ +++ +++ +++ +++ ++ ++ + -
1000 - - +++ +++ +++ +++ ++ + + + -
5000 - - +++ +++ +++ ++ + + + + -
10.000 - - +++ +++ ++ + - + - - -
50.000 - - ++ ++ ++ + - - - - -
100.000 - - ++ ++ + + - - - - -
500.000 - - + + + - - - - - -
1.000.000 - - - - - - - - - - -
Keterangan : Reaksi positif jika nilai absorbansi (405 nm) 3 kali nilai kontrol negatif
(Verma et al. 2003).
* Tanaman krisan sehat sebagai kontrol negatif
+++ Reaksi kuat bila nilai absorbansi > 6 kali nilai kontrol negatif
++ Reaksi sedang bila nilai absorbansi (4,5< ⊕ <6) kali nilai
kontrol negatif
+ Reaksi lemah bila nilai absorbansi < 4,5 atau > 3 kali nilai
kontrol negatif
- Reaksi negatif

26
Uji laboratorium menunjukkan bahwa TBIA terbukti cukup sensitif untuk
mendeteksi keberadaan CVB pada tanaman krisan. Reaksi positif pada pengujian TBIA,
yang ditunjukkan dengan perubahan warna ungu pada tissue blot setelah pemberian
substrat BCIP/NBT, masih nampak pada pengenceran antiserum 10.000 kali (Gambar 11).
Sampel tanaman sehat (kontrol) bersih, menandakan kespesifikan antiserum terhadap CVB.
Melalui I-ELISA dan TBIA diketahui bahwa antiserum yang dihasilkan dapat
mengidentifikasi CVB yang menginfeksi krisan dari berbagai lokasi (Cianjur, Medan,
Malang dan Bali). Reaktivitas antiserum ternyata sama terhadap sampel yang berasal dari
berbagai lokasi yang berbeda. Reaktivitas yang sama pada isolat yang berbeda tersebut
disebabkan keempat isolat memiliki hubungan yang sangat dekat dengan susunan asam
amino protein selubung yang memiliki tngkat homologi tinggi (95-99%) (Temaja, 2008).
Di samping itu antiserum yang digunakan dalam pengujian adalah antiserum poliklonal
sehingga mampu mengenali berbagai epitop dari protein selubung virus. Jenis epitop yang
ada sangat dipengaruhi oleh jenis protein yang tersusun dari beberapa macam asam
amino. Gen penyandi protein selubung dari genus carlavirus diketahui mempunyai
runutan susunan DNA dengan derajat kesamaan yang tinggi (conserved) antar anggotanya.
Gambar 11. Reaktivitas antiserum pada pengenceran 1/100, 1/500, 1/1000, 1/5000 dan
1/10.000 terhadap isolat CVB dari berbagai lokasi pada uji serologi dengan
TBIA. S : sampel terinfeksi; K : kontrol negatif
Uji serologi secara TBIA lebih menguntungkan dibandingkan dengan teknik
ELISA. Metode TBIA lebih mudah dan cepat, biayanya lebih murah karena hanya
diperlukan reagen yang lebih sedikit dan dapat menguji sampel lebih banyak. Pada
selembar membran nitroselulosa yang berukuran 7 cm x 4 cm dapat menampung 96
Cianjur
Medan
Malang
Bali
1/100 1/500 1/1.000 1/5.000 1/10.000
S K S K S K S K S K

27
sampel, dengan jarak antar sampel sekitar 5 mm (Gambar 12). Hal ini penting untuk
deteksi massal terhadap bahan tanaman krisan terinfeksi CVB, terutama pada perusahaan
eksportir bahan tanaman krisan dan Karantina Tumbuhan yang memerlukan pengujian
sampel dalam jumlah besar. Pengujian secara massal sampel dapat mengurangi
pengeluaran untuk pengujian karena lebih efisien memanfaatkan reagen dan di samping itu
dapat dikerjakan lebih cepat.
Metode ini juga memungkinkan penyiapan (blotting) sampel dilakukan di lapangan,
kemudian membran disimpan untuk proses selanjutnya di laboratorium. Penelitian ini
mencoba mengerjakan blotting sampel di laboratorium dan di lapangan. Sampel dari
Cianjur diblot di laboratorium (Gambar 12), sedangkan sampel Bali diblot di tempat
pengambilan sampel (Gambar 12). Proses TBIA selanjutnya dilakukan setelah 7 hari.
Kedua cara tersebut memberikan pengaruh yang tidak berbeda pada hasil TBIA.
Gambar 12. Deteksi serologis CVB pada tanaman krisan dengan teknik TBIA dengan
pengenceran antiserum 1/10.000. Kiri : sampel dari Cianjur. Kanan : sampel
dari Bali. 1a dan 1b : kontrol negatif. 1c dan 1d : kontrol positif.
A B
1
2
3
4
5
6
7
8
9
10
11
12
a b c d e f g h a b c d e f
Cianjur Bali
4 cm 3 cm
7 cm

