
Role of perforin in controlling B-cell hyperactivity and humoral autoimmunity

Apoptosis Regulators Fas and Bim Cooperate in Shutdown ofChronic Immune Responses and Prevention of Autoimmunity
Peter D. Hughes1,2, Gabrielle T. Belz1, Karen A. Fortner3, Ralph C. Budd3, AndreasStrasser1,4∗, and Philippe Bouillet1,4∗∗1Molecular Genetics of Cancer, The Walter and Eliza Hall Institute of Medical Research, Melbourne, 3050,Australia.
2Department of Medical Biology, The University of Melbourne, Melbourne 3010, Australia.
3Immunobiology Program, The University of Vermont College of Medicine, Burlington, VT 05405-0068, USA.
SummaryApoptotic death of T lymphocytes is critical for shutdown of immune responses and hemopoieticcell homeostasis. Both death receptor (Fas) activation and mitochondrial apoptosis triggered by theBH3-only protein Bim have been implicated in the killing of antigen-stimulated T cells. We examinedmice lacking the gene encoding Bim (Bcl2l11) and with the inactivating lpr mutation in the geneencoding Fas (Fas), designated Bcl2l11−/−Faslpr/lpr mice. Shutdown of an acute T cell response toherpes simplex virus involved only Bim with no contribution by Fas, whereas both pathwayssynergized in killing antigen-stimulated T cells in chronic infection with murine γ-herpesvirus.Bcl2l11−/−Faslpr/lpr mice developed remarkably enhanced and accelerated fatal lymphadenopathyand autoimmunity compared to mice lacking only one of these apoptosis inducers. These resultsidentify critical overlapping roles for Fas and Bim in T cell death in immune response shutdown andprevention of immunopathology and thereby resolve a long-standing controversy.
KeywordsMOLIMMUNO
IntroductionIn response to infection or immunization, T cells that express antigen-specific T cell receptors(TCR) become activated and proliferate, and some differentiate into effector cells (Sprent andTough, 2001). Activated T cells then produce cytokines that help coordinate the immuneresponse aimed at eliminating the pathogen. Clearance of the antigen is accompanied by theshutdown of T cell immune responses and involves apoptosis of a large fraction of antigen-activated T cells. This prevents accumulation of no-longer-needed and potentially dangerouseffector cells and thereby precludes immunopathology. Two distinct but ultimately convergingpathways control apoptosis initiation in mammalian cells (Strasser et al., 1995). Ligation of“death receptors,” members of the tumor necrosis factor receptor (TNF-R) family with an
© 2008 Elsevier Inc.∗Corresponding author [email protected]. ∗∗Corresponding author [email protected] authors contributed equally to this work.This document was posted here by permission of the publisher. At the time of deposit, it included all changes made during peer review,copyediting, and publishing. The U.S. National Library of Medicine is responsible for all links within the document and for incorporatingany publisher-supplied amendments or retractions issued subsequently. The published journal article, guaranteed to be such by Elsevier,is available for free, on ScienceDirect.
Sponsored document fromImmunity
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

intracellular “death domain” (e.g., Fas or the tumor necrosis factor receptor 1 [TNF-R1])triggers formation of the “death-inducing signaling complex” (DISC), which promotes celldemolition through activation of caspase-8 (Ashkenazi and Dixit, 1998; Krammer, 2000;Wallach et al., 1998). Conversely, the “mitochondrial” (also called “intrinsic” or “B celllymphoma [Bcl-2]-regulated”) apoptotic pathway, activated by developmental cues orcytotoxic stimuli, such as cytokine deprivation, is independent of caspase-8 and Fas-associateddeath domain protein (FADD), but instead involves mitochondrial release of cytochrome c,which promotes Apaf-1-mediated activation of caspase-9 (Green and Kroemer, 2004; Wang,2001). This pathway is controlled by the Bcl-2 protein family, which consists of threesubgroups: the Bcl-2-like prosurvival proteins essential for cell survival; the Bax and Bak-likeproteins required for mitochondrial outer membrane disruption, release of apoptogenic proteins(e.g., cytochrome c), and activation of the caspase cascade (Green and Kroemer, 2004; Wang,2001); and the proteins containing only the Bcl-2 homology domain 3 (BH3-only proteins,e.g., Bim), which are activated by distinct death stimuli and are essential for initiation ofapoptosis signaling (Huang and Strasser, 2000).
Both death receptor signaling and the mitochondrial apoptotic pathway have been shown to becritical for lymphocyte homeostasis (Krammer, 2000; Sprent and Tough, 2001). Mice(Watanabe-Fukunaga et al., 1992) or humans (ALPS, autoimmune lymphoproliferationsyndrome patients) (Rieux-Laucat et al., 1995) defective for Fas (Faslpr/lpr mice) or its ligand,FasL (Faslgld/gld mice), develop progressive lymphadenopathy and splenomegaly resultingfrom accumulation of excess mature T and B cells as well as “unusual”αβTCR+CD4−CD8−B220+ T cells. Bcl-2-deficient mice (Veis et al., 1993) and mice lackingmyeloid cell leukemia 1 (Mcl-1) in the hematopoietic system (Opferman et al., 2005) exhibitprofound lymphopenia, whereas mice overexpressing a Bcl-2 transgene throughout thehematopoietic compartment show marked lymphadenopathy (Ogilvy et al., 1999). Micelacking the BH3-only protein Bim also accumulate excess lymphocytes and myeloid cells andthese cells are abnormally resistant to cytokine deprivation, deregulated calcium flux, and ERstress (Bouillet et al., 1999; Puthalakath et al., 2007). Moreover, Bim is critical for apoptosisduring negative selection of autoreactive thymocytes (Bouillet et al., 2002), mature T cells(Davey et al., 2002), and B cells (Enders et al., 2003).
The relative contributions of the two distinct apoptotic pathways in the termination of T cellimmune responses has been a matter of controversy for a long time. FasL and Fas have beenimplicated because activation-induced cell death (AICD), an in vitro model in which mitogen-activated T cell blasts are killed by TCR restimulation, which causes FasL upregulation, isinhibited by FasL or Fas inactivation (Brunner et al., 1995; Dhein et al., 1995; Russell et al.,1993). Moreover, clearance of Staphylococcus Enterotoxin B (SEB)-activated TCR Vβ8+ Tcells was reported to depend partially on Fas (Mogil et al., 1995; Van Parijs et al., 1996). Themitochondrial pathway has been implicated in immune response shutdown, because Bcl-2overexpression (Strasser et al., 1991), loss of Bim (Hildeman et al., 2002; Pellegrini et al.,2003), or Bax and Bak deficiency (Rathmell et al., 2002) inhibited death of T cells stimulatedin vitro or in vivo by a single dose of SEB or in vivo after infection with human herpes simplexvirus (HSV-1). It is conceivable that Fas and Bim may have overlapping roles in T cell killing,with their relative contribution determined by the nature of the immune response (acute versuschronic). In acute immune responses, apoptosis is triggered by the drop in cytokine amounts,after clearance of the pathogen or injected immunogen (Hildeman et al., 2002; Krammer, 2000;Sprent and Tough, 2001). A single administration of SEB, which is known to be eliminatedquickly from the body (Vabulas et al., 1996), would therefore mimic an acute infection. Incontrast, TCR restimulation of activated T cells in vitro (AICD) or repeated administration ofSEB to mice is more likely to imitate the repeated TCR activation that is thought to occurduring chronic immune responses, such as persistent infections or stimulation with self-antigens (Krammer, 2000; Sprent and Tough, 2001).
Hughes et al. Page 2
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

The removal of effector T lymphocytes prevents immunopathology, but the mechanisms forapoptosis induction remain contentious (Krammer, 2000; Sprent and Tough, 2001). In thepresent study, we have generated mice lacking Bim and harboring the inactivating lpr mutationin the Fas gene, and we used acute and chronic viral infection models to decipher the individualand combined roles of Fas and Bim in the shutdown of T cell immune responses and inimmunopathology.
