
Basal and Fasting/Refeeding-regulated Tissue Levels of Endogenous PPAR-α Ligands in Zucker Rats
8
OBESITY | VOLUME 18 NUMBER 1 | JANUARY 2010 55 nature publishing group ARTICLES INTEGRATIVE PHYSIOLOGY INTRODUCTION e peroxisome proliferator–activated receptor-α (PPAR-α) is a nuclear receptor deeply involved in the control of metabolism and its dysfunctions (1). PPAR-α activation by various ligands, including derivatives of fatty acids and fibrates, upregulates the expression of several catabolic enzymes that are involved in mitochondrial and peroxisomal β-oxidation and micro- somal ω-oxidation, as well as in the transcriptional regulation of genes that are necessary for the maintenance of the redox balance during the oxidative catabolism of fatty acids (2,3). WY14,643, a well-known specific PPAR-α agonist, increases fatty acid oxidation by increasing the expressions of peroxi- somal and mitochondrial fatty acid β-oxidation enzymes (4). Accordingly, PPAR-α agonists are used in the treatment of hypertriglyceridemia (1). WY14,643 also reduces liver insulin resistance more efficiently than muscle insulin resistance and normalizes the circulating triglyceride levels and blood glu- cose levels in lipoatrophic mice (4,5). Finally, PPAR-α agonists have anti-inflammatory effects in vascular cells and macro- phages (6,7), which, together with their antihyperlipidemic effects, might explain why they inhibit atherosclerotic plaque formation (7,8). Among the several endogenous ligands proposed so far for PPAR-α, N-palmitoylethanolamine (PEA) and, particularly, N-oleoylethanolamine (OEA) are among the most potent with affinity constants estimated in the range of ~3 μmol/l and ~120 nmol/l, respectively (9,10). e PPAR-α-mediated effects of OEA have been widely investigated (see refs. 11,12 for reviews) and include: (i) inhibition of food intake via action on sensory neurons originating in the duodenum and terminating in the brainstem (13) or through fatty acid translocase/CD36-mediated stimulation of fatty acid uptake by enterocytes (14,15); (ii) reduction of body weight gain and triacylglycerol content in the liver and adipose tissue of diet- induced obese rats and wild-type (WT) mice, but not in obese PPAR-α −/− mice (16); (iii) stimulation of lipolysis and fatty acid Basal and Fasting/Refeeding-regulated Tissue Levels of Endogenous PPAR-α Ligands in Zucker Rats Angelo A. Izzo 1 , Fabiana Piscitelli 2 , Raffaele Capasso 1 , Pietro Marini 2 , Luigia Cristino 3 , Stefania Petrosino 2 and Vincenzo Di Marzo 2 N-oleoylethanolamine (OEA) and N-palmitoylethanolamine (PEA) are endogenous lipids that activate peroxisome proliferator–activated receptor-α with high and intermediate potency, and exert anorectic and anti-inflammatory actions in rats, respectively. We investigated OEA and PEA tissue level regulation by the nutritional status in lean and obese rats. OEA and PEA levels in the brainstem, duodenum, liver, pancreas, and visceral (VAT) or subcutaneous (SAT) adipose tissues of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum or following overnight food deprivation, with and without refeeding, were measured by liquid chromatography–mass spectrometry. In WT rats, duodenal OEA, but not PEA, levels were reduced by food deprivation and restored by refeeding, whereas the opposite was observed for OEA in the pancreas, and for both mediators in the liver and SAT. In ad lib fed Zucker rats, PEA and OEA levels were up to tenfold higher in the duodenum, slightly higher in the brainstem, and lower in the other tissues. Fasting/refeeding-induced changes in OEA levels were maintained in the duodenum, liver, and SAT, and lost in the pancreas, whereas fasting upregulated this compound also in the VAT. The observed changes in OEA levels in WT rats are relevant to the actions of this mediator on satiety, hepatic and adipocyte metabolism, and insulin release. OEA dysregulation in Zucker rats might counteract hyperphagia in the duodenum, but contribute to hyperinsulinemia in the pancreas, and to fat accumulation in adipose tissues and liver. Changes in PEA levels might be relevant to the inflammatory state of Zucker rats. Obesity (2009) 18, 55–62. doi:10.1038/oby.2009.186 1 Endocannabinoid Research Group, Department of Experimental Pharmacology, University of Naples “Federico II”, Naples, Italy; 2 Endocannabinoid Research Group, Institute of Biomolecular Chemistry, Consiglio Nazionale delle Ricerche, Pozzuoli, Italy; 3 Endocannabinoid Research Group, Institute of Cybernetics, Consiglio Nazionale delle Ricerche, Pozzuoli, Italy. Correspondence: Vincenzo Di Marzo ([email protected]) Received 30 December 2008; accepted 5 May 2009; published online 11 June 2009. doi:10.1038/oby.2009.186
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Basal and Fasting/Refeeding-regulated Tissue Levels of Endogenous PPAR-α Ligands in Zucker Rats

obesity | VOLUME 18 NUMBER 1 | jaNUaRy 2010 55
nature publishing group articlesintegrative Physiology
IntroductIonThe peroxisome proliferator–activated receptor-α (PPAR-α) is a nuclear receptor deeply involved in the control of metabolism and its dysfunctions (1). PPAR-α activation by various ligands, including derivatives of fatty acids and fibrates, upregulates the expression of several catabolic enzymes that are involved in mitochondrial and peroxisomal β-oxidation and micro-somal ω-oxidation, as well as in the transcriptional regulation of genes that are necessary for the maintenance of the redox balance during the oxidative catabolism of fatty acids (2,3). WY14,643, a well-known specific PPAR-α agonist, increases fatty acid oxidation by increasing the expressions of peroxi-somal and mitochondrial fatty acid β-oxidation enzymes (4). Accordingly, PPAR-α agonists are used in the treatment of hypertriglyceridemia (1). WY14,643 also reduces liver insulin resistance more efficiently than muscle insulin resistance and normalizes the circulating triglyceride levels and blood glu-cose levels in lipoatrophic mice (4,5). Finally, PPAR-α agonists
have anti-inflammatory effects in vascular cells and macro-phages (6,7), which, together with their antihyperlipidemic effects, might explain why they inhibit atherosclerotic plaque formation (7,8).