28
VI. KESIMPULAN
1. N. benthamiana adalah tanaman yang paling prospektif digunakan sebagai inang
perbanyakan CVB. Total virus yang dihasilkan dari 200 g daun segar N. benthamiana
adalah 6,250 mg.
2. Imunisasi marmut menggunakan 150 µg CVB murni dapat menghasilkan antiserum
yang cukup baik kualitasnya. Antiserum yang dihasilkan rata-rata 10,75 ml/ekor.
3. Dengan metode ELISA didapatkan bahwa titer antiserum adalah 1/100.000.
4. Reaktivitas antiserum terhadap antigen sangat kuat dan dapat dibedakan dengan jelas
antara sampel tanaman sakit dengan tanaman yang sehat.
5. TBIA merupakan metode deteksi CVB pada tanaman krisan yang memenuhi
persyaratan sensitifitas, massal dan ekonomis.

29
DAFTAR PUSTAKA
Abouzid AM, Freitas-Astua J, Purcifull DE, Polston JE, Beckham KA, Crawford WE,
Petersen MA, Peyser B, Patte C, Hiebert E. 2002. Serological studies using
polyclonal antisera prepared against the viral coat protein of four Begomovirus
expressed in Escherichia coli. Plant Dis 86:1109-1114.
Agrios GN. 1997. Plant Pathology. 4 th ed. California : Academic Press, Inc.
Attathom S, Chiemsombat P, Sutabutra T, and Pongpanitanond R. 1990. Characterization
of nucleic acid of Tomato yellow leaf curl virus. Kasetsart J. Nat. Sci. 24:1-5.
Badge J, Brunt A, Carson R, Dagless E, Karamagioli M, Phillips S, Seal S, Turner R,
Foster G D. 1996. A carlavirus-specitic PCR primer and partial rnicleotidc sequence
provides further evidence for the recognition of cowpea mild mottle virus as a
whitefly-transmitted carlavirus. Eur. J. Plant Pathol. 102: 305-310.
Chen J, Chen JP, Adam MJ. 2002. Characterization of some carla- and potyvirus from bulb
in China. Arch. Virol. 147:419-428.
Choi SH, Ryu KH. 2003. The complete nucleotide sequence of the genom RNA Lily
symtompless virus and its compariron with that of the other carlaviruses. Arch.
Virol. 148:1943-1955.
Clark MF, Adams AN, 1977. Characteristics of the microplate method of enzyme-linked
immunosorbent assay for the detection of plant viruses. J. Virol. Methods 34,
475-483.
Cohen, J., Zeidan, M., Rosner, A., Gera, A. 2000. Biological and molecular characterization
of a new carlavirus isolated from an Aconitum sp. Phytopathology 90:340-344.
Dijkstra J, de Jager CP. 1998. Pratical Plant Virology. Protocol and Exercises. New
York: Springer-Verlag Berlin Heidelberg.
Foster, G. D. 1992. The structure and expression of the genome of Carlaviruses. Res.
Virol. 143: 103-112.
Foster GD. 1998. Carlavirus isolation and RNA extraction. In: Foster GD, Taylor SC.
(Eds.), Plant Virology Protocols, from Virus Isolation to Transgenic Resistance.
Totowa : Humans Press. pp. 145-150.
Foster GD, Mills PR. 1992. The 3'-Nucleotide Sequence of an Ordinary Strain of Potato
Virus S. Virus Gene 6(3): 213-220.
Fuji S, Yamamoto H, Inoue M, Yamashita K, Fukui Y, Furuya H, Naito H. 2002. Complete
nucleotide sequence of the genomic RNA of Aconitum latent virus (genus
Carlavirus) isolated from Delphinium sp. Arch Virol 147: 865-870.
Hakkaart, F.A., Maat, D.Z., 1974. Variation of chrysanthemum virus B. Netherland J.
Plant Pathol. 80, 97-103.

