
Assessment of the impact of heavy metal pollution from a ferro-nickel smelting plant using...
12
Ecotoxicology and Environmental Safety 66 (2007) 232–243 Assessment of the impact of heavy metal pollution from a ferro-nickel smelting plant using biomarkers Catherine Tsangaris , Evangelos Papathanasiou, Efthimia Cotou Hellenic Centre for Marine Research, 46.7 klm, Athinon-Souniou Ave., P.O. Box 712, 19013 Anavyssos, Greece Received 13 September 2005; received in revised form 9 March 2006; accepted 25 March 2006 Available online 6 June 2006 Abstract A set of biomarkers was used to assess the impact of heavy metal pollution by a ferro-nickel smelting plant in Larymna bay (North Evoikos Gulf, Greece). These included a biomarker reflecting health status of an organism (scope for growth, SFG), a cellular biomarker of heavy metal exposure (composition of metal-containing granules), and two biochemical biomarkers reflecting oxidative stress (glutathione peroxidase, GPX) and neurotoxicity (acetylcholinesterase, AChE) measured in mussels (Mytilus galloprovincialis) both native and transplanted for 1 and 6 months at the coastal area of Larymna. All biomarkers in mussels at Larymna revealed differences from mussels at a reference site, signaling effects of the increased heavy metal levels on the biota. While effects on SFG and GPX in Larymna mussels were obvious on short-term exposure and persistent during chronic exposure, only chronic exposure induced a possibly cumulative effect on AChE. To validate the causal relationship between heavy metal exposure and effects observed in Larymna, SFG, GPX, and ACHE were examined in mussels exposed to a mixture of heavy metals (Ni, Cr, and Fe) under controlled laboratory conditions. The laboratory experiment verified the causal relationship between SFG and GPX responses and heavy metals but this was not demonstrated for AChE. Results from field-collected and laboratory-exposed mussels indicated a potential of GPX as predictive biomarker of population-level effects of heavy metal exposure. r 2006 Elsevier Inc. All rights reserved. Keywords: Biomarker; Metals; Pollution effects; Scope for growth; Glutathione peroxidase; Acetylcholinesterase; X-ray microanalysis; Mytilus galloprovincialis 1. Introduction Responses to pollution at various levels of biological organization are widely used in environmental monitoring studies since, by definition, pollution implies hazard to living resources. Measuring changes at the community and population levels is ecologically relevant but may provide only indirect assumptions with regard to the causes of ecosystem-level effects. On the other hand biomarkers represent changes from the molecular to the organism level that can be related to toxic effects of and/or exposure to chemical contaminants. In addition, biomarker responses occur prior to alterations at the population and community levels so they can be predictive and anticipatory. Thus biomarkers have the ability to diagnose causes and may act as early warning signals of ecosystem-level damage. Biochemical and cellular biomarkers may indicate expo- sure to specific classes of contaminants (Bucheli and Kent, 1995; Viarengo et al., 1999; Marigo´mez et al., 2002) or reflect specific toxicity mechanisms (Livingstone, 2001; Fulton and Key, 2001; Bolognesi et al., 2004) but, to signal effects at the population and the community levels, they should be related to a degree of impairment of growth, reproductive output, or metabolic function signifying degradation in the health of the organism (Depledge et al., 1995). Hence linking biochemical and cellular biomarkers with biomarkers of the overall health of individual organisms is important for the evaluation of their utility in predicting population-level effects (Allen and Moore, 2004). The small bay of Larymna located in North Evoikos Gulf Greece, is contaminated with heavy metals by a ferro- nickel smelting plant situated on the coast within the bay. The plant extracts nickel from laterite ore and dumps the ARTICLE IN PRESS www.elsevier.com/locate/ecoenv 0147-6513/$ - see front matter r 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.ecoenv.2006.03.011 Corresponding author. Fax: +30 2 2910 76 347.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Assessment of the impact of heavy metal pollution from a ferro-nickel smelting plant using...

ARTICLE IN PRESS
0147-6513/$ - se
doi:10.1016/j.ec
�Correspond
Ecotoxicology and Environmental Safety 66 (2007) 232–243
www.elsevier.com/locate/ecoenv
Assessment of the impact of heavy metal pollution from a ferro-nickelsmelting plant using biomarkers
Catherine Tsangaris�, Evangelos Papathanasiou, Efthimia Cotou
Hellenic Centre for Marine Research, 46.7 klm, Athinon-Souniou Ave., P.O. Box 712, 19013 Anavyssos, Greece
Received 13 September 2005; received in revised form 9 March 2006; accepted 25 March 2006
Available online 6 June 2006
Abstract
A set of biomarkers was used to assess the impact of heavy metal pollution by a ferro-nickel smelting plant in Larymna bay (North
Evoikos Gulf, Greece). These included a biomarker reflecting health status of an organism (scope for growth, SFG), a cellular biomarker
of heavy metal exposure (composition of metal-containing granules), and two biochemical biomarkers reflecting oxidative stress
(glutathione peroxidase, GPX) and neurotoxicity (acetylcholinesterase, AChE) measured in mussels (Mytilus galloprovincialis) both
native and transplanted for 1 and 6 months at the coastal area of Larymna. All biomarkers in mussels at Larymna revealed differences
from mussels at a reference site, signaling effects of the increased heavy metal levels on the biota. While effects on SFG and GPX in
Larymna mussels were obvious on short-term exposure and persistent during chronic exposure, only chronic exposure induced a possibly
cumulative effect on AChE. To validate the causal relationship between heavy metal exposure and effects observed in Larymna, SFG,
GPX, and ACHE were examined in mussels exposed to a mixture of heavy metals (Ni, Cr, and Fe) under controlled laboratory
conditions. The laboratory experiment verified the causal relationship between SFG and GPX responses and heavy metals but this was
not demonstrated for AChE. Results from field-collected and laboratory-exposed mussels indicated a potential of GPX as predictive
biomarker of population-level effects of heavy metal exposure.
r 2006 Elsevier Inc. All rights reserved.
Keywords: Biomarker; Metals; Pollution effects; Scope for growth; Glutathione peroxidase; Acetylcholinesterase; X-ray microanalysis; Mytilus
galloprovincialis
1. Introduction
Responses to pollution at various levels of biologicalorganization are widely used in environmental monitoringstudies since, by definition, pollution implies hazard toliving resources. Measuring changes at the community andpopulation levels is ecologically relevant but may provideonly indirect assumptions with regard to the causes ofecosystem-level effects. On the other hand biomarkersrepresent changes from the molecular to the organism levelthat can be related to toxic effects of and/or exposure tochemical contaminants. In addition, biomarker responsesoccur prior to alterations at the population and communitylevels so they can be predictive and anticipatory. Thusbiomarkers have the ability to diagnose causes and may actas early warning signals of ecosystem-level damage.
e front matter r 2006 Elsevier Inc. All rights reserved.
oenv.2006.03.011
ing author. Fax: +30 2 2910 76 347.
Biochemical and cellular biomarkers may indicate expo-sure to specific classes of contaminants (Bucheli and Kent,1995; Viarengo et al., 1999; Marigomez et al., 2002) orreflect specific toxicity mechanisms (Livingstone, 2001;Fulton and Key, 2001; Bolognesi et al., 2004) but, to signaleffects at the population and the community levels, theyshould be related to a degree of impairment of growth,reproductive output, or metabolic function signifyingdegradation in the health of the organism (Depledgeet al., 1995). Hence linking biochemical and cellularbiomarkers with biomarkers of the overall health ofindividual organisms is important for the evaluation oftheir utility in predicting population-level effects (Allen andMoore, 2004).The small bay of Larymna located in North Evoikos
Gulf Greece, is contaminated with heavy metals by a ferro-nickel smelting plant situated on the coast within the bay.The plant extracts nickel from laterite ore and dumps the

ARTICLE IN PRESS
Fig. 1. Location of the sampling sites. Larymna, the metal-contaminated
site in Evoikos Gulf is marked by a black arrow and the reference site in
Saronikos Gulf by a white arrow.
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 233
metalliferous slag at a dumping site in the North EvoikosGulf in the vicinity of Larymna. Spillages of dust, coolingwater, and slag also contribute to the contamination of thebay with heavy metals. Elevated concentrations mainly ofNi, Cr, and Fe but also of Co and Mn have been recordedin sediments, sea grass, and benthic organisms of thecoastal zone (Nicolaidou and Nott, 1989, 1990) andsediments at the dumping site (Voutsinou-Taliadouriet al., 1995). Nicolaidou et al. (1989) have previouslyreported changes at the community level which wereattributed to the indirect effects of dumping of metallifer-ous slag, causing a change in the particle size compositionof the sediment, but could not be related to toxic effects ofheavy metals in the slag. Biomarkers that can be related totoxic effects of and/or exposure to chemical contaminantshave not previously been applied in Larymna.
Mussels are commonly used as sentinel organisms forbiomarker measurements due to their wide distribution,sedentary state, ease of collection, transplantation, andmaintenance in the laboratory, and ability to accumulatehigh concentrations of a wide range of contaminantsincluding heavy metals (Viarengo and Canesi, 1991).
