
Ascorbate distribution during hibernation is independent of ascorbate redox state
10
Original Contribution ASCORBATE DISTRIBUTION DURING HIBERNATION IS INDEPENDENT OF ASCORBATE REDOX STATE YI LONG MA, * MARGARET E. RICE, y MEI LAN CHAO, y P ATRICIA M. RIVERA, * HUIWEN W. ZHAO, * AUSTIN P. ROSS, * XIONGWEI ZHU, z MARK A. SMITH, z and KELLY L. DREW * ,z * Institute of Arctic Biology, University of Alaska Fairbanks, Fairbanks, AK, USA; y Department of Physiology and Neuroscience, New York University School of Medicine, New York, NY, USA; and z Institute of Pathology, Case Western Reserve University, Cleveland, OH, USA (Received 19 November 2003; Revised 16 April 2004; Accepted 22 April 2004) Available online 25 May 2004 Abstract—Distribution of ascorbate into tissues is an essential process in ascorbate antioxidant defense. Hibernating animals are studied as a model of tolerance to ischemia – reperfusion because of their tolerance to fluctuations in blood flow associated with prolonged torpor and periodic arousal episodes. Throughout hibernation, plasma ascorbate concentration ([Asc] p ) repetitively increases during torpor, then falls during periodic arousal bouts. We previously proposed that high [Asc] p provides a ready source of antioxidant protection for distribution to the central nervous system and peripheral tissues during arousal. Here we tested whether deliberate oxidation of plasma ascorbate by intravenous administration of ascorbate oxidase (AO), prior to arousal, compromised tissue levels of ascorbate or the other water- soluble antioxidants, glutathione (GSH) and urate. Although AO decreased [Asc] p to below the level of detection during torpor and after arousal, ascorbate oxidation did not decrease post-arousal tissue levels of reduced ascorbate, glutathione, or urate in any tissue examined, except liver. The data imply that ascorbate is taken up equally well into brain and other tissues as either ascorbate or its oxidized product dehydroascorbate, with subsequent intracellular reduction of dehydroascorbate. Lack of effect of ascorbate oxidation on tissue levels of GSH or urate indicates that dehydroascorbate uptake and reduction do not compromise tissue concentrations of these other water-soluble antioxidants. Thus, we show equal availability of reduced and oxidized plasma ascorbate during metabolically demanding thermogenesis and reperfusion associated with arousal from hibernation. D 2004 Elsevier Inc. All rights reserved. Keywords—Vitamin C, Ascorbic acid, Reperfusion, Hypothermia, Torpor, Dehydroascorbate, Glutathione, Urate, Uric acid, Free radicals INTRODUCTION Ascorbate is the most important antioxidant in plasma [1] and has been shown to minimize ischemic damage in models of cerebral ischemia–reperfusion, both in vivo [2–5] and in vitro [6,7]. Hibernation is neuro- protective [8] and is studied as a model of tolerance to ischemia, reperfusion, neurodegenerative disease, and brain injury [9,10]. During hibernation, ascorbate con- centration increases 4-fold in plasma ([Asc] p ) and doubles in cerebralspinal fluid [11]. On arousal, [Asc] p decreases to levels seen in euthermic animals (37jC body temperature), at the same time as O 2 consumption [12] and cerebral blood flow [13] reach maximum levels. We have proposed previously that during the 300-fold surge in oxidative metabolism that is neces- sary to rewarm from 2 to 37jC, ascorbate is redis- tributed to metabolically active tissues, including the brain, where it provides protection against oxidative stress [12]. To address this hypothesis, we assessed whether oxidation of ascorbate in plasma prior to resurgence of oxidative metabolism during arousal from Address correspondence to: Dr. K. L. Drew, Institute of Arctic Biology, Alaska Basic Neuroscience Program, PO Box 757000, University of Alaska Fairbanks, Fairbanks, AK 99775-7000, USA; Fax: (907) 474-6050; E-mail: [email protected]. Free Radical Biology & Medicine, Vol. 37, No. 4, pp. 511–520, 2004 Copyright D 2004 Elsevier Inc. Printed in the USA. All rights reserved 0891-5849/$-see front matter doi:10.1016/j.freeradbiomed.2004.04.025 511
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Ascorbate distribution during hibernation is independent of ascorbate redox state

Free Radical Biology & Medicine, Vol. 37, No. 4, pp. 511–520, 2004Copyright D 2004 Elsevier Inc.
Printed in the USA. All rights reserved0891-5849/$-see front matter
doi:10.1016/j.freeradbiomed.2004.04.025
Original Contribution
ASCORBATE DISTRIBUTION DURING HIBERNATION IS INDEPENDENT OF
ASCORBATE REDOX STATE
YI LONG MA,* MARGARET E. RICE,y MEI LAN CHAO,y PATRICIA M. RIVERA,* HUIWEN W. ZHAO,* AUSTIN P. ROSS,*
XIONGWEI ZHU,z MARK A. SMITH,z and KELLY L. DREW *,z
*Institute of Arctic Biology, University of Alaska Fairbanks, Fairbanks, AK, USA; yDepartment of Physiology and Neuroscience,New York University School of Medicine, New York, NY, USA; and z Institute of Pathology,
Case Western Reserve University, Cleveland, OH, USA
(Received 19 November 2003; Revised 16 April 2004; Accepted 22 April 2004)
Available online 25 May 2004
Add
Biolog
Univer
Fax: (9
Abstract—Distribution of ascorbate into tissues is an essential process in ascorbate antioxidant defense. Hibernating
animals are studied as a model of tolerance to ischemia–reperfusion because of their tolerance to fluctuations in blood
flow associated with prolonged torpor and periodic arousal episodes. Throughout hibernation, plasma ascorbate
concentration ([Asc]p) repetitively increases during torpor, then falls during periodic arousal bouts. We previously
proposed that high [Asc]p provides a ready source of antioxidant protection for distribution to the central nervous system
and peripheral tissues during arousal. Here we tested whether deliberate oxidation of plasma ascorbate by intravenous
administration of ascorbate oxidase (AO), prior to arousal, compromised tissue levels of ascorbate or the other water-
soluble antioxidants, glutathione (GSH) and urate. Although AO decreased [Asc]p to below the level of detection during
torpor and after arousal, ascorbate oxidation did not decrease post-arousal tissue levels of reduced ascorbate, glutathione,
or urate in any tissue examined, except liver. The data imply that ascorbate is taken up equally well into brain and other
tissues as either ascorbate or its oxidized product dehydroascorbate, with subsequent intracellular reduction of
dehydroascorbate. Lack of effect of ascorbate oxidation on tissue levels of GSH or urate indicates that dehydroascorbate
uptake and reduction do not compromise tissue concentrations of these other water-soluble antioxidants. Thus, we show
equal availability of reduced and oxidized plasma ascorbate during metabolically demanding thermogenesis and
reperfusion associated with arousal from hibernation. D 2004 Elsevier Inc. All rights reserved.
