
Atypical presentation of Leigh syndrome associated with a Leber hereditary optic neuropathy primary...
8
Atypical presentation of Leigh syndrome associated with a Leber hereditary optic neuropathy primary mitochondrial DNA mutation Gary Fruhman a,b,1 , Megan L. Landsverk a,1 , Timothy E. Lotze c , Jill V. Hunter d , Michael F. Wangler a , Adekunle M. Adesina e , Lee-Jun C. Wong a , Fernando Scaglia a, ⁎ a Department of Molecular and Human Genetics, Baylor College of Medicine, Houston, TX, USA b Department of Obstetrics and Gynecology, Baylor College of Medicine, Houston, TX, USA c Department of Pediatrics, Division of Pediatric Neurology, Baylor College of Medicine, Baylor College of Medicine, Houston, TX, USA d Department of Radiology, Baylor College of Medicine, Houston, TX, USA e Department of Pathology, Baylor College of Medicine, Houston, TX, USA abstract article info Article history: Received 21 February 2011 Accepted 21 February 2011 Available online 26 February 2011 Keywords: Atypical Leigh syndrome Leber hereditary optic neuropathy Mitochondrial DNA mutation Phenotypic variation Leber hereditary optic neuropathy (LHON) is caused by point mutations in mitochondrial DNA (mtDNA), and is characterized by bilateral, painless sub-acute visual loss that develops during the second decade of life. Here we report the case of a five year old girl who presented with clinical and neuroradiological findings reminiscent of Leigh syndrome but carried a mtDNA mutation m.11778GNA (p.R340H) in the MTND4 gene usually observed in patients with LHON. This case is unusual for age of onset, gender, associated neurological findings and evolution, further expanding the clinical spectrum associated with primary LHON mtDNA mutations. © 2011 Elsevier Inc. All rights reserved. 1. Introduction Mutations in mitochondrial DNA (mtDNA) are associated with a spectrum of clinical syndromes including among others, mitochon- drial encephalomyopathy with lactic acidosis and stroke-like episodes (MELAS), myoclonic epilepsy with ragged-red fibers (MERRF), neurogenic muscle weakness, ataxia and retinitis pigmentosa (NARP) and Leber hereditary optic neuropathy (LHON) [1]. The m.11778GNA transition changes a conserved arginine to a histidine residue within the most evolutionarily conserved region in the MTND4 gene encoding a component of complex I of the mitochondrial respiratory chain [2]. This mutation is typically associated with the LHON phenotype, and is by far the most common LHON mutation, accounting for approximately 70% of cases observed among Caucasian populations [3]. It generally causes the most severe visual failure with only about 4% of patients experiencing visual recovery. The pene- trance for developing optic atrophy differs depending on the gender of the individual carrying this mutation; the risk is 43% for males and 11% for females [4]. However, the risk of visual failure in both genders increases when this mutation arises in haplogroup J, and decreases when the mutation arises in haplogroup H [5]. The m.3460GNA is the second most common mtDNA mutation causing LHON, and also causes severe visual failure, although more than 20% of patients experience visual recovery. The m.14459GNA is the most pathogenic mtDNA mutation causing LHON, and is associated with visual loss, dystonia, a generalized movement disorder, pseudobulbar syndrome, and myopathic features among other clinical manifestations [6]. Leigh syndrome is usually a devastating neurodegenerative disease (also called subacute necrotizing encephalopathy) that presents mostly in infancy. From a neuroradiological perspective, it consists of almost identical lesions in the central nervous system, e.g., focal, bilaterally symmetrical lesions, particularly in the basal ganglia, thalamus, and brainstem [7,8]. The clinical presentation of Leigh syndrome is highly variable with considerable heterogeneity and is characterized by a wide variety of clinical features. Most frequently, the central nervous system is affected, with psychomotor retardation, epilepsy, nystagmus, ophthalmoparesis, optic atrophy, ataxia, dysto- nia, or respiratory failure associated with brainstem dysfunction. Some patients also present with involvement of the peripheral nervous system, including polyneuropathy, myopathy, or non- neurological abnormalities such as diabetes, short stature, hyper- trichosis, cardiomyopathy, anemia, renal failure, vomiting, or diarrhea (atypical Leigh syndrome) [9]. The disease is usually fatal [10]. Leigh syndrome can be caused by mutations affecting either the mitochon- drial or the nuclear genome leading to dysfunction of subunits of the respiratory chain (particularly complexes I, II, IV, or V), coenzyme Q 10 deficiency, pyruvate dehydrogenase complex or pyruvate carboxylase deficiencies [9,11]. Molecular Genetics and Metabolism 103 (2011) 153–160 ⁎ Corresponding author at: Baylor College of Medicine, Department of Molecular and Human Genetics, 6701 Fannin Street, Suite 1560, Houston, TX 77030, USA. Fax: + 1 832 825 4294. E-mail address: [email protected] (F. Scaglia). 1 Coauthors. 1096-7192/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.ymgme.2011.02.014 Contents lists available at ScienceDirect Molecular Genetics and Metabolism journal homepage: www.elsevier.com/locate/ymgme
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Atypical presentation of Leigh syndrome associated with a Leber hereditary optic neuropathy primary...

Molecular Genetics and Metabolism 103 (2011) 153–160
Contents lists available at ScienceDirect
Molecular Genetics and Metabolism
j ourna l homepage: www.e lsev ie r.com/ locate /ymgme
Atypical presentation of Leigh syndrome associated with a Leber hereditary opticneuropathy primary mitochondrial DNA mutation
Gary Fruhman a,b,1, Megan L. Landsverk a,1, Timothy E. Lotze c, Jill V. Hunter d, Michael F. Wangler a,Adekunle M. Adesina e, Lee-Jun C. Wong a, Fernando Scaglia a,⁎a Department of Molecular and Human Genetics, Baylor College of Medicine, Houston, TX, USAb Department of Obstetrics and Gynecology, Baylor College of Medicine, Houston, TX, USAc Department of Pediatrics, Division of Pediatric Neurology, Baylor College of Medicine, Baylor College of Medicine, Houston, TX, USAd Department of Radiology, Baylor College of Medicine, Houston, TX, USAe Department of Pathology, Baylor College of Medicine, Houston, TX, USA
⁎ Corresponding author at: Baylor College of MedicineHuman Genetics, 6701 Fannin Street, Suite 1560, Housto825 4294.
E-mail address: [email protected] (F. Scaglia).1 Coauthors.