In the present study, an established biomarker reflectingthe overall health of an organism (scope for growth SFG);and two biochemical biomarkers reflecting differenttoxicity mechanisms, namely oxidative stress (glutathioneperoxidase GPX); and neurotoxicity (acetylcholinesteraseAChE); were measured in mussels (Mytilus galloprovincia-
lis) native and transplanted at the coastal area of Larymna.The same biomarkers were also measured in musselsexposed to a mixture of Ni, Cr, and Fe under controlledlaboratory conditions. Additionally, a cellular biomarkerof heavy metal exposure (composition of metal-containinggranules) was studied in native mussels from Larymna.Scope for growth, a physiological condition index, is ameasure of the energy balance of an organism, widely usedin environmental monitoring studies (Widdows and Don-kin, 1992; Widdows et al., 1995a, b, 2002). Glutathioneperoxidase is an enzyme of the antioxidant defense systemagainst reactive oxygen species (ROS) that can be producedby a wide range of contaminants including organicxenobiotics and heavy metals (Lemaire and Livingstone,1993). Variations in antioxidant enzyme activities havebeen proposed as biomarkers of oxidative stress (Living-stone, 2001). Acetylcholinesterase is an enzyme of neuro-transmission and its inhibition reflects neurotoxicity.AChE inhibition has been considered a biomarker ofexposure to organophosphate and carbamate pesticides(Bocquene and Galgani, 1991) but recent studies suggestthat other contaminants including heavy metals can inhibitAChE (Payne et al., 1996; Najimi et al., 1997; Guilherminoet al., 1998; Hamza-Chaffai et al., 1998; Elumalai et al.,2002). Lysosomal and inorganic granules accumulateheavy metals as part of the cellular detoxification process(Marigomez et al., 2002). X-ray microanalysis of thesegranules provides information on the bioavailable fractionof heavy metals and has been proposed as a biomarker of
heavy metal exposure (Vesk and Byrne, 1999; Correia,et al., 2002; Marigomez et al., 2002).The aim of the present study was to assess the impact of
heavy metal pollution in the coastal area of Larymna usingmultiple level biomarkers in M. galloprovincialis and to linkbiochemical biomarker responses to the health status ofindividual organisms to assess whether these biochemicalresponses show potential as predictive biomarkers ofpopulation-level effects of heavy metal exposure.
2. Materials and methods
2.1. Source of animals—field sampling
Native mussels from Larymna (North Evoikos Gulf) and farmed
mussels from Megara (Saronikos Gulf) transplanted to the same area in
Larymna and to a reference site at Agios Kosmas (Saronikos Gulf) were
used (Fig. 1). The Agios Kosmas site is considered as an area of good
environmental conditions and heavy metal concentrations in surface
sediments are low (Cotou et al., 2002; Karageorgis and Hatzianestis,
2003). Both Larymna and Agios Kosmas are areas of oligotrophic
character.
Farmed mussels of similar shell length were purchased in spring, placed
in plastic cages (approximately 100 individuals), and immersed in water to
a depth of 1–2m at the coastal areas of Larymna and the reference site
where they remained for 6 months. Additionally, farmed mussels were
purchased in autumn and transplanted as above to the Larymna site
were they remained for 1 month. Native, ‘‘1-month’’, and ‘‘6-month’’
transplanted mussel populations were collected during autumn and were
transported to the laboratory under moist/cool conditions for measure-
ments of SFG, GPX, AChE, and heavy metal concentrations in whole
tissues. Native mussels from Larymna were also collected in the winter and
transported to the University of Plymouth and Plymouth Marine
Laboratory for X-ray microanalysis.
2.2. Laboratory experiment
Farmed mussels of similar shell length were purchased from Megara in
autumn. These were placed in 2-L tanks (5mussels/L) and were acclimated
in aerated synthetic seawater for 7 days prior to heavy metal exposure.
The mussels were exposed to a mixture of Ni, Cr, and Fe in two

ARTICLE IN PRESS
123.5 J/mg ash-free dry weight is the energy content of phytoplankton
cells (Widdows et al., 1979).20.456 J/mmol O2 is the energetic equivalent of respiratory oxygen
consumption (Gnaiger, 1983).
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243234
combinations of concentrations for 4 weeks. The low (L) level of
concentrations comprised 20mg Ni/L, 0.5 mg Cr/L, and 400mg Fe/L and
the high (H) level of concentrations comprised 200mg Ni/L, 5 mg Cr/L, and4000mg Fe/L. The low level exposure concentrations were chosen as being
representative of the highly dissolved Ni, Cr, and Fe levels reported in
Larymna (22.6mg Ni/L, 0.48mg Cr(VI)/L, 434mg Fe/L, Nakopoulou et al.,
1985) and in highly contaminated estuaries and coastal waters (1–16mgNi/L: Cambell et al., 1988, 0.52–0.79mg Cr(VI)/L; Dahab and Al-Madfa,
1997, 170–890mg Fe/L, Chan et al., 1974). Heavy metals were added to
artificial seawater as solutions of NiCl2, NaCr2O4, and FeCl2. Both H and
L level concentrations and the control (artificial seawater with no added
metal) were used in triplicate. The seawater was renewed every 2nd day
and the mussels were fed daily with Dunaniella tertiolecta (60� 106 cells/
mussel). After 4 weeks of heavy metal exposure, SFG, GPX, and AChE
activities and heavy metal Ni, Cr, and Fe concentrations in whole mussels
were measured.
2.3. Heavy metal analysis
Whole mussel tissues were dissected, freeze-dried, and digested with
concentrated HNO3 in Teflon digestion vials under pressure at 120 1C for
12 h (UNEP, 1984). Ni, Cr, and Fe analyses were performed by atomic
absorption spectrophotometry on a Varian Spectra AA 20plus spectro-
photometer (Varian, Victoria, Australia). Heavy metal concentrations
were expressed asmg/g dry weight tissue. Analytical quality control was
achieved using certified reference material provided by the National
Research Council of Canada.
2.4. Body condition index
The body condition index was calculated according to Bayne and
Widdows (1978) as dry whole tissue weight (mg) versus shell length (mm)
per individual.
2.5. Scope for growth measurements
The procedure described by Widdows and Salked (1992) with the
modification of Cotou et al. (2002) was used to measure SFG. Clearance
rate, respiration rate, and food absorption efficiency were measured for
each mussel individually under standardized laboratory conditions. Prior
to all measurements mussels were left undisturbed in the chambers for 12 h
to recover from transportation and resume feeding. For clearance rate and
absorption efficiency measurements, mussels were placed in individual
chambers in synthetic seawater filtered through 1 and 3-mm activated
carbon cartridge filters in a closed circulated system at a flow rate of
160–180mL/min. The unicellular algae D. tertiolecta was added in the
chambers at a concentration of 6500–7000 cells/mL. Subsequently algal
cell concentrations were measured from the outfall of all chambers
including the controls (two chambers without mussels) four times at 45-
min intervals using a Z1 Coulter Counter (Coulter Electronics, Luton,
UK) calibrated to count particles above 3 mm.
Clearance rate (Cl) was calculated as
Cl ¼ FlðC0 �C1Þ=C0,
where Fl is flow rate, C0 is inflow concentration (control chamber), and C1
is outflow concentration (experimental chamber).
Feces were collected from the chambers after the mussels remained in
the system for 12 h. Feces produced during the initial 12-h recovery period
were discarded. Absorption efficiency (AE) was measured by the ratio of
Conover (1966):
AE ¼ ðF � EÞ=ðð1� EÞ � F Þ,
where F is ash-free dry mass/dry mass of food and E is ash-free dry mass/
dry mass of feces.
Respiration rate was measured for each mussel in a transparent plastic
respirometer (i.e., modified Quickfit chamber) containing 750mL of air-
saturated synthetic seawater circulated with a magnetic stirrer. Each
mussel was settled in the respirometer for 15min and subsequently the
decline in oxygen tension was monitored over a period of 30–45min using
a Strathkelvin Model 781 oxygen meter (Strathkelvin Instruments,
Glasgow, UK). Physiological rates were converted to energy equivalents
and were used to calculate SFG, the energy available for growth and
reproduction, according to the equation
SFG ¼ A� R ¼ ðC � absorption efficiencyÞ � R,
where A is energy absorbed from food, C is energy consumed from food,
and R is energy lost via respiration.
Energy lost via excretion was not included in the above equation
because it comprised less than 5% of the total energy loss.
Calculation of C, A and R were as follows:
C (J/g/h) ¼ clearance rate (L/g/h)� algae cells concentration (mg ash-
free dry weight/L)� 23.5 J/mg ash free dry weight,1
A (J/g/h) ¼ C� absorption efficiency, and
R (J/g/h) ¼ respiration rate (mmol O2/g/h)� 0.456 J/mmol O2.2
2.6. Determination of enzyme activities
2.6.1. Glutathione peroxidase activity
Samples were prepared as described by Livingstone et al. (1992).