Keywords—Vitamin C, Ascorbic acid, Reperfusion, Hypothermia, Torpor, Dehydroascorbate, Glutathione, Urate, Uric
acid, Free radicals
INTRODUCTION
Ascorbate is the most important antioxidant in plasma
[1] and has been shown to minimize ischemic damage
in models of cerebral ischemia–reperfusion, both in
vivo [2–5] and in vitro [6,7]. Hibernation is neuro-
protective [8] and is studied as a model of tolerance to
ischemia, reperfusion, neurodegenerative disease, and
ress correspondence to: Dr. K. L. Drew, Institute of Arctic
y, Alaska Basic Neuroscience Program, PO Box 757000,
sity of Alaska Fairbanks, Fairbanks, AK 99775-7000, USA;
07) 474-6050; E-mail: [email protected].
511
brain injury [9,10]. During hibernation, ascorbate con-
centration increases 4-fold in plasma ([Asc]p) and
doubles in cerebralspinal fluid [11]. On arousal, [Asc]pdecreases to levels seen in euthermic animals (37jCbody temperature), at the same time as O2 consumption
[12] and cerebral blood flow [13] reach maximum
levels. We have proposed previously that during the
300-fold surge in oxidative metabolism that is neces-
sary to rewarm from 2 to 37jC, ascorbate is redis-
tributed to metabolically active tissues, including the
brain, where it provides protection against oxidative
stress [12]. To address this hypothesis, we assessed
whether oxidation of ascorbate in plasma prior to
resurgence of oxidative metabolism during arousal from

Y. L. Ma et al.512
hibernation compromises tissue ascorbate content. Our
results indicate that oxidizing ascorbate in the plasma
compartment prior to arousal from hibernation does not
affect tissue ascorbate content after arousal in brain or
any tissue studied (except liver), nor does it affect
tissue concentrations of the other water-soluble anti-
oxidants, glutathione (GSH) and urate, in brain or any
tissue examined.
MATERIALS AND METHODS
Animal handling
All procedures were approved by the Institutional
Animal Care and Use Committee of the University of
Alaska Fairbanks and were in accordance with National
Research Council’s Guide for the Care and Use of
Laboratory Animals. Arctic ground squirrels (AGS,
Spermophilus parryii) were trapped during mid-July in
the northern foothills of the Brooks Range, AK, USA,
approximately 30 miles south of the Toolik Field Station
of the University of Alaska, Fairbanks (68j 38V W,
elevation 809 m), and transported to Fairbanks. The
squirrels were housed individually at 18jC, under a
natural light cycle for 64jC latitude, and fed rodent
chow, sunflower seeds, and fresh carrots and apples ad
libidum until mid-September. They were then moved to
an environmental chamber set to an ambient temperature
(Ta) of 2jC and 4:20 h light:dark cycle. Animal experi-
ments were conducted December through April. Hiber-
nation was assessed from core body temperature or by
inactivity (indicated by the lack of disturbance of wood
shavings placed on an animal’s back 24 h previously).
Deep hibernation was verified by rectal temperature at
the time of euthanasia.
Surgical procedures
Telemetry transmitters, used to monitor core body
temperature, and indwelling femoral arterial and venous
cannulas were implanted in all AGS treated with AO or
saline. Prior to surgery, AGS were placed at 21jC and
left overnight to arouse from hibernation. General
anesthesia induced with halothane (Halocarbon Lab,
Riveredge, NJ, USA) at 5% was maintained at 2.5–
3% mixed with 100% medical grade O2 at a flow rate
of 1.5 l/min. Surgery was performed under strictly
aseptic conditions. Body temperature was kept between
36.5 and 37.5jC with a fluid-filled heating pad (Omni
Medical Equipment, Cincinnati, OH, USA) throughout
the surgery. Cannulas were inserted into the femoral
artery and vein (Tygon tubing; 0.375 mm i.d., 0.75 mm
o.d.; Norton, Akron, OH, USA), passed subcutaneously
and dorsally to an incision at the back of the neck, and
secured with suture as described previously [12]. Pre-
calibrated, wax-coated telemetry transmitters (Model
VM-FH, 2 cm disk, Mini Mitter Co., Inc., Sunriver,
OR, USA) were implanted intraperitoneally via a
midline incision through linea alba, and sealed with
three layers of sutures. Animals were allowed 1 day
postoperative recovery at 18jC before being returned
to the environmental chamber to resume hibernation.
Ascorbate oxidase experiments
To determine the effect of ascorbate redox state
during arousal on postarousal tissue levels of reduced
ascorbate, GSH, and urate, plasma ascorbate was
deliberately oxidized with AO administered by intra-
venous injection. AO (EC 1.10.3.3), isolated from
zucchini, oxidizes ascorbate to dehydroascorbate
[14,15]. Control AGS received an equal i.v. volume
of saline (0.9% NaCl) vehicle. For these experiments,
AO (96 IU/kg in a volume of 0.6 ml/kg saline) or
vehicle was administered immediately before arousal
from hibernation during four consecutive bouts of
torpor with mean F SEM torpor bout lengths of
6.1 F 0.88 days (AO group) and 5.5 F 0.54 days
(saline group). In each of the four episodes, arterial
blood was sampled immediately prior to i.v. injection
(T0) and 30 min after injection (T1). An additional
blood sample was collected after the fourth arousal
bout (4T2) when an animal had reached a body tem-
perature of at least 35jC (see Fig. 1). At each sampling
point, approximately 150 Al of blood was collected into
heparinized capillary tubes and spun for 5 min in a
microcapillary centrifuge, and three 20 Al samples of
plasma were collected and immediately frozen on dry
ice. These samples were stored at �80jC until ascor-
bate analysis.
Tissue collection
Tissues were collected immediately after arterial
blood was sampled on the fourth arousal bout. Tissue
was sampled from four groups of animals as described
above (euthermic, hibernating, aroused-AO, and
aroused-saline). Before initial sampling in euthermic
and aroused AGS, animals were lightly anesthetized
with halothane (5% halothane mixed with O2 delivered
at 1.5 l/min); hibernators were not initially anesthetized.
Rectal temperatures were measured with a copper–
constantan thermocouple. After decapitation, brains
and peripheral tissues (spleen, liver, kidney, heart, skel-
etal muscle, brown fat, white fat, adrenal, small intes-
tine) were removed immediately, dissected, and frozen
in liquid N2. Brain samples were taken from frontal
cortex, hippocampus, and cerebellum, and frozen in
liquid N2 as in previous studies [11,12]. Frozen tissue
samples were stored at �80jC. Time from decapitation
to freezing was less than 10 min.