1096-7192/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.ymgme.2011.02.014
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 February 2011Accepted 21 February 2011Available online 26 February 2011
Keywords:Atypical Leigh syndromeLeber hereditary optic neuropathyMitochondrial DNA mutationPhenotypic variation
Leber hereditary optic neuropathy (LHON) is caused by point mutations in mitochondrial DNA (mtDNA), andis characterized by bilateral, painless sub-acute visual loss that develops during the second decade of life. Herewe report the case of a five year old girl who presented with clinical and neuroradiological findingsreminiscent of Leigh syndrome but carried a mtDNA mutation m.11778GNA (p.R340H) in the MTND4 geneusually observed in patients with LHON. This case is unusual for age of onset, gender, associated neurologicalfindings and evolution, further expanding the clinical spectrum associated with primary LHON mtDNAmutations.
, Department of Molecular andn, TX 77030, USA. Fax: +1 832
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
Mutations in mitochondrial DNA (mtDNA) are associated with aspectrum of clinical syndromes including among others, mitochon-drial encephalomyopathywith lactic acidosis and stroke-like episodes(MELAS), myoclonic epilepsy with ragged-red fibers (MERRF),neurogenic muscle weakness, ataxia and retinitis pigmentosa(NARP) and Leber hereditary optic neuropathy (LHON) [1]. Them.11778GNA transition changes a conserved arginine to a histidineresidue within the most evolutionarily conserved region in theMTND4 gene encoding a component of complex I of the mitochondrialrespiratory chain [2]. This mutation is typically associated with theLHON phenotype, and is by far the most common LHON mutation,accounting for approximately 70% of cases observed among Caucasianpopulations [3]. It generally causes the most severe visual failure withonly about 4% of patients experiencing visual recovery. The pene-trance for developing optic atrophy differs depending on the genderof the individual carrying this mutation; the risk is 43% for males and11% for females [4]. However, the risk of visual failure in both gendersincreases when this mutation arises in haplogroup J, and decreaseswhen the mutation arises in haplogroup H [5]. The m.3460GNA is the
second most common mtDNA mutation causing LHON, and alsocauses severe visual failure, although more than 20% of patientsexperience visual recovery. The m.14459GNA is the most pathogenicmtDNA mutation causing LHON, and is associated with visual loss,dystonia, a generalized movement disorder, pseudobulbar syndrome,and myopathic features among other clinical manifestations [6].
Leigh syndrome is usually a devastating neurodegenerativedisease (also called subacute necrotizing encephalopathy) thatpresents mostly in infancy. From a neuroradiological perspective, itconsists of almost identical lesions in the central nervous system, e.g.,focal, bilaterally symmetrical lesions, particularly in the basal ganglia,thalamus, and brainstem [7,8]. The clinical presentation of Leighsyndrome is highly variable with considerable heterogeneity and ischaracterized by a wide variety of clinical features. Most frequently,the central nervous system is affected, with psychomotor retardation,epilepsy, nystagmus, ophthalmoparesis, optic atrophy, ataxia, dysto-nia, or respiratory failure associated with brainstem dysfunction.Some patients also present with involvement of the peripheralnervous system, including polyneuropathy, myopathy, or non-neurological abnormalities such as diabetes, short stature, hyper-trichosis, cardiomyopathy, anemia, renal failure, vomiting, or diarrhea(atypical Leigh syndrome) [9]. The disease is usually fatal [10]. Leighsyndrome can be caused by mutations affecting either the mitochon-drial or the nuclear genome leading to dysfunction of subunits of therespiratory chain (particularly complexes I, II, IV, or V), coenzyme Q10
deficiency, pyruvate dehydrogenase complex or pyruvate carboxylasedeficiencies [9,11].
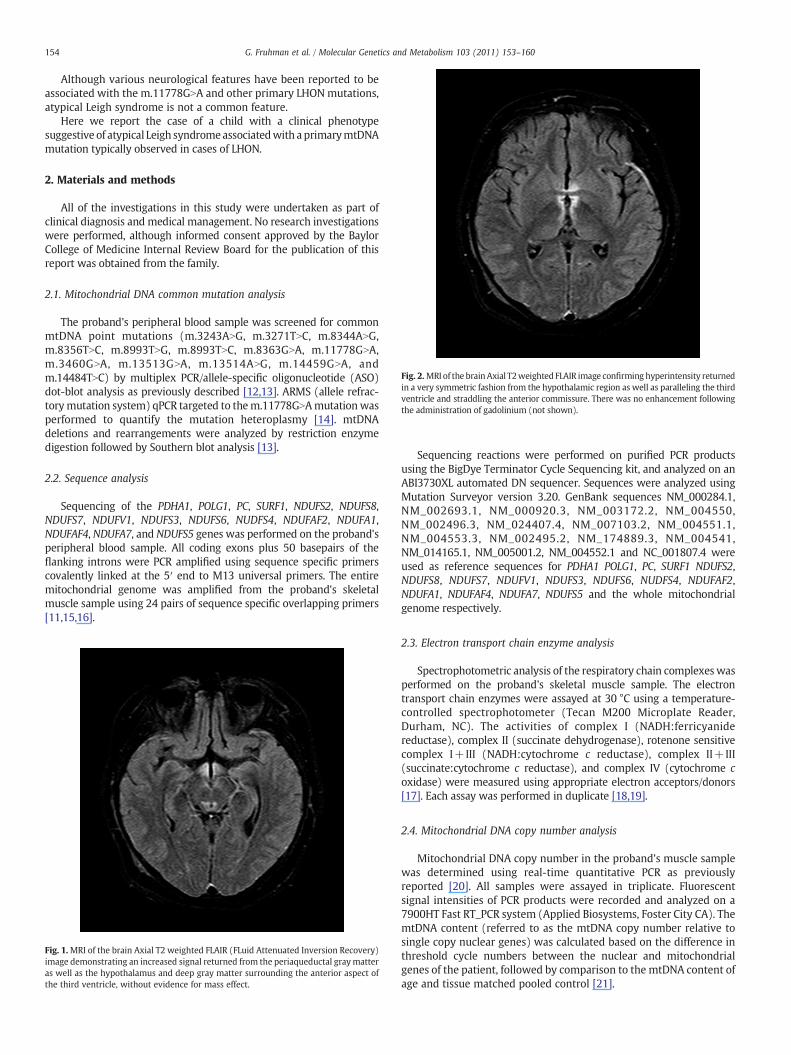
Fig. 2.MRI of the brainAxial T2weighted FLAIR image confirminghyperintensity returnedin a very symmetric fashion from the hypothalamic region as well as paralleling the thirdventricle and straddling the anterior commissure. There was no enhancement followingthe administration of gadolinium (not shown).