Digestive glands from four to six individuals were pooled and
homogenized using a Potter–Elvehjem homogenizer (Heidolph Electro
GmbH, Kelheim, Germany) in 1:4 (w:v) 20mM Tris–HCl buffer, pH 7.6,
containing 1mM EDTA, 1mM dithiothietrol, 0.5M sucrose, and 0.15M
KCl. Homogenates were centrifuged at 500g for 15min and 10,000g for
30min. All preparation procedures were carried out at 4 1C. GPX activity
was assayed by the rate of NADPH oxidation at 340 nm by the coupled
reaction with glutathione reductase using H2O2 as a substrate
(GSSG+NADPH+H+-2GSH+NADP+, NADPH extinction
coefficient ¼ 6.2mM�1/cm) (Wendel, 1980). Just before the assay, a
reagent containing 48mM sodium phosphate buffer, pH 7, 0.38mM
EDTA, 0.12mM NADPH, 0.95mM sodium azide, 3.2 units glutathione
reductase, and 1mM GSH was prepared; 1mL of this reagent and 17mLof sample supernatant were added in a cuvette and the reaction was
initiated by addition of 17mL H2O2 solution. Nonenzymic rate was
measured in the absence of sample and subtracted from total rate. The
assay was carried out at 25 1C. GPX activity was expressed as nmol
NADPH consumed per min per mg protein.
2.6.2. Acetylcholinesterase activity
Samples were prepared as described by Bocquene and Galgani (1992).
Adductor muscles from four to six individuals were pooled and
homogenized using a Potter–Elvehjem homogenizer in 1:5 (w:v) 0.1M
Tris–HCl buffer, pH 7. Homogenates were centrifuged at 15,000g for
20min. All preparation procedures were carried out at 4 1C. AChE activity
was assayed at 412 nm by the method of Ellman et al. (1961); 1.5mL of
0.1M Tris–HCl buffer, pH 7, 100mL of 0.01M 5,50-dithiobis(2-
nitrobenzoic acid)and 50 mL of sample supernatant were added in a
cuvette and the reaction was initiated by addition of 50 mL of 0.075M
acetylthiocholine. The assay was carried out at 25 1C. AChE activity was
expressed as nmol of acetylthiocholine hydrolyzed per min per mg protein.
2.6.3. Total protein
The total protein content in the homogenate supernatants was
measured by the Bradford method (1976). Protein concentration was
calculated using a bovine serum albumin standard curve.

ARTICLE IN PRESSC. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 235
2.7. X-ray microanalysis of granules
Analytical microscopy was applied to kidneys and digestive glands of
mussels. Tissues were dissected out, cut in to small pieces, and immediately
fixed by cryofixation to minimize loss of elements during chemical fixation
procedures (Nott, 1991).
The following techniques were used: (a) metal mirror cryofixation on a
Reichert MM80E (Leica, Milton Keynes, UK) followed by freeze
substitution on a Reichert AFS (Leica) and embedding in Lowcril
HM20 after UV polimerization; (b) cryofixation by plunging samples into
liquid nitrogen followed by freeze-drying and embedding in Spur’s resin.
Thick sections (2mm) were cut dry, placed on Ti grids, carbon-coated,
and examined on a JEOL 200CX (STEM mode) (Jeol Ltd.) at 200KV
equipped with a Link 860 analyzer. X-ray microanalysis was performed on
two types of granules (amorphous granules in lysosomes and spherical
granules). Background spectra of resin and tissue areas were recorded.
Background spectra of Lowcril resin showed Ti and Cu peaks that derived
from the grid and the microscope, respectively. Background spectra of
Spur’s resin revealed Ti, Cu, and Cl peaks.
2.8. Statistical analysis
Data are presented as mean7standard error of the mean. The
Kolmogorov–Smirnoff test and Levene’s test were applied to test normal
distribution and homogeneity of variance, respectively. One-way analysis
of variance (ANOVA) followed by the Tukey HSD multiple comparison
test was applied to determine differences between populations (field data)
or treatments (laboratory data). For SFG data from the field that
exhibited heterogeneity of variance, the Kruskall–Wallis and Tamhanes
multiple comparison tests were performed. To examine relationships
between GPX, AChE, and SFG from combined field and laboratory data,
values were expressed as percentage of reference site or control mussels
and linear regression was applied. Statistical analysis was performed using
the SPSS statistical package. Significance level was set at Po0:05.
3. Results
3.1. Heavy metal concentrations in mussel tissues
Ni, Cr, and Fe concentrations in mussels from Larymnaand the reference site are shown in Table 1. Both nativeand transplanted mussels at Larymna showed significantly
Table 1
Ni, Cr, and Fe concentrations (mg/g dry weight) in whole tissues of mussels (
exposed to a mixture of Ni, Cr, and Fe in the laboratory
Ni (mg/g dw)
Field mussel populations
Larymna native 21.171.3a
Larymna transplanted 6-months 50.378.6a,b
Larymna transplanted 1-month 47.073.8a,b
Reference site 4.970.4
Laboratory exposed mussels
High exposure level (H) 42.572.4a
Low exposure level (L) 8.870.8a
Control 2.570.3
Mean7standard error, n ¼ 10 (field experiment), n ¼ 8 or 9 (laboratory exper
exposure level (H): 200mg Ni/L, 50 mg Cr/L, 4000mg Fe/L.aSignificantly different from reference site or control, Po0.05.bSignificantly different from native mussels in Larymna, Po0.05.
higher levels of Ni (21.1–50.3 mg/g dry weight) and Cr(17.0–25.0 mg/g dry weight) than transplanted mussels atthe reference site (4.9 mg Ni/g dry weight and 0.8 mg Cr/gdry weight). Fe concentrations showed high individualvariability but, though they were elevated in Larymnamussels (824–1082 mg/g dry weight) compared to mussels atthe reference site (280 mg/g dry weight), the differences werenot significant. Among mussel populations in Larymna,significantly higher Ni concentrations were found intransplanted mussels (47.0 and 50.3 mg/g dry weight) thanin native mussels (21.1 mg/g dry weight), whereas there wereno differences in Cr and Fe concentrations.Ni, Cr, and Fe concentrations in mussels exposed to a
mixture of Ni, Cr, and Fe in the laboratory increasedaccording to exposure concentrations and were signifi-cantly different from corresponding concentrations incontrol mussels (Table 1). At the high exposure level,laboratory-treated mussels reached Ni concentrations(42.5 mg/g dry weight) similar to those recorded in thetransplanted mussels at Larymna (47.0 and 50.3 mg/g dryweight). On the contrary, Cr and Fe concentrations inlaboratory-treated mussels, even at the high exposure level,were one order of magnitude lower (2.5 mg Cr/g dry weight,183 mg Fe/g dry weight) than those in native andtransplanted mussels at Larymna (17–25 mg Cr/g dryweight, 824–1082 mg Fe/g dry weight).
3.2. Body condition index
The body condition index was similar in musselpopulations from Larymna and the reference site with theexception of 6-month transplanted mussels in Larymnawhich showed a significantly higher value than all othermussel populations (Table 2).In laboratory-treated mussels similar values of the body
condition index were found among those exposed to themixture of Ni, Cr, and Fe and controls (Table 2).
M. galloprovincialis) from Larymna and the reference site and of mussels
Cr (mg/g dw) Fe (mg/g dw)
20.270.9a 10827138
25.073.2a 8987298
17.072.4a 824747
0.870.3 280731
2.570.0a 183716a
1.470.0a 10578a
0.970.0 5677
iment). Low exposure level (L): 20mg Ni/L, 5mg Cr/L, 400mg Fe/L. High
ARTICLE IN PRESS
Table 2
Body condition index of mussels (M. galloprovincialis) from Larymna and
the reference site and of mussels exposed to a mixture of Ni, Fe, and Cr in
the laboratory
Body condition index
Field mussel populations
Larymna native 6.7670.25
Larymna transplanted 6-months 10.4870.52a
Larymna transplanted 1-month 5.9870.63
Reference site 6.1070.20
Laboratory-exposed mussels
High exposure level (H) 4.5670.58
Low exposure level (L) 4.6570.21
Control 5.3270.65
Mean7standard error, n ¼ 15 (field experiment), n ¼ 4 (laboratory
experiment). Low exposure level (L): 20 mg Ni/L, 5mg Cr/L, 400mg Fe/
L. High exposure level (H): 200mg Ni/L, 50 mg Cr/L, 4000mg Fe/L.aSignificantly different from all mussel populations, Po0.05.