Fig. 1. Core body temperature over time in an individual AGS illustrates time of blood sampling and AO or saline injection. Arterialblood was sampled at T0 followed immediately by intravenous injection of AO (96 IU/kg) or saline. Thirty minutes later (T1), blood wassampled again, before body temperature began to increase. Emergence from hibernation was initiated during blood sampling and wasrepeated four times to mimic natural periodic arousal from hibernation. Numbers 1 through 4 indicate arousal bouts. Blood was sampledafter the fourth injection, and body temperature reached 35jC (4T2) just prior to tissue collection. Torpor bout length averaged (FSEM)5.7 F 0.4 days and did not differ between AO- and saline-treated groups.
Ascorbate distribution and redox state 513
HPLC analysis
Tissue samples. On the day of analysis, tissues
(typically 10 mg) were weighed and sonicated while
still frozen in ice-cold, deoxygenated eluent (see be-
low). Samples were then centrifuged, and the superna-
tant was injected directly onto the HPLC system.
HPLC analysis was based on methods described pre-
viously [12,16] using a gold amalgam electrode for
ascorbate and GSH and a glassy carbon electrode for
urate. Eluent contained 16 mM monochloroacetic acid,
with 150 mg L�1 EDTA and 300 mg L�1 myristyl
dimethylbenzylammonium chloride, pH 5, and up to
20% methanol. Retention time for urate is similar to
that for GSH under these conditions, however, GSH is
not detected on the carbon electrode and thus does not
interfere with urate analysis. Standards were prepared
in ice-cold, deoxygenated eluent at regular intervals
throughout the analysis. Tissue contents of ascorbate,
urate, and GSH are expressed as micromoles per gram
wet weight of tissue.
Plasma samples. On the day of analysis, plasma sam-
ples were diluted 1:4 in ice cold, deoxygenated eluent (see
above) and spun at 14,000 rpm for 1 min, and 10 Al of thesupernatant was again injected directly onto the HPLC.
Reduced plasma [Asc]p and urate ([Urate]p) were ana-
lyzed simultaneously using methods similar to those for
tissue samples with an ESA 5100A coulometric detector
with the analytical cell (Model 5011; ESA, Inc., Chelm-
ford, MA, USA) set at 0.4 V versus Ag/AgCl. Plasma
levels of GSH were not determined. The limit of detection
for ascorbate (signal-to noise ratio of 3:1) with this assay
was 1.0 AM per injection (i.e., 4 AM [Asc]p).
Chemicals
All chemicals were analytical grade. Myristyl dime-
thylbenzylammonium chloride (Myris-100) was a gift
from Jame Fine Chemicals, (Bound Brook, NJ). EDTA,
monochloroacetic acid (MCA), ascorbate, uric acid, and
GSH were purchased from Sigma–Aldrich (St. Louis,
MO, USA). Ascorbate oxidase (L-ascorbate oxidoreduc-
tase, EC 1.10.3.3) was purchased from Calzyme Labo-
ratories, Inc (San Luis Obispo, CA, USA). All glassware
used for HPLC analysis was treated with concentrated
HNO3, followed by thorough rinsing with deionized
water [17].
Statistics
All plasma and tissue samples were analyzed by
HPLC in triplicate. Subsequent statistical analyses were
performed on means of triplicates with n equal to the
number of animals. Plasma urate concentrations were
compared using repeated-measures ANOVA, paired t
test, or t test where indicated. Tissue contents of ascor-

Y. L. Ma et al.514
bate, urate, and glutathione in peripheral tissues were
compared using one-way ANOVA and Tukey post-hoc
comparisons. Differences in ascorbate content among
brain regions were analyzed using two-way ANOVA
followed by pairwise multiple comparisons (Tukey test)
(SigmaStat, SPSS Science, Chicago, IL, USA). Data are
expressed as group means F SEM. The criterion for
statistical significance was p < .05.
RESULTS
Effect of AO on [Asc]p
The effect of AO treatment on [Asc]p and [Urate]p is
shown in Fig. 2. By 30 min after AO injection (T1),
[Asc]p was below the detection limit in 14 of 16 samples
(Fig. 2A) (1T1 through 4T1 for each of four animals).
Although five animals received AO through patent
venous cannulas, intermittent plasma sampling was not
possible in one animal because the arterial cannula was
temporarily blocked). [Asc]p was also below the limit of
detection in all AO-treated animals sampled at 4T2immediately prior to tissue collection. Thus, enzymatic
Fig. 2. Effects of intravenous AO injection on [Asc]p and [Urate]p. (A)Thirty minutes after AO injection (T1), plasma ascorbate declined tobelow-detectable levels in the majority of samples. Although [Asc]pincreased again prior to the next injection, low [Asc]p at 4T2 indicatesthat [Asc]p remained low during arousal from hibernation. Decrease in[Asc]p in the saline group at 4T2 illustrates the typical decline in [Asc]pseen during arousal from hibernation. (B) The difference in [Urate]pbetween AO- and saline-treated groups at 4T2 was not statisticallysignificant ( p = .110, t test, n = 4–5).
oxidation of [Asc]p persisted after arousal (4T2 sampling
point), whereas in saline-treated control AGS, [Asc]preturned to a level (mean F SEM: 47.3F14.8 AM, n =
5) typical of euthermic animals [11,12].
We have suggested previously that [Urate]p may be an
indicator of oxidative stress [12]. Notably, AO adminis-
tration did not cause an increase in [Urate]p (Fig. 2B). On
the other hand, there was a tendency for [Urate]p to
increase after arousal from hibernation (at 4T2) in control
AGS, but not in AO-treated animals. The difference in
[Urate]p between groups at this sampling point was not
significant ( p = .110, t test, n = 4–5). Furthermore, no
significant difference was observed in [Urate]p analyzed
using a repeated-measures ANOVA or a paired t test to
compare [Urate]p sampled at 4T1 (prior to arousal) and
4T2 (after arousal) within saline-treated animals ( p =
.229, paired t test). Although some animals showed a
pronounced increase in [Urate]p, concentration decreased
in two of five animals after arousal. The tendency for
[Urate]p to increase at 4T2 in saline-treated AGS contrasts
with previous results in which [Urate]p peaked at the
same time as oxygen consumption and returned to basal
levels before core Tb reached 35jC (Fig. 2) [12]. The
large degree of variability in [Urate]p at 4T2 (Fig. 2B)
suggests the increase reflects variability in the time
course of the transient rise in [Urate]p rather than a
specific affect of AO.
Effect of hibernation, arousal, and [Asc]p oxidation on
brain ascorbate, GSH, and urate
Depletion of [Asc]p by AO had no effect on brain
ascorbate, GSH, or UA content in any brain region exam-
ined (Fig. 3). Regarding effects of hibernation and arousal,
ascorbate levels in brain were greatest in animals after
arousal, whether treated with saline or AO (Fig. 3A) (main
effect of group, p < .001). Brain ascorbate levels were
similar in aroused-saline and aroused-AO groups, but
differed from both euthermic and hibernation groups,
(Tukey test, p < .05 euthermic = hibernating < aroused-
saline = aroused-AO, n = 5). Although statistical compar-
ison indicates an overall increase in brain ascorbate levels,
results (Fig. 3A) suggest the increase in brain ascorbate was
restricted to frontal cortex. Nonetheless, the interaction
between treatment and brain region was not significant.