30
Harlow, Lane D. 1999. Using Antibodies. A Laboratorium Manual. New York: Cold
Springer Harbor Laboratory Press.
Haryadi D. 2006. Produksi antibodi poliklonal geminivirus penyebab penyakit daun
keriting kuning cabai,dan kajian serologinya. [tesis]. Sekolah Pascasarjana IPB.
Hataya T, Uchino K, Arimoto R, Suda N, Sano T, Shikata E, Uyeda I. 2000. Molecular
characterization of Hop latent virus and phylogenetic relationships among viruses
closely related to carlaviruses. Arch Virol. 145: 2503–2524.
Hollings, M. 1957. Investigation of chrysanthemum viruses. II. Virus B (mild mosaic) and
chrysanthemum latent virus. Ann. Appl. Biol. 45:589-602.
Hollings, M., Stone OM. 1972. Chrysanthemum virus B. CMI/AAB Description of Plant
Viruses No. 110.
Hull R. 2002. Matthews’ Plant Virology. Fourth Ed. San Diego : Academic Press.
Koenig R. 1982. Carlavirus group. CMI/AAB. Descriptions of Plant Viruses No. 259.
Lawrence D.M., M.N. Rozanov, B.I. Hillman. 1995. Autocatalytic processing of the 223
kDa protein of blueberry scorch carlavirus by papain-like proteinase. Virology 207:
127-135.
Lee BY, Choi SH, Ryu KH. 2003. Characterization of the 3’-terminal nucleotide sequence
of two Korean isolates of Daphne virus S support its placement as a distinct species
of the genus Carlavirus. Arch. Virol. 148:1915-1924.
Mahmood T, Hein GL, French RC. 1997. Development of serological procedures for
rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite.
Plant Dis 81:250-253.
Moran, J.R. 1987. Chrysanthemum B carlavirus. Cite this publication as : Brunt, A.A.,
Crabtree, K., Dallwitz, M.J., Watson, L. and Zurcher, E.J. (eds) (1996 onwards).
'Plant Viruses Online Descriptions and Lists from the VIDE Database. Version :
20th
August 1996.
Ram R, Verma N, Singh AK, Singh L, Hallan V, Zaidi AA. 2005. Indexing and production
of virus-free chrysanthemums. Biologia Plantarum 49(1):149-152.
Sharma A, Mahinghara BK, Singh AK, Kulshretha S, Raikhy G, Singh L, Verma N,
Hallan V, Ram R, Zaidi AA. 2005. Identification, detection and frequency of lily
viruses in Northern India. Scientia Horticulturae 106: 213-227.
Somowiyarjo S, Sako N, Nonaka F. 1989. Dot-immunobinding assay for Zucchini yellow
mosaic virus using polyclonal and monoclonal antibodies. Ann Phytopathol Soc
55:56-63.

31
Suastika G, Kurihara J, Natsuaki KT, Tomaru K. 1997. A strain of Chrysanthemum B
carlavirus causing flower colour breaking on Gymnaster savatieri (Makino)
Kitamura. Ann. Phytopathol. Soc. Jpn. 63:1-7.
Sudarshana MR, Wang HL, Lucas WJ, Gilbertson RL. 1997. Dynamics of Bean -
dwarf mosaic geminivirus cell-to-cell and long distance movement in Phaseolus
vulgaris revealed, using the green fluorescent protein. MPMI 4:277-291.
Sulandari S, Suseno R, Hidayat SH, Sosromarsono S, Harjosudarmo J. 2004.
Pembuatan antiserum dan kajian serologi virus penyebab penyakit daun
keriting kuning cabai. J Perlind Tan Ind 10(1):42-52.
Van Lent JWM, Wit AJ, Dijkstra J. 1980. Characterization of carlavirus in elderberry
(Sambucus spp.). Neth. J. Plant Pathol. 86:117-134.
Verma N, Sharma A, Ram R, Hallan V, Zaidi AA, Garg ID. 2003. Detection, identification
and incidence of Chrysanthemum B carlavirus in chrysanthemum in India. Crop
Protect. 22:415-429.
Zavriev SK, Kanyuka KV, Levay KE. 1991. The genome organization of potato virus
M RNA. J Gen Virol 72: 9-14.




















