
Automated segmentation of neuroanatomical structures in multispectral MR microscopy of the mouse...
11
Automated segmentation of neuroanatomical structures in multispectral MR microscopy of the mouse brain Anjum A. Ali, a, * Anders M. Dale, b Alexandra Badea, a and G. Allan Johnson a a Center for In Vivo Microscopy, Box 3302, Duke University Medical Center, Durham, NC 27710, USA b Department of Neurosciences and Radiology, University of California, San Diego, 9500 Gilman Drive MC0662, La Jolla, CA 92093-0662, USA Received 27 December 2004; revised 24 March 2005; accepted 5 April 2005 Available online 23 May 2005 We present the automated segmentation of magnetic resonance micro- scopy (MRM) images of the C57BL/6J mouse brain into 21 neuro- anatomical structures, including the ventricular system, corpus callosum, hippocampus, caudate putamen, inferior colliculus, internal capsule, globus pallidus, and substantia nigra. The segmentation algorithm operates on multispectral, three-dimensional (3D) MR data acquired at 90-Mm isotropic resolution. Probabilistic information used in the segmentation is extracted from training datasets of T2-weighted, proton density-weighted, and diffusion-weighted acquisitions. Spatial information is employed in the form of prior probabilities of occurrence of a structure at a location (location priors) and the pairwise probabilities between structures (contextual priors). Valida- tion using standard morphometry indices shows good consistency between automatically segmented and manually traced data. Results achieved in the mouse brain are comparable with those achieved in human brain studies using similar techniques. The segmentation algorithm shows excellent potential for routine morphological pheno- typing of mouse models. D 2005 Elsevier Inc. All rights reserved. Keywords: Automated segmentation; Magnetic resonance microscopy; Morphological phenotyping; Mouse brain Introduction Volumetric measurements, spatial organization of structures in the brain, and 3D shape analysis are now recognized as critical metrics of altered brain function and development (Caviness et al., 1999). Human studies have shown deviation from normal structure volume and shape in the presence of Alzheimer’s (Wang et al., 2003), depression (Steffens et al., 2000), and schizophrenia (Gaser et al., 2004). Magnetic resonance imaging (MRI) with its non- invasive nature, exceptional soft tissue contrast, and 3D digital format enables one to delineate structures and detect abnormalities between normal and diseased brains. Suitable selection of imaging parameters accentuates selected tissue properties based on proton density (PD), T1, T2, T2*, and diffusion contrast. MRI thus lends itself exceptionally well to morphometric studies in the brain where clear distinction between structures is required. The field of molecular genetics has produced animal models that replicate disease found in humans. Conventional histological sectioning of mouse models requires sectioning of specimens, which can be time-consuming and labor-intensive. Magnetic resonance microscopy (MRM), which is MRI with ultra-high spatial resolution of 10 – 100 Am, provides us with an alternative that is non-destructive and its 3D digital nature allows reslicing of data in any plane. It also obviates the need for reconstruction and alignment from sectioned slices. MRM can thus provide us with a reliable measure of structure volume and shape in the study of mouse models. We use the term magnetic resonance histology (MRH) to describe the characterization of tissue structure (Johnson et al., 1993). A normal adult mouse brain at 460 mg is approximately 3000 times smaller than an adult human brain at 1400 g. One needs to scale the spatial resolution accordingly from the 1-mm 3 voxels used in the clinical arena to voxel volumes <0.001 mm 3 . A number of studies have demonstrated the value of 3D MR morphometry in mouse models; the onset of atrophy in brains of C57BL/6J and ApoE-deficient mice (McDaniel et al., 2001); reduction in the volume of the anterior striatum of the Dopamine transporter knock-out; and reduction in the dentate gyrus before the onset of plaques in the PDAPP mouse (Redwine et al., 2003). A precursor to the morphometric measurement of brain structures, however, is their segmentation. Segmentation in the human brain using multispectral MR, initiated by Vannier et al. (1985), has been used extensively (Andersen et al., 2002; Cline et al., 1990; Fletcher et al., 1993; Lundervold and Storv, 1995; Pham et al., 1997). Data collected from multiple channels aid in differentiating between structure and tissue types that have overlapping intensity histograms in one spectral space and sufficient separation in another. Automated segmentation in the human brain, until very recently, concentrated only on classifica- tion of brain tissue into gray matter, white matter, and cerebrospi- 1053-8119/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.neuroimage.2005.04.017 * Corresponding author. Fax: +1 919 6847158. E-mail address: [email protected] (A.A. Ali). Available online on ScienceDirect (www.sciencedirect.com). www.elsevier.com/locate/ynimg NeuroImage 27 (2005) 425 – 435
Transcript of Automated segmentation of neuroanatomical structures in multispectral MR microscopy of the mouse...

www.elsevier.com/locate/ynimg
NeuroImage 27 (2005) 425 – 435
Automated segmentation of neuroanatomical structures in
multispectral MR microscopy of the mouse brain
Anjum A. Ali,a,* Anders M. Dale,b Alexandra Badea,a and G. Allan Johnsona
aCenter for In Vivo Microscopy, Box 3302, Duke University Medical Center, Durham, NC 27710, USAbDepartment of Neurosciences and Radiology, University of California, San Diego, 9500 Gilman Drive MC0662, La Jolla, CA 92093-0662, USA
Received 27 December 2004; revised 24 March 2005; accepted 5 April 2005
Available online 23 May 2005
We present the automated segmentation of magnetic resonance micro-
scopy (MRM) images of the C57BL/6J mouse brain into 21 neuro-
anatomical structures, including the ventricular system, corpus
callosum, hippocampus, caudate putamen, inferior colliculus, internal
capsule, globus pallidus, and substantia nigra. The segmentation
algorithm operates on multispectral, three-dimensional (3D) MR data
acquired at 90-Mm isotropic resolution. Probabilistic information used
in the segmentation is extracted from training datasets of T2-weighted,
proton density-weighted, and diffusion-weighted acquisitions. Spatial
information is employed in the form of prior probabilities of
occurrence of a structure at a location (location priors) and the
pairwise probabilities between structures (contextual priors). Valida-
tion using standard morphometry indices shows good consistency
between automatically segmented and manually traced data. Results
achieved in the mouse brain are comparable with those achieved in
human brain studies using similar techniques. The segmentation
algorithm shows excellent potential for routine morphological pheno-
typing of mouse models.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Automated segmentation; Magnetic resonance microscopy;
Morphological phenotyping; Mouse brain
Introduction
Volumetric measurements, spatial organization of structures in
the brain, and 3D shape analysis are now recognized as critical
metrics of altered brain function and development (Caviness et al.,
1999). Human studies have shown deviation from normal structure
volume and shape in the presence of Alzheimer’s (Wang et al.,
2003), depression (Steffens et al., 2000), and schizophrenia (Gaser
et al., 2004). Magnetic resonance imaging (MRI) with its non-
invasive nature, exceptional soft tissue contrast, and 3D digital
format enables one to delineate structures and detect abnormalities
1053-8119/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuroimage.2005.04.017
* Corresponding author. Fax: +1 919 6847158.