Among the several endogenous ligands proposed so far for PPAR-α, N-palmitoylethanolamine (PEA) and, particularly, N-oleoylethanolamine (OEA) are among the most potent with affinity constants estimated in the range of ~3 μmol/l and ~120 nmol/l, respectively (9,10). The PPAR-α-mediated effects of OEA have been widely investigated (see refs. 11,12 for reviews) and include: (i) inhibition of food intake via action on sensory neurons originating in the duodenum and terminating in the brainstem (13) or through fatty acid translocase/CD36-mediated stimulation of fatty acid uptake by enterocytes (14,15); (ii) reduction of body weight gain and triacylglycerol content in the liver and adipose tissue of diet-induced obese rats and wild-type (WT) mice, but not in obese PPAR-α−/− mice (16); (iii) stimulation of lipolysis and fatty acid
Basal and Fasting/Refeeding-regulated Tissue Levels of Endogenous PPaR-α Ligands in Zucker RatsAngelo A. Izzo1, Fabiana Piscitelli2, Raffaele Capasso1, Pietro Marini2, Luigia Cristino3, Stefania Petrosino2 and Vincenzo Di Marzo2
N-oleoylethanolamine (OEA) and N-palmitoylethanolamine (PEA) are endogenous lipids that activate peroxisome proliferator–activated receptor-α with high and intermediate potency, and exert anorectic and anti-inflammatory actions in rats, respectively. We investigated OEA and PEA tissue level regulation by the nutritional status in lean and obese rats. OEA and PEA levels in the brainstem, duodenum, liver, pancreas, and visceral (VAT) or subcutaneous (SAT) adipose tissues of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum or following overnight food deprivation, with and without refeeding, were measured by liquid chromatography–mass spectrometry. In WT rats, duodenal OEA, but not PEA, levels were reduced by food deprivation and restored by refeeding, whereas the opposite was observed for OEA in the pancreas, and for both mediators in the liver and SAT. In ad lib fed Zucker rats, PEA and OEA levels were up to tenfold higher in the duodenum, slightly higher in the brainstem, and lower in the other tissues. Fasting/refeeding-induced changes in OEA levels were maintained in the duodenum, liver, and SAT, and lost in the pancreas, whereas fasting upregulated this compound also in the VAT. The observed changes in OEA levels in WT rats are relevant to the actions of this mediator on satiety, hepatic and adipocyte metabolism, and insulin release. OEA dysregulation in Zucker rats might counteract hyperphagia in the duodenum, but contribute to hyperinsulinemia in the pancreas, and to fat accumulation in adipose tissues and liver. Changes in PEA levels might be relevant to the inflammatory state of Zucker rats.
Obesity (2009) 18, 55–62. doi:10.1038/oby.2009.186
1Endocannabinoid Research Group, Department of Experimental Pharmacology, University of Naples “Federico II”, Naples, Italy; 2Endocannabinoid Research Group, Institute of Biomolecular Chemistry, Consiglio Nazionale delle Ricerche, Pozzuoli, Italy; 3Endocannabinoid Research Group, Institute of Cybernetics, Consiglio Nazionale delle Ricerche, Pozzuoli, Italy. Correspondence: Vincenzo Di Marzo ([email protected])
Received 30 December 2008; accepted 5 May 2009; published online 11 June 2009. doi:10.1038/oby.2009.186

56 VOLUME 18 NUMBER 1 | jaNUaRy 2010 | www.obesityjournal.org
articlesintegrative Physiology
oxidation in skeletal muscle strips, dissociated hepatocytes and primary cardiomyocyte cultures (16); and (iv) reduction of body weight, plasma triacylglycerol, cholesterol levels, and fatty liver in obese Zucker rats and corresponding enhanced transcription of PPAR-α and its target genes, including fatty acid translocase/CD36, liver fatty acid binding protein, and uncoupling protein-2 in these rodents (17). However, OEA also produces glucose intolerance in vivo without decreasing insulin levels, whereas, ex vivo, it inhibits insulin-stimulated glucose uptake in isolated adipocytes (18). Furthermore, similar to the synthetic PPAR-α agonist WY-14,623, OEA decreases glucose- induced calcium oscillations and insulin release in intact mouse pancreatic islets (19). Possibly due to its lower efficacy as a PPAR-α agonist, PEA does not share these metabolic effects with OEA, although it causes anti-inflammatory actions mediated by this nuclear receptor (10). Furthermore, PEA inhibits lipopolysaccharide-induced lep-tin and tumor necrosis factor-α from human subcutaneous adipocytes, although these effects do not seem to be mediated by PPAR-α (20,21).
Both OEA and PEA are metabolically related to another N-acylethanolamine (NAE), i.e., N-arachidonoylethanolamine (anandamide, AEA), an endogenous agonist of cannabinoid CB1 receptors profoundly involved in the peripheral control of metabolism (22). However, OEA and PEA are completely inactive at cannabinoid receptors (11,22). Furthermore, we have shown that the regulation of OEA and PEA levels in mouse adipocytes during differentiation, in rat pancreatic β-cells grown in different concentrations of glucose, in the pancreas and visceral (VAT) and subcutaneous (SAT) adipose tissues of mice fed with a high-fat diet, and in human vol-unteers after a meal or with type 2 diabetes, are not always regulated in the same way as AEA (23). It is also known that the levels of OEA, PEA, and AEA in the duodenum are regu-lated in different ways following brief food deprivation (the levels of OEA being reduced, and those of AEA increased with PEA being unaltered (24,25)), and following 1 week supplementation of the diet with different fatty acids (26). These differences probably reflect NAE-specific differences in the accumulation of the NAE precursors, the N-acyl-phosphatidylethanolamines (NAPEs), or in the routes con-verting N-acyl-phosphatidylethanolamines into NAEs, as well as in the type of NAE hydrolytic enzymes (27). We have recently described the effects on the levels of AEA and of the other endocannabinoid, 2-arachidonoylglycerol, in peripheral tissues of the rat, of brief food deprivation and refeeding, and the alterations of such effects in obese Zucker rats, an ani-mal model of dyslipidemia and other metabolic disturbances occurring during obesity (28). Furthermore, several studies have been published so far on PPAR-α effects in various tis-sues of Zucker rats (29–32). Therefore, we have undertaken the present study in order to investigate the effects of food deprivation and refeeding on OEA and PEA levels in both central and peripheral tissues that control metabolism in lean rats, as well as the modifications of such effects in 7-week-old Zucker rats.
Methods and ProceduresanimalsSeven-week-old male WT Wistar rats (Harlan, Correzzana, Italy; ~280 g body weight) and age-matched male Zucker fa/fa rats (from Charles River Laboratories, Calco, Italy; ~320 g body weight) underwent differ-ent feeding regiments, after 1 week of acclimation. After treatments and killing, the brainstem, duodenum, liver, pancreas, and adipose (sub-cutaneous and mesenteric) tissues were removed and immersed into liquid nitrogen, to be stored at −70 °C until extraction and purification of OEA and PEA.
treatmentsThe experimental procedures were approved by the Animal Ethics Committee of the University of Naples. Animal care was in compliance with Italian (D.L. 116/92) and EEC (O.J. of E.C. L358/1 18/12/86) regu-lations on the protection of laboratory animals. All efforts were made to minimize the suffering of animals and to reduce the number of animals used. Groups of Wistar (n = 5) or Zucker rats (n = 5) were either fed ad libitum overnight and before killing (“ad lib” groups, killed at 7:30 am), or kept without food overnight and then killed (“fasted” groups, killed at 7:30 am), or kept without food overnight until 7 am, then fed for 30 min and then killed.