ResultsBcl2l11−/−Faslpr/lpr Lymphoid Cells Are Resistant to Both Fas-Transduced as well as Bim-Dependent Apoptotic Stimuli
To investigate the functional interaction of Bim and Fas in the control of lymphocytehomeostasis, we generated mice lacking both of these apoptosis inducers (with both parentalstrains backcrossed to C57BL/6 >20 generations prior to intercrossing). Bim is required forthe killing of lymphocytes induced by growth factor withdrawal, deregulated calcium flux,DNA damage, or treatment with dexamethasone (Bouillet et al., 1999). Bcl2l11−/− thymocytesare, however, normally sensitive to treatment with phorbol myristyl acetate (PMA, a deathstimulus that can be countered by Bcl-2 overexpression) or FasL, which triggers the deathreceptor pathway (Bouillet et al., 1999). To determine the consequences of the combinedmutations on cell survival, we sorted several lymphocyte populations and subjected them inculture to cytokine deprivation, treatment with FasL, the calcium ionophore ionomycin, PMA,or the DNA damage-inducing drug etoposide. CD4+CD8+ thymocytes (Figure 1A) as well asmature B cells (see Figures S1E–S1G available online) and T cells (CD4+, CD8+, andαβTCR+CD4−CD8−B220+; Figures 1B and 1C; Figure S1) from Bcl2l11−/−Faslpr/lpr micewere resistant to both FasL as well as Bim-dependent apoptotic stimuli, such as growth factorwithdrawal or treatment with ionomycin. Thymocytes remained, however, normally sensitiveto PMA (Figure 1), which was shown to require the BH3-only protein Puma for cell killing(Villunger et al., 2003). These results demonstrate that Fas signaling and Bim exertnonredundant roles in the apoptosis of hemopoietic cells.
Clearance of HSV-Specific CD8+ T Cells in Acute Infection Requires the BH3-Only ProteinBim but Is Fas Independent
Although the importance of Fas and Bim in lymphocyte homeostasis is widely accepted, theirrespective roles in immune response shutdown remain controversial (Hildeman et al., 2002;Krammer, 2000; Sprent and Tough, 2001). We addressed the existing controversy bymeasuring the numbers of antigen-stimulated CD8+ T cells in spleen and lymph nodes duringacute and chronic immune responses in Bcl2l11−/−Faslpr/lpr as well as Bcl2l11+/−Faslpr/lpr,Faslpr/lpr, Bcl2l11−/−, and WT mice (Figures 2A–2D and 3A–3D). Consistent with the acutenature of an HSV-1 (HSV1 KOS) infection and the ensuing immune response (Pellegrini et al.,2003), all mice were able to clear the virus rapidly (not shown). Previous studies have shownthat the number of HSV-specific CD8+ T cells in the spleen of WT and Faslpr/lpr mice peaksat day 7, after which the majority of these effectors are eliminated by cell death (Pellegriniet al., 2003). Consistent with this, flow cytometric analysis showed that the spleens of WT andFaslpr/lpr mice contained only low numbers (<106) of HSV-specific CD8+ T cells on days 17and 35 after HSV infection (Figures 2A and 2B). In contrast, in Bcl2l11−/− mice, the numbersof HSV-specific CD8+ T cells in the spleen did not fall substantially after pathogen clearance(Figure 2E). Notably, the additional loss of Fas (in Bcl2l11−/−Faslpr/lpr mice) did not furtherincrease the accumulation of viral antigen-stimulated CD8+ T cells (Figure 2E). HSV-specificCD8+ cell numbers were also similar in nonlymphoid tissues (lungs and liver) of Bcl2l11−/−
and Bcl2l11−/−Faslpr/lpr mice 17 and 35 days after infection with HSV-1 (Figure S2). Theseresults demonstrate that Fas activation, which requires repeated TCR stimulation to upregulate
Hughes et al. Page 3
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

FasL in T cells, is not involved in the shutdown of acute immune responses; instead, this deathis mediated predominantly by Bim.
Bim and Fas Cooperate to Eliminate MHV-OVA-Specific T Cells in Chronic InfectionBoth mitochondrial as well as Fas-mediated apoptosis have been hypothesized to mediate thedeath of CD4+ as well as CD8+ T cells during chronic antigenic stimulation (Krammer, 2000;Sprent and Tough, 2001). We therefore examined the individual and combined roles of Bimand Fas in the T cell immune response to mouse γ-herpes virus (MHV) transgenicallyexpresssing ovalbumin (MHV 68-OVA) (Figure 3). This virus causes a chronic infectionpredominantly in the lung and B cells, with persistent antigen presentation in the lung(Figure S3; Rajcani and Kudelova, 2005). Total numbers of leukocytes as well as CD8+ T cellswere determined 14, 40, and 60 days after infection (Figures 3A–3D). Surface staining for viralantigen (ovalbumin)-specific TCR (Figures 3E and 3F) or intracellular staining for IFN-γproduction after brief antigenic (ovalbumin) stimulation in culture (Figures 3G and 3H)demonstrated that in spleens and draining lymph nodes of WT mice, MHV-OVA-specificCD8+ T cells (undetectable prior to infection) were greatly expanded on days 14 and 40 afterinfection, followed by a loss of nearly 50% of these cells by day 60. Loss of Bim resulted in asignificant (p < 0.05), albeit relatively small, increase in MHV-OVA-specific CD8+ T cells ondays 40 and 60, which was particularly prominent in the spleen (Figures 3E and 3G). Fas-deficient mice contained similarly increased numbers of these cells (Figures 3E and 3G; p <0.05). Remarkably, the combined absence of both apoptosis inducers had dramatic effects,causing a 43- or 63-fold increase of MHV-OVA-specific CD8+ T cells in the spleen comparedto WT mice on days 40 and 60, respectively. The increase of MHV-OVA-specific CD8+ Tcells in the mediastinal lymph nodes was even more spectacular, with an 83- or 845-folddifference compared to WT mice observed on days 40 and 60, respectively (Figures 3F and3H). This preferential increase of antigen-specific CD8+ T cells in the draining lymph nodesmight be a consequence of the ongoing antigen release from the lungs resulting in lymphocytehoming to this site. These results demonstrate that Fas-mediated death receptor signaling andthe Bim-mediated mitochondrial apoptotic pathway cooperate to prevent deregulatedexpansion of activated T cells during chronic antigenic stimulation.
Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr Mice Develop Extreme LymphadenopathyLoss of either Bim or Fas both lead to marked lymphadenopathy. Whereas Bcl2l11−/− miceshow an increase in most lymphoid and myeloid cell populations in the periphery (Bouilletet al., 1999), lymphadenopathy in Faslpr/lpr mice is due in large part to the accumulation ofunusual αβTCR+CD4−CD8−B220+ T cells (Watanabe-Fukunaga et al., 1992). These unusualT cells are not found in WT or Bcl2l11−/− mice. Bcl2l11−/−Faslpr/lpr and evenBcl2l11+/−Faslpr/lpr mice developed lymphadenopathy and splenomegaly at a dramaticallyincreased magnitude and rate compared to mice lacking either apoptosis initiator alone(Figure 4; Figure S5). In particular, the αβTCR+CD4−CD8−B220+ T cell population wasincreased 4–5 times in the spleen of Bcl2l11−/−Faslpr/lpr mice compared to Faslpr/lpr mice.Precise quantification in the lymph nodes was made impossible by huge variations betweenindividual mice (one particular Bcl2l11−/−Faslpr/lpr mouse was found to have 12.7 g of lymphnodes; Figure S6), but very large lymph nodes were a hallmark of all Bcl2l11−/−Faslpr/lpr andBcl2l11+/−Faslpr/lpr mice (Figures S5A and S5B). Conventional CD4+ and CD8+ T cells, Bcells, and, unexpectedly, granulocytes were also markedly increased in spleens and lymphnodes of Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr mice compared to WT mice or animalslacking only one of these apoptosis inducers (Figure 4). This shows that the Fas-activated deathreceptor and the Bim-mediated mitochondrial apoptotic pathways act synergistically to controlhomeostasis of all mature lymphocyte subpopulations and granulocyte turnover.
Hughes et al. Page 4
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

The thymi of Bcl2l11−/−Faslpr/lpr, Bcl2l11+/−Faslpr/lpr, Bcl2l11−/−, Faslpr/lpr, and WT micewere all similar in size and cellularity. The distribution of the four major cellular subsets(defined by the expression of CD4 and CD8) within the thymus of Bcl2l11−/−Faslpr/lpr micewas comparable to that of Bcl2l11−/− mice (Bouillet et al., 1999), with a reduction inCD4+CD8+ cells and a 2- to 3-fold increase in CD4−CD8− as well as mature (CD4+CD8− andCD4−CD8+) T cells (Figure S4). Thus, loss of Bim and Fas do not cooperate to causeabnormalities in the thymus, demonstrating that the enhanced lymphadenopathy andsplenomegaly seen in Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr mice are a consequence ofdefects in peripheral lymphoid organs.