E-mail address: [email protected] (A.A. Ali).
Available online on ScienceDirect (www.sciencedirect.com).
between normal and diseased brains. Suitable selection of imaging
parameters accentuates selected tissue properties based on proton
density (PD), T1, T2, T2*, and diffusion contrast. MRI thus lends
itself exceptionally well to morphometric studies in the brain where
clear distinction between structures is required.
The field of molecular genetics has produced animal models
that replicate disease found in humans. Conventional histological
sectioning of mouse models requires sectioning of specimens,
which can be time-consuming and labor-intensive. Magnetic
resonance microscopy (MRM), which is MRI with ultra-high
spatial resolution of 10–100 Am, provides us with an alternative
that is non-destructive and its 3D digital nature allows reslicing of
data in any plane. It also obviates the need for reconstruction and
alignment from sectioned slices. MRM can thus provide us with a
reliable measure of structure volume and shape in the study of
mouse models. We use the term magnetic resonance histology
(MRH) to describe the characterization of tissue structure (Johnson
et al., 1993). A normal adult mouse brain at 460 mg is
approximately 3000 times smaller than an adult human brain at
1400 g. One needs to scale the spatial resolution accordingly from
the 1-mm3 voxels used in the clinical arena to voxel volumes
<0.001 mm3. A number of studies have demonstrated the value of
3D MR morphometry in mouse models; the onset of atrophy in
brains of C57BL/6J and ApoE-deficient mice (McDaniel et al.,
2001); reduction in the volume of the anterior striatum of the
Dopamine transporter knock-out; and reduction in the dentate
gyrus before the onset of plaques in the PDAPP mouse (Redwine
et al., 2003).
A precursor to the morphometric measurement of brain
structures, however, is their segmentation. Segmentation in the
human brain using multispectral MR, initiated by Vannier et al.
(1985), has been used extensively (Andersen et al., 2002; Cline et
al., 1990; Fletcher et al., 1993; Lundervold and Storv, 1995; Pham
et al., 1997). Data collected from multiple channels aid in
differentiating between structure and tissue types that have
overlapping intensity histograms in one spectral space and
sufficient separation in another. Automated segmentation in the
human brain, until very recently, concentrated only on classifica-
tion of brain tissue into gray matter, white matter, and cerebrospi-

Fig. 1. Large flip angle simulation for two tissues with different T1s (1100
ms and 900 ms) for 90- flip and 140- flip at different TRs.
A.A. Ali et al. / NeuroImage 27 (2005) 425–435426
nal fluid. Methods are either solely intensity-based (Gonzalez
Ballester et al., 2000; Kovacevic et al., 2002; Van Leemput et al.,
1999; Wells et al., 1996) or coupled with spatial constraints (Held
et al., 1997; Zhang et al., 2001). Increasing insights about
neurodegenerative disorders and brain development anomalies that
target a group of cells or a specific structure have focused research
on the sub-segmentation of gray matter and white matter into
physiologically relevant constituent structures. Individual struc-
tures are characteristic in their anatomical shape, size, and
orientation, which makes the segmentation complicated. Techni-
ques focusing on the automated segmentation of only specific
structures like the ventricular system (Schnack et al., 2001), the
thalamus and mediodorsal nucleus (Spinks et al., 2002), and
hippocampus (Crum et al., 2001) have been demonstrated. One of
the first studies for delineating multiple sub-structures in the gray
matter and white matter of brain was done using multispectral T1,
T2, proton density, Gd + T1, and perfusion imaging (Zavaljevski et
al., 2000). A complete labeling of each voxel in an MR volume of
the human brain into 37 left and right parts of neuroanatomical
structures has also been accomplished (Fischl et al., 2002). Feature
vectors from individual MR contrasts can be transformed into
statistically independent features using independent component
analysis (ICA) and subsequently used for classification (Amato
et al., 2003).
Although considerable progress has been made in automated
human brain segmentation, segmentation in the mouse brain has
received little attention. Studies that require volume analysis of
structures have used manual tracings, which are tedious and error-
prone. Consequently, methods have also relied on warping of brain
datasets into manually segmented volumes and extracting structure
labels. Diffusion tensor data have been used to distinguish white
matter structures in the mouse brain (Zhang et al., 2002); however,
an automated quantification of structure volumes has not been
attempted. The challenges to develop segmentation methods in the
mouse brain are enormous. The gray–white matter differentiation
that one sees in the human brain is much reduced in the mouse
brain, particularly at the high field required for MRH. Further, at
the high spatial resolution used for MRH, one sees much more
detail and complexity in structures, making a single approach
inefficient. Thus, there is a need for automated segmentation
methods specifically catered to the mouse brain, not only to
identify structures reliably, but also to report subtle differences in
volume and shape not visible to the human eye.
In this work, the classification of voxels in the mouse brain into
structures is accomplished by combining the MR intensity features
with spatial priors into a discriminant Bayes classifier. The spatial
priors integrate information about the location of structures in the
brain as well as the contextual relationship between structures. This
contextual information is modeled using the concept of Markov
random fields (MRFs). An MRF is a statistical model that enables
the probability of a label at a voxel location to be expressed as a
function of the labels in a predefined neighborhood of the voxel.
Modeling each neighborhood of voxels as a conditionally depend-
ent MRF with the properties of anisotropy and non-stationarity
allows the labels to be direction and location dependent. This
approach follows the algorithm developed by Fischl et al. (2002)
for automated segmentation in the human brain. There are
fundamental differences however; the technique here has been
explicitly adapted for the mouse brain where structure classes have
been chosen and inter-structure relationships have been derived
accordingly; the tissue is fixed and not live unlike the human brain
study. Finally, we use intensity information from five MR
channels. The Bayes classifier classifies each voxel into the most
probable label from the 21 structure classes defined.