154 G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
Although various neurological features have been reported to beassociated with the m.11778GNA and other primary LHON mutations,atypical Leigh syndrome is not a common feature.
Here we report the case of a child with a clinical phenotypesuggestive of atypical Leigh syndromeassociatedwith aprimarymtDNAmutation typically observed in cases of LHON.
2. Materials and methods
All of the investigations in this study were undertaken as part ofclinical diagnosis and medical management. No research investigationswere performed, although informed consent approved by the BaylorCollege of Medicine Internal Review Board for the publication of thisreport was obtained from the family.
2.1. Mitochondrial DNA common mutation analysis
The proband's peripheral blood sample was screened for commonmtDNA point mutations (m.3243ANG, m.3271TNC, m.8344ANG,m.8356TNC, m.8993TNG, m.8993TNC, m.8363GNA, m.11778GNA,m.3460GNA, m.13513GNA, m.13514ANG, m.14459GNA, andm.14484TNC) by multiplex PCR/allele-specific oligonucleotide (ASO)dot-blot analysis as previously described [12,13]. ARMS (allele refrac-torymutation system) qPCR targeted to them.11778GNAmutationwasperformed to quantify the mutation heteroplasmy [14]. mtDNAdeletions and rearrangements were analyzed by restriction enzymedigestion followed by Southern blot analysis [13].
2.2. Sequence analysis
Sequencing of the PDHA1, POLG1, PC, SURF1, NDUFS2, NDUFS8,NDUFS7, NDUFV1, NDUFS3, NDUFS6, NUDFS4, NDUFAF2, NDUFA1,NDUFAF4, NDUFA7, and NDUFS5 genes was performed on the proband'speripheral blood sample. All coding exons plus 50 basepairs of theflanking introns were PCR amplified using sequence specific primerscovalently linked at the 5′ end to M13 universal primers. The entiremitochondrial genome was amplified from the proband's skeletalmuscle sample using 24 pairs of sequence specific overlapping primers[11,15,16].
Fig. 1.MRI of the brain Axial T2 weighted FLAIR (FLuid Attenuated Inversion Recovery)image demonstrating an increased signal returned from the periaqueductal graymatteras well as the hypothalamus and deep gray matter surrounding the anterior aspect ofthe third ventricle, without evidence for mass effect.
Sequencing reactions were performed on purified PCR productsusing the BigDye Terminator Cycle Sequencing kit, and analyzed on anABI3730XL automated DN sequencer. Sequences were analyzed usingMutation Surveyor version 3.20. GenBank sequences NM_000284.1,NM_002693.1, NM_000920.3, NM_003172.2, NM_004550,NM_002496.3, NM_024407.4, NM_007103.2, NM_004551.1,NM_004553.3, NM_002495.2, NM_174889.3, NM_004541,NM_014165.1, NM_005001.2, NM_004552.1 and NC_001807.4 wereused as reference sequences for PDHA1 POLG1, PC, SURF1 NDUFS2,NDUFS8, NDUFS7, NDUFV1, NDUFS3, NDUFS6, NUDFS4, NDUFAF2,NDUFA1, NDUFAF4, NDUFA7, NDUFS5 and the whole mitochondrialgenome respectively.
2.3. Electron transport chain enzyme analysis
Spectrophotometric analysis of the respiratory chain complexeswasperformed on the proband's skeletal muscle sample. The electrontransport chain enzymes were assayed at 30 °C using a temperature-controlled spectrophotometer (Tecan M200 Microplate Reader,Durham, NC). The activities of complex I (NADH:ferricyanidereductase), complex II (succinate dehydrogenase), rotenone sensitivecomplex I+III (NADH:cytochrome c reductase), complex II+III(succinate:cytochrome c reductase), and complex IV (cytochrome coxidase) were measured using appropriate electron acceptors/donors[17]. Each assay was performed in duplicate [18,19].
2.4. Mitochondrial DNA copy number analysis
Mitochondrial DNA copy number in the proband's muscle samplewas determined using real-time quantitative PCR as previouslyreported [20]. All samples were assayed in triplicate. Fluorescentsignal intensities of PCR products were recorded and analyzed on a7900HT Fast RT_PCR system (Applied Biosystems, Foster City CA). ThemtDNA content (referred to as the mtDNA copy number relative tosingle copy nuclear genes) was calculated based on the difference inthreshold cycle numbers between the nuclear and mitochondrialgenes of the patient, followed by comparison to the mtDNA content ofage and tissue matched pooled control [21].

Fig. 3. Short TE H-1MR spectroscopy performed through the plane of the basal ganglia, demonstrating a lactate doublet at 1.35 ppm returned from the deep gray nuclei (basal gangliaand thalami) as well as the CSF.
155G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
2.5. PDHC enzyme activity
The activity of PDHC in skeletal muscle was performed at the Centerfor Inherited Disorders of Energy Metabolism (CIDEM) at the Depart-ment of Pediatrics, Rainbow and Children's Hospital, Cleveland, Ohio.PDHC activity was assayed on skeletal muscle by thiamine pyrophos-phate dependent decarboxylation of 1-14C-pyruvate in the presence ofCoASH and NAD+, following activation (dephosphorylation) by pre-incubation with dichloroacetate/fluoride as previously published[22,23].
2.6. Mitochondrial immunofluorescence assay
The pattern of expression of specific respiratory chain complexproteins was determined using standard immunohistochemical meth-ods. The primary antibody cocktails included the followingantibodies torespiratory chain enzyme complex proteins: Complex I (30 kDaNDUFS3, IgG1), Complex II (30 kDa FeS protein, IgG2a), Complex III(core 2, IgG1), Complex IV subunit I (IgG2a), Complex IV subunit IV(IgG2a), Complex V (OSCP, IgG1), and pyruvate dehydrogenase (PDH
E1α, IgG1). Antibodies to porin (anti-porin IgG 2b) served as referencecontrol for mitochondrial density and distribution. The secondaryantibody cocktail included Alexa Fluor 594 goat anti-mouse IgG2bspecific antibody and either Alexa Fluor 488 GAM IgG1 for complexes I,III, V and PDH or Alexa Fluor 488 GAM IgG2a for complexes II and IV. Allprimary antibodies were from Mitosciences, Eugene, Oregon.