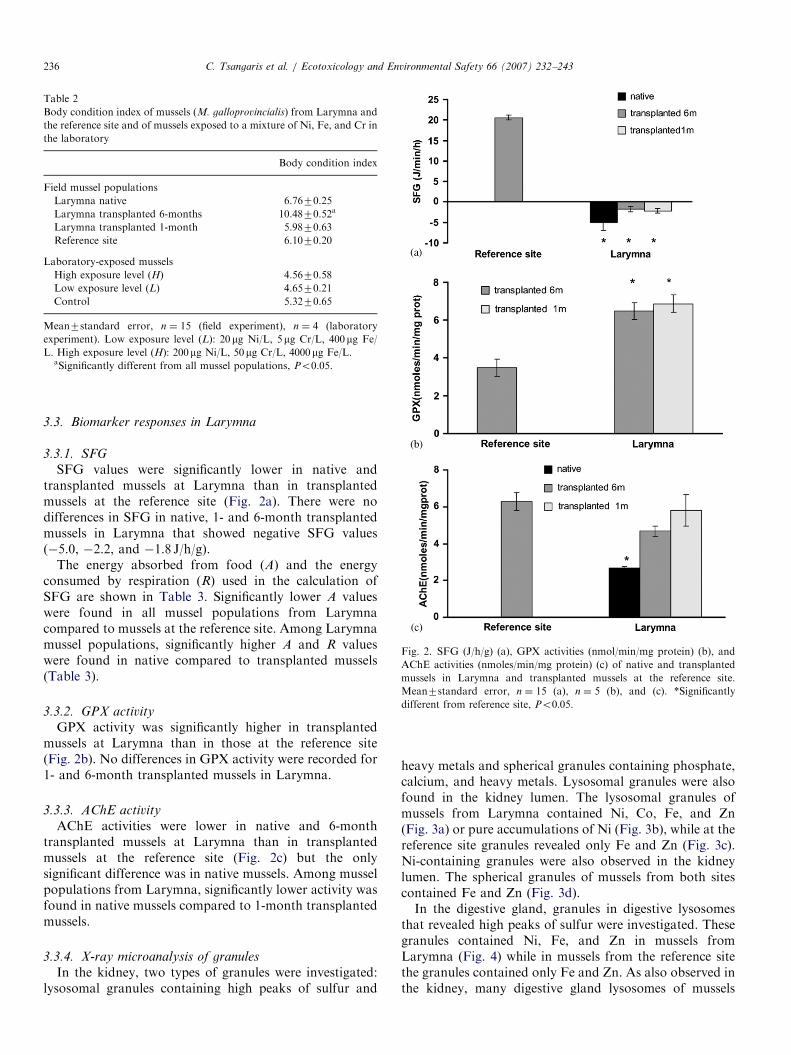
Fig. 2. SFG (J/h/g) (a), GPX activities (nmol/min/mg protein) (b), and
AChE activities (nmoles/min/mg protein) (c) of native and transplanted
mussels in Larymna and transplanted mussels at the reference site.
Mean7standard error, n ¼ 15 (a), n ¼ 5 (b), and (c). *Significantly
different from reference site, Po0:05.
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243236
3.3. Biomarker responses in Larymna
3.3.1. SFG
SFG values were significantly lower in native andtransplanted mussels at Larymna than in transplantedmussels at the reference site (Fig. 2a). There were nodifferences in SFG in native, 1- and 6-month transplantedmussels in Larymna that showed negative SFG values(�5.0, �2.2, and �1.8 J/h/g).
The energy absorbed from food (A) and the energyconsumed by respiration (R) used in the calculation ofSFG are shown in Table 3. Significantly lower A valueswere found in all mussel populations from Larymnacompared to mussels at the reference site. Among Larymnamussel populations, significantly higher A and R valueswere found in native compared to transplanted mussels(Table 3).
3.3.2. GPX activity
GPX activity was significantly higher in transplantedmussels at Larymna than in those at the reference site(Fig. 2b). No differences in GPX activity were recorded for1- and 6-month transplanted mussels in Larymna.
3.3.3. AChE activity
AChE activities were lower in native and 6-monthtransplanted mussels at Larymna than in transplantedmussels at the reference site (Fig. 2c) but the onlysignificant difference was in native mussels. Among musselpopulations from Larymna, significantly lower activity wasfound in native mussels compared to 1-month transplantedmussels.
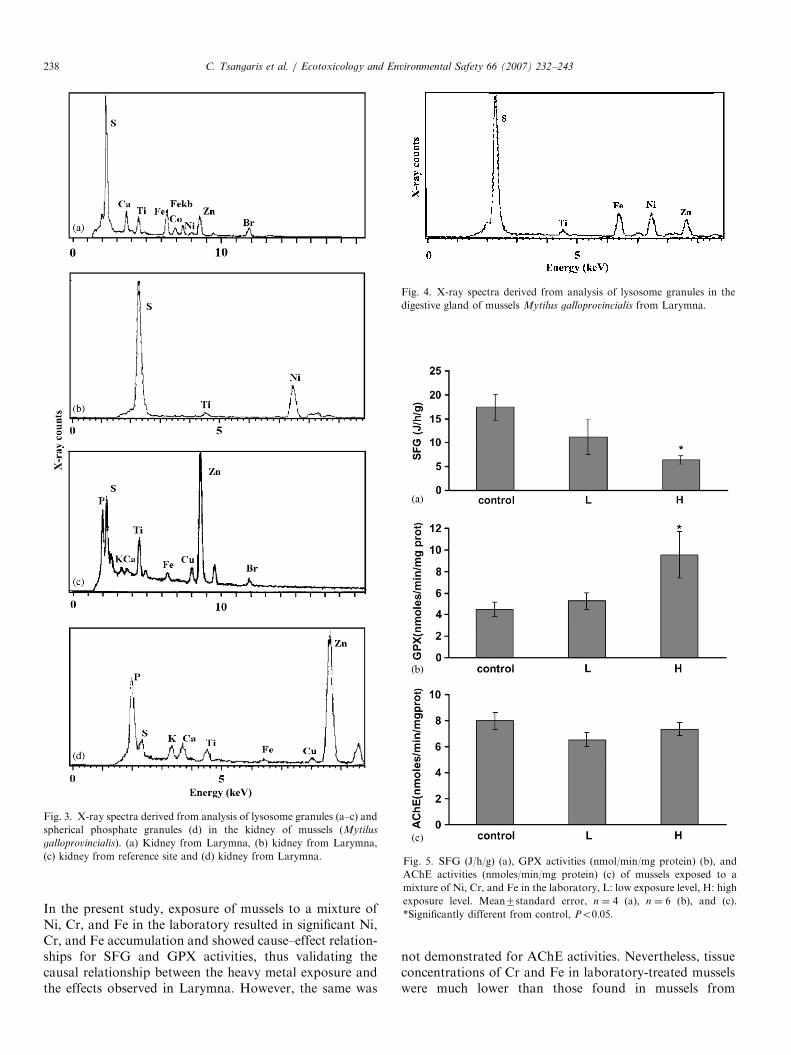
3.3.4. X-ray microanalysis of granules
In the kidney, two types of granules were investigated:lysosomal granules containing high peaks of sulfur and
heavy metals and spherical granules containing phosphate,calcium, and heavy metals. Lysosomal granules were alsofound in the kidney lumen. The lysosomal granules ofmussels from Larymna contained Ni, Co, Fe, and Zn(Fig. 3a) or pure accumulations of Ni (Fig. 3b), while at thereference site granules revealed only Fe and Zn (Fig. 3c).Ni-containing granules were also observed in the kidneylumen. The spherical granules of mussels from both sitescontained Fe and Zn (Fig. 3d).In the digestive gland, granules in digestive lysosomes
that revealed high peaks of sulfur were investigated. Thesegranules contained Ni, Fe, and Zn in mussels fromLarymna (Fig. 4) while in mussels from the reference sitethe granules contained only Fe and Zn. As also observed inthe kidney, many digestive gland lysosomes of mussels

ARTICLE IN PRESS
Table 3
Components of the energy budget (A, R) used for calculation of SFG of mussels (M. galloprovincialis) from Larymna and the reference site and of mussels
exposed to a mixture of Ni, Fe, and Cr in the laboratory
A (J/h/g) R (J/h/g) SFG (J/h/g)
Field mussel populations
Larymna native 10.870.5a 15.871.9 �5.072.1a
Larymna transplanted 6-months 7.870.4a,b 10.070.6b �2.270.6a
Larymna transplanted 1-month 7.570.4a,b 9.270.8b �1.770.6a
Reference site 33.271.3 12.671.8 20.673.1
Laboratory-exposed mussels
High exposure level (H) 19.370.9 12.970.6a 6.470.9a
Low exposure level (L) 22.573.9 11.371.2 11.273.7
Control 28.072.9 10.670.2 17.472.2
Mean7standard error, n ¼ 15 (field experiment), n ¼ 4 (laboratory experiment). Low exposure level (L): 20mg Ni/L, 5mg Cr/L, 400mg Fe/L.
High exposure level (H): 200mg Ni/L, 50mg Cr/L, 4000mg Fe/L.aSignificantly different from reference site or control, Po0.05.bSignificantly different from native mussels in Larymna, Po0.05.
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 237
from the metal-contaminated site contained pure accumu-lations of Ni.
3.4. Biomarker responses on exposure to Ni, Cr, and Fe
mixture in the laboratory
3.4.1. SFG
Mussels exposed to the mixture of Ni, Cr, and Feshowed dose dependent decreases in SFG (Fig. 5a). Thesedecreases were significant at the highest concentration ofmetal mixture where SFG values decreased to 6.4 J/h/gwith regard to the components of the energy budget usedfor calculation of SFG, a gradual decrease in A and agradual increase in R with increasing concentration of themetal mixture was noted, but only the increase in R at thehighest concentration of metal mixture was statisticallysignificant (Table 3).
3.4.2. GPX activity
Mussels exposed to the mixture of Ni, Cr, and Feshowed significant increases in GPX activity compared tocontrol mussels at the highest concentration of metalmixture (Fig. 5b). At the low level of metal concentrationsa slight increase in GPX activity compared to control levelswas noted but this difference was not significant.