Lymphadenopathy in Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr Mice Is Not Associatedwith Lymphoid Malignancy
Deregulation of apoptosis is thought to contribute to the development of many, perhaps all,types of cancer (Hanahan and Weinberg, 2000). We have previously shown that loss of Bimaccelerates lymphoma development in mice expressing a Myc transgene in their B cellcompartment (Egle et al., 2004). Because the dramatic expansion of the lymphocytecompartment in Bcl2l11−/−Faslpr/lpr and even the Bcl2l11+/−Faslpr/lpr mice was due to thedisruption of apoptotic pathways, we examined whether the T cells accumulating in theseanimals were monoclonal and hence most likely lymphomatous. However, immunofluorescentstaining with a panel of monoclonal antibodies to different TCR-Vβ subtypes demonstratedthat T cells from multiple lymph nodes of different Bcl2l11−/−Faslpr/lpr mice had a normalpolyclonal repertoire of TCR specificities (Figure 5; Figure S7). To confirm the absence ofhematological (or other) malignancy in Bcl2l11−/−Faslpr/lpr mice, we verified that C57BL/6mice injected with 2 × 106 lymph node cells from several Bcl2l11−/−Faslpr/lpranimals did notdevelop tumors over a 12 month observation period (data not shown). These results show thatthe extreme lymphadenopathy and splenomegaly in Bcl2l11−/−Faslpr/lpr andBcl2l11+/−Faslpr/lpr mice are caused by the polyclonal expansion of nontransformedlymphocytes.
Autoimmune Pathology in Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr MiceBim (Davey et al., 2002) as well as Fas (Van Parijs et al., 1998) have been implicated in thekilling of T cells chronically activated by self-antigens, so it appears predictable that loss ofboth may synergize to cause severe immunopathology. Although Faslpr/lpr (Watanabe-Fukunaga et al., 1992) as well as Bcl2l11−/− (Bouillet et al., 1999) mice generally produceautoantibodies, they develop immune-complex deposits and fatal SLE-like autoimmunedisease only on particular genetic backgrounds (e.g., MRL for lpr mice or mixed C57BL/6x129SV for Bcl2l11−/− mice). On the C57BL/6 background, Bcl2l11−/− mice developlymphocyte infiltrates in lung, liver, and kidneys around 1 year of age, but these have nodiscernible consequences for their health. All Faslpr/lpr and Bcl2l11−/− mice examined between16 and 42 weeks were healthy, and histological examination did not show any signs of severeautoimmunity. In contrast, all Bcl2l11−/−Faslpr/lpr and even the Bcl2l11+/−Faslpr/lpr mice hadbecome moribund by 250 days of age, with a median survival of 114.5 days forBcl2l11−/−Faslpr/lpr mice and 159 days for Bcl2l11+/−Faslpr/lpr mice (Figure 6). Almost all ofthese animals had to be sacrificed because they presented with labored breathing resulting fromfilling of the chest cavity by large lymph nodes. Histological examination revealed the presenceof massive lymphocyte infiltrates in the lungs, kidneys, and liver of Bcl2l11−/−Faslpr/lpr andBcl2l11+/−Faslpr/lpr mice (Figure 6A). All moribund mice also tested positive for the presenceof anti-DNA autoantibodies (not shown). 20%–30% of the Bcl2l11−/−Faslpr/lpr and ∼10% ofthe Bcl2l11+/−Faslpr/lpr mice also presented with urticarial rash between the ears and in theneck area. In contrast, all Faslpr/lpr, Bcl2l11−/−, and WT mice survived well beyond 250 dayswithout symptoms. These observations demonstrate that Bim-dependent mitochondrial
Hughes et al. Page 5
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

apoptosis and Fas death receptor signaling cooperate in hematopoietic cell homeostasis andprevention of immunopathology.
DiscussionImmune responses to an infectious pathogen or injection of a foreign substance result in theclonal expansion of antigen-specific T cells and, upon pathogen or antigen clearance, this isfollowed by contraction of this T cell population (Sprent and Tough, 2001). The shutdown ofT cell immune responses is mediated by their programmed death (Kawabe and Ochi, 1991),but the mechanisms for apoptosis induction remain controversial (Hildeman et al., 2002;Krammer, 2000). This is (at least in part) due to the use of different model systems. Culturingantigen- or mitogen-activated T cells with IL-2 and restimulating them through the TCR orwith PMA plus ionomycin has long been used as the standard in vitro model to mimic theremoval of activated T cells in vivo (Krammer, 2000). Experiments with blocking antibodiesor FasL- or Fas-deficient mutant mice demonstrated that in this setting, TCR restimulationcauses the expression of FasL, which induces autocrine and/or paracrine killing of Fas-expressing T cells (Brunner et al., 1995; Dhein et al., 1995; Russell et al., 1993). This led tothe conclusion that FasL-Fas signaling is responsible for the shutdown of T cell immuneresponses. Some studies showed that deletion of SEB-activated T cells in mice was in partFasL-Fas dependent (Mogil et al., 1995; Renno et al., 1996), whereas others found that Fasdeficiency (in Faslpr/lpr mutant mice) had no impact but that loss of Bim provided profoundprotection (Hildeman et al., 2002). These discrepancies might be due to differences in theregimes of SEB injection. This is supported by the finding that effects of Fas deficiency ondeletion of activated T cells in vivo were observed most consistently when mice wereadministered several doses of SEB (Strasser et al., 1995). Interestingly, the death of antigen-activated CD8+ T cells during shutdown of and acute immune response to a viral (HSV-1)infection was substantially inhibited by loss of Bim but was not affected by Fas deficiency(Pellegrini et al., 2003). We therefore reasoned that depending on the nature of an immuneresponse—acute versus chronic—Fas-mediated death receptor signaling and Bim-dependentmitochondrial apoptosis might contribute to different extents to the killing of activated T cells(Strasser and Pellegrini, 2004). Accordingly, a single low-dose challenge with SEB wouldmimic a response to an acute infection, whereas high-dose and/or repeated challenge with thissuperantigen would be expected to resemble the response to a chronic infection.
Our analysis of Bcl2l11−/−Faslpr/lpr mice demonstrates that killing of activated T cells duringtermination of a response to acute infection relies predominantly on Bim with no apparentcontribution by Fas death receptor signaling. In contrast, these two pathways synergize in thekilling of T cells during shutdown of an immune response to chronic infection, and they alsocooperate to maintain homeostasis of T cells, B cells, as well as granulocytes, and to preventautoimmune pathology. These results help reconcile current controversies regarding therelative contributions of the death receptor versus the mitochondrial apoptotic pathway to theshutdown of T cell immune responses. They are consistent with the notion that during shutdownof an acute immune response, killing of activated T cells is not elicited by repeated TCRligation, which would have been required for activation of the Fas pathway (Hildeman et al.,2002; Pellegrini et al., 2003). Instead, apoptosis requires the BH3-only protein Bim, which isthought to be activated by the drop in cytokine levels that occurs after pathogen clearance(Strasser and Pellegrini, 2004). This fits with the observations that Bim is critical for cytokinedeprivation-induced apoptosis (Bouillet et al., 1999) and transcriptionally (Dijkers et al.,2000) as well as post-translationally (reviewed in Ley et al., 2005) activated by this deathstimulus. It is possible that a drop in the levels of prosurvival Bcl-2 family members,particularly Bcl-2 (Hildeman et al., 2002), and/or the induction of the BH3-only protein Puma(encoded by the Bbc3 gene), which is also involved in growth factor withdrawal-inducedapoptosis of hemopoietic cells (Jeffers et al., 2003; Villunger et al., 2003), are also involved
Hughes et al. Page 6
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

in the killing of antigen-activated T cells during shutdown of an acute immune response.Indeed, Bim and Puma doubly deficient T cells survive better in culture in the absence ofcytokines than those lacking only Bim or Puma (Erlacher et al., 2006). Analysis of HSV-1-infected Bcl2l11−/−Puma−/− mice could help determine whether these two BH3-only proteinshave overlapping functions in the shutdown of acute T cell immune responses.