2.7. Case report and results
The proband presented at 5 years of age to Texas Children's Hospitalwith a 9-month history of increasing lethargy, fatigue, difficulty inwalking andaweightgain of 20 kg. Prior to this 9-monthperiod, shehadnormal motor and cognitive development. During this time period, sheexhibited a voracious appetite alternating with nausea, vomiting, andnocturnal enuresis. The patient also developed a moon facies, a buffalohump, acne, and hair thinning prompting the diagnostic considerationof Cushing syndrome. Therewas no significant pastmedical history, andno known family history of neurological disease including encephalop-athy, epilepsy, ophthalmoplegia, or cardiomyopathy, although theparentswere not available to answer questions about the family history.

Fig. 4. Immunofluorescence images illustrating variable expression of ETC complexes including regions of normal expression for complexes 1 (A), II (C), III (E), IV-I (G), V (I) and regions ofreduced expression for complexes I (B), II (D), III (F), IV-I (H) and V (J), respectively. [Magnification A, B, G, H ×400; C, D, E, F, I, J ×100.]
156 G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
She was born at full term gestation via cesarean section secondary tofailure to progress with a birth-weight of 2.7 kg to nonconsanguineousparents of Northern European descent. The pregnancy history was onlyremarkable for maternal methamphetamine and nicotine use.
Physical exam revealed a height of 113 cm (75th percentile), aweight of 38.9 kg (N95th percentile) which was increased from 20 kgninemonths prior, and a BMI of 30.5. Her neurological examinationwasnotable for fatigue. The remainder of her neurological examination was

Fig. 5. Immunofluorescence image showing normal expression for PDHA1[magnification×100].
157G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
otherwise unremarkable including normal extraocular movements,deep tendon reflexes and power, and no ataxia.
A low dose dexamethasone suppression test and a TSH level wereobtained as part of the initial diagnostic work-up to evaluate thepatient's physical features and rapid weight gain. The TSH level wasmildly elevated to 10µIU/mL (0.32–5.00). The lowdose dexamethasonesuppression test was abnormal, with a mildly elevated ACTH level of54 pg/mL(6–48),withanAMcortisol level of 13.4 μg/dL (it shouldbeb2if there is a normal response). A repeat low dose dexamethasone testwas obtained 2 months later and was normal, with an AM cortisol levelof 0.7 μg/dL.
MRI of the brain at the time of the initial presentation was withinnormal limits and CT of the abdomen revealed no adrenal masses. Twomonths after the initial evaluation she was noticed to be increasinglylethargic with the appearance of drowsiness. At that time, a repeat MRIof the brain revealed a non-enhancing T2/Flair hyperintense, T1hypointense signal abnormality in the hypothalamus, medial thalami,periaqueductal region, the dorsolateralmedulla, and cerebellar nodules.The presence of these extensive bilateral CNS lesions prompted theconsideration of atypical Leigh syndrome as a working diagnosis(Figs. 1 and 2).
A metabolic diagnostic work-up that included plasma amino acids,urine organic acids, and an acylcarnitine profile was normal. The serumvenous lactate was 2.2 mmol/L (0.2–2.0). Given the brain MRI findingssuggestive of atypical Leigh syndrome, a blood sample was obtained toperform a panel of common mtDNA point mutations and singledeletions. The results of this screen revealed an apparently homo-plasmic mtDNA mutation, m.11778GNA, most commonly associatedwith LHON.
A suspicion of mitochondrial dysfunction as an underlying cause ofher symptoms led to the initiation of carnitine and coenzyme Q10
supplementation. Since the patient improved clinically, she was
Table 1Electron transport chain enzymatic activities — skeletal muscle.
Electron transportchain enzymatic activities
ETC complexes Patient(% of mean)
Control+/−SD⁎
NADH:ferricyanidedehydrogenase
I 272 (114) 239+/−82.5⁎
NADH:cytochrome c reductase I+IIITotal 60.0 (73) 82.2+/−16.2⁎
Rotenone sensitive 31.7 (86) 37.0+/−7.6⁎
Succinate dehydrogenase II 9.34 (129) 7.22+/−1.32⁎
Succinate:cytochromec reductase
II+III 3.43 (66) 5.21+/−2.63⁎
Cytochrome c oxidase IV 15.6 (64) 24.4+/−9.9⁎
Citrate synthase 26.1 (108) 242+/−69⁎
ETC: electron transport chain.⁎ nmol/min/mg protein.
discharged home. One month later, her symptoms worsened, and shepresented with increasing lethargy, tachycardia, fever to 105 °F andshortness of breath. Her respiratory compromise required intubation.Echocardiogram was normal. A positive nasal adenovirus test wasnoted. The serumvenous lactate levelwas0.8 mmol/L (0.2–2.0). Furtherwork-up included a repeat MRI of the brain that showed no significantinterval change in the signal abnormalities that had been previouslyvisualized and a normal MRA of the brain. Brain MR spectroscopyrevealed elevated lactate peaks in the basal ganglia and thalami (Fig. 3).
Although theproband's brainMRI andMRSfindingswere suggestiveof ongoingmitochondrial dysfunction in the central nervous system, onthe other hand, the patient's clinical phenotype did not fit the classicalLHON phenotype caused by the m.11778GNA mutation. Serial ophthal-mological examinations revealed no papilledema, ocular signs ofelevated intracranial pressure or optic nerve atrophy. In order todetermine whether there were additional biochemical and molecularcauses for the observedatypical Leighencephalopathy, a skeletalmusclebiopsy was performed. Results revealed a moderate predominance oftype II fibers, moderately increased neutral lipids, and a mild tomoderate increase in the number of mitochondria. No abnormalities ofthe cristae or paracrystalline inclusionswere observed. Focal to regionalreduced immunostaining for complexes I, II, III, IV (subunits COI andCOII) and V was also observed (Figs. 4 and 5). However, deficiencies inthe mitochondrial electron transport chain enzyme activities were notnoted (Table 1). To establish if othermtDNAmutations or variants werepresent, the whole mitochondrial genome from skeletal muscle wassequenced. The results confirmed the presence of the apparentlyhomoplasmic m.11778GNA mutation and haplogroup H in this patient.However, additional deleterious mtDNA mutations were not detected(Table 2). In addition, real-time quantitative PCRwas performed on themuscle sample to estimate mtDNA copy number and was found to be84% of the mean value for age and tissue matched controls, ruling outmtDNA depletion. Array Comparative Genomic Hybridization (aCGH)(Baylor version 8.0) did not detect any copy number variants in nuclearencoded genes. Sequencing of the genes encoding the E1 alpha subunitof thepyruvate dehydrogenase complex (PDHA1), pyruvate carboxylase(PC), DNA polymerase gamma (POLG1) and surfeit locus protein 1(SURF1) did not show any deleterious mutations. In addition, sequenceanalysis of a panel of genes encoding complex I proteins includingNDUFS2, NDUFS8, NDUFS7, NDUFV1, NDUFS3, NDUFS6, NUDFS4, NDU-FAF2, NDUFA1, NDUFAF4, NDUFA7, and NDUFS5 did not detectdeleterious mutations either. Pyruvate dehydrogenase complex(PDHC) activity was measured in the muscle biopsy sample, and was50% of the mean of the control values, a value in the low normal rangebut not consistent with PDHC deficiency.