3.4.3. AChE activity
There were no significant differences in AChE activitiesamong mussels exposed to the mixture of Ni, Cr, and Feand among controls (Fig. 5c).
3.5. Relationship between biochemical and organism
biomarker responses
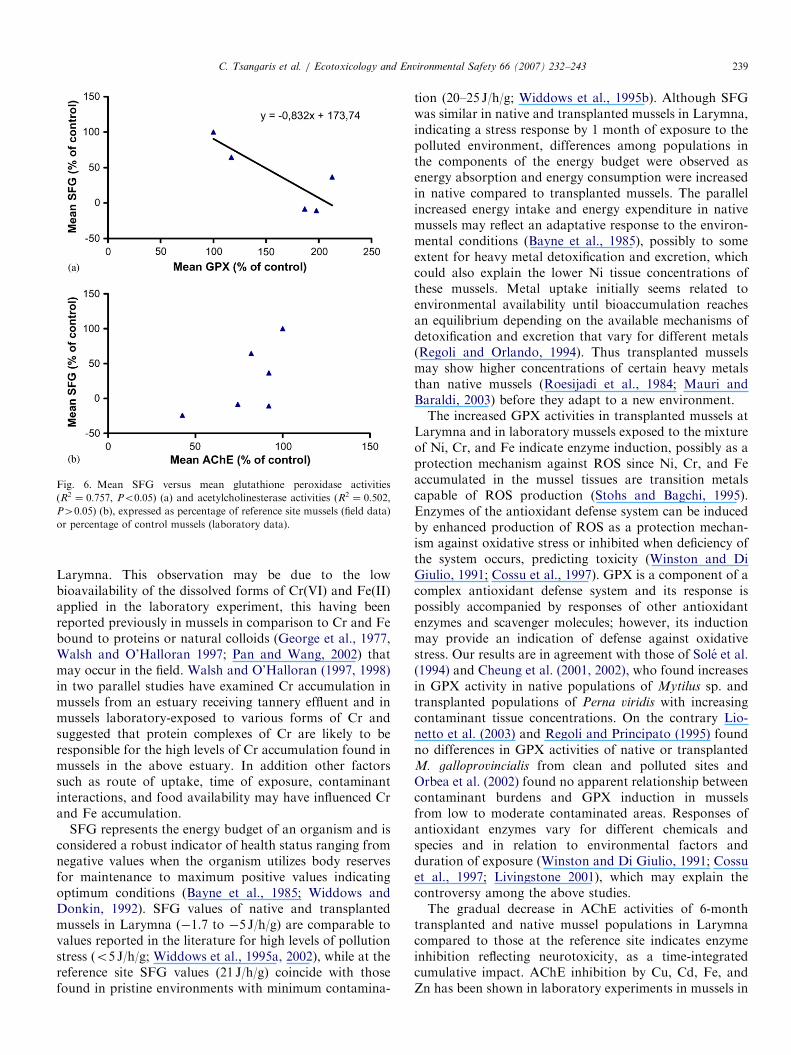
A significant linear regression relationship was foundbetween GPX activities and SFG when data from field-collected and laboratory-treated mussels were combined
and expressed as percentage of reference site or controlmussels (R2 ¼ 0:757, Po0:05, Fig. 6a) whereas there wasno significant relationship between AChE activities andSFG (R2 ¼ 0:502, P40:05, Fig. 6b).
4. Discussion
The use of biomarkers is recognized as an importantapproach for the assessment of pollution, as chemicalanalysis of environmental samples alone does not provideevidence of impacts of contaminants in biota. In this study,results obtained by chemical analysis showed elevated Ni,Cr, and Fe concentrations in mussels from Larymnacompared to mussels from the reference site and to valuesreported in Mytilus sp. from other metal-contaminatedareas (Giusti et al., 1999; Szefer et al., 2004). Accordingly,the biomarkers applied in mussels from Larymna werealtered compared to mussels from the reference site,signaling effects of the increased heavy metal concentra-tions at the organism, cellular, and biochemical levels.The parallel use of native and transplanted mussel
populations can provide information on the time courseeffects of pollution, as responses in native mussels willreflect time-integrated cumulative impacts, while in trans-planted mussels responses signal effects of recent exposure(Bolognesi et al., 2004). Our results showed that recent andchronic exposure of mussels to the polluted conditions inLarymna induced persistent SFG and GPX responses whileonly chronic exposure induced an effect on AChE.Laboratory studies can help establish cause and effect
relationships that can be used to verify and predict effectson natural populations in the field; however, it has to betaken into account that abiotic and biotic factors in thefield will modify contaminant accumulation and biomarkerresponses (Philips, 1976; Sheehan and Power, 1999; Vidalet al., 2002; Serafim et al., 2002; Reid and MacFarlane,2003; Luedeking and Koehler, 2004; Khessiba et al., 2005).
ARTICLE IN PRESS
Fig. 3. X-ray spectra derived from analysis of lysosome granules (a–c) and
spherical phosphate granules (d) in the kidney of mussels (Mytilus
galloprovincialis). (a) Kidney from Larymna, (b) kidney from Larymna,
(c) kidney from reference site and (d) kidney from Larymna.
Fig. 4. X-ray spectra derived from analysis of lysosome granules in the
digestive gland of mussels Mytilus galloprovincialis from Larymna.
Fig. 5. SFG (J/h/g) (a), GPX activities (nmol/min/mg protein) (b), and
AChE activities (nmoles/min/mg protein) (c) of mussels exposed to a
mixture of Ni, Cr, and Fe in the laboratory, L: low exposure level, H: high
exposure level. Mean7standard error, n ¼ 4 (a), n ¼ 6 (b), and (c).
*Significantly different from control, Po0:05.
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243238
In the present study, exposure of mussels to a mixture ofNi, Cr, and Fe in the laboratory resulted in significant Ni,Cr, and Fe accumulation and showed cause–effect relation-ships for SFG and GPX activities, thus validating thecausal relationship between the heavy metal exposure andthe effects observed in Larymna. However, the same was
not demonstrated for AChE activities. Nevertheless, tissueconcentrations of Cr and Fe in laboratory-treated musselswere much lower than those found in mussels from
ARTICLE IN PRESS
Fig. 6. Mean SFG versus mean glutathione peroxidase activities
(R2 ¼ 0:757, Po0:05) (a) and acetylcholinesterase activities (R2 ¼ 0:502,P40:05) (b), expressed as percentage of reference site mussels (field data)
or percentage of control mussels (laboratory data).
C. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 239
Larymna. This observation may be due to the lowbioavailability of the dissolved forms of Cr(VI) and Fe(II)applied in the laboratory experiment, this having beenreported previously in mussels in comparison to Cr and Febound to proteins or natural colloids (George et al., 1977,Walsh and O’Halloran 1997; Pan and Wang, 2002) thatmay occur in the field. Walsh and O’Halloran (1997, 1998)in two parallel studies have examined Cr accumulation inmussels from an estuary receiving tannery effluent and inmussels laboratory-exposed to various forms of Cr andsuggested that protein complexes of Cr are likely to beresponsible for the high levels of Cr accumulation found inmussels in the above estuary. In addition other factorssuch as route of uptake, time of exposure, contaminantinteractions, and food availability may have influenced Crand Fe accumulation.