The finding that Fas-mediated death receptor signaling and the Bim-dependent mitochondrialapoptosis both contribute to shutdown of chronic immune responses is consistent with theobservations that loss of Bim (Grayson et al., 2006), Bcl-2 overexpression (Strasser et al.,1995), or loss of Fas (Kawabe and Ochi, 1991), each on their own, enhanced (to a relativelysmall extent) survival of T cells chronically stimulated by repeated SEB injection or LCMVinfection. These results are consistent with the notion that the death receptor and themitochondrial apoptotic pathways are distinct and that they can cooperate in the removal of acell population (Strasser et al., 1995). Our results are reminiscent of the effects observed uponthe overexpression of Bcl-2 in Faslpr/lpr mice. It is highly probable that Bcl-2 might havecountered Bim function in these mice. The importance of the genetic background on theseverity of the Faslpr/lpr and Bcl2l11−/− phenotypes is well established. Direct comparison ofthe Bcl2-tg Faslpr/lpr mice and our present study is limited by the fact that the mice used in theformer study were on the C3H/HeJ background, whereas the Bcl2l11−/−Faslpr/lpr mice are onthe C57BL/6 background. It is, however, possible that other BH3-only proteins may beinvolved in the killing of chronically activated T cells, and lymphadenopathy and splenomegalymay therefore be even more severe in Bcl2-tg Faslpr/lpr mice than in Bcl2l11−/−Faslpr/lpr mice.
It appears likely that repeated TCR stimulation causes activation of the FasL-Fas signalingpathway in T cells during chronic infection (Brunner et al., 1995; Dhein et al., 1995), but howBim is induced in this setting is less clear. This might be a consequence of a drop in the amountsof cytokines, similar to shutdown of acute immune responses. Alternatively, the calcium fluxelicited by repeated TCR ligation might activate Bim (Bouillet et al., 2002), and, pertinently,Bim is essential for deregulated calcium flux-induced apoptosis (Bouillet et al., 1999).
Combined loss of Fas and Bim also synergized in causing lymphadenopathy, demonstratingthat these two pathways cooperate not only in the killing of T cells chronically activated bypathogen-borne antigens but also those stimulated by self-antigens. The unusualαβTCR+CD4−CD8−B220+ T lymphoid cells are seen only in Faslpr/lpr but not in Bcl2l11−/−
mice, consistent with the view that they are descendants of chronically activated“conventional” T cells, which previously expressed CD4 or CD8 (Renno et al., 1996) andwould normally be killed by FasL-Fas signaling (Brunner et al., 1995; Dhein et al., 1995).These unusual T cells are as sensitive as conventional T cells to cytokine deprivation but canbe protected by loss of Bim, providing a likely explanation as to why their numbers are greatlyincreased in Bcl2l11−/−Faslpr/lpr and even Bcl2l11+/−Faslpr/lpr mice compared to animalslacking only Fas. Although loss of either Fas or Bim causes fatal autoimmune disease on certaingenetic backgrounds (e.g., MRL for Faslpr/lpr [Watanabe-Fukunaga et al., 1992] or mixedC57BL/6x129SV for Bcl2l11−/− [Bouillet et al., 1999]), such pathology is not apparent foreither mutation on an inbred C57BL/6 background. The immunopathology seenin Bcl2l11−/−Faslpr/lpr and Bcl2l11+/−Faslpr/lpr mice on this background thereforedemonstrates that these two apoptotic pathways have overlapping functions in the preventionof autoimmune disease.
These findings have implications for clinical medicine. For example, abnormally increasedlevels of Bim and enhanced sensitivity to FasL (Gougeon, 2003) have been detected inleukocytes from patients infected with human immunodeficiency virus (HIV) or otherpathogens. Thus, Bim- and Fas-mediated apoptosis may cooperate to cause the excessivekilling of antigen-specific T cells that is thought to impair clearance of the infectious microbes
Hughes et al. Page 7
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

in these settings. Conversely, the inability to kill T cells that are chronically stimulated by self-antigens might underlie some autoimmune conditions. For example, because loss of one alleleof Bim synergizes with Fas deficiency to cause immunopathology, it is possible thatcombinations of mutant alleles of Bim and Fas, which by themselves do not cause readilyidentifiable abnormalities, may underlie certain autoimmune diseases in humans.
In summary, two major pathways regulate the life and death of lymphocytes. The death receptorpathway involves membrane receptors of the TNFR1 family, whereas the mitochondrialpathway is regulated by proteins of the Bcl-2 family. Here, we show that both pathwayssynergize to kill activated T cells in chronic immune responses, whereas death of activated Tcells in acute immune responses relies only on the mitochondrial pathway.
Experimental ProceduresMice
All animal experiments were performed in accordance with the guidelines of the MelbourneDirectorate Animal Ethics Committee. Bcl2l11−/− and Faslpr/lpr mice had been backcrossedonto a C57BL/6 genetic background for >20 generations prior to intercrossing and were housedin specific pathogen-free (SPF) conditions prior to infectious challenge.
Immunofluorescent Staining, FACS Analysis, Cell Sorting, Tissue Culture, and Cell-SurvivalAssays
Single-cell suspensions were prepared from thymus, spleen, and lymph nodes (axillary,brachial, and inguinal) of 6- to 10-week-old mice and total leukocyte numbers counted.Leukocyte subpopulations were identified by immunofluorescent staining with surface marker-specific mAbs coupled to FITC, R-PE, APC, Cy5, or biotin (the latter revealed by stainingwith streptavidin coupled to R-PE or Tricolor; Caltag) followed by flow cytometric analysisin a FACScan (Becton Dickinson) or sorting in a FACS DIVA (Becton Dickinson) or MoFlo(Cytomation). Sorted cells were resuspended in DMEM supplemented with 10% FCS, 50 μM2-mercaptoethanol (2-ME), and 100 μM asparagine and cultured at 37°C in 96-well flat-bottomplates. Cells were incubated in medium alone (no treatment, i.e., cytokine deprivation) ortreated with FLAG-tagged human recombinant FasL 10 ng/mL (Alexis) crosslinked with 1μg/mL M2 FLAG monoclonal antibody (Sigma), ionomycin (1 μg/mL), PMA (10 ng/mL),etoposide 1 μg/mL, or γ-irradiated (1.25 Gy). Cell survival was quantified daily by stainingwith propidium iodide (PI; 2 μg/mL) and FACS analysis.
HSV Infection and Quantification of HSV Antigen-Specific CD8+ T CellsMice were injected subcutaneously (s.c.) in each hind foot with 4 × 105 plaque-forming units(PFU) of HSV-1 KOS strain under general anesthesia. After 17 and 35 days, mice were killed,and popliteal lymph nodes, spleen, liver, lungs, and feet were removed. Single-cell suspensionswere prepared from these tissues, and those from the lymph nodes of mice of the samegenotypes were pooled. Lymphocytes were purified from liver and lung on Percoll densitygradients. To measure the percentages of viral antigen-specific T cells, single-cell suspensionsfrom spleen and lymph nodes were first enriched for CD8+ T cells with magnetic bead depletionby staining with M1/70 (anti-Mac-1), F4/80 (macrophage marker), Ter 119 (erythroid marker),RB6-8C5 (anti-Gr-1), M5/114 (anti-class II MHC), and GK 1.5 (anti-CD4) antibodies followedby incubation with goat anti-rat IgG antibody-coupled magnetic beads (Dynal, Oslo andQIAGEN). Enriched CD8+ cells were washed and stained for 45 min at room temperature withPE-streptavidin-conjugated Kb-gB tetramers containing the immuno-dominant HSVgB498-505 peptide. Cells were then stained for 30 min on ice with APC-conjugated CD8 mAb(53-6.7) in balanced salt solution containing 2% FBS. PI staining was used to exclude dead
Hughes et al. Page 8
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

cells and data were acquired on at least 104 viable cells with a FACSCalibre flow cytometerand analyzed with CellQuest Pro (BD Biosciences).
MHV Infection and Quantification of Viral Antigen-Specific CD8+ T Cells by Staining with PE-Streptavidin-Conjugated Kb-OVA Tetramers
Mice were infected under brief general anesthesia intranasally with 3 × 104 PFU of recombinantmurine γ-herpesvirus that coexpressed ovalbumin (OVA) from an intergenic expressioncassette driven by an ectopic MHV-68 lytic promoter (MHV 68-OVA). Infected mice werekilled after 14, 40, or 60 days, and lungs, mediastinal lymph nodes, and spleen were collected.Single-cell suspensions were prepared and leukocytes within the lymph nodes and spleen werecounted. Cells collected from the lymph nodes from mice of the same genotype were pooled.Total numbers of CD4+ and CD8+ T cells were measured by staining cells for 30 min on icewith PE-coupled anti-CD4 mAb (CT-CD4) and FITC-coupled anti-CD8 mAb (CT-CD8), andanalysis was performed as described above. To enumerate viral antigen-specific T cells,suspensions from lymph nodes and spleen were enriched for CD8+ T cells as described above,labeled for 30 min on ice with FITC-coupled anti-CD8 mAb, washed, stained for 45 min atRT with PE-coupled streptavidin conjugated to Kb class I MHC tetramers loaded withSIINFEKL peptide, and then analyzed as described above.