Measurement of oea and Pea levelsThe extraction, purification, and quantification of OEA and PEA from tissues require several biochemical steps as described previously (23). First, tissues were dounce-homogenized and extracted with chloro-form/methanol/Tris–HCl 50 mmol/l pH 7.5 (2:1:1, vol/vol) containing internal standards ([2H]4 PEA and [2H]4 OEA, 50 pmol each). The lipid-containing organic phase was dried down, weighed, and prepurified by open-bed chromatography on silica gel. Fractions were obtained by eluting the column with 9:1 (by vol) chloroform/methanol. After lipid extraction and separation, the prepurified lipids were analyzed by liq-uid chromatography–atmospheric pressure chemical ionization–mass spectrometry by using a Shimadzu high-performance liquid chroma-tography apparatus (LC-10ADVP) coupled to a Shimadzu (LCMS-2010) quadrupole mass spectrometry via a Shimadzu atmospheric pressure chemical ionization interface as previously described (23). The amounts of OEA and PEA in tissues, quantified by isotope dilution with the abovementioned deuterated standards, are expressed as pmol per gram of wet tissue weight.
statistical analysesANOVA followed by the Bonferroni’s test was used for comparisons of OEA and PEA levels among the various groups. P values <0.05 were considered significant.
Immunostaining of PPar-α in the brainstem of Wt ratsThree WT adult male rats were used. The animals were deeply anes-thetized (sodium pentobarbital, 60 mg/kg, intraperitoneal) and per-fused transcardially with saline followed by 4% paraformaldehyde in 0.1 mol/l phosphate buffer, pH 7.4. Each brain was removed from the skull, fixated for 2 h and then washed, and soaked for cryoprotection in 30% sucrose in phosphate buffer at 4 °C until it sank. Adjacent coronal frozen sections were cut through the brainstem at 12 µm and processed, alternately, for PPAR-α immunohistochemistry by means of peroxidase revelation, and Nissl-staining for the cytoarchitectonic analysis. All the sections were dehydrated with increasing concentrations of alcohol and coverslipped with De.P.X, mixture of distyrene, a plasticizer (tricresyl phosphate), and xylene. For anti-PPAR-α immunoperoxidase labeling, the sections were reacted for 10 min in 0.1% H2O2 to inactivate endog-enous peroxidase activity and incubated for 60 min at room tempera-ture in 10% normal rabbit serum (Vector Laboratories, Burlingame, CA) in 0.1 mol/l phosphate-buffered saline, pH 7.4, containing 0.3% Triton X-100 and 0.05% sodium azide (Sigma-Aldrich, St Louis, MO). The sections were then incubated overnight in a cold room with goat polyclonal antibody that recognizes the N terminus of PPAR-α receptor
obesity | VOLUME 18 NUMBER 1 | jaNUaRy 2010 57
articlesintegrative Physiology
(Santa Cruz Biotechnology, Santa Cruz, CA) diluted 1:500 in normal rabbit serum. After several rinses, the sections were incubated for 2 h in biotinylated rabbit anti-goat IgGs (Vector Laboratories) diluted 1:250 in normal rabbit serum, followed by incubation for 1 h at room tem-perature in the avidin–biotin–peroxidase solution (Vectastain ABC Kit; Vector Laboratories) in phosphate-buffered saline, and then for 10 min in 0.05‰ 3-3′-diaminobenzidine (Sigma Fast; Sigma-Aldrich) in 0.01 mol/l Tris–HCl-buffered saline, pH 7.6.
Controls of immunoreactivity reaction included preabsorption of diluted antibody with immunizing peptide (Santa Cruz Biotechnology, Santa Cruz, CA) or omission of the primary antisera or the second-ary antibody. These control experiments did not show staining. All the sections were studied under bright-field illumination (Leica DM IRB; Leica Microsystems, Wetzlar, Germany) and images were acquired using the digital camera Leica DFC 320 connected to the microscope and the image analysis software Leica IM500. Digital images were proc-essed in Adobe Photoshop, with brightness and contrast being the only adjustments made.
resultslevels of oea and Pea in the duodenumSimilar to what reported previously (24,25,33), the levels of OEA, but not those of PEA, in the duodenum of WT rats were significantly reduced following food deprivation and increased after refeeding (Figure 1). We report for the first time, how-ever, that in Zucker rats fed ad libitum, the duodenal levels of both PEA and OEA were ~3- and ~12-fold higher than in WT rats (Figure 1). Whereas PEA levels were upregulated follow-ing food deprivation, to return to basal after refeeding, OEA levels were reduced and enhanced by food deprivation and refeeding, respectively, as observed in WT rats.
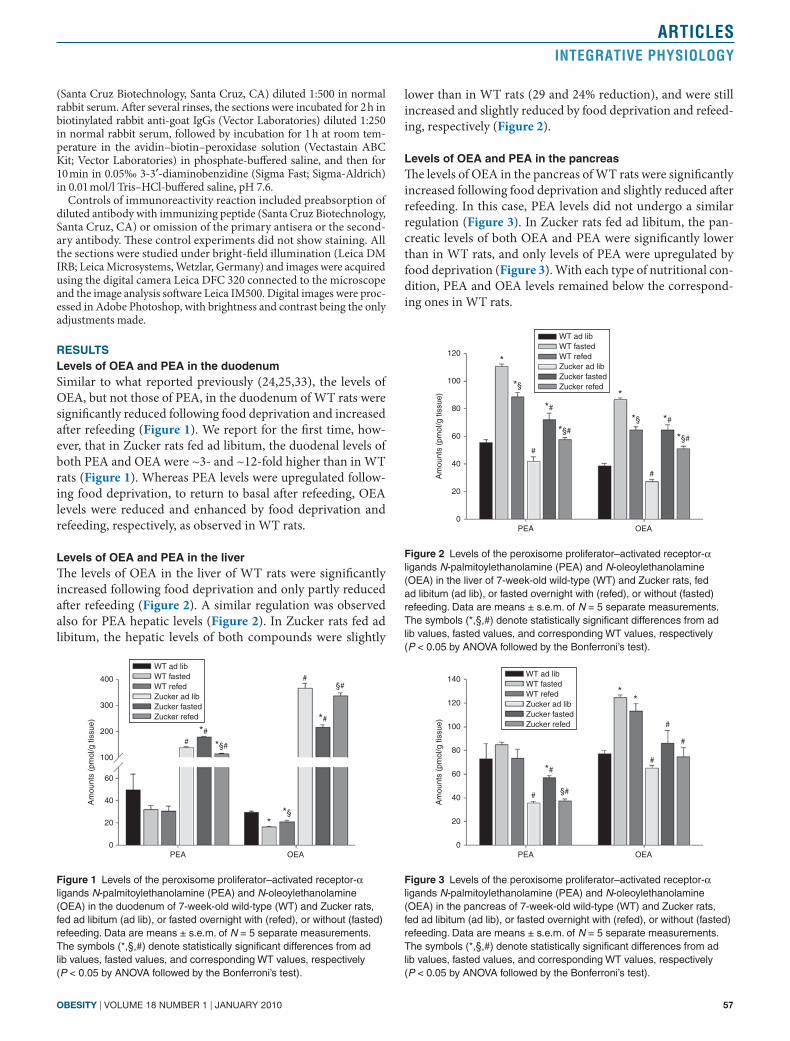
levels of oea and Pea in the liverThe levels of OEA in the liver of WT rats were significantly increased following food deprivation and only partly reduced after refeeding (Figure 2). A similar regulation was observed also for PEA hepatic levels (Figure 2). In Zucker rats fed ad libitum, the hepatic levels of both compounds were slightly
lower than in WT rats (29 and 24% reduction), and were still increased and slightly reduced by food deprivation and refeed-ing, respectively (Figure 2).