To determine if the patient's asymptomatic mother also carried them.11778GNA mutation, real-time Allele Refractory Mutation System(ARMS) qPCR was performed on her blood sample. These studiesshowed that the patient's mother's blood is 56% heteroplasmic forthe m.11778GNA mutation. Additional matrilineal relatives were notavailable for testing.
3. Discussion
Leber hereditary optic neuropathy (LHON) is a degenerative eyedisease characterized by bilateral, painless, subacute visual failure thatoccurs usually in young adult life. There is variable penetrance, andmales are more likely to be affected than females [24,25]. Affectedsubjects are typically asymptomatic until visual blurring developsaffecting the central visual field in one eye, with similar symptomsappearing in the other eye about eight weeks later. The vast majority ofcases of LHON fall under one of four mutations in the genes encodingdifferent protein subunits of the mitochondrial NADH dehydrogenase(complex I of respiratory chain): m.3460GNA (p.A52T) in the MTND1genes, m.11778GNA (p.R340H) in the MTND4 gene, m.14484TNC

Table 2mtDNA polymorphisms detected in this patient using whole mitochondrial genome sequencing on a skeletal muscle sample.
Nucleotide number Gene Base change Codon Amino acid change Reported in MITOMAP Base pair ratios as reported in mtDB
73 HV2 ANG Reported mtDB, 309:1555263 HV2 ANG Reported mtDB, 6:1861303 OH, CSB2 insC Reported N/A311 OH, CSB2 insC Reported N/A750 MTRNR1 (12S rRNA) ANG Reported mtDB, 22:26821438 MTRNR1 (12S rRNA) ANG Reported mtDB, 84:26203010 MTRNR2 (16S rRNA) GNA Reported mtDB, 2154:5504769 ND2 ANG ATANATC M100M Reported mtDB, 30:26748860 ATP6 ANG ACANGCA T112A Reported mtDB, 6:269815326 CYTB ANG ACANGCA T194A Reported mtDB, 17:268716162 HV1, d-loop ANG Reported mtDB, 1834:3116519 D-loop TNC Reported mtDB, 1115:752
HV: Hypervariable Region; D-loop: displacement loop 2; OH: Origin of Heavy Strand; CSB2: Conserved Sequence Block 2.mtDB: Human Mitochondrial Genome Database.
158 G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
(p.M64V) in theMTND6 gene, andm.14459GNA (p.A72V) in theMTND6gene [3].
In addition to optic atrophy, additional abnormalities includingmovement disorders, peripheral neuropathy, brainstem involve-ment and sensorineural hearing loss have been observed in subjectsaffected with LHON [4,26]. Clinical variability has been reported notonly among the common LHON mutations, but among individualswith the same LHONmutation and within the same family (Table 3),[27,28]. For example, the m.14459GNA mutation was reported in 3siblings: an asymptomatic male, one male showing isolated LHON,and one female with isolated spasticity/dystonia and no signs ofoptic neuropathy [27]. However, precise measurements of the de-gree of heteroplasmy were not described in that study. In anotherfamily with multiple individuals homoplasmic for the m.14459GNAmutation, the major clinical presentations were spasticity, hemipar-esis, dystonia, and dysarthria. Some individuals were completelyasymptomatic, and no family members manifested LHON at the timeof assessment [28].
Sensorineural hearing loss (SNHL) has also been reported withcommon LHON mutations with and without additional clinicalphenotypes [29,30]. Ceranic et al. reported two patients with LHONcaused by the m.11778GNA mutation with progressive auditory neu-ropathy [30]. One of the patients, a 59 year old male, presented withprogressive and significant bilateral hearing loss in addition to LHON.The second patient, a 45 year old female, also had severe optic atrophyyet had amore subtle SNHL and also presentedwith amultiple-sclerosislike illness. Recently, Kato et al. screened blood samples from 373unrelated Japanese patients with SNHL for 29 common mtDNA muta-tions, which included the 4 common LHONmutations [29]. One of thesepatients, a 28 year old female, carried the m.11778GNA mutation andhad not yet developed any visual difficulties. The patient's mother, twosisters, and two nieces all suffered from SNHL with no optic atrophy aswell, and presumably carried them.1178GNAmutation, although it wasnot reported [29].
Harding et al. described 8 female patients with LHON caused by them.11778GNAmutation, 6 of whom had clinical and brain MRI findingsindistinguishable from multiple sclerosis [4]. Thus far, less than 20subjects with LHON and multiple sclerosis-like (MSL) symptoms havebeen reported. Eighteen of these individuals are either homoplasmic,or have levels of heteroplasmy above 95%, for the m.11778GNAmutation [31]. Interestingly, unlike the classical LHON phenotype, theratio of males to females with LHON-MSL is approximately 1:2. Theearliest age of presentation of these individuals is 17 years, with amajority becoming affected in their late 20 s to early 30 s. Moreover,the onset of neurological features often occurs years after the opticneuritis first presents.
A few cases of Leigh disease associated with some primary LHONmutations have been reported. Kirby et al. reported 3 patients withLeigh disease and the m.14459GNA LHON mutation [32]. All three
individuals (twomale siblings andoneunrelated female)first presentedat less than one year of age with lactic acidosis, developmentalregression, and symptoms of brainstemdysfunction. One of the patientsdied of respiratory arrest at age 6 years 11 months, the other twodied ofdisease related complications prior to the age of 1 year. No evidence orfamily history of optic neuropathy or dystonia was noted. Funalot et al.reported three male subjects with Leigh-like encephalopathy, each ofwhich had visual loss followed by neurological regression and bilateralbrainstem lesions on MRI [33]. One patient suffered from vision loss atthe age of 4 years, another at the age of 18 years, and the third at age34 years. All three developed neurological symptoms in their early 30 s.Sequence analysis indicated that two of the affected individuals were~70% and ~40% heteroplasmic for the m.14484TNC and m.14459GNAmutations respectively, the third carried a homoplasmic m.3460GNAmutation [33]. The m.3460GNA mutation has also been reported in a23 year old male who suffered from severe visual impairment followedshortly by thedevelopmentof palatalmyoclonusandvertical gazepalsy,and the appearance of brainstem lesions observed by MRI [34].Interestingly, the patient's mother and grandmother were alsodiagnosed with LHON due to vision loss, yet never suffered the centralnervous system defects observed in the patient. An additional malepatient harboring the m.3460GNA mutation with LHON plus spasticataxia with dystonia, and periventricular multiple sclerosis-like whitematter lesions has also been reported [35].