Intracellular FACS Analysis of Cytokine ProductionApproximately 1 × 106 cells from lymph nodes or spleen of Bcl2l11−/−Faslpr/lpr,Bcl2l11+/−Faslpr/lpr, Faslpr/lpr, Bcl2l11−/−, and WT mice were incubated for 5 hr at 37°C inRPMI 1640 supplemented with 50 μM 2-ME, 10% FBS, 5 μg/mL brefeldin A (which was alsoprovided in all subsequent washes and incubations), and 10 U/mL recombinant mouse IL-2plus either 1 μM HSV gB498-505 peptide for mice infected with HSV or 1 μMOvalbumin257-265 (SIINFEKL) peptide for mice infected with MHV-OVA or, as a control,with no peptide. Cells were harvested, washed, stained for 30 min on ice with FITC-coupledanti-CD8 mAb, washed again, fixed for 15 min at 4°C with 1% formalin, permeabilized for10 min at RT in PBS containing 0.5% saponin and 1% BSA, stained for 30 min on ice withPE-coupled rat anti-IFN-γ mAb (C1 XMG1.2), washed, and analyzed on a LSR orFACSCalibre flow cytometer. The frequency of virus-specific CD8+ T cells was calculated bysubtracting the frequency of CD8+ IFN-γ+ T cells in samples stimulated with no peptide fromthe frequency of CD8+ IFN-γ+ T cells in samples with the antigenic peptide.
MHV-OVA Viral Titer AssaysTiters of infectious MHV-OVA virus were determined by plaque assay. In brief, 10-folddilutions of (virus containing) spleen cell suspensions from infected mice were incubated for2 hr on BHK-21 cell monolayers. The inoculum was then removed and the monolayers overlaidwith Dulbecco's modified Eagle medium containing 0.3% carboxymethylcellulose. After 5days, the monolayers were fixed in 10% formaldehyde and stained with 10% Giemsa (Merck)for 30 min before rinsing with water. Monolayers were dried at room temperature overnightbefore plaques were counted with a plate microscope.
Analysis of Persisting Antigen Presentation during MHV-OVA InfectionPersistent antigen presentation was evaluated by analyzing the induction of proliferation ofCFSE-labeled OVA-specific CD8+ T cells (from OT-I TCR transgenic mice) after injectioninto mice that had been infected with MHV-OVA for various lengths of time. In brief,lymphocyte suspension from lymph nodes (inguinal, axillary, sacral, cervical, and mesenteric)of OT-I TCR transgenic mice were enriched for CD8+ T cells by incubation with antibodiesspecific for Mac-1 (M1/70), Mac-3 (F4/80), erythrocytes (Ter-119), B220 (RA3-6B2), MHCclass II (M5/114), and CD4 (GK1.5). Antibody-coated cells were removed with goat anti-rat
Hughes et al. Page 9
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

IgG antibody-coated magnetic beads (QIAGEN). Purity was verified by FACS analysis andwas routinely found to be ∼95%. Enriched CD8+ T cell preparations were resuspended at107 cells/mL in PBS containing 0.1% BSA and incubated for 10 min at 37°C with 5 μM CFSEstock solution (5 mM in DMSO, Molecular Probes). 1 × 106 CFSE-labeled OT-I TCRtransgenic T cells were injected into naive or MHV-OVA-infected mice, and proliferation wasanalyzed by FACS analysis after 3 days to determine the reduction in CFSE staining intensity.
TCR-Vβ Expression AnalysisTCR-Vβ expression was analyzed by preparing cell suspensions from multiple separate lymphnodes from Bcl2l11−/−Faslpr/lpr with massive lymphadenopathy (aged approximately 12weeks). Cells were stained with FITC-coupled Thy-1 mAb (T24.31.2) and biotinylated mAbsspecific for TCR Vβ3, 4, 5, 6, 7, 8, or 10 (BD PharMingen), followed by R-phycoerythrin (PE)-streptavidin. PI was used to exclude dead cells and cells were analyzed in a FACScan.
Supplemental DataRefer to Web version on PubMed Central for supplementary material.
Acknowledgments
We are grateful to J. Adams, S. Cory, A. Harris, and D. Vaux for advice and discussions. We thank J. Coughlin, G.Siciliano, J. Allen, and S. Shehata for expert animal care; B. Helbert and C. Young for genotyping; M. Robati fortechnical assistance; F. Battye, V. Lapatis, and C. Tarlinton for cell sorting; and S. Mihajlovic, E. Tsui, and A. Hasaneinfor histology. This work was supported by fellowships and grants from the National Health and Medical ResearchCouncil (Australia; #257502, #461263, and #356214), the Charles and Sylvia Viertel Foundation, the Leukemia andLymphoma Society (New York; SCOR grant #7015), the Wellcome Trust (UK), the Howard Hughes Medical Institute,and the National Cancer Institute (NIH, US; CA 80188 and CA 43540).
ReferencesAshkenazi and Dixit, 1998. Ashkenazi A. Dixit V.M. Death receptors: signaling and modulation. Science
1998;281:1305–1308. [PubMed: 9721089]Bouillet et al, 1999. Bouillet P. Metcalf D. Huang D.C.S. Tarlinton D.M. Kay T.W.H. Köntgen F. Adams
J.M. Strasser A. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocytehomeostasis, and to preclude autoimmunity. Science 1999;286:1735–1738. [PubMed: 10576740]
Bouillet et al, 2002. Bouillet P. Purton J.F. Godfrey D.I. Zhang L.-C. Coultas L. Puthalakath H. PellegriniM. Cory S. Adams J.M. Strasser A. BH3-only Bcl-2 family member Bim is required for apoptosisof autoreactive thymocytes. Nature 2002;415:922–926. [PubMed: 11859372]
Brunner et al, 1995. Brunner T. Mogil R.J. LaFace D. Yoo N.J. Mahboubi A. Echeverri F. Martin S.J.Force W.R. Lynch D.H. Ware C.F. Cell-autonomous Fas (CD95)/Fas-ligand interaction mediatesactivation-induced apoptosis in T-cell hybridomas. Nature 1995;373:441–444. [PubMed: 7530336]
Davey et al, 2002. Davey G.M. Kurts C. Miller J.F. Bouillet P. Strasser A. Brooks A.G. Carbone F.R.Heath W.R. Peripheral deletion of autoreactive CD8 T cells by cross presentation of self-antigenoccurs by a Bcl-2-inhibitable pathway mediated by Bim. J. Exp. Med. 2002;196:947–955. [PubMed:12370256]
Dhein et al, 1995. Dhein J. Walczak H. Baumler C. Debatin K.-M. Krammer P.H. Autocrine T-cell suicidemediated by APO-1/(Fas/CD95). Nature 1995;373:438–441. [PubMed: 7530335]
Dijkers et al, 2000. Dijkers P.F. Medema R.H. Lammers J.J. Koenderman L. Coffer P.J. Expression ofthe pro-apoptotic Bcl-2 family member Bim is regulated by the forkhead transcription factor FKHR-L1. Curr. Biol. 2000;10:1201–1204. [PubMed: 11050388]
Egle et al, 2004. Egle A. Harris A.W. Bouillet P. Cory S. Bim is a suppressor of Myc-induced mouse Bcell leukemia. Proc. Natl. Acad. Sci. USA 2004;101:6164–6169. [PubMed: 15079075]
Enders et al, 2003. Enders A. Bouillet P. Puthalakath H. Xu Y. Tarlinton D.M. Strasser A. Loss of thepro-apoptotic BH3-only Bcl-2 family member Bim inhibits BCR stimulation-induced apoptosis anddeletion of autoreative B cells. J. Exp. Med. 2003;198:1119–1126. [PubMed: 14517273]
Hughes et al. Page 10
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Erlacher et al, 2006. Erlacher M. Laabi V. Manzl C. Bock G. Tzankov A. Haecker G. Strasser A. VillungerA. Puma cooperates with Bim, the rate-limiting BH3-only protein in cell death during lymphocytedevelopment, in apoptosis induction. J. Exp. Med. 2006;203:2939–2951. [PubMed: 17178918]