levels of oea and Pea in the pancreasThe levels of OEA in the pancreas of WT rats were significantly increased following food deprivation and slightly reduced after refeeding. In this case, PEA levels did not undergo a similar regulation (Figure 3). In Zucker rats fed ad libitum, the pan-creatic levels of both OEA and PEA were significantly lower than in WT rats, and only levels of PEA were upregulated by food deprivation (Figure 3). With each type of nutritional con-dition, PEA and OEA levels remained below the correspond-ing ones in WT rats.
0PEA OEA
20
40
60
100
200
300
Am
ount
s (p
mol
/g ti
ssue
)
400
WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed
#
#
*#
*#
*§#
§#
*§*
Figure 1 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the duodenum of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. The symbols (*,§,#) denote statistically significant differences from ad lib values, fasted values, and corresponding WT values, respectively (P < 0.05 by ANOVA followed by the Bonferroni’s test).
0PEA OEA
20
40
60
80
100
Am
ount
s (p
mol
/g ti
ssue
)
120
WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed
#
#
*#
*#*§#
*§#
*§
*§
*
*
Figure 2 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the liver of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. The symbols (*,§,#) denote statistically significant differences from ad lib values, fasted values, and corresponding WT values, respectively (P < 0.05 by ANOVA followed by the Bonferroni’s test).
0PEA OEA
20
40
60
80
100
Am
ount
s (p
mol
/g ti
ssue
)
140
120
WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed
#
#*#
#
§#
#
**
Figure 3 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the pancreas of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. The symbols (*,§,#) denote statistically significant differences from ad lib values, fasted values, and corresponding WT values, respectively (P < 0.05 by ANOVA followed by the Bonferroni’s test).

58 VOLUME 18 NUMBER 1 | jaNUaRy 2010 | www.obesityjournal.org
articlesintegrative Physiology
levels of oea and Pea in the satThe levels of OEA in the SAT of WT rats were significantly increased following food deprivation and reduced by refeed-ing. A similar regulation was observed also for PEA levels (Figure 4). Importantly, in Zucker rats fed ad libitum, the SAT levels of both OEA and PEA were approximately twofold lower than in WT rats and were still increased by food deprivation and reduced by refeeding (Figure 4). Also in this organ, with each type of nutritional condition, the levels of PEA and OEA remained well below the corresponding ones in WT rats.
levels of oea and Pea in the VatUnlike what observed in the SAT, OEA and PEA levels in the VAT did not undergo any changes following food deprivation and refeeding in WT rats (Figure 5). Also in the VAT of Zucker
rats fed ad libitum, the levels of both OEA and PEA were approximately twofold lower than in WT rats. Interestingly, in these obese rats, the VAT levels of both compounds became increased by food deprivation and reduced by refeeding (Figure 5). Also in this organ, however, with each type of nutritional condition, the levels of PEA and OEA remained well below the corresponding ones in WT rats.
levels of oea and Pea in the brainstemThe levels of OEA in the brainstem of WT rats were unaltered following food deprivation and refeeding, and a similar lack of regulation was observed also for PEA levels (Figure 6). In Zucker rats, the brainstem concentrations of both OEA and PEA were slightly higher than in WT rats and still did not change following food deprivation and reduced by refeeding (Figure 6).
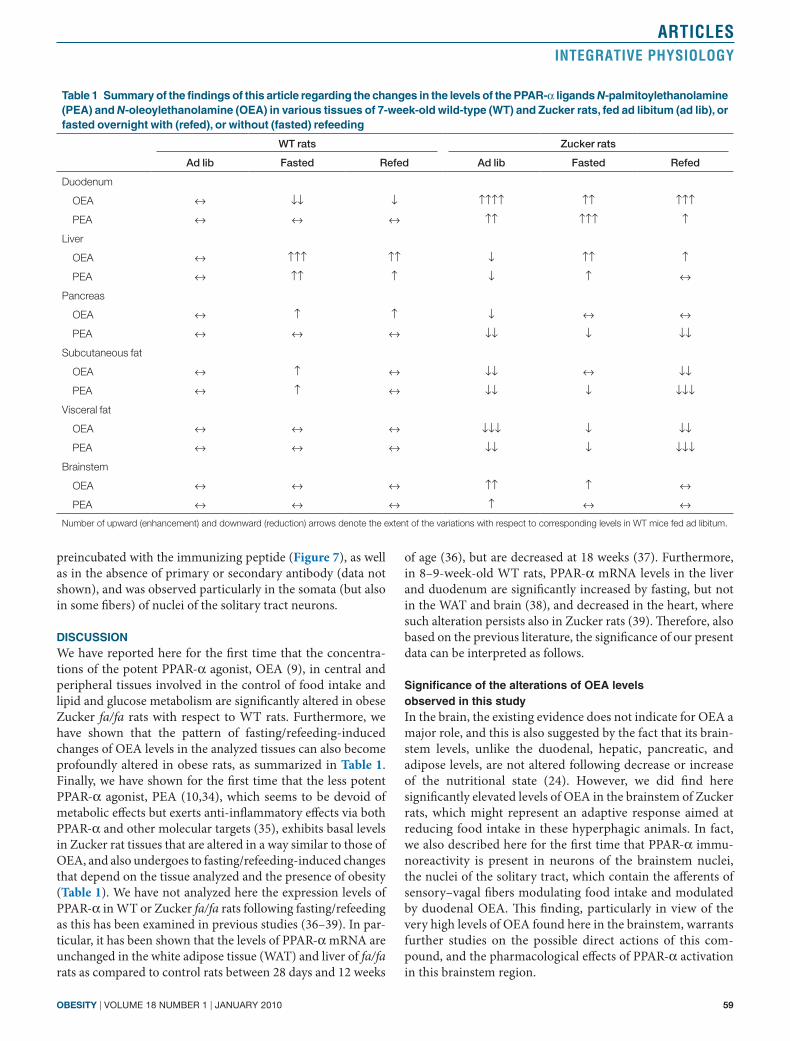
PPar-α expression in the nuclei of the solitary tract of Wt ratsFor the first time we describe here that an immunoreactiv-ity signal for PPAR-α is present in the nuclei of the solitary tract of WT rats, where sensory/vagal fibers originating in the duodenum and stomach, and activated by OEA (13) are known to terminate (Figure 7). The signal was absent in slices
0PEA OEA
100
200
300
Am
ount
s (p
mol
/g ti
ssue
)
400
WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed
#
#
*#
*#
*§#
§#§
§
*
*
Figure 4 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the subcutaneous adipose tissue of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. The symbols (*,§,#) denote statistically significant differences from ad lib values, fasted values, and corresponding WT values, respectively (P < 0.05 by ANOVA followed by the Bonferroni’s test).