Similar analysis of brain GSH levels showed no
effect of hibernation state or arousal with or without
AO, although there were some regional differences in
absolute level, with higher GSH levels in hippocampus
than in frontal cortex or cerebellum (Fig. 3B) (main
effect of region, p < .001, n = 5; Tukey test comparing
regions, p < .05). Paradoxically, hibernation was asso-
ciated with a pronounced increase in urate levels, with
significantly higher levels during hibernation than in any
other state (Fig. 3C) (main effects of group, region, and
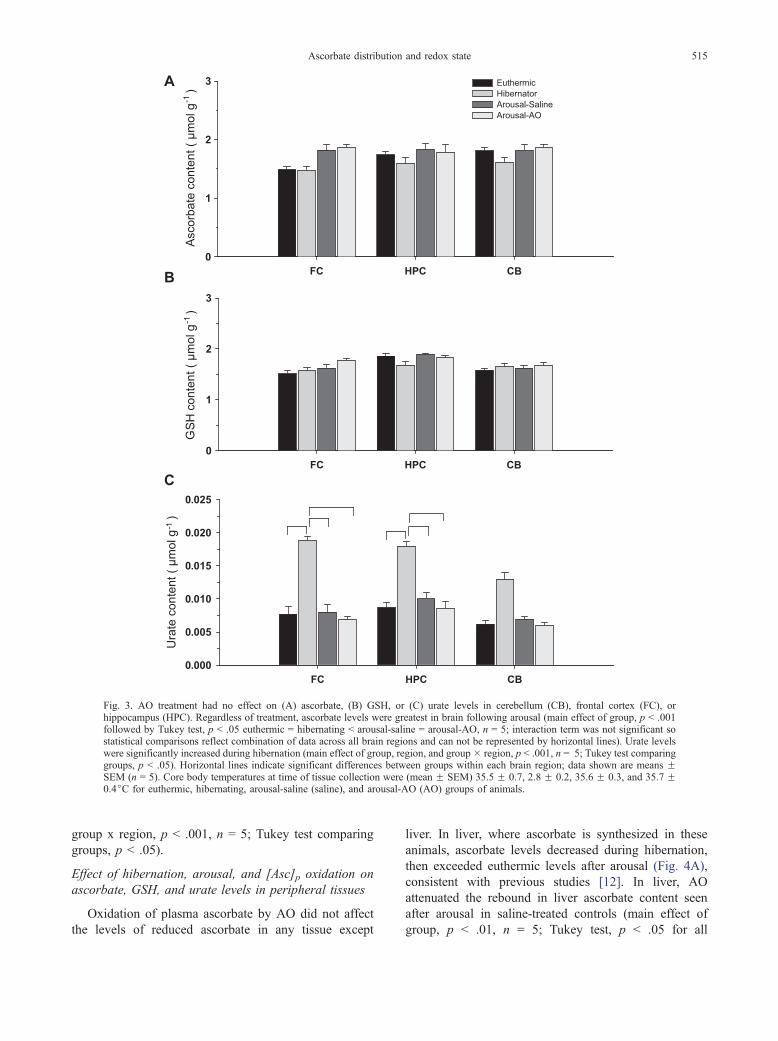
Fig. 3. AO treatment had no effect on (A) ascorbate, (B) GSH, or (C) urate levels in cerebellum (CB), frontal cortex (FC), orhippocampus (HPC). Regardless of treatment, ascorbate levels were greatest in brain following arousal (main effect of group, p < .001followed by Tukey test, p < .05 euthermic = hibernating < arousal-saline = arousal-AO, n = 5; interaction term was not significant sostatistical comparisons reflect combination of data across all brain regions and can not be represented by horizontal lines). Urate levelswere significantly increased during hibernation (main effect of group, region, and group � region, p < .001, n = 5; Tukey test comparinggroups, p < .05). Horizontal lines indicate significant differences between groups within each brain region; data shown are means FSEM (n = 5). Core body temperatures at time of tissue collection were (mean F SEM) 35.5 F 0.7, 2.8 F 0.2, 35.6 F 0.3, and 35.7 F0.4jC for euthermic, hibernating, arousal-saline (saline), and arousal-AO (AO) groups of animals.
Ascorbate distribution and redox state 515
group x region, p < .001, n = 5; Tukey test comparing
groups, p < .05).
Effect of hibernation, arousal, and [Asc]p oxidation on
ascorbate, GSH, and urate levels in peripheral tissues
Oxidation of plasma ascorbate by AO did not affect
the levels of reduced ascorbate in any tissue except
liver. In liver, where ascorbate is synthesized in these
animals, ascorbate levels decreased during hibernation,
then exceeded euthermic levels after arousal (Fig. 4A),
consistent with previous studies [12]. In liver, AO
attenuated the rebound in liver ascorbate content seen
after arousal in saline-treated controls (main effect of
group, p < .01, n = 5; Tukey test, p < .05 for all
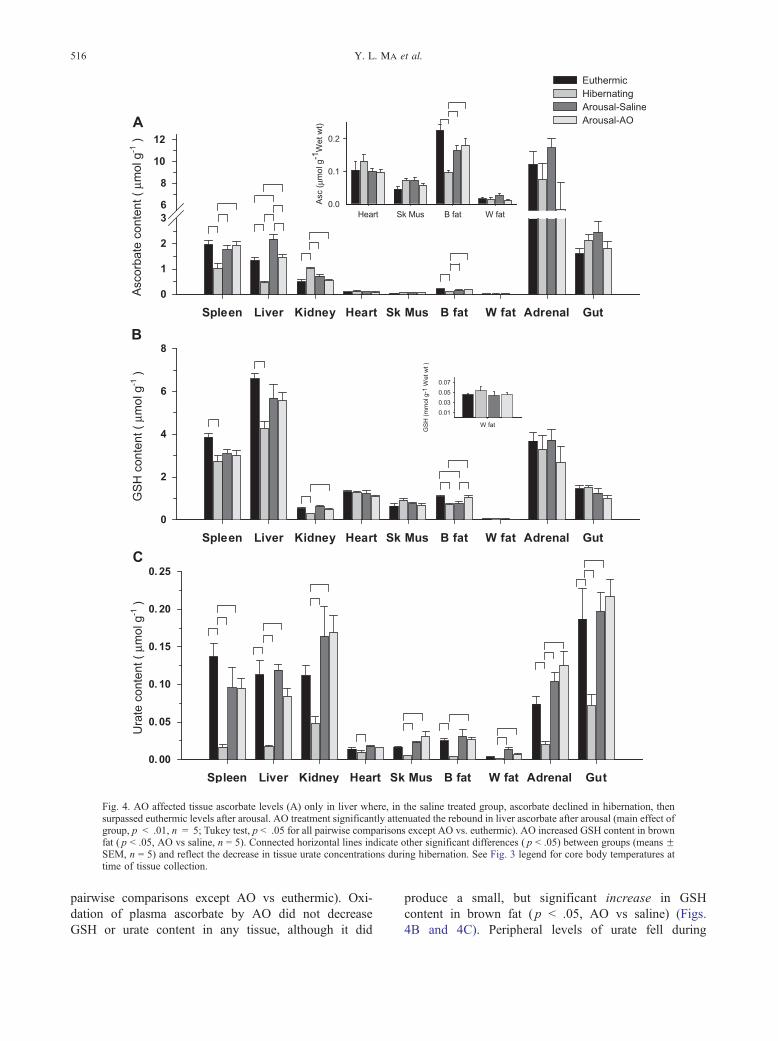
Fig. 4. AO affected tissue ascorbate levels (A) only in liver where, in the saline treated group, ascorbate declined in hibernation, thensurpassed euthermic levels after arousal. AO treatment significantly attenuated the rebound in liver ascorbate after arousal (main effect ofgroup, p < .01, n = 5; Tukey test, p < .05 for all pairwise comparisons except AO vs. euthermic). AO increased GSH content in brownfat ( p < .05, AO vs saline, n = 5). Connected horizontal lines indicate other significant differences ( p < .05) between groups (means FSEM, n = 5) and reflect the decrease in tissue urate concentrations during hibernation. See Fig. 3 legend for core body temperatures attime of tissue collection.