Gougeon, 2003. Gougeon M.L. Apoptosis as an HIV strategy to escape immune attack. Nat. Rev.Immunol. 2003;3:392–404. [PubMed: 12766761]
Grayson et al, 2006. Grayson J.M. Weant A.E. Holbrook B.C. Hildeman D. Role of Bim in regulatingCD8+ T-cell responses during chronic viral infection. J. Virol. 2006;80:8627–8638. [PubMed:16912311]
Green and Kroemer, 2004. Green D.R. Kroemer G. The pathophysiology of mitochondrial cell death.Science 2004;305:626–629. [PubMed: 15286356]
Hanahan and Weinberg, 2000. Hanahan D. Weinberg R.A. The hallmarks of cancer. Cell 2000;100:57–70. [PubMed: 10647931]
Hildeman et al, 2002. Hildeman D.A. Zhu Y. Mitchell T.C. Bouillet P. Strasser A. Kappler J. MarrackP. Activated T cell death in vivo mediated by pro-apoptotic Bcl-2 family member, Bim. Immunity2002;16:759–767. [PubMed: 12121658]
Huang and Strasser, 2000. Huang D.C.S. Strasser A. BH3-only proteins—essential initiators of apoptoticcell death. Cell 2000;103:839–842. [PubMed: 11136969]
Jeffers et al, 2003. Jeffers J.R. Parganas E. Lee Y. Yang C. Wang J. Brennan J. MacLean K.H. Han J.Chittenden T. Ihle J.N. Puma is an essential mediator of p53-dependent and -independent apoptoticpathways. Cancer Cell 2003;4:321–328. [PubMed: 14585359]
Kawabe and Ochi, 1991. Kawabe Y. Ochi A. Programmed cell death and extrathymic reduction of Vβ8+CD4− T cells in mice tolerant to Staphylococcus aureus enterotoxin B. Nature 1991;349:245–248.[PubMed: 1670963]
Krammer, 2000. Krammer P.H. CD95's deadly mission in the immune system. Nature 2000;407:789–795. [PubMed: 11048730]
Ley et al, 2005. Ley R. Ewings K.E. Hadfield K. Cook S.J. Regulatory phosphorylation of Bim: sortingout the ERK from the JNK. Cell Death Differ. 2005;12:1008–1014. [PubMed: 15947788]
Mogil et al, 1995. Mogil R.J. Radvanyi L. Gonzalez-Quintial R. Miller R. Mills G. Theofilopoulos A.N.Green D.R. Fas (CD95) participates in peripheral T cell deletion and associated apoptosis in vivo.Int. Immunol. 1995;7:1451–1458. [PubMed: 7495753]
Ogilvy et al, 1999. Ogilvy S. Metcalf D. Print C.G. Bath M.L. Harris A.W. Adams J.M. Constitutivebcl-2 expression throughout the hematopoietic compartment affects multiple lineages and enhancesprogenitor cell survival. Proc. Natl. Acad. Sci. USA 1999;96:14943–14948. [PubMed: 10611317]
Opferman et al, 2005. Opferman J. Iwasaki H. Ong C.C. Suh H. Mizuno S. Akashi K. Korsmeyer S.J.Obligate role of anti-apoptotic MCL-1 in the survival of hematopoietic stem cells. Science2005;307:1101–1104. [PubMed: 15718471]
Pellegrini et al, 2003. Pellegrini M. Belz G. Bouillet P. Strasser A. Shut down of an acute T cell immuneresponse to viral infection is mediated by the pro-apoptotic Bcl-2 homology 3-only protein Bim.Proc. Natl. Acad. Sci. USA 2003;100:14175–14180. [PubMed: 14623954]
Puthalakath et al, 2007. Puthalakath H. O'Reilly L.A. Gunn P. Lee L. Kelly P.N. Huntington N.D. HughesP.D. Michalak E.M. McKimm-Breschkin J. Motoyama N. ER stress triggers apoptosis by activatingBH3-only protein Bim. Cell 2007;129:1337–1349. [PubMed: 17604722]
Rajcani and Kudelova, 2005. Rajcani J. Kudelova M. Murine herpesvirus pathogenesis: a model for theanalysis of molecular mechanisms of human gamma herpesvirus infections. Acta Microbiol.Immunol. Hung. 2005;52:41–71. [PubMed: 15957234]
Rathmell et al, 2002. Rathmell J.C. Lindsten T. Zong W.-X. Cinalli R.M. Thompson C.B. Deficiency inBak and Bax perturbs thymic selection and lymphoid homeostasis. Nat. Immunol. 2002;3:932–939.[PubMed: 12244308]
Renno et al, 1996. Renno T. Hahne M. Tschopp J. MacDonald H.R. Peripheral T cells undergoingsuperantigen-induced apoptosis in vivo express B200 and upregulate Fas and Fas ligand. J. Exp.Med. 1996;183:431–437. [PubMed: 8627156]
Rieux-Laucat et al, 1995. Rieux-Laucat F. Le Deist F. Hivroz C. Roberts I.A.G. Debatin K.M. FischerA. de Villartay J.P. Mutations in Fas associated with human lymphoproliferative syndrome andautoimmunity. Science 1995;268:1347–1349. [PubMed: 7539157]
Hughes et al. Page 11
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Russell et al, 1993. Russell J.H. Rush B. Weaver C. Wang R. Mature T cells of autoimmune lpr/lpr micehave a defect in antigen-stimulated suicide. Proc. Natl. Acad. Sci. USA 1993;90:4409–4413.[PubMed: 8506280]
Sprent and Tough, 2001. Sprent J. Tough D.F. T cell death and memory. Science 2001;293:245–248.[PubMed: 11452113]
Strasser and Pellegrini, 2004. Strasser A. Pellegrini M. T-lymphocyte death during shutdown of animmune response. Trends Immunol. 2004;25:610–615. [PubMed: 15489190]
Strasser et al, 1991. Strasser A. Harris A.W. Cory S. Bcl-2 transgene inhibits T cell death and perturbsthymic self-censorship. Cell 1991;67:889–899. [PubMed: 1959134]
Strasser et al, 1995. Strasser A. Harris A.W. Huang D.C.S. Krammer P.H. Cory S. Bcl-2 and Fas/APO-1regulate distinct pathways to lymphocyte apoptosis. EMBO J. 1995;14:6136–6147. [PubMed:8557033]
Vabulas et al, 1996. Vabulas R. Bittlingmaier R. Heeg K. Wagner H. Miethke T. Rapid clearance of thebacterial superantigen staphylococcal enterotoxin B in vivo. Infect. Immun. 1996;64:4567–4573.[PubMed: 8890208]
Van Parijs et al, 1996. Van Parijs L. Ibraghimov A. Abbas A.K. The roles of costimulation and Fas in Tcell apoptosis and peripheral tolerance. Immunity 1996;4:321–328. [PubMed: 8624822]
Van Parijs et al, 1998. Van Parijs L. Peterson D.A. Abbas A.K. The Fas/Fas ligand pathway and Bcl-2regulate T cell responses to model self and foreign antigens. Immunity 1998;8:265–274. [PubMed:9492007]
Veis et al, 1993. Veis D.J. Sorenson C.M. Shutter J.R. Korsmeyer S.J. Bcl-2-deficient mice demonstratefulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell 1993;75:229–240.[PubMed: 8402909]
Villunger et al, 2003. Villunger A. Michalak E.M. Coultas L. Müllauer F. Böck G. Ausserlechner M.J.Adams J.M. Strasser A. p53- and drug-induced apoptotic responses mediated by BH3-only proteinsPuma and Noxa. Science 2003;302:1036–1038. [PubMed: 14500851]
Wallach et al, 1998. Wallach D. Kovalenko A.V. Varfolomeev E.E. Boldin M.P. Death-inducingfunctions of ligands of the tumor necrosis factor family: a Sanhedrin verdict. Curr. Opin. Immunol.1998;10:279–288. [PubMed: 9638364]
Wang, 2001. Wang X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001;15:2922–2933. [PubMed: 11711427]
Watanabe-Fukunaga et al, 1992. Watanabe-Fukunaga R. Brannan C.I. Copeland N.G. Jenkins N.A.Nagata S. Lymphoproliferation disorder in mice explained by defects in Fas antigen that mediatesapoptosis. Nature 1992;356:314–317. [PubMed: 1372394]
Hughes et al. Page 12
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
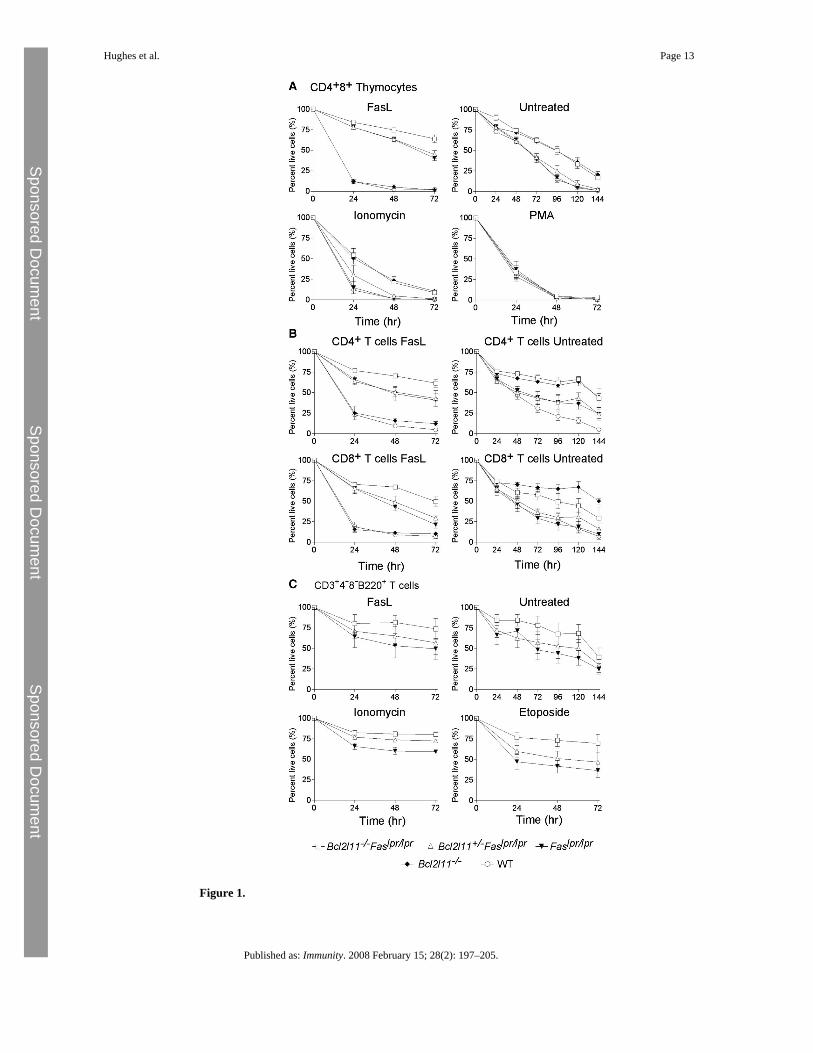
Figure 1.