0PEA OEA
100
200
300
Am
ount
s (p
mol
/g ti
ssue
)
400WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed
#
#
*#
*#
§#
*§#
Figure 5 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the visceral (mesenteric) adipose tissue of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. The symbols (*,§,#) denote statistically significant differences from ad lib values, fasted values, and corresponding WT values, respectively (P < 0.05 by ANOVA followed by the Bonferroni’s test).
0PEA OEA
200
400
600
Am
ount
s (p
mol
/g ti
ssue
)
800
1,000WT ad libWT fastedWT refedZucker ad libZucker fastedZucker refed#
# #
#
#
Figure 6 Levels of the peroxisome proliferator–activated receptor-α ligands N-palmitoylethanolamine (PEA) and N-oleoylethanolamine (OEA) in the brainstem of 7-week-old wild-type (WT) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding. Data are means ± s.e.m. of N = 5 separate measurements. #Statistically significant differences from corresponding WT values (P < 0.05 by ANOVA followed by the Bonferroni’s test).
PPAR-α PPAR-α + BP
Figure 7 Peroxisome proliferator–activated receptor-α (PPAR-α) immunoreactivity in neurons of the nuclei of the solitary tract in wild-type rats. Note the absence of the signal in slices preincubated with the immunizing blocking peptide (PPAR-α + BP). Scale bar = 200 μm.
obesity | VOLUME 18 NUMBER 1 | jaNUaRy 2010 59
articlesintegrative Physiology
preincubated with the immunizing peptide (Figure 7), as well as in the absence of primary or secondary antibody (data not shown), and was observed particularly in the somata (but also in some fibers) of nuclei of the solitary tract neurons.
dIscussIonWe have reported here for the first time that the concentra-tions of the potent PPAR-α agonist, OEA (9), in central and peripheral tissues involved in the control of food intake and lipid and glucose metabolism are significantly altered in obese Zucker fa/fa rats with respect to WT rats. Furthermore, we have shown that the pattern of fasting/refeeding-induced changes of OEA levels in the analyzed tissues can also become profoundly altered in obese rats, as summarized in Table 1. Finally, we have shown for the first time that the less potent PPAR-α agonist, PEA (10,34), which seems to be devoid of metabolic effects but exerts anti-inflammatory effects via both PPAR-α and other molecular targets (35), exhibits basal levels in Zucker rat tissues that are altered in a way similar to those of OEA, and also undergoes to fasting/refeeding-induced changes that depend on the tissue analyzed and the presence of obesity (Table 1). We have not analyzed here the expression levels of PPAR-α in WT or Zucker fa/fa rats following fasting/refeeding as this has been examined in previous studies (36–39). In par-ticular, it has been shown that the levels of PPAR-α mRNA are unchanged in the white adipose tissue (WAT) and liver of fa/fa rats as compared to control rats between 28 days and 12 weeks
of age (36), but are decreased at 18 weeks (37). Furthermore, in 8–9-week-old WT rats, PPAR-α mRNA levels in the liver and duodenum are significantly increased by fasting, but not in the WAT and brain (38), and decreased in the heart, where such alteration persists also in Zucker rats (39). Therefore, also based on the previous literature, the significance of our present data can be interpreted as follows.
significance of the alterations of oea levels observed in this studyIn the brain, the existing evidence does not indicate for OEA a major role, and this is also suggested by the fact that its brain-stem levels, unlike the duodenal, hepatic, pancreatic, and adipose levels, are not altered following decrease or increase of the nutritional state (24). However, we did find here significantly elevated levels of OEA in the brainstem of Zucker rats, which might represent an adaptive response aimed at reducing food intake in these hyperphagic animals. In fact, we also described here for the first time that PPAR-α immu-noreactivity is present in neurons of the brainstem nuclei, the nuclei of the solitary tract, which contain the afferents of sensory–vagal fibers modulating food intake and modulated by duodenal OEA. This finding, particularly in view of the very high levels of OEA found here in the brainstem, warrants further studies on the possible direct actions of this com-pound, and the pharmacological effects of PPAR-α activation in this brainstem region.
table 1 summary of the findings of this article regarding the changes in the levels of the PPar-α ligands N-palmitoylethanolamine (Pea) and N-oleoylethanolamine (oea) in various tissues of 7-week-old wild-type (Wt) and Zucker rats, fed ad libitum (ad lib), or fasted overnight with (refed), or without (fasted) refeeding
WT rats Zucker rats
Ad lib Fasted Refed Ad lib Fasted Refed
Duodenum
OEA ↔ ↓↓ ↓ ↑↑↑↑ ↑↑ ↑↑↑
PEA ↔ ↔ ↔ ↑↑ ↑↑↑ ↑
Liver
OEA ↔ ↑↑↑ ↑↑ ↓ ↑↑ ↑
PEA ↔ ↑↑ ↑ ↓ ↑ ↔
Pancreas
OEA ↔ ↑ ↑ ↓ ↔ ↔
PEA ↔ ↔ ↔ ↓↓ ↓ ↓↓
Subcutaneous fat
OEA ↔ ↑ ↔ ↓↓ ↔ ↓↓
PEA ↔ ↑ ↔ ↓↓ ↓ ↓↓↓
Visceral fat
OEA ↔ ↔ ↔ ↓↓↓ ↓ ↓↓
PEA ↔ ↔ ↔ ↓↓ ↓ ↓↓↓
Brainstem
OEA ↔ ↔ ↔ ↑↑ ↑ ↔
PEA ↔ ↔ ↔ ↑ ↔ ↔
Number of upward (enhancement) and downward (reduction) arrows denote the extent of the variations with respect to corresponding levels in WT mice fed ad libitum.

