
Apo-Calmodulin Binds with its C-terminal Domain to the N-Methyl-D-aspartate Receptor NR1 C0 Region
11
Apo-Calmodulin Binds with its C-terminal Domain to the N-Methyl-D-aspartate Receptor NR1 C0 Region* Received for publication, March 12, 2003, and in revised form, September 30, 2003 Published, JBC Papers in Press, October 6, 2003, DOI 10.1074/jbc.M302542200 Zeynep Akyol‡, Jason A. Bartos§, Michelle A. Merrill§, Laurel A. Faga‡, Olav R. Jaren‡, Madeline A. Shea‡¶, and Johannes W. Hell§ From the Departments of ‡Biochemistry and §Pharmacology, Roy J. and Lucille A. Carver College of Medicine, University of Iowa, Iowa City, Iowa 52242-1109 Calmodulin (CaM) is the major Ca 2 sensor in eukary- otic cells. It consists of four EF-hand Ca 2 binding mo- tifs, two in its N-terminal domain and two in its C-ter- minal domain. Through a negative feedback loop, CaM inhibits Ca 2 influx through N-methyl-D-aspartate-type glutamate receptors in neurons by binding to the C0 region in the cytosolic tail of the NR1 subunit. Ca 2 - depleted (apo)CaM is pre-associated with a variety of ion channels for fast and effective regulation of channel activities upon Ca 2 influx. Using the NR1 C0 region for fluorescence and circular dichroism spectroscopy stud- ies we found that not only Ca 2 -saturated CaM but also apoCaM bound to NR1 C0. In vitro interaction assays showed that apoCaM also binds specifically to full- length NR1 solubilized from rat brain and to the com- plete C terminus of the NR1 splice form that contains the C0 plus C2 domain. The Ca 2 -independent interac- tion of CaM was also observed with the isolated C- but not N-terminal fragment of calmodulin in the independ- ent spectroscopic assays. Fluorescence polarization studies indicated that apoCaM associated via its C-ter- minal domain with NR1 C0 in an extended conformation and collapsed to adopt a more compact conformation of faster rotational mobility in its complex with NR1 C0 upon addition of Ca 2 . Our results indicate that apoCaM is associated with NR1 and that the complex of CaM bound to NR1 C0 undergoes a dramatic conformational change when Ca 2 binds to CaM. Glutamate is the prevailing excitatory neurotransmitter in the mammalian brain. The ionotropic N-methyl-D-aspartate (NMDA) 1 -type glutamate receptor is a major source of Ca 2 influx into neurons in the central nervous system. NMDA re- ceptors in the cerebral cortex largely consist of two NR1 and two or three NR2A and 2B subunits, which are homologous to each other (1– 4). Each subunit has an extracellular N termi- nus, four membrane regions (M1– 4), and an intracellular C terminus. M1, M3, and M4 are transmembrane regions but M2 loops into and back out of the plasma membrane at its cyto- plasmic face. The C-terminal domain of NR1 contains the 30 residue-long membrane-proximal C0 region common to all NR1 isoforms, followed in some NR1 isoforms by the C1 region (37 residues), which can be spliced in or out, and by either the C2 (38 residues) or the C2 (22 residues) region, which is down- stream of C2 in the genome. The C2 region can be spliced out but, when present, C2 will not be translated because C2 car- ries its own stop codon. In a negative feedback loop calmodulin (CaM) inhibits Ca 2 influx through the NMDA receptor by strongly binding to the NR1 C0 region in the presence of Ca 2 (5, 6). The same region interacts with -actinin, which can be displaced by Ca 4 2 -CaM (7, 8). Displacement of -actinin appears to reduce NMDA receptor activity by increasing desensitization, i.e. closing of the receptor pore while the agonist is still present (9). The Ca 4 2 -CaM-mediated negative feedback mechanism may help to reduce the otherwise uncontrolled Ca 2 influx through the NMDA receptor during ischemia as occurs during stroke and other pathological conditions (10, 11). Ischemia triggers a mas- sive release of glutamate from neurons, which results in over- activation of NMDA receptors. The ensuing unchecked Ca 2 influx ultimately leads to neuronal dysfunction and death. CaM constitutes the major Ca 2 sensor in eukaryotes and is ubiquitously expressed. It spans 148 residues and consists of an N- and a C-terminal domain, each containing two EF-hand Ca 2 binding sites (I–IV). It is highly conserved with only 3 residues differing between Drosophila and man. Ca 2 binds cooperatively, with sites III and IV in the C-domain usually having a 10-fold higher affinity than sites I and II in the N-domain. Ca 2 binding induces rearrangement of the tertiary structure of each domain, stabilizing the exposure of hydropho- bic clefts. These surfaces promote the association of CaM with a wide array of target proteins including kinases, phosphata- ses, ion channels, esterases, and metabolic enzymes (12–14). Early genetic studies of avoidance reactions of Paramecium led to the realization that (a) CaM was a subunit of two classes of ion channels in those organisms and (b) the two domains of CaM had separable roles in regulating ion channels (15, 16). Out of more than a dozen mutants, those that disrupted Ca 2 - dependent K -channel regulation in Paramecium and caused an exaggerated response to a chemical stimulus all mapped to the C-domain: either within sites III and IV (e.g. S101F, H135R) or to the terminal helix in that domain (e.g. M145V). In contrast, regulatory mutations that hindered Ca 2 -dependent Na -channel regulation, causing an abbreviated response to negative stimuli, mapped to positions in the N-domain of CaM, primarily between sites I and II (e.g. G40E, E54K) (15, 16). The effects of the mutations on Ca 2 binding were different: muta- tions in sites III and IV diminished Ca 2 binding while all * These studies were supported by Grants R01 GM57001 and R01 NS35563 from the National Institutes of Health (to M. A. S. and J. W. H.) and by the American Heart Association Established Investi- gator Award 0040151N (to J. W. H.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. ¶ To whom correspondence should be addressed. E-mail: madeline- [email protected]. To whom correspondence should be addressed. E-mail: johannes- [email protected]. 1 The abbreviations used are: NMDA, N-methyl-D-aspartate; CaM, calmodulin; CaMKII, calcium-calmodulin-dependent kinase II; CD, cir- cular dichroism; NR1C0p, NMDA receptor NR1 subunit C-terminal region covering residues 838 – 865; NTA, nitrilotriacetic acid; GST, glu- tathione S-transferase. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 3, Issue of January 16, pp. 2166 –2175, 2004 © 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 2166 by guest on August 10, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Apo-Calmodulin Binds with its C-terminal Domain to the N-Methyl-D-aspartate Receptor NR1 C0 Region

Apo-Calmodulin Binds with its C-terminal Domain to theN-Methyl-D-aspartate Receptor NR1 C0 Region*
Received for publication, March 12, 2003, and in revised form, September 30, 2003Published, JBC Papers in Press, October 6, 2003, DOI 10.1074/jbc.M302542200
Zeynep Akyol‡, Jason A. Bartos§, Michelle A. Merrill§, Laurel A. Faga‡, Olav R. Jaren‡,Madeline A. Shea‡¶, and Johannes W. Hell§�
From the Departments of ‡Biochemistry and §Pharmacology, Roy J. and Lucille A. Carver College of Medicine,University of Iowa, Iowa City, Iowa 52242-1109
Calmodulin (CaM) is the major Ca2� sensor in eukary-otic cells. It consists of four EF-hand Ca2� binding mo-tifs, two in its N-terminal domain and two in its C-ter-minal domain. Through a negative feedback loop, CaMinhibits Ca2� influx through N-methyl-D-aspartate-typeglutamate receptors in neurons by binding to the C0region in the cytosolic tail of the NR1 subunit. Ca2�-depleted (apo)CaM is pre-associated with a variety ofion channels for fast and effective regulation of channelactivities upon Ca2� influx. Using the NR1 C0 region forfluorescence and circular dichroism spectroscopy stud-ies we found that not only Ca2�-saturated CaM but alsoapoCaM bound to NR1 C0. In vitro interaction assaysshowed that apoCaM also binds specifically to full-length NR1 solubilized from rat brain and to the com-plete C terminus of the NR1 splice form that containsthe C0 plus C2� domain. The Ca2�-independent interac-tion of CaM was also observed with the isolated C- butnot N-terminal fragment of calmodulin in the independ-ent spectroscopic assays. Fluorescence polarizationstudies indicated that apoCaM associated via its C-ter-minal domain with NR1 C0 in an extended conformationand collapsed to adopt a more compact conformation offaster rotational mobility in its complex with NR1 C0upon addition of Ca2�. Our results indicate that apoCaMis associated with NR1 and that the complex of CaMbound to NR1 C0 undergoes a dramatic conformationalchange when Ca2� binds to CaM.
Glutamate is the prevailing excitatory neurotransmitter inthe mammalian brain. The ionotropic N-methyl-D-aspartate(NMDA)1-type glutamate receptor is a major source of Ca2�
influx into neurons in the central nervous system. NMDA re-ceptors in the cerebral cortex largely consist of two NR1 andtwo or three NR2A and 2B subunits, which are homologous toeach other (1–4). Each subunit has an extracellular N termi-nus, four membrane regions (M1–4), and an intracellular C
terminus. M1, M3, and M4 are transmembrane regions but M2loops into and back out of the plasma membrane at its cyto-plasmic face. The C-terminal domain of NR1 contains the 30residue-long membrane-proximal C0 region common to all NR1isoforms, followed in some NR1 isoforms by the C1 region (37residues), which can be spliced in or out, and by either the C2(38 residues) or the C2� (22 residues) region, which is down-stream of C2 in the genome. The C2 region can be spliced outbut, when present, C2� will not be translated because C2 car-ries its own stop codon.