Materials and methods
Tissue preparation
Six C57BL/6J male mice approximately 9 weeks in age were
used in this work.
Mice were anesthetized with an intraperitoneal injection of
pentobarbital (100 mg/kg). The animals were transcardially
perfused with an initial flush of 36 ml of 0.9% NaCl (37-C),followed by 36 ml of 10% buffered formalin. The head was then
stored overnight in formalin. Brains were excised the following
morning and stored in 0.9% saline for 3 days after which they were
scanned using the acquisition protocol outlined in the next section.
MR acquisition protocol
All imaging was performed on a 9.4-T (400 MHz) vertical bore
Oxford superconducting magnet. A 14 mm in diameter solenoid
RF coil was used. Accurate T2 weighting of tissue requires
complete recovery of the longitudinal magnetization before the
next RF excitation, thereby eliminating T1 dependence. This is
ensured with the repetition time TR of a pulse sequence set to 2–3
times the T1 of tissue of interest. In the mouse brain, tissue T1s are
of the order of 1000 ms (Johnson et al., 2002b), which makes the
ideal TR 2–3 s. For a 3D 100-Am volumetric acquisition, this
would result in a scan time of about 36 h for a spin-echo
experiment with a matrix size of 128 � 128 � 256 and NEX = 2. A
solution to achieve T2-weighted images at short TRs has been the
use of large flip angle (>90) pulses (Bogdan and Joseph, 1990;
DiIorio et al., 1995; Elster and Provost, 1993; Ma et al., 1996). Fig.
1 simulates the signal intensity curve for a conventional 90- flip
and 140- flip for different TRs in two tissues having T1 values of
1100 ms and 900 ms. Comparing the two continuous curves, we
see that the large flip angle and short TR combination ensures
higher equilibrium signal in the longitudinal axis and a faster
recovery to steady state. We also see that the T1 dependence
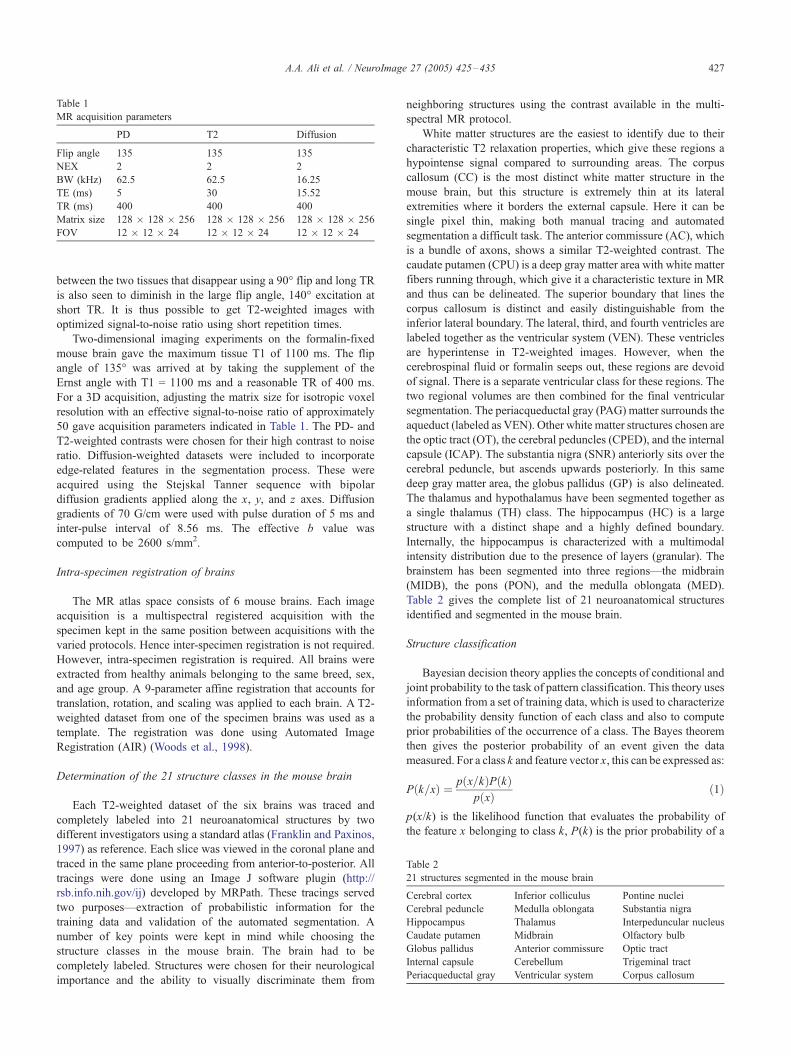
Table 1
MR acquisition parameters
PD T2 Diffusion
Flip angle 135 135 135
NEX 2 2 2
BW (kHz) 62.5 62.5 16.25
TE (ms) 5 30 15.52
TR (ms) 400 400 400
Matrix size 128 � 128 � 256 128 � 128 � 256 128 � 128 � 256
FOV 12 � 12 � 24 12 � 12 � 24 12 � 12 � 24
Table 2
21 structures segmented in the mouse brain
Cerebral cortex Inferior colliculus Pontine nuclei
Cerebral peduncle Medulla oblongata Substantia nigra
Hippocampus Thalamus Interpeduncular nucleus
Caudate putamen Midbrain Olfactory bulb
Globus pallidus Anterior commissure Optic tract
Internal capsule Cerebellum Trigeminal tract
Periacqueductal gray Ventricular system Corpus callosum
A.A. Ali et al. / NeuroImage 27 (2005) 425–435 427
between the two tissues that disappear using a 90- flip and long TRis also seen to diminish in the large flip angle, 140- excitation at
short TR. It is thus possible to get T2-weighted images with
optimized signal-to-noise ratio using short repetition times.
Two-dimensional imaging experiments on the formalin-fixed
mouse brain gave the maximum tissue T1 of 1100 ms. The flip
angle of 135- was arrived at by taking the supplement of the
Ernst angle with T1 = 1100 ms and a reasonable TR of 400 ms.
For a 3D acquisition, adjusting the matrix size for isotropic voxel
resolution with an effective signal-to-noise ratio of approximately
50 gave acquisition parameters indicated in Table 1. The PD- and
T2-weighted contrasts were chosen for their high contrast to noise
ratio. Diffusion-weighted datasets were included to incorporate
edge-related features in the segmentation process. These were
acquired using the Stejskal Tanner sequence with bipolar
diffusion gradients applied along the x, y, and z axes. Diffusion
gradients of 70 G/cm were used with pulse duration of 5 ms and
inter-pulse interval of 8.56 ms. The effective b value was
computed to be 2600 s/mm2.