McFarland et al. [40] reported the case of a 22 year old woman whoinitially presentedwith signs and symptoms of atypical Leigh syndromeat 17 months of age. These symptoms primarily included progressivedystonia involving the trunk, face, and all four limbs. Her 19 year oldbrother presentedwith similar signs and symptomsat3 years of age andeventually became wheelchair dependent. Brain MRI scans demon-strated bilateral symmetric putaminal necrosis in both individuals. Them.11778ANG mutation was detected at 86% heteroplasmy in theproband's muscle sample, and at 87% heteroplasmy in a blood sampleof her brother. Interestingly, the m.11778ANG mutation was alsodetected at 85% heteroplasmy in a blood sample of their asymptomaticmother. Both individuals exhibited an atypical clinical course for Leighdisease, surviving well into adulthood and completing school, with thebrother going on to college.
The case herein presented is atypical because of the clinicalpresentation, age, and gender of the patient, in conjunction with thepresence of them.11778ANGmutation. Theprobandcurrently at 6 yearsof age exhibits a normal cognitive status. In general, subjects harboringprimarymtDNAmutations associatedwith LHON become symptomaticat a later age. In addition, the penetrance for thesemutations is lower infemales and in particular for the m.11778GNA mutation (9–11%). Theproband's mother also harbors the m.11778GNA mutation at 56%heteroplasmy in blood, yet at the age of 44 years is currentlyasymptomatic. Moreover, based on the results of the real-time qPCRand ETC analyses, mtDNA depletion has been ruled out. Other known

Table 3Atypical clinical features of patients with primary LHON mutations described in this manuscript.
Family–individual Gender Mutation Ophthalmologic abnormalities Extra-ophthalmologic findings Reference
1–1 M m.14459GNA No None [27]1–2 M m.14459GNA LHON None [27]1–3 F m.14459GNA No Spasticity, dystonia [27]2–1 F m.14459GNA No None [28]2–2 F m.14459GNA No Hemiparesis, lactic acidemia [28]2–3 F m.14459GNA No Stroke, dystonia, developmental delay, spasticity,
hearing loss, lactic acidemia[28]
2–4 M m.14459GNA No Global delay, dysarthria, spasticity, lactic acidemia [28]2–5 M m.14459GNA No None [28]3–1 M m.14459GNA No Leigh disease [32]3–2 M m.14459GNA No Leigh disease [32]4 F m.14459GNA No Leigh disease [32]5 M m.14459GNA LHON SNHL Leigh-like encephalopathy [33]6 M m.14484TNC LHON Leigh-like encephalopathy [33]7 M m.11778GNA LHON Hearing loss [30]8 F m.11778GNA LHON Hearing loss MS-like illness [30]9 F m.11778GNA No SNHL [29]10…15 All F m.11778GNA LHON MS-like illness [4]16 M m.11778GNA LHON MS-like illness [31]17–1 F m.11778GNA Left eye Keratoconus Leigh syndrome [40]17–2 M m.11778GNA No Leigh syndrome [40]18 M m.3460GNA LHON Atypical Leigh syndrome [33]
MS: multiple sclerosis; SNHL: sensorineural hearing loss.
159G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
possible causes of Leigh syndrome such as defects in PDHC activity andmutations in POLG1, PDHA, PC, SURF1, and nuclear genes encodingcomplex I proteins have also been excluded in our proband.
Immunohistochemistry demonstrated variable focal to regionalreduced expression of mitochondrial proteins for complexes I, II, III, IVand V. This reduction reflects a “global” depression of the expressionof themajor ETC complexes, albeit in a subset of fibers. The proportionof affected fibers was not large enough to reduce the overall bio-chemical activity of ETC complexes below 30% of the mean activity.Since biochemical assays are often based on non-selective tissuehomogenates, they are therefore unlikely to detect reduced proteinexpression affecting only a small subset of fibers in a tissue.
Since a causative effect of the m.11778ANG mutation cannot becompletely proven in our patient, modifying factors such as mitochon-drial haplotype, nuclear modifier genes, and possible environmentalfactors causing atypical Leigh syndrome in this patient may be possible.A synergistic role ofmitochondrial variants depending on the haplotypeof the individual has previously been proposed in cases of LHON withand without additional clinical phenotypes [5,36]. Studies of two HanChinese families have suggested that m.4435GNA (tRNA Met) andm.15951GNA (tRNA Thr) may increase the penetrance of LHON causedby the m.11778GNA mutation by decreasing the expression of tRNAsmethionine and threonine respectively, leading to altered mitochon-drial tRNA metabolism and increased mitochondrial dysfunction[37,38]. As previously noted, the mitochondrial haplotype J, whichincludes secondary LHONmutations 4216TNC and 13708GNA, increasesthe risk of visual failure [5]. In contrast, mitochondrial haplotype H hasbeen associated with a decreased risk of visual failure [5]. These datasuggest that our patient, who has haplotype H, may actually be at adecreased risk of developing optic atrophy. In the case of the siblingswith atypical Leigh syndrome [40], the presence of m.4716CNG (whichchanges glutamine to glutamic acid at amino acid position 83) waspostulated to influence the pathogenicity of them.11778GNAmutation.However, no other notable variants, including m.4716CNG, wereobserved in our patient's mitochondrial whole genome (Table 2).
Because LHON mutations are more penetrant among men thanwomen, anX-linked susceptibility gene has been hypothesized. Hudsonet al. identified a region of the X chromosome (Xp21–Xq21) thatcontains a high risk haplotype (markers DXS8090 and DXS106) whichmay contain a nuclear modifier gene [39]. Males homoplasmic for them.11778ANG and 14484TNC mutations and hemizygous for this highrisk haplotype on the X chromosomeweremuchmore likely to develop
visual impairment than those not carrying the haplotype. Females withthe m.11778ANG mutation who were homozygous for the high riskhaplotype were also much more likely to develop visual impairmentthan heterozygous females. However, the presence of an X-linkedsusceptibility locus does not exclude the possibility of autosomal factorshaving an influence on disease, although to date no such factors havebeen identified.