In a negative feedback loop calmodulin (CaM) inhibits Ca2�
influx through the NMDA receptor by strongly binding to theNR1 C0 region in the presence of Ca2� (5, 6). The same regioninteracts with �-actinin, which can be displaced by Ca4
2�-CaM(7, 8). Displacement of �-actinin appears to reduce NMDAreceptor activity by increasing desensitization, i.e. closingof the receptor pore while the agonist is still present (9). TheCa4
2�-CaM-mediated negative feedback mechanism may helpto reduce the otherwise uncontrolled Ca2� influx through theNMDA receptor during ischemia as occurs during stroke andother pathological conditions (10, 11). Ischemia triggers a mas-sive release of glutamate from neurons, which results in over-activation of NMDA receptors. The ensuing unchecked Ca2�
influx ultimately leads to neuronal dysfunction and death.CaM constitutes the major Ca2� sensor in eukaryotes and is
ubiquitously expressed. It spans 148 residues and consists ofan N- and a C-terminal domain, each containing two EF-handCa2� binding sites (I–IV). It is highly conserved with only 3residues differing between Drosophila and man. Ca2� bindscooperatively, with sites III and IV in the C-domain usuallyhaving a 10-fold higher affinity than sites I and II in theN-domain. Ca2� binding induces rearrangement of the tertiarystructure of each domain, stabilizing the exposure of hydropho-bic clefts. These surfaces promote the association of CaM witha wide array of target proteins including kinases, phosphata-ses, ion channels, esterases, and metabolic enzymes (12–14).
Early genetic studies of avoidance reactions of Parameciumled to the realization that (a) CaM was a subunit of two classesof ion channels in those organisms and (b) the two domains ofCaM had separable roles in regulating ion channels (15, 16).Out of more than a dozen mutants, those that disrupted Ca2�-dependent K�-channel regulation in Paramecium and causedan exaggerated response to a chemical stimulus all mapped tothe C-domain: either within sites III and IV (e.g. S101F,H135R) or to the terminal helix in that domain (e.g. M145V). Incontrast, regulatory mutations that hindered Ca2�-dependentNa�-channel regulation, causing an abbreviated response tonegative stimuli, mapped to positions in the N-domain of CaM,primarily between sites I and II (e.g. G40E, E54K) (15, 16). Theeffects of the mutations on Ca2� binding were different: muta-tions in sites III and IV diminished Ca2� binding while all
* These studies were supported by Grants R01 GM57001 and R01NS35563 from the National Institutes of Health (to M. A. S. andJ. W. H.) and by the American Heart Association Established Investi-gator Award 0040151N (to J. W. H.). The costs of publication of thisarticle were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
¶ To whom correspondence should be addressed. E-mail: [email protected].
� To whom correspondence should be addressed. E-mail: [email protected].
1 The abbreviations used are: NMDA, N-methyl-D-aspartate; CaM,calmodulin; CaMKII, calcium-calmodulin-dependent kinase II; CD, cir-cular dichroism; NR1C0p, NMDA receptor NR1 subunit C-terminalregion covering residues 838–865; NTA, nitrilotriacetic acid; GST, glu-tathione S-transferase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 3, Issue of January 16, pp. 2166–2175, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org2166
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

other mutations had little effect (17, 18). Because all of theN-domain mutants exhibited Ca2� binding affinities similar tothose of wild type, it appeared that the physiological defectscaused by these mutations were likely to be related to targetbinding properties (17, 18). Studies of these mutants binding toa helical peptide showed that all of the mutants could bindunder apo (Ca2�-depleted) conditions and also suggested thatCaM may engage in a stepwise Ca2�-dependent association ofits domains with a target (19). Such a mechanism contradictedthe long held view of CaM as an “all-or-none” regulator oftarget proteins, “on” in the presence of Ca2� and “off” in itsabsence. In fact, it is now known that CaM interactions varygreatly between its many targets (Fig. 1) (12, 14).
Pre-association of apoCaM with various ion channels thatare controlled by Ca4
2�-CaM is critical for a swift and potentresponse (e.g. (13, 20–26)). An initial study by Ehlers et al. (5)suggested that the interaction of CaM with NR1 C0 only occursin the presence of Ca2� (5) raising the possibility that theNMDA receptor may be an exception to this rule. Such anexception would be critical because it would question the gen-eral relevance of pre-association of CaM with various ionchannels for fast and effective signaling. Therefore and be-cause of the central role the NMDA receptor plays duringischemia-triggered neuronal damage and the antagonistic ef-fect of Ca4
2�-CaM on NMDA receptor activity we want to gain abetter understanding of the molecular interplay between CaMand NR1. We investigated whether Ca2�-depleted (apo)CaM ispre-associated with the NMDA receptor. In the earlier study byEhlers et al. (5) CaM binding under Ca2�-depleted conditionshad only been studied by pull-down and co-immunoprecipita-tion assays, during which protein-protein interactions with anoff-rate in the minute range may be lost. We employed severalspectroscopic methods to study CaM association with a peptidespanning most of the NR1 C0 region (NR1C0p; residues 838–
865) under Ca2�-depleted (apo) and Ca2�-saturating condi-tions. Fluorescence and circular dichroism (CD) spectroscopyprovided strong evidence for association of CaM with NR1C0pnot only in the presence of Ca2� but also under apo conditions.ApoCaM also bound specifically to the full-length NR1 subunitisolated from rat brain and to a GST fusion protein of the Cterminus of the C0-C2� splice form of NR1 when a fast washingprotocol was used. Experiments with isolated domain frag-ments of CaM further indicated that the C- but not the N-domain of CaM selectively interacts with NR1C0p under apoconditions. Fluorescence anisotropy of NR1C0p in the presenceof saturating concentrations of apoCaM is larger than in thepresence of Ca4
2�-CaM demonstrating that the rotational mo-bility of NR1C0p is lower when bound to apoCaM than toCa4
2�-CaM. These results suggest that apoCaM is pre-associ-ated with NR1 C0 via its C-domain and that Ca2� induces thebind of the N-domain to NR1 C0. The latter interaction resultsin a CaM-NR1C0p complex that is more compact than theapoCaM-containing complex.
EXPERIMENTAL PROCEDURES
Materials—Fluorescein-labeled NR1 C0 peptide (NR1C0p; Fluores-cein-SRHKDARRKQMQLAFAAVNVWRKNLQDR) was custom-syn-thesized by the W. M. Keck Biotechnology Resource Center, Yale Uni-versity (New Haven, CT), and purified by HPLC. NR1C0p correspondedto residues 838–865 of NR1, containing a single tryptophan residue atits position 21. A biotinylated form of a peptide with a nearly identicalsequence (biotin-NR1C0p: biotin-RHKDARRKQMQLAFAAVNVWRK-NLQ) was obtained from the same source. Recombinant rat calmodulin(CaM1–148) and N- and C-domain fragments (CaM1–80 and CaM76–148,respectively) were overexpressed in Escherichia coli using the pT7-7vector in Lys-S cells (U. S. Biochemicals, Cleveland, OH) (27). Purifica-tion was performed on phenyl-Sepharose CL-4B (Amersham Bio-sciences) (28). Recombinant proteins were characterized by reversed-phase high-performance liquid chromatography and by SDS-polyacrylamide gel electrophoresis followed by silver staining (29). CaMsamples were depleted of Ca2� by successive dialyzes at 4 °C against 50mM HEPES, 100 mM KCl, 1 mM MgCl2, 0.5 mM NTA, with 5 mM EGTAand then against the same buffer containing only 0.05 mM EGTA.
Fluorescence Emission Wavelength Scans—Fluorescence emissionstudies were performed with an SLM-4800 spectrofluorimeter (SLM,Urbana, IL) with a Xenon short arc lamp (Ushio, Inc.), using 8 nmbandpasses with NR1C0p (2.54 �M) and CaM (�2.3 �M) for Trp fluo-rescence, and a Fluorolog 3 spectrofluorimeter (Jobin Yvon, Horiba)with a 450 watt xenon lamp using 3 nm bandpasses with NR1C0p (0.22�M) and CaM (�2.5 �M) for fluorescein fluorescence. Emission spectraof NR1C0p were obtained at �ex of 297 nm for tryptophan and �ex of 496nm for fluorescein. The following buffer conditions were used: 50 mM
HEPES, 100 mM KCl, 1 mM MgCl2, 0.05 mM EGTA, 5 mM NTA, �10 mM
CaCl2, pH 7.4, 22 °C. Data were corrected for contributions from thebuffer but not corrected for instrument response. Each experiment wasrepeated at least three times with consistent results.
Circular Dichroism—CD spectra were collected with an Aviv 62DSCD instrument using 2-s increments, a 1.5-nm bandpass, and 0.5-nmwavelength spacing from 260 to 215 nm. CD spectra were recorded in aquartz cuvette (1-cm pathlength) with 6 �M recombinant CaM or itsfragments and 10.24 �M or 13.54 �M NR1C0p. The following bufferconditions were used: 2 mM HEPES, 100 mM KCl, 1 mM MgCl2, 0.05 mM
EGTA, 5 mM, �10 mM CaCl2, pH 7.4 22 °C. Each experiment wasrepeated three times under both conditions. In each individual experi-ment, readings from three successive scans were averaged, and datawere corrected for contribution from buffer. To observe changes in theellipticity of biotin-NR1C0p, CD spectra were collected using 0.5-nmstep size and 1.0-nm bandwidth in a 1-cm quartz cuvette at 22 °C.Ellipticity of 120 �M biotin-NR1C0p was monitored in ddH2O withincreasing fractional concentrations (v/v) of TFE (2,2,2-trifluoro-etha-nol; Sigma Chemicals).