Intra-specimen registration of brains
The MR atlas space consists of 6 mouse brains. Each image
acquisition is a multispectral registered acquisition with the
specimen kept in the same position between acquisitions with the
varied protocols. Hence inter-specimen registration is not required.
However, intra-specimen registration is required. All brains were
extracted from healthy animals belonging to the same breed, sex,
and age group. A 9-parameter affine registration that accounts for
translation, rotation, and scaling was applied to each brain. A T2-
weighted dataset from one of the specimen brains was used as a
template. The registration was done using Automated Image
Registration (AIR) (Woods et al., 1998).
Determination of the 21 structure classes in the mouse brain
Each T2-weighted dataset of the six brains was traced and
completely labeled into 21 neuroanatomical structures by two
different investigators using a standard atlas (Franklin and Paxinos,
1997) as reference. Each slice was viewed in the coronal plane and
traced in the same plane proceeding from anterior-to-posterior. All
tracings were done using an Image J software plugin (http://
rsb.info.nih.gov/ij) developed by MRPath. These tracings served
two purposes—extraction of probabilistic information for the
training data and validation of the automated segmentation. A
number of key points were kept in mind while choosing the
structure classes in the mouse brain. The brain had to be
completely labeled. Structures were chosen for their neurological
importance and the ability to visually discriminate them from
neighboring structures using the contrast available in the multi-
spectral MR protocol.
White matter structures are the easiest to identify due to their
characteristic T2 relaxation properties, which give these regions a
hypointense signal compared to surrounding areas. The corpus
callosum (CC) is the most distinct white matter structure in the
mouse brain, but this structure is extremely thin at its lateral
extremities where it borders the external capsule. Here it can be
single pixel thin, making both manual tracing and automated
segmentation a difficult task. The anterior commissure (AC), which
is a bundle of axons, shows a similar T2-weighted contrast. The
caudate putamen (CPU) is a deep gray matter area with white matter
fibers running through, which give it a characteristic texture in MR
and thus can be delineated. The superior boundary that lines the
corpus callosum is distinct and easily distinguishable from the
inferior lateral boundary. The lateral, third, and fourth ventricles are
labeled together as the ventricular system (VEN). These ventricles
are hyperintense in T2-weighted images. However, when the
cerebrospinal fluid or formalin seeps out, these regions are devoid
of signal. There is a separate ventricular class for these regions. The
two regional volumes are then combined for the final ventricular
segmentation. The periacqueductal gray (PAG) matter surrounds the
aqueduct (labeled as VEN). Other white matter structures chosen are
the optic tract (OT), the cerebral peduncles (CPED), and the internal
capsule (ICAP). The substantia nigra (SNR) anteriorly sits over the
cerebral peduncle, but ascends upwards posteriorly. In this same
deep gray matter area, the globus pallidus (GP) is also delineated.
The thalamus and hypothalamus have been segmented together as
a single thalamus (TH) class. The hippocampus (HC) is a large
structure with a distinct shape and a highly defined boundary.
Internally, the hippocampus is characterized with a multimodal
intensity distribution due to the presence of layers (granular). The
brainstem has been segmented into three regions—the midbrain
(MIDB), the pons (PON), and the medulla oblongata (MED).
Table 2 gives the complete list of 21 neuroanatomical structures
identified and segmented in the mouse brain.
Structure classification
Bayesian decision theory applies the concepts of conditional and
joint probability to the task of pattern classification. This theory uses
information from a set of training data, which is used to characterize
the probability density function of each class and also to compute
prior probabilities of the occurrence of a class. The Bayes theorem
then gives the posterior probability of an event given the data
measured. For a class k and feature vector x, this can be expressed as:
P k=xð Þ ¼ p x=kð ÞP kð Þp xð Þ ð1Þ
p(x/k) is the likelihood function that evaluates the probability of
the feature x belonging to class k, P(k) is the prior probability of a

A.A. Ali et al. / NeuroImage 27 (2005) 425–435428
class, and p(x) is the marginal probability of the occurrence of
feature x. P(k/x) is the probability of the state of nature being k
given the data measured in x. This is also known as the posteriori
probability. Having evaluated this expression for different k’s, the
k that corresponds to the maximum posterior probability is the
most likely class to which the measured data belong. This is also
known as the maximum a posteriori (MAP) estimate of the
classification.
In the context of image classification, the Bayes theorem can
be used to arrive at the MAP solution for the segmentation.
The posterior probability of the segmentation can be expressed as
P(L/Y), which is the probability of label configuration L given the
multivariate intensity measure Y. The prior probability of the
configuration of labels is given by P(L). The Bayes expression can
be formulated as:
P L=Yð Þ ¼ p Y=Lð ÞP Lð Þ ð2Þ
p(Y/L) is the likelihood of observing intensity vector Y for the
label configuration L. The likelihood measure that class k
generated a feature vector Y can be computed using the multi-
variate Gaussian distribution function and the maximum likelihood
estimates of mean lk(v) and covariance matrix ~k(v) derived from
the training data as follows:
p Y½ �k L vð Þ ¼ kð Þ ¼ 1ffiffiffiffiffiffi2p
pðk~
k
vð ÞkÞd=2
� exp� 1
2Y½ � vð Þ � lk vð Þð ÞT~
k
vð Þ�1Y½ � vð Þ � lk vð Þð Þ
� ���ð3Þ
p([ Y]kL(v) = k) is the probability of observing image intensity
vector Y if the label at v belonged to class k. d is the dimensionality
of the feature space. Intensity data from T2, proton-density, and
diffusion-weighted (x, y, and z gradients) acquisition at each voxel
form a five-dimensional feature vector [ Y1, Y2, Y3, Y4, Y5]v. The
exponential term in Eq. (3) is the Mahalanobis distance measure.