60 VOLUME 18 NUMBER 1 | jaNUaRy 2010 | www.obesityjournal.org
articlesintegrative Physiology
In the duodenum, chronically administered OEA exerts anorectic effects via PPAR-α (40) via increased feeding latency and postmeal interval—these effects might be both due to an action at sensory neurons terminating in the brainstem (13) and to release of fatty acids in enterocytes (15). We confirmed here that duodenal OEA levels decrease during fasting and increase after refeeding in rats (24,25,33). However, in WT rats, the tissue concentration of OEA detected here in ad lib fed rats or after refeeding (~30 nmol/l) might not be sufficient to fully activate PPAR-α (half-maximal effective concentration (EC50) for OEA ~120 nmol/l, (9)), the activity of which might, therefore, not be reduced after fasting. Accordingly, in WT rats, PPAR-α expression was previously reported to be increased, rather than reduced, after fasting (38). Instead, the dramati-cally higher (approximately tenfold) levels of OEA in the duodenum of Zucker rats are likely to fully activate PPAR-α, and can be interpreted as a strong adaptive reaction aiming at inhibiting food intake. In view of the stimulatory effect of die-tary oleic acid on OEA biosynthesis (15), this strong increase can be explained with the higher fatty acid intake that is likely to occur in these hyperphagic animals. Interestingly, the capa-bility of fasting to reduce and refeeding to enhance the duo-denal levels of OEA was not impaired in Zucker rats in which, probably, this signal keeps being as important as a negative regulator of metabolism as in WT rats, as suggested also by the capability of Zucker rats to respond to OEA by eating less and losing weight (41). Interestingly, the levels of OEA were previously shown to be elevated also in the whole stomach of diet-induced obese mice, and this finding was related to the capability of this compound to inhibit gastric emptying (an effect that might contribute to inhibition of food intake), and to the observed reduction of basal gastric emptying in these obese animals (42).
In the liver, OEA, by activating PPAR-α, reduces triacylglyc-erol accumulation and enhances fatty acid β-oxidation (16,17). We found here that hepatic OEA levels, which are slightly higher than in the duodenum (~40 nmol/l), increase up to over ~85 nmol/l following food deprivation, and decrease after refeeding. Given the similar fasting-induced increase previ-ously observed for PPAR-α expression in this organ (38), it is tempting to speculate that OEA does act as an activator of hepatic PPAR-α during fasting, possibly to induce fatty acid β-oxidation. Importantly, in Zucker rats, the hepatic OEA lev-els, although still responding to fasting/refeeding, were lower than in WT rats at all nutritional statuses studies, thus suggest-ing that, in this organ, reduction of both OEA and, particularly at an older age (37), PPAR-α levels contribute to one of the typical consequences of obesity, eventually observed also in aged Zucker rats, that is fatty liver.
In the pancreas, the levels of OEA were higher than in the duodenum (~80 nmol/l), and increased up to over ~120 nmol/l following food deprivation, to be reduced following refeeding. In this organ, PPAR-α agonists inhibit insulin release via a fast, nongenomic mechanism (18,19), and, therefore, the fasting/feeding-induced changes in the levels of OEA might underlie in part the decrease and increase of insulin levels occurring
during fasting and after feeding. Furthermore, pancreatic OEA levels were decreased in Zucker rats at all nutritional statuses studied, possibly prompting a role for PPAR-α in the hyperin-sulinemia typical of these obese rodents. In diet-induced obese mice, instead, no changes in the pancreatic levels of OEA levels were observed previously (23), indicating that there might be species differences in the regulation of this lipid.
In the WAT, OEA inhibits triacylglycerol accumulation and enhances fatty acid β-oxidation (16), possibly also through inhibition of glucose uptake (18), which, by preventing glu-cose oxidation, negates the biosynthesis of glycerol necessary for triacylglycerol biosynthesis. Indeed, the WAT is, among the peripheral tissues analyzed in this study, the one with the high-est concentrations of OEA (~300 nmol/l), in agreement with a role of this compound as a PPAR-α agonist in this organ. In fact, following fasting, OEA levels increased further (up to ~400 nmol/l), although only in the SAT, possibly in agree-ment with the necessity of releasing, from this fat store, fatty acids available for β-oxidation during the lack of energy intake. However, we found, at all three nutritional statuses analyzed here, a strong reduction of OEA levels in both the SAT and VAT of Zucker rats, which, in the absence of any compensative changes in WAT PPAR-α expression in these obese animals (36), and given the stimulatory effects on β-oxidation of OEA and PPAR-α in the WAT (16), might contribute to fat accumu-lation in these obese animals.
significance of the alterations in Pea levels observed in this studyWe found here that, of the six tissues analyzed, PEA levels were sensible to the nutritional status only in the liver and SAT, being in both cases elevated with fasting and reduced with refeeding. The changes in the liver cannot, to date, be prop-erly interpreted due to the lack of data on the effects of PEA in this organ, except for some actions on nonenzymatic lipid per-oxidation (supporting the idea that this compound possesses hepatoprotective properties (43)). The changes observed in the SAT might be related to the anti-inflammatory effects observed with PEA in human subcutaneous adipocytes stimulated with lipopolysaccharide (20,21). This might be particularly true for Zucker rats, which are characterized by an overall inflamma-tory state, especially in the adipose tissue, liver, and pancreas, and in which PEA levels became higher during fasting also in the duodenum, pancreas, and VAT. Within the framework of altered metabolism, as in Zucker rats, it is likely that fast-ing as well as refeeding, might either transiently worsen or ameliorate inflammation in tissues involved in metabolism via effects on the hypothalamic–pituitary–adrenal axis (which becomes overactive in fasted Zucker rats) or on reactive oxy-gen species. Indeed, possibly reflecting a common biosynthetic pathway with OEA but not necessarily with AEA, in all tis-sues from obese rats, basal PEA levels, like OEA levels, were either higher (brainstem, duodenum) or lower (pancreas, liver, WAT depots) than in the corresponding tissues from WT rats, in agreement with previous findings of either reduced (44,45) or increased (46–49) PEA levels in inflamed vs. healthy tissues.

obesity | VOLUME 18 NUMBER 1 | jaNUaRy 2010 61
articlesintegrative Physiology
However, until more data are available on potential metabolic effects of PEA, we cannot rule out the possibility that the changes observed here in the levels of this compound, as with OEA and AEA, are relevant to the control of food intake in WT rats, and to metabolic dysfunctions in Zucker rats.
Possible causes of nutritional status–dependent and obesity-induced changes of oea and Pea tissue concentrationsApart from the aforementioned effects of dietary oleic acid on OEA biosynthesis, observed selectively in enterocytes (15), and the fasting/refeeding-induced effects on NAE biosynthetic precursors and biosynthetic and inactivating enzymes in the duodenum (24), not much is known about the regulation of OEA and PEA levels in mammalian tissues. Both compounds are biosynthesized from the correspond-ing N-acyl-phosphatidylethanolamines, but not necessarily using the same biosynthetic enzyme (50). Furthermore, while OEA can be degraded by both fatty acid amide hydrolase-1 and 2, PEA is degraded by fatty acid amide hydrolase-1 and N-acylethanolamine acid amidohydrolase (12,27). Therefore, it was not surprising to see that the levels of the two com-pounds change in different tissue-specific and nutritional status–dependent ways. It is likely that OEA and PEA tissue levels are the complex result of combined as well as differen-tial effects of: (i) nutritional status–sensitive hormones, such as insulin, glucocorticoids, ghrelin, and leptin, on the activity of NAE biosynthetic and degrading enzymes (see refs. 51–56 for examples) and (ii) the amount and quality of the food ingested, which, possibly also through different amounts of essential fatty acids, affects the availability of NAE biosynthetic precur-sors in a NAE- and tissue-dependent way (57–59).
conclusionsWe have reported here for the first time both that the basal and fasting/refeeding-regulated levels of endogenous AEA–metabolically related PPAR-α ligands, one, OEA, deeply involved in the control of metabolism and the other, PEA, more efficacious at counteracting inflammation, are altered in Zucker rats. OEA, in particular, is upregulated in the duodenum, pos-sibly to antagonize hyperphagia, but seems to be dysfunctional in other organs in its capability of counteracting the metabolic consequences of obesity. These observations might underlie some of the metabolic and inflammation-related alterations observed in Zucker rats.
acknoWledgMentWe are grateful to Davide Castelluccio for his technical assistance.
dIsclosureThe authors declared no conflict of interest.