Y. L. Ma et al.516
pairwise comparisons except AO vs euthermic). Oxi-
dation of plasma ascorbate by AO did not decrease
GSH or urate content in any tissue, although it did
produce a small, but significant increase in GSH
content in brown fat ( p < .05, AO vs saline) (Figs.
4B and 4C). Peripheral levels of urate fell during

Ascorbate distribution and redox state 517
hibernation, in contrast to the significant increase in
urate that occurred in brain (Fig. 4C).
DISCUSSION
Maintenance of tissue ascorbate levels by reduced or
oxidized plasma ascorbate
Ascorbate, an essential component of the antioxidant
network, is synthesized in the liver in mammals and
transported in plasma to all other tissues, where it is then
concentrated in the intracellular compartment by active
uptake [18]. Ascorbate is the most important antioxidant
in plasma [1] and has been shown to minimize ischemic
damage in models of cerebral ischemia–reperfusion,
both in vivo [3–5] and in vitro [6]. During hibernation,
ascorbate concentration increases 4-fold in plasma
([Asc]p) and doubles in cerebralspinal fluid [11]. On
arousal, [Asc]p declines to levels seen in euthermic
animals (37jC body temperature), and the rate of fall
peaks at the time of maximal O2 consumption [12] and
cerebral blood flow [13]. We have proposed that ascor-
bate is redistributed to vulnerable tissues during arousal
where it serves as an antioxidant and cytoprotectant. In
the present study, we asked if oxidizing ascorbate in the
plasma compartment prior to arousal from hibernation
would affect tissue ascorbate concentrations. We show
that recovery of tissue ascorbate content during arousal
does not require plasma ascorbate to remain in the
reduced form ([Asc]p). After arousal, tissue levels of
[Asc]p were the same in AO- and saline-treated AGS in
all tissues examined except liver. This suggests that
ascorbate and its oxidation product, dehydroascorbate
(DHA), are taken up equally well into tissues, with ready
reduction of DHA back to ascorbate by GSH and other
thiols [19,20] and by GSH-dependent dehydroascorbate
reductase [21,22]. The finding that the post-arousal tissue
levels of reduced GSH and urate were the same in AO-
and saline-treated animals argues that intracellular anti-
oxidant capacity is sufficient to allow for reduction of
DHA without compromising concentrations of these
other water-soluble antioxidants.
During arousal in AGS, ascorbate is presumably taken
up by active transport. In peripheral tissues, this uptake is
mediated by SVCT1 or SVCT2 depending on the organ
examined [23]. In the central nervous system, uptake of
reduced ascorbate occurs exclusively via SVCT2, first
from plasma via the choroid plexus (the site of CSF
production), then from the extracellular fluid into neu-
rons [18,23]; glial cells do not appear to express either
SVCT1 or SVCT2 [23,24]. Additionally, DHA can be
taken up in most cells via facilitated diffusion of DHA
through glucose transporters GLUT 1, GLUT 3, and
GLUT 4 [25–27]. Other studies have shown that if
sodium-dependent uptake of ascorbate becomes limiting,
such as after ischemia, uptake of DHA via glucose
transporters may enhance ascorbate distribution and
cytoprotection [5]. The results reported here suggest that
during arousal from hibernation, ascorbate and DHA are
equally well distributed to provide cytoprotection in
brain and other tissues studied.
In liver, ascorbate content returned to euthermic levels
after arousal in AO-treated animals, but did not rebound
to exceed euthermic levels as it did in saline-treated
controls. Although liver is the site of ascorbate synthesis
in mammals [28], hepatic levels are maintained by active
uptake, as indicated by the high density of SVCT1 in
liver [23] and the ready uptake of ascorbate into hep-
atocytes [29] and liver microsomes [30]. Results suggest
that in liver, uptake of ascorbate during arousal exceeds
that of DHA, most likely because of a high density of
SVCT1. A direct effect of AO on ascorbate synthesis in
liver is unlikely, as AO is a 70.4 kDa protein that would
be trapped in the plasma compartment after intravenous
injection.
AO treatment did not compromise tissue content of
ascorbate, GSH, or urate in other peripheral tissues,
such as spleen and brown fat, where ascorbate de-
creased during hibernation but returned to euthermic
levels on arousal. These results suggest, that as in brain,
DHA in peripheral tissues is taken up and subsequently
reduced to ascorbate without affecting concentrations of
other water-soluble antioxidants.
Redox balance during hibernation and after arousal
Tissue levels of ascorbate and GSH reflect oxidative
status of the tissue such that decreases in either one or
more of these antioxidants would be consistent with
decreased antioxidant capacity and oxidative stress.
Ascorbate in brain is tightly regulated [18] and fluctuates
little throughout hibernation and arousal [12]. A trend for
brain tissue levels of ascorbate to increase after arousal
observed previously [12] was statistically significant in
this independent set of samples. This increase could be
derived, in part, from ascorbate stores in cerebral spinal
fluid that double during hibernation [11]. Although it is
unclear if the slight increase in brain ascorbate provides a
physiologically relevant increase in antioxidant capacity,
preservation of brain ascorbate levels is consistent with a
lack of oxidative stress during hibernation or arousal.