The intensity distribution of each structure class Fk_ is modeled as a
multivariate Gaussian at each location Fv_ in the atlas. The
Gaussian is characterized by mean lk(v) and covariance matrix
Rk(v) specified for each atlas location. Means and covariance
estimates can be computed as follows:
lk vð Þ ¼ 1
N~N
i¼1
Y1; Y2; Y3; Y4; Y5½ � vð Þ ð4Þ
~k
vð Þ ¼ 1
N � 1~N
i¼1
Y1; Y2; Y3; Y4; Y5½ � vð Þ � lk vð Þð Þ
� Y1; Y2; Y3; Y4; Y5½ � vð Þ � lk vð Þð ÞT ð5Þ
From Eqs. (4) and (5) we see that the class statistics vary with
location. This accounts for the multimodal nature of intensity
histograms in structures, where a single mean and covariance
matrix cannot adequately represent the data. Thus in the
classification step, intensity values at each voxel are evaluated
only against means and covariance values that describe the MR
intensity distribution data of structures occurring at that location in
the atlas. N is the number of manually labeled training sets that
make up the atlas.
lk(v) and ~k(v) are the maximum likelihood estimates of
structure class k at every location in the atlas and form the
multivariate intensity model for classification.
The P(L) term in the Bayes expression has to incorporate both
the location and the contextual priors of structure labels at every
location. Location prior is the prior probability of the occurrence of
a label at a voxel location. For every voxel location in the atlas, the
prior probability of a structure k occurring at location v was
computed. For a voxel at location Fv_, the location prior is
evaluated as the ratio of the number of times class k occurs at Fv_ tothe number of times a brain voxel is mapped to Fv_ in the training
datasets. This is computed as:
p L vð Þ ¼ kð Þ L vð Þ ¼ kð Þvox
ð6Þ
where L(v) indicates the label at location v in the atlas.
The contextual priors are estimated for each voxel location with
the adjoining 6 nearest voxels in a first order neighborhood system.
For a voxel at location v, the first order neighborhood system can
be described as Nv = {vn, vs, ve, vw, vf, vb}. vn, vs, ve, and vw are
the neighboring voxels in the 4 cardinal directions in the plane. vfand vb are the voxels in front and behind the plane, respectively.
Pairwise probabilities are computed for every combination of
structure pairs between the voxel under consideration and the six
neighboring voxels taken one at a time in the first order
neighborhood:
P vð Þkikj ¼vox L vð Þ ¼ kikL Nvð Þ ¼ kj
� �vox L vð Þ kif g ð7Þ
In Eq. (7), P(v)kikj is the probability of observing the class
ki at location v when class kj occurs at a voxel in Nv. A
histogram function computes the number of times the particular
label configuration has occurred; probabilities are then stored in
a K � K matrix for every atlas location v for all 6 directions
around the voxel of interest. K is 22 for 21 structure classes and one
miscellaneous class. Currently, the information is stored at a scale of
90 Am and is averaged in a 3 � 3 grid around each voxel.
Contextual priors are modeled using the concepts of Markov
random field (MRF) theory. A particular MRF model favors the
arrangement of labels that conform to itself by associating them
with larger probabilities (Li, 2001). A set of random variables or
fields L can be defined for an image domain S where L is a label
arrangement. Each field is a subset of the image S and its elements
can take on specific configuration of labels. L can be modeled as a
Markov random field at a location v with respect to its selected
neighborhood Nv and will satisfy the following two criteria:
P Lð Þ > 0 ð8Þ
P Lv kLS�vð Þ ¼ P Lv kLNvð Þ ð9Þ
where (S � v) is the set of all voxels in the image excluding the
voxel under consideration at v. The prior probability of the label
at voxel location v being dependent on remaining voxels in an
image is equivalent to it being dependent only on the labels of
voxels in a predefined neighborhood Nv. This is the Markov
property. P(L) now, in this case, is the joint probability of the
MRF characterized by the label pattern L. P(LvkLS�v) is the
local conditional probability at voxel v. An MRF can be
evaluated globally using the joint probability or locally using
conditional probabilities. Furthermore, the equivalence between

Fig. 2. Proton density (PD), T2, and diffusion-weighted images from
registered 3D multispectral acquisition of formalin-fixed mouse brain at
level of the thalamus (left column) and midbrain (right column).
A.A. Ali et al. / NeuroImage 27 (2005) 425–435 429
the MRF conditional probability and the Gibbs joint probability
distributions enables the probability of the MRF to be expressed
as an energy function of many local Fclique_ potentials. A clique
is a subset of voxels in the image domain. The first order
neighborhood Nv of a voxel can thus be represented as a clique
and the clique potentials as a combination of the probabilities of
labels at each location given their neighborhood. Applying the
Fig. 3. Automated segmentation results in a slice from 3D MRM of the mouse br
defined labels.
Markovian property, the local conditional dependence can be
expressed as:
p L vð ÞkL S � vð Þð Þ ¼ p L vð ÞkL v1ð Þ; L v2ð Þ; L v3ð Þ; N ; L við Þð Þ; viZNv
ð10ÞThis reduces the global classification task in an image to a set of
smaller local classifications where the prior probability of a label at a
voxel location v is affected only by the labels at voxels defined for a
neighborhood Nv and independent of the neighborhood of other
voxels. As a result, the prior probability of a segmentation pattern
P(L) can be expressed as the product of local conditional
probabilities of each voxel with its neighborhood:
p Lð Þ ¼ PvZS
p L vð ÞkL v1ð Þ; L v2ð Þ; L v3ð Þ; N ; L við ÞÞ; viZNvð ð11Þ
Applying Bayes rule on the right-hand side of Eq. (11), the
conditional probabilities can be interchanged with the product of
the location prior at each voxel and the conditional probability of
labels in a neighborhood given the label at the center voxel:
p Lð Þ” PvZS
p L vð Þð Þp L v1ð Þ; L v2ð Þ; N ; L við ÞkL vð ÞÞ; viZNvð ð12Þ
Fischl et al. (2002) have simplified Eq. (12) by assuming only a first
order dependence between labels in a neighborhood in which case
the probability of a label given the labels in a neighborhood can be
computed by taking the product of the conditional probabilities with
respect to each of the neighboring labels at a time. The expression
can be written as:
p Lð Þ” PvZS
p L vð Þð Þ Pk
i¼1p L við ÞkL vð Þ; við Þ ð13Þ
Eq. (13) evaluates the prior probability of a segmentation pattern L
and incorporates both the location priors for each structure class at
each location v and the contextual priors between structures at
different voxels. Applying the iterated conditional modes algorithm
(Besag, 1986) for a faster convergence to the MAP solution of the
equation, the segmentation is initialized using only the multivariate
intensity model. Using this initial estimate, the segmentation label at
each voxel is iteratively updated by estimating the label that
maximizes the conditional posterior probability given the condi-
tional density of the multivariate intensity vector and the current
label estimate at each voxel.