Fluorescence Anisotropy—Fluorescence anisotropy was measured us-ing a Fluorolog 3 (Jobin Yvon, Horiba) spectrofluorimeter with an 8-nmbandpass. Fluorescein fluorescence was detected at �ex of 496 nmand �em of 520 nm. Anisotropy R was calculated with r � (IVV� G �IVH)/(IVV � 2 � GIVH) [IVV and IVH: intensity of vertically or horizontallyemitted light, respectively, when vertically excited; IHV and IHH: inten-sity of vertically or horizontally emitted light, respectively, when hori-zontally excited; G � IHV/IHH]. Three readings were taken for each
FIG. 1. Ribbon diagrams of apo- and Ca42�-CaM and CaM-target
complexes. The EF-hand Ca2� binding sites and Ca2� ions are alldepicted in yellow. In CaM, N-domain residues 1–75 are blue, linkerresidues 76–80 are black, and C-domain residues 81–148 are red.Target binding domains and proteins are colored green. Shown areapoCaM (1cfd.pdb; Ref. 45); Ca4
2�-CaM (1clm.pdb, Ref. 46); Ca42�-CaM
in a complex with the Anthrax edema factor (1k90.pdb, Ref. 47);Ca4
2�-CaM in complex with a peptide from a plasma membrane calciumATPase (1cff.pdb, Ref. 48); and Ca4
2�-CaM in complex with a peptidefrom skeletal muscle myosin light chain kinase (2bbm.pdb, Ref. 49).Ribbon drawings were made with Molscript (50).
Apo-Calmodulin Binds to the NMDA Receptor 2167
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

measurement and averages calculated and recorded. Each signal wasmonitored for 2 s. Buffer consisted of 50 mM HEPES, 100 mM KCl, 1 mM
MgCl2, 0.05 mM EGTA, 5 mM NTA, pH 7.4, 22 °C. Aliquots were addedfrom concentrated apoCaM samples (0.5–1.2 mM) to titrate NR1C0pwith a starting concentration of 0.22 �M; the total dilution of theNR1C0p solution in the cuvette was �3% over the course of the titra-tions. To determine the affinity of apoCaM1–148 and apoCaM76–148 forNR1C0p, the anisotropy data from the titration curves with increasingamounts of CaM shown in Fig. 6, A and C were fit to a simple bindingmodel, treating the peptide-CaM complex as having a 1:1 stoichiometry.Fractional saturation of the peptide by CaM was represented as Y �(Ka[X])/(1 � Ka[X]) (29), where Ka represents the intrinsic associationconstant (Ka � 1/Kd) for CaM binding to NR1C0p and [X] is the CaMconcentration. Titration curves were analyzed with NONLIN (30). Theabscissa in Fig. 6 is shown on a log scale. To indicate the value ofanisotropy in the absence of apoCaM (i.e. at 0 M), the abscissa value waschosen arbitrarily to be 5 � 10�8 M; fitting was done using the actualvalue.
For the second phase of these experiments (right panels in Fig. 6), thefinal solution of the titration of NR1C0p with apoCaM was titratedsubsequently with 10 mM CaCl2 in 50 mM HEPES, 100 mM KCl, 1 mM
MgCl2, 0.05 mM EGTA, 5 mM NTA, pH 7.4, 22 °C. The abscissa repre-sents the total concentration of Ca2� added; the concentration of freeCa2� would be lower because of competition between CaM and theEGTA used to deplete the sample of calcium initially. Each experimentwas repeated at least three times with consistent results.
Pull-down of CaM with Purified NR1 and GST-NR1C0-C2�—NR1and the AMPA-type glutamate receptor subunit GluR1 were solubilizedfrom crude rat forebrain membrane fractions and immunoisolated asdescribed (31, 32). Briefly, membrane fractions were treated with 1%SDS at 60 °C for 30 min. This procedure efficiently extracts GluR1 andNR1 from the membranes and dissociates NR1 from NR2 subunits andGluR1 from the other AMPA receptor subunits (i.e. GluR2 and GluR3;Refs. 31–33). After addition of an 8-fold excess of a 1% Triton X-100solution, immunoprecipitation with anti-NR1 and anti-GluR1 antibod-ies (31–33) and washing, immobilized glutamate receptor subunits wereincubated for 2 h at 4 °C with 50 �M purified CaM �10 mM CaCl2 in
HEPES buffer (50 mM HEPES, 100 mM KCl, 1 mM MgCl2, 5 mM NTA,0.05 mM EGTA). Samples were washed twice as fast as possible, usuallyfor a total of 1.5–2 min, with HEPES buffer containing 0.1% TritonX-100 and 10 mM CaCl2 if present during the incubation with CaM. Theproteins were extracted using standard SDS sample buffer (31–33)supplemented with 50 mM EGTA to effectively remove Ca2� form CaMin samples incubated in the presence of CaCl2; otherwise, apoCaM andCa2�-CaM would migrate with different speed. After SDS-PAGE, pro-teins were transferred onto polyvinylidene difluoride membranes.Membranes were cut into half, the upper portion probed with anti-NR1,stripped, and re-probed with anti-GluR1 (31–33) and the lower partprobed with a mouse monoclonal anti-CaM antibody (Zymed Laborato-ries Inc.). Immunosignals were detected with ECL plus (AmershamBiosciences) (31–33).
GST and a GST fusion protein containing the C-terminal C0 and C2�regions of NR1 were expressed in E. coli, extracted and immobilized onglutathione-Sepharose as described (8, 32, 33). The samples werewashed three times with HEPES buffer, incubated with 0.1–50 �M
purified CaM (Sigma) in HEPES buffer � 10 mM CaCl2 for 2 h at 4 °C,and washed two times as quickly as possible (1.5–2 min total time) withHEPES buffer containing 0.1% Triton X-100 and also CaCl2 if the latterwas present during the incubation with CaM. SDS-PAGE and immu-noblotting was performed as described above. Quantification of theCaM immunosignals was conducted by determining the mean densityas measured by Adobe Photoshop after digitizing exposed films with aUMAX UTA1100 scanner.
RESULTS
CaM regulates the activity of a variety of ion channels in-cluding voltage-gated K� channels (20), Ca2� channels (21–23,25), and glutamate-gated NMDA receptors (5). In the presenceof Ca2�, CaM inhibits NMDA receptor activity via its interac-tion with the C0 region in the cytosolic C terminus of the NR1subunit (5, 6). The previous biochemical characterization of theCaM-NMDA receptor interaction suggested that CaM binding
FIG. 2. Tryptophan fluorescence emission spectra of NR1C0p. All samples contained 2.5 �M NR1C0p, 50 mM HEPES, 100 mM KCl, 1 mM
MgCl2, 0.05 mM EGTA, 5 mM NTA, �10 mM CaCl2, pH 7.4, at 22 °C. Emission spectra were recorded at �ex of 297 nm before (black traces) or afteraddition of 2.5 �M CaM1–148 (A, D; green traces), CaM1–80 (B, E; blue traces), or CaM76–148 (C, F; red traces) in Ca2�-depleted (A–C), andCa2�-saturated (D–F) buffer. Emission spectra were normalized by setting the maximum of each spectrum (at �350 nm) measured before additionof CaM equal to 1. The black arrow reflects the increase in emission intensity at 330 nm upon addition of the indicated CaM polypeptides.Schematic diagrams, CaM residues 1–75 (blue circle), CaM residues 81–148 (red circle), CaM linker (�; residues 76–80, present in all CaMconstructs). Calcium binding sites (small circles): apo (open) or saturated (closed). Similar results were obtained in three independent experimentswith fluorescein-labeled NR1C0p and in two experiments with the biotinylated NR1C0p.
Apo-Calmodulin Binds to the NMDA Receptor2168
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from
to the NR1 C0 region strictly requires Ca2� (5). This conclusionrelied on pull-down experiments, in which an interaction that isspecific but has a fast off-rate would be lost. Because apoCaM isassociated with other ion channels for fast and efficient signaling,we investigated with complementary spectroscopic methods andpull-down assays whether CaM may be able to interact in asimilar Ca2�-independent manner with NR1 C0.
Tryptophan Fluorescence of NR1C0p—Because CaM doesnot have a tryptophan, the fluorescent properties of tryptophanat position 21 (Trp-21) of the NR1C0p peptides can be utilizedto detect changes in its immediate environment that occurwhen the peptide binds to CaM. The NR1C0p tryptophan waseffectively excited at 297 nm; at this wavelength contributionby tyrosine or phenylalanine to the fluorescence signal is min-imal. The resulting emission spectrum of the NR1C0p had a�max of 350 nm in the absence of CaM (Fig. 2 A–F, black traces).Addition of full-length CaM1–148 to NR1C0p changed the emis-sion spectrum of Trp-21 of NR1C0p not only under Ca2�-satu-rating (Fig. 2A) but also under Ca2�-depleted conditions (Fig.2D). There was a blue-shift (to lower wavelength) in �max of theemission spectrum by slightly less than 20 nm under Ca2�-depleted conditions and slightly more than 20 nm under Ca2�-saturating conditions. At the same time the maximal intensityof the emission signals increased by about 2- and 4-fold,respectively. These findings suggest that apoCaM as well asCa4
2�-CaM can interact with NR1C0p.To investigate whether the N- or C-domain of CaM is ne-
cessary or sufficient for interaction with NR1C0p under apo orCa2�-saturated conditions, we used recombinant puri-fied CaM1–80 (N-domain followed by the linker region) andCaM76–148 (C-domain preceded by the linker region) for similarmeasurements. Both fragments bound to NR1C0p in the pres-ence of Ca2� (Fig. 2, E and F). A shift of slightly less than �20nm in �max was observed upon addition of the N- as well as the
C-domain. CaM76–148, but not CaM1–80, also interacted withthe peptide under Ca2�-depleted conditions (Fig. 2, B and C).Binding of the apo C-domain caused a shift of �max by �10 nmto 340 nm and a modest increase in the maximal intensity ofthe emission signal (Fig. 2C). Similar results were obtainedstudying the same forms of CaM binding to biotin-NR1C0p(data not shown).