SFG represents the energy budget of an organism and isconsidered a robust indicator of health status ranging fromnegative values when the organism utilizes body reservesfor maintenance to maximum positive values indicatingoptimum conditions (Bayne et al., 1985; Widdows andDonkin, 1992). SFG values of native and transplantedmussels in Larymna (�1.7 to �5 J/h/g) are comparable tovalues reported in the literature for high levels of pollutionstress (o5 J/h/g; Widdows et al., 1995a, 2002), while at thereference site SFG values (21 J/h/g) coincide with thosefound in pristine environments with minimum contamina-
tion (20–25 J/h/g; Widdows et al., 1995b). Although SFGwas similar in native and transplanted mussels in Larymna,indicating a stress response by 1 month of exposure to thepolluted environment, differences among populations inthe components of the energy budget were observed asenergy absorption and energy consumption were increasedin native compared to transplanted mussels. The parallelincreased energy intake and energy expenditure in nativemussels may reflect an adaptative response to the environ-mental conditions (Bayne et al., 1985), possibly to someextent for heavy metal detoxification and excretion, whichcould also explain the lower Ni tissue concentrations ofthese mussels. Metal uptake initially seems related toenvironmental availability until bioaccumulation reachesan equilibrium depending on the available mechanisms ofdetoxification and excretion that vary for different metals(Regoli and Orlando, 1994). Thus transplanted musselsmay show higher concentrations of certain heavy metalsthan native mussels (Roesijadi et al., 1984; Mauri andBaraldi, 2003) before they adapt to a new environment.The increased GPX activities in transplanted mussels at
Larymna and in laboratory mussels exposed to the mixtureof Ni, Cr, and Fe indicate enzyme induction, possibly as aprotection mechanism against ROS since Ni, Cr, and Feaccumulated in the mussel tissues are transition metalscapable of ROS production (Stohs and Bagchi, 1995).Enzymes of the antioxidant defense system can be inducedby enhanced production of ROS as a protection mechan-ism against oxidative stress or inhibited when deficiency ofthe system occurs, predicting toxicity (Winston and DiGiulio, 1991; Cossu et al., 1997). GPX is a component of acomplex antioxidant defense system and its response ispossibly accompanied by responses of other antioxidantenzymes and scavenger molecules; however, its inductionmay provide an indication of defense against oxidativestress. Our results are in agreement with those of Sole et al.(1994) and Cheung et al. (2001, 2002), who found increasesin GPX activity in native populations of Mytilus sp. andtransplanted populations of Perna viridis with increasingcontaminant tissue concentrations. On the contrary Lio-netto et al. (2003) and Regoli and Principato (1995) foundno differences in GPX activities of native or transplantedM. galloprovincialis from clean and polluted sites andOrbea et al. (2002) found no apparent relationship betweencontaminant burdens and GPX induction in musselsfrom low to moderate contaminated areas. Responses ofantioxidant enzymes vary for different chemicals andspecies and in relation to environmental factors andduration of exposure (Winston and Di Giulio, 1991; Cossuet al., 1997; Livingstone 2001), which may explain thecontroversy among the above studies.The gradual decrease in AChE activities of 6-month
transplanted and native mussel populations in Larymnacompared to those at the reference site indicates enzymeinhibition reflecting neurotoxicity, as a time-integratedcumulative impact. AChE inhibition by Cu, Cd, Fe, andZn has been shown in laboratory experiments in mussels in

ARTICLE IN PRESSC. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243240
vitro (Najimi et al., 1997) and in clams in vivo (Hamza-Chaffai et al., 1998) while certain field studies with Mytilus
sp. suggest that a decrease in AChE activity may be anindication of metal pollution (Amiard-Triquet et al., 1998;Dellali et al., 2001). Yet no evidence of AChE inhibition inmussels after exposure to the mixture of Ni, Cr, and Fe inthe laboratory was found, which may be related to theshort duration of exposure, the lower Cr and Fe tissueconcentrations in these mussels compared to Larymnamussels which showed AChE inhibition, or the presence ofother contaminants and contaminant interactions in thefield. The sampling area in Larymna receives domestic andurban runoff wastes from the neighboring town that maycontain contaminants capable of inhibiting AchE, forexample extracts of used engine oil (Payne et al., 1996) anddetergents (Guilhermino et al., 1998). Consequently,laboratory results could not verify whether the AChEinhibition in Larymna is associated with elevated levels ofNi, Cr, and Fe and this thus remains a subject for furtherresearch.
Abiotic and biotic factors such as temperature, salinity,food supply, age, and reproductive status influence heavymetal accumulation and biomarker responses and thushave to be considered in pollution assessment studies(Philips, 1976; Sheehan and Power, 1999; Vidal et al., 2002;Serafim et al., 2002; Khessiba et al., 2005). Both theLarymna site and the study reference site are oligotrophicareas and, during the samplings in autumn, differencesin temperature and salinity were limited. The bodycondition index, which is influenced primarily by thereproductive cycle and food availability, was similar amongthe mussel populations from both areas with the exceptionof 6-month transplanted mussels in Larymna. These hadvisibly larger gonads that nevertheless did not resultin a marked dilution effect on heavy metal whole-tissueconcentrations.
X-ray microanalysis of heavy-metal-containing granuleswas performed only in native mussels from Larymna andreference mussels collected during the winter. Thereforeseasonal variations in biotic and abiotic factors precludedirect comparisons between the physiological and bio-chemical biomarker responses and the composition ofheavy-metal-containing granules. Nevertheless, X-ray mi-cronalysis results contribute to the elucidation of cellulardetoxification mechanisms in mussels chronically exposedto heavy metals in Larymna. At the cellular level,compartmentalization of heavy metals in lysosomes andtheir accumulation in inorganic granules are detoxificationmechanisms that isolate the toxic metals to prevent toxicinteractions with cellular constituents (Nott, 1993). Subse-quently metal-containing granules can be excreted by thekidney cells (Marigomez et al., 2002). In the present study,the high accumulations of Ni found in lysosome granulesof the kidney and the digestive gland in native mussels fromLarymna imply that the elevated Ni levels in the tissuesresulted in an effort to achieve Ni detoxification in thesegranules and possibly Ni excretion, as Ni-containing
granules were observed in the kidney lumen. Similarly,traces of Co previously reported at increased environ-mental levels in the area of Larymna (Voutsinou-Talia-douri et al., 1995) were also detected in lysosome kidneygranules of Larymna mussels. Fe and Zn were found inlysosome and spherical granules of mussels both fromLarymna and from the reference site, as also previouslyreported in M. edulis from a clean reference site by Georgeand Pirie (1980), implying a regulation capacity for thesemetals. On the contrary, Cr was not detected in intracel-lular granules of Larymna mussels, suggesting lowconcentration of Cr in the granules (below the detectionlimit of the method, 0.1%) and/or different cellularlocalization of Cr. Previous studies on M. edulis exposedto Cr have located Cr in kidney and digestive glandlysosome granules (Chassard-Bouchaud et al., 1989; Walshand O’Halloran, 1998), in kidney extracellular sphericalcrystals (Chassard-Bouchaud et al., 1989), and in thecytosol bound to proteins (Walsh and O’Halloran, 1998),however at higher Cr exposure or tissue concentrationsthan those in the present study. Consequently, X-raymicroanalysis results of the present study suggest that thecomposition of lysosome granules was indicative ofexposure to elevated levels of Ni and to a lesser extent ofCo but could not provide information on Fe and Crexposure.It is generally considered that pollution effects are
manifested initially at the biochemical level and subse-quently at the organismal level, leading to effects inpopulations, and thus biochemical biomarkers are sug-gested to exhibit predictive capacities of population-leveleffects. The ability of biochemical markers to provide earlywarning signals of ecosystem-level damage can be evalu-ated by their correlation with ecologically relevant bio-markers such as SFG (Sanders et al., 1991; Allen andMoore, 2004). In the present study significant relationshipsbetween GPX and SFG responses and exposure to heavymetals in the environment of Larymna and underlaboratory conditions were found, suggesting that thedecline in the overall health of the organisms may be due tosome extent to ROS production by heavy metals. Incontrast, AChE inhibition did not parallel the SFG decline.This was evident in mussels after short-term exposure toheavy metals both in the field and in the laboratory, whereit was found that when SFG decreased there was nosignificant AChE inhibition. Accordingly Rickwood andGalloway (2004) found no measurable AChE inhibition inmussels exposed in the laboratory to an organophosphatepesticide when biomarkers of physiological status, cellularintegrity, and immunotoxicity were impacted. Recently theuse of AChE inhibition as a biomarker of adverse effecthas been questioned as previous studies in fish indicate thatthe degree of inhibition is not readily correlated withtoxicological effects (Chambers et al., 2002). Our resultssuggest that GPX but not AChE shows potential as apredictive biomarker of population-level effects of heavymetal exposure.

ARTICLE IN PRESSC. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 241
5. Conclusions
This study applied biomarker measurements in Larym-na, an area impacted by ferro-nickel smelting operations,and revealed effects at the organism (SFG), cellular(composition of granules), and biochemical (GPX, AChE)levels. SFG and GPX responses reflecting general healthstatus and oxidative stress, respectively, were moresensitive to the polluted conditions in Larymna. Effectson these two biomarkers were obvious on short-termexposure and persistent during chronic exposure, whileonly chronic exposure induced a possibly cumulative effecton AChE indicative of neurotoxicity. Laboratory experi-ments verified the causal relationship between GPX andSFG responses and heavy metals but this was notdemonstrated for AChE. The relationship between GPXand SFG responses on exposure to heavy metals suggests apotential of GPX as predictive biomarker of population-level effects of heavy metal exposure.
Acknowledgments
The authors thank R. Moate at the University ofPlymouth and L. Mavin at Plymouth Marine Laboratoryfor assistance in preparation of samples for X-raymicroanalysis, J.A. Nott at Plymouth Marine Laboratoryfor X-ray microanalysis, and C. Edge for English correc-tion of the manuscript. We are grateful to the anonymousreferees for their helpful comments on this manuscript.This work was partially supported by an ERASMUSgrant.
References
Allen, J.I., Moore, M.N., 2004. Environmental prognostics: Is the current
use of biomarkers appropriate for environmental risk evaluation? Mar.
Environ. Res. 58, 227–232.
Amiard-Triquet, C., Altmann, S., Amiard, J.C., Ballan-Dufranc-ais, C.,
Baumard, P., Budzinski, H., Crouzet, C., Garrigues, P., His, E.,
Jeantet, A.Y., Menasria, R., Mora, P., Mouneyrac, C., Narbonne,
J.F., Pavillon, J.F., 1998. Fate and effects of micropollutants in the
Gironde estuary, France: a multidisciplinary approach. Hydrobiologia
373/374, 259–279.
Bayne, B.L., Widdows, J., 1978. The physiological ecology of two
populations of Mytilus edulis L. Oecologia 37, 137–162.
Bayne, B.L., Brown, D.A., Burns, K., Dixon, D.R., Ivanovici, A.,
Livingstone, D.R., Lowe, D.M., Moore, M.N., Stebbing, A.R.D.,
Widdows, J., 1985. The Effects of Stress and Pollution on Marine
Animals. Praeger Press, New York.