The complete expression is given as follows:
arg max p L vð Þ ¼ k kL við Þ; Y½ � vð Þ; við Þ¼ p Y½ � vð ÞkL vð Þ ¼ kð Þp L vð Þ ¼ kð Þ P
k
i¼1p L við ÞkL vð Þ¼ k; við Þ ð14Þ
p([ Y](v)kL(v) = k) is the conditional probability of observing
an intensity vector Y for a particular class k. p(L(v) = k) is
ain. (A) Gray scale, (B) automated segmentation results, and (C) manually

Fig. 4. Automated segmentation results in a slice from 3D MRM of the mouse brain. (A) Gray scale, (B) automated segmentation results, and (C) manually
defined labels.
A.A. Ali et al. / NeuroImage 27 (2005) 425–435430
the prior probability of the voxel at v being labeled as
belonging to structure k. Pi=1
kp(L(vi)kL(v) = k,vi) is the joint
conditional probability given the current estimate of labels at
location v and vi where viZNv. Eq. (14) is evaluated until no
Fig. 5. 3D volume-rendered structures automatically segmented and overlaid on M
ant—anterior, pos—posterior, rt—right, lt—left.
more voxels change labels. This is followed by a post-processing
step of connected component labeling. Isolated groups of voxels
are relabeled according to the majority of labels in their
neighborhood.
RM volume of mouse brain in dorsal (A), ventral (B), and sagittal (C) view;

A.A. Ali et al. / NeuroImage 27 (2005) 425–435 431
Results and discussion
MR acquisition
Six mouse brains were scanned using the acquisition protocol
outlined in Table 1. A total of 30 3D volume datasets were
obtained with five contrasts; PD weighting, T2 weighting, and
diffusion weighting (x, y, z) per brain. Slices at two different
coronal levels in the brain are displayed in Fig. 2. The left column
is at the level of the thalamus and the column at the right is in the
midbrain area.
Segmentation and validation
The algorithm was evaluated on each of the six mouse brain
volumes. Probabilistic information containing multivariate inten-
Fig. 6. 3D volume-rendered structures automatically segmented and overlaid o
Magnified view; ant—anterior, pos—posterior, rt—right, lt—left.
sity distributions and the local and contextual priors is extracted
and stored for each brain. However, for the segmentation of each
brain, only the data from the remaining five volumes are used as
part of the training set. Thus, the test volume was kept out of the
atlas set. Each brain was segmented automatically and then
compared with results from its manual tracings. Segmentation by
two raters was used for computing the inter-rater reliability metrics.
Figs. 3 and 4 illustrate the labels for a slice from 3D volumetric
data of a mouse brain from automated and manual segmentation.
By visual inspection, we see that the segmentation works well even
in areas of complicated neuroanatomy where a number of
structures are present in close vicinity to each other. This is
illustrated in Fig. 4 where the hippocampus first appears anteriorly.
In the coronal slices of this area we find from medial to lateral a
close proximity of the hippocampus, lateral ventricles, caudate
putamen, and inferiorly the thalamus, internal capsule, and globus
n MRM volume of mouse brain in dorsal (A) and ventral (B) view. (C)

A.A. Ali et al. / NeuroImage 27 (2005) 425–435432
pallidus. All structures here are segmented in a spatial config-
uration consistent with the manual segmentation. Misclassifica-
tions of voxels predominantly take place at the boundaries between
different structures, which is a result of partial volume effects. Figs.
5 and 6 give 3D volume-rendered visualization of the segmented
structures overlaid in an MR volume generated using Volocity
software (http://www.bucher.ch/improvision/volocity.html). The
opacity of the MR volume has been adjusted to show structures
inside the brain. In each set, a selection of structures is highlighted
with the others turned off. Highlighted structures in Fig. 5 include
the cerebellum, inferior colliculus, thalamus, ventricular system,
midbrain, cerebral peduncles, pontine nuclei, globus pallidus,
internal capsule, and the substantia nigra. Fig. 6 shows the
ventricular system, caudate putamen, periacqueductal grey, hippo-
campus, internal capsule, and globus pallidus. Fig. 6C illustrates
the ventricular system and associated structures magnified.
Fig. 7. Reliability of automated segmentation with manual tracing for 21 structures
structures. Bottom: percent difference overlap for same structures. Key: CORTEX
caudate putamen, GP—globus pallidus, ICAP—internal capsule, PAG—periacque
thalamus, MIDB—midbrain, AC—anterior commissure, CBLM—cerebellum, V
INTPEDN—interpeduncular nucleus, OLFBLB—olfactory bulb, OPT—optic trac
The segmentation was statistically validated using morphom-
etry indices of percent volume overlap and percent difference
overlap. Letting LM and LA indicate labeling of a structure from the
manual tracing and automated segmentation, respectively, the
volume overlap percentage between them for a structure can be
computed using the equation:
O LA kLMð Þ ¼ V LA7LMð ÞV LAð Þ þ V LMð Þð Þ=2 � 100 ð15Þ
The intersection term in the numerator makes this metric sensitive
to differences in spatial location to a greater extent than differences
in volume or size. This is also known as the j index or the Dice
coefficient. Higher values of this metric indicate greater conformity
between the automated and manual segmentation. The overlap
percentage is not influenced by structure size or sample size and,
. Error bars are standard errors of the mean. Top: percent volume overlap for
—cerebral cortex, CPED—cerebral peduncle, HC—hippocampus, CPU—
ductal gray, INFC—inferior colliculus, MED—medulla oblongata, THAL—
EN—ventricular system, PON—pontine nuclei, SNR—substantia nigra,
t, TRI—trigeminal tract, CC—corpus callosum.

Table 3
Results comparing automated segmentation in mouse brain with human
brain
Structure Percent
overlap
(mouse)
Percent
overlap
(human)
Hippocampus 87 80
Thalamus 93 87
Caudate putamen 88 90
Ventricular system 70 90
A.A. Ali et al. / NeuroImage 27 (2005) 425–435 433
thus, is a good measure of efficacy of the segmentation especially
in our case.