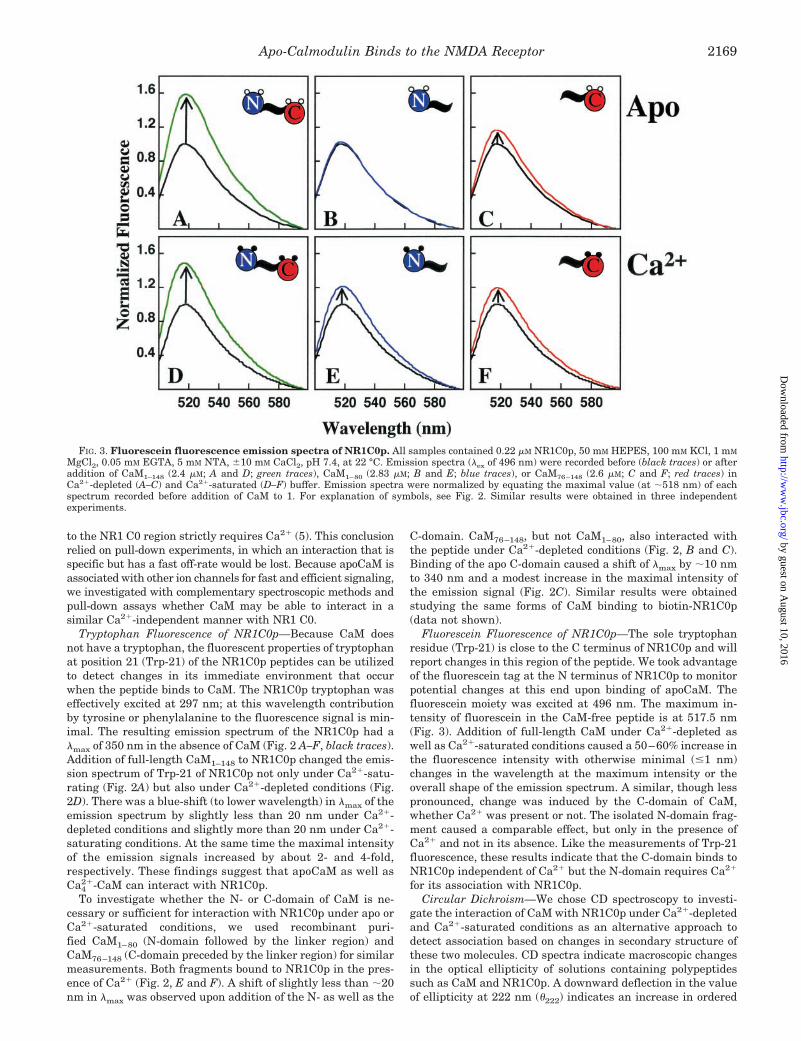
Fluorescein Fluorescence of NR1C0p—The sole tryptophanresidue (Trp-21) is close to the C terminus of NR1C0p and willreport changes in this region of the peptide. We took advantageof the fluorescein tag at the N terminus of NR1C0p to monitorpotential changes at this end upon binding of apoCaM. Thefluorescein moiety was excited at 496 nm. The maximum in-tensity of fluorescein in the CaM-free peptide is at 517.5 nm(Fig. 3). Addition of full-length CaM under Ca2�-depleted aswell as Ca2�-saturated conditions caused a 50–60% increase inthe fluorescence intensity with otherwise minimal (�1 nm)changes in the wavelength at the maximum intensity or theoverall shape of the emission spectrum. A similar, though lesspronounced, change was induced by the C-domain of CaM,whether Ca2� was present or not. The isolated N-domain frag-ment caused a comparable effect, but only in the presence ofCa2� and not in its absence. Like the measurements of Trp-21fluorescence, these results indicate that the C-domain binds toNR1C0p independent of Ca2� but the N-domain requires Ca2�
for its association with NR1C0p.Circular Dichroism—We chose CD spectroscopy to investi-
gate the interaction of CaM with NR1C0p under Ca2�-depletedand Ca2�-saturated conditions as an alternative approach todetect association based on changes in secondary structure ofthese two molecules. CD spectra indicate macroscopic changesin the optical ellipticity of solutions containing polypeptidessuch as CaM and NR1C0p. A downward deflection in the valueof ellipticity at 222 nm (�222) indicates an increase in ordered
FIG. 3. Fluorescein fluorescence emission spectra of NR1C0p. All samples contained 0.22 �M NR1C0p, 50 mM HEPES, 100 mM KCl, 1 mM
MgCl2, 0.05 mM EGTA, 5 mM NTA, �10 mM CaCl2, pH 7.4, at 22 °C. Emission spectra (�ex of 496 nm) were recorded before (black traces) or afteraddition of CaM1–148 (2.4 �M; A and D; green traces), CaM1–80 (2.83 �M; B and E; blue traces), or CaM76–148 (2.6 �M; C and F; red traces) inCa2�-depleted (A–C) and Ca2�-saturated (D–F) buffer. Emission spectra were normalized by equating the maximal value (at �518 nm) of eachspectrum recorded before addition of CaM to 1. For explanation of symbols, see Fig. 2. Similar results were obtained in three independentexperiments.
Apo-Calmodulin Binds to the NMDA Receptor 2169
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

secondary structure, often interpreted as an increase in �-hel-ical content of polypeptides in the solution. Under both Ca2�-depleted and Ca2�-saturating conditions, NR1C0p alone hadan ellipticity reading close to zero, reflecting that NR1C0p hadno detectable persistent secondary structure by itself (Fig. 4,black traces). In contrast, CaM possesses a high degree ofellipticity under both conditions reflecting the primarily �-hel-ical structure of CaM (see Fig. 1). As expected for CaM (34),Ca2� binding induces an increase in ordered secondary struc-ture in CaM (compare Fig. 4, A and B, green solid tracesreflecting spectra of CaM before addition of NR1C0p). Underapo conditions, the addition of NR1C0p (final concentration of10.24 �M) to 6 �M CaM1–148 lowered �222 by 6.54 � 2.4% (Fig.4A, compare the solid with the dashed green trace). In thepresence of Ca2�, the decrease in �222 was greater than 10.69 �2.40% (Fig. 4B, green). The change in ellipticity upon NR1C0paddition reflects an increase in ordered secondary structure ofthe solutes. This change cannot be assigned to individual com-ponents (i.e. NR1C0p may adopt an �-helical conformationupon binding to CaM and/or the secondary structure of CaMmay change upon peptide binding). However, the decrease in�222 confirms a direct interaction between NR1C0p and CaMunder apo conditions. Comparable results were obtained whenthe biotin-NR1C0p instead of fluorescein-NR1C0p was used tostudy the interaction of CaM with NR1C0p by CD spectroscopy(data not shown).
We performed similar measurements with the isolated N-and C-domain fragments of CaM: CaM1–80 and CaM76–148.Results were consistent with those obtained using fluorescence
to monitor the environment of Trp-21 of NR1C0p or its fluores-cein label. In the absence of Ca2�, the CD spectrum of theN-domain (CaM1–80) did not show detectable and consistentchanges upon addition of NR1C0p (signal differences of 1.49 �1.65%; Fig. 4A, blue). However, the value of �222 decreased by21.99 � 4.42% when NR1C0p was added to CaM1–80 underCa2�-saturating conditions (Fig. 4B). In contrast to the resultsfor CaM1–80, the CD spectra obtained for CaM76–148 exhibiteda clear and reproducible decrease in ellipticity upon addition ofNR1C0p both in the presence and the absence of Ca2� (Fig. 4,red). NR1C0p induced a decrease in �222 of 12.66 � 2.73% whenadded to apoCaM76–148 and a deflection of 45.81 � 4.22% in thepresence of Ca2�-saturated CaM76–148. To corroborate thesefindings, we repeated this set of experiments in triplicate witha higher NR1C0p concentration. When NR1C0p (final concen-tration of 13.54 �M) was added to 6 �M CaM under apo condi-tions, the signal for CaM1–148 dropped by 8.57 � 2.1%, thesignal for CaM1–80 dropped 0.12 � 0.66% (negligible change)and the signal for CaM CaM76–148 dropped by 16.59 � 2.29%(Fig. 4C). In the presence of saturating levels of Ca2�, thecorresponding decreases in �222 were 11.96 � 3.53% for CaM1–148,27.82 � 4.55% for CaM1–80 and 50.53 � 1.95% for CaM76–148 (Fig.4D). These results confirmed that the C- but not the N-domain ofCaM associated with NR1C0p under apo conditions, while bothbound under calcium-saturating conditions. Similar results wereobtained with the biotinylated version of NR1C0p (data not shown).
The CD spectra of biotin-NR1C0p alone and in the presenceof increasing fractions of TFE, a helicogenic solvent, showedthat the peptide underwent a progressive conformationalchange that was consistent with it adopting an �-helical struc-ture (e.g. minima at 208 and 222 nm; Fig. 5). This finding isconsistent with the change in ellipticity seen when NR1C0pwas added to CaM and could be promoted by its conformationaltransition upon associating with CaM.