Similarly, maintenance of GSH levels in brain is consis-
tent with redox homeostasis; however, because levels of
GSSG (and thus total GSH equivalents) were not mea-
sured in the present study, the question of GSH redox
balance remains open and requires further study.
Curiously, brain urate levels increased dramatically
during hibernation, consistent with previous results [12].
Reactive oxygen species rapidly convert xanthine dehy-

Y. L. Ma et al.518
drogenase to xanthine oxidase, the enzyme responsible
for metabolism of xanthine to urate. For this reason
transient increases in urate concentrations have been
interpreted as evidence of increased oxidative stress
[12]. Elevations in brain urate concentrations during
hibernation may likewise reflect increased oxidative
stress. Alternatively, increases in urate concentration in
brain may reflect enhanced nucleotide metabolism or
decreased urate clearance from brain tissue. More direct
and comprehensive measures of oxidative stress in brain
across the hibernation cycle are necessary to confirm
redox status during hibernation and arousal.
GSH decreased in spleen, liver, and brown fat during
hibernation, consistent with a shift toward a more oxi-
dized environment. Although preliminary studies [31,32]
have not revealed other evidence of oxidative stress in
these tissues during hibernation, these tissues warrant
further study. Carey et al. reported that the GSH/GSSG
ratio decreases 5-fold in intestinal mucosa during hiber-
nation in 13-lined ground squirrels (Spermophilus tride-
cemlineatus), an effect attributed to a 50% decrease in
GSSG reductase activity and subsequent increase in
GSSG [33].
Role of ascorbate as an antioxidant during arousal from
hibernation
Ascorbate is an essential water-soluble antioxidant
that has been shown to be an effective scavenger of
reactive oxygen species [18]. Although controversy over
a neuroprotective role of ascorbate stems from evidence
of pro-oxidant properties in vitro [34], the in vivo
findings reported here provide evidence of ascorbate
redistribution during arousal from hibernation and sug-
gest this antioxidant may protect tissues from oxidative
stress during arousal.
Frei et al. previously reported that in plasma, neither
protein thiols, a-tocopherol, nor urate can prevent initia-
tion of lipid peroxidation and formation of micromolar
concentrations of lipid hydroperoxides induced by
aqueous peroxyl radicals in human plasma [1]. Rather,
these antioxidants merely lower the rate at which lipid
peroxidation occurs. In contrast, ascorbate was found to
be the most effective aqueous-phase antioxidant under
experimental conditions of oxidative stress. Ascorbate not
only completely protected plasma lipids from peroxida-
tive damage, but also spared a-tocopherol, urate, and
bilirubin [1]. These data led Frei et al. to conclude that
ascorbate is an outstanding antioxidant in human plasma.
Ascorbate has selective antioxidant properties that
make it unique among the antioxidant network. For
example, ascorbate limits peroxynitrite formation by
reacting selectively with superoxide, which spares NO
signaling, and helps maintain enhanced vascular integrity
[35,36]. Interestingly, Haung et al. have suggested that
DHA may also provide vascular protection [5]. In the
brain, ascorbate selectively scavenges hydroxyl radicals
formed from hydrogen peroxide (H2O2) to prevent neu-
rotoxicity [37], while sparing H2O2 signaling pathways
[38]. Definitive assessment of neuroprotective effects of
ascorbate during arousal from hibernation, as well as the
relative roles of ascorbate and DHA, requires specific
inhibitors, which are not yet available. SVCT2 knockout
mice die at birth, in part from intraparenchymal brain
hemorrhage that cannot be accounted for by a scorbutic
effect [36].
Evidence of redistribution of ascorbate during arousal,
regardless of ascorbate redox state, supports our model
that enhanced antioxidant defense during hibernation is
an important cytoprotective component of hibernation
physiology. Hibernating animals tolerate extreme fluctu-
ation in brain and body temperature, cerebral blood flow,
and other physiological parameters associated with het-
erothermy [9]. Moreover, these animals endure a variety
of events including traumatic brain injury [8], hypoxia
[39], and oxygen-glucose deprivation [40] better than
homeothermic mammals such as rats and humans. Un-
derstanding neuroprotective aspects of hibernation phy-
siology thus has potential to translate into improved
therapies and strategies for treating cerebral ischemia,
brain injury, and neurological disease. Results reported
here demonstrate that ascorbate is distributed to tissues
equally well in either the reduced or oxidized state
without compromising concentrations of other water
soluble antioxidants. Thus, from a human therapeutic
perspective, where ascorbate must be provided exoge-
nously, administration of ascorbate or DHA should be
equally efficacious and distinguished only by patholo-
gical conditions that could influence distribution and
uptake. Moreover, our findings emphasize that depletion
of [Asc]p alone, without consideration of tissue levels of
ascorbate is a poor indicator of ascorbate availability. In
addition to reports for protection from reperfusion injury
by ascorbate, and DHA discussed above, there is ample
evidence for neuroprotection by other antioxidants in
other forms of central nervous system injury [41–43], as
well as in neurodegenerative diseases [44–46]. In the
human population, low plasma antioxidant activity is
associated with greater neurological impairment after
central nervous system insult [47] and a lifelong dietary
pattern poor in antioxidants correlates with a higher
propensity for neurological disease [48]. Moreover, low
dietary intake of ascorbate and other antioxidants is a risk
factor for morbidity and mortality from heart disease
[49], whereas ascorbate supplementation leads to im-
provement in endothelial function in a variety of cardiac
pathologies [50].
We show here that elevated plasma ascorbate levels
during hibernation provide an important source of anti-

Ascorbate distribution and redox state 519
oxidant in hibernating mammals. These findings should
guide use of ascorbate and other antioxidants, in combi-
nation with other protective factors found in hibernation
[10,51], in the treatment of heart disease and other
diseases of the periphery, as well as neurological disor-
ders, including cerebral ischemia and trauma.
Acknowledgments—The authors thank Shufen Wu for superb technicalassistance. This work was funded by NS-34115 and NS-40169 supportedby NINDS, NIMH, NCRR, NCMHD, and the NYU Ricciardi ResearchFund.
REFERENCES
[1] Frei, B.; England, L.; Ames, B.N. Ascorbate is an outstandingantioxidant in human blood plasma. Proc. Natl. Acad. Sci. USA86:6377–6381; 1989.
[2] Sciamanna, M.A.; Lee, C.P. Ischemia/reperfusion-induced injuryof forebrain mitochondria and protection from ascorbate. Arch.Biochem. Biophys. 305:215–224; 1993.
[3] Henry, P.T.; Chandy, M.J. Effect of ascorbic acid on infarct size inexperimental focal cerebral ischemia and reperfusion in a primatemodel. Acta. Neurochir. (Wien) 140:977–980; 1998.
[4] Ranjan, A.; Theodore, D.; Haran, R.D.; Chandy, M.J. Ascorbicacid and focal cerebral ischemia in a primate model. Acta. Neuro-chir. (Wien) 123:87–91; 1993.