Volumetric differences between labelings independent of the
spatial position can be verified by a difference function given
as:
D LA; LMð Þ ¼ kV LAð Þ � V LMð ÞkV LAð Þ þ V LMð Þð Þ=2 � 100 ð16Þ
Fig. 7 illustrates the percent volume overlap and difference
overlap between the automated segmentation and manual tracing
for the 21 structures. The algorithm does exceedingly well for
large structures like the cortex, hippocampus, caudate putamen,
medulla oblongata, thalamus, midbrain, inferior colliculus, and
cerebellum (80–90%). Small and thin structures like the anterior
commissure, optic tract, and trigeminal tract are penalized for their
size (50–60%). The corpus callosum is large in size, but its width
decreases to single pixel laterally where it lines the external
capsule and hence does not segment out as well (70%). The
ventricles suffer from high variability in their size and shape in the
excised brains, which could be due to perfusion and handling
differences. In cases where fluid seeps out of the ventricle after
perfusion, the ventricles are hypointense and collapsed. Fluid
retained in the ventricles makes them appear expanded and
hyperintense with contrast. Results for deep cortical structures like
the internal capsule, globus pallidus, periacqueductal grey, and
substantia nigra are encouraging. The difference overlap indicated
the difference in actual volume measurements. All values here are
sufficiently low, which indicate good agreement between manual
and automated segmentation. Fig. 8 gives the inter-rater reliability
from percent volume overlap of manual segmentations from the
two raters. Table 3 shows the percent volume overlap measures
comparing results for the mouse brain with those seen using the
same technique (Fischl et al., 2002) in human brain studies. While
the sample dataset (n = 6) is limited, results indicate that we will
be able to autosegment the mouse brain with accuracy comparable
to that achieved now in human studies. The software code
currently executes the automated segmentation of a 90-Am
Fig. 8. Inter-rater reliability for manual segmentation for the 21 structures. Key: C
CPU—caudate putamen, GP—globus pallidus, ICAP—internal capsule, PAG—p
THAL—thalamus, MIDB—midbrain, AC—anterior commissure, CBLM—cereb
nigra, INTPEDN—interpeduncular nucleus, OLFBLB—olfactory bulb, OPT—op
resolution mouse brain dataset in 15 min on a 2.5-GHz Macintosh
system.
Conclusions and future work
We have successfully implemented automated segmentation of
the complete mouse brain into 21 structures, including sub-cortical
structures like the corpus callosum, hippocampus, internal capsule,
globus pallidus, periacqueductal grey, and substantia nigra. The
algorithm is trained using multispectral MR intensity data, prior
probabilities of the spatial location of structures, and inter-structure
contextual relationships. The use of T2, PD, and diffusion-weighted
contrasts enables the algorithm to incorporate characteristics from
multiple channels in the classification. Although a linear registration
has been used in this work, the application of higher dimensional and
non-linear registration techniques would improve the segmentation.
This is especially relevant when the genetic mutation used severely
affects the anatomical relationships in the resultant mouse model.
The current work achieves a segmentation reliability of 70–90%
between manual and automated segmentation.
Though the current work uses multiple MR contrasts, it would
be useful to determine the contribution of each contrast in the
segmentation. A future directive is to quantify the contribution of
each MR channel in the segmentation and estimate an optimum
subset of contrasts required. The contrasts can be further optimized
ORTEX—cerebral cortex, CPED—cerebral peduncle, HC—hippocampus,
eriacqueductal gray, INFC—inferior colliculus, MED—medulla oblongata,
ellum, VEN—ventricular system, PON—pontine nuclei, SNR—substantia
tic tract, TRI—trigeminal tract, CC—corpus callosum.

A.A. Ali et al. / NeuroImage 27 (2005) 425–435434
for the segmentation of a specific structure or a group of structures.
The 90-Am resolution used in this work is still at the lower end for
studying detailed neuroanatomy in the mouse brain. The average
width of the granular layer in the mouse brain hippocampus is, for
example, 30 Am, making high resolution indispensable. An
isotropic resolution of 10–15 Am is what a neuroanatomist would
like to look at for effectively extracting histologically relevant
information from MR microscopy. Recently, 20-Am resolution has
been attained with the use of novel Factive staining_ techniques thatinvolve perfusion fixation combined with a T1 reducing contrast
agent (Johnson et al., 2002a). Due to the high signal-to-noise ratio
gained by T1 shortening agents and the enhanced resolution,
structure that we do not normally see in the formalin-fixed brain
appears in exquisite detail in the actively stained brain. Thus, the
next step is the segmentation of structures in the higher resolution
MRM of the mouse brain. The high resolution might enable us to
further subdivide already segmented structures into constituent
components. Ultimately, this would enable robust and expedited
morphometry analysis of structures to aid in the morphological
phenotyping of mouse models.
Acknowledgments
The authors thank Boma Fubara and Will Kurylo for their
contribution. All work was performed at the Duke Center for In
Vivo Microscopy, an NCRR/NCI National Resource (P41
RR05959 and R24 CA92656). This work was made possible
through collaborations with the Mouse Bioinformatics Research
Network (MBIRN).
References
Amato, U., Larobina, M., Antoniadis, A., Alfano, B., 2003. Segmentation
of magnetic resonance images through discriminant analysis. J. Neuro-
sci. Methods 131, 65–74.
Andersen, A.H., Zhang, Z., Avison, M.J., Gash, D.M., 2002. Automated
segmentation of multispectral brain MR images. J. Neurosci. Methods
122, 13–23.
Besag, J., 1986. On the statistical analysis of dirty pictures (with
discussion). J. R. Stat. Soc. 48, 259–302.
Bogdan, A.R., Joseph, P.M., 1990. RASEE: a rapid spin-echo pulse
sequence. Magn. Reson. Imaging 8, 9–13.
Caviness, V., Lange, N., Makris, M., Herbert, M., Kennedy, D., 1999. MRI-
based brain volumetrics: emergence of a developmental brain science
from a tool. Brain Develop. 21, 289–295.
Cline, H.E., Lorensen, W.E., Kikinis, R., Jolesz, F., 1990. Three-dimen-
sional segmentation of MR images of the head using probability and
connectivity. J. Comput. Assist. Tomogr. 14, 1037–1045.
Crum, W.R., Scahill, R.I., Fox, N.C., 2001. Automated hippocampal
segmentation by regional fluid registration of series MRI: validation and
application in Alzheimers disease. NeuroImage 13, 847–855.
DiIorio, G., Brown, J.J., Borrello, J.A., Perman, W.H., Shu, H.H., 1995.
Large angle spin-echo imaging. Magn. Reson. Imaging 13, 39–44.
Elster, A.D., Provost, T.J., 1993. Large-tip-angle spin-echo imaging. Theory
and applications. Invest. Radiol. 10, 944–953.