Fluorescence Anisotropy of NR1C0p—We looked for changesin fluorescence anisotropy of NR1C0p by monitoring its fluo-rescein signal as a third approach to investigate interactions ofCaM with NR1 C0 under apo conditions and to test whetheraddition of Ca2� may change the conformation of the hypoth-esized complex of NR1C0p with apoCaM. If polarized light isused to excite NR1C0p in solution, most of the polarization islost due to the fast tumbling rate of monomeric NR1C0p. Uponbinding of CaM, the rotational mobility of NR1C0p will bereduced, thereby increasing the degree of polarization of the
FIG. 4. Circular dichroism of CaM upon addition of NR1C0p.Ellipticity values were determined for 6 �M CaM1–148 (green), CaM76–148(red), and CaM1–80 (blue) under Ca2�-depleted (A and C) and Ca2�-saturated (B and D) conditions in 2 mM HEPES, 100 mM KCl, 1 mM
MgCl2, 0.05 mM EGTA, 5 mM NTA, � 10 mM CaCl2, pH 7.4, at 22 °C, inthe absence (solid traces) and presence (dashed traces) of 10.24 �M
NR1C0p as a function of wavelength (A, B). Buffer (orange traces) andNR1C0p alone (black traces) did not exhibit any detectable ellipticity.Average of three independent experiments were taken to determine thechanges in the % ellipticity upon addition of 13.54 �M NR1C0p to 6 �M
CaM1–148 (green), CaM76–148 (red), and CaM1–80 (blue) under the samebuffer conditions in the absence (C) and presence (D) of Ca2�. Similarresults were obtained in three independent experiments for both 10.24and 13.54 �M fluorescein-labeled NR1C0p and in two experiments withthe biotinylated version of NR1C0p. For explanation of symbols seeFig. 2.
FIG. 5. Secondary structure of biotin-NR1C0p. Circular dichro-ism spectra showing that the ellipticity at 222 nm of biotin-NR1C0p(120 �M in ddH2O; black, thin solid curve) decreased upon addition ofthe helicogenic solvent TFE to a final concentration of 25% (v/v) (green,dotted), 50% (v/v) (red, dashed) and 91.6% (v/v) (purple, thick solid).
Apo-Calmodulin Binds to the NMDA Receptor2170
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

emitted light. NR1C0p alone has an anisotropy value of �0.06.Addition of increasing amounts of CaM1–148 to NR1C0p re-sulted in a saturation curve with a maximal increase of 69% inpeptide anisotropy under apo conditions (Fig. 6A, left). Whenthe solution of NR1C0p saturated with apoCaM was then ti-trated with Ca2�, a final decrease of �22% in anisotropy wasobserved (Fig. 6A, right). An initial plateau in anisotropy isobserved because EGTA in the bulk solution competes withCaM for added Ca2�.
Titration of NR1C0p with apoCaM76–148 also showed a sat-uration curve with a maximal increase in anisotropy of 46.5%(Fig. 6C, left). In contrast to the experiments with full-lengthCaM, addition of Ca2� to the solution containing apoCaM76–148
and NR1C0p did not cause any further changes in anisotropy(Fig. 6C, right). Titration of NR1C0p with apoCaM1–80 did notalter its anisotropy (Fig. 6B) but titration of the mixture ofCaM1–80 and NR1C0p with Ca2� elevated the anisotropy by upto 21.4% in a saturable fashion. Nonlinear least squares anal-
ysis of the titrations of NR1C0p with apoCaM1–148 (one of threetrials is shown in Fig. 6) yielded an apparent Kd of 2.25 � 0.08�M while the Kd for apoCaM76–148 binding to NR1C0p was1.21 � 0.10 �M. The difference between the two dissociationconstants is much larger than the error for each.
Binding of ApoCaM to Full-length NR1—To test whetherapoCaM can interact with full-length NR1 expressed in brain,NR1 was solubilized from a crude rat brain membrane fractionwith 1% SDS at 60 °C to dissociate NR1 from NR2 subunits.After immunoisolation, the resin-bound NR1 was incubatedwith CaM with and without Ca2� added. After two subsequentquick washing steps CaM was detectable in the pellet not onlyunder Ca2�-saturated but also Ca2�-depleted conditions (Fig.7). Neither Ca2�-CaM nor apoCaM bound to the AMPA recep-tor subunit GluR1, which was extracted and immunoisolatedfrom rat brain in parallel with NR1.
Binding of ApoCaM to the C Terminus of NR1—To excludethat regions outside of the C terminus of NR1 or the C-terminal
FIG. 6. Fluorescence anisotropy of NR1C0p and model of its interactions with the N- and the C-domain of CaM under Ca2�-depleted and Ca2�-saturated conditions. Fluorescence anisotropy of the fluorescein-labeled NR1C0p was monitored (�ex of 496 nm, �em of 520nm) as a function of increasing concentrations of apoCaM1–148 (A left; green), CaM1–80 (B left; blue), or CaM76–148 (C left; red) in 50 mM HEPES,100 mM KCl, 1 mM MgCl2, 0.05 mM EGTA, 5 mM NTA, pH 7.4, at 22 °C. Nonlinear analysis of the titration curves of NR1C0p with apoCaM1–148and apoCaM76–148 indicated apparent Kd values of 2.25 and 1.21 �M, respectively (respective EC50 values were 1.99 and 1.05 �M). At a finalconcentration of �20 �M for each CaM polypeptide (at this concentration binding of apoCaM1–148 and apoCaM76–148 is in the saturating range; notethe log scale of the abscissa), the samples were titrated with Ca2� (right side). The total CaCl2 concentration is indicated. Similar results wereobtained in 3 independent experiments. For explanation of schematic diagrams of CaM, see Fig. 2. The gray bar represents NR1C0p; an overlapindicates the proposed direct interactions with N- and C-domains under specific conditions. Accordingly, under Ca2�-depleted conditions theC-domain is associated with NR1C0 and CaM is mainly in an extended conformation. The progressive reduction in the fluorescent anisotropy signalupon addition of increasing amounts of Ca2� to the apoCaM1–148-NR1C0p complex likely reflects the formation of a second, Ca2�-dependentinteraction between these two polypeptides. This Ca2�-dependent interaction is likely mediated by the N-domain, and causes a collapse of theextended apoCaM1–148 conformation to adopt a more compact structure in its complex with NR1 C0, thereby reducing the hydrodynamic radius andincreasing the rotational mobility.
Apo-Calmodulin Binds to the NMDA Receptor 2171
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

C1 region of NR1 rather than the C0 region bind apoCaM infull-length NR1, we expressed the shortest naturally occurringsplice form of the C terminus of NR1 as GST fusion protein,
which consists of C0 and C2�. Specific binding of Ca2�-CaM toGST-NR1C0-C2� was detectable at all concentrations of CaMtested and saturated around 2 �M (Fig. 8). At the higher con-centration range specific binding by apoCaM was also ob-served. Immunosignals were quantified after digitalization andnonspecific binding, which was determined with GST alone(Fig. 8A, lower panels) and subtracted.
DISCUSSION
An elegant study by Ehlers et al. (5) provided early evidencethat CaM interacts with NR1 C0 in the presence of Ca2�. TheKd value of the NR1 C0 region for full-length CaM fluorescentlylabeled with a dansyl moiety, a well established tool for report-ing CaM interactions with various targets, is �87 nM underCa2�-saturated conditions (5). Using unmodified wild typeCaM1–148 for titrating the fluorescein-tagged NR1C0p in thepresence of Ca2� we determined a comparable Kd value of �80nM by fluorescence polarization (data not shown). The similar-ity in these two Kd values suggests that labeling CaM with adansyl group or alternatively our NR1C0-derived polypeptidewith a fluorescein group has little if any effect on their bindingconstants and properties. Confidence that the NR1C0p reflectsproperties of the C0 region as present in full-length NR1 isfurther increased as two other proteins that interact with theC0 domain, �-actinin, and Ca2�- and CaM-dependent kinaseCaMKII (7, 8, 32), bound to NR1C0p in a saturable mannerwith Kd values in the range of �500 nM and 1–2 �M, respec-tively (data not shown). The Kd value of CaMKII matches thecorresponding Kd value for the interaction of CaMKII withfull-length NR1 from rat brain (32).
Titration of the fluorescence anisotropy of the fluorescein-labeled NR1C0p with apoCaM1–148 revealed a much weakerinteraction (Kd of 2.25 �M; see Fig. 6A and “Results”) than withCa2�-CaM. This more than 25-fold difference in affinity uponremoval of Ca2� provides an explanation as to why apoCaMbinding to the NR1 C-terminal region might not be readilydetectable in pull-down and co-immunoprecipitation experi-ments and was missed in the earlier study by Ehlers et al. (5).Such methods usually involve washing procedures that take atleast several minutes and that time period would allow disso-ciation of apoCaM from the fusion proteins. However, a Kd
value in the low micromolar range can be quite sufficient forsubstantial prelocalization and enrichment of CaM at theNMDA receptor complex under basal Ca2� concentrations. Be-cause a similar pre-association of CaM with other ion channelsis critical for their effective regulation by Ca2� (13, 20–23, 25,26), we scrutinized and further characterized binding ofapoCaM to NR1 C0 with several spectroscopic methods. Wedetected significant changes in four different spectroscopicproperties upon mixing NR1C0p with CaM under Ca2�-de-pleted conditions (Figs. 2–4 and 6), each of which stronglyargues that apoCaM interacts with NR1C0p.