In summary, our report adds further evidence to the existing body ofliterature that atypical Leigh syndrome may also be associated withprimarymtDNAmutations typically associatedwith a LHONphenotype.However, we cannot definitively rule out an as yet unknown nuclearmodifier gene interactingwith them.11778ANGmutation leading to anatypical Leigh encephalopathy. Further casesmayneed tobe analyzed inorder to establish the causality of this association.
References
[1] D.R. Thorburn, H.H. Dahl, Mitochondrial disorders: genetics, counseling, prenataldiagnosis and reproductive options, Am. J. Med. Genet. 106 (2001) 102–114.
[2] G. Singh, M.T. Lott, D.C. Wallace, A mitochondrial DNA mutation as a cause ofLeber's hereditary optic neuropathy, N. Engl. J. Med. 320 (1989) 1300–1305.
[3] D.A. Mackey, R.J. Oostra, T. Rosenberg, E. Nikoskelainen, J. Bronte-Stewart, J.Poulton, A.E. Harding, G. Govan, P.A. Bolhuis, S. Norby, Primary pathogenic mtDNAmutations in multigeneration pedigrees with Leber hereditary optic neuropathy,Am. J. Hum. Genet. 59 (1996) 481–485.
[4] A.E. Harding, M.G. Sweeney, D.H. Miller, C.J. Mumford, H. Kellar-Wood, D. Menard,W.I. McDonald, D.A. Compston, Occurrence of a multiple sclerosis-like illness inwomen who have a Leber's hereditary optic neuropathy mitochondrial DNAmutation, Brain 115 (Pt 4) (1992) 979–989.
[5] G. Hudson, V. Carelli, L. Spruijt, M. Gerards, C. Mowbray, A. Achilli, A. Pyle, J. Elson,N. Howell, C. La Morgia, M.L. Valentino, K. Huoponen, M.L. Savontaus, E.Nikoskelainen, A.A. Sadun, S.R. Salomao, R. Belfort Jr., P. Griffiths, P.Y. Man, R.F.de Coo, R. Horvath, M. Zeviani, H.J. Smeets, A. Torroni, P.F. Chinnery, Clinicalexpression of Leber hereditary optic neuropathy is affected by the mitochondrialDNA-haplogroup background, Am. J. Hum. Genet. 81 (2007) 228–233.
[6] D.C. Wallace, M.T. Lott, M.D. Brown, K. Kerstann, Mitochondria and neuroophthal-mologic diseases, in: C.R. Scriver, A.L. Beaudet, D. Valle, W.S. Sly (Eds.), TheMetabolic and Molecular Bases of Inherited Disease, McGraw-Hill, New York,2001, pp. 2425–2509.
[7] D. Leigh, Subacute necrotizing encephalomyelopathy in an infant, J. Neurol.Neurosurg. Psychiatry 14 (1951) 216–221.
[8] R.P. Saneto, S.D. Friedman, D.W. Shaw, Neuroimaging of mitochondrial disease,Mitochondrion 8 (2008) 396–413.
[9] J. Finsterer, Leigh and Leigh-like syndrome in children and adults, Pediatr. Neurol.39 (2008) 223–235.
[10] S. Rahman, R.B. Blok, H.H. Dahl, D.M. Danks, D.M. Kirby, C.W. Chow, J.Christodoulou, D.R. Thorburn, Leigh syndrome: clinical features and biochemicaland DNA abnormalities, Ann. Neurol. 39 (1996) 343–351.
[11] A. Brautbar, J. Wang, J.E. Abdenur, R.C. Chang, J.A. Thomas, T.A. Grebe, C. Lim, S.W.Weng, B.H. Graham, L.J. Wong, The mitochondrial 13513GNA mutation is

160 G. Fruhman et al. / Molecular Genetics and Metabolism 103 (2011) 153–160
associated with Leigh disease phenotypes independent of complex I deficiency inmuscle, Mol. Genet. Metab. 94 (2008) 485–490.
[12] L.J. Wong, D. Senadheera, Direct detection of multiple point mutations inmitochondrial DNA, Clin. Chem. 43 (1997) 1857–1861.
[13] S. Shanske, L.J. Wong, Molecular analysis for mitochondrial DNA disorders,Mitochondrion 4 (2004) 403–415.
[14] R.K. Bai, L.J. Wong, Detection and quantification of heteroplasmic mutantmitochondrial DNA by real-time amplification refractory mutation systemquantitative PCR analysis: a single-step approach, Clin. Chem. 50 (2004)996–1001.
[15] J. Wang, A. Brautbar, A.K. Chan, T. Dzwiniel, F.Y. Li, P.J. Waters, B.H. Graham, L.J.Wong, Two mtDNAmutations 14487TNC (M63V, ND6) and 12297TNC (tRNA Leu)in a Leigh syndrome family, Mol. Genet. Metab. 96 (2009) 59–65.
[16] S.M.Ware, N. El-Hassan, S.G. Kahler, Q. Zhang, Y.W.Ma, E.Miller, B.Wong, R.L. Spicer,W.J. Craigen, B.A. Kozel, D.K. Grange, L.J.Wong, Infantile cardiomyopathy causedby amutation in the overlapping region of mitochondrial ATPase 6 and 8 genes, J. Med.Genet. 46 (2009) 308–314.
[17] D.M. Kirby, D.R. Thorburn, D.M. Turnbull, R.W. Taylor, Biochemical assays ofrespiratory chain complex activity, Methods Cell Biol. 80 (2007) 93–119.
[18] Y.Ma, R.K. Bai, R. Trieu, L.J.Wong,Mitochondrial dysfunction in human breast cancercells and their transmitochondrial cybrids, Biochim. Biophys. Acta 1797 (2010)29–37.
[19] L.J. Wong, N. Brunetti-Pierri, Q. Zhang, N. Yazigi, K.E. Bove, B.B. Dahms, M.A.Puchowicz, I. Gonzalez-Gomez, E.S. Schmitt, C.K. Truong, C.L. Hoppel, P.C. Chou, J.Wang, E.E. Baldwin, D. Adams, N. Leslie, R.G. Boles, D.S. Kerr, W.J. Craigen, Mutationsin the MPV17 gene are responsible for rapidly progressive liver failure in infancy,Hepatology 46 (2007) 1218–1227.