© 2009 The Obesity Society
reFerences1. Staels B, Maes M, Zambon A. Fibrates and future PPARalpha agonists in
the treatment of cardiovascular disease. Nat Clin Pract Cardiovasc Med 2008;5:542–553.
2. Yu S, Rao S, Reddy JK. Peroxisome proliferator-activated receptors, fatty acid oxidation, steatohepatitis and hepatocarcinogenesis. Curr Mol Med 2003;3:561–572.
3. Cho MC, Lee K, Paik SG, Yoon DY. Peroxisome Proliferators-Activated Receptor (PPAR) Modulators and Metabolic Disorders. PPAR Res 2008;2008:679137.
4. Chou CJ, Haluzik M, Gregory C et al. WY14,643, a peroxisome proliferator-activated receptor alpha (PPARalpha) agonist, improves hepatic and muscle steatosis and reverses insulin resistance in lipoatrophic A-ZIP/F-1 mice. J Biol Chem 2002;277:24484–24489.
5. Tsunoda M, Kobayashi N, Ide T et al. A novel PPARalpha agonist ameliorates insulin resistance in dogs fed a high-fat diet. Am J Physiol Endocrinol Metab 2008;294:E833–E840.
6. Chinetti-Gbaguidi G, Fruchart JC, Staels B. Pleiotropic effects of fibrates. Curr Atheroscler Rep 2005;7:396–401.
7. Zandbergen F, Plutzky J. PPARalpha in atherosclerosis and inflammation. Biochim Biophys Acta 2007;1771:972–982.
8. Srivastava RA, Jahagirdar R, Azhar S, Sharma S, Bisgaier CL. Peroxisome proliferator-activated receptor-alpha selective ligand reduces adiposity, improves insulin sensitivity and inhibits atherosclerosis in LDL receptor-deficient mice. Mol Cell Biochem 2006;285:35–50.
9. Fu J, Gaetani S, Oveisi F et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-alpha. Nature 2003;425:90–93.
10. Lo Verme J, Fu J, Astarita G et al. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol Pharmacol 2005;67:15–19.
11. Gaetani S, Kaye WH, Cuomo V, Piomelli D. Role of endocannabinoids and their analogues in obesity and eating disorders. Eat Weight Disord 2008;13:e42–e48.
12. Thabuis C, Tissot-Favre D, Bezelgues JB et al. Biological functions and metabolism of oleoylethanolamide. Lipids 2008;43:887–894.
13. Rodríguez de Fonseca F, Navarro M, Gómez R et al. An anorexic lipid mediator regulated by feeding. Nature 2001;414:209–212.
14. Yang Y, Chen M, Georgeson KE, Harmon CM. Mechanism of oleoylethanolamide on fatty acid uptake in small intestine after food intake and body weight reduction. Am J Physiol Regul Integr Comp Physiol 2007;292:R235–R241.
15. Schwartz GJ, Fu J, Astarita G et al. The lipid messenger OEA links dietary fat intake to satiety. Cell Metab 2008;8:281–288.
16. Guzmán M, Lo Verme J, Fu J et al. Oleoylethanolamide stimulates lipolysis by activating the nuclear receptor peroxisome proliferator-activated receptor alpha (PPAR-alpha). J Biol Chem 2004;279:27849–27854.
17. Fu J, Oveisi F, Gaetani S, Lin E, Piomelli D. Oleoylethanolamide, an endogenous PPAR-alpha agonist, lowers body weight and hyperlipidemia in obese rats. Neuropharmacology 2005;48:1147–1153.
18. González-Yanes C, Serrano A, Bermúdez-Silva FJ et al. Oleylethanolamide impairs glucose tolerance and inhibits insulin-stimulated glucose uptake in rat adipocytes through p38 and JNK MAPK pathways. Am J Physiol Endocrinol Metab 2005;289:E923–E929.
19. Ropero AB, Juan-Picó P, Rafacho A et al. Rapid non-genomic regulation of Ca2+ signals and insulin secretion by PPAR alpha ligands in mouse pancreatic islets of Langerhans. J Endocrinol 2009;200:127–138.
20. Hoareau L, Ravanan P, Gonthier MP et al. Effect of PEA on LPS inflammatory action in human adipocytes. Cytokine 2006;34:291–296.
21. Hoareau L, Buyse M, Festy F et al. Anti-inflammatory effect of palmitoylethanolamide on human adipocytes. Obesity (Silver Spring) 2009;17:431–438.
22. Di Marzo V. CB(1) receptor antagonism: biological basis for metabolic effects. Drug Discov Today 2008;13:1026–1041.
23. Matias I, Gonthier MP, Petrosino S et al. Role and regulation of acylethanolamides in energy balance: focus on adipocytes and beta-cells. Br J Pharmacol 2007;152:676–690.
24. Fu J, Astarita G, Gaetani S et al. Food intake regulates oleoylethanolamide formation and degradation in the proximal small intestine. J Biol Chem 2007;282:1518–1528.
25. Gómez R, Navarro M, Ferrer B et al. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J Neurosci 2002;22:9612–9617.
26. Artmann A, Petersen G, Hellgren LI et al. Influence of dietary fatty acids on endocannabinoid and N-acylethanolamine levels in rat brain, liver and small intestine. Biochim Biophys Acta 2008;1781:200–212.

62 VOLUME 18 NUMBER 1 | jaNUaRy 2010 | www.obesityjournal.org
articlesintegrative Physiology
27. De Petrocellis L, Di Marzo V. An introduction to the endocannabinoid system: from the early to the latest concepts. Best Pract Res Clin Endocrinol Metab 2009;23:1–15.
28. Izzo AA, Piscitelli F, Capasso R et al. Peripheral endocannabinoid dysregulation in obesity: relation to intestinal motility and energy processing induced by food deprivation and re-feeding. Br J Pharmacol 2009; e-pub ahead of print 3 April 2009.
29. Zhou YT, Shimabukuro M, Wang MY et al. Role of peroxisome proliferator-activated receptor alpha in disease of pancreatic beta cells. Proc Natl Acad Sci USA 1998;95:8898–8903.