Bocquene, G., Galgani, F., 1991. L’acetylcholinesterase chez les orga-
nismes marins, outil de surveillance des effets des pesticides
organophosphores et carbamates. Oceanis 17, 439–448.
Bocquene, G., Galgani, F., 1992. Assessment of biological water quality
using acetylcholinesterase (AChE) inhibition measurement. In Report
of the FAO/IOC/UNEP Training Workshop on the Techniques for
Monitoring Biological Effects of Pollutants in Marine Organisms,
pp. 133–139.
Bolognesi, C., Fenzilli, G., Lasagna, C., Perrone, E., Roggieri, P., 2004.
Genotoxicity biomarkers in Mytilus galloprovincialis: wild versus caged
mussels. Mutat. Res. 552, 153–162.
Bradford, M., 1976. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding. Anal. Biochem. 772, 248–264.
Bucheli, T.D., Kent, K., 1995. Induction of cytochrome P450 as a
biomarker for environmental contamination in aquatic ecosystems.
Crit. Rev. Environ. Sci. Technol. 25, 201–268.
Cambell, J.A., Whitelaw, K., Riley, J.P., Head, P.C., Jones, P.D., 1988.
Contrasting behaviour of dissolved and particulate nickel and zinc in a
polluted estuary. Sci. Total Environ. 71, 141–155.
Chan, J.P., Cheung, M.T., Lau, F.P., 1974. Trace metals in Hong Kong
waters. Mar. Pollut. Bull. 5, 171–174.
Chambers, J.E., Boone, J.S., Carr, R.L., Chambers, H.W., Straus, D.L.,
2002. Biomarkers as predictors in health and ecological risk assess-
ment. Hum. Ecol. Risk Assess. 8, 165–176.
Chassard-Bouchaud, C., Boutin, J.F., Hallegot, B., Galle, P., 1989.
Chromium uptake, distribution and loss in the mussel Mytilus edulis: a
structural, ultrastructural and microanalytical study. Dis. Aquat.
Organ. 7, 117–136.
Cheung, C.C.C., Zheng, G.J., Li, A.M.Y., Richardson, B.J., Lam, P.K.S.,
2001. Relationships between tissue concentrations of polycyclic
aromatic hydrocarbons and antioxidant responses of marine mussels,
Perna viridis. Aquat. Toxicol. 52, 189–203.
Cheung, C.C.C., Zheng, G.J., Lam, P.K.S., Richardson, B.J., 2002.
Relationships between tissue concentrations of chlorinated hydro-
carbons (polychlorinated biphenyls and chlorinated pesticides) and
antioxidative responses of marine mussels, Perna viridis. Mar. Pollut.
Bull. 45, 181–191.
Conover, R.J., 1966. Assimilation of organic matter by zooplankton.
Limnol. Oceanogr. 11, 338–354.
Correia, A.D., Costa, M.H., Ryan, K.P., Nott, J.A., 2002. Studies
on biomarkers of copper exposure and toxicity in the marine
amphipod Gammarus locusta (Crustacea): I. Copper-containing
granules within the midgut gland. J. Mar. Biol. Assoc. UK 82,
827–834.
Cossu, C., Doyotte, A., Jacquin, M.C., Babut, M., Exinger, A., Vasseur,
P., 1997. Glutathione reductase, selenium-dependent glutathione
peroxidase, glutathione levels and lipid peroxidation in freshwater
bivalves, Unio tumidus, as biomarkers of aquatic contamination in field
studies. Ecotoxicol. Environ. Saf. 38, 122–131.
Cotou, E., Papathanassiou, E., Tsangaris, C., 2002. Assessing the quality
of marine coastal environments: comparison of scope for growth and
Microtoxs bioassay results of pollution gradient areas in eastern
Mediterranean (Greece). Environ. Pollut. 119, 141–149.
Dahab, O.A., Al-Madfa, H., 1997. Chromium distribution in waters and
sediments of the eastern side of the Qatari Peninsula. Sci. Total
Environ. 196, 1–11.
Dellali, M., Gnassia Barelli, M., Romeo, M., Aissa, P., 2001. The use of
acetylcholinesterase activity in Ruditapes decussates and Mytilus
galloprovincialis in the biomonitoring of Bizerta lagoon. Comp.
Biochem. Physiol. Part C 130, 227–235.
Depledge, M.H., Aagard, A., Gyorkos, P., 1995. Assessment of trace
metal toxicity using molecular, physiological and behavioural biomar-
kers. Mar. Pollut. Bull. 31, 19–27.
Ellman, G.L., Courtney, K.D., Andres Jr., V., Featherstone, R.M., 1961.
A new and rapid colorimeric determination of acetylcholinesterase
activity. Biochem. Pharmacol. 7, 88–95.
Elumalai, M., Antunes, C., Guilhermino, L., 2002. Single metals and their
mixtures on selected enzymes of Carcinus maenas. Water Air Soil
Pollut. 141, 273–280.
Fulton, M.H., Key, P.B., 2001. Acetylcholinesterase inhibition in
estuarine fish and invertebrates as an indicator of organophosphorus
insecticide exposure and effects. Environ. Toxicol. Chem. 20, 37–45.
George, S.G., Pirie, B.J.S., 1980. Metabolism of zinc in the mussel Mytilus
edulis (L.): a combined ultrastructural and biochemical study. J. Mar.
Biol. Assoc. UK 60, 575–590.
George, S.G., Pirie, B.J.S., Coombs, T.L., 1977. Absorption, accumula-
tion and excretion of iron-protein complexes by Mytilus edulis (L).
Proceedings of the International Conference on Heavy Metal

ARTICLE IN PRESSC. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243242
Environment, Toronto, Canada, Natural Research Council, Canada,
Publ. No. 2, pp. 887–900.
Giusti, L., Williamson, A.C., Mistry, A., 1999. Biologically available trace
metals in Mytilus edulis from the coast of Northeast England. Environ.
Int. 25, 969–981.
Gnaiger, E., 1983. Heat dissipation and energetic efficiency in animal
anoxibiosis: economy contra power. J. Exp. Zool. 228, 471–490.
Guilhermino, L., Barros, P., Silva, M.C., Soares, A.M.V.M., 1998. Should
the use of inhibition of cholinesterases as a specific biomarker for
organophosphate and carbamate pesticides be questioned? Biomarkers
3, 157–163.
Hamza-Chaffai, A., Romeo, M., Gnassia-Barelli, M., El Abed, A., 1998.
Effect of copper and lindane on some biomarkers measured in the clam
Ruditapes decussates. Bull. Environ. Contam. Toxicol. 61, 397–404.
Karageorgis, A.P., Hatzianestis, I., 2003. Surface sediment chemistry in
the Olympic Games 2004 Sailing Center (Saronikos Gulf). Medit. Mar.
Sci. 4, 5–22.
Khessiba, A., Romeo, M., Aıssa, P., 2005. Effects of some environmental
parameters on catalase activity measured in the mussel (Mytilus
galloprovincialis) exposed to lindane. Environ. Pollut. 133, 275–281.
Lemaire, P., Livingstone, D.R., 1993. Pro-oxidant/antioxidant processes
and organic xenobiotic interactions in marine organisms, in particular
the flounder Platicthys flesus and the mussel Mytilus edulis. Trends
Comp. Biochem. Physiol. 1, 1119–1150.
Lionetto, M.G., Caricato, R., Giordano, M.E., Pascariello, M.F.,
Marinosci, L., Schettino, T., 2003. Integrated use of biomarkers
(acetylcholinesterase and antioxidant enzymes activities) in Mytilus
galloprovincialis and Mullus barbatus in an Italian coastal marine area.
Mar. Pollut. Bull. 46, 324–330.
Livingstone, D.R., 2001. Contaminant-stimulated reactive oxygen species
production and oxidative damage in aquatic organisms. Mar. Pollut.
Bull. 42, 656–666.
Livingstone, D.R., Lips, F., Garcia Martinez, P., Pipe, R.K., 1992.
Antioxidant enzymes in the digestive gland of the common mussel
Mytilus edulis. Mar. Biol. 112, 265–276.
Luedeking, A., Koehler, A., 2004. Regulation of expression of multi-
xenobiotic resistance (MXR) genes by environmental factors in the
blue mussel Mytilus edulis. Aquat. Toxicol. 69, 1–10.
Marigomez, I., Soto, M., Cajaraville, M.P., Angulo, A., Giamberini, L.,
2002. Cellular and subcellular distribution of metals in molluscs.
Microsc. Res. Techniq. 56, 358–392.
Mauri, M., Baraldi, E., 2003. Heavy metal bioaccumulation in Mytilus
galloprovincialis: a transplantation experiment in Venice Lagoon.
Chem. Ecol. 19, 79–90.
Najimi, S., Bouhaimi, A., Daubeze, M., Zekhini, A., Pellerin, J.,
Narbonne, J.F., Moukrim, A., 1997. Use of Acetylcholinesterase in
Perna perna and Mytilus galloprovincialis as a biomarker of pollution
in Agadir Marine Bay (South of Morocco). Bull. Environ. Contam.
Toxicol. 58, 901–908.