Fischl, B., Salat, D.H., Busa, E., Albert, M., Dieterich, M., Haselgrove, C.,
Van der Kouwe, A., Killiany, R., Kennedy, D., Klaveness, S., et al.,
2002. Whole brain segmentation: automated labeling of neuroanato-
mical structures in the human brain. Neuron 33, 341–355.
Fletcher, L.M., Barsotti, J.B., Hornak, J.P., 1993. A multispectral analysis
of brain tissues. Magn. Reson. Med. 29, 623–630.
Franklin, K.B.J., Paxinos, G., 1997. The Mouse Brain in Stereotaxic
Coordinates. Academic Press.
Gaser, C., Nenadic, I., Buchsbaum, B.R., Hazlett, E.A., Buchsbaum, M.S.,
2004. Ventricular enlargement in schizophrenia related to volume
reduction of the thalamus, striatum, and superior temporal cortex. Am.
J. Psychiatry 161, 154–156.
Gonzalez Ballester, M.A., Zisserman, A., Brady, M., 2000. Segmentation
and measurement of brain structures in MRI including confidence
bounds. Med. Image Anal. 4, 189–200.
Held, K., Kops, E.R., Krause, B.J., Wells, W.M., Kikinis, R., Muller-
Gartner, H.W., 1997. Markov random field segmentation of brain MR
images. IEEE Trans. Med. Imag. 16, 878–886.
Johnson, G.A., Benveniste, H., Black, R.D., Hedlund, L.W., Maronpot,
R.R., Smith, B.R., 1993. Histology by magnetic resonance microscopy.
Magn. Reson. Q. 9, 1–30.
Johnson, G.A., Cofer, G.P., Fubara, B., Gewalt, S.L., Hedlund, L.W.,
Maronpot, R.R., 2002a. Magnetic resonance histology for structural
phenotyping. Journal of Magnetic Resonance Imaging 16, 423–429.
Johnson, G.A., Cofer, G.P., Gewalt, S.L., Hedlund, L.W., 2002b.
Morphologic phenotyping with magnetic resonance microscopy: the
visible mouse. Radiology 222, 789–793.
Kovacevic, N., Lobaugh, N.J., Bronskill, M.J., Levine, B., Feinstein, A.,
Black, S.E., 2002. A robust method for extraction and automatic
segmentation of brain images. NeuroImage 17, 1087–1100.
Li, S.Z., 2001. Markov Random Field Modeling in Image Analysis, 2nd edR
Springer, Verlag.
Lundervold, A., Storv, G., 1995. Segmentation of brain parenchyma and
cerebrospinal fluid in multispectral magnetic resonance image. IEEE
Trans. Med. Imag. 14, 339–349.
Ma, J., Wehrli, F.W., Song, H.K., 1996. Fast 3D large-angle spin-echo
Imaging (3D FLASE). Magn. Reson. Med. 35, 903–910.
McDaniel, B., Sheng, H., Warner, D.S., Hedlund, L.W., Benveniste, H.,
2001. Tracking brain volume changes in C57BL/6J and ApoE-deficient
mice in a model of neurodegeneration: a 5 week longitudinal micro-
MRI study. NeuroImage 14, 1244–1255.
Pham, D.L., Prince, J.L., Dagher, A.P., Xu, C., 1997. An automated
technique for statistical characterization of brain tissues in magnetic
resonance imaging. Int. J. Pattern Recogn. Artif. Intell. 11, 1189–1211.
Redwine, J.M., Kosofsky, B., Jacobs, R.E., Games, D., Reilly, J.F.,
Morrison, J.H., Young, W.G., Bloom, F.E., 2003. Dentate gyrus volume
is reduced before onset of plaque formation in PDAPP mice: a magnetic
resonance microscopy and stereologic analysis. Proc. Natl. Acad. Sci.
100, 1381–1386.
Schnack, H.G., Hulshoff Pol, H.E., Baare, W.F.C., Viergever, M.A., Kahn,
R.S., 2001. Automatic segmentation of the ventricular system from MR
images of the human brain. NeuroImage 14, 95–104.
Spinks, R., Magnotta, V.A., Andreasen, N.C., Albright, K.C., Ziebell, S.,
Nopoulos, P., Cassell, M., 2002. Manual and automated measurement of
the whole thalamus and mediodorsal nucleus using magnetic resonance
imaging. NeuroImage 17, 631–642.
Steffens, D.C., Byrum, C.E., McQuoid, D.R., Greenberg, D.L., Payne,
M.E., Blitchington, T.F., MacFall, J.R., Krishnan, K.R., 2000.
Hippocampal volume in geriatric depression. Biol. Psychiatry 48,
301–309.
Van Leemput, K., Maes, F., Vandermeulen, D., P, S., 1999. Automated
model-based tissue classification of MR images of the brain. IEEE
Trans. Med. Imag. 18, 897–908.
Vannier, M.W., Butterfield, R.L., Jordan, D., Murphy, W.A., Levitt, R.G.,
Gado, M., 1985. Multispectral analysis of magnetic resonance images.
Radiology 154, 221–224.
Wang, L., Swank, J.S., Glick, I.E., Gado, M.H., Miller, M.I., Morris, J.C.,
Sernansky, J.G., 2003. Changes in hippocampal volume and shape
across time distinguish dementia of the Alzheimer type from healthy
aging. NeuroImage 20, 667–682.
Wells, W.M., Grimson, E.L., Kinkinis, R., Jolesz, F.A., 1996. Adaptive
segmentation of MRI data. IEEE Trans. Med. Imag. 15, 429–442.
Woods, R.P., Grafton, S.T., Holmes, C.J., Cherry, S.R., Mazziotta, J.C.,

A.A. Ali et al. / NeuroImage 27 (2005) 425–435 435
1998. Automated image registration: I. General methods and intra-
subject, intramodality validation. Journal of Computer Assisted Tomog-
raphy 22, 139–152.
Zavaljevski, A., Dhawan, A.P., Gaskil, M., Ball, W., Johnson, J.D., 2000.
Multi-level adaptive segmentation of multi-parameter MR brain images.
Comput. Med. Imag. Graph. 24, 87–98.
Zhang, Y., Smith, S., Brady, M., 2001. Segmentation of brain MR images
through a hidden Markov random field model and the expectation–
maximization algorithm. IEEE Trans. Med. Imag. 20, 45–57.
Zhang, J., Van Zijl, P.C., Mori, S., 2002. Three-dimensional diffusion tensor
magnetic resonance microimaging of adult mouse brain and hippo-
campus. NeuroImage 15, 892–901.




