30. Dana SL, Hoener PA, Bilakovics JM et al. Peroxisome proliferator-activated receptor subtype-specific regulation of hepatic and peripheral gene expression in the Zucker diabetic fatty rat. Metab Clin Exp 2001;50:963–971.
31. Reifel-Miller A, Otto K, Hawkins E et al. A peroxisome proliferator-activated receptor alpha/gamma dual agonist with a unique in vitro profile and potent glucose and lipid effects in rodent models of type 2 diabetes and dyslipidemia. Mol Endocrinol 2005;19:1593–1605.
32. Lanne B, Dahllöf B, Lindahl C et al. PPARalpha and PPARgamma regulation of liver and adipose proteins in obese and dyslipidemic rodents. J Proteome Res 2006;5:1850–1859.
33. Petersen G, Sørensen C, Schmid PC et al. Intestinal levels of anandamide and oleoylethanolamide in food-deprived rats are regulated through their precursors. Biochim Biophys Acta 2006;1761:143–50; discussion 141.
34. D’Agostino G, La Rana G, Russo R et al. Acute intracerebroventricular administration of palmitoylethanolamide, an endogenous peroxisome proliferator-activated receptor-alpha agonist, modulates carrageenan-induced paw edema in mice. J Pharmacol Exp Ther 2007;322: 1137–1143.
35. Re G, Barbero R, Miolo A, Di Marzo V. Palmitoylethanolamide, endocannabinoids and related cannabimimetic compounds in protection against tissue inflammation and pain: potential use in companion animals. Vet J 2007;173:21–30.
36. Gorla-Bajszczak A, Siegrist-Kaiser C, Boss O, Burger AG, Meier CA. Expression of peroxisome proliferator-activated receptors in lean and obese Zucker rats. Eur J Endocrinol 2000;142:71–78.
37. Oana F, Takeda H, Hayakawa K et al. Physiological difference between obese (fa/fa) Zucker rats and lean Zucker rats concerning adiponectin. Metab Clin Exp 2005;54:995–1001.
38. Escher P, Braissant O, Basu-Modak S et al. Rat PPARs: quantitative analysis in adult rat tissues and regulation in fasting and refeeding. Endocrinology 2001;142:4195–4202.
39. Young ME, Guthrie PH, Razeghi P et al. Impaired long-chain fatty acid oxidation and contractile dysfunction in the obese Zucker rat heart. Diabetes 2002;51:2587–2595.
40. Fu J, Kim J, Oveisi F, Astarita G, Piomelli D. Targeted enhancement of oleoylethanolamide production in proximal small intestine induces across-meal satiety in rats. Am J Physiol Regul Integr Comp Physiol 2008; 295:R45–R50.
41. Serrano A, Del Arco I, Javier Pavón F et al. The cannabinoid CB1 receptor antagonist SR141716A (Rimonabant) enhances the metabolic benefits of long-term treatment with oleoylethanolamide in Zucker rats. Neuropharmacology 2008;54:226–234.
42. Aviello G, Matias I, Capasso R et al. Inhibitory effect of the anorexic compound oleoylethanolamide on gastric emptying in control and overweight mice. J Mol Med 2008;86:413–422.
43. Gulaya NM, Kuzmenko AI, Margitich VM et al. Long-chain N-acylethanolamines inhibit lipid peroxidation in rat liver mitochondria under acute hypoxic hypoxia. Chem Phys Lipids 1998;97:49–54.
44. Petrosino S, Palazzo E, de Novellis V et al. Changes in spinal and supraspinal endocannabinoid levels in neuropathic rats. Neuropharmacology 2007;52:415–422.
45. Maione S, De Petrocellis L, de Novellis V et al. Analgesic actions of N-arachidonoyl-serotonin, a fatty acid amide hydrolase inhibitor with antagonistic activity at vanilloid TRPV1 receptors. Br J Pharmacol 2007;150:766–781.
46. Darmani NA, Izzo AA, Degenhardt B et al. Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and neuropathic conditions: review of the available pre-clinical data, and first human studies. Neuropharmacology 2005;48:1154–1163.
47. D’Argenio G, Petrosino S, Gianfrani C et al. Overactivity of the intestinal endocannabinoid system in celiac disease and in methotrexate-treated rats. J Mol Med 2007;85:523–530.
48. Loría F, Petrosino S, Mestre L et al. Study of the regulation of the endocannabinoid system in a virus model of multiple sclerosis reveals a therapeutic effect of palmitoylethanolamide. Eur J Neurosci 2008;28:633–641.
49. Garcia-Ovejero D, Arevalo-Martin A, Petrosino S et al. The endocannabinoid system is modulated in response to spinal cord injury in rats. Neurobiol Dis 2009;33:57–71.
50. Leung D, Saghatelian A, Simon GM, Cravatt BF. Inactivation of N-acyl phosphatidylethanolamine phospholipase D reveals multiple mechanisms for the biosynthesis of endocannabinoids. Biochemistry 2006;45:4720–4726.
51. Jo YH, Chen YJ, Chua SC Jr, Talmage DA, Role LW. Integration of endocannabinoid and leptin signaling in an appetite-related neural circuit. Neuron 2005;48:1055–1066.
52. Maccarrone M, Di Rienzo M, Finazzi-Agrò A, Rossi A. Leptin activates the anandamide hydrolase promoter in human T lymphocytes through STAT3. J Biol Chem 2003;278:13318–13324.
53. Waleh NS, Cravatt BF, Apte-Deshpande A, Terao A, Kilduff TS. Transcriptional regulation of the mouse fatty acid amide hydrolase gene. Gene 2002;291:203–210.
54. Malcher-Lopes R, Di S, Marcheselli VS et al. Opposing crosstalk between leptin and glucocorticoids rapidly modulates synaptic excitation via endocannabinoid release. J Neurosci 2006;26:6643–6650.
55. Murdolo G, Kempf K, Hammarstedt A et al. Insulin differentially modulates the peripheral endocannabinoid system in human subcutaneous abdominal adipose tissue from lean and obese individuals. J Endocrinol Invest 2007;30:RC17–RC21.
56. Kola B, Farkas I, Christ-Crain M et al. The orexigenic effect of ghrelin is mediated through central activation of the endogenous cannabinoid system. PLoS ONE 2008;3:e1797.
57. Berger A, Crozier G, Bisogno T et al. Anandamide and diet: inclusion of dietary arachidonate and docosahexaenoate leads to increased brain levels of the corresponding N-acylethanolamines in piglets. Proc Natl Acad Sci USA 2001;98:6402–6406.
58. Matias I, Léonhardt M, Lesage J et al. Effect of maternal under-nutrition on pup body weight and hypothalamic endocannabinoid levels. Cell Mol Life Sci 2003;60:382–389.
59. Matias I, Carta G, Murru E et al. Effect of polyunsaturated fatty acids on endocannabinoid and N-acyl-ethanolamine levels in mouse adipocytes. Biochim Biophys Acta 2008;1781:52–60.




