Nakopoulou, Ch., Hatzias, V., Hadjigeorgiou, E., 1985. Levels of heavy
metals in the seawater in Northern Evoikos Gulf. In: Environmental
Effects of Dumping Metalliferous Slag in the Southern Part of Evoikos
Gulf. Technical Report. National Centre for Marine Research,
Athens, Greece, pp. 143–158 [in Greek].
Nicolaidou, A., Pancucci, M.A., Zenetos, A., 1989. The impact of
dumping coarse metalliferous waste on the benthos in Evoikos gulf,
Greece. Mar. Pollut. Bull. 20, 28–33.
Nicolaidou, A., Nott, J.A., 1989. Heavy metal pollution induced by a
ferro-nickel smelting plant in Greece. Sci. Total Environ. 84, 113–117.
Nicolaidou, A., Nott, J.A., 1990. Mediterranean pollution from a ferro-
nickel smelter: differential uptake of metals by some gastropods. Mar.
Pollut. Bull. 21, 137–143.
Nott, J.A., 1991. Cytology of pollutant metals in marine invertebrates: a
review of microanalytical applications. Scan. Microsc. 5, 191–205.
Nott, J.A., 1993. X-ray microanalysis in pollution studies. In: Sigee, D.C.,
Morgan, A.J., . Summer, A.T., Warley, A. (Eds.), X-ray Microanalysis
in Biology: Experimental Techniques and Applications. Cambridge
University Press, Cambridge, pp. 257–281.
Orbea, A., Maren, O-Z., Sole, M., Porte, C., Cajaraville, M.P., 2002.
Antioxidant enzymes and peroxisome proliferation in relation to
contaminant body burdens of PAHs and PCBs in bivalve molluscs,
crabs and fish from the Urdaibai and Plentzia estuaries (Bay of
Biscay). Aquat. Toxicol. 58, 75–98.
Pan, J-F., Wang, W-X., 2002. Comparison of the bioavailability of Cr and
Fe bound with natural colloids of different origins and sizes to two
marine bivalves. Mar. Biol. 141, 915–924.
Payne, J.F., Mathieu, A., Melvin, W., Fancey, L.L., 1996. Acetylcholi-
nesterase, an old biomarker with a new future? Field trials in
association with two urban rivers and a paper mill in Newfoundland.
Mar. Pollut. Bull. 32, 225–231.
Philips, D.J.H., 1976. The common mussel Mytilus edulis as an indicator
of pollution by zinc, cadmium, lead and copper. I Effects of
environmental variables on uptake of metals. Mar. Biol. 38, 59–69.
Regoli, F., Orlando, E., 1994. Accumulation and subcellular distribution
of metals (Cu, Fe, Mn, Pb and Zn) in the Mediterranean mussel
Mytilus galloprovincialis during a field transplant experiment. Mar.
Pollut. Bull. 28, 592–600.
Regoli, F., Principato, G., 1995. Glutathione, glutathione-dependent
and antioxidant enzymes in mussel, Mytilus galloprovincialis,
exposed to metals under field and laboratory conditions: implica-
tions for the use of biochemical biomarkers. Aquat. Toxicol. 31,
143–164.
Reid, D.J., MacFarlane, G.R., 2003. Potential biomarkers of crude oil
exposure in the gastropod mollusc Austrocochlea porcata: laboratory
and manipulative field studies. Environ. Pollut. 123, 147–155.
Rickwood, C.J., Galloway, T.S., 2004. Acetylcholinesterase inhibition as a
biomarker of adverse effect. A study of Mytilus edulis exposed to the
priority pollutant chlorfenvinphos. Aquat. Toxicol. 67, 45–56.
Roesijadi, G., Young, J.S., Drum, A.S., Gurtisen, J.M., 1984. Behavior of
trace metals in Mytilus edulis during a reciprocal transplant field
experiment. Mar. Ecol. Prog. Ser. 18, 155–170.
Sanders, B.M., Martin, L.S., Nelson, W.G., Phelps, D.K., 1991.
Relationships between accumulation of a 60 kDa stress protein and
Scope for Growth in Mytilus edulis exposed to a range of copper
concentrations. Mar. Environ. Res. 31, 81–97.
Serafim, M.A., Company, R.M., Bebianno, M.J., Langston, W.J., 2002.
Effect of temperature and size on metallothionein synthesis in the gill
of Mytilus galloprovincialis exposed to cadmium. Mar. Environ. Res.
54, 361–365.
Sheehan, D., Power, A., 1999. Effects of seasonality on xenobiotic and
antioxidant defense mechanisms of bivalve molluscs. Comp. Biochem.
Physiol. 123C, 193–199.
Sole, M., Porte, C., Albaiges, J., 1994. Mixed function oxygenase system
components and antioxidant enzymes in different marine bivalves: its
relation with contaminant body burdens. Aquat. Toxicol. 30, 271–283.
Stohs, S.J., Bagchi, D., 1995. Oxidative mechanisms in the toxicity of
metal ions. Free Radical Biol. Med. 18, 321–336.
Szefer, P., Kim, B.S., Kim, C.K., Kim, E.H., Lee, C.B., 2004. Distribution
and coassociations of trace elements in soft tissue and byssus of
Mytilus galloprovincialis relative to the surrounding seawater and
suspended matter of the southern part of the Korean Peninsula.
Environ. Pollut. 129, 209–228.
UNEP, 1984. Determination of total Cd, Zn, Pd and Cu in selected marine
organisms by atomic absorption spectrophotometry. Reference Meth-
ods for Marine Pollution Studies, No. 11, Rev. 1.
Vesk, P.A., Byrne, M., 1999. Metal levels in tissue granules of the
freshwater bivalve Hyridella depressa (Unionida) for biomonitoring:
the importance of cryopreparation. Sci. Total Environ. 225, 219–229.
Viarengo, A., Burlando, B., Dondero, F., Marro, A., Fabbri, R., 1999.
Metallothionein as a tool in biomonitoring programmes. Biomarkers
4, 455–466.
Viarengo, A., Canesi, L., 1991. Mussels as biological indicators of
pollution. Aquaculture 94, 225–243.
Vidal, M.L., Basseres, A., Narbonne, J.F., 2002. Seasonal variations of
pollution biomarkers in two populations of Corbicula fluminea
(Muller). Comp. Biochem. Physiol. 131C, 133–151.

ARTICLE IN PRESSC. Tsangaris et al. / Ecotoxicology and Environmental Safety 66 (2007) 232–243 243
Voutsinou-Taliadouri, F., Psyllidou-Giouranovits, R., Georgakopoulou-
Gregoriadou, E., 1995. Active growth of a submarine slag disposal ore
deposit in the northern Euboekos Gulf, Greece. Fresen. Environ. Bull.
4, 195–200.
Walsh, A.R., O’Halloran, J., 1997. The accumulation of chromium by
mussels Mytilus edulis (L) as a function of valency, solubility and
ligation. Mar. Environ. Res. 43, 41–53.
Walsh, A.R., O’Halloran, J., 1998. Accumulation of chromium by a
population of mussels (Mytilus edulis (L.)) exposed to leather tannery
effluent. Environ. Toxicol. Chem. 17, 1429–1438.
Wendel, A., 1980. Glutathione peroxidase. In: Jakoby, W.B., Bend, J.R.,
Caldwell, J. (Eds.), Enzymatic Basis of Detoxication. Academic Press,
NewYork, pp. 333–348.
Widdows, J., Donkin, P., 1992. Mussels and environmental contaminants:
bioaccumulation and physiological aspects. In: Gosling, E. (Ed.), The
Mussel Mytilus: Ecology, Physiology, Genetics and Culture. Elsevier,
Amsterdam, pp. 383–424.
Widdows, J., Donkin, P., Brinsley, M.D., Evans, S.V., Salked, P.N.,
Franklin, A., Law, R.J., Waldock, M.J., 1995a. Scope for growth and
contaminant levels in North Sea mussels Mytilus edulis. Mar. Ecol.
Prog. Ser. 127, 131–148.
Widdows, J., Donkin, P., Evans, S.V., Page, S., Salked, P.N., 1995b.
Sublethal biological effects and chemical contaminant monitoring of
Sullom Voe (Shetland) using mussels (Mytilus edulis). R. Soc.
(Edinburgh) Proc. 103B, 99–112.
Widdows, J., Donkin, P., Staff, F.J., Matthiessen, P., Law, R.J., Allen, Y.T.,
Thain, J.E., Allchin, C.R., Jones, B.R., 2002. Measurement of stress
effects (scope for growth) and contaminant levels in mussels (Mytilus
edulis) collected from the Irish Sea. Mar. Environ. Res. 53, 327–356.
Widdows, J., Fieth, P., Worrall, C.M., 1979. Relationships between
seston, available food and feeding activity in the common mussel
Mytilus edulis. Mar. Biol. 50, 195–207.
Widdows, J., Salked, P., 1992. Practical procedures for measurement of
Scope for Growth. In: Report of the FAO/IOC/UNEP Training
Workshop on the Techniques for Monitoring Biological Effects of
Pollutants in Marine Organisms, pp. 175–205.
Winston, G.W., Di Giulio, R.T., 1991. Prooxidant and antioxidant
mechanisms in aquatic organisms. Aquat. Toxicol. 19, 137–161.




