Our first approach took advantage of the single tryptophan atposition 21 in NR1C0p (Trp-21; Fig. 2). Because CaM does notcontain any tryptophan, changes in the spectroscopic and therebyenvironmental properties of Trp-21 can be easily monitored. Weobserved a strong shift in the fluorescence emission maximum(�max) of Trp-21 to lower wavelengths and a concomitant strongincrease in the intensity of this maximum upon addition ofapoCaM1–148. These effects indicate a change in the Trp-21 en-vironment, which could be due to either a direct interactionbetween CaM and Trp-21 or an indirect effect of CaM binding toNR1C0p as this binding likely caused a change in the overallNR1C0p structure (see below). Similar though smaller effectswere obtained with apoCaM76–148 but not with apoCaM1–80 in-dicating that the C- but not the N-domain can bind by itself toNR1 C0 in the absence of Ca2�. With Ca2� present, both domains
FIG. 8. Binding of Ca2�-depleted and Ca2�-saturated CaM to afull-length NR1 C-terminal GST fusion protein. GST and the GSTfusion protein carrying the C-terminal C0 and C2� regions of NR1 wereimmobilized on glutathione-Sepharose, washed, incubated with 0.1–50�M CaM � 10 mM CaCl2, quickly washed twice and analyzed by immu-noblotting with anti-CaM. A shows the immunosignals obtained afterpull-down with the C terminus of NR1 (top) and GST alone (bottom)with equal exposure times. B, immunosignals were quantified afterdigitalization by determining the mean density with Adobe Photoshop.Specific binding of CaM to NR1-GST was determined by subtracting thevalues obtained for GST alone from those of NR1-GST for the respectiveCaM concentration. Bars represent averages from three independentexperiments � S.D.
FIG. 7. Binding of Ca2�-depleted and Ca2�-saturated CaM tofull-length NR1. Glutamate receptor complexes were solubilized anddissociated with 1% SDS before immunoprecipitation of NR1 or GluR1,in vitro incubation with 50 �M purified CaM � 10 mM Ca2� (as indicatedat the top), two quick washing steps, and immunoblotting with anti-NR1 (top panel; 100 kDa range of blot), anti-GluR1 (middle panel;reprobing of 100 kDa range of blot) and anti-CaM (17 kDa range). Aspositive control for the anti-CaM antibody, 20 �l of extract was directlyapplied to SDS-PAGE. CaM specifically interacted with NR1 underCa2�-saturated and Ca2�-depleted conditions. Immunosignals for cal-modulin were quantified subsequent to digitalization with a UMAXUTA1100 scanner with Adobe Photoshop. Background signals from thefilm were determined from neighboring unstained fields and subtracted(less than 10% of total signals). Mean density values obtained afterpull-down with NR1 and GluR1 (control for nonspecific binding) immu-noprecipitates were 96.5 � 11.0 and 30.3 � 12.4 (n � 4, p � 0.01,Student’s t test), respectively.
Apo-Calmodulin Binds to the NMDA Receptor2172
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

bound independently from each other to NR1C0p suggesting thatthe C-domain is not necessary for N-domain binding. Because theN-domain is capable of binding NR1C0p on its own and becausebinding of Ca2�-CaM1–80 causes a pronounced effect in the fluo-rescence spectrum of Trp-21, the lack of an effect of apoCaM1–80
suggests that CaM1–80 might not interact with NR1C0p underCa2�-depleted conditions.
The changes in the NR1C0p spectrum are substantiallylarger upon addition of apoCaM1–148 than apoCaM76–148.These results suggest that even though apoCaM1–80 does notbind NR1C0p by itself, the presence of the N-domain does affectthe interaction between the C-domain and NR1C0p. TheTrp-21 spectra in Fig. 2 also are shifted slightly farther to theleft and show a much stronger emission upon addition of Ca2�-CaM than of apoCaM. This difference may imply that theapoCaM-NR1C0p complex undergoes an additional conforma-tional change upon Ca2� binding (see also Figs. 4 and 6 asdiscussed below). Although this spectroscopic effect could bedue to increased binding as the affinity of CaM for NR1C0p isstrongly augmented by Ca2�, further observations and consid-erations discussed below support the model of conformationalalterations within the apoCaM -NR1C0p complex upon addi-tion of Ca2�.
Results similar to those for Trp-21 were obtained when thefluorescence emission of the fluorescein of NR1C0p was moni-tored. We found clear increases in emission intensity uponaddition of Ca2�-saturated CaM1–148, CaM1–80, or CaM76–148
and of apoCaM1–148 or apoCaM76–148 but not apoCaM1–80 (Fig.3). Again, these changes indicate that the C-domain can bind toNR1C0p independent of Ca2� and the N-domain can bind toNR1C0p by itself without the C-domain but only when Ca2� istk;2present. Because the fluorescein tag is located at the Nterminus of NR1C0p, these results also show that binding ofCaM76–148 affects that end of the peptide. The increase inemission is substantially larger with the addition of full-lengthCaM. Because apoCaM76–148 has a slightly higher apparentaffinity for NR1C0p than apoCaM1–148 (Kd of 1.21 �M versus2.25 �M; see “Results”), this increase likely reflects that thepresence or absence of the N-domain in the context of full-length CaM does influence the interaction between the C-domain and NR1C0p (see also Fig. 2 and preceding discussionof the Trp-21 fluorescence). In contrast to the Trp-21 spectra,addition of Ca2� to NR1C0p complexed to apoCaM1–148 orapoCaM76–148 does not cause any substantial further increasesin the fluorescein spectra. However, this behavior of the fluo-rescein group is not unexpected based on the nature and posi-tion of this fluorophore. It is likely that fluorescein at the Nterminus of the peptide would report in general terms on thepresence or absence of CaM bound to the peptide. In contrast,the side chain of Trp-21 presumably interacts with a hydropho-bic cleft of CaM, and accordingly its environment would besensitive to a Ca2�-induced conformational switch of CaM,while remaining associated with NR1C0p throughout thetransition.
Because similar molar concentrations of CaM1–148 andCaM76–148 were used for both experiments, these observationsargue against the possibility that the increase in Trp-21 fluo-rescence emission upon addition of Ca2� is due to increasedaffinity of CaM for the peptide and therefore more completebinding of CaM. If the fractional abundance of CaM-NR1C0pcomplex had increased due to Ca2� binding, an elevation offluorescence emission should also have been detected for thefluorescein spectra when Ca2� was added. Collectively thesefindings suggest instead that binding of Ca2� to a complexconsisting of apoCaM bound to NR1C0p causes additional con-formational changes that have strong effects on Trp-21, but
little if any impact on the fluorescein moiety at the N terminusof this peptide.
Circular dichroism showed that the largely �-helical struc-ture of apoCaM1–148 becomes further ordered upon addition ofCa2� (Fig. 4; compare solid green traces in A and B) as has beenobserved for many years (34). In contrast, NR1C0p did notshow any ellipticity by itself whether Ca2� was present or not(Fig. 4, black traces in A and B). When NR1C0p was added toCaM1–148, the decrease in ellipticity indicated that the peptideinduced ordered secondary structure both in the presence andin the absence of Ca2�. Although this change in ellipticitycannot be assigned to specific changes in the secondary struc-ture of CaM or NR1C0p or both, it is likely that binding to CaMinduces an �-helical conformation in NR1C0p because (a) theaddition of TFE to biotin-NR1C0p demonstrated that the se-quence can adopt a helical structure (Fig. 5) and (b) structuresof CaM-containing protein complexes show that the CaM bind-ing sequences form an �-helix when bound to Ca4
2�-CaM (Fig.1) (12, 14). Regardless of the microscopic details of the struc-ture, changes in ellipticity of the CaM solution upon addition ofNR1C0p clearly demonstrate that CaM1–148 and NR1C0p in-teract not only under Ca2�-saturated but also under Ca2�-depleted (apo) conditions.
The same observations and considerations hold true forCaM76–148; however, experiments with CaM1–80 revealed achange in ellipticity only in the presence (Fig. 4, B and D) butnot in the absence of Ca2� (Fig. 4, A and C). Like the Trp-21and fluorescein fluorescence measurements discussed above,these data indicate that the interaction of NR1C0p with theC-domain occurs independent of Ca2� but NR1C0p binding tothe N-domain requires Ca2�. The decreases in ellipticity uponaddition of NR1C0p to either CaM1–148 or CaM76–148 weremuch smaller in the absence than in the presence of Ca2�.These findings support the model that adding Ca2� to a pre-existing complex of NR1C0p bound to apoCaM causes a distinctconformational change in addition to that observed upon theinteraction of apoCaM with NR1C0p.
Fluorescence anisotropy monitors the rotational mobility ofNR1C0p. Addition of both apoCaM1–148 and apoCaM76–148 toNR1C0p increased its fluorescence anisotropy, indicating theformation of a complex of CaM-NR1C0p having larger radiusand lower mobility than NR1C0p alone (Fig. 6, A and C, leftpanels). This effect was concentration-dependent and saturableconfirming the specificity of these apoCaM-NR1C0p interac-tions. When the CaM-saturated apoCaM1–148-NR1C0p complexwas titrated with Ca2� (Fig. 6A, right panel), the anisotropy ofthe complex substantially dropped with increasing Ca2� con-centrations. This effect was also saturable. In contrast, theanisotropy of the apoCaM76–148-NR1C0p complex did notchange upon addition of Ca2�. The simplest explanation forthese observations is that apoCaM1–148 bound to NR1C0p viathe C-domain alone. In this complex, apoCaM1–148 existedmainly in an extended conformation with a larger hydrody-namic radius and therefore reduced mobility (see schematic inFig. 6). Upon binding of Ca2�, CaM1–148-NR1C0p collapsed to amuch more compact conformation with concomitant associationof the N-domain with NR1C0p. In support of this model is theobservation that the fluorescence anisotropy of NR1C0p did notchange with increasing amounts of apoCaM1–80, indicatingthat the N-domain requires Ca2� to bind to NR1C0p.