[20] R.K. Bai, L.J. Wong, Simultaneous detection and quantification of mitochondrialDNA deletion(s), depletion, and over-replication in patients with mitochondrialdisease, J. Mol. Diagn. 7 (2005) 613–622.
[21] D. Dimmock, L.Y. Tang, E.S. Schmitt, L.J. Wong, Quantitative evaluation of themitochondrial DNA depletion syndrome, Clin. Chem. 56 (2010) 1119–1127.
[22] K.F. Sheu, C.W. Hu, M.F. Utter, Pyruvate dehydrogenase complex activity innormal and deficient fibroblasts, J. Clin. Invest. 67 (1981) 1463–1471.
[23] D.S. Kerr, L. Ho, C.M. Berlin, K.F. Lanoue, J. Towfighi, C.L. Hoppel, M.M. Lusk, C.M.Gondek, M.S. Patel, Systemic deficiency of the first component of the pyruvatedehydrogenase complex, Pediatr. Res. 22 (1987) 312–318.
[24] N.J. Newman, M.T. Lott, D.C. Wallace, The clinical characteristics of pedigrees ofLeber's hereditary optic neuropathy with the 11778 mutation, Am. J. Ophthalmol.111 (1991) 750–762.
[25] D.R. Johns, K.L. Heher, N.R. Miller, K.H. Smith, Leber's hereditary optic neuropathy.Clinical manifestations of the 14484 mutation, Arch. Ophthalmol. 111 (1993)495–498.
[26] E.K. Nikoskelainen, R.J. Marttila, K. Huoponen, V. Juvonen, T. Lamminen, P.Sonninen, M.L. Savontaus, Leber's “plus”: neurological abnormalities in patientswith Leber's hereditary optic neuropathy, J. Neurol. Neurosurg. Psychiatry 59(1995) 160–164.
[27] M.A. Tarnopolsky, S.K. Baker, T. Myint, C.E. Maxner, J. Robitaille, B.H. Robinson,Clinical variability in maternally inherited Leber hereditary optic neuropathy withthe G14459A mutation, Am. J. Med. Genet. A 124A (2004) 372–376.
[28] A. Gropman, T.J. Chen, C.L. Perng, D. Krasnewich, E. Chernoff, C. Tifft, L.J. Wong,Variable clinical manifestation of homoplasmic G14459A mitochondrial DNAmutation, Am. J. Med. Genet. A 124A (2004) 377–382.
[29] T. Kato, Y. Nishigaki, Y. Noguchi, H. Ueno, H. Hosoya, T. Ito, Y. Kimura, K. Kitamura, M.Tanaka, Extensive and rapid screening formajormitochondrial DNApointmutationsin patients with hereditary hearing loss, J. Hum. Genet. 55 (2010) 147–154.
[30] B. Ceranic, L.M. Luxon, Progressive auditory neuropathy in patients with Leber'shereditary optic neuropathy, J. Neurol. Neurosurg. Psychiatry 75 (2004) 626–630.
[31] F. Perez, O. Anne, S. Debruxelles, P. Menegon, V. Lambrecq, D. Lacombe, M.L. Martin-Negrier, B. Brochet, C. Goizet, Leber's optic neuropathy associatedwith disseminatedwhite matter disease: a case report and review, Clin. Neurol. Neurosurg. 111 (2009)83–86.
[32] D.M. Kirby, S.G. Kahler,M.L. Freckmann, D. Reddihough, D.R. Thorburn, Leigh diseasecaused by the mitochondrial DNA G14459A mutation in unrelated families, Ann.Neurol. 48 (2000) 102–104.
[33] B. Funalot, P. Reynier, A. Vighetto, D. Ranoux, J.P. Bonnefont, C. Godinot, Y.Malthiery, J.L. Mas, Leigh-like encephalopathy complicating Leber's hereditaryoptic neuropathy, Ann. Neurol. 52 (2002) 374–377.
[34] W. Paulus, A. Straube, W. Bauer, A.E. Harding, Central nervous systeminvolvement in Leber's optic neuropathy, J. Neurol. 240 (1993) 251–253.
[35] F.M. Meire, R. Van Coster, P. Cochaux, B. Obermaier-Kusser, C. Candaele, J.J. Martin,Neurological disorders in members of families with Leber's hereditary opticneuropathy (LHON) caused by different mitochondrial mutations, OphthalmicGenet. 16 (1995) 119–126.
[36] C. LaMorgia, A.Achilli, L. Iommarini, P. Barboni,M. Pala, A.Olivieri, C. Zanna, S.Vidoni,C. Tonon, R. Lodi, R. Vetrugno, B. Mostacci, R. Liguori, R. Carroccia, P. Montagna, M.Rugolo, A. Torroni, V. Carelli, Rare mtDNA variants in Leber hereditary opticneuropathy families with recurrence of myoclonus, Neurology 70 (2008) 762–770.
[37] J. Qu, R. Li, X. Zhou, Y. Tong, F. Lu, Y. Qian, Y. Hu, J.Q. Mo, C.E. West, M.X. Guan, Thenovel A4435G mutation in the mitochondrial tRNAMet may modulate thephenotypic expression of the LHON-associated ND4 G11778A mutation, Invest.Ophthalmol. Vis. Sci. 47 (2006) 475–483.
[38] R. Li, J. Qu, X. Zhou, Y. Tong, Y. Hu, Y. Qian, F. Lu, J.Q. Mo, C.E. West, M.X. Guan, Themitochondrial tRNA(Thr) A15951G mutation may influence the phenotypicexpression of the LHON-associated ND4 G11778A mutation in a Chinese family,Gene 376 (2006) 79–86.
[39] G. Hudson, S. Keers, P. Yu Wai Man, P. Griffiths, K. Huoponen, M.L. Savontaus, E.Nikoskelainen, M. Zeviani, F. Carrara, R. Horvath, V. Karcagi, L. Spruijt, I.F. de Coo, H.J.Smeets, P.F. Chinnery, Identification of an X-chromosomal locus and haplotypemodulating the phenotype of a mitochondrial DNA disorder, Am. J. Hum. Genet. 77(2005) 1086–1091.
[40] R. McFarland, P.F. Chinnery, E.L. Blakely, A.M. Schaefer, A.A. Morris, S.M. Foster, H.A.Tuppen, V. Ramesh, P.J. Dorman, D.M. Turnbull, R.W. Taylor, Homoplasmy,heteroplasmy, and mitochondrial dystonia, Neurology 69 (2007) 911–916.




