Hughes et al. Page 13
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Leukocytes from Bcl2l11−/−Faslpr/lpr Mice Are Resistant to Both FasL as well as to Bim-Dependent Apoptotic Stimuli(A) CD4+8+ thymocytes from mice of the genotypes indicated were purified by FACS afterstaining with antibodies to CD4 and CD8. Cells were challenged in culture with the apoptoticstimuli indicated and survival assessed daily by propidium iodide staining and FACS analysis.(B and C) Similar experiments were performed with CD4+CD8− and CD4−CD8+ T cells (B)and αβTCR+CD4−CD8−B220+ T cells (C) purified from lymph nodes of mice of the genotypesindicated. Data represent the mean ± SEM of 3–7 mice for each genotype.
Hughes et al. Page 14
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
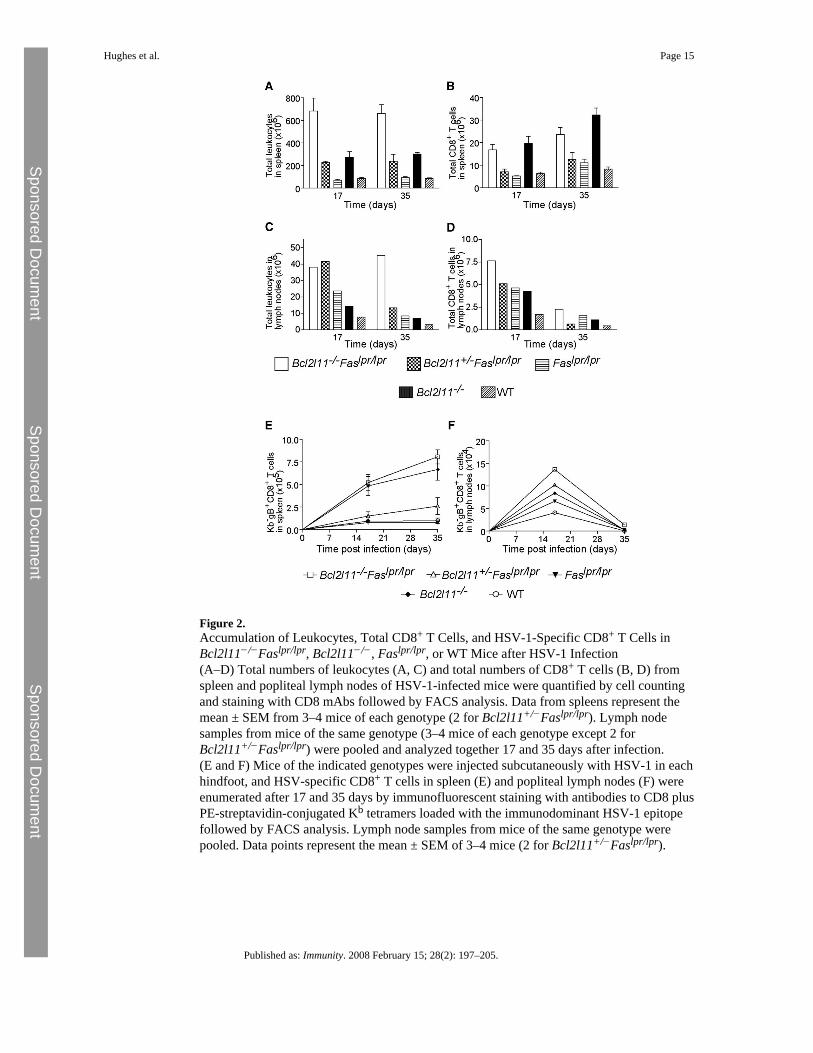
Figure 2.Accumulation of Leukocytes, Total CD8+ T Cells, and HSV-1-Specific CD8+ T Cells inBcl2l11−/−Faslpr/lpr, Bcl2l11−/−, Faslpr/lpr, or WT Mice after HSV-1 Infection(A–D) Total numbers of leukocytes (A, C) and total numbers of CD8+ T cells (B, D) fromspleen and popliteal lymph nodes of HSV-1-infected mice were quantified by cell countingand staining with CD8 mAbs followed by FACS analysis. Data from spleens represent themean ± SEM from 3–4 mice of each genotype (2 for Bcl2l11+/−Faslpr/lpr). Lymph nodesamples from mice of the same genotype (3–4 mice of each genotype except 2 forBcl2l11+/−Faslpr/lpr) were pooled and analyzed together 17 and 35 days after infection.(E and F) Mice of the indicated genotypes were injected subcutaneously with HSV-1 in eachhindfoot, and HSV-specific CD8+ T cells in spleen (E) and popliteal lymph nodes (F) wereenumerated after 17 and 35 days by immunofluorescent staining with antibodies to CD8 plusPE-streptavidin-conjugated Kb tetramers loaded with the immunodominant HSV-1 epitopefollowed by FACS analysis. Lymph node samples from mice of the same genotype werepooled. Data points represent the mean ± SEM of 3–4 mice (2 for Bcl2l11+/−Faslpr/lpr).
Hughes et al. Page 15
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Figure 3.After Intranasal MHV-OVA Infection, Bcl2l11−/−Faslpr/lpr Mice Accumulate More ViralAntigen-Specific CD8+ T Cells than Bcl2l11−/−, Faslpr/lpr, or WT MiceMice were infected intranasally with MHV-OVA and sacrificed after 14, 40, or 60 days, andmediastinal lymph nodes and spleen were analyzed.(A–D) Total leukocytes (A, B) and total numbers of CD8+ T cells (C, D) from spleen andmediastinal lymph nodes were quantified by cell counting and staining with CD8 mAbsfollowed by FACS analysis. Data represent the mean ± SEM from 3–4 mice of each genotype.Lack of data at day 14 for the Bcl2l11+/−Faslpr/lpr genotype is indicated with an asterisk.(E–H) MHV-OVA-specific CD8+ T cells were measured by immunofluorescent staining withPE-streptavidin-conjugated Kb/SIINFEKL tetramers plus antibodies to CD8 (E, F), or, afterbrief antigenic stimulation in vitro, by intracellular staining for IFN-γ (G, H), both followedby FACS analysis. Lymph node samples from mice of the same genotype were pooled. Datapoints represent the mean ± SEM of 3–4 mice (2 mice at some time points forBcl2l11+/−Faslpr/lpr and Faslpr/lpr mice).