Together with results from the other spectroscopic methodsthis finding indicates that apoCaM1–80 does not bind by itself toNR1C0p under these conditions. The lack of effect ofapoCaM1–80 on NR1C0p anisotropy is perhaps the strongestindication that apoCaM1–80 does not associate by itself withNR1C0p, because any binding should result in reduced rota-
Apo-Calmodulin Binds to the NMDA Receptor 2173
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

tional mobility and thereby elevated anisotropy of NR1C0p.However, addition of Ca2� to the solution containingapoCaM1–80 and NR1C0p did increase the anisotropy ofNR1C0p in a concentration-dependent manner reflecting aCa2�-dependent interaction of CaM1–80 with NR1C0p, as seenwith the other spectroscopic approaches. Accordingly, the sim-plest and therefore most attractive model for the interaction ofCaM with NR1C0p is that the C- but not the N-domain directlybinds to NR1C0p under Ca2�-depleted conditions causingapoCaM1–148 to adopt an extended structure in its complexwith NR1C0p. Addition of Ca2� promotes binding of the N-domain to NR1C0p, which results in a more compact structure.This model is identical to one proposed by us previously forCaM binding to a helical peptide (19).
Changes in the fluorescence emission spectra of Trp-21 andfluorescein, as well as decreases in ellipticity observed withapoCaM1–148 were higher than with apoCaM76–148. These dif-ferences are not likely to be due to increased binding ofapoCaM1–148 because apoCaM76–148 has an affinity similar tothat of CaM1–148 (apparent Kd of 1.21 versus 2.25 �M; Fig. 6).The possibility that the N-domain binds in addition to theC-domain in the apoCaM1–148-NR1C0p complex also appearsunlikely because the fluorescence anisotropy of the CaM1–148-NR1C0p complex is much larger under Ca2�-depleted thanCa2�-saturated conditions indicating an extended confor-mation of apoCaM in its complex with NR1C0p, with the N-domain likely to be disordered in its orientation relative toNR1C0p and therefore occupying a greater volume. Further-more, all these experiments support the conclusion thatCaM1–80 does not bind by itself under Ca2�-depleted condi-tions to NR1C0p. The higher (less favorable) Kd value ofapoCaM1–148 as compared with that of apoCaM76–148 for bind-ing to NR1C0p also argues against a simultaneous associationof the N-domain in the apoCaM1–148-NR1C0p complex, as asecond interaction would be expected to reduce rather thanincrease the Kd value. However, the difference in Kd valuesindicates that the presence of the N-domain affects the inter-action of the C-domain with NR1C0p under apo conditions.Therefore, the observation that several spectroscopic effectsare stronger for apoCaM1–148 than for apoCaM76–148 is inter-preted to mean that the presence of the N-domain in the full-length CaM alters the conformation of the C-domain within aCaM-NR1C0p structure, making it different from that ofapoCaM76–148 bound to NR1C0p.
In addition to the studies of peptides binding to CaM, weused full-length NR1 isolated from brain and a GST fusionprotein of the shortest splice variant of the C terminus of NR1for pull-down experiments of CaM. Specific binding was ob-served under Ca2�-saturated and Ca2�-depleted conditionswhen a fast washing protocol was applied (Figs. 7 and 8). Thecontamination level of our Ca2�-free buffer was 2.6 �M Ca2� asdetermined by atomic absorption spectroscopy; using the pro-gram for calculating the amount of free Ca2� in various buffersin IgorPro/PatcherPowertools we estimate a free Ca2� concen-tration of 2.2 nM in this buffer. Because the detection limit forbinding of Ca2�-saturated calmodulin to NR1 C0 is in the rangeof 100 nM (Fig. 8A, upper left panel), the bulk of calmodulindetected after pull-down experiments with either full-lengthNR1 from rat brain (Fig. 7) or GST-NR1 C0 (Fig. 8) is apo-calmodulin. From a physiological point of view the most criticalquestion is not necessarily whether calmodulin can bind to theNMDA receptor in the total absence of Ca2� but whether it ispre-associated with NR1 C0 under resting conditions inside thecell, i.e. at Ca2� concentrations in the range of 50 nM. Our dataindicate that the NR1 C0 region can act as an attachment sitefor calmodulin at resting conditions.
Accurate determination of binding constants for CaM usingthe pull-down procedure is not possible because of the rela-tively fast off rates and therefore the inevitable loss of a largeportion of CaM during the washing steps. This loss was muchless dramatic under Ca2�-saturated conditions suggesting thatapoCaM has a much faster off rate than Ca2�-CaM. The ob-served binding of apoCaM to NR1 extracted from brain and toGST-C0-C2� after a 1.5-min washing procedure suggests thatthe dwell time for docking of CaM at the NR1 subunit underour conditions is in the range of at least 5 s but more likelybetween 10–30 s. If the dwell time were shorter than 5 s, allspecifically bound apoCaM should have completely disap-peared as the washing procedure would have exceeded 20 half-lives. The dwell time may be further increased in vivo due tomolecular crowding and because NMDA receptors likely con-sist of two NR1 subunits, providing at least 2 binding sites perreceptor complex for apoCaM and thereby increasing avidity.Even short-lived interactions should help to increase the con-centration of apoCaM in the vicinity of the NMDA receptor overtime. Unfortunately it is currently not possible to determinethe true dwell time of apoCaM next to or associated with theNMDA receptor in vivo. Furthermore, interactions with Kd
values in the �M range are often considered as being physio-logically relevant (e.g. Ref. 35). Two examples that are well-characterized both functionally and biochemically are the in-teractions of the Ca2� channel Cav2.1 with syntaxin andSNAP25, both of which showed apparent Kd values in therange of 1–2 �M (e.g. (36)).
Physiologically the most important finding of this study isthat apoCaM has the potential to pre-associate with the NR1subunit of the NMDA receptor. Analogous interactions ofapoCaM with various voltage-gated ion channels help to medi-ate their regulation by Ca4
2�-CaM in an efficient and specificmanner (20–23, 25). Association of apoCaM with NR1C0p maybe of physiological importance for two reasons. The area prox-imal to the NMDA receptor experiences the largest and fastestincrease in free Ca2� as it enters the cell through this receptor.Accordingly, NMDA receptor-associated CaM can be activatedmore quickly and effectively than free CaM or CaM bound toother proteins. In addition, apoCaM associated with the NMDAreceptor would be able to exert its effect on the receptor fasterand more efficiently than CaM that has to first diffuse and thenbind to the receptor. Furthermore, the pre-association of CaMwith the NMDA receptor may help to sharpen the specificity ofsignaling by Ca4
2�-CaM for the same two reasons, i.e. becauseCaM that is associated with the NMDA receptor is more effec-tively activated upon Ca2� influx through the NMDA receptorthan CaM bound to other proteins and because receptor-asso-ciated CaM is more likely to influence the activity of the NMDAreceptor than diffuse to other proteins and regulate theiractivity.
Earlier results indicated that CaM reduces the NMDA re-ceptor activity by displacing �-actinin and thereby increasingits desensitization upon Ca2� influx (5–7,9). We recently dem-onstrated that �-actinin and CaM bind to the same segment ofthe NR1 C0 region (8). In addition, CaMKII, another criticalplayer in synaptic signaling, interacts with a segment in thatregion that overlaps with the �-actinin and CaM interactionsite (8, 32). Ca2� influx through the NMDA receptor and theresulting stimulation of CaMKII are critical events in synapticplasticity and especially in a long lasting change in thestrength of synaptic signaling known as long term potentiation(37–39). Long term potentiation is thought to underlie learningand memory and those brain functions critically depend onNMDA receptor activity and CaMKII as well. We are onlybeginning to understand the precise interplay of �-actinin,
Apo-Calmodulin Binds to the NMDA Receptor2174
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

CaM, and CaMKII with the NMDA receptor but it appearslikely that it may be critical in fine-tuning synaptic signalingand plasticity.
Excessive Ca2� influx through the NMDA receptor (10, 11)and subsequent overactivation of CaMKII (40) have been im-plicated in neuronal damage during stroke. We do not know theeffect of CaMKII binding to NR1 on the NMDA receptor activ-ity, but the inhibitory effect of Ca4
2�-CaM on the NMDA recep-tor, possibly regulated further by CaMKII binding, may pro-vide a critical negative feedback loop that reduces the NMDAreceptor activity during periods of overstimulation. Under-standing the precise molecular mechanisms of the regulation ofNMDA receptor activity by CaM, CaMKII, and �-actinin willlead to a better understanding of the role of these proteins andtheir interactions with the NMDA receptor complex underphysiological and pathological conditions. Such insight will becritical for a rational approach to the development of treat-ments that will inhibit the downstream effects of increasedCa2� influx through NMDA receptors during ischemia andthereby the resulting neuronal damage.
In this study, we present the first evidence that apoCaM canpre-associate with a fast acting ionotropic neurotransmitterreceptor. Our observations suggest that apoCaM associateswith its C-terminal domain to NR1C0 while the N-domain isnot bound, resulting in an extended conformation. The dataobtained by monitoring the solvated size by fluorescence ani-sotropy indicate that the addition of the Ca2� induces a struc-tural transition such that the N-domain of CaM collapses ontothe NR1C0 region, while the C-domain remains bound. TheCa2�-saturated endstate of this model fits very well with awealth of earlier findings that demonstrate that Ca4
2�-CaMmay adopt a compact structure when bound to a target protein(cf., Fig. 1, 2bbm.pdf (Ca4
2�-CaM bound to MLCK) and analyt-ical ultracentrifugation studies of a peptide bound to Ca4
2�-CaM, Ref. 19). To the best of our knowledge we provide for thevery first time evidence for Ca2�-induced compaction of a com-plex of apoCaM1–148 bound via its C- but not N-terminal do-main to a physiologically relevant target when titrating Ca2�
in (Fig. 6). Augmented by studies of the individual domains,these findings reveal their different roles in associating withNR1 C0.