[5] Huang, J.; Agus, D.B.; Winfree, C.J.; Kiss, S.; Mack, W.J.;McTaggart, R.A.; Choudhri, T.F.; Kim, L.J.; Mocco, J.; Pinsky,D.J.; Fox, W.D.; Israel, R.J.; Boyd, T.A.; Golde, D.W.; Connolly,E.S., Jr. Dehydroascorbic acid, a blood–brain barrier transportableform of vitamin C, mediates potent cerebroprotection in experi-mental stroke. Proc. Natl. Acad. Sci. USA 98:11720–11724; 2001.
[6] Perez-Pinzon, M.A.; Mumford, P.L.; Rosenthal, M.; Sick, T.J.Antioxidants, mitochondrial hyperoxidation and electrical recov-ery after anoxia in hippocampal slices. Brain Res. 754:163–170;1997.
[7] MacGregor, D. G.; Avshalumov, M. V.; Rice, M. E. Brain edemainduced by in vitro ischemia: causal factors and neuroprotection.J. Neurochem. 85:1402–1411; 2003.
[8] Zhou, F.; Zhu, X.; Castellani, R. J.; Stimmelmayr, R.; Perry, G.;Smith, M. A.; Drew, K. L. Hibernation, a model of neuroprotec-tion. Am. J. Pathol. 158:2145–2151; 2001.
[9] Frerichs, K.U.; Kennedy, C.; Sokoloff, L.; Hallenbeck, J.M. Localcerebral blood flow during hibernation, a model of natural tole-rance to ‘‘cerebral ischemia.’’J. Cereb. Blood Flow Metab.14:193–205; 1994.
[10] Drew, K.L.; Rice, M.E.; Kuhn, T.B.; Smith, M.A. Neuroprotectiveadaptations in hibernation: therapeutic implications for ischemia–reperfusion, traumatic brain injury and neurodegenerative dis-eases. Free Radic. Biol. Med. 31:563–573; 2001.
[11] Drew, K.L.; Osborne, P.G.; Frerichs, K.U.; Hu, Y.; Koren, R.E.;Hallenbeck, J.M.; Rice, M.E. Ascorbate and glutathione regula-tion in hibernating ground squirrels. Brain Res. 851:1–8; 1999.
[12] Tøien, O.; Drew, K.L.; Chao, M.L.; Rice, M.E. Ascorbate dynam-ics and oxygen consumption during arousal from hibernation inArctic ground squirrels. Am. J. Physiol. Reg. I 281:R1–R11; 2001.
[13] Osborne, P. G.; Hashimoto, M. State-dependent regulation of cor-tical blood flow and respiration in hamsters: response to hyper-capnia during arousal from hibernation. J. Physiol. 547: (Pt. 3)963–970; 2003.
[14] Messerschmidt, A.; Rossi, A.; Ladenstein, R.; Huber, R.; Bologne-si, M.; Gatti, G.; Marchesini, A.; Petruzzelli, R.; Finazzi-Agro, A.X-ray crystal structure of the blue oxidase ascorbate oxidase fromzucchini: analysis of the polypeptide fold and a model of the coppersites and ligands. J. Mol. Biol. 206:513–529; 1989.
[15] Messerschmidt, A.; Steigemann, W.; Huber, R.; Lang, G.; Kro-neck, P. M. X-ray crystallographic characterisation of type-2-depleted ascorbate oxidase from zucchini. Eur. J. Biochem.209:597–602; 1992.
[16] Rice, M.E.; Lee, E.J.; Choy, Y. High levels of ascorbic acid,not glutathione, in the CNS of anoxia-tolerant reptiles con-trasted with levels in anoxia-intolerant species. J. Neurochem.64:1790–1799; 1995.
[17] Rice, M.E. Ascorbate compartmentalization in the CNS. Neuro-tox. Res. 1:81–90; 1999.
[18] Rice, M.E. Ascorbate regulation and its neuroprotective role in thebrain. Trends Neurosci. 23:209–216; 2000.
[19] Meister, A. Glutathione–ascorbic acid antioxidant system in ani-mals. J. Biol. Chem. 269:9397–9400; 1994.
[20] Winkler, B. S.; Orselli, S. M.; Rex, T. S. The redox couple bet-ween glutathione and ascorbic acid: a chemical and physiologicalperspective. Free Radic. Biol. Med. 17:333–349; 1994.
[21] Fornai, F.; Saviozzi, M.; Piaggi, S.; Gesi, M.; Corsini, G. U.;Malvaldi, G.; Casini, A. F. Localization of a glutathione-depend-ent dehydroascorbate reductase within the central nervous systemof the rat. Neuroscience 94:937–948; 1999.
[22] Fornai, F.; Piaggi, S.; Gesi, M.; Saviozzi, M.; Lenzi, P.; Paparelli,A.; Casini, A. F. Subcellular localization of a glutathione-depen-dent dehydroascorbate reductase within specific rat brain regions.Neuroscience 104:5–31; 2001.
[23] Tsukaguchi, H.; Tokui, T.; Mackenzie, B.; Berger, U.V.; Chen,X.Z.; Wang, Y.; Brubaker, R.F.; Hediger, M.A. A family of mam-malian Na+-dependent l-ascorbic acid transporters. Nature399:70–75; 1999.
[24] Berger, U. V.; Lu, X. C.; Liu, W.; Tang, Z.; Slusher, B. S.; He-diger, M. A. Effect of middle cerebral artery occlusion on mRNAexpression for the sodium-coupled vitamin C transporter SVCT2in rat brain. J. Neurochem. 86:896–906; 2003.
[25] Agus, D.B.; Gambhir, S.S.; Pardridge, W.M.; Spielholz, C.; Base-lga, J.; Vera, J.C.; Golde, D.W. Vitamin C crosses the blood–brain barrier in the oxidized form through the glucose transporters.J. Clin. Invest. 100:2842–2848; 1997.
[26] Rumsey, S. C.; Daruwala, R.; Al-Hasani, H.; Zarnowski, M. J.;Simpson, I. A.; Levine, M. Dehydroascorbic acid transport byGLUT4 in Xenopus oocytes and isolated rat adipocytes. J. Biol.Chem. 275:28246–28253; 2000.
[27] Padayatty, S.J.; Levine, M. New insights into the physiology andpharmacology of vitamin C. Can. Med. Assoc. J. 164:353–355;2001.
[28] Chatterjee, I. B.; Majumder, A. K.; Nandi, B. K.; Subramanian, N.Synthesis and some major functions of vitamin C in animals. Ann.N.Y. Acad. Sci. 258:24–47; 1975.
[29] Ha, T.Y.; Otsuka, M.; Arakawa, N. Ascorbate indirectly stimulatesfatty acid utilization in primary cultured guinea pig hepatocytes byenhancing carnitine synthesis. J. Nutr. 124:732–737; 1994.