Hughes et al. Page 16
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Figure 4.Loss of Bim and Fas Synergize in the Development of Lymphadenopathy and SplenomegalyMice (WT, Bcl2l11−/−, Faslpr/lpr, Bcl2l11+/−Faslpr/lpr, and Bcl2l11−/−Faslpr/lpr) wereanalyzed between the ages of 6 and 10 weeks. Spleen and lymph node cells were counted andanalyzed by immunofluorescent staining with surface marker-specific antibodies and FACSanalysis. Data represent mean ± SEM from 3–6 mice of each genotype.
Hughes et al. Page 17
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
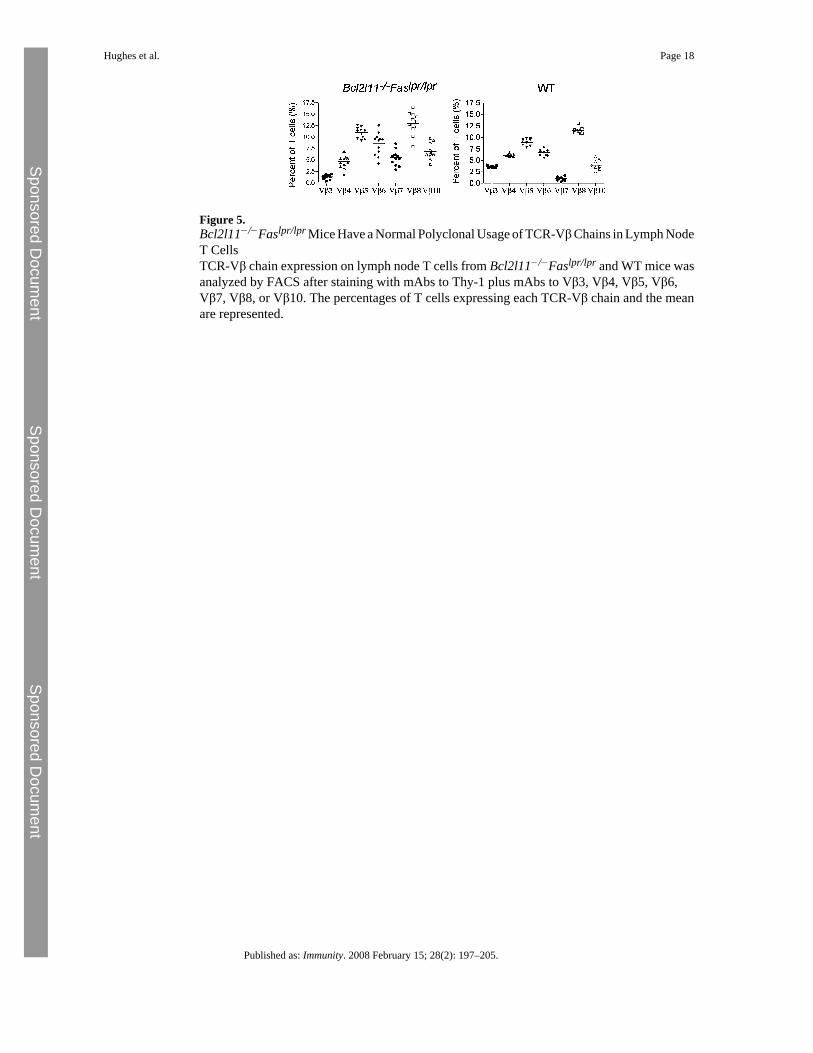
Figure 5.Bcl2l11−/−Faslpr/lpr Mice Have a Normal Polyclonal Usage of TCR-Vβ Chains in Lymph NodeT CellsTCR-Vβ chain expression on lymph node T cells from Bcl2l11−/−Faslpr/lpr and WT mice wasanalyzed by FACS after staining with mAbs to Thy-1 plus mAbs to Vβ3, Vβ4, Vβ5, Vβ6,Vβ7, Vβ8, or Vβ10. The percentages of T cells expressing each TCR-Vβ chain and the meanare represented.
Hughes et al. Page 18
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
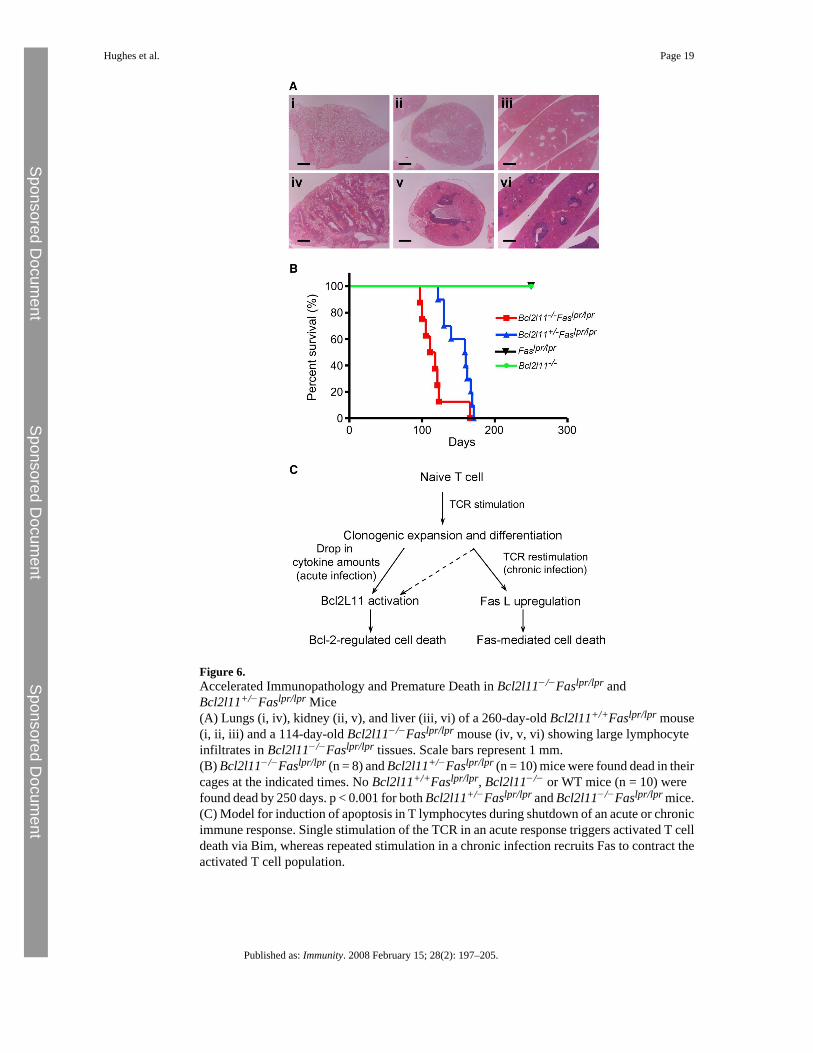
Figure 6.Accelerated Immunopathology and Premature Death in Bcl2l11−/−Faslpr/lpr andBcl2l11+/−Faslpr/lpr Mice(A) Lungs (i, iv), kidney (ii, v), and liver (iii, vi) of a 260-day-old Bcl2l11+/+Faslpr/lpr mouse(i, ii, iii) and a 114-day-old Bcl2l11−/−Faslpr/lpr mouse (iv, v, vi) showing large lymphocyteinfiltrates in Bcl2l11−/−Faslpr/lpr tissues. Scale bars represent 1 mm.(B) Bcl2l11−/−Faslpr/lpr (n = 8) and Bcl2l11+/−Faslpr/lpr (n = 10) mice were found dead in theircages at the indicated times. No Bcl2l11+/+Faslpr/lpr, Bcl2l11−/− or WT mice (n = 10) werefound dead by 250 days. p < 0.001 for both Bcl2l11+/−Faslpr/lpr and Bcl2l11−/−Faslpr/lpr mice.(C) Model for induction of apoptosis in T lymphocytes during shutdown of an acute or chronicimmune response. Single stimulation of the TCR in an acute response triggers activated T celldeath via Bim, whereas repeated stimulation in a chronic infection recruits Fas to contract theactivated T cell population.
Hughes et al. Page 19
Published as: Immunity. 2008 February 15; 28(2): 197–205.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
Copyright © 2022 FDOKUMEN




