Pre-association of apoCaM with ion channels via the C-do-main has been studied at a biochemical level in only a couple ofother cases, including SK channels (41) and ryanodine recep-tors (26). However, the NMDA receptor likely has a CaM do-main-specific molecular mechanism of regulation differentfrom those channels. A crystallographic study of the SK chan-nel (42) shows that the CaM-SK channel complex does not haveCa2� bound to the C-domain of CaM (Ca2� is present only inthe N-domain), whereas Ca2� does affect binding of the C-domain fragment of CaM to NR1 C0 (e.g. Figs. 2–4). Theryanodine receptor can also bind apoCaM, but in contrast toNR1 C0 it appears that the N- and C-domains of CaM areassociated with different regions of the ryanodine receptorunder Ca2�-depleted and Ca2�-saturating conditions (43) andthat Ca2� binding shifts the conformation of CaM rather thancausing the N-domain to associate per se (26, 44).
In conclusion, the studies in this work are the first to dem-onstrate apoCaM binding to the NR1 C0 region and thereby toprovide evidence for apoCaM binding to a fast neurotransmit-ter receptor, the first to demonstrate separable roles for thedomains of CaM binding to the NMDA receptor, and the first toshow that Ca2� binding changes the conformation of the C-domain from that assumed when it is pre-associated with theNR1 C0 region.
Acknowledgments—We thank W. S. VanScyoc, B. R. Sorensen, andR. Hultman for purifying the CaM used in these studies, and L. Teeschand E. Rus for amino acid analysis (Molecular Analysis Facility, Uni-versity of Iowa, Iowa City, IA).
REFERENCES
1. Seeburg, P. H. (1993) Trends Pharmacol. Sci. 14, 297–3032. Hollmann, M., and Heinemann, S. (1994) Annu. Rev. Neurosci. 17, 31–1083. Sheng, M., Cummings, J., Roldan, L. A., Jan, Y. N., and Jan, L. Y. (1994)
Nature 368, 144–1474. Blahos, J., II, and Wenthold, R. J. (1996) J. Biol. Chem. 271, 15669–156745. Ehlers, M. D., Zhang, S., Bernhardt, J. P., and Huganir, R. L. (1996) Cell 84,
745–7556. Zhang, S., Ehlers, M. D., Bernhardt, J. P., Su, C. T., and Huganir, R. L. (1998)
Neuron 21, 443–4537. Wyszynski, M., Lin, J., Rao, A., Nigh, E., Beggs, A. H., Craig, A. M., and Sheng,
M. (1997) Nature 385, 439–4428. Leonard, A. S., Bayer, K.-U., Merrill, M. A., Lim, I. A., Shea, M. A., Schulman,
H., and Hell, J. W. (2002) J. Biol. Chem. 277, 48441–484489. Krupp, J. J., Vissel, B., Thomas, C. G., Heinemann, S. F., and Westbrook, G. L.
(1999) J Neurosci 19, 1165–117810. Lee, J.-M., Zipfel, G. J., and Choi, D. W. (1999) Nature 399, (suppl.) A7–A1411. Rothman, S. M., and Olney, J. W. (1987) Trends Neurosci. 10, 299–30212. Hoeflich, K. P., and Ikura, M. (2002) Cell 108, 739–74213. Saimi, Y., and Kung, C. (2002) Annu. Rev. Physiol. 64, 289–31114. Vetter, S. W., and Leclerc, E. (2003) Eur. J. Biochem. 270, 404–41415. Kink, J. A., Maley, M. E., Preston, R. R., Ling, K. Y., Wallen-Friedman, M. A.,
Saimi, Y., and Kung, C. (1990) Cell 62, 165–17416. Kung, C., Preston, R. R., Maley, M. E., Ling, K. Y., Kanabrocki, J. A., Seavey,
B. R., and Saimi, Y. (1992) Cell Calcium 13, 413–42517. VanScyoc, W. S., and Shea, M. A. (2001) Protein Sci. 10, 1758–176818. Jaren, O. R., Harmon, S., Chen, A. F., and Shea, M. A. (2000) Biochemistry 39,
6881–689019. Sorensen, B. R., Eppel, J. T., and Shea, M. A. (2001) Biochemistry 40, 896–90320. Xia, X. M., Fakler, B., Rivard, A., Wayman, G., Johnson-Pais, T., Keen, J. E.,
Ishii, T., Hirschberg, B., Bond, C. T., Lutsenko, S., Maylie, J., and Adelman,J. P. (1998) Nature 395, 503–507
21. DeMaria, C. D., Soong, T. W., Alseikhan, B. A., Alvania, R. S., and Yue, D. T.(2001) Nature 411, 484–489
22. Peterson, B. Z., DeMaria, C. D., Adelman, J. P., and Yue, D. T. (1999) Neuron22, 549–558
23. Lee, A., Wong, S. T., Gallagher, D., Li, B., Storm, D. R., Scheuer, T., andCatterall, W. A. (1999) Nature 399, 155–159
24. Zuhlke, R. D., Pitt, G. S., Deisseroth, K., Tsien, R. W., and Reuter, H. (1999)Nature 399, 159–162
25. Lee, A., Scheuer, T., and Catterall, W. A. (2000) J. Neurosci. 20, 6830–683826. Xiong, L. W., Newman, R. A., Rodney, G. G., Thomas, O., Zhang, J. Z.,
Persechini, A., Shea, M. A., and Hamilton, S. L. (2002) J. Biol. Chem. 277,40862–40870
27. Sorensen, B. R., and Shea, M. A. (1998) Biochemistry 37, 4244–425328. Putkey, J. A., Slaughter, G. R., and Means, A. R. (1985) J. Biol. Chem. 260,
4704–471229. Pedigo, S., and Shea, M. A. (1995) Biochemistry 34, 10676–1068930. Johnson, M. L., and Frasier, S. G. (1985) Methods Enzymol. 117, 301–34231. Leonard, A. S., and Hell, J. W. (1997) J. Biol. Chem. 272, 12107–1211532. Leonard, A. S., Lim, I. A., Hemsworth, D. E., Horne, M. C., and Hell, J. W.
(1999) Proc. Natl. Acad. Sci. U. S. A. 96, 3239–324433. Leonard, A. S., Davare, M. A., Horne, M. C., Garner, C. C., and Hell, J. W.
(1998) J. Biol. Chem. 273, 19518–1952434. Klevit, R. E. (1983) Methods Enzymol. 102, 82–10435. Pawson, T., and Nash, P. (2003) Science 300, 445–45236. Rettig, J., Sheng, Z. H., Kim, D. K., Hodson, C. D., Snutch, T. P., and Catterall,
W. A. (1996) Proc. Natl. Acad. Sci. U. S. A. 93, 7363–736837. Lisman, J. E., and Zhabotinsky, A. M. (2001) Neuron 31, 191–20138. Bliss, T. V., and Collingridge, G. L. (1993) Nature 361, 31–3939. Malenka, R. C., and Nicoll, R. A. (1999) Science 285, 1870–187440. Hajimohammadreza, I., Probert, A. W., Coughenour, L. L., Borosky, S. A.,
Marcoux, F. W., Boxer, P. A., and Wang, K. K. W. (1995) J. Neurosci. 15,4093–4101
41. Keen, J. E., Khawaled, R., Farrens, D. L., Neelands, T., Rivard, A., Bond, C. T.,Janowsky, A., Fakler, B., Adelman, J. P., and Maylie, J. (1999) J. Neurosci.19, 8830–8838
42. Schumacher, M. A., Rivard, A. F., Bachinger, H. P., and Adelman, J. P. (2001)Nature 410, 1120–1124
43. Zhang, H., Zhang, J. Z., Danila, C. I., and Hamilton, S. L. (2003) J. Biol. Chem.278, 8348–8355
44. Samso, M., and Wagenknecht, T. (2002) J. Biol. Chem. 277, 1349–135345. Kuboniwa, H., Tjandra, N., Grzesiek, S., Ren, H., Klee, C. B., and Bax, A.
(1995) Nat. Struct. Biol. 2, 768–77646. Rao, S. T., Wu, S., Satyshur, K. A., Ling, K. Y., Kung, C., and Sundaralingam,
M. (1993) Protein Sci. 2, 436–44747. Drum, C. L., Yan, S. Z., Bard, J., Shen, Y. Q., Lu, D., Soelaiman, S., Grabarek,
Z., Bohm, A., and Tang, W. J. (2002) Nature 415, 396–40248. Elshorst, B., Hennig, M., Forsterling, H., Diener, A., Maurer, M., Schulte, P.,
Schwalbe, H., Griesinger, C., Krebs, J., Schmid, H., Vorherr, T., andCarafoli, E. (1999) Biochemistry 38, 12320–12332
49. Ikura, M., Clore, G. M., Gronenborn, A. M., Zhu, G., Klee, C. B., and Bax, A.(1992) Science 256, 632–638
50. Kraulis, P. J. (1991) J. Appl. Crystallogr. 24, 946–950
Apo-Calmodulin Binds to the NMDA Receptor 2175
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Madeline A. Shea and Johannes W. HellZeynep Akyol, Jason A. Bartos, Michelle A. Merrill, Laurel A. Faga, Olav R. Jaren,
Receptor NR1 C0 Region-Methyl-d-aspartateNApo-Calmodulin Binds with its C-terminal Domain to the
doi: 10.1074/jbc.M302542200 originally published online October 6, 20032004, 279:2166-2175.J. Biol. Chem.
10.1074/jbc.M302542200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/279/3/2166.full.html#ref-list-1
This article cites 50 references, 17 of which can be accessed free at
by guest on August 10, 2016
http://ww
w.jbc.org/
Dow
nloaded from




