[30] Banhegyi, G.; Marcolongo, P.; Puskas, F.; Fulceri, R.; Mandl,J.; Benedetti, A. Dehydroascorbate and ascorbate transport inrat liver microsomal vesicles. J. Biol. Chem. 273:2758–2762;1998.
[31] Rivera, P. M.; Ma, Y. L.; Zhu, X.; Ross, A.; Castellani, R.; Smith,M. A.; Rice, M. E.; Drew, K. L. Study of oxidative stress in CA1neurons and liver during euthermia, torpor and arousal from tor-por in arctic ground squirrels (Spermophilus parryii). In: 3rdAnnual Conference of Specialized Programs in Neuroscience,Honolulu, Hawaii; May 28–30, 2003.
[32] Orr, A.; Rivera, P. M.; Drew, K. L.; Hermes-Lima, M. Is arousalfrom hibernation stress free? In: 3rd Annual Conference of Spe-cialized Centers in Neuroscience, Honolulu, Hawaii; May 27–29,2003: 39.
[33] Carey, H. V.; Rhoads, C. A.; Aw, T. Y. Hibernation induces glu-tathione redox imbalance in ground squirrel intestine. J. Comp.Physiol. B 173:269–276; 2003.
[34] Lee, S. H.; Oe, T.; Blair, I. A. Vitamin C-induced decompositionof lipid hydroperoxides to endogenous genotoxins. Science292:2083–2086; 2001.
[35] Maeda, N.; Hagihara, H.; Nakata, Y.; Hiller, S.; Wilder, J.; Red-dick, R. Aortic wall damage in mice unable to synthesize ascorbicacid. Proc. Natl. Acad. Sci. USA 97:841–846; 2000.
[36] Sotiriou, S.; Gispert, S.; Cheng, J.; Wang, Y.; Chen, A.; Hoog-straten-Miller, S.; Miller, G.; Kwon, O.; Levine, M.; Guttentag, S.;Nussbaum, R. Ascorbic-acid transporter Slc23a1 is essential for

Y. L. Ma et al.520
vitamin C transport into the brain and for perinatal survival. Nat.Med. 8:514–517; 2002.
[37] Avshalumov, M. V.; Chen, B. T.; Rice, M. E. Mechanismsunderlying H2O2-mediated inhibition of synaptic transmissionin rat hippocampal slices. Brain Res. 882:86–94; 2000.
[38] Avshalumov, M. V.; Chen, B. T.; Marshall, S. P.; Pena, D. M.;Rice, M. E. Glutamate-dependent inhibition of dopamine releasein striatum is mediated by a new diffusible messenger, H2O2.J. Neurosci. 23:2744–2750; 2003.
[39] D’Alecy, L. G.; Lundy, E. F.; Kluger, M. J.; Harker, C. T.; LeMay,D. R.; Shlafer, M. Beta-hydroxybutyrate and response to hypoxiain the ground squirrel, Spermophilus tridecimlineatus. Comp. Bi-ochem. Physiol. B 96:189–193; 1990.
[40] Frerichs, K. U.; Hallenbeck, J. M. Hibernation in ground squirrelsinduces state and species-specific tolerance to hypoxia and agly-cemia: an in vitro study in hippocampal slices. J. Cereb. BloodFlow Metab. 18:168–175; 1998.
[41] Yang, Y.; Li, Q.; Shuaib, A. Neuroprotection by 2-h postischemiaadministration of two free radical scavengers, alpha-phenyl-n-tert-butyl-nitrone (PBN) and N-tert-butyl-(2-sulfophenyl)-nitrone (S-PBN), in rats subjected to focal embolic cerebral ischemia. Exp.Neurol. 163:39–45; 1993.
[42] Juurlink, B.H.; Paterson, P.G. Review of oxidative stress inbrain and spinal cord injury: suggestions for pharmacologicaland nutritional management strategies. J. Spinal Cord Med.21:309–334; 1998.
[43] Knuckey, N.W.; Palm, D.; Primiano, M.; Epstein, M.H.; Johanson,C.E. N-Acetylcysteine enhances hippocampal neuronal survivalafter transient forebrain ischemia in rats. Stroke 26:305–310; 1995.
[44] Fahn, S. A pilot trial of high-dose alpha-tocopherol and ascor-bate in early Parkinson’s disease. Ann. Neurol. 32:S128–S132;1992.
[45] Sano, M.; Ernesto, C.; Thomas, R.G.; Klauber, M.R.; Schafer, K.;Grundman, M.; Woodbury, P.; Growdon, J.; Cotman, C.W.;Pfeiffer, E.; Schneider, L.S.; Thal, L.J. The Alzheimer’s DiseaseCooperative Study. A controlled trial of selegiline, alpha-toco-pherol, or both as treatment for Alzheimer’s disease. N. Engl. J.Med. 336:1216–1222; 1997.
[46] Chan, P. Reactive oxygen radicals in signaling and damage in theischemic brain. J. Cereb. Blood Flow Metab. 21:2–14; 2001.
[47] Leinonen, J.S.; Ahonen, J.P.; Lonnrot, K.; Jehkonen, M.; Dastidar,P.; Molnar, G.; Alho, H. Low plasma antioxidant activity isassociated with high lesion volume and neurological impairmentin stroke. Stroke 31:33–39; 2000.
[48] Smith, M.A.; Petot, G.J.; Perry, G. Diet and oxidative stress: anovel synthesis of epidemiological data on Alzheimer’s disease. J.Alzheimer’s Dis. 1:203–206; 1999.
[49] Joshipura, K. J.; Hu, F. B.; Manson, J. E.; Stampfer, M. J.;Rimm, E. B.; Speizer, F. E.; Colditz, G.; Ascherio, A.; Rosner,B.; Spiegelman, D.; Willett, W. C. The effect of fruit and veg-etable intake on risk for coronary heart disease. Ann. Intern.Med. 134:1106–1114; 2001.
[50] Erbs, S.; Gielen, S.; Linke, A.; Mobius-Winkler, S.; Adams, V.;Baither, Y.; Schuler, G.; Hambrecht, R. Improvement of peripheralendothelial dysfunction by acute vitamin C application: differenteffects in patients with coronary artery disease, ischemic, and di-lated cardiomyopathy. Am. Heart J. 146:280–285; 2003.
[51] Carey, H. V.; Andrews, M. T.; Martin, S. T. Mammalian hiberna-tion: cellular and molecular responses to depressed metabolismand low temperature. Physiol. Rev. 83:1153–1181; 2003.
ABBREVIATIONS
AGS—arctic ground squirrels
AO—L-ascorbate oxidase
Asc—ascorbate
[Asc]p—ascorbate concentration in plasma
DHA—dehydroascorbate
GSH—glutathione
GSSG—oxidized glutathione
MCA—monochloroacetic acid
[Urate]p—urate concentration in plasma




















